
Submitted:
28 August 2023
Posted:
30 August 2023
You are already at the latest version
Abstract
Currently, multiple studies have indicated that CD8+ T lymphocytes play a role in causing damage to the exocrine glands through acinar injury in Primary Sjogren’s syndrome (pSS). The aim of this research was to assess the imbalance of subsets within CD8+ cells in peripheral blood. We examined blood samples from 34 patients with pSS and 34 healthy individuals as controls. We used flow cytometry to enumerate CD8+ T cell maturation stages using as markers CD62L, CD28, CD27, CD4, CD8, CD3, CD45RA, and CD45. For immunophenotyping of ‘polarized’ CD8+ T cell subsets we used the following monoclonal antibodies:CXCR5, CCR6,CXCR3 and CCR4. The findings revealed that both the relative and absolute numbers of 'naïve' CD8+ T cells were higher in pSS patients compared to the healthy volunteers. Conversely, the proportions of effector memory CD8+ T cells were notably lower. Furthermore, our data suggested that among patients with pSS, the levels of cytotoxic Tc1 CD8+ T cells were reduced, while the frequencies of regulatory cytokine-producing Tc2 and Tc17 CD8+ T cells were significantly elevated. Simultaneously, the Tc1 cell subsets displayed a negative correlation with immunoglobulin G, Rheumatoid factor, the Schirmer test, and unstimulated saliva flow. On the other hand, the Tc2 cell subsets exhibited a positive correlation with these parameters. In summary, our study indicated that immune dysfunction within CD8+ T cells, including alterations in Tc1 cells, plays a significant role in the development of pSS.
Keywords:
Sjögren's syndrome
; T cell subsets
; СD8+ T Cells
1. Introduction
Primary Sjogren’s syndrome (pSS) is an autoimmune epithelitis, which is characterized by a lymphocytic infiltration of the lacrimal and salivary glands, lead to the loss of appropriate tear and saliva production. The clinical hallmarks of pSS are keratoconjunctivitis sicca, xerostomia and parotid gland swelling [1]. Extra-glandular manifestations of pSS develop in one-third of patients, leading to immune complex deposition and chronic inflammation in various organs such as skin, joints, lungs, liver, kidneys, blood vessels and even nervous system. Glandular lesions in patients with pSS are characterized by the massive infiltration of inflammatory cells, which include CD3+ T cells, CD19+ B cells, natural killer cells, dendritic cells and macrophages [2]. In 18-59% of patients, lymphocytic infiltration of the lacrimal and salivary glands may become alike secondary lymphoid organs, forming ectopic germinal centers. Extensive inflammation of the glands leads to progressive acinar epithelial cell atrophy and fibrosis [3].
Previously, there was an idea of predominance of activated CD4+helper T cells (Th) in the infiltrates, and the role of CD8+ cytotoxic T cells (Tcyt) in disease pathogenesis has been unexplored [4]. Currently, the role of Tcyt is strongly revised. CD8+ T cells are a complex group of lymphocytes, which are known to play a critical role in particular in autoimmune disease. CD8+ T cells provide their effector function in three ways: by releasing granules filled with cytotoxic molecules, through Fas-FasL interactions, and by producing cytokines. Overactivity or abnormal proliferation of CD8+ T lymphocytes can be detected in the peripheral circulation and target tissues of pSS patients [5].
A pathogenic role of CD8+ T cells in lacrimal gland autoimmunity was shown in fundamental work with Nonobese diabetic (NOD) mouse, which is a well-characterized model of pSS. Transfer of purified CD8 T cells isolated from the cervical lymph nodes of NOD mice into NOD-SCID (Severe combined immunodeficiency) recipients resulted in inflammation of the lacrimal glands. This data demonstrate a pathogenic role of CD8 T cells in lacrimal gland autoimmunity [6].
In the study Mingueneau M et al., patients with pSS had increased CD8+ HLA-DR+ cells in peripheral blood. Likewise, salivary gland biopsy revealed abundant infiltration of CD8+ T-lymphocytes, which mostly had the CD8+HLA-DR+ phenotype. HLA-DR+ CD8+ T cells in salivary gland biopsy and peripheral blood positively correlated with the activity of the disease, showing their important role in pSS [7].
Fujihara T et al. showed that CD8+but not CD4+T lymphocytes were located around the acinar epithelial cells and the ascensive effector molecules of cytotoxic CD8+ T lymphocytes such as granzyme B, perforin, interferon-g, and tumor necrosis factor-a can be detected. These results indicate the presence of a population of activated CD8+ T lymphocytes with cytotoxicity that are competent to lead the death or apoptosis of the cells [8].
In the study Kaneko N et. al., granzyme-A positive CD8+ T cells were the most prominent T cells in the labial salivary glands from untreated patients with primary Sjögren’s syndrome. It was noted, that acinar cells and ductal cells undergo apoptosis, and CD8+ T cells contribute to apoptotic cell death, using both granule exocytosis and Fas signaling to destroy target cells [9].
However, Zhang X et al. investigated the regulatory function of CD8+ cells in pSS pathogenesis, using murine desiccating stress model that resembles Sjögren syndrome [10].
Here we report a study of CD8+ cells peripheral blood subsets of patients with pSS. We used flow cytometry to immunophenotype peripheral blood from 34 patients with pSS and 34 control subjects. Then we analyzed the relationship between CD8+ T cells subsets, clinical manifestations of the disease and disease activity.
2. Materials and Methods
All patients with pSS (n=34) met the 2016 ACR-EULAR Classification Criteria for pSS [11]. Control subjects (n=34) were either healthy volunteers with no signs of autoimmune disease. The groups were comparable in gender and age. ESSDAI (EULAR Sjögren’s Syndrome Disease Activity Index) and ESSPRI (EULAR Sjögren’s Syndrome Patient Reported Index) were calculated to assess the activity of the disease [12,13]. The Schirmer test was performed without the use of local anesthetics using special test strips (Contacare, India). Unstimulated sialometry was measured according to generally accepted recommendations [14]. Minor labial salivary glands biopsies were obtained from all pSS patients to confirm the diagnosis. Clinical characteristics of pSS patients in this study presented in the Table 1.

Table 1.
Clinical characteristics of pSS patients in this study.
| Sjogren’s Syndrome (n = 34) | |
|---|---|
| Male/female ratio | 2/32 |
| Age, years, median (IQR) | 46 [41; 55] |
| Disease duration (years), median (IQR) | 8 [2,5; 11] |
| ESSDAI, median (IQR) | 5 [2; 9] |
| ESSPRI, median (IQR) | 6,7 [5,6; 7,3] |
| Clinical manifestations / ESSDAI domain (percentage) | |
| Constitutional domain | 5 (14,7) |
| Lymphadenopathy and lymphoma domain | 6 (17,7) |
| Glandular domain | 5 (14,7) |
| Articular domain | 17 (50) |
| Cutaneous domain | 5 (14,7) |
| Pulmonary domain | 2 (5,8) |
| Renal domain | 0 |
| Muscular domain | 2 (5,8) |
| PNS domain | 4 (11,6) |
| Central nervous system domain | 0 |
| Haematological domain | 14 (41,2) |
| Biological domainSchirmer test, mm/5 min., median (IQR)Unstimulated sialometry, ml/15 min., median (IQR) | 3 (9,7)3 [1; 9]1,5 [1;3] |
| Laboratory manifestations (percentage) | |
| Complement C3 levels below the lower limit of normal value | 6 (17,6) |
| Complement C4 levels below the lower limit of normal value | 3 (8,7) |
| Anti-DNA Ab | 4 (11,8) |
| Rheumathoid factor | 10 (29,4) |
| Ro-52 Ab | 20 (58,8) |
| SSA Ab | 25 (73,5) |
| SSB Ab | 10 (29,4) |
| CENT B Ab | 2 (5,8) |
| Medications | 28 (82,4) |
| Prednisone dose, mg/day, median (IQR) | 3,75 [0,0; 10,0] |
| Hydroxychloroquine (percentage) | 23 (67,7) |
| Azathioprine (percentage) | 1 (2,9) |
| Cyclosporine (percentage) | 1 (2,9) |
| Mycophenolate mofetil (percentage) | 1 (2,9) |
| Leflunomide (percentage) | 2 (5,8) |
| Methotrexate (percentage) | 5 (14,7) |
All participants provided signed informed consent. All samples were obtained for scientific research according to the Helsinki Declaration and approved by the local ethics committee (protocol no 03-20 16.03.2020).
2.2. Laboratory and Pathological Determination
The level of total Rheumathoid factor (RF), immunoglobulin G (IgG), C-reactive protein (CRP), C3, C4 complement was investigated using an automatic Abbott analyzer. Erythrocyte sedimentation rate was investigated by Westergren method.
The method of indirect immunofluorescence on a commercial substrate (Euroimmun AG, Germany) was used to detect the antinuclear factor (ANF). The line-blot method with commercial reagent kits (EuroimmunAG, Germany) was used to identify the spectrum of antinuclear autoantibodies, suchas double-stranded DNA (Anti-DNA Ab), anti SSAantibodies (SSA Ab), anti- SS-Bantibodies (SSB Ab), anti- Ro-52antibodies (Ro-52 Ab), Anti-centromere B antibodies (CENP B).
2.3. Sample Collection
Peripheral blood samples from the patients were collected before treatment initiation. Five milliliters of peripheral blood were collected from each patient in K3EDTA anticoagulant tubes. Collected peripheral blood samples were processed immediately. CD8+ T cell subsets immunophenotyping was performed within a several hours (less that 6 h) after blood collection.For cytokines measurement, cell-free plasma samples were obtained after whole blood centrifugation at 300× g for 7 min at + 4 °C, followed by placing it into pure 1.5 mL tubes and subsequent centrifugation at 300× g for 15 min at + 4 °C again to sediment residual platelets and other blood cells. Finally, each plasma sample was aliquoted and stored at −80 °C until use.
2.4. Immunophenotyping of Peripheral Blood CD8+ T Cell Subset Maturation Stages and ‘Polarized’ CD8+ T Cell Subsets
For immunophenotyping of CD8+ T cell subset maturation stages, 200 μL of whole peripheral blood samples were stained with the following monoclonal antibodies: ECD-labeled mouse anti-human CD62L, PC5.5-labeled mouse anti-humanCD28, PC7-labeled mouse anti-human CD27, APC-labeled mouse anti-humanCD4, APC-AF700-labeled mouse anti-human CD8, APC-AF750-labeled mouse anti-human CD3, Pacific Blue-labeled mouse anti-human CD45RA, and Krome Orange-labeled mouse anti-human CD45 (all antibodies were manufactured by Beckman Coulter, Indianapolis, IN, USA, and were used according to the manufacturer’s recommendations). Next, all samples were incubated at room temperature in the dark for 15min followed by red blood cell lysis for 15min in the dark with 2 mL of the VersaLyse Lysing Solution (Beckman Coulter, Inc., USA) supplied with 50 μL of the IOTest 3 Fixative Solution (Beckman Coulter, Inc., USA). Finally, 200 μL of Flow-Count Fluorospheres (Beckman Coulter, Indianapolis, IN, USA) were added, and sample acquisition was performed using a 3/10 Navios flow cytometer (Beckman Coulter, Indianapolis, IN, USA). At least 20,000 CD8+ T cells were analyzed in each sample.The gating strategy for CD8+ T cell subset maturation stages was described previously in details [15].
For immunophenotyping of ‘polarized’ CD8+ T cell subsets 200 μL of whole peripheral blood samples were stained with the following monoclonal antibodies:FITC-labeled mouse anti-human CD45RA (Beckman Coulter, Indianapolis, IN, USA), PE-labelled mouse anti-human CD62L (Beckman Coulter, Indianapolis, IN, USA), PerCP/Cy5.5-labeled mouse anti-human CXCR5 (CD185, BioLegend, Inc., San Diego, CA, USA), PE/Cy7-labeled mouse anti-human CCR6 (BioLegend, Inc., San Diego, CA, USA), APC-labeled mouse anti-human CXCR3 (CD183, BioLegend, Inc., San Diego, CA, USA), APC-Alexa Fluor 750-labeled mouse anti-human CD3 (Beckman Coulter, Indianapolis, IN, USA), Pacific-Blue-labeled mouse anti-human CD8 (BioLegend, Inc., San Diego, CA, USA), and Brilliant Violet 510-labeled mouse anti-human CCR4 (CD194, BioLegend, Inc., San Diego, CA USA). All antibodies were used according to the manufacturer’s recommendations). Blood samples were stained with antibodies at room temperature for 15 min in the dark. Then, erythrocytes were lysed by adding 1 mL of VersaLyse Lysing Solution (Beckman Coulter, Inc., Indianapolis, IN, USA) with 25 µL of IOTest 3 Fixative Solution (Beckman Coulter, Inc., Indianapolis, IN, USA) in the dark at room temperature for 15 min. Next, all samples were washed (330× g for 8 min) twice with sterile PBS supplemented with 2% of fetal calf serum (FCS) (Sigma-Aldrich Co., Saint Louis, MO, USA), resuspended in 500 µL of fresh PBS with 2% neutral formalin (Sigma-Aldrich Co., Saint Louis, MO, USA), and subjected to a flow cytometry analysis. At least 20,000 CD3+CD8+ Th cells were collected from each sample.The gating strategy for the major ‘polarized’ CD8+ T cell subsets was described previously in details [15].
2.5. Cytokine and Chemokine Measurement
The levels of 47 cytokines/chemokines/growth factors in plasma samples were assessed by multiplex analysis performed on the fluorescently labeled magnetic microsphere beads using the MILLIPLEX® MAP Human Cytokine/Chemokine/Growth Factor Panel A (HCYTA-60K-PX48, MilliporeSigma, Burlington, MA, USA) according to the manufacturer’s instructions on the Luminex MAGPIX® (RUO) Instrument (Luminex, Austin, TX, USA) as it was described earlier in details [16].
2.6. Statistical Analysis
The flow cytometry data were analyzed using Kaluza software v2.3 (Beckman Coulter, Indianapolis, IN, USA). All the multiplex analysis data were generated with xPONENT software and analyzed with Milliplex Analyst 5.1 Flex software(Luminex, Austin, TX, USA).Statistical analysis was performed with Statistica 7.0 (StatSoft, Tulsa, OK, USA) and GraphPad Prism 8 (GraphPad software Inc., San Diego, CA, USA) software packages. Normality was checked using Pearson’s chi-squared test. Flow cytometry data were presented as a percentage of cells, and the absolute number of CD8+ T cell subsets was calculated using Flow-Count Fluorospheres (Beckman Coulter, Indianapolis, IN, USA), that were used to determine absolute counts on the flow cytometer, and were presented as the number of cells per 1 μL of whole peripheral blood. All the multiplex analysis data were presented as acytokines/chemokines/growth factors concentration in pg/ml. All data were presented as the median and interquartile range, Me (Q25;Q75). The inter-group differences were analyzed using a nonparametric Mann–Whitney U-testwith a p < 0.05 value. A correlation analysis was performed using the nonparametric Spearman rank test, and significance was set at p < 0.05.
3. Results
3.1. Main Peripheral Blood CD3+ T Cell Subsets in Patients with Sjögren’s Syndrome
Preliminary, peripheral blood samples from 34 patients with pSS and 34 samples from healthy controls (HC) were analyzed for the main T cell subsets. Our results indicated these two groups had similar percentages and absolute numbers of CD3+cells as it was shown of the Figure 1. We found that the frequency of CD3+ cells in patients with SS was 80.13% (71.13; 83.23), and very similar results were obtained for HC (76.77% (72.58; 80.44) with p = 0.211), while the absolute numbers of CD3+ cells were 1204 cells/1μL (909; 1811) vs. 1083 cells/1μL (967; 1521), respectively, with p = 0.492. Next, we compared the frequencies of CD4+ T cells, and found that in SS group the relative number of Th cell was 46.15% (37.34; 54.63) vs. 47.83% (43.27; 52.42) in healthy controls (p = 0.404). We also found no significant differences in CD4+ T cell concentrations between the groups (650 cells/1μL (475; 1167) vs. 670 cells/1μL (580; 855), respectively, with p = 0.650). Finally, we analyzed the frequencies of CD8+ T cells, and, as it was shown on the Figure 1, no significant differences were detected in relative and absolute numbers between the groups (25.21% (19.86; 30.14) vs. 24.26% (19.26; 28.13) with p = 0.418, and 459 cells/1μL (305; 628) vs. 350 cells/1μL (267; 487) with p = 0.093, respectively).
Next, we consider Spearman correlations between the levels of CD3+, CD4+ and CD8+ T cell and disease parameters of pSS.The relative number of Tcyt had significant direct correlation with IgG level (r = 0.408, p = 0.032).
3.2. Alterations in CD8+ T Cell Maturation Subsets in Patients with Sjögren’s Syndrome
Next, using multicolor flow cytometry, we classified peripheral blood CD8+ T cell subsetsby CD45RA and CD62L antigens co-expression. Our combination ofCD45RA, and CD62L allowed identification of ‘naïve’ CD8+ T cells (co-expressing CD45RA and CD62L), central memory CD8+ T cells (CM, with CD45RA–CD62L+ phenotype), effector memory CD8+ T cells (EM, CD45RA and CD62L double-negative cells), and terminally differentiated CD45RA-positive effector memory CD8+ T cells (TEMRA, with CD45RA+CD62L−phenotype) within total CD8+ T cell subsets [17]. We assessed the percentage and relative numbers of four main CD8+ T cell maturation subsets, and data obtained are summarized in Figure 2. Interestingly, we found that the relative and absolute numbers of ‘naïve’ CD8+ T cell were increased in patients with Sjögren’s syndrome if compared to HC (34.02% (18.86; 42.97) vs. 21.49% (14.94; 28.47) with p = 0.002, and 147 cells/1μL (62; 226) vs. 69 cells/1μL (51; 122) with p = 0.002, respectively). We also noticed that the percentages of EM CD8+ T cells was significantly decreased in patients with SS (19.25% (15.93; 25.52) vs. 27.73% (22.96; 38.21) with p < 0.001), as it was shown on the Figure 2.
Conducting a correlation between peripheral blood maturation CD8+ T cell subsets and disease parameters, we found, that ‘naïve’ Tcyt was inversed correlated with CRP level (r = –0.382, p = 0.034), EM Tcyt was directly correlated with IgG (r = 0.486, p = 0.009) and CM Tcyt was directly correlated with SSA Ab (r = 0.361, p = 0.043).
3.3. Imbalance in EM and TEMRA CD8+ T Cell Subsets Inpatients with Sjögren’s Syndrome
EM and TEMRA had many characteristic of effector cells, were able to migrate to inflamed tissues and could be closely associated with the functions of CD8+ T cells. Preliminary, we investigated the expression of CD27 and CD28 co-stimulatory molecules on the cell surface of CD45RA–CD62L– CD8+ T cell subset, and defined the four distinct effector memory subsets, as it was described previously by Romero et al. [18]. Thus, CD27 and CD28 co-expression resulted in identifying CD27+CD28+EM1 cells, CD27+CD28−EM2 cells, double-negative CD27−CD28− EM3 cells, as well as CD27−CD28+EM4 cell subset (Figure 3). Our data revealed a significantly higher increase in both relative and absolute numbers of EM3 and decreased EM1 CD8+ T cells in patients with Sjögren’s syndrome compared to healthy controls (45.19% (34.91; 61.30) vs. 18.43% (10.96; 31.24) with p < 0.001 and 39 cells/1μL (19; 56) vs. 16 cells/1μL (10; 38) with p = 0.009 for EM3 subsets, respectively, and 30.86% (20.04; 45.41) vs. 61.67% (51.19; 67.97) with p < 0.001 and 24 cells/1μL (16; 39) vs. 57 cells/1μL (40; 91) with p < 0.001 for EM1 cells, respectively).
Finally, based on CD27 and CD28 expression the TEMRA CD8+ T cells were subdivide into pre-effector type 1 cells (pE1, with CD27+CD28+ phenotype), pre-effector type 2 cells (pE2, with CD27+CD28– phenotype), and effector cells (Eff, with CD27−CD28− phenotype), as it was suggested by Rufer et al. [19] and Koch et al. [20].We found that the relative and absolute numbers of two immature TEMRA CD8+ T cell subsets – pE1 nad pE2 – were significantly decreased in peripheral blood samples from patients with Sjögren’s syndrome if compared to healthy controls (5.13% (2.39; 11.27) vs. 16.88% (8.36; 23.74) and 6 cells/1μL (3; 12) vs. 18 cells/1μL (10; 32) with p < 0.001 in both cases for pE1 cells, and 9.91% (5.39; 18.07) vs. 18.19% (12.61; 22.76) with p = 0.001 and 10 cells/1μL (5; 22) vs. 18 cells/1μL (13; 28) with p = 0.004 for pE2 cells, respectively). Furthermore, the frequency of mature CD27–CD28– effector TEMRA CD8+ T cells was increased in patients with Sjögren’s syndrome vs. healthy controls (81.52% (72.83; 87.68) vs. 64.42% (52.78; 77.55), p < 0.001). The results was shown on the Figure 4.
Thereafter, we assessed the correlations of the levels of maturation TEMRA CD8+ T cell subsets and disease parameters of pSS. There were significant positive correlations for relative number of EM3 cells and IgG, SSA and SSB Ab levels (r = 0.491, p = 0.008, r = 0.350, p = 0.049, and r = 0.425, p = 0.015, respectively). Also, EM2 CD8+ T cells frequency was directly correlated with erythrocyte sedimentation rate (r = 0.477, p = 0.016), while EM1 cells level was inversely correlated with CRP (r = –0.564, p < 0.001). As well, there were significant negative correlations for relative pE1 Tc and CRP (r = –0.396, p = 0.028).
Thus, our data suggest that EM and TEMRA CD8+ T cell subsets were enriched with most mature cells with effector functions, as well as effector CD8+ T cells may function in the pathogenesis of pSS.
3.4. Imbalance in Peripheral Blood CD8+ T cells ‘Polarization’ in Patients with Sjögren’s Syndrome
To assess relevant ‘polarized’ CD8+ T cell subsets, we studied CXCR3 and CCR6 co-expression, as it was proposed earlier [21,22]. Thus, we identified for major CD8+ T cell subsets – Tc1 (CCR6–CXCR3+), Tc2 (CCR6–CXCR3–), Tc17 (CCR6+CXCR3–), and double-positive Tc17.1 (CCR6+CXCR3+) – within total CD8+ T cell subsets (Figure 5). We noticed that the relative numbers of CXCR3-expressing CD8+ T cells – Tc1 and Tc17.1 – were decreased in patients with Sjögren’s syndrome (59.66% (50.66; 67.47) vs. 71.38% (64.85; 77.76) with p , 0.001 and 2.56% (1.58; 3.39) vs. 3.99% (2.71; 7.18) with p = 0.003, respectively), while the frequencies of Tc2 and Tc17 CD8+ T cell were elevated in patients with Sjögren’s syndrome if compared to healthy controls (30.53% (25.49; 41.10) vs. 19.26% (25.49; 41.10) with p < 0.001 and 1.42% (0.84; 2.86) vs. 0.82% (0.56; 1.50) with p = 0.013, respectively). Similarly, the absolute numbers of CD8+ T cells Tc2 and Tc17 CD8+ T cell were elevated in patients with Sjögren’s syndrome vs. healthy control group (149 cells/1μL (76; 218) vs. 71 cells/1μL (59; 113) with p , 0.001 and 7 cells/1μL (4; 11) vs. 3 cells/1μL (2; 6) with p = 0.006), while the levels of Tc17.1 was decreased (11 cells/1μL (7; 18) vs 14 cells/1μL (11; 23) with p = 0.036).
Thus, our data indicated that in circulating blood from patients with Sjögren’s syndrome the level of cytotoxic Tc1 CD8+ T cells that were able to kill infected cells and to secrete effector cytokines (IFNg and TNFa) was decreased while the frequencies of regulatory cytokine-producing Tc2 and Tc17 CD8+ T cells were significantly elevated.
To further explore the relationship between the peripheral blood CD8+ T cells ‘polarization’ and disease activity, correlation with the disease activity index and markers of disease activity were tested. There were significant negative correlations for relative number of Tc1 and IgG level and level of RF (r = –0.398, p = 0.036 and r = –0.608, p=0.040, respectively). On the contrary, relative number of Tc2 was directly correlated with RF (r = 0.634, p = 0.030). Moreover, ‘naïve’ Tc1 cells, EM Tc1 and TEMRA Tcyt1 were inversely correlated with RF (r = –0.504, p=0.023, r = –0.613, p=0.004 and r = –0.753, p < 0.001, respectively), while ‘naïve’ Tcyt2, EM Tcyt2 and TEMRA Tcyt2 cells were directly correlated with it (r = 0.512, p = 0.021, r = 0.717, p < 0.001 and r = 0.771, p < 0.001, respectively). In addition, IgG level was inversely correlated with EM Tc1 and TEMRA Tc1 (r = –0.396, p = 0.037 and r = -0.463, p = 0.013, respectively), while it wasdirectly correlated with EM Tcyt2 and TEMRA Tcyt2 (r = 0.393, p = 0,038 and r = 0.459, p = 0.014, respectively).
Next, Tc17 and Tc17.1 were inversely correlated with the CRP level (r = -0,430, p = 0,016 and r = –0.375, p = 0.038, respectively) and SSB antibody (r = –0.482, p = 0.005 and r = –0.621, p < 0.001, respectively). Also, CM Tcyt17.1, EM Tc17, EM Tc17.1 and TEMRA Tc17 were inversely correlated with the CRP level (r = –0.440, p = 0.013, r = –0.431, p = 0.015, r = –0.369, p = 0.004 and r = –0.364, p = 0,044, respectively). SSB Ab was inversely correlated with ‘naïve’ Tc17, CM Tc17.1, EM Tc17, EM Tc17.1, TEMRA Tc17. TEMRA T17.1 (r = –0.408, p = 0.020; r = –0.495, p = 0.004; r = –0.350, p = 0.049; r = –0.492, p = 0.004; r = –0.406, p = 0.020 and r = –0.525, p = 0.002, respectively). SSA Ab was inversely correlated with CM Tc17.1 (r = –0.396, p = 0.025). Anti-DNA Abs level was directly correlated with naïve Tc17.1 and CM Tc17 (r = 0.599, p = 0.007 and r = 0.633, p = 0.007, respectively), while inversely correlated with CM Tc1 and EM Tc1 (r = –0.499, p=0.029 and r = –0.468, p = 0.043, respectively).
Given the correlation between CD8+T cell subsets and markers of disease activity, we asked if the former were linked to organ involvement. Loss of appropriate tear and saliva production are the clinical hallmarks of pSS. We found significant negative correlation for relative number of Tc1, Tc1 naive, Tc1 EM, Tc1 TEMRA and Shirmer test (r = –0.483, p = 0.02; r = –0.54, p = 0.008; r = –0.501, p = 0.015 and r = –0.518, p = 0.011). Also, there were significant positive correlation of Tc2, Tc2 naïve , Tc2 EM and Shirmer test (r = 0.484, p = 0.02; r = 0.509, p = 0.013 and r = 0.468, p = 0.024 ). Moreover, unstimulated saliva production flow was inversely correlated with relative numbers of Tc1, Tc1 naïve, Tc1 CM, Tc1 EM, EM2 Tc, EM3 Tc and E Tcyt(r = –0.709, p = 0.0003; r = –0.597, p = 0.004; r = –0.718, p = 0.0002 ; r = –0.774, p = 0.00004; r = –0.543, p = 0.011; r = –0.457, p = 0.037 and r = –0.546, p = 0.01), while it was directly correlated with Tc17, Tc17.1, Tc2 naïve, Tc2 CM, Tc17 CM, Tc17 EM, Tc17 TEMRA, Tc17.1 TEMRA and pE1 Tcyt (r = 0.598, p = 0.004; r = 0.448, p = 0.042; r = 0.605, p = 0.004; r = 0.531, p = 0.013; r = 0.653, p = 0.001; r = 0.652, p = 0.001; r = 0.490, p = 0.024; r = 0.495, p = 0.022 and r = 0.535, p = 0.012).
As for other domenes, relative amount of CD3+ T cells was inversely correlated with lung damage in the form of fibrosing alveolitis (R = -0,583, p≤0.05). Moreover, relative value of Tc1 was directly correlated with it, while relative value of Tc2 and Tc TEMRA had inversed correlations (R = 0.648, p≤0.05, R = -0.648, p≤0.05 and R = -0.583, p≤0.05, respectively). There were significant negative correlation EM2 Tc and Tc1 CM with peripheral nervous system involvement (R = -0,692, p≤0.05 and R = -0,780, p≤0.05, respectively). Also, pE2 Tc was inversely correlated with joint damage (R = -0,534, p≤0.05).
3.5. Blood Level of ‘Polarized’ CD8+ T Cell Subsets and Correlations with Cytokines and Chemokines
We determined the correlations of peripheral blood ‘polarized’ CD8+ T cell subsets with main serum chemokines and cytokines. Therefore, we used multiplex analysis to measure the serum levels of 47 cytokines/chemokines/growth factors in plasma samples 20 patients with pSS.
Preliminary, we analyzed possible correlations between IP-10 (CXCL10), MCP-1 (CCL2), MCP-3 (CCL7), MDC (CCL22), MIG (CXCL9), MIP-1a (CCL3), MIP-1b (CCL4), IL-8 (CXCL8), eotaxin (CCL11), fractalkine (CX3CL1) and main ‘polarized’ CD8+ T cell subsets (Figure 6). Wenoticedthatserum levels of two CXCR3 ligands – CXCRL9 andCXCRL10 – negatively correlated with total Tc17.1 and ‘naïve’ Tc17.1 frequencies. Similarly, the negative correlations were detected between total and effector memory Tc17, as well as total, central memory, effector memory and TEMRA Tc17.1 cells and serum levels of CCL2, that acts during physiological immune defense and chronic inflammationto activate the migrationmyeloid and lymphoid cells [23]. Next, the frequencies circulating of Tc1 CD8+ T cell subsets of different maturation state negatively correlated with serum levels of CCL7 andCCL3, that play an important part in inflammatory events by attracting macrophages and monocytes to further amplify inflammatory processes [24], and in regulating lymph node homing of dendritic cell subsets, as well as induces antigen-specific CD4+ and CD8+ T cell responses [25]. Furthermore, CCL3 serum levels positively correlated with the increased frequencies of circulating ‘naïve’ Tc2 cells, as well as effector memory Тс2, Тс17 andТс17.1 CD8+ T cell subsets, that were unregulated in patients with pSS, as it was shown previously.
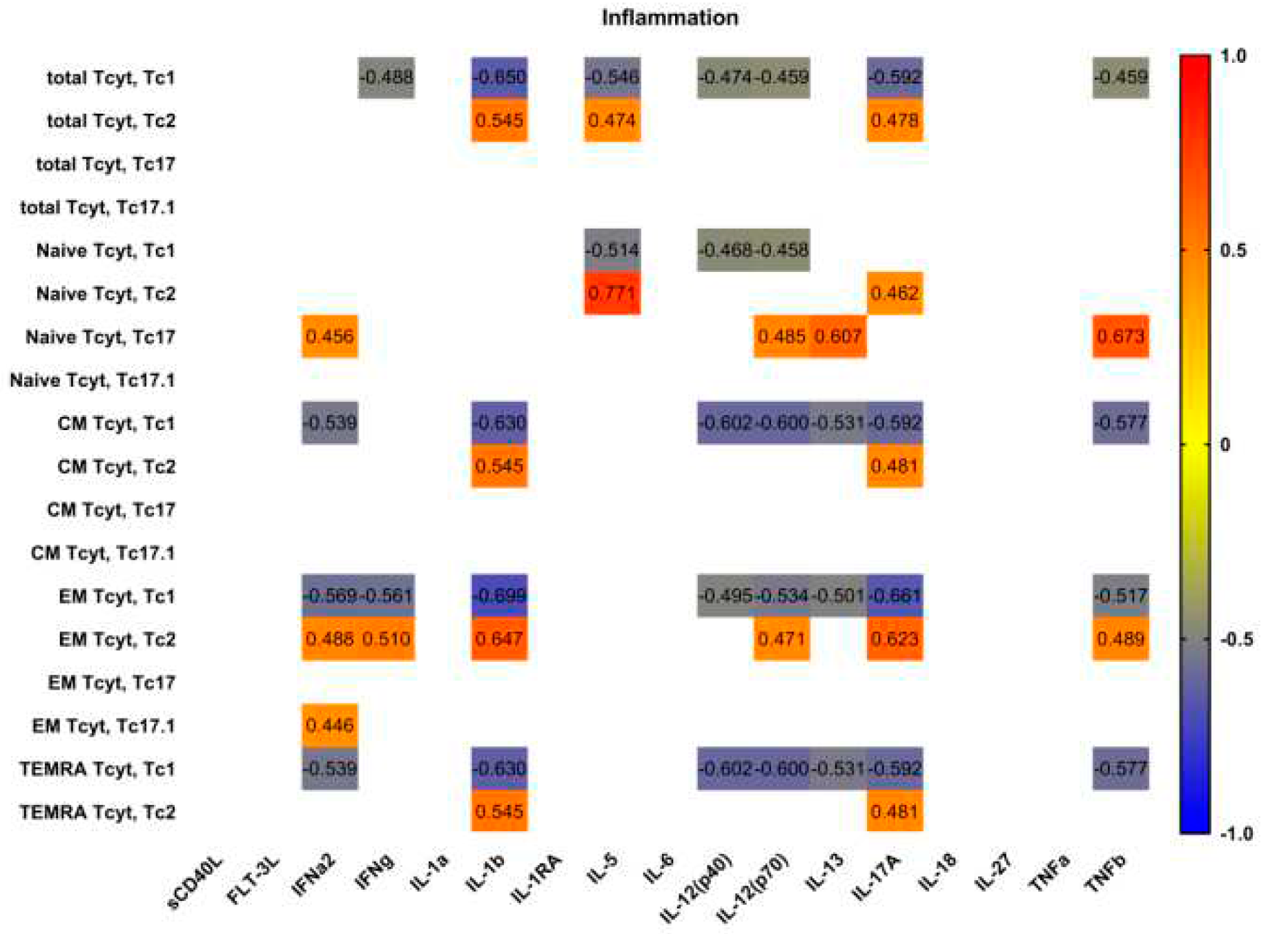
Next, we analyzed possible correlations between main proinflammatory and effector cytokines and ‘polarized’ CD8+ T cell subsets in patients with pSS (Figure 7). Interestingly, we found no significant correlations between serum levels of main proinflammatory cytokines – IL-1a, IL-6, IL-18, and TNFa – and ‘polarized’ CD8+ T cell subsets in patients with pSS. However, we noticed positive corretation between IL-1b and several Tc2 subsets (including total Tc2 cells and CM, EM and TEMRA Tc2 cells); oppositely, we found negative correlations between IL-1b levels and Tc1 subsets frequencies, including total Tc1 cells and CM, EM and TEMRA Tc1 cells. Similarly, ‘non naïve’ Tc1 CD8+ T cells negatively correlated with such cytokines as IL-12p70, IL-13 andIL-17A serum levels, that regulate type 2 and type 3 inflammation [26].
4. Discussion
In our studies described here, we focused on maturation and ‘polarization’ subsets of peripheral blood CD8+ T cell in patients with pSS. Primarily, we found no differences in related and absolute numbers of CD3+CD8+ cell between pSS group and healthy controls. Several research groups also noticed that patients with pSS and controls groups had no differences in CD8+ T cell frequency [27,28] or absolute count [29]. However, Sudzius et al. showed that the absolute counts of CD8+ T cells were significantly lower in pSS patients in comparison to controls [30]. Oppositely, Li et al. noticed that pSS patients had an increased level of circulating peripheral blood CD8+ T cells thatpositively correlated with multiple disease parameters as well as the serum levels of main effector cytokine IFN-γ [31]. Furthermore, these CD8+ T cell were highly activated and expressed elevated levels of CD38 and HLA-DR on their cell membranes.
Next, we found that the relative and absolute numders of ‘naïve’ CD8+ T cell were increased in patients with pSS, pointing to impaired differentiation and maturation of CD8+ T cells in the thymus. Interestingly, in patients with pSS thymus enlargement was identified in approximately 22% of cases [32]. Interestingly, several autoimmune diseases such as systemic lupus erythematosus, myasthenia gravis and rheumatoid arthritis are usually accompanied by thymic hyperplasia, and there are several reports on thymic hyperplasia that could be combined with pSS [33,34,35]. Furthermore, thymectomy in patients with pSS did not appear to improve the clinical symptoms or to decrease the serum level of anti-SSA/Ro antigens in the patient, described by Minato et al. [36]. Similarly, Izumi et al. also reported that after thymectomy in two patients with pSS the serum ANA levels remained increased vs. control levels [37].
We also noticed the decreased frequency of circulating effector memory CD8+ T cell in patients with pSS. Similarly, Narkeviciute et al. also showed that effector memory CD27+CD57− CD8+ T cells were significantly reduced in the peripheral blood of pSS patients [38]. According to some studies, EM CD8+ T cells were able to migrate to inflamed sites located in peripheral tissues during the effector phase of immune response and look part in pathogens clearance [39]. Furthermore, we found that EM CD8+ T cells were enriched with potentially highly cytotoxic EM3 cells, while EM1 cells were decreased. It is known, that EM1 (CD27+CD28+) and EM4 (CD27-CD28+) T cells express low levels of effector mediators such as granzyme B and perforin and high levels of CD127 [15], are closely related to ‘non-effector’ CM CD8+ cells. Oppositely, EM2 (CD27+CD28-) and EM3 (CD27-CD28-) CD8+ T cells express numerous markers of mature effector cells, whereby EM3 cells display stronger ex vivo cytolytic activity and effector cytokine production, resembling TEMRA CD8+ cells. Thus, EM CD8+ T cell subset in patients with pSS was enriched with effector cell that had high cytolytic activity and were more efficient in migrating to inflamed peripheral tissues than other EM subsets.
Next, we also found that most mature CD8+ T cells – TEMRA cells – were also altered. We have shown that the frequencies of pre-effector type 1 and pre-effector type 2 cells were decreased, while the levels of CD27+CD28–effector cells were increased in peripheral blood samples from patients with pSS. Prewiously it was shown by Sudzius et al. that the levels of effector CD8+ T cells were increased patients with pSS [30]. Oppositely, Narkeviciute et al. found that the percentages of effector CD27−CD57−/+CD45RA+CD8+ T cells in pSS patients were lower than in healthy subjects [28]. Therefore, the imbalance of EM and TEMRA CD8+ T cells could be closely linked with the efficient generation of the effector cell subset peripheral lymphoid tissues as well as the migration of mature effector cell to the sites of autoimmune inflammation. Moreover, inexperimental mouse model of pSS (nonobese diabetic mouse model) lacrimal gland infiltrating CD8+ Tcel lsshowed an effector cytotoxic phenotype and took part in epithelial cell damage [6] . Our findings also support the idea that CD8+ T cells could be actively involved in the development of pSS.
Finally, we identified four main ‘polarized’ CD8+ T cell in peripheral blood samples from patients with pSS, and we found imbalance in Tc1, Tc2, Tc17 and Tc17.1 cell in our patients if compared to healthy controls. Primarily, we noticed decreased relative level of Tc1 cell and found negative correlations of Tc1 subsets with IgG, RF levels, Shirmer test and unstimulated saliva flow. The reduced amount of Tc in the peripheral blood of patients with pSS can be explained by the migration of effector cells to the foci of autoimmune inflammation. It was shown, that CXCR3+CCR6– Tc1 cell were capable of killing target cells by secretory degranulation of cytotoxic molecules – granzymes and perforin – releasing, as well as effector cytokines secretion – IFN-γ and TNF-α, that accelerated the tissue inflammation [40]. Interestingly, Jing Zhou et al.l showed that administration of a neutralizing anti-TNF-α antibody to female NOD mice during the stage prior to disease onset significantly improved salivary secretion, indicating a remission of clinical symptoms of SS [41]. Нowever, in Randomized Controlled Trial Etanercept was an ineffective therapeutic agent in pSS consistent with the absence of suppression of TNFalpha and other indicators of immune activation in this patient population [42]. Also, increased levels of CXCR3 ligands – CXCL9, CXCL-10 andCXCL-11 – were detected in tears of the patients with pSS compared with those of the patients with non-Sjögren’s dry eye [43]. Furthermore, it was found that Gal1-null mutant mice had high expression of CXCL9 and CXCL10 chemokines in submandibular glands, as well as CXCR3+ CD8+ T cells effectively infiltrated glands tissues if compared with wild-type mice [44]. All the mentioned finding pointing to the important part of CXCR3-expressing CD8+ T cells in progressive damage to target tissues drying pSS, as well as targeting CXCR3+ CD8+ T cells may be a new strategy for disease immunotherapy.
Moreover, Tс2 are known for their production of type 2 cytokines (IL-4, IL-5, and IL-13), in conjunction with diminished production of IFN-γ and low cytolytic activity, while IL-17-producing CD8+ Tc17 cells are a distinct subset of T cells known for the high production of IL-17A [45]. Our data indicated that in circulating blood from patients with pSS the level of cytotoxic Tc1 CD8+ T cells that were able to kill infected cells and to secrete effector cytokines (IFNg and TNFa) was decreased while the frequencies of regulatory cytokine-producing Tc2 and Tc17 CD8+ T cells were significantly elevated. We found positive correlations of Tc2, Tc17, Tc17.1 subsets with IgG, RF levels, Shirmer test and unstimulated saliva flow. Little is known about the cellular characteristics and functional activityof Tc2 and Tc17 CD8+ T cells in autoimmune disorders, but, for instance, in patients with rheumatoid arthritis Tc2 cells were significantly higher than in healthy controls [46]. Similarly, IL-17-producing Tc17 cells are detectible in multiple sclerosis lesions, and patients with early-stage multiple sclerosis had elevated numbers of Tc17 cells in the cerebrospinal fluid than in peripheral blood [47]. Also, X. Zhang showed that in a murine desiccating stress mouse model, that resembles pSS, CD8+ cell-depletion augmented pathogenic Th17 cell generation, and consequently worsened IL-17A-induced disruption of corneal barrier function [10]. However, we were the group that determined these ‘polarized’ CD8+ T cell subsets in patients with Sjogren’s syndrome, and need further exploration. We also assumed that imbalance in ‘polarized’ CD8+ T cellsubsets may be involved in the pathogenesis of Sjogren’s syndromeby regulating immune functions of cytolytic and cytokine-producing subsets. Herein, we show for the first time that a subset of CD8+ Tregs can significantly mitigate.
6. Conclusions
Collectively, our study suggested that CD8+ T cell immune dysfunction plays an important role in the pathogenesis of pSS. Subsets of Tc 1, Tc2 and Tc17 play a role in the salivary and lacrimal gland damage, which is confirmed by links with the degree of keratoconjunctivitis sicca and xerostomia. It is still unclear to what extent CD8+ T cells and their distinct cell subsetscontribute to the pathology and progression of pSS. Furthermore, the involvement of CD8+T cells in disease initiation and progression remains to be unclear.
Author Contributions
Conceptualization: Artem Rubinstein; methodology: Alexey Maslyanskiy; software: Maria Serebriakova, validation: Dmitri Motorin; formal analysis: Evgenii Kuvardin; investigation: Anna Budkova; resources: Alexey Golovkin; data curation: Olga Kalinina; writing—original draft preparation: Stanislava Benevolenskaya; writing—review and editing: Igor Kudryavtsev; visualization: Darina Zammoeva; supervision: IrinaGrigor’yeva, project administration: Ekaterina Zaikova; funding acquisition: Sergey Lapin.
Funding:
This research was financially supported by the Ministry of Science and Higher Education of the Russian Federation (Agreement No. 075-15-2022-301).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the local Ethics Committee of Almazov National Medical Research Centre (protocol No. 03-20 16.03.2020). All patients included in the study provided informed consent.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gary S. Firestein, Ralph C Budd, Sherine E. Gabriel, Iain B. McInnes, James R. O’Dell,Kelley and Firestein’s Textbook of Rheumatology, 10th ed.Elsevier Health Sciences,Philadelphia, USA, 2017,pp. 1221- 1244.
- Nocturne, G. , & Mariette, X. B cells in the pathogenesis of primary Sjögren syndrome. Nature Reviews Rheumatology, 2018, 14, 133–145. [Google Scholar] [PubMed]
- Fisher, B. A. , Brown, R. M. et al. A review of salivary gland histopathology in primary Sjögren’s syndrome with a focus on its potential as a clinical trials. Ann Rheum Dis, 2015, 74, 1645–1650. [Google Scholar] [CrossRef] [PubMed]
- Kassan, S.S.; Moutsopoulos, H.M. Clinical manifestations and early diagnosis of Sjogren syndrome. Arch. Intern. Med. 2004, 164, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Zhou H, Yang J, et al. CD8+ T Lymphocytes: Crucial Players in Sjögren’s Syndrome. Front. Immunol. 2021 11:602823.
- Jennifer, Y. Barr, Xiaofang Wang et al. CD8 T cells contribute to lacrimal gland pathology in the nonobese diabetic mouse model of Sjögren syndrome. Immunol Cell Biol. 2017, 95, 684–694. [Google Scholar]
- Mingueneau M, Boudaoud S, Haskett S, et al. Cytometry by time-of-flight immunophenotyping identifies a blood Sjögren’s signature correlating with disease activity and glandular inflammation. J Allergy Clin Immunol. 2016, 137, 1809–1821. [Google Scholar] [CrossRef]
- Fujihara T, Fujita H, Tsubota K, et al. Preferential localization of CD8+ alpha E beta 7+ T cells around acinar epithelial cells with apoptosis in patients with Sjögren’s syndrome. J Immunol. 1999, 163, 2226–2235. [Google Scholar] [CrossRef]
- Kaneko N, Chen H, Perugino CA, et al. Cytotoxic CD8+ T cells may be drivers of tissue destruction in Sjögren’s syndrome. Sci Rep. 2022, 12, 15427. [Google Scholar] [CrossRef]
- Zhang X, Schaumburg CS, Coursey TG, Siemasko KF, Volpe EA, Gandhi NB, et al. CD8(+) cells regulate the T helper-17 response in an experimental murine model of Sjogren syndrome. Mucosal Immunol 2014, 7, 417–27. [Google Scholar] [CrossRef]
- Caroline, H. Shiboski, Stephen C. Shiboski, Raphaèle Seror, et al. ACR-EULAR Classification Criteria for primary Sjögren’s Syndrome: A Consensus and Data-Driven Methodology Involving Three International Patient Cohorts. Arthritis Rheumatol. 2017, 69, 35–45. [Google Scholar]
- : Seror R, Bowman SJ, Brito-Zeron P, et al. EULAR Sjögren’s syndrome disease activity index (ESSDAI): a user guide. RMD Open 2015, 1, e000022. [Google Scholar] [CrossRef]
- Seror R, Ravaud P, Mariette X, et al. EULAR Sjögren’s Syndrome Patient Reported Index (ESSPRI): development of a consensus patient index for primary Sjögren’s syndrome. Annals of the Rheumatic Diseases 2011, 70, 968–972. [Google Scholar] [CrossRef] [PubMed]
- Navazesh M, Kumar SKS. Measuring salivary flow. Dent Assist J. 2008, 139, 35S–40S. [Google Scholar]
- Kudryavtsev, I.V.; Arsentieva, N.A.; Korobova, Z.R.; Isakov, D.V.; Rubinstein, A.A.; Batsunov, O.K.; Khamitova, I.V.; Kuznetsova, R.N.; Savin, T.V.; Akisheva, T.V.; et al. Heterogenous CD8+ T Cell Maturation and ‘Polarization’ in Acute and Convalescent COVID-19 Patients. Viruses 2022, 14, 1906. [Google Scholar] [CrossRef] [PubMed]
- Kalinina, O.; Golovkin, A.; Zaikova, E.; Aquino, A.; Bezrukikh, V.; Melnik, O.; Vasilieva, E.; Karonova, T.; Kudryavtsev, I.; Shlyakhto, E. Cytokine Storm Signature in Patients with Moderate and Severe COVID-19. Int. J. Mol. Sci. 2022, 23, 8879. [Google Scholar] [CrossRef]
- Sallusto, F; Lenig, D; Förster, R; Lipp, M; Lanzavecchia, A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature. 1999, 401, 708–712. [Google Scholar] [CrossRef]
- Romero, P.; Zippelius, A.; Kurth, I.; Pittet, M.J.; Touvrey, C.; Iancu, E.M.; Corthesy, P.; Devevre, E.; Speiser, D.E.; Rufer, N. Four functionally distinct populations of human effector-memory CD8+ T lymphocytes. J. Immunol. 2007, 178, 4112–4119. [Google Scholar] [CrossRef]
- Rufer N, Zippelius A, Batard P, Pittet MJ, Kurth I, Corthesy P, Cerottini JC, Leyvraz S, Roosnek E, Nabholz M, Romero P. Ex vivo characterization of human CD8+ T subsets with distinct replicative history and partial effector functions. Blood. 2003, 102, 1779–87. [Google Scholar] [CrossRef]
- Koch S, Larbi A, Derhovanessian E, Ozcelik D, Naumova E, Pawelec G. Multiparameter flow cytometric analysis of CD4 and CD8 T cell subsets in young and old people. Immun Ageing. 2008, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Loyal L, Warth S, Jürchott K, Mölder F, Nikolaou C, Babel N, Nienen M, Durlanik S, Stark R, Kruse B, Frentsch M, Sabat R, Wolk K, Thiel A. SLAMF7 and IL-6R define distinct cytotoxic versus helper memory CD8+ T cells. Nat Commun. 2020, 11, 6357. [Google Scholar] [CrossRef]
- Trombke, J.; Loyal, L.; Braun, J.; Pleyer, U.; Thiel, A.; Pohlmann, D. Analysis of peripheral inflammatory T cell subsets and their effector function in patients with Birdshot Retinochoroiditis. Sci. Rep. 2021, 11, 8604. [Google Scholar] [CrossRef]
- Gschwandtner M, Derler R, Midwood KS. More Than Just Attractive: How CCL2 Influences Myeloid Cell Behavior Beyond Chemotaxis. Front Immunol. 2019, 10, 2759. [Google Scholar] [CrossRef] [PubMed]
- Chang TT, Chen C, Chen JW. CCL7 as a novel inflammatory mediator in cardiovascular disease, diabetes mellitus, and kidney disease. Cardiovasc Diabetol. 2022, 21, 185. [Google Scholar] [CrossRef] [PubMed]
- Schaller TH, Batich KA, Suryadevara CM, Desai R, Sampson JH. Chemokines as adjuvants for immunotherapy: implications for immune activation with CCL3. Expert Rev Clin Immunol. 2017, 13, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Annunziato F, Romagnani C, Romagnani S. The 3 major types of innate and adaptive cell-mediated effector immunity. J Allergy Clin Immunol. 2015, 135, 626–35. [Google Scholar] [CrossRef] [PubMed]
- Martin-Gutierrez L, Peng J, Thompson NL, Robinson GA, Naja M, Peckham H, Wu W, J’bari H, Ahwireng N, Waddington KE, Bradford CM, Varnier G, Gandhi A, Radmore R, Gupta V, Isenberg DA, Jury EC, Ciurtin C. Stratification of Patients With Sjögren’s Syndrome and Patients With Systemic Lupus Erythematosus According to Two Shared Immune Cell Signatures, With Potential Therapeutic Implications. Arthritis Rheumatol. 2021, 73, 1626–1637. [Google Scholar] [CrossRef]
- Narkeviciute I, Sudzius G, Mieliauskaite D, Mackiewicz Z, Butrimiene I, Viliene R, Dumalakiene I. Are cytotoxic effector cells changes in peripheral blood of patients with Sjögren’s syndrome related to persistent virus infection: Suggestions and conundrums. Cell Immunol. 2016, 310, 123–130. [Google Scholar] [CrossRef]
- Mingueneau M, Boudaoud S, Haskett S, Reynolds TL, Nocturne G, Norton E, Zhang X, Constant M, Park D, Wang W, Lazure T, Le Pajolec C, Ergun A, Mariette X. Cytometry by time-of-flight immunophenotyping identifies a blood Sjögren’s signature correlating with disease activity and glandular inflammation. J Allergy Clin Immunol. 2016, 137, 1809–1821. [Google Scholar] [CrossRef]
- Sudzius G, Mieliauskaite D, Siaurys A, Viliene R, Butrimiene I, Characiejus D, Dumalakiene I. Distribution of Peripheral Lymphocyte Populations in Primary Sjögren’s Syndrome Patients. J Immunol Res. 2015, 2015, 854706. [Google Scholar]
- Li H, Zhou Y, Wang P, Wang Y, Feng Y, Zhang Y, Wu Z. Alterations of CD8+ T cells in the blood and salivary glands of patients with primary Sjögren’s syndrome. Clin Rheumatol. 2023 May, 42, 1327-1338. 1327.
- Murata O, Suzuki K, Takeuchi T. Thymus variants on imaging of patients with primary Sjögren’s syndrome and polymyositis/dermatomyositis: clinical and immunological significance. Immunol Med. 2023, 46, 25–31. [Google Scholar] [CrossRef]
- Kobayashi H, Ozeki Y, Aida S. Pulmonary and thymic lymphoid hyperplasia in primary Sjögren’s syndrome. Jpn J Radiol. 2009, 27, 107–10. [Google Scholar] [CrossRef]
- Kondo K, Miyoshi T, Sakiyama S, Shimosato Y, Monden Y. Multilocular thymic cyst associated with Sjögren’s syndrome. Ann Thorac Surg. 2001, 72, 1367–9. [Google Scholar] [CrossRef] [PubMed]
- Xin Y, Cai H, Li Y, Cui Y. Thymic hyperplasia associated with primary Sjogren’s syndrome cured by thymectomy. J Thorac Dis. 2017, 9, E130–E132. [Google Scholar] [CrossRef] [PubMed]
- Minato H, Kinoshita E, Nakada S, Nojima T, Tanaka M, Usuda K, Sagawa M, Iwao H, Tanaka M, Doai M, Takahashi T, Shibata N. Thymic lymphoid hyperplasia with multilocular thymic cysts diagnosed before the Sjögren syndrome diagnosis. Diagn Pathol. 2015, 10, 103. [Google Scholar] [CrossRef] [PubMed]
- Izumi H, Nobukawa B, Takahashi K, Kumasaka T, Miyamoto H, Yamazaki A, Sonobe S, Uekusa T, Suda K. Multilocular thymic cyst associated with follicular hyperplasia: clinicopathologic study of 4 resected cases. Hum Pathol. 2005, 36, 841–4. [Google Scholar] [CrossRef]
- Narkeviciute I, Sudzius G, Mieliauskaite D, Mackiewicz Z, Butrimiene I, Viliene R, Dumalakiene I. Are cytotoxic effector cells changes in peripheral blood of patients with Sjögren’s syndrome related to persistent virus infection: Suggestions and conundrums. Cell Immunol. 2016, 310, 123–130. [Google Scholar] [CrossRef]
- Nizharadze T, Becker NB, Höfer T. Quantitating CD8+ T cell memory development. Trends Immunol. 2023, 44, 519–529. [Google Scholar] [CrossRef]
- Mittrücker HW, Visekruna A, Huber M. Heterogeneity in the differentiation and function of CD8+ T cells. Arch Immunol Ther Exp (Warsz). 2014, 62, 449–58. [Google Scholar] [CrossRef]
- Jing Zhou, Toshihisa Kawai, and Qing Yu. Pathogenic role of endogenous TNF-α in the development of Sjögren’s-like sialadenitis and secretory dysfunction in nonobese diabetic mice. Lab Invest. 2017, 97, 458–467. [Google Scholar] [CrossRef]
- N M Moutsopoulos 1, G E Katsifis, N Angelov, R A Leakan, V Sankar, S Pillemer, S M Wahl. Lack of efficacy of etanercept in Sjögren syndrome correlates with failed suppression of tumour necrosis factor alpha and systemic immune activation. Ann Rheum Dis. 2008, 67, 1437–43.
- Yoon KC, Park CS, You IC, Choi HJ, Lee KH, Im SK, Park HY, Pflugfelder SC. Expression of CXCL9, -10, -11, and CXCR3 in the tear film and ocular surface of patients with dry eye syndrome. Invest Ophthalmol Vis Sci. 2010, 51, 643–50. [Google Scholar] [CrossRef]
- Martínez Allo VC, Hauk V, Sarbia N, Pinto NA, Croci DO, Dalotto-Moreno T, Morales RM, Gatto SG, Manselle Cocco MN, Stupirski JC, Deladoey Á, Maronna E, Marcaida P, Durigan V, Secco A, Mamani M, Dos Santos A, Catalán Pellet A, Pérez Leiros C, Rabinovich GA, Toscano MA. Suppression of age-related salivary gland autoimmunity by glycosylation-dependent galectin-1-driven immune inhibitory circuits. Proc Natl Acad Sci U S A. 2020, 117, 6630–6639. [Google Scholar] [CrossRef] [PubMed]
- Michael, St. Paul1,2 and Pamela S. Ohashi1,2,* The Roles of CD8+ T Cell Subsets in Antitumor Immunity. Trends Cell Biol 2020, 30, 695–704. [Google Scholar]
- Cho BA, Sim JH, Park JA, Kim HW, Yoo WH, Lee SH, Lee DS, Kang JS, Hwang YI, Lee WJ, Kang I, Lee EB, Kim HR. Characterization of effector memory CD8+ T cells in the synovial fluid of rheumatoid arthritis. J Clin Immunol. 2012, 32, 709–20. [Google Scholar] [CrossRef] [PubMed]
- Huber M, Heink S, Pagenstecher A, Reinhard K, Ritter J, Visekruna A, Guralnik A, Bollig N, Jeltsch K, Heinemann C, Wittmann E, Buch T, Prazeres da Costa O, Brüstle A, Brenner D, Mak TW, Mittrücker HW, Tackenberg B, Kamradt T, Lohoff M. IL-17A secretion by CD8+ T cells supports Th17-mediated autoimmune encephalomyelitis. J Clin Invest. 2013, 123, 247–60. [Google Scholar] [CrossRef]
Figure 1.
Comparison of relative numbers and concentrations of main circulating T cell subsets patients with pSS. Scatter plots (A–C) and (D–F) showing the percentages (the percentage of T cell subset within total lymphocyte subset) and absolute numbers (number of cells per 1 μL of whole peripheral blood) of T cells (CD3+), T-helpers (Th, CD3+CD4+), and CD8+ T cells (Tcyt, CD3+CD8+), respectively.Black circles denote patients with Sjögren’s syndrome (SS, n = 34); white circles—healthy control (HC, n = 34). Each dot represents individual subjects, and horizontal bars depict the group medians and quartile ranges (Med (Q25; Q75)). The statistical analysis was performed with the Mann–Whitney U test.
Figure 1.
Comparison of relative numbers and concentrations of main circulating T cell subsets patients with pSS. Scatter plots (A–C) and (D–F) showing the percentages (the percentage of T cell subset within total lymphocyte subset) and absolute numbers (number of cells per 1 μL of whole peripheral blood) of T cells (CD3+), T-helpers (Th, CD3+CD4+), and CD8+ T cells (Tcyt, CD3+CD8+), respectively.Black circles denote patients with Sjögren’s syndrome (SS, n = 34); white circles—healthy control (HC, n = 34). Each dot represents individual subjects, and horizontal bars depict the group medians and quartile ranges (Med (Q25; Q75)). The statistical analysis was performed with the Mann–Whitney U test.
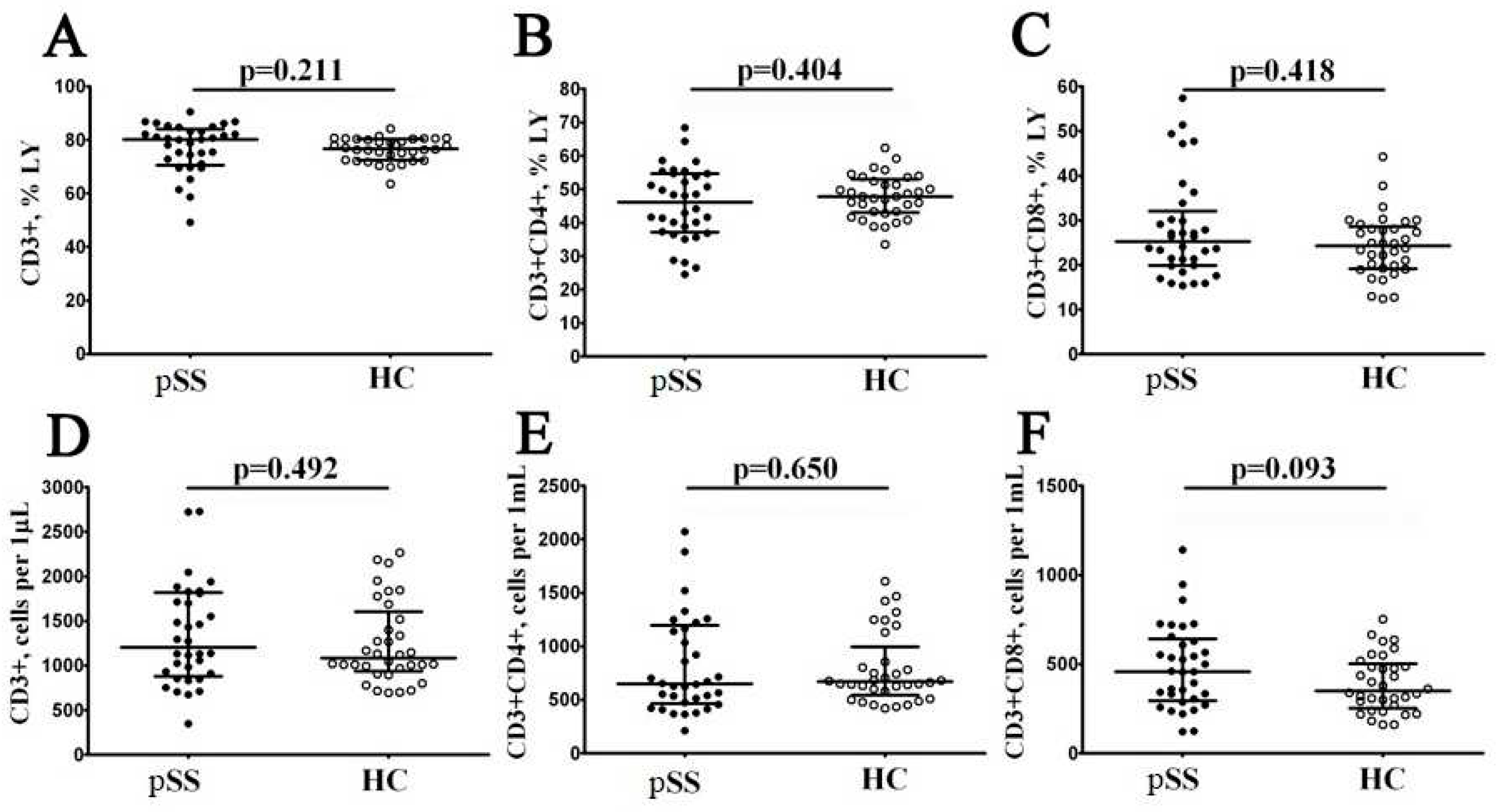
Figure 2.
Alteration in relative and absolute numbers of peripheral blood maturation CD8+ T cell subsets in patients with pSS. Scatter plots (A–D) and (E–H) show the percentages and absolute numbers of ‘naïve’ (CD45RA+CD62L+), central memory (CM, CD45RA−CD62L+), effector memory (EM, CD45RA−CD62L−), and terminally differentiated CD45RA-positive effector memory (TEMRA, CD45RA+CD62L−) CD8+ T cells, respectively. Black circles denote patients with Sjögren’s syndrome (SS, n = 34); white circles—healthy control (HC, n = 34). Each dot represents individual subjects, and horizontal bars depict the group medians and quartile ranges (Med (Q25; Q75)). The statistical analysis was performed with the Mann–Whitney U test.
Figure 2.
Alteration in relative and absolute numbers of peripheral blood maturation CD8+ T cell subsets in patients with pSS. Scatter plots (A–D) and (E–H) show the percentages and absolute numbers of ‘naïve’ (CD45RA+CD62L+), central memory (CM, CD45RA−CD62L+), effector memory (EM, CD45RA−CD62L−), and terminally differentiated CD45RA-positive effector memory (TEMRA, CD45RA+CD62L−) CD8+ T cells, respectively. Black circles denote patients with Sjögren’s syndrome (SS, n = 34); white circles—healthy control (HC, n = 34). Each dot represents individual subjects, and horizontal bars depict the group medians and quartile ranges (Med (Q25; Q75)). The statistical analysis was performed with the Mann–Whitney U test.
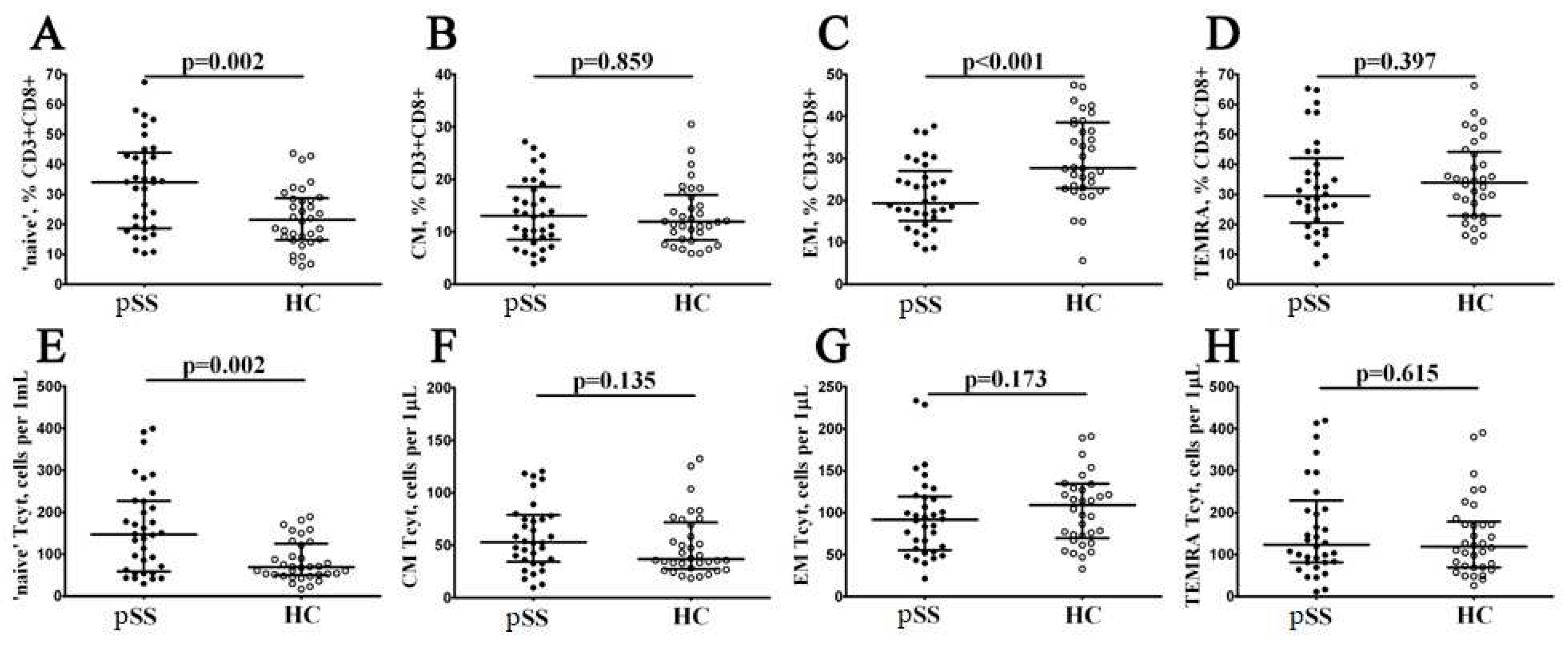
Figure 3.
Alterations in relative and absolute number of EM CD8+ T cell subsets with different patterns of CD27 and CD28 expression in patients with pSS. Scatter plots (A–D) and (E–H) —EM CD8+ T cell were subdivided into EM1 (CD27+CD28+), EM2 (CD27+CD28−), EM3 (CD27−CD28−), and EM4 (CD27−CD28+) subsets, respectively. Scatter plots (A–D) – the relative numbers of EM1, EM2, EM3 and EM4 cells within EM CD8+ T cell subset; scatter plots (E–H) – EM1, EM2, EM3 and EM4 cells concentrations (number of cell per 1μL of peripheral whole blood). Black circles denote patients with Sjögren’s syndrome (SS, n = 34); white circles—healthy control (HC, n = 34). Each dot represents individual subjects, and horizontal bars depict the group medians and quartile ranges (Med (Q25; Q75)). The statistical analysis was performed with the Mann–Whitney U test.
Figure 3.
Alterations in relative and absolute number of EM CD8+ T cell subsets with different patterns of CD27 and CD28 expression in patients with pSS. Scatter plots (A–D) and (E–H) —EM CD8+ T cell were subdivided into EM1 (CD27+CD28+), EM2 (CD27+CD28−), EM3 (CD27−CD28−), and EM4 (CD27−CD28+) subsets, respectively. Scatter plots (A–D) – the relative numbers of EM1, EM2, EM3 and EM4 cells within EM CD8+ T cell subset; scatter plots (E–H) – EM1, EM2, EM3 and EM4 cells concentrations (number of cell per 1μL of peripheral whole blood). Black circles denote patients with Sjögren’s syndrome (SS, n = 34); white circles—healthy control (HC, n = 34). Each dot represents individual subjects, and horizontal bars depict the group medians and quartile ranges (Med (Q25; Q75)). The statistical analysis was performed with the Mann–Whitney U test.
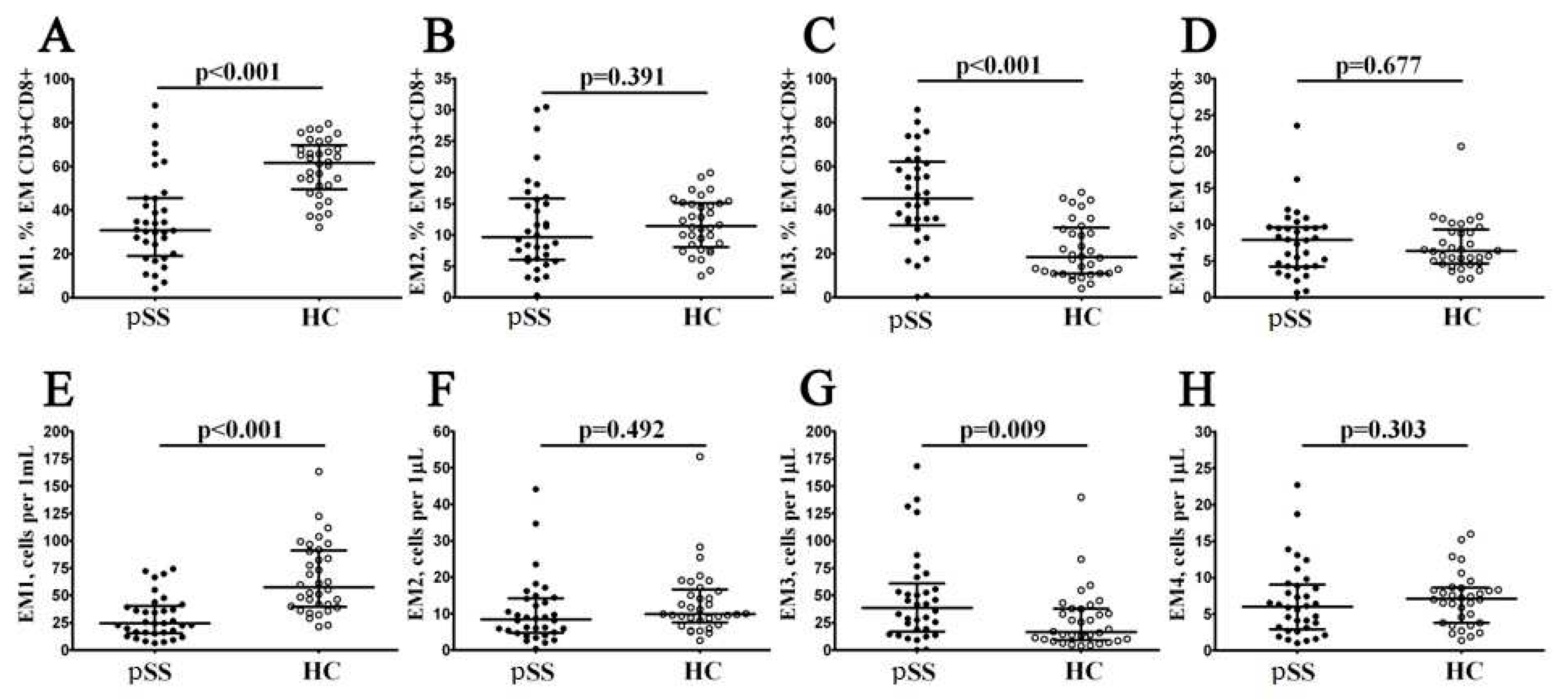
Figure 4.
Alterations in relative and absolute number of TEMRA CD8+ T cell subsets with different patterns of CD27 and CD28 expression in patients with pSS. Scatter plots (A–C) and (D-F) —TEMRA CD8+ T cells were subdivided into CD27+CD28+ pE1, CD27+CD28− pE2, and CD27–CD28– E subsets, respectively. Scatter plots (A–C) – the relative numbers of pE1, pE2 and effector cells within TEMRA CD8+ T cell subset; scatter plots (D-F) – pE1, pE2 and effector cells concentrations (number of cell per 1μL of peripheral whole blood). Black circles denote patients with Sjögren’s syndrome (SS, n = 34); white circles—healthy control (HC, n = 34). Each dot represents individual subjects, and horizontal bars depict the group medians and quartile ranges (Med (Q25; Q75)). The statistical analysis was performed with the Mann–Whitney U test.
Figure 4.
Alterations in relative and absolute number of TEMRA CD8+ T cell subsets with different patterns of CD27 and CD28 expression in patients with pSS. Scatter plots (A–C) and (D-F) —TEMRA CD8+ T cells were subdivided into CD27+CD28+ pE1, CD27+CD28− pE2, and CD27–CD28– E subsets, respectively. Scatter plots (A–C) – the relative numbers of pE1, pE2 and effector cells within TEMRA CD8+ T cell subset; scatter plots (D-F) – pE1, pE2 and effector cells concentrations (number of cell per 1μL of peripheral whole blood). Black circles denote patients with Sjögren’s syndrome (SS, n = 34); white circles—healthy control (HC, n = 34). Each dot represents individual subjects, and horizontal bars depict the group medians and quartile ranges (Med (Q25; Q75)). The statistical analysis was performed with the Mann–Whitney U test.
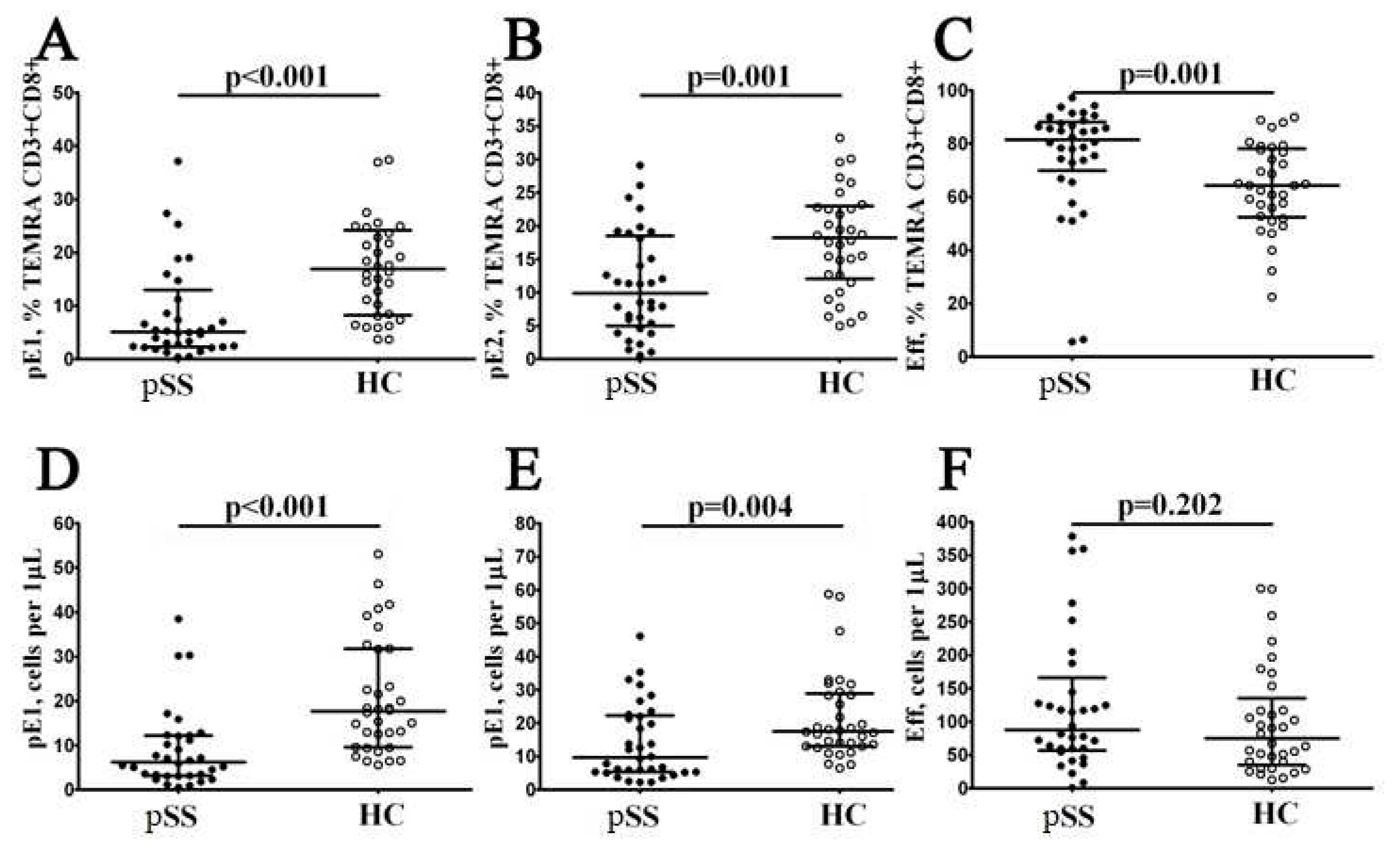
Figure 5.
Imbalance in relative and absolute numbers of peripheral blood Tc1, Tc2, Tc17, and double-positive Tc17.1 cells inpatients with pSS. Scatter plots (A–D) and (E–H) —Tc1 (CCR6−CXCR3+), Tc2 (CCR6−CXCR3−), Tc17 (CCR6+CXCR3−), and double-positive Tc17.1 (CCR6+CXCR3+) frequencies, respectively. Scatter plots (A–D) – the relative numbers of Tc1, Tc2, Tc17, and Tc17.1 within total CD8+ T cell subset; scatter plots (E–H) – Tc1, Tc2, Tc17, and Tc17.1 concentrations (number of cell per 1μL of peripheral whole blood). Black circles denote patients with Sjögren’s syndrome (SS, n = 34); white circles—healthy control (HC, n = 34). Each dot represents individual subjects, and horizontal bars depict the group medians and quartile ranges (Med (Q25; Q75)). The statistical analysis was performed with the Mann–Whitney U test.
Figure 5.
Imbalance in relative and absolute numbers of peripheral blood Tc1, Tc2, Tc17, and double-positive Tc17.1 cells inpatients with pSS. Scatter plots (A–D) and (E–H) —Tc1 (CCR6−CXCR3+), Tc2 (CCR6−CXCR3−), Tc17 (CCR6+CXCR3−), and double-positive Tc17.1 (CCR6+CXCR3+) frequencies, respectively. Scatter plots (A–D) – the relative numbers of Tc1, Tc2, Tc17, and Tc17.1 within total CD8+ T cell subset; scatter plots (E–H) – Tc1, Tc2, Tc17, and Tc17.1 concentrations (number of cell per 1μL of peripheral whole blood). Black circles denote patients with Sjögren’s syndrome (SS, n = 34); white circles—healthy control (HC, n = 34). Each dot represents individual subjects, and horizontal bars depict the group medians and quartile ranges (Med (Q25; Q75)). The statistical analysis was performed with the Mann–Whitney U test.
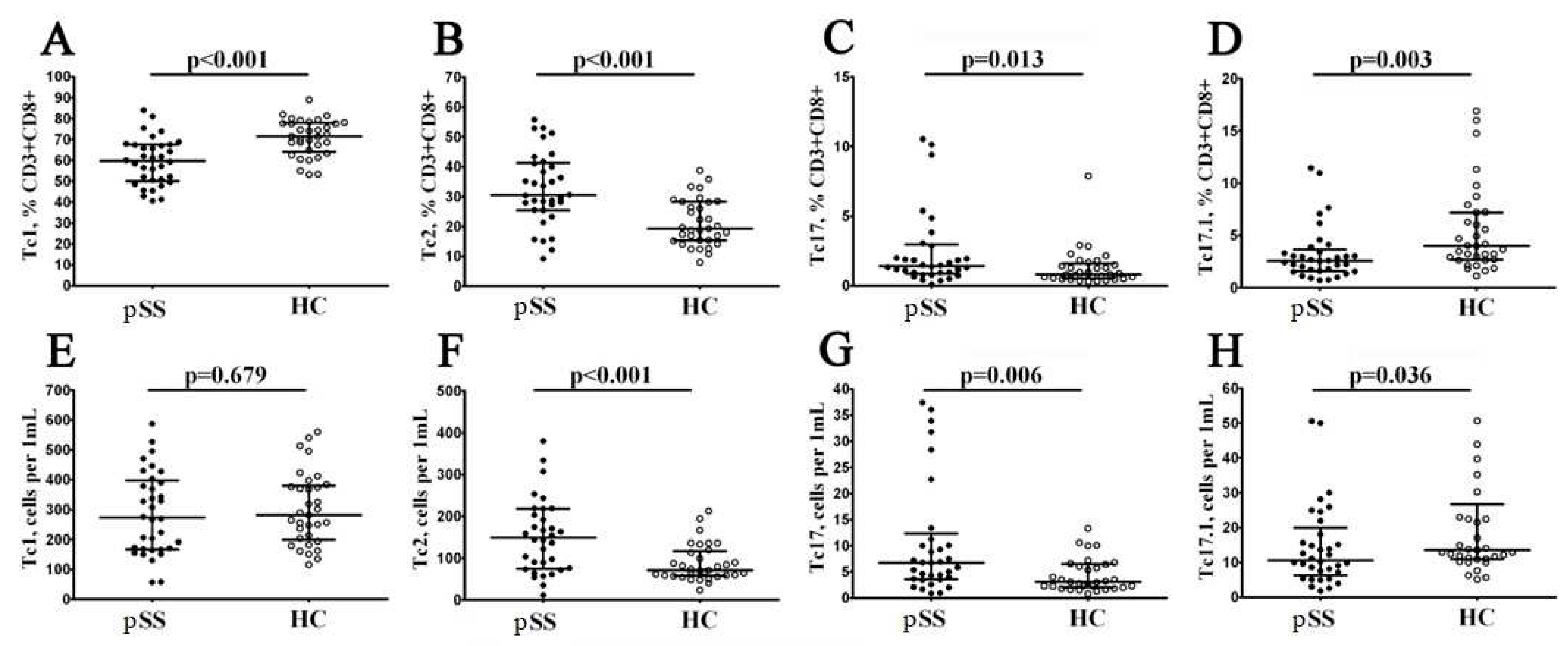
Figure 6.
Heat map of correlations between chemokines levels and frequencies of ‘polarized’ CD8+ T cell subsetsin patients with pSS (n = 20). Color scale bar shows a range of correlation coefficients (r). Correlation analysis was performed using nonparametric Spearman rank test, only significant correlation are shown (significance was set at p < 0.05). The red color represents a high positive correlation, decreasing to the blue color bar, which represents a negative correlation.
Figure 6.
Heat map of correlations between chemokines levels and frequencies of ‘polarized’ CD8+ T cell subsetsin patients with pSS (n = 20). Color scale bar shows a range of correlation coefficients (r). Correlation analysis was performed using nonparametric Spearman rank test, only significant correlation are shown (significance was set at p < 0.05). The red color represents a high positive correlation, decreasing to the blue color bar, which represents a negative correlation.
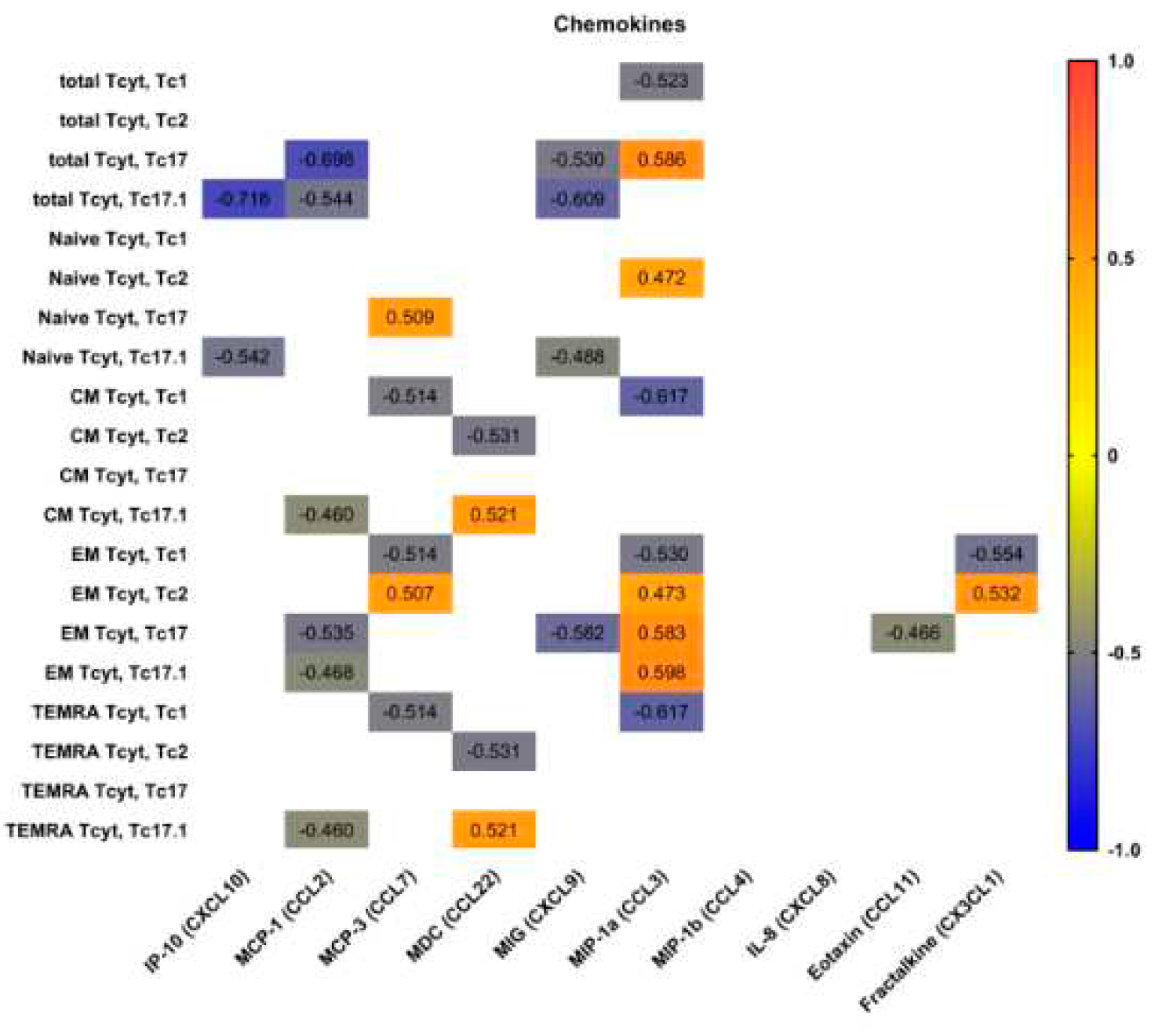
Figure 7.
Heat map of correlations between main proinflammatory and effector cytokine levels and frequencies of ‘polarized’ CD8+ T cell subsetsin patients with pSS (n = 20). Color scale bar shows a range of correlation coefficients (r). Correlation analysis was performed using nonparametric Spearman rank test, only significant correlation are shown (significance was set at p < 0.05). The red color represents a high positive correlation, decreasing to the blue color bar, which represents a negative correlation.
Figure 7.
Heat map of correlations between main proinflammatory and effector cytokine levels and frequencies of ‘polarized’ CD8+ T cell subsetsin patients with pSS (n = 20). Color scale bar shows a range of correlation coefficients (r). Correlation analysis was performed using nonparametric Spearman rank test, only significant correlation are shown (significance was set at p < 0.05). The red color represents a high positive correlation, decreasing to the blue color bar, which represents a negative correlation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



