
Submitted:
30 August 2023
Posted:
30 August 2023
You are already at the latest version
Abstract
The aim was to determine the effect of Sechium edule var. nigrum spinosum (chayote) on gene ex-pression related to antioxidant protection mechanisms and the inflammatory process in older adults with metabolic syndrome (MetS). A quasi-experimental study was carried out in a con-venience sample of 46 older adults diagnosed with MetS: (i) placebo group (PG; n=20), (ii) ex-perimental group (EG; n=26). Clinical, biochemical, and anthropometric parameters, SOD, GPx, and CAT enzyme activity, total oxidant status (TOS), total antioxidant status (TAS), oxidative stress index (OSI), cytokines (IL-6, IL-8 and TNF-α) and mRNA expression of (SOD, GPx, CAT, IL-6, IL-8, TNF-α, Nrf2, NFkB p50 and NFkB p65), were measured at baseline and 6 months post-intervention. A statistically significant decrease was observed in TOS (baseline, 28.9 ± 3.6 vs. post, 23.7 ± 3.4, p < 0.01) and OSI (baseline, 24.1 ± 3.8 vs. post, 17.7 ± 4). As well as the increase in IL-6 (baseline, 10.7 ± 1.1 vs. post, 12.3 ± 2, p = 0.03), SOD activity (baseline, 167.1 ± 11.9 vs. post, 180.6 ± 7.6, p < 0.05), CAT activity (baseline, 1.0 ± 0.2 vs. post, 1.3 ± 0.2, p < 0.01) and TAS (baseline, 1.1 ± 0.1 vs. post, 1.4 ± 0.1, p < 0.01) in the EG compared to the PG. Regarding the expression of Nrf2, SOD and IL-6, the EG showed a significant increase vs basal levels (47%, 44% and 43%, re-spectively). Our findings suggest that Sechium edule supplementation promotes the antioxidant response and decreases oxidative stress via Nrf2.
Keywords:
Sechium edule
; metabolic syndrome
; antioxidants
; markers inflammatory
; aging
; RNAm
1. Introduction
In recent years, there has been an increase in the number of older adults worldwide; projections estimate that by 2030, 16% of the population will be 60 years of age or older [1]. In this sense, human aging is the result of a gradual and adaptive process characterized by a decrease in the biological response accompanied by social, psychological, morphological, physiological, and biochemical modifications, favored by the genetic load and accumulated wear and tear in the face of the challenges that the person faces over time throughout its history [2]. Aging can predispose to the development of non-communicable chronic diseases; among them metabolic syndrome (MetS).
The MetS leads to high rates of morbidity and mortality, represents a major threat to public health systems; since it increases the risk of presenting cardiovascular disease, type II diabetes mellitus (T2DM), development of neurodegenerative diseases and ultimately death [3,4,5]. Its prevalence in Latin America is almost 25%, with women and older adults being the most compromised [6]. Metabolic abnormalities that accompany this syndrome include at least three of the following disorders: (i) arterial hypertension, (ii) increased blood glucose, (iii) central (abdominal) obesity, and (iv) dyslipidemia [increased triglycerides and decreased high-density lipoporotein cholesterol (HDL-c)] [7].
Oxidative stress (OxS) and chronic inflammation (CI) play a significative role in the pathogenesis of MetS [4,8]. The increase in reactive oxygen species (ROS) in MetS is pointed out as an underlying mechanism for mitochondrial dysfunction, accumulation of oxidative damage at the level of lipids, proteins and nucleic acids, a consequence of a poor antioxidant status compared to those who do not suffer MetS [9,10,11].
Likewise, various studies have shown that MetS is associated with a state of low-grade inflammation, characterized by abnormal production of proinflammatory cytokines and acute phase reactants such as interleukin-6 (IL-6), tumor necrosis factor alpha (TNF -α), fibrinogen and C reactive protein (CRP).
It has been pointed out that individually each of the MetS alterations by themselves are associated with both OxS and chronic inflammation [12]. Hence, the importance of finding strategies to avoid the complications that the presence of MetS entails; pharmacotherapy has been proposed, as well as the implementation of healthy lifestyles such as regular physical activity and a healthy diet [10,11,13]. However, adopting healthy habits is usually difficult for the population, so it is still necessary to explore new strategies such as nutraceutical supplements, which are well accepted among the elderly.
In this regard, Sechium edule (chayote) is an edible fruit of the Cucurbitaceae family, to which various benefits on human health had attributed due to its antioxidant, anti-inflammatory, antitumor and hypotensive properties [14,15,16]. Our research team has shown that consumption of Sechium edule reduces OxS and IC related to MetS and [17]. However, the transcripts and regulators that could be involved in this decrease are still unknown. Therefore, the aim of the present study was to determine the effect of Sechium edule supplementation on gene expression related to antioxidant protection mechanisms and the inflammatory process in older adults with MetS.
2. Materials and Methods
2.1. Experimental design
This study was approved by the Bioethics in Research and Biosafety Committee of the Faculty of Higher Studies Zaragoza UNAM (23/02-SO/2.4.2) with trial registration number (ISRCTN: 43215432). Informed consent was obtained from each of the participants. All procedures were carried out following the ethical principles of the Declaration of Helsinki of the World Medical Association. The fruit Sechium edule var. nigrum spinosum was donated by the Interdisciplinary Group for the study of Sechium edule of Mexico A.C. (GISeM). The biological material was collected in a state of horticultural maturity; subsequently, it was selected, washed, sectioned into slices, dried at 40 °C, and pulverized (epidermis, seeds, and spines). The formulation of both the active (Sechium edule) and placebo capsules was carried out in the pharmaceutical development laboratory of the FES-Z. The placebo group (PG) received capsules of lactose monohydrate and pharmaceutical talc, both United States Pharmacopeia (USP) grade (Sigma, St. Louis, MO, USA) and with physical characteristics identical to those of the nutraceutical. The particle size was standardized to obtain a fine powder of the fruit and with it, there were elaborate capsules based on rheological studies to guarantee the optimal conditions of filling, homogeneity, and long-term stability of the powder. After the design, the treatments were manufactured and packaged by a pharmaceutical company specialized in the field of nutraceuticals.
2.2. Intervention
The intervention consisted of consuming three 500 mg/day capsules of Sechium edule or placebo (one before each meal) for six months. The selection of the dose according to the phytochemical and safety profile has already been described by our work group [17,18]. The type of sampling was for convenience in a population of 46 older adults, with an average age of 67.6 ± 5.85. All participants were diagnosed with MetS according to the National Cholesterol Education Program Adult Treatment Panel (NCEP/ATP III) criteria: (i) waist circumference ≥ 102 cm for men or ≥ 88 cm for women; (ii) blood pressure ≥ 130/85 mmHg; (iii) glucose > 110 mg/dL; (iv) High-density lipoprotein cholesterol (HDL-c) < 40 mg/dL in men or <50 mg/dL in women; and (v) triglycerides ≥ 150 mg/dL. MetS was diagnosed with presence of at least three of five criteria [19].
Participants were randomly assigned to the experimental group with Sechium edule intervention (EG; n=50) or to the placebo group (PG; n=50). Of these patients, only certain participants wished to donate venous blood for gene expression determinations (PG; n=20) and (EG; n=26). All measurements in both groups were made before the treatments (baseline measurements) and six months after the intervention (post-treatment measurements).
2.3. Anthropometric and clinic measurements
Measurements of body weight and waist circumference were made. For body weight, a calibrated medical scale (SECA, Hamburg, Germany) was used; while the distribution of abdominal fat was obtained by measuring the waist circumference with a medical tape measure (SECA, Hamburg, Germany), which was placed at the level of the navel. All measurements were performed by trained FES-Z personnel [20].
To determinate systolic (SBP) and diastolic (DBP) blood pressure, a calibrated mercury baumanometer was used. The patient remained at rest for at least five minutes before taking the blood pressure, sitting in a seat with a backrest, with a straight back, feet resting on the floor and legs uncrossed. In addition, the Osler technique was used to identify pseudohypertension [21].
2.4. Biochemical analysis
2.4.1. Samples
After fasting for 8 hours, blood samples were obtained by venipuncture, collected in vacuum tubes without anticoagulant for clinical chemistry determinations (glucose, high-density lipoprotein cholesterol (HDL-c) and triglycerides), and serum cytokines; in tubes with EDTA as anticoagulant for the determination of catalase activity (CAT) and extraction of lymphocytes and, in tubes with sodium heparin for the determination of erythrocyte superoxide dismutase (SOD) and glutathione peroxidase (GPx) activity while in heparinized plasma was determined total antioxidant status (TAS) and total oxidant status (TOS). The SOD, GPx, CAT, TAS, TOS techniques were performed at microscale in multiwell plates, which were read on a Multiskan Go spectrophotometer, version 1.00.40 (Thermo Scientific, Denver, CO, USA). The lipid and glycemic profile were determined by colorimetric techniques with an automated chemical-clinical analyzer Selectra Junior (Vital Scientific, Dieren, Netherlands).
2.4.2. SOD enzyme activity
For the activity of the SOD enzyme, the commercial kit (Randox Laboratories Ltd., Antrim, UK) was used, following the manufacturer’s instructions. Xanthine oxidase acts as a generator of superoxide radicals, which react with 2-(4-iodophenyl)-3-(4-nitrophenol)-5-phenyl-tetrazolium chloride to form the formazan dye, which is read at 505 nm. The inhibition of this reaction in the presence of SOD allows us to determine its activity.
2.4.3. GPx enzyme activity
In the presence nicotinamide adenine dinucleotide phosphate (NADPH) and glutathione reductase, glutathione peroxidase (GPx) catalyzes the oxidation of glutathione (GSH) by cumene hydroperoxide. Oxidized glutathione (GSSG) is immediately converted into the reduced form with the subsequent oxidation of NADPH to NADP (Randox Laboratories, Ltd., Crumlin Co., UK). The decrease in absorbance was measured at 340 nm UV-spectrophotometer with a Multiskan TM Go Microplate (Thermo Scientific TM).
2.4.4. Catalase activity
The catalase activity was determined by spectrophotometry, using hydrogen peroxide (H2O2) as substrate (Sigma, St. Louis, MO, USA). For this, was made a mixture of 190 μL of working solution (0.1 M phosphate buffer, pH = 7.0 and 20 mM H2O2) and 10 μL of the sample. The decrease in H2O2 concentration was measured every 15 s for 3 min at a wavelength of 240 nm [22].
2.4.5. Total oxidant status (TOS)
Plasma TOS determination was performed using the commercial kit (Rel Assay Diagnostics, Gaziantep, TR), following the supplier’s instructions. Oxidants present in the sample oxidize the ferrous ion chelating complex to ferric ion. The latter forms a colored complex with the chromogen in an acid medium. In such a way, that the intensity of the color is directly proportional to the amount of oxidant molecules present in the sample. Measurements were made at a wavelength of 530 nm.
2.4.6. Total antioxidant status (TAS)
Total antioxidant status was determined using the commercial kit (Randox Laboratories Ltd., Antrim, UK); based on the reaction of 2,2′-azino-bis (3-ethylbenzthiazoline-6-sulfonic acid) (ABTS) with metmyoglobin and H2O2. This reaction stimulates the formation of the cationic radical ABTS+; which produces a blue-green color. The intensity of the color is inversely proportional to the amount of antioxidants present in the sample. The kinetics of the reaction was determined at a wavelength of 600 nm.
2.4.7. Inflammatory cytokines
Serum concentrations of interleukin-6 (IL-6), interleukin-8 (IL-8), and tumor necrosis factor alpha (TNF-α) were determined using the Human Inflammatory Cytokine Cytometric Bead Array (CBA) kit (BD, Biosciences, San Jose, CA, USA), which consists of a sandwich capture method with beads conjugated with specific antibodies and a detection reagent (phycoerythrin mixture-conjugated antibodies), which results in a fluorescent signal in proportion to the number of bound analytes, which were detected by the flow cytometer (BD Biosciences, San Jose, CA, USA), and the FCAP ArrayTM version 3.0 software.
2.4.8. Lymphocyte isolation and RNA extraction
Lymphocyte isolation was performed from 5 mL of venous blood, diluted 1:1 with phosphate buffered saline (PBS) (Sigma, St. Louis, MO, USA)/2% fetal bovine serum (FBS) (ThermoFisher Scientific, Waltham, MA, USA). Subsequently, 4 mL of Ficoll-paque (Gibco ThermoFisher Scientific, Waltham, MA, USA) were added. It was centrifuged at 200 g and the opaque interface of interest was removed. From 2 × 106 lymphocytes, RNA extraction was performed using the RNeasy Mini kit isolation kit (Qiagen, Hilden, Düssseldorf, Germany), following the manufacturer’s recommendations. RNA extraction was performed just after lymphocyte isolation, aliquoted, and stored at -70°C until further use. The RNA was quantified and from 5 µg its integrity was determined in a 1% agarose gel with ethidium bromide and tris-acetate-EDTA buffer visualized in a Kodak Molecular Imaging Software (v.4.5.1) imaging system; while its purity was calculated using the A260/A280 ratio. All the RNA samples used in the present study were considered intact and with an optimal purity value.
2.4.9. Gene expression analysis
The reactions were performed from 10 ng of RNA and forward and reverse primers at a final concentration of 100 nM (IDT, IA, USA) (Table 1). It should be noted that the primers were designed with the Primer-BLAST-NCBI- tool NIH [23]. For the analysis of gene expression, the QuantiFast SYBR Green RT-PCR kit (one-step RT-PCR) kit (Qiagen, Hilden, Düssseldorf, Germany) was used, which allows the simultaneous execution of the reverse transcription and the PCR reaction. Reaction conditions were as follows: 50 °C for 10 min for reverse transcription, 95 °C for 5 min for PCR initial activation step, followed by 40 three-segment cycles to amplify the specific PCR product: denaturation at 95 °C for 10 s, annealing at 60°C for 30 s and extension at 72°C for 15 s. The mean crossing threshold (Ct) of each gene was normalized to the mean Ct of the housekeeping gene β-actin.
3. Results
Table 2 shows the clinical and anthropometric parameters of both study groups: experimental group (EG) and placebo group (PG) pre and post-treatment. The EG showed a statistically significant decrease in both SBP (baseline, 141.4 ± 12.1 vs. post, 131.2 ± 11.2, p < 0.01) and DBP (baseline, 95 ± 9.8 vs. post, 83.8 ± 8, p < 0.001); as well as in body weight at six months post-treatment (baseline, 74.4 ± 16.5 vs. post, 72.4 ± 16.4, p < 0.01). On the other hand, no statistically significant changes were observed in waist circumference regardless of the intervention.
Regarding the biochemical parameters related to the pathophysiology of MetS, only the concentration of HDL-c in the EG increased at six months post-treatment (baseline, 42.8 ± 7.5 vs post, 47.4 ± 7.4, p < 0.01) compared to the PG. Meanwhile, glucose and triglyceride levels did not show apparent changes in both post-treatment groups (Table 3).
Regarding the antioxidant activity markers, a statistically significant increase was observed in the post-treatment in the EG compared to the PG in the activity of enzymes with antioxidant function SOD (baseline, 167.1 ± 11.9 vs post, 180.6 ± 7.6, p < 0.05 ) and CAT (baseline, 1.0 ± 0.2 vs post, 1.3 ± 0.2, p < 0.01); as well as in the total antioxidant capacity (TAS) (baseline, 1.1 ± 0.1 vs post, 1.4 ± 0.1, p < 0.01); on the contrary, there was a decrease in the total oxidant status (TOS) (baseline, 28.9 ± 3.6 vs post, 23.7 ± 3.4, p < 0.01) and the OSI (EG: baseline, 24.1 ± 3.8 vs post, 17.7 ± 4 vs PG: baseline, 19.2 ± 2.1 vs post, 26.1 ± 7, p < 0.01) in the EG (Table 4).
Regarding the interleukins, a statistically significant increase was observed in the IL-6 concentration of the EG post-treatment (baseline, 10.7 ± 1.1 vs post, 12.3 ± 2, p = 0.03), compared to the PG. The levels of IL-8 and TNF-α did not present statistically significant changes (Table 5).
Figure 1 shows the relative expression of the RNAm of genes that code for proteins that participate in antioxidant protection mechanisms and in the inflammatory process. SOD (Figure 1A), Nrf2 (Figure 1D) and IL-6 (Figure 1E) gene expressions showed a statistically significant increase in post-treatment GA of 44%, 47%, and 43% with respect to their basal levels.
4. Discussion
MetS is a set of systemic alterations whose pathophysiology has been linked to a pro oxidative and inflammatory state [7]. It has been pointed out that individually each of the MetS alterations are linked to OxS [12]. On the other hand, various studies have shown that MetS is associated with a state of low-grade inflammation, characterized by abnormal production of proinflammatory cytokines and acute phase reactants such as tumor necrosis factor alpha (TNF-α), fibrinogen and C reactive protein (CRP). Given the high prevalence of this syndrome, and the difficulty of treating the set of alterations that compose it, it is necessary to have alternatives that help in its control. In this sense, Sechium edule has been shown to have hypotensive, lipolytic, antioxidant, and anti-inflammatory effects, which makes it a feasible option for older adults in whom there is also a greater acceptance of natural treatments [24].
The results of this research show that the consumption of Sechium edule has positive effects at a biochemical and systemic level. On the one hand, we observed that it promoted weight loss in the study population, a finding consistent with previous results reported by our research team, which can be explained by its lipolytic effect. It has been reported that the polyphenols present in Sechium edule activate signaling pathways that decrease the activity of lipogenic relative enzymes, such as FAS (fatty acid synthase), HMGCoR (HMG-CoA reductase), and SREBPs (sterol regulatory element binding proteins), and increase the expression of CPT-I (carnitine palmitoyltransferase I) and PPARα (peroxisome proliferators activated receptor α), which are critical regulators of hepatic lipid metabolism [25,26], this, associated with the direct effect on the digestive process due to its low caloric content and high amount of insoluble fiber allow weight loss, which is a significant result given the role of obesity in the pathophysiology of high-frequency non-communicable chronic diseases among this population group.
Likewise, regarding blood pressure, we observed a significant decrease, a finding that may be due to the activity of flavonoids, such as quercetin and coumaric acid which affect the renin-angiotensin system and the modulation of calcium release with an effect on endothelial vasodilation and consequently on blood pressure, this result is also consistent with previous findings reported by our research group [16,17,18].
Regarding circulating lipids, in this investigation a significant increase in HDL-cholesterol was observed in the group that consumed Sechium edule. In this regard, it has been pointed out that compounds such as naringenin and quercetin influence HDL function beyond HDL cholesterol concentration by regulating cellular cholesterol efflux from macrophages and hepatic paraoxonase 1 expression and activity [27].
On the other hand, the determined OxS markers, it was observed that the consumption of Sechium edule had a marked antioxidant effect, a finding that coincides with and corroborates previous results of our research group. In this case, we found a significant increase in the activity of the SOD and CAT enzymes, as well as an increase in TAS, a marker that reflects extracellular antioxidant activity. Likewise, there was a decrease in the TOS oxidation marker coupled with a decrease in OSI, an index that reflects the TOS/TAS relationship; together, these results show that the group that consumed Sechium edule had an increase in antioxidant activity with a concomitant decrease in oxidation. Likewise, an effect on IL6 was observed, which increased significantly in the EG, which may suggest an impact on the inflammatory process, which is closely related to OxS, since the regulatory mechanisms frequently present intersection points. These effects, as we have pointed out in detail in a previous investigation [18], have been consistently reported and are due to the variety of bioactive compounds specifically, phenolic acids, cucurbitacins, and flavonoids present in chayote capsules, which they synergistically affect the activity and even expression of antioxidant molecules.
Regarding the mechanisms behind this effect, there is evidence of the signaling pathways in which these bioactives interfere, in various in vivo and in vitro studies the capacity of some of these molecules has been demonstrated, for example, quercetin and naringenin, to bind to the master regulator of the antioxidant response, erythroid-derived nuclear factor 2 (Nrf2) to DNA [28,29,30].
Regarding cucurbitacins, it has been reported that they present antioxidant and chemoprotective activity [31,32,33]. Specifically, cucurbitacins I, D, B, and E, promote the expression of phase II detoxification enzymes heme oxygenase-1 (HO-1) and NAD(P)H dehydrogenase quinone 1 (NQO-1) through modulation of the Nrf2 transcription factor, which in turn regulates the expression of SOD, GPx, and CAT. Likewise, cucurbitacins cause the inhibition of nuclear factor enhancing the kappa light chain of activated B cells (NFkB) [34].
Also, myricetin, a flavonoid, increases the nuclear accumulation of Nrf2 and inhibits NFkB, generating a protective effect [35,36]. For its part, phlorizin stimulates the translocation of Nrf2 from the cytoplasm to the nucleus and upregulates its down-stream antioxidant response element (ARE), which includes the enzymes HO-1 and NQO-1 in an animal model with oxidative injury [37]. For its part, naringenin reduces ROS levels through the Nrf2/ARE pathway [38]. About the phenolic acids present in Sechium edule, they have been reported to have antioxidant and anti-inflammatory effects mediated by Nrf2 and NFkB, respectively. In the case of chlorogenic acid, it exerts a nephroprotective effect by inhibiting OxS and inflammation [39]; gallic acid increases nuclear Nrf2 levels and attenuates oxidative damage [40].
These findings are consistent with those previously reported by our research group and are consistent with the results obtained in this work regarding the expression of the genes of the molecules involved in the antioxidant response.
In this sense, it has been shown in experimental models of aging that the expression of genes that code for proteins that participate in antioxidant defense mechanisms decreases; suggesting that this alteration is involved in oxidative damage, possibly due to a decrease in cell signaling [41,42,43].
In the present study, we were able to observe a parallel increase in the relative expression levels of SOD mRNA by around 12% and enzymatic activity by 8% in the post-treatment EG. That is, the consumption of Sechium edule improves the efficiency of the SOD enzyme from transcriptional levels, which makes the superoxide anion less dangerous for the cell [44], which leads to the reduction of oxidative damage at the of lipids, proteins, and DNA, as previously reported [17,18,45,46].
On the other hand, we observed that gene expression and GPx activity did not present statistically significant changes in the study groups. While CAT activity increased by 30%, without significant changes in its mRNA levels; these results lead us to assume that the increase in SOD and plasmatic antioxidants (TAS) were sufficient to counteract the reactive species, so that there was no increase in the de novo synthesis of CAT and neither in GPx because it was not necessary [41,47].
About the expression and content of IL-6 it increased by 43% and 20%, respectively, in the post-treatment EG. Regarding this cytokine, a possible duality has been pointed out depending on the conditions of the organism, it has been reported that in pathologies such as DMT2, there is a loss of IL-6 signaling leading to oxidative damage and cell death [48]. While the increase in its levels protects from this type of damage at the DNA level, coupled with a positive regulation of SOD, via Nrf2; which agrees with our results [49]. Therefore, our findings suggest that, in this case, IL-6 exerts an antioxidant-like protective effect against OxS mediated by Nrf2 [50].
In this sense, in the present study, an increase in Nrf2 mRNA levels was observed by 47% in the post-treatment EG, compared to its basal levels, which is consistent with the overall increase in the antioxidant response observed.
Our results since there were no significant changes in the mRNAs of the NFκB p50/p65 factor. This allows us to assume that the transcriptional regulation of genes involved in the inflammatory process is more difficult to modify through nutraceutical supplementation.
Finally, it is worth mentioning that, as explained in detail in a previous study, practically all the secondary metabolites present in Sechium edule modulate the expression of Nrf2 mRNA; for example, cucurbitacins, naringen, caffeic, and ferulic acids have been related to an increase in Nrf2 gene expression and inhibition of NFκB-mediated signaling [51,52,53,54]. Apigenin, myrecetin, and gallic acid improve translocation and/or nuclear accumulation [55,56,57]. Quercetin only temporarily stimulates its expression [58]. Phlorizin, rutin, and chlorogenic and protocatechuic acids enhance antioxidant protection through this same pathway [59,60,61,62] and finally the hypolipidemic effects exerted by p-coumaric acid are directly related to the expression of Nrf2 [63].
5. Conclusions
The results of our research, that is, the increase in the levels of gene expression, activity, or content of SOD, CAT, and IL-6, as well as the expression of Nrf2, in the group that consumed Sechium edule, suggest that the antioxidant effect global observed is mediated by the transcriptional factor Nrf2.
Author Contributions
V.M.M.-N designed the study, wrote the manuscript, and analyzed the data. J.R.-P., D.H.-A., T.L.A.-U., I.A.-S., E.S.-O., and G.G.-G. performed the study and analyzed the data. All authors reviewed the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the General Directorate of Academic Personnel Affairs, National Autonomous University of Mexico (DGAPA-UNAM) (PAPIIT IN215821), and the Secretariat of Science and Technology and Innovation Project of Mexico City (SECITI) (SECITI/045/ 2018).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Ethics Committee of the “National Autonomous University of Mexico (UNAM) - Zaragoza Campus”’.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author, [VMM-N], upon reasonable request.
Acknowledgments
We appreciate the support from National Council for Science and Technology (CONACyT) for the scholarship granted to Graciela Gavia-García for postdoctoral stay.
Conflicts of Interest
No potential conflict of interest was reported by the authors.
References
- WHO. Ageing and health. 2022. Available online: https://www.who.int/news-room/fact-sheets/detail/ageing-and-health.
- Mendoza-Núñez, V.M.; Martínez-Maldonado, M.L.; Vivaldo-Martínez, M. What Is the onset age of human aging and old age? Int J Gerontol 2016, 10, 56. [Google Scholar] [CrossRef]
- Caterson, I.D.; Hubbard, V.; Bray. G.A.; Grunstein, R.; Hansen, B.C.; Hong, Y.; Labarthe, D.; Seidell, J.C.; Smith, S.C. American Heart Association. Prevention Conference VII: Obesity, a worldwide epidemic related to heart disease and stroke: Group III: worldwide comorbidities of obesity. Circulation 2004, 110, e476–e483. [Google Scholar] [CrossRef] [PubMed]
- Soares, R.; Costa, C. Oxidative stress, inflammation and angiogenesis in the metabolic syndrome. Amsterdam, The Netherlands: Springer 2009, 85-121.
- Gouveia, É.R.; Gouveia, B.R.; Marques, A.; Peralta, M.; França, C.; Lima, A.; Campos, A.; Jurema, J.; Kliegel, M.; Ihle, A. Predictors of metabolic syndrome in adults and older adults from Amazonas, Brazil. Int J Environ Res Public Health 2021, 18, 1303. [Google Scholar] [CrossRef] [PubMed]
- Márquez-Sandoval, F.; Macedo-Ojeda, G.; Viramontes-Hörner, D.; Fernández, B.J.D.; Salas, S.J.; Vizmanos, B. The prevalence of metabolic syndrome in Latin America: a systematic review. Public Health Nutr 2011, 14, 1702–1713. [Google Scholar] [CrossRef] [PubMed]
- Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z. The metabolic syndrome. Lancet 2005, 365, 1415–1428. [Google Scholar] [CrossRef]
- Adair, T.H.; Montani, J.P. Angiogenesis. San Rafael (CA): Morgan & Claypool Life Sciences. 2010.
- Vona, R.; Gambardella, L.; Cittadini, C.; Straface, E.; Pietraforte, D. Biomarkers of oxidative stress in metabolic syndrome and associated diseases. Oxid Med Cell Longev 2019, 8267234. [Google Scholar] [CrossRef]
- Sharma, P.; Mishra, S.; Ajmera, P.; Mathur, S. Oxidative stress in metabolic syndrome. Indian J Clin Biochem 2005, 20, 145–149. [Google Scholar] [CrossRef]
- Fernandez-Garcia, J.C; Cardona, F.J.; Tinahones, F. Inflammation, oxidative stress and metabolic syndrome: dietary modulation. Curr Vasc Pharmacol 2013, 11, 906–919. [Google Scholar] [CrossRef]
- Pruchniak, M.P.; Aražna, M.; Demkow, U. Biochemistry of oxidative stress. Adv Exp Med Biol 2016, 878, 9–19. [Google Scholar]
- Krzemińska, J.; Wronka, M.; Młynarska, E.; Franczyk, B.; Rysz, J. Arterial hypertension—oxidative stress and inflammation. Antioxidants 2022, 11, 172. [Google Scholar] [CrossRef]
- Ordonez, A.A.L.; Gomez, J.D.; Vattuone, M.A. Antioxidant activities of Sechium edule (Jacq.) Swartz extracts. Food Chem 2006, 97, 452–458. [Google Scholar] [CrossRef]
- MinJin, K.; YouChul, C.; SangSuk, K.; ChanKyu, L.; KyungJin, P.; YoungHun, C.; Taejin, P.; SeungYoung, K.; ChangGu, H. Anti-inflammatory effect of Sechium edule extract in LPS-stimulated RAW 264.7 cells via p-JNK and p-p38 down-regulation. KSBB J 2019, 34, 99–106. [Google Scholar]
- Lombardo-Earl, G.; Roman-Ramos, R.; Zamilpa, A.; Herrera-Ruiz, M.; Rosas-Salgado, G.; Tortoriello, J.; Jiménez-Ferrer, E. Extracts and fractions from edible roots of Sechium edule (Jacq.) Sw. with antihypertensive activity. ECAM 2014, 594326. [Google Scholar]
- Arista-Ugalde, T.L.; Santiago-Osorio, E.; Monroy-García, A.; Rosado-Pérez, J.; Aguiñiga-Sánchez, I.; Cadena-Iñiguez, J.; Gavia-García, G.; Mendoza-Núñez, V.M. Antioxidant and anti-inflammatory effect of the consumption of powdered concentrate of Sechium edule var. nigrum spinosum in mexican older adults with metabolic syndrome. Antioxidants (Basel) 2022, 11, 1076. [Google Scholar] [PubMed]
- Gavia-García, G.; Rosado-Pérez, J.; Arista-Ugalde, T.L.; Aguiñiga-Sánchez, I.; Santiago-Osorio, E.; Mendoza-Núñez, V.M. The consumption of Sechium edule (chayote) has antioxidant effect and prevents telomere attrition in older adults with metabolic syndrome. Redox Rep 2023, 28, 2207323. [Google Scholar] [CrossRef]
- NCEP. Executive summary of the third report of the National Cholesterol Education Program (NCEP) expert panel on detection, evaluation, and treatment of high blood cholesterol in adults (Adult treatment panel III). JAMA 2001, 285, 2486–2497. [Google Scholar] [CrossRef] [PubMed]
- Secretaría de Salud. Toma de medidas clínicas y antropométricas en el adulto mayor. Mexico City, (Mexico): Subsecretaría de prevención y protección de la salud; 2002.
- Secretaría de Salud. Norma Oficial Mexicana NOM-030-SSA-1999. Para la prevención, tratamiento y control de la hipertensión arterial. Mexico City, (Mexico): Secretaría de Salud; 1999.
- Aebi, H. Catalase in vitro. Methods Enzymol 1984, 105, 121–126. [Google Scholar] [PubMed]
- Primer-BLAST-NCBI-NIH (Primer-BLAST; https://www.ncbi.nlm.nih.gov/tools/primer-blast/).
- Agbabiaka, T.; Wider, B.; Watson, L.K.; Goodman, C. Concurrent use of prescription drugs and herbal medicinal products in older adults: A systematic review. Drugs Aging 2017, 34, s40017–s40266. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Ou, T.T.; Chang, C.H.; Chang, X.Z.; Yang, M.Y.; Wang, C.J. The polyphenol extract from Sechium edule shoots inhibits lipogenesis and stimulates lipolysis via activation of AMPK signals in HepG2 cells. J Agric Food Chem 2014, 62, 750–759. [Google Scholar] [CrossRef]
- Yang, S.H.; Kim, J.; Lee, M.J.; Kim, Y. Abnormalities of plasma cytokines and spleen in senile APP/PS1/Tau transgenic mouse model. Sci Rep 2015, 5, 15703. [Google Scholar] [CrossRef]
- Millar, C.L.; Duclos, Q.; Blesso, C.N. Effects of dietary flavonoids on reverse cholesterol transport, HDL metabolism, and HDL function. Adv Nutr 2017, 8, 226–239. [Google Scholar] [CrossRef] [PubMed]
- Weng, C.J.; Chen, MJ, Yeh, CT, Yen, G. C. Hepatoprotection of quercetin against oxidative stress by induction of metallothionein expression through activating MAPK and PI3K pathways and enhancing Nrf2 DNA-binding activity. N Biotechnol 2011, 28, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Dreger, H.; Westphal, K.; Weller, A.; Baumann, G.; Stangl, V.; Meiners, S.; Stangl, K. Nrf2-dependent upregulation of antioxidative enzymes: a novel pathway for proteasome inhibitor-mediated cardioprotection. Cardiovasc Res 2009, 83, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, A.K. Nrf2 signaling in coordinated activation of antioxidant gene expression. Free Radic Biol Med 2004, 36, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Kotakeyama, Y.; Li, J.; Pan, Y.; Matsuura, A.; Ohya, Y.; Yoshida, M.; Xiang, L.; Qi, J. Cucurbitacin B exerts antiaging effects in yeast by regulating autophagy and oxidative stress. Oxid Med Cell Longev 2019, 2019, 4517091. [Google Scholar] [CrossRef]
- Liu, Z.; Kumar, M.; Kabra, A. Cucurbitacin B exerts neuroprotection in a murine Alzheimer’s disease model by modulating oxidative stress, inflammation, and neurotransmitter levels. Front Biosci 2022, 27, 71. [Google Scholar] [CrossRef]
- Ranjan, A.; Ramachandran, S.; Gupta, N.; Kaushik, I.; Wright, S.; Srivastava, S.; Das, H.; Srivastava, S.; Prasad, S.; Srivastava, S.K. Role of phytochemicals in cancer prevention. Int J Mol Sci 2019, 20, 4981. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, Y.H.; Park, G. Cucurbitacins attenuate microglial activation and protect from neuroinflammatory injury through Nrf2/ ARE activation and STAT/NF-κB inhibition. Neurosci Lett 2015, 609, 129–136. [Google Scholar] [CrossRef]
- Qin, S.; Chen, J.; Tanigawa, S.; Hou, D.X. Microarray and pathway analysis highlight Nrf2/ARE-mediated expression profiling by polyphenolic myricetin. Mol Nutr Food Res 2013, 57, 435–446. [Google Scholar] [CrossRef]
- Liao, H.H.; Zhu, J.X.; Feng, H.; Ni, J.; Zhang, N.; Chen, S.; Liu, H.J.; Yang, Z.; Deng, W.; Tang, Q.Z. Myricetin possesses potential protective effects on diabetic cardiomyopathy through inhibiting IκBα/NFκB and enhancing Nrf2/HO-1. Oxid Med Cell Longev 2017, 2017, 8370593. [Google Scholar] [CrossRef]
- Ma, C.; Deng Y, Xiao R, Xu, F. ; Li, M.; Gong, Q.; Gao, J.. Anti-fatigue effect of phlorizin on exhaustive exercise-induced oxidative injury mediated by Nrf2/ARE signaling pathway in mice. Eur J Pharmacol 2022, 918, 174563. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Chen, Z.; Huang, L.; Meng, B.; Zhou, X.; Wen, X.; Ren, D. Naringenin reduces oxidative stress and improves mitochondrial dysfunction via activation of the Nrf2/ARE signaling pathway in neurons. Int J Mol Med 2017, 40, 1582–1590. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Li, J.; Zha, D.; Zhang, L.; Gao, P.; Yao, T.; Wu, X. Chlorogenic acid prevents diabetic nephropathy by inhibiting oxidative stress and inflammation through modulation of the Nrf2/HO-1 and NF-ĸB pathways. Int Immunopharmacol 2018, 54, 245–253. [Google Scholar] [CrossRef]
- Yeh, C.T.; Yen, G.C. Involvement of p38 MAPK and Nrf2 in phenolic acid-induced P-form phenol sulfotransferase expression in human hepatoma HepG2 cells. Carcinogenesis 2006, 27, 1008–1007. [Google Scholar] [CrossRef]
- Ji, L.L. Exercise-induced modulation of antioxidant defense. Ann N Y Acad Sci 2002, 959, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Jinhwan, L.; Ulrike, L. Oxidative damage increases and antioxidant gene expression decreases with aging in the mouse ovary. Biol Reprod 2011, 84, 775–782. [Google Scholar]
- Rao, G.; Xia, E.; Nadakavukaren, M.J.; Richardson, A. Effect of dietary restriction on the age-dependent changes in the expression of antioxidant enzymes in rat liver. J Nutr 1990, 120, 602–609. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alexandria Med J 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Rosado-Pérez, J.; Aguiñiga-Sánchez, I.; Santiago-Osorio, E, Mendoza-Núñez, V. M. Effect of Sechium edule var. nigrum spinosum (Chayote) on oxidative stress and pro-inflammatory markers in older adults with metabolic syndrome: an exploratory study. Antioxidants 2019, 8, 146. [Google Scholar]
- Gavia-García, G.; Rosado-Pérez, J.; Aguiñiga-Sánchez, I.; Santiago-Osorio, E.; Mendoza-Núñez, V.M. Effect of Sechium edule var. nigrum spinosum (chayote) on telomerase levels and antioxidant capacity in older adults with metabolic syndrome. Antioxidants 2020, 9, 634. [Google Scholar]
- Oh-Ishi, S.; Kizaki, T.; Yamashita, H.; Nagata, N.; Suzuki, K.; Taniguchi, N.; Ohno, H. Alterations of superoxide dismutase iso-enzyme activity, content, and mRNA expression with aging in rat skeletal muscle. Mech Ageing Dev 1995, 84, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Marasco, M.R.; Conteh, A.M.; Reissaus, C.A.; Cupit, J.E. 5th.; Appleman, E.M.; Mirmira, R.G.; Linnemann, A.K. Interleukin-6 reduces β-cell oxidative stress by linking autophagy with the antioxidant response. Diabetes 2018, 67, 1576–1588. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, Y.; Nakayama, H.; Yoshida, R.; Hirosue, A.; Nagata, M.; Tanaka, T.; Kawahara, K.; Sakata, J.; Arita, H.; Nakashima, H.; Shinriki, S.; Fukuma, D.; Ogi, H.; Hiraki, A.; Shinohara, M.; Toya, R.; Murakami, R. IL-6 controls resistance to radiation by suppressing oxidative stress via the Nrf2-antioxidant pathway in oral squamous cell carcinoma. Br J Cancer 2016, 115, 1234–1244. [Google Scholar] [CrossRef] [PubMed]
- Wruck, C.J.; Streetz, K.; Pavic, G.; Götz, M.E.; Tohidnezhad, M.; Brandenburg, L.O.; Varoga, D.; Eickelberg, O.; Herdegen, T.; Trautwein, C.; Cha, K.; Kan, Y.W.; Pufe, T. Nrf2 induces interleukin-6 (IL-6) expression via an antioxidant response element within the IL-6 promoter. J Biol Chem 2011, 286, 4493–4499. [Google Scholar] [CrossRef]
- Mohamed, G.A.; Ibrahim, S.R. , El-Agamy, D.S.; Elsaed, W.M.; Sirwi, A.; Asfour, H.Z.; Elhady, S.S. Cucurbitacin E glucoside alleviates concanavalin A-induced hepatitis through enhancing SIRT1/Nrf2/HO-1 and inhibiting NF-ĸB/NLRP3 signaling pathways. J Ethnopharmacol 2022, 292, 115223. [Google Scholar] [CrossRef]
- Li, J.; Mao, B.; Tang, X.; Zhang, Q.; Zhao, J.; Zhang, H.; Cui, S. Protective effects of naringenin and apigenin in ameliorating skin damage via mediating the Nrf2 and NF-κB pathways in mice. Foods 2023, 12, 2120. [Google Scholar] [CrossRef]
- Lee, Y.; Shin, D.H.; Kim, J.H.; Hong, S.; Choi, D.; Kim, Y.J.; Kwak, M.K.; Jung, Y. Caffeic acid phenethyl ester-mediated Nrf2 activation and IκB kinase inhibition are involved in NFκB inhibitory effect: structural analysis for NFκB inhibition. Eur J Pharmacol 2010, 643, 21–28. [Google Scholar] [CrossRef]
- Lampiasi, N.; Montana, G. An in vitro inflammation model to study the Nrf2 and NF-κB crosstalk in presence of ferulic acid as modulator. Immunobiology 2018, 223, 349–355. [Google Scholar] [CrossRef]
- Paredes-Gonzalez, X.; Fuentes, F.; Su, Z.Y.; Kong, A.N.T. Apigenin reactivates Nrf2 anti-oxidative stress signaling in mouse skin epidermal JB6 P+ cells through epigenetics modifications. AAPS J 2014, 16, 727–735. [Google Scholar] [CrossRef]
- Wang, X.; Sun, Y.; Li, P.; Wu, Z.; Chen, Y.; Fu, Y.; Wu, H.; Ye, Y.; Wang, J.; Yang, Z.; Zhou, E. The protective effects of myricetin against acute liver failure via inhibiting inflammation and regulating oxidative stress via Nrf2 signaling. Nat Prod Res 2023, 37, 798–802. [Google Scholar] [CrossRef]
- Chi-Tai, Y.; Gow-Chin, Y. Involvement of p38 MAPK and Nrf2 in phenolic acid-induced P-form phenol sulfotransferase expression in human hepatoma HepG 2 cells. Carcinogenesis 2006, 27, 1008–1017. [Google Scholar]
- Granado-Serrano, A.B.; Martín, M.A.; Bravo, L.; Goya, L.; Ramos, S. Quercetin modulates Nrf2 and glutathione-related defenses in HepG2 cells: Involvement of p38. Chem Biol Interact 2012, 195, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Y.; Guo, Y.; Xu, L.; Wang, H. Phlorizin exerts potent effects against aging induced by D-galactose in mice and PC12 cells. Food Funct 2021, 12, 2148–2160. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, S.; Zhao, A.; Mi, Y.; Zhang, C. Protective effect of rutin on ferroptosis-induced oxidative stress in aging laying hens through Nrf2/HO-1 signaling. Cell Biol Int 2023, 47, 598–611. [Google Scholar] [CrossRef] [PubMed]
- Shi, A.; Shi, H.; Wang, Y.; Liu, X.; Cheng, Y.; Li, H.; Zhao, H.; Wang, S.; Dong, L. Activation of Nrf2 pathway and inhibition of NLRP3 inflammasome activation contribute to the protective effect of chlorogenic acid on acute liver injury. Int Immunopharmacol 2018, 54, 125–130. [Google Scholar] [CrossRef]
- Varì, R.; D’Archivio, M.; Filesi, C.; Carotenuto, S.; Scazzocchio, B.; Santangelo, C.; Giovannini, C.; Masella, R. Protocatechuic acid induces antioxidant/detoxifying enzyme expression through JNK-mediated Nrf2 activation in murine macrophages. J Nutr Biochem 2011, 22, 409–417. [Google Scholar] [CrossRef]
- Shen, Y.; Song, X.; Li, L.; Sun, J.; Jaiswal, Y.; Huang, J.; Li, C.; Yang, W.; Williams, L.; Zhang, H.; Guan, Y. Protective effects of p-coumaric acid against oxidant and hyperlipidemia-an in vitro and in vivo evaluation. Biomed Pharmacother 2019, 111, 579–587. [Google Scholar] [CrossRef]
Figure 1.
Relative expression of the mRNA of genes that code for proteins that participate in processes of antioxidant protection and inflammation, among the study subjects. Data are expressed as means ± standard deviation. ANOVA of repeated measures test, significance level 95%, p < 0.05. SOD, superoxide dismutase; GPx, glutathione peroxidase; CAT, catalase; Nrf2, nuclear factor (erythroid-derived 2)-like 2; IL-6, interleukin-6; IL-8, interleukin-8; TNF-α, tumor necrosis factor-alpha; NFκB p50, Nuclear Factor kappa B p50; NFκB p65, Nuclear Factor kappa B p65. The mRNA relative expression levels were determined after normalization against β-actin.
Figure 1.
Relative expression of the mRNA of genes that code for proteins that participate in processes of antioxidant protection and inflammation, among the study subjects. Data are expressed as means ± standard deviation. ANOVA of repeated measures test, significance level 95%, p < 0.05. SOD, superoxide dismutase; GPx, glutathione peroxidase; CAT, catalase; Nrf2, nuclear factor (erythroid-derived 2)-like 2; IL-6, interleukin-6; IL-8, interleukin-8; TNF-α, tumor necrosis factor-alpha; NFκB p50, Nuclear Factor kappa B p50; NFκB p65, Nuclear Factor kappa B p65. The mRNA relative expression levels were determined after normalization against β-actin.
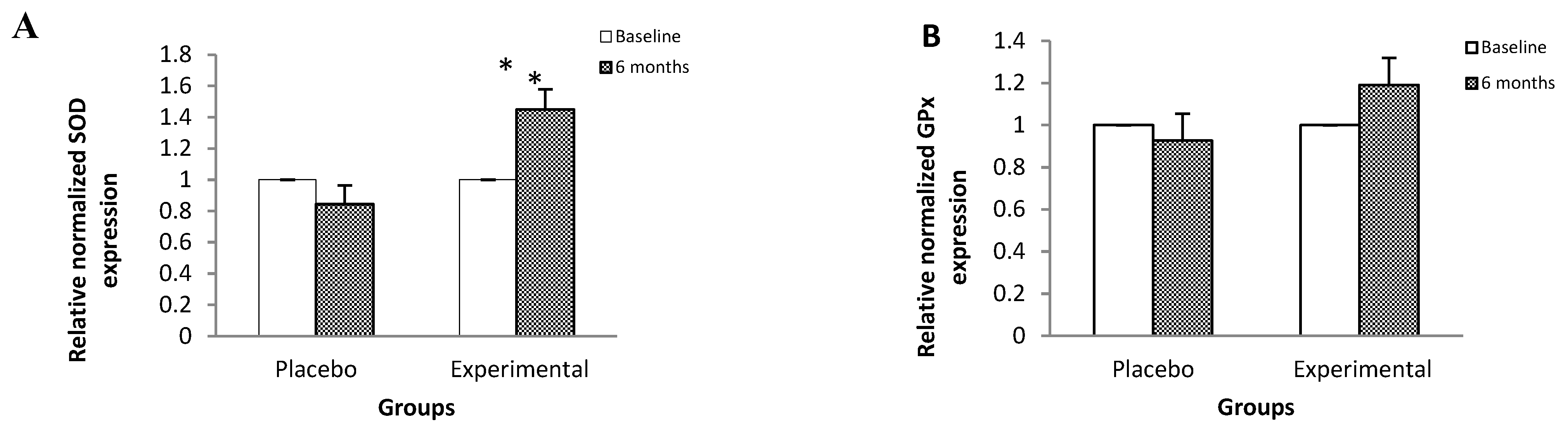
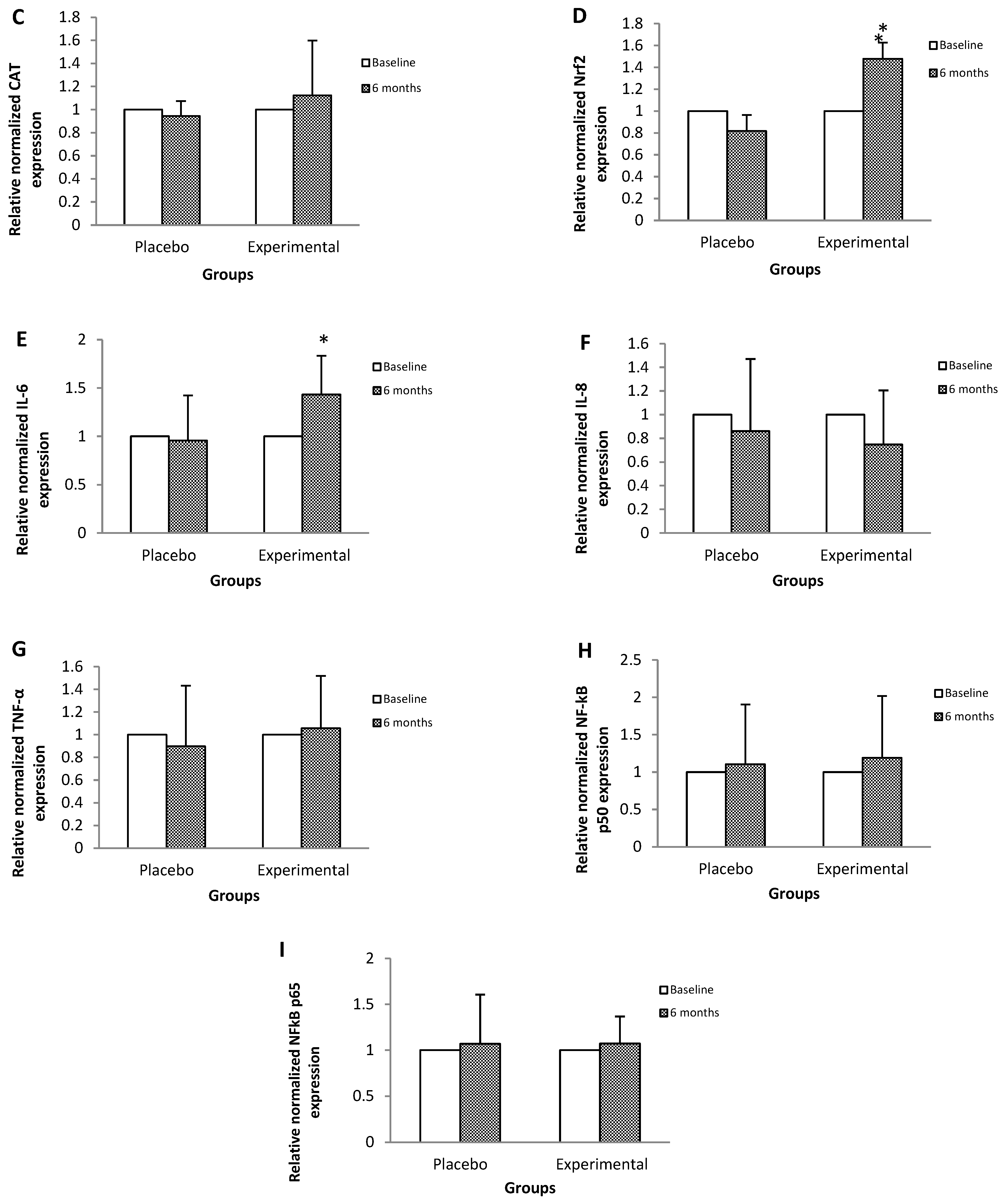

Table 1.
Detailed primers used for real-time PCR assays.
| Gene | Primer Name |
Primer sequence |
|---|---|---|
| SOD1 | SOD-F | GGTGGGCCAAAGGATGAAGA |
| SOD-R | ATAGACACATCGGCCACACC | |
| GPX1 | GPX-F | ACACCCAGATGAACGAGCTG |
| GPX-R | CTTCGTTCTTGGCGTTCTCC | |
| CAT | CAT-F | TGAAGATGCGGCGAGACTTT |
| CAT-R | GAGGGGTACTTTCCTGTGGC | |
| NFE2L2 | NRF2-F | AGGTTGCCCACATTCCCAAA |
| NRF2-R | ACGTAGCCGAAGAAACCTCA | |
| IL-6 | IL6-F | CCACCGGGAACGAAAGAGAA |
| IL6-R | GAGAAGGCAACTGGACCGAA | |
| IL-8 | IL8-F | GAAGAGAGCTCTGTCTGGACC |
| IL8-R | TGAATTCTCAGCCCTCTTCAAAAAC | |
| TNF-α | TNF-F | GAAGAGAGCTCTGTCTGGACC |
| TNF-R | TGAATTCTCAGCCCTCTTCAAAAAC | |
| NFKB-p50 | NFKBP50-F | GGAGGCCGAACGCGG |
| NFKBP50-R | AAACATTTGTTCAGGCCTTCCC | |
| NFKB-p65 | NFKB65-F | CAGTGTGTGAAGAAGCGGGA |
| NFKB65-R | CCACGCTGCTCTTCTTGGAA | |
| β-ACTIN | ACTIN-F | GAGCACAGAGCCTCGCC |
| ACTIN-R | CGCGGCGATATCATCATCCA |
SOD1, copper/zinc-superoxide dismutase; GPX1, glutathione peroxidase 1; CAT, catalase; NFE2L2, nuclear factor, erythroid 2-like 2; IL-6; interleukin 6, IL-8, interleukin 8; TNF-α, tumor necrosis factor-alpha; NFκB-p50, nuclear factor kappa B subunit-p50; NFκB-p65, nuclear factor kappa B subunit-p65.
Table 2.
Clinical and anthropometric measurements by study group.
| Placebo n = 20 |
Experimental n =26 |
||
|---|---|---|---|
| Parameter | p-Value | ||
| Age (years) | 68.4 ± 5.7 | 67.9 ± 6.6 | |
| SBP (mmHg) | |||
| Baseline | 132.5 ± 15.5 | 141.4 ± 12.1 | |
| 6 months | 132.3 ± 12.1 | 131.2 ± 11.2* | 0.01 |
| DBP (mmHg) | |||
| Baseline | 90.2 ± 9.9 | 95.0 ± 9.8 | |
| 6 months | 85.6 ± 6.4 | 83.8 ± 8.0* | 0.001 |
| Weight (kg) | |||
| Baseline | 76.5 ± 14.6 | 74.4 ± 16.5 | |
| 6 months | 76.2 ± 12.8 | 72.4 ± 16.4* | 0.01 |
| Waist circumference (cm) | |||
| Baseline | 103.6 ± 11.4 | 99.9 ± 12.2 | |
| 6 months | 104.2 ± 11.9 | 102.1 ± 12.1 | 0.55 |
TThe data are expr The data are expressed as the average ± standard deviation. ANOVA of repeated measures, significance level 95%, p < 0.05. SBP, systolic blood pressure; DBP, diastolic arterial pressure. *Baseline statistical significance vs 6 months inter-group.
Table 3.
Pre and post-treatment biochemical parameters by study group.
| Placebo n = 20 |
Experimental n = 26 |
||
|---|---|---|---|
| Parameter | p-Value | ||
| Glucose [mg/dL] | |||
| Baseline | 137.3 ± 61.3 | 140.7 ± 48.1 | |
| 6 months | 139.4 ± 69.1 | 140.0 ± 57.3 | 0.91 |
| HDL-c [mg/dL] | |||
| Baseline | 47.8 ± 9.5 | 42.8 ± 7.5 | |
| 6 months | 47.4 ± 7.5 | 47.4 ± 7.4* | 0.01 |
| Triglycerides [mg/dL] | |||
| Baseline | 169.6 ± 36.6 | 174.1 ± 54.8 | |
| 6 months | 125.7 ± 30.4 | 149.2 ± 45.6 | 0.19 |
The data are expressed as the mean ± standard deviation. ANOVA of repeated measures, significance level 95%, p < 0.05. HDL-c, high-density lipoprotein colesterol. *Baseline statistical significance vs 6 months inter-group.
Table 4.
Antioxidant capacity and oxidative state by study group.
| Placebo n = 20 |
Experimental n = 26 |
||
|---|---|---|---|
| Parameter | p-Value | ||
| SOD [U/mL] | |||
| Baseline | 176.8 ± 10.3 | 167.1 ± 11.9 | |
| 6 months | 174.6 ± 4.9 | 180.6 ± 7.6* | 0.05 |
| GPx [U/L] | |||
| Baseline | 5740 ± 938 | 5221 ± 822 | |
| 6 months | 5023 ± 1885 | 5559 ± 2007 | 0.33 |
| CAT [U/mL] | |||
| Baseline | 1.2 ± 0.1 | 1.0 ± 0.2 | |
| 6 months | 1.2 ± 0.2 | 1.3 ±0.2* | 0.01 |
| TOS [µmol H2O2 Equiv./L] | |||
| Baseline | 24.5 ± 2.3 | 28.9 ± 3.6 | |
| 6 months | 28.0 ± 3.7 | 23.7 ± 3.4* | 0.01 |
| TAS [mmol/L] | |||
| Baseline | 1.2 ± 0.1 | 11.1 ± 0.1 | |
| 6 months | 1.1 ± 0.2 | 1.4 ± 0.1* | 0.01 |
| OSI | |||
| Baseline | 19.2 ± 2.1 | 24.1 ± 3.8 | |
| 6 months | 26.1 ± 7.0 | 17.7 ± 4.0* | 0.01 |
The data are expressed as the mean ± standard deviation. ANOVA of repeated measures, significance level 95%, p < 0.05. SOD, superoxide dismutase; GPx, glutathione peroxidase; CAT, catalase; TOS, total oxidizing state; TAS, total antioxidant status; OSI, oxidative stress index (TOS/TAS). *Baseline statistical significance vs 6 months inter-group
Table 5.
Pre and post-treatment inflammatory markers by study group.
| Placebo n = 20 |
Experimental n = 26 |
||
|---|---|---|---|
| Parameter | p-Value | ||
| IL-6 [pg/dL] | |||
| Baseline | 10.7 ± 2.1 | 10.7 ± 1.1 | |
| 6 months | 11.0 ± 0.9 | 12.3 ± 2.0* | 0.03 |
| IL-8 [pg/dL] | |||
| Baseline | 27.6 ± 4.4 | 37.7 ± 9.9 | |
| 6 months | 25.8 ± 6.0 | 30.9 ± 11.1 | 0.12 |
| TNF-α [pg/dL] | |||
| Baseline | 8.4 ± 1.2 | 8.2 ± 0.6 | 0.61 |
| 6 months | 8.8 ± 1.6 | 9.2 ± 1.1 |
Data are expressed as means ± standard deviation. ANOVA of repeated measures test, significance level 95%, p < 0.05. IL-6, interleukin-6; IL-8, interleukin-8; TNF-α, tumor necrosis factor-alpha. *Baseline statistical significance vs 6 months inter-group.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



