
Submitted:
28 August 2023
Posted:
29 August 2023
You are already at the latest version
Abstract
Melanoma is a highly aggressive type of skin cancer produced by malignant transformation of melanocytes and it is usually associated with a poor prognosis. Clinically, melanoma has several stages associated with migration and invasion of the cells through the skin layers, rapid cell spreading and formation of tumors in multiple organs. The main problem is the emergence of resistance of melanoma to the applied methods of treatment, thus it is of primary importance to find more crucial signaling pathways that control the progression of this type of cancer and could be targeted to prevent melanoma spreading. The main life-threatening stage of melanoma is metastasis formation, and 3D cell spheroids are better suited for the study of this process. Using a combinative approach (RT-PCR, immunofluorescence, patch-clamp and calcium imaging) we showed the functional expression of mechanosensitive Piezo1 channels in SK-MEL-2 aggressive melanoma cell line. We found that chemical activation of Piezo1 by its selective agonist Yoda1 completely prevents melanoma spheroid formation, and could be considered as a novel approach for the prevention of melanoma development.
Keywords:
piezo1 channel
; melanoma
; Yoda1
; 3D spheroids
Introduction
Melanoma is a highly aggressive type of skin cancer produced by malignant transformation of melanocytes and it is usually associated with a poor prognosis. Clinically, melanoma has several stages associated with migration and invasion of the cells through the skin layers, rapid cell spreading and formation of tumors in multiple organs. Nowadays, active development of novel targeted drugs is underway, but the main problem is the emergence of resistance of melanoma to the applied methods of treatment [1,2]. Therefore, it is of primary importance to find more crucial signaling pathways that control the progression of this type of cancer. Mechanosensitive ion channels and molecules, especially those that mediate Ca2+ influx into the cell, regulate cancer development, metabolism and progression [3,4]. Recently, it was shown that mechanosensitive TRPV2 channel regulates melanoma invasiveness and metastatic potential [3]. The high expression of Piezo1, another important mechanosensitive Ca2+- permeable ion channel, was detected in melanoma [3], and was shown to control its malignant progression [4]. The main life-threatening stage of melanoma is metastasis formation, and 3D spheroids are better suited for the study of this process due to imitation in vivo conditions that are observed during melanoma progression [5]. Here, we evidenced functional activity of Piezo1 channels in aggressive melanoma SK-MEL-2 cell line. We found that chemical activation of Piezo1 by its selective agonist Yoda1 completely prevents melanoma spheroid formation and thus could be considered as a novel targeted approach for the prevention of melanoma development.
Materials and Methods
Cells and reagents
SK-MEL-2 human malignant melanoma cell line (ATCC® HTB-68™, RRID: CVCL_0069) was purchased from Biolot, St. Petersburg, Russia. Cells were cultured in RPMI 1640 medium (Biolot, Russia) containing 10% fetal bovine serum (Biowest, Rui de Caille, France) and antibiotic gentamicin (Biolot, Russia) in humidified incubator at 37o C and 5% CO2. For patch-clamp studies, fluorescent staining and Ca2+-imaging, the cells were plated on glass coverslips in 2-3 days before the experiments. Selective chemical Piezo1 activator Yoda1 was purchased from Tocris Bioscience (Bristol, United Kingdom). Ca2+ ionophore ionomycin was purchased from Thermo Fischer Scientific (Waltham, MA, USA).
RT-PCR
Total RNA and cDNA were obtained as described in previous studies [6,7]. The primer sequences for hPIEZO1 were: 3’-CCAGAACAGGTATCGGAAG-5’ (reverse) and 5’-TGCTGTACCAGTACCTGCTG-3’ (forward). The cycling conditions were as in previous studies [8]. The number of PCR cycles was 30, and the optimal annealing temperature for hPIEZO1 primers is 57 °C.
Electrophysiology
The detailed description of the whole-cell mode of patch-clamp technique used to record single ion channels in the plasma membrane is available in our publications [8,9]. Cytosol-like solution contained (in mM): 140 KAsp, 5 NaCl, 1 MgCl2, 2 EGTA, 20 HEPES/TrisOH and CaCl2 to establish intracellular Ca2+ concentration ([Ca2+]i) at 10 nM. The extracellular chamber solution contained: 145 NaCl, 2 CaCl2, 1 MgCl2, and 10 HEPES/TrisOH. pH of all solutions was set at 7.3. Yoda1 was applied to the extracellular solution.
Immunofluorescence
Polyclonal primary antibodies against extracellular domain of Piezo1 were used (KD/KO validated, from Proteintech, Manchester, United Kingdom, Cat No 15939-1-AP, RRID: AB_2231460). Cells were fixed with 4% paraformaldehyde (15 min at room temperature, RT), blocked with 10% Goat serum (1 h, RT) and incubated with 1:100 Anti-Piezo1 antibodies (overnight, at 4o C). Then the cells were incubated with 1:200 secondary goat anti-rabbit-Cy3 fluorescent antibodies (Santa Cruz, TX, USA, for 1 h, RT). Cell nuclei were counterstained with DAPI (0.05 μg/ml, 30 min, RT). Cells were mounted on glass slides using Vectashield mounting media (Vector Laboratories, Newark, CA, USA) and visualized using Olympus FV3000 (Olympus Corporation, Shinjuku, Tokyo, Japan) confocal microscope.
Calcium imaging
Ca2+-measurements were performed as described earlier [7]. Briefly, the cells were loaded with 5 μM of Fluo8-AM fluorescent Ca2+ probe (AAT Bioquest, Pleasanton, CA, USA) for 45 min in Ca2+-containing solution (at 37o C and 5% CO2, in dark), then the cells were incubated for 15 min without the dye. The Ca2+-containing solution consists of (in mM): 150 NaCl, 4.5 KCl, 2 CaCl2, 1 MgCl2 and 10 HEPES/TrisOH. At the start of the experiment, cells were placed in Ca2+-containing solution, and basal fluorescence signals were recorded for 15 s. After that, 10 µM Yoda1 was applied to the cells. The experiment was performed at least 3 times.
Spheroid formation assay
The melanoma spheroids were formed using a hanging drop method. 5 independent experiments with total number of 140 hanging drops for each experimental condition were conducted. Firstly, SK-MEL-2 cells were trypsinized, collected in culture medium and counted using automatic Cell Counter Countess II (Thermo Fisher Scientific, Waltham, MA, USA). The cell concentration in suspension was adjusted to 2x105 cells/ml (equal to 7.000 of cells in 35 µl drop). The total cell volume was divided into two, and 10 µM of Yoda1 or equal amount of DMSO (vehicle, 0.1%) was added. The drops were deposited onto the cover of the Petri dish, the drops were overlaid, transferred to the CO2 incubator, and monitored daily using the upright digital microscope. After 14 days of formation, the drops were collected and plated on Petri dishes to obtain microphotographs of the spheroids.
Cell spreading and morphology assays
Cells were trypsinized, resuspended in full medium, and vehicle (0.3% DMSO) or 30 µM Yoda1 were added to the cells. After that, cell suspensions were plated on Petri dishes. At least five fields of view for control or Yoda1-treated cells were captured every 20 min, and a number of spreaded/non-spreaded cells was counted. For morphology analysis, the contours of the cells were traced manually in ImageJ using the “Freehand selection” tool. Cell area and shape descriptors were measured using the “Analyze - Measure” commands. A total number of 100-110 cells was analyzed for each experimental condition at each timepoint.
Statistics
The statistical analysis was performed in GraphPad Prism 8.0 (GraphPad Software, USA), p<0.05 was considered as significant. The statistical tests used for data comparison are indicated in the corresponding figure legends.
Results and Discussion
Firstly, we performed RT-PCR to confirm the presence of hPIEZO1 mRNA in SK-MEL-2 cells (Figure 1A). Further, to record the functional activity of endogenous Piezo1 channels in the plasma membrane, we used the patch-clamp technique. Previously, we have shown that 1 µM Yoda1, a selective agonist of Piezo1, could evoke single Piezo1-mediated channel activity in whole-cell experiments [8]. Consistently, we recorded single-channel Piezo1 currents in melanoma cells in response to the application of 1 µM Yoda1. The single channel properties of Yoda1-induced Piezo1 channels (Figure 1B) were similar to those reported earlier [8]. The channels were activated in 7 out of 9 experiments indicating that several cells seem to have no functionally active Piezo1 in the plasma membrane. Consistently, immunofluorescent staining revealed that part of SK-MEL-2 cells was Piezo1-negative (Figure 1C). As Piezo1 channels provide physiologically relevant pathways for Ca2+ entry in various cells and tissues, the Ca2+- measurements were done for additional verification of obtained results (Figure 1D). We found that Yoda1 application resulted in [Ca2+]i increase further confirming the presence of functional Piezo1 channels in SK-MEL-2 cells. Interestingly, Ca2+ imaging allowed to distinguish three types of cell responses: (1) cells with significant increases in [Ca2+]i (about 2-fold, red line), (2) with slow moderate increases (about 1.3 fold, observed in majority of cell population, blue line), and (3) cells with no Ca2+ entry in response to Yoda1 (green line). The results confirm the variations in Piezo1 expression in SK-MEL-2 cells. Taken together, our data show that SK-MEL-2 cells have functionally active Piezo1 channels in the plasma membrane, whose activity could be evoked by Yoda1, a selective chemical Piezo1 activator. The intriguing result is the presence of a number of the cells in which no Piezo1 staining or activity was detected. This phenomenon could be explained by various factors that could be revealed and analyzed in further studies.
Cell aggregation in 3D spheroids is an important step in melanoma progression where cells primarily have to change their microenvironment and mechanical properties as well as morphology, polarization and cell-cell contacts. These steps could be critically controlled by the functioning of various ion transporters, including mechanosensitive channels. Thus, we probed if selective chemical activation of Piezo1 could have any effect on melanoma spheroids by adding 10 µM Yoda1 to SK-MEL-2 suspension during the formation of spheroids. Importantly, we detected the perturbation of spheroid formation in the presence of Yoda1: firstly, no compaction of the cells was observed (Figure 2A), whereas control cells became compacted after 7 days from the start of spheroid formation, and the compaction continued (to the 14th day). Further (after 14 days), the control- and Yoda1-containing drops were transferred to the Petri dish to confirm stable spheroid formation. Evidently, no spheroids were formed in the presence of Yoda1, whereas control cells formed distinct compact spheroids (Figure 2B). After 24 hours on the adherent surface, the cells from the control spheroids have spread out, whereas the Yoda1-treated cells remain round, are unable to spread and are non-viable (Figure 2B). Thus, the addition of Yoda1 completely blocked the spheroid formation from SK-MEL-2 aggressive melanoma cell line.
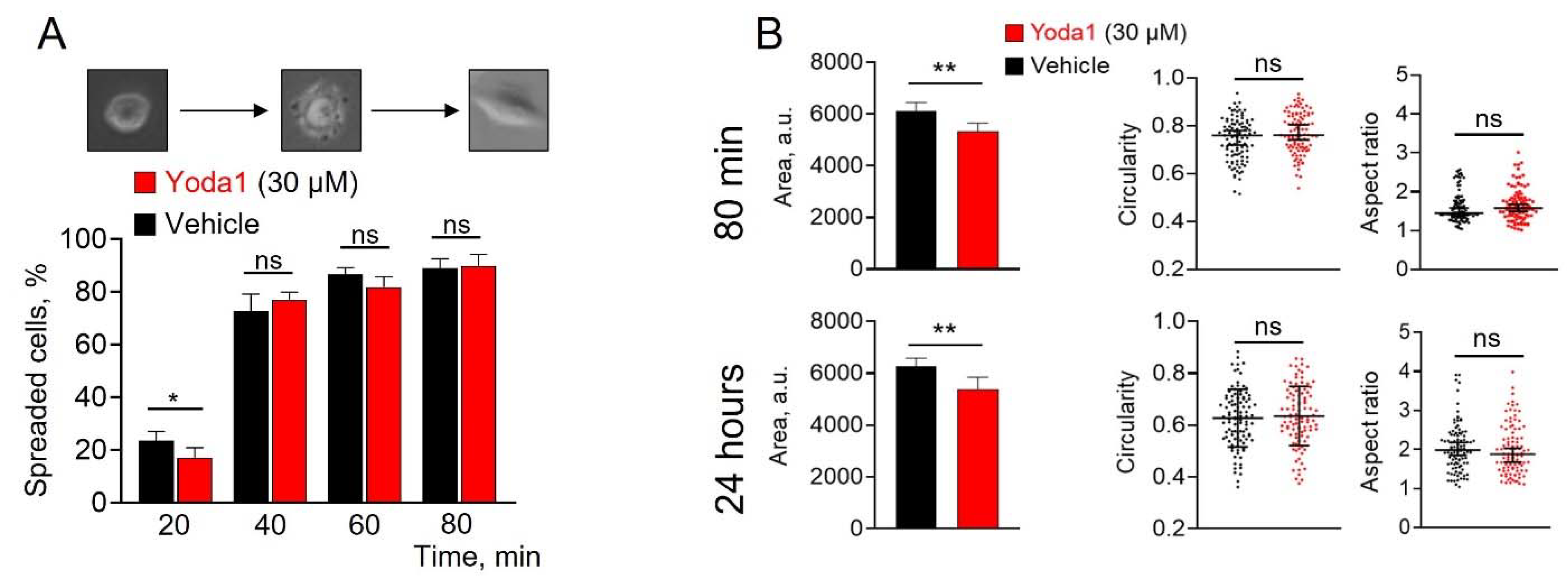
The first key step of spheroid formation is the transfer of the cells to suspension, which significantly affects their shape and plasma membrane curvature compared to adherent cells. As membrane curvature was shown to strongly affect Piezo1 distribution [10], as well as the activating potential of Yoda1 [11], we analyzed the action of Yoda1 on the SK-MEL-2 cells in the suspension. We hypothesized, that Yoda1 could have a strong effect on suspension melanoma cells, and this could potentially underlie the prevention of spheroid formation. Thus, we tested if the addition of Yoda1 to cell suspension could affect their spreading and viability (Figure 3). Our assays showed only a small (or non-significant effect) of 30 µM Yoda1 (note the higher concentration used compared to spheroid formation) on SK-MEL-2 cells. Particularly, the number of spreaded cells in the presence of Yoda1 was slightly lower at the early stage (after 20 minutes), however no significant changes were detected at later time points (Figure 3A) that also confirms the viability of the cells. Also, Yoda1 had no effect on cell shape, but caused a slight decrease in cell size (Figure 3B). Thus, the lack of significant effect of Yoda1 on melanoma cell suspension could indicate, that the perturbation of melanoma spheroid formation in the presence of Yoda1 occurs at latter stages of the process. It should be noted that no specific cell-cell interactions occur between the cells in suspension (at least during a short timescale) or during cell spreading, where cell-surface interactions prevail. Thus, we speculate that Piezo1 activation probably affects cell-cell interactions that are observed in 3D culture of SK-MEL-2 cells. It is of specific interest to analyze the mechanisms that underlie the inhibition of melanoma spheroid formation by selective activation of Piezo1 channels. 3D structure formation occurs through the sequential establishment of cell-cell contacts in which adhesion molecules such as integrins and cadherins are the key players [12]. Interestingly, both cadherins and integrins were shown to interact with or be regulated by Piezo1, and v.v. [13,14]. Thus, the study of the possible regulation of integrin/cadherin activity and interactions by Piezo1 during spheroid formation could be a promising experimental goal of future research in the prevention of tumor formation.
Author Contributions
Valeria Y. Vasileva: Conceptualization; Data curation; Formal analysis; Investigation; Validation; Visualization; Writing - original draft; review & editing. Zuleikha M. Khairullina: Data curation; Investigation; Visualization. Vladislav I. Chubinskiy-Nadezhdin: Conceptualization; Data curation; Formal analysis; Funding acquisition; Investigation; Methodology; Project administration; Resources; Supervision; Validation; Visualization; Writing: original draft, review & editing.
Grants
The Russian Science Foundation, grant number: 22-74-10037 (to C-N VI).
Data Availability
The data is available upon request.
Disclosures
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Winder, M.; Virós, A. Mechanisms of Drug Resistance in Melanoma. Handb Exp Pharmacol. 2018, 249, 91–108. [Google Scholar] [CrossRef] [PubMed]
- Kozar, I.; Margue, C.; Rothengatter, S.; Haan, C.; Kreis, S. Many ways to resistance: How melanoma cells evade targeted therapies. Biochim Biophys Acta Rev Cancer. 2019, 1871, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Shoji, K.F.; Bayet, E.; Leverrier-Penna, S.; Le Devedec, D.; Mallavialle, A.; Marionneau-Lambot, S.; Rambow, F.; Perret, R.; Joussaume, A.; Viel, R.; Fautrel, A.; Khammari, A.; Constantin, B.; Tartare-Deckert, S.; Penna, A. The mechanosensitive TRPV2 calcium channel promotes human melanoma invasiveness and metastatic potential. EMBO Rep. 2023, 24, e55069. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Cao, S.; Gong, M.; Zhang, W.; Zhang, W.; Zhu, Z.; Wu, S.; Yue, Y.; Qian, W.; Ma, Q.; Wang, S.; Wang, Z. Mechanically activated ion channel Piezo1 contributes to melanoma malignant progression through AKT/mTOR signaling. Cancer Biol Ther. 2022, 23, 336–347. [Google Scholar] [CrossRef]
- Ana I. Soares, Rui L. Reis, Alexandra P. Marques. 3D in vitro cutaneous melanoma models, in: Subhas C. Kundu, Rui L. Reis, In Materials Today, Biomaterials for 3D Tumor Modeling, Elsevier, 2020, pp. 287–303.
- Vasileva, V.Y.; Morachevskaya, E.A.; Negulyaev, Y.A. Analysis of Possible Mechanisms of Endometrial Stem Cell Migration Suppression by Selective Chemical Activation of Piezo1 Mechanosensitive Channels. Cell Tiss. Biol. 2022, 16, 599–607. [Google Scholar] [CrossRef]
- Vasileva, V.Y.; Khairullina, Z.M.; Sudarikova, A.V.; Chubinskiy-Nadezhdin, V.I. Role of Calcium-Activated Potassium Channels in Proliferation, Migration and Invasion of Human Chronic Myeloid Leukemia K562 Cells. Membranes 2023, 13, 583. [Google Scholar] [CrossRef] [PubMed]
- Vasileva, V.; Morachevskaya, E.; Sudarikova, A.; Negulyaev, Y.; Chubinskiy-Nadezhdin, V. Selective Chemical Activation of Piezo1 in Leukemia Cell Membrane: Single Channel Analysis. Int J Mol Sci. 2021, 22, 7839. [Google Scholar] [CrossRef] [PubMed]
- Sudarikova, A.V.; Vasileva, V.Y.; Vassilieva, I.O.; Negulyaev, Y.A.; Morachevskaya, E.A.; Chubinskiy-Nadezhdin, V.I. Extracellular protease trypsin activates amiloride-insensitive sodium channels in human leukemia cells. J Cell Biochem. 2019, 120, 461–469. [Google Scholar] [CrossRef]
- Yang, S.; Miao, X.; Arnold, S.; Li, B.; Ly, A.T.; Wang, H.; Wang, M.; Guo, X.; Pathak, M.M.; Zhao, W.; Cox, C.D.; Shi, Z. Membrane curvature governs the distribution of Piezo1 in live cells. Nat Commun. 2022, 13, 7467. [Google Scholar] [CrossRef] [PubMed]
- Botello-Smith, W.M.; Jiang, W.; Zhang, H.; Ozkan, A.D.; Lin, Y.C.; Pham, C.N.; Lacroix, J.J.; Luo, Y. A mechanism for the activation of the mechanosensitive Piezo1 channel by the small molecule Yoda1. Nat Commun. 2019, 10, 4503. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Hartanto, Y.; Zhang, H. Advances in multicellular spheroids formation. J R Soc Interface. 2017, 14, 20160877. [Google Scholar] [CrossRef] [PubMed]
- Aglialoro, F.; Hofsink, N.; Hofman, M.; Brandhorst, N.; van den Akker, E. Inside Out Integrin Activation Mediated by PIEZO1 Signaling in Erythroblasts. Front Physiol. 2020, 11, 958. [Google Scholar] [CrossRef] [PubMed]
- Chuntharpursat-Bon, E.; Povstyan, O.V.; Ludlow, M.J.; Carrier, D.J.; Debant, M.; Shi, J.; Gaunt, H.J.; Bauer, C.C.; Curd, A.; Simon Futers, T.; Baxter, P.D.; Peckham, M.; Muench, S.P.; Adamson, A.; Humphreys, N.; Tumova, S.; Bon, R.S.; Cubbon, R.; Lichtenstein, L.; Beech, D.J. PIEZO1 and PECAM1 interact at cell-cell junctions and partner in endothelial force sensing. Commun Biol. 2023, 6, 358. [Google Scholar] [CrossRef]
Figure 1.
Functional expression of mechanosensitive Piezo1 channels in SK-MEL-2 cells. A: RT-PCR showing the presence of hPIEZO1 mRNA in SK-MEL-2 cell lysates. The obtained length of product (360 bp) was as expected. B: The representative whole-cell recordings of Yoda1-induced Piezo1 activity in response to 1 µM Yoda1. Single-channel conductance and reversal potential were similar to Yoda1-induced Piezo1 channels described earlier [9]. c - closed state (zero current, baseline). C: Immunofluorescent staining of Piezo1 in SK-MEL-2 cells. Red channel- Piezo1, blue channel - DAPI. Note that the number of cells is Piezo1-negative (marked by the arrows). Scale bar is 50 μm. As a control, the staining of the cells only with fluorescent secondary antibodies was used (no pAb, no red fluorescent signal was observed). D: Ca2+-responses of SK-MEL-2 cells to Yoda1 application in the representative experiment (from three independent Ca2+-measurements) with n=67 cells. Note that three populations of the cells could be distinguished (see Results and Discussion). Shown are mean fold changes (±S.D.) of Fluo8 fluorescence, normalized to the fluorescence at the starting point (first frame). Note that all cell populations had similar reactions to calcium ionophore Ionomycin (5 μM), indicating the specificity of Yoda1-induced Ca2+-responses.
Figure 1.
Functional expression of mechanosensitive Piezo1 channels in SK-MEL-2 cells. A: RT-PCR showing the presence of hPIEZO1 mRNA in SK-MEL-2 cell lysates. The obtained length of product (360 bp) was as expected. B: The representative whole-cell recordings of Yoda1-induced Piezo1 activity in response to 1 µM Yoda1. Single-channel conductance and reversal potential were similar to Yoda1-induced Piezo1 channels described earlier [9]. c - closed state (zero current, baseline). C: Immunofluorescent staining of Piezo1 in SK-MEL-2 cells. Red channel- Piezo1, blue channel - DAPI. Note that the number of cells is Piezo1-negative (marked by the arrows). Scale bar is 50 μm. As a control, the staining of the cells only with fluorescent secondary antibodies was used (no pAb, no red fluorescent signal was observed). D: Ca2+-responses of SK-MEL-2 cells to Yoda1 application in the representative experiment (from three independent Ca2+-measurements) with n=67 cells. Note that three populations of the cells could be distinguished (see Results and Discussion). Shown are mean fold changes (±S.D.) of Fluo8 fluorescence, normalized to the fluorescence at the starting point (first frame). Note that all cell populations had similar reactions to calcium ionophore Ionomycin (5 μM), indicating the specificity of Yoda1-induced Ca2+-responses.
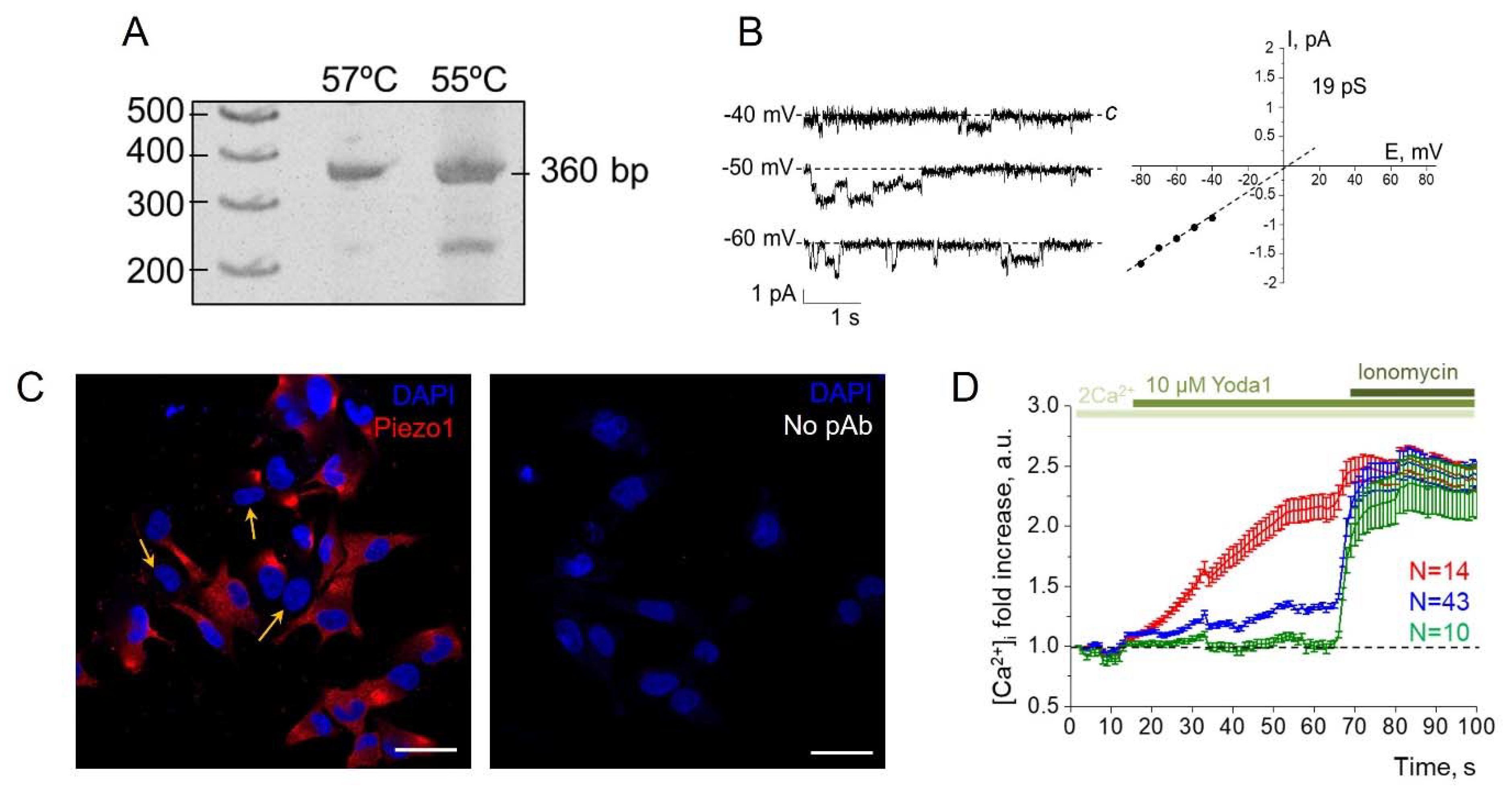
Figure 2.
Yoda1-induced Piezo1 activation prevents spheroid formation from malignant melanoma SK-MEL-2 cells. A: Shown are representative microphotographs of SK-MEL-2 spheroids in control (vehicle, 0.1% DMSO) and in the presence of 10 μM Yoda1 at 7th and 14th day after hanging drop initiation. Note the more compact size of melanoma spheroids under control conditions (with further compaction from 7th and 14th day), compared to Yoda1-treated cells. Scale bar is 300 μm. White circles are the light source (light-emitting diodes) of the upright microscope. B: Representative images of the melanoma spheroids at 0 h and 24 h after their collection and plating on the Petri dishes. Note that no spheroids were formed in the presence of Yoda1 (cell suspension was observed). After 24 h, the melanoma cells migrated from the control spheroids indicating their viable status. At the same time, the cells collected from Yoda1-containing hanging drops remained round (were not able to spread) and were non-viable (were positively stained with propidium iodide, data not shown). Scale bar is 300 μm.
Figure 2.
Yoda1-induced Piezo1 activation prevents spheroid formation from malignant melanoma SK-MEL-2 cells. A: Shown are representative microphotographs of SK-MEL-2 spheroids in control (vehicle, 0.1% DMSO) and in the presence of 10 μM Yoda1 at 7th and 14th day after hanging drop initiation. Note the more compact size of melanoma spheroids under control conditions (with further compaction from 7th and 14th day), compared to Yoda1-treated cells. Scale bar is 300 μm. White circles are the light source (light-emitting diodes) of the upright microscope. B: Representative images of the melanoma spheroids at 0 h and 24 h after their collection and plating on the Petri dishes. Note that no spheroids were formed in the presence of Yoda1 (cell suspension was observed). After 24 h, the melanoma cells migrated from the control spheroids indicating their viable status. At the same time, the cells collected from Yoda1-containing hanging drops remained round (were not able to spread) and were non-viable (were positively stained with propidium iodide, data not shown). Scale bar is 300 μm.
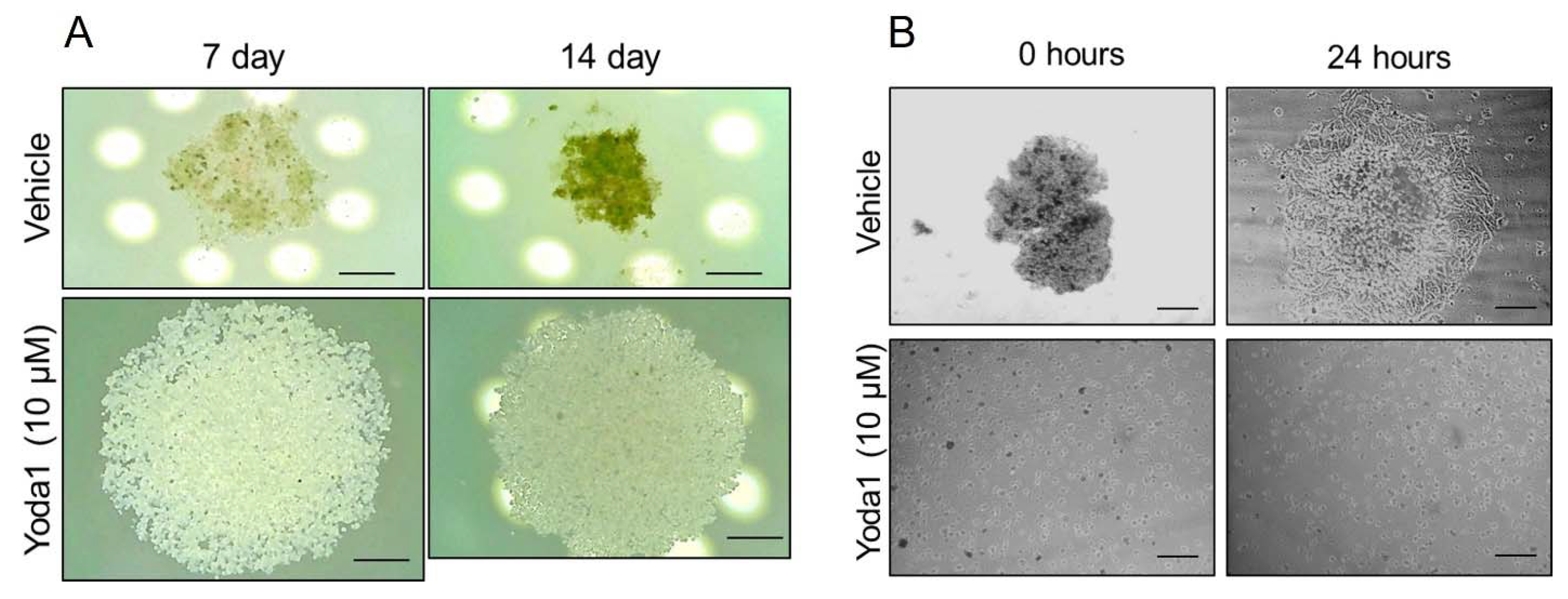
Figure 3.
The effect of Yoda1 on SK-MEL-2 spreading and morphology. A: Microphotographs showing the subsequent steps of SK-MEL-2 spreading. Round cells were considered as non-spreaded (left box), cells at center and right boxes were considered as spreaded. Shown are percentages of spreaded cells (± S.D.) at each timepoint. A slightly lower (significantly different, p=0.0236, Student’s t-test) percent of spreaded cells in the presence of Yoda1 after 20 min from the start of the experiment whereas no significant differences between experimental conditions were detected at later time points (40, 60 and 80 min). Successful spreading of the cells indicates their viability. B: The effect of Yoda1 on SK-MEL-2 morphology. The cell area, circularity and aspect ratio of melanoma cells after 80 min and 24 hours from the beginning of the experiment were calculated. A small, but significant (**p<0.01, Mann-Whitney test) decrease (by 15-17%, compared to control) of cell area in the presence of Yoda1, whereas cell circularity and aspect ratio were not significantly different (Student’s t-test and Mann-Whitney test, respectively). Shown are medians and 95% confidence intervals for cell area and aspect ratio, and mean ± S.D. for cell circularity. ns - the differences are non-significant.
Figure 3.
The effect of Yoda1 on SK-MEL-2 spreading and morphology. A: Microphotographs showing the subsequent steps of SK-MEL-2 spreading. Round cells were considered as non-spreaded (left box), cells at center and right boxes were considered as spreaded. Shown are percentages of spreaded cells (± S.D.) at each timepoint. A slightly lower (significantly different, p=0.0236, Student’s t-test) percent of spreaded cells in the presence of Yoda1 after 20 min from the start of the experiment whereas no significant differences between experimental conditions were detected at later time points (40, 60 and 80 min). Successful spreading of the cells indicates their viability. B: The effect of Yoda1 on SK-MEL-2 morphology. The cell area, circularity and aspect ratio of melanoma cells after 80 min and 24 hours from the beginning of the experiment were calculated. A small, but significant (**p<0.01, Mann-Whitney test) decrease (by 15-17%, compared to control) of cell area in the presence of Yoda1, whereas cell circularity and aspect ratio were not significantly different (Student’s t-test and Mann-Whitney test, respectively). Shown are medians and 95% confidence intervals for cell area and aspect ratio, and mean ± S.D. for cell circularity. ns - the differences are non-significant.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



