
Submitted:
30 August 2023
Posted:
31 August 2023
You are already at the latest version
Abstract
Flaxseed has been recognized as a valuable source of nutrients and bioactive compounds, including proteins that possess various health benefits. In recent years, studies have shown that flaxseed proteins, including albumins, globulins, glutelin, and prolamins, possess anti-cancer properties. These properties are attributed to their ability to inhibit cancer cell proliferation, induce apoptosis, and interfere with cancer cell signaling pathways, ultimately leading to the inhibition of metastasis. Moreover, flaxseed proteins have been reported to modulate cancer cell mechanobiology, leading to changes in cell behavior and reduced cancer cell migration and invasion. This review provides an overview of the anti-cancer properties of flaxseed proteins, with a focus on their potential use in cancer treatment. Additionally, it highlights the need for further research to fully establish the potential of flaxseed proteins in cancer therapy.
Keywords:
metastasis
; flaxseed proteins
; cancer treatment
; radiotherapy
; mechanobiology
; proteoform level analysis
; flaxseed amino-acids
Introduction
Cancer is the second leading cause of mortality worldwide. The majority of cancer-related deaths (90%) are attributed to metastasis, making its reduction a critical factor in improving patients’ survival rates [1]. As metastases from primary tumors often occur in sites that are not easily amenable to surgery, such as bones and the brain, chemotherapy remains the primary therapeutic option. However, conventional chemotherapy is associated with significant side effects due to its impact on all fast-proliferating cells.
One broadly utilized indicator of cancer's propensity to metastasize is cell invasiveness. During the invasion to the surrounding tissues, cancer cells are able rapidly change their shape and apply forces to facilitate the invasion process. Cells with high metastatic potential (MP) are typically more receptive to internalizing materials from their surroundings, displaying a more dynamic intercellular composition than benign or low MP cells. The cytoskeleton of highly metastatic cells is denser [2,3,4]. While common chemotherapeutics primarily target cancer cell proliferation and growth, addressing metastatic spread requires more intricate and innovative approaches, such as radiotherapy. Anti-metastatic drugs aim to inhibit processes like vascularization, microenvironment-dependent cell growth, cell binding to the extracellular matrix (ECM), and stem-like properties [5,6]. In clinical cases where metastasis is expected, an aggressive treatment strategy combining tumor growth and metastasis targeting is typically employed. Unfortunately, these aggressive treatments often compromise patients' quality of life and increase the risk of kidney or liver-related toxicity [7], and in some cases, inappropriate treatment might even elevate the chance of metastasis development. Thus, accurate assessment of metastatic risk, followed by the identification of specific personalized treatments that can potentially reduce or inhibit metastasis formation, holds paramount importance.
Treatment with flaxseed proteins may induce changes in cytoskeleton dynamics that influence a cell's ability to change shape and apply force, affecting its invasive capacity. During penetration through the extracellular matrix (ECM) and endothelial transmigration (intravasation/extravasation), invading cancer cells must alter their morphology based on the local microenvironment conditions. Leading cells often display pseudopods and filament-based protrusions, aiding them in navigating through narrow pores [8,9]. The mechanobiological response to chemotherapy, coupled with changes in cell viability, dictates the treatment's success and overall effectiveness [10].
According to reports from the World Health Organization (WHO) [11,12], unhealthy eating habits and lack of physical activity contribute to about 30% of cancer-related deaths. Approximately one-third of diagnosed cancers could be prevented or mitigated by promoting physical activity and adopting healthier dietary habits. Various studies, including those conducted by the European Medicines Agency, support the potential benefits of plants used in traditional cancer medicines. Flax, a globally significant agricultural crop, is abundant in bioactive molecules that may possess anti-cancer properties. The biological activity of dietary proteins often leads to the development of bioactive peptides released during digestion in the gastrointestinal tract. These peptides can enter the bloodstream by crossing the digestive epithelial barrier, enabling them to reach distant organs and provide beneficial effects for the organism [13].
Flaxseeds proteins
Generally, whole flaxseed comprises 30–41% fat, 20–35% dietary fiber, 20–30% protein, 4–8% moisture, 3–4% ash, and 1% simple sugars [14]. Recently, researchers have begun exploring the anti-cancer properties of flaxseed proteins, uncovering various ways in which they may exhibit such properties. For instance, flaxseed stands out as a significant dietary source of lignans, a type of phytoestrogen recognized for its anti-cancer attributes [15]. Lignans have been found to inhibit the growth of hormone-sensitive cancers, such as breast and prostate cancers, by interfering with the effects of estrogen on these tissues [16]. Additionally, flaxseed offers an abundant supply of alpha-linolenic acid—an omega-3 fatty acid. Omega-3 fatty acids are known for their anti-inflammatory effects, potentially reducing cancer risk [17]. Moreover, there is speculation that omega-3 fatty acids could hinder or limit the proliferation and dissemination of certain types of cancer cells [18]. The rich antioxidant content in flaxseed contributes to cellular protection against damage induced by free radicals [19,20]. Free radicals, unstable molecules, can induce cellular damage and escalate cancer risk. Certain studies have even indicated that flaxseed proteins might possess anti-angiogenic properties, impeding the formation of new blood vessels essential for tumor growth [21].
The composition of flaxseed proteins can fluctuate based on factors such as flax variety, growth conditions, and processing techniques. However, on average, flaxseed proteins comprise roughly 15-25% albumins, 43-80% globulins, 4-10% prolamins, and up to 10% other proteins [22,23,24,25,26,27,28,29], as illustrated in Figure 1.
The level and composition of flaxseed proteome depends on many factors, for example cultivars, environmental conditions, and processing methods [28]. Flaxseed proteins mainly consists of 11S globulin and 2S albumin (Table 1). The 11S globulin is a salt-soluble protein, it has high molecular weight (252-298 kDa). The 2S albumin is a water-soluble protein, it has low molecular weight (16-17 kDa) [24,28,30].
Albumins of flaxseed
Albumin, a type of protein present in various foods such as egg whites and whey protein, is recognized for its water-soluble nature and high digestibility, making it a valuable source of essential amino acids. While flaxseed albumin has not been as extensively studied as other components like lignans or omega-3 fatty acids, there exists some evidence to suggest its potential anti-cancer properties. Functioning as a potent antioxidant, albumin aids in neutralizing free radicals—unstable molecules responsible for cellular damage and heightened cancer risk [19]. Furthermore, albumin plays a pivotal role in immune system functionality, safeguarding against endothelial dysfunction through immunomodulation and antioxidant mechanisms [47]. Diminished albumin levels could potentially trigger inflammation and an increase in leukocyte count [48]. As a significant source of amino acids, albumin contributes to essential protein synthesis required for healthy cell growth and division; deficiencies in this synthesis have been correlated with an elevated cancer risk [49,50]. Albumin is also responsible for transporting vital nutrients, including vitamins and minerals, throughout the body [51], which is crucial for maintaining a robust immune system and minimizing cancer risk. Beyond these roles, albumin can serve as a carrier for chemotherapy drugs, enhancing their targeted delivery to tumor sites and potentially lowering metastasis risk [52].
The extracellular matrix (ECM), a complex network of proteins and molecules surrounding cells, plays a pivotal role in cancer progression and metastasis. Evidence indicates that albumin interacts with and regulates ECM components, potentially inhibiting cancer cell invasion and metastasis. Albumin's interactions encompass key proteins involved in cancer cell invasion, like matrix metalloproteinases (MMPs) and urokinase-type plasminogen activator (uPA) [53,54]. Moreover, albumin can engage with other ECM proteins, such as laminin and fibronectin, contributing to the inhibition of cancer cell invasion [55,56,57].
The primary form of albumin in flaxseed is 2S albumin, a seed storage protein. Although the precise physiological and metabolic role of 2S albumins remains to be definitively described, evidence based on their amino acid composition and mobilization during germination suggests their function as nitrogen and sulfur donors [61]. Typically existing as heterodimers, these proteins consist of 8–16 kDa water-soluble polypeptides [58], connected by 2 disulfide bonds resistant to pepsin and trypsin. While the subunit compositions and structures of 2S albumins differ, their 3D form is generally a compact sphere enriched in α-helices [59,60]. An extensively studied anti-cancer peptide derived from plants is lunasin, a small peptide from the 2S albumin family, containing 43-44 amino acid residues [36]. Lunasin encompasses multiple functional domains, including an aspartic acid tail, an RGD domain, and a chromatin-binding helical domain [62,63]. This protein holds the potential to hinder cancer cell invasion and migration (Table 2) and represents a promising avenue for further research in developing novel cancer treatments.
Overall, while further research is required to comprehensively comprehend the potential anti-cancer effects of 2S albumin, this protein shows promise in potentially inhibiting cancer cell invasion and migration. Thus, it stands as a promising realm of investigation for the advancement of novel cancer treatments.
Globulins of flaxseed
Globulins form a diverse family of proteins present in various foods, including legumes, nuts, and animal products. Among flaxseed proteins, globulin takes precedence as the principal component, its size being reported in the range of 252-298 kDa (for 11-12 S Globulins). Comprising about 3% α-helical and 17% β-structures [75,76,77], globulins, akin to albumins, have not undergone extensive examination for their anti-cancer attributes. Nevertheless, some evidence indicates the potential anti-cancer properties of globulins.
Globulins play a pivotal role in the immune system, actively combating infections and diseases, including cancer. There's a suggestion that globulins might stimulate white blood cell production, contributing to the battle against cancer cells [78]. Certain globulins have been shown to impede the activity of enzymes linked to tumor growth and metastasis. For instance, soybean globulins have been observed to hinder the action of tyrosine kinase, an enzyme central to cancer cell proliferation [79,80]. Certain globulins, like whey protein, boast significant antioxidant content that aids in neutralizing free radicals and guarding against cellular damage that could lead to cancer [81]. Moreover, globulins partake in regulating hormone levels within the body [82,83,84]. Given that hormones can influence the development and progression of specific cancers, modulating their levels might yield anti-cancer effects as well.
Predominantly, flaxseed's major globulin type is the 11S globulin (comprising over 85% of all globulins), while the 7S (less than 2%) and 2S Vicilin-like globulins (less than 4%) are considered minor [85]. The non-reduced 11S globulin reveals five polypeptide bands, including a basic subunit (18-20 kDa), an acidic subunit (30-40 kDa), and additional polypeptides with molecular weights of 47, 80, 120, and 160 kDa. Studies exploring the distinct biological properties of flaxseed protein digests have yielded varied results, contingent on the methods employed. For example, the highest antioxidant activity (90%) and effective fungal inhibition were observed with glutelin hydrolysates, whereas intact glutelin protein exhibited the highest angiotensin-converting enzyme inhibition (60%) activity [25]. There's some evidence suggesting that 11S globulin (also referred to as glutelin [86]), a major storage protein in flaxseed, might possess anti-cancer properties, potentially inhibiting cancer cell proliferation and inducing apoptosis (Table 3). However, it's crucial to acknowledge that these investigations were conducted in vitro (within a laboratory setting using cell cultures), and further research is necessary to ascertain the potential effects of glutelin in vivo (within living organisms).
The proteoform level analysis of major proteins from flaxseed
While studies on 2S albumin have focused on its characterization, structural properties, allergenicity, and functional aspects, detailed proteoform-level analysis has been somewhat limited in comparison. Nevertheless, certain studies underscore the significance of proteoform-level analysis in comprehending the structural diversity, post-translational modifications (PTMs), and functional attributes of 11S globulins sourced from different plants.
For instance, research delving into proteoform-level analysis of soybean 11S globulin, referred to as glycinin [94], and peanut 11S globulin, known as arachin, was conducted using mass spectrometry-based proteomics [95]. For glycinin, various proteoforms exhibiting distinct PTMs—like phosphorylation, acetylation, methylation, and disulfide bond formation—were identified through two-dimensional gel electrophoresis (2DE) and mass spectrometry [96]. Similarly, multiple proteoforms of lupin 11S globulin, characterized by varying isoelectric points (pI) and molecular weights, were detected [97]. These lupin 11S globulin proteoforms displayed differences in solubility, digestion patterns, and IgE-binding capacity, hinting at potential implications in terms of allergenicity [98].
Extraction and characterization of flaxseed`s amino acids
The isolation and characterization of amino acids from flaxseed proteins generally involve a series of steps, including protein extraction, hydrolysis, and analysis (Figure 2). Flaxseed proteins are initially extracted from the seeds using appropriate extraction buffers or solvents. Common methods encompass saline extraction, phosphate buffer extraction, or sodium dodecyl sulfate (SDS) extraction [99,100,101]. These methods aid in solubilizing and separating proteins from other seed components. Following successful extraction, hydrolysis is conducted to disintegrate the proteins into individual amino acids. Two frequently employed techniques are acid hydrolysis and enzymatic hydrolysis. Acid hydrolysis involves treating the protein sample with controlled conditions using hydrochloric acid (HCl) or sulfuric acid (H2SO4) [98]. On the other hand, enzymatic hydrolysis employs proteolytic enzymes like trypsin, chymotrypsin, or pepsin to cleave proteins into smaller peptide fragments and eventually amino acids [102,103,104]. The hydrolyzed amino acids are subsequently quantified and characterized using diverse analytical methods. The primary technique is high-performance liquid chromatography (HPLC) coupled with UV detection or fluorescence detection [105]. Amino acids are separated based on their chemical properties, such as polarity, using a suitable chromatographic column and elution gradient. The resulting peaks corresponding to individual amino acids are identified and quantified with the aid of calibration curves created using amino acid standards. To verify the identity of the amino acids, mass spectrometry (MS) can be employed [96].
Gathering both quantitative and qualitative insights into the amino acid composition of flaxseed proteins holds considerable value in comprehending their nutritional quality, functional attributes, and potential applications in cancer treatment. It's important to acknowledge that the amino acid makeup within flaxseed proteins can vary based on the specific flaxseed variety and the analytical methods employed for amino acid analysis (Table 4).
In the realm of cancer therapy, specific amino acids can serve as supplements or be incorporated into treatment strategies to bolster efficacy and alleviate treatment-related side effects. Notably, glutamic acid, a predominant amino acid found in flaxseed proteins, exhibits anticancer properties and has even been proposed as an anticancer agent [106]. Due to its adept penetration capability, high compatibility, and low immunogenicity, glutamic acid has been regarded as a drug nano-delivery platform [107]. A combination of aspartic and glutamic acids has been shown to inhibit tumor cell proliferation, induce tumor cell death, and hold substantial promise for clinical utilization as an anti-cancer agent [108]. Modifying the widely accepted anti-tumor treatment with peptides containing arginine, glycine, and aspartic acid has demonstrated a noteworthy antiproliferative impact [109]. Arginine, known for its immunostimulatory effect [110], has been extensively explored as a cell-penetrating agent for drug delivery in the form of arginine-containing peptides [111,112]. Additionally, isoleucine, when administered at high doses, has been observed to suppress the proliferation of breast and lung cancer cells [113]. While leucine is not typically employed as a direct cancer treatment, it assumes pivotal roles in diverse metabolic processes and might bear implications for cancer metabolism and treatment strategies.

Table 4.
Major amino-acids of flaxseed proteins. The schematic structure prepared using @3dchem.com.
Table 4.
Major amino-acids of flaxseed proteins. The schematic structure prepared using @3dchem.com.
| Amino Acid | Description | Structure | Composition from flaxseed proteins |
| Glutamic Acid | An aliphatic, acidic, conditionally non-essential, crystalline α-amino acid | 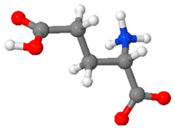 |
19-27 %[114,115,116] |
| Aspartic Acid | An aliphatic, acidic, non-essential, crystalline α-amino acid | 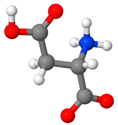 |
8-21 % [117,118,119] |
| Arginine | An aliphatic, non-aromatic, conditionally non-essential basic/cationic amino acid | 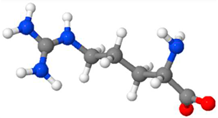 |
8-12 % [120,121,122] |
| Isoleucine | An aliphatic, non-polar, essential, crystalline α-amino acid | 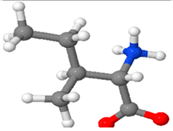 |
4-8 %[118,123,124] |
| Leucine | An aliphatic, non-polar, essential, crystalline α-amino acid | 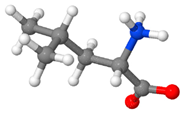 |
4-7 %[116,117,119,121,124] |
Presence of flaxseed proteome in databases
Despite the evident significance of flaxseed proteins as potential anti-cancer agents, their comprehensive study and description remain somewhat lacking when compared to proteins from other plant sources. For instance, Linum usitatissimum (Flax) has a total of 936 entries within the UniProt database, with merely 11 of these entries reviewed by the Swiss-Prot database. In contrast, Glycine max (Soybean), another prominent seed source, boasts a staggering 84,712 entries in the UniProt database, with 431 of these entries reviewed by Swiss-Prot. Additionally, while the proteomes of soybeans have been extensively described and documented in protein databases, the proteomes of flax are notably absent from these databases.
Among the few flax proteins covered by UniProt databases and reviewed by Swiss-Prot are those involved in diverse functions such as lignan biosynthesis, disease resistance against pathogens, enzymatic activities in the catabolism of cyanogenic glycosides, protease inhibition, and more. These proteins play roles in biosynthesis of secondary metabolites (59%), conferring resistance to pathogens (33%), and are rarely employed in biotechnological applications (8%).
The existing NLM database mainly highlights protein families like Glycosyltransferase 2, DEAD box helicase, LTN1, SCAMP, RNA polymerase alpha/beta chains, ATPase alpha/beta chains, Peroxidase, among others. However, limited information is available about the core components of the flaxseed proteome (Table 1), especially when compared to the diversity of protein coverage for various other plants (Table 5).
While specialized databases like the Plant Proteome Database (developed by ©Klaas J. van Wijk Lab, Cornell University) exist for plants such as Arabidopsis, maize, and rice, there is currently no such database dedicated to flax. Individual databases have been established for specific plant species including Arabidopsis (TAIR database), maize (maizeGDB), tomatoes (TOMATOMICS), rice (Rice genome annotation project database), soybean (SoyBase), and wheat (Wheat proteome database) [125], but a dedicated database for flax is notably absent.
The sequencing and profiling of flaxseed proteins
Protein sequencing is a pivotal process that involves determining the exact sequence of amino acids constituting a protein. This step is indispensable for comprehending the protein's structure, function, and potential applications. However, the sequences and comprehensive profiles of flaxseed proteins have yet to be documented in databases. Nonetheless, significant advancements have been made in sequencing the main proteins of flaxseed, such as 2S-albumin and 11S-globulin, in comparison to their counterparts from other plant sources.
Figure 3 demonstrates a remarkable degree of similarity in protein profiles across various plant sources. This suggests that computational models could potentially be employed to predict the typical structures and sequences of the primary flaxseed proteins. While some progress has been made in profiling and sequencing flaxseed proteins, it's worth noting that only 47 proteins have been identified using the BlastP database, with sequence coverage ranging from 6% to 49% [126].
Further exploration and comprehensive profiling of flaxseed proteins, including a focus on post-translational modifications, will undoubtedly contribute to a better understanding of protein functionality, stability, and their potential applications in anti-cancer strategies.
The combined anti-cancer action of flaxseed proteins
Flaxseed proteins can be consumed as part of the diet and subsequently absorbed within the digestive tract. Once absorbed, they can enter the bloodstream and circulate throughout the body, potentially reaching cancer cells. There's also the possibility of delivering flaxseed proteins directly to cancer cells through targeted drug delivery systems or nanoparticle formulations. Moreover, the topical application of flaxseed oil, which contains certain proteins present in flaxseed, has been explored as a potential method for delivering these proteins to skin cancer cells [127,128].
The efficacy of flaxseed proteins in various cancer cell lines, such as breast [129], skin [62], prostate, and colon [65] cancer cells, has been demonstrated. Flaxseed proteins have exhibited anti-tumor activity by inducing cell cycle arrest and promoting apoptosis (programmed cell death) [23]. Notably, flaxseed proteins possess the capacity to modulate the immune system, which holds a pivotal role in cancer surveillance and elimination [127]. Furthermore, flaxseed proteins have been observed to enhance the effectiveness of chemotherapy drugs across different cancer cell lines [21,130].
In essence, the potential advantages of utilizing flaxseed proteins in cancer treatment are indeed promising (Figure 4). However, further research is imperative to fully elucidate their underlying mechanisms of action and gauge their clinical effectiveness in treating cancer.
Flaxseed proteome effect on anti-cancer radiotherapy
Radiotherapy (RT) stands as a prominent method for cancer treatment, functioning by inducing DNA damage within cancer cells. This complex medical procedure necessitates meticulous planning and monitoring. However, despite advancements in RT techniques, certain tumors display notable radiore sistance, leading to elevated rates of treatment failure and tumor relapse [131,132]. Upon encountering DNA damage, cancer cells elevate their antioxidant activity [133] and metabolic processes, encompassing glucose flux, amino acid metabolism, and fatty acid utilization. This metabolic response provides the necessary substrates and energy for the repair of DNA damage [134].
Although research specifically focusing on the impact of flaxseed proteins on radiotherapy remains limited, it's crucial to recognize that flaxseed proteins could potentially offer benefits during radiotherapy due to their antioxidant and anti-inflammatory properties. Flaxseed 2S-albumin, akin to other albumin proteins, possesses antioxidant attributes (Table 1 and Table 2). Antioxidants play a role in neutralizing free radicals generated during radiotherapy [135], thereby reducing oxidative stress and potential harm to healthy tissues. Through its scavenging action against free radicals, flaxseed 2S-albumin might confer protective effects on cells exposed to radiation.
Some studies propose that specific globulin proteins exhibit anti-inflammatory characteristics (Table 1 and Table 3). Inflammation is a natural reaction to radiation exposure [136], but excessive or prolonged inflammation could contribute to tissue damage. By lessening inflammation, flaxseed 11S-globulin might potentially alleviate some of the detrimental effects of radiotherapy on healthy tissues.
Flaxseed proteins, including 2S-albumin and 11S-globulin, furnish essential amino acids and contribute to nutritional support for the body. Maintaining sufficient protein intake is vital during radiotherapy to facilitate tissue repair and overall well-being [137]. Flaxseed proteins can play a role in meeting the nutritional requirements of individuals undergoing radiotherapy, potentially supporting recovery and minimizing treatment-related complications.
Previous research has demonstrated that the intake of flaxseed oil increased the survival rate of rats exposed to 8 Gy gamma radiation. However, this effect was hypothesized to stem from indirect biological mechanisms, such as impacts on the gut microbiota, rather than the direct radioprotective qualities of flaxseed oil [138]. For instance, after being metabolized by the intestinal microbiota, flax components could modify mammary gland miRNAs, potentially reducing the risk of adult breast cancer [139]. Flaxseed has also shown potential in mitigating side effects induced by radiotherapy. In mice subjected to a combination of thoracic radiation therapy and a flaxseed diet, a decrease in p53-responsive miR-34a was observed. This miRNA is associated with cellular maturity and apoptosis regulation [140]. Mice fed with flaxseed exhibited reduced expression of lung injury biomarkers (Bax, p21, and TGF-beta1), oxidative lung damage, lung fibrosis, and inflammatory cell influx into lungs following 13.5 Gy thoracic X-ray radiation exposure. However, the radioprotective effect of flaxseed on Lewis Lung carcinoma was not observed [141,142]. Consequently, targeting tumor metabolism through the flaxseed proteome presents a promising therapeutic avenue for safeguarding normal cells during radiation while sensitizing cancer cells to its effects.
Further research
Delivering proteins to cancer cells is a complex field of research, and the effectiveness of different delivery methods can vary depending on the specific protein and type of cancer cells. More investigation is necessary to optimize the delivery of flaxseed proteins to cancer cells. Nanoparticles, for instance, can be tailored to encapsulate and deliver proteins to specific cells or tissues, including cancer cells. Functionalized nanoparticles can enhance cellular uptake and intracellular delivery of proteins. The successful delivery of anticancer proteins using biocompatible nanoparticles has been demonstrated before [143].
Liposomal delivery is another approach worth considering. Liposomes are lipid-based vesicles capable of encapsulating proteins for targeted delivery. They can be engineered to improve stability, cellular uptake, and controlled release of proteins at the intended site. Liposomal delivery systems have been explored for various therapeutic proteins, including those with anticancer properties [144]. While specific studies involving flaxseed proteins are scarce, liposomal delivery methods can potentially be adapted for their delivery.
Furthermore, proteins can be modified or engineered to enhance their stability, cellular uptake, and affinity for cancer cells. For example, fusion with targeting peptides or antibodies specific to cancer cell surface receptors can facilitate the selective binding and internalization of proteins into cancer cells. This approach has been successfully employed for the targeted delivery of diverse therapeutic proteins [52]. Identifying crucial proteoforms is of utmost importance as they contribute to the development of potential anti-cancer treatments. Highlighting the significance of proteoform-level analyses is crucial for advancing research in this direction most effectively.
Finally, in order to establish cancer treatment with flaxseed proteins, more research is needed (Figure 5) to: (a) Investigate the mechanisms of action - unveiling the specific mechanisms through which individual flaxseed proteins inhibit cancer cell growth and metastasis is vital. This understanding will aid in developing more targeted and effective therapeutic strategies; (b) Determine the optimal dose and duration of treatment - Determining the appropriate dose and treatment duration is critical for the efficacy of flaxseed protein treatment against cancer cells. Tailoring the dosage and duration for different cancer types is essential; (c) Evaluate the potential of flaxseed proteins in combination with other treatments - exploring the potential synergies between flaxseed proteins and other anti-cancer treatments, like chemotherapy and radiation therapy, could enhance treatment outcomes while minimizing side effects; (d) Evaluate the safety and toxicity of flaxseed protein - despite the promising anti-cancer effects observed, the safety and toxicity profiles of flaxseed proteins in humans need thorough evaluation through clinical trials; (e) Conduct clinical trials - conducting well-designed clinical trials is paramount to assessing the efficacy and safety of flaxseed proteins in cancer treatment. Larger sample sizes, longer treatment periods, and placebo-controlled designs are imperative for obtaining reliable and robust results.
Conclusion
Nutrition plays a crucial role in the overall well-being of cancer patients and can significantly contribute to reducing complications during treatment [145,146]. In recent years, there has been a growing focus on plant-based nutrition as a means to protect against a range of leading causes of death, including various types of cancer such as breast, prostate, colorectal, and gastrointestinal cancers. Plant-based diets comprising whole foods have demonstrated substantial protective effects against these cancers and other chronic diseases. These diets can serve as disease-modifying agents, enhancing the management and treatment of these conditions. Consequently, interventions involving nutrition in the prevention of diverse cancers offer a notable complement to existing medical therapies.
The development of a compatible nutraceutical enriched with the protein fraction of flaxseeds and the analysis of its potential anti-metastatic effects represent valuable contributions towards promoting healthy aging within the population. Investigating the biological activities of flaxseed proteins in vivo and assessing their impact on the initial stages of metastasis formation hold promise for advancing our understanding of their therapeutic potential. By exploring these avenues, researchers can uncover novel strategies that not only enhance cancer treatment but also contribute to overall health and wellness.
Author Contributions
Conceptualization Y.M., J.R. and S.L.; Databases A.A.; Original Draft Preparation Y.M.; Review & Editing M.P, J.R., S.L. and Y.M.; Visualization E.K. and Y.M.; Supervision S.L. and Y.M..; Project Administration M.P.; Funding Acquisition, S.L. and M.P.
Funding
This research was funded by the Ministry of Science and Higher Education of the Russian Federation (Goszadaniye) 075-03-2023-106, project № FSMG-2023-0015.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Merkher, Y.; Weihs, D. Proximity of Metastatic Cells Enhances Their Mechanobiological Invasiveness. Ann. Biomed. Eng. 2017, 45, 1399–1406. [Google Scholar] [CrossRef] [PubMed]
- Merkher, Y.; Horesh, Y.; Abramov, Z.; Shleifer, G.; Ben-Ishay, O.; Kluger, Y.; Weihs, D. Rapid Cancer Diagnosis and Early Prognosis of Metastatic Risk Based on Mechanical Invasiveness of Sampled Cells. Ann. Biomed. Eng. 2020, 48, 2846–2858. [Google Scholar] [CrossRef]
- Merkher Yulia; Kontareva Elizaveta; Bogdan Elizaveta; Achkasov Konstantin; Grolman Joshua; Leonov Sergey Nanoparticle Cellular Endocytosis as Potential Prognostic Biomarker for Cancer Progression. FEBS Open Bio 2021, 11, 429–430. [CrossRef]
- Merkher, Y.; Kontareva, E.; Melekhova, A.; Leonov, S. Abstract PO-042: Nanoparticles Imaging for Cancer Metastasis Diagnosis. Clin. Cancer Res. 2021, 27, PO–042. [Google Scholar] [CrossRef]
- Weber, G.F. Molecular Mechanisms of Metastasis. Cancer Lett. 2008, 270, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Eckhardt, B.L.; Francis, P.A.; Parker, B.S.; Anderson, R.L. Strategies for the Discovery and Development of Therapies for Metastatic Breast Cancer. Nat. Rev. Drug Discov. 2012, 11, 479–497. [Google Scholar] [CrossRef] [PubMed]
- Barney, L.E.; Jansen, L.; Polio, S.R.; Galarza, S.; Lynch, M.E.; Barney, L.E.; Jansen, L.; Polio, S.R.; Galarza, S.; Lynch, M.E.; et al. The Predictive Link between Matrix and Metastasis. Curr. Opin. Chem. Eng. 2016, 11, 85–93. [Google Scholar] [CrossRef]
- Van Zijl, F.; Krupitza, G.; Mikulits, W. Initial Steps of Metastasis: Cell Invasion and Endothelial Transmigration. Mutat. Res. - Rev. Mutat. Res. 2011, 728, 23–34. [Google Scholar] [CrossRef]
- Clark, A.G.; Vignjevic, D.M. Modes of Cancer Cell Invasion and the Role of the Microenvironment. Curr. Opin. Cell Biol. 2015, 36, 13–22. [Google Scholar] [CrossRef]
- Zustiak, S.; Nossal, R.; Sackett, D.L. Multiwell Stiffness Assay for the Study of Cell Responsiveness to Cytotoxic Drugs. Biotechnol. Bioeng. 2014, 111, 396. [Google Scholar] [CrossRef]
- Majcherek, D.; Weresa, M.A.; Ciecierski, C. A Cluster Analysis of Risk Factors for Cancer across EU Countries: Health Policy Recommendations for Prevention. Int. J. Environ. Res. Public Health 2021, 18, 8142. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Yust, M.M.; Pedroche, J.; Girón-Calle, J.; Alaiz, M.; Millán, F.; Vioque, J. Production of Ace Inhibitory Peptides by Digestion of Chickpea Legumin with Alcalase. Food Chem. 2003, 81, 363–369. [Google Scholar] [CrossRef]
- Bernacchia, R.; Preti, R.; Vinci, G. Chemical Composition and Health Benefits of Flaxseed. AUSTIN J. Nutr. FOOD Sci. 2022, 2(8), 1–9. [Google Scholar]
- Lowcock, E.C.; Cotterchio, M.; Boucher, B.A. Consumption of Flaxseed, a Rich Source of Lignans, Is Associated with Reduced Breast Cancer Risk. Cancer Causes Control 2013, 24, 813–816. [Google Scholar] [CrossRef] [PubMed]
- McCann, S.E.; Hootman, K.C.; Weaver, A.M.; Thompson, L.U.; Morrison, C.; Hwang, H.; Edge, S.B.; Ambrosone, C.B.; Horvath, P.J.; Kulkarni, S.A. Dietary Intakes of Total and Specific Lignans Are Associated with Clinical Breast Tumor Characteristics. J. Nutr. 2012, 142, 91. [Google Scholar] [CrossRef]
- Freitas, R.D.S.; Campos, M.M. Protective Effects of Omega-3 Fatty Acids in Cancer-Related Complications. Nutrients 2019, 11. [Google Scholar] [CrossRef]
- D’Eliseo, D.; Velotti, F. Omega-3 Fatty Acids and Cancer Cell Cytotoxicity: Implications for Multi-Targeted Cancer Therapy. J. Clin. Med. 2016, 5. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free Radicals, Antioxidants and Functional Foods: Impact on Human Health. Pharmacogn. Rev. 2010, 4, 118. [Google Scholar] [CrossRef]
- Pruteanu, L.L.; Bailey, D.S.; Grădinaru, A.C.; Jäntschi, L. The Biochemistry and Effectiveness of Antioxidants in Food, Fruits, and Marine Algae. Antioxidants 2023, Vol. 12, Page 860 2023, 12, 860. [Google Scholar] [CrossRef]
- Pal, P.; Hales, K.; Petrik, J.; Hales, D.B. Pro-Apoptotic and Anti-Angiogenic Actions of 2-Methoxyestradiol and Docosahexaenoic Acid, the Biologically Derived Active Compounds from Flaxseed Diet, in Preventing Ovarian Cancer. J. Ovarian Res. 2019, 12. [Google Scholar] [CrossRef] [PubMed]
- Kajla, P.; Goyal, N.; Bangar, S.P.; Chaudhary, V.; Lorenzo, J.M. Flaxseed Proteins (Linum Usitassimum): Thermal, Functional and Spectroscopic Characterization. Food Anal. Methods 2023, 16, 459–467. [Google Scholar] [CrossRef]
- Mueed, A.; Madjirebaye, P.; Shibli, S.; Deng, Z. Flaxseed Peptides and Cyclolinopeptides: A Critical Review on Proteomic Approaches, Biological Activity, and Future Perspectives. J. Agric. Food Chem. 2022, 70, 14600–14612. [Google Scholar] [CrossRef]
- Sammour, R.H. Proteins of Linseed (Linum Usitatissimum L.), Extraction and Characterization by Electrophoresis. Bot. Bull. Acad. Sin. 1999. [Google Scholar]
- Ayad, A.A. Characterization and Properties of Flaxseed Protein Fractions, 2010.
- Singh, K.K.; Mridula, D.; Rehal, J.; Barnwal, P. Flaxseed: A Potential Source of Food, Feed and Fiber. 2011, 51, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Saini, C.S. Amino Acid Composition, Nutritional Profiling, Mineral Content and Physicochemical Properties of Protein Isolate from Flaxseeds (Linum Usitatissimum). J. Food Meas. Charact. 2022, 16, 829–839. [Google Scholar] [CrossRef]
- Peng, D.; Ye, J.; Jin, W.; Yang, J.; Geng, F.; Deng, Q. A Review on the Utilization of Flaxseed Protein as Interfacial Stabilizers for Food Applications. JAOCS, J. Am. Oil Chem. Soc. 2022, 99, 723–737. [Google Scholar] [CrossRef]
- Oomah, B.D.; Mazza, G. Flaxseed Proteins—a Review. Food Chem. 1993, 48, 109–114. [Google Scholar] [CrossRef]
- Arntfield, S.D. Proteins from Oil-Producing Plants. Proteins Food Process. Second Ed. 2018, 187–221. [Google Scholar] [CrossRef]
- Xie, M.; Liu, D.; Yang, Y. Anti-Cancer Peptides: Classification, Mechanism of Action, Reconstruction and Modification. Open Biol. 2020, 10. [Google Scholar] [CrossRef]
- Fillería, S.G.; Nardo, A.E.; Paulino, M.; Tironi, V. Peptides Derived from the Gastrointestinal Digestion of Amaranth 11S Globulin: Structure and Antioxidant Functionality. Food Chem. Mol. Sci. 2021, 3, 100053. [Google Scholar] [CrossRef]
- Taghizadeh, S.F.; Rezaee, R.; Mehmandoust, M.; Badibostan, H.; Karimi, G. Assessment of in Vitro Bioactivities of Pis v 1 (2S Albumin) and Pis v 2.0101 (11S Globulin) Proteins Derived from Pistachio (Pistacia Vera L.). J. Food Meas. Charact. 2020, 14, 1054–1063. [Google Scholar] [CrossRef]
- Sandoval-Sicairos, E.S.; Milán-Noris, A.K.; Luna-Vital, D.A.; Milán-Carrillo, J.; Montoya-Rodríguez, A. Anti-Inflammatory and Antioxidant Effects of Peptides Released from Germinated Amaranth during in Vitro Simulated Gastrointestinal Digestion. Food Chem. 2021, 343, 128394. [Google Scholar] [CrossRef]
- Martínez, J.H.; Velázquez, F.; Burrieza, H.P.; Martínez, K.D.; Paula Domínguez Rubio, A.; dos Santos Ferreira, C.; del Pilar Buera, M.; Pérez, O.E. Betanin Loaded Nanocarriers Based on Quinoa Seed 11S Globulin. Impact on the Protein Structure and Antioxidant Activity. Food Hydrocoll. 2019, 87, 880–890. [Google Scholar] [CrossRef]
- Luo, X.; Wu, W.; Feng, L.; Treves, H.; Ren, M. Short Peptides Make a Big Difference: The Role of Botany-Derived Amps in Disease Control and Protection of Human Health. Int. J. Mol. Sci. 2021, 22, 11363. [Google Scholar] [CrossRef]
- Paterson, S.; Fernández-Tomé, S.; Hernández-Ledesma, B. Modulatory Effects of a Lunasin-Enriched Soybean Extract on Immune Response and Oxidative Stress-Associated Biomarkers. Biol. Life Sci. Forum 2022, Vol. 12, Page 10 2022, 12, 10. [Google Scholar] [CrossRef]
- Caponio, G.R.; Wang, D.Q.H.; Di Ciaula, A.; De Angelis, M.; Portincasa, P. Regulation of Cholesterol Metabolism by Bioactive Components of Soy Proteins: Novel Translational Evidence. Int. J. Mol. Sci. 2021, 22, 1–18. [Google Scholar] [CrossRef]
- Capraro, J.; De Benedetti, S.; Heinzl, G.C.; Scarafoni, A.; Magni, C. Bioactivities of Pseudocereal Fractionated Seed Proteins and Derived Peptides Relevant for Maintaining Human Well-Being. Int. J. Mol. Sci. 2021, 22, 3543. [Google Scholar] [CrossRef]
- Ateeq, M.; Adeel, M.M.; Kanwal, A.; Qamar, M.T.U.; Saeed, A.; Khaliq, B.; Saeed, Q.; Atiq, M.N.; Bilal, M.; Alharbi, M.; et al. In Silico Analysis and Functional Characterization of Antimicrobial and Insecticidal Vicilin from Moth Bean (Vigna Aconitifolia (Jacq.) Marechal) Seeds. Molecules 2022, 27, 3251. [Google Scholar] [CrossRef]
- Chen, Y.J.; Chen, Y.Y.; Wu, C.T.; Yu, C.C.; Liao, H.F. Prolamin, a Rice Protein, Augments Anti-Leukaemia Immune Response. J. Cereal Sci. 2010, 51, 189–197. [Google Scholar] [CrossRef]
- Ji, Z.; Mao, J.; Chen, S.; Mao, J. Antioxidant and Anti-Inflammatory Activity of Peptides from Foxtail Millet (Setaria Italica) Prolamins in HaCaT Cells and RAW264.7 Murine Macrophages. Food Biosci. 2020, 36, 100636. [Google Scholar] [CrossRef]
- Jimenez-Pulido, I.J.; Daniel, R.; Perez, J.; Martínez-Villaluenga, C.; De Luis, D.; Martín Diana, A.B. Impact of Protein Content on the Antioxidants, Anti-Inflammatory Properties and Glycemic Index of Wheat and Wheat Bran. Foods 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Montserrat-de la Paz, S.; Rodriguez-Martin, N.M.; Villanueva, A.; Pedroche, J.; Cruz-Chamorro, I.; Millan, F.; Millan-Linares, M.C. Evaluation of Anti-Inflammatory and Atheroprotective Properties of Wheat Gluten Protein Hydrolysates in Primary Human Monocytes. Foods 2020, 9, 854. [Google Scholar] [CrossRef] [PubMed]
- Aqeel, T.; Gurumallu, S.C.; Bhaskar, A.; Hashimi, S.M.; Lohith, N.C.; Javaraiah, R. Protective Role of Flaxseed Lignan Secoisolariciresinol Diglucoside against Lead-Acetate-Induced Oxidative-Stress-Mediated Nephrotoxicity in Rats. Phytomedicine Plus 2021, 1, 100038. [Google Scholar] [CrossRef]
- Rajesha, J.; Ranga Rao, A.; Madhusudhan, B.; Karunakumar, M. Antibacterial Properties of Secoisolariciresinol Diglucoside Isolated from Indian Flaxseed Cultivars. Curr. Trends Biotechnol. Pharm. 2010, 4, 551–560. [Google Scholar]
- Nguyen, N.P.T.; Cong, T.L.; Tran, T.T.H.; Do, B.N.; Nguyen, S.T.; Vu, B.T.; Nguyen, L.H.T.; Van Ngo, M.; Dinh, H.T.; Huy, H.D.; et al. Lower Plasma Albumin, Higher White Blood Cell Count and High-Sensitivity C-Reactive Protein Are Associated with Femoral Artery Intima-Media Thickness Among Newly Diagnosed Patients with Type 2 Diabetes Mellitus. Int. J. Gen. Med. 2022, 15, 2715–2725. [Google Scholar] [CrossRef]
- Lessomo, F.Y.N.; Fan, Q.; Wang, Z.Q.; Mukuka, C. The Relationship between Leukocyte to Albumin Ratio and Atrial Fibrillation Severity. BMC Cardiovasc. Disord. 2023, 23, 1–8. [Google Scholar] [CrossRef]
- Gupta, D.; Lis, C.G. Pretreatment Serum Albumin as a Predictor of Cancer Survival: A Systematic Review of the Epidemiological Literature. Nutr. J. 2010, 9, 69. [Google Scholar] [CrossRef]
- Nazha, B.; Moussaly, E.; Zaarour, M.; Weerasinghe, C.; Azab, B. Hypoalbuminemia in Colorectal Cancer Prognosis: Nutritional Marker or Inflammatory Surrogate? World J. Gastrointest. Surg. 2015, 7, 370. [Google Scholar] [CrossRef]
- Van De Wouw, J.; Joles, J.A. Albumin Is an Interface between Blood Plasma and Cell Membrane, and Not Just a Sponge. Clin. Kidney J. 2022, 15, 624–634. [Google Scholar] [CrossRef]
- Cho, H.; Jeon, S.I.; Ahn, C.H.; Shim, M.K.; Kim, K. Emerging Albumin-Binding Anticancer Drugs for Tumor-Targeted Drug Delivery: Current Understandings and Clinical Translation. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, Z.; Yu, S.; Chen, S.; Zhou, Y.; Qu, Y.; Xu, P.; Jiang, L.; Yuan, C.; Huang, M. Albumin-Based Drug Carrier Targeting Urokinase Receptor for Cancer Therapy. Int. J. Pharm. 2023, 634. [Google Scholar] [CrossRef] [PubMed]
- Ranaivo, H.R.; Hodge, J.N.; Choi, N.; Wainwright, M.S. Albumin Induces Upregulation of Matrix Metalloproteinase-9 in Astrocytes via MAPK and Reactive Oxygen Species-Dependent Pathways. J. Neuroinflammation 2012, 9, 68. [Google Scholar] [CrossRef] [PubMed]
- von Au, A.; Vasel, M.; Kraft, S.; Sens, C.; Hackl, N.; Marx, A.; Stroebel, P.; Hennenlotter, J.; Todenhöfer, T.; Stenzl, A.; et al. Circulating Fibronectin Controls Tumor Growth. Neoplasia (United States) 2013, 15, 925–938. [Google Scholar] [CrossRef]
- Zhao, P.; Wang, Y.; Wu, A.; Rao, Y.; Huang, Y. Roles of Albumin-Binding Proteins in Cancer Progression and Biomimetic Targeted Drug Delivery. ChemBioChem 2018, 19, 1796–1805. [Google Scholar] [CrossRef]
- Givant-Horwitz, V.; Davidson, B.; Reich, R. Laminin-Induced Signaling in Tumor Cells. Cancer Lett. 2005, 223, 1–10. [Google Scholar] [CrossRef]
- Radovic, R.S.; Maksimovic, R. V.; Brkljacic, M.J.; Varkonji Gasic, I.E.; Savic, P.A. 2S Albumin from Buckwheat (Fagopyrum Esculentum Moench) Seeds. J. Agric. Food Chem. 1999, 47, 1467–1470. [Google Scholar] [CrossRef]
- Souza, P.F.N. The Forgotten 2S Albumin Proteins: Importance, Structure, and Biotechnological Application in Agriculture and Human Health. Int. J. Biol. Macromol. 2020, 164, 4638–4649. [Google Scholar] [CrossRef]
- Bueno-Díaz, C.; Martín-Pedraza, L.; Parrón, J.; Cuesta-Herranz, J.; Cabanillas, B.; Pastor-Vargas, C.; Batanero, E.; Villalba, M. Characterization of Relevant Biomarkers for the Diagnosis of Food Allergies: An Overview of the 2s Albumin Family. Foods 2021, 10. [Google Scholar] [CrossRef]
- Khan, S.; Ali, S.A.; Yasmin, T.; Ahmed, M.; Khan, H. Purification and Characterization of 2S Albumin from Nelumbo Nucifera. Biosci. Biotechnol. Biochem. 2016, 80, 2109–2114. [Google Scholar] [CrossRef]
- Shidal, C.; Inaba, J.I.; Yaddanapudi, K.; Davis, K.R. The Soy-Derived Peptide Lunasin Inhibits Invasive Potential of Melanoma Initiating Cells. Oncotarget 2017, 8, 25525–25541. [Google Scholar] [CrossRef] [PubMed]
- Vuyyuri, S.B.; Shidal, C.; Davis, K.R. Development of the Plant-Derived Peptide Lunasin as an Anticancer Agent. Curr. Opin. Pharmacol. 2018, 41, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Seber, L.E.; Barnett, B.W.; McConnell, E.J.; Hume, S.D.; Cai, J.; Boles, K.; Davis, K.R. Scalable Purification and Characterization of the Anticancer Lunasin Peptide from Soybean. PLoS One 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Dia, V.P.; Gonzalez de Mejia, E. Lunasin Potentiates the Effect of Oxaliplatin Preventing Outgrowth of Colon Cancer Metastasis, Binds to α 5β 1 Integrin and Suppresses FAK/ERK/NF-ΚB Signaling. Cancer Lett. 2011, 313, 167–180. [Google Scholar] [CrossRef]
- Fernández-tomé, S.; Xu, F.; Han, Y.; Hernández-ledesma, B.; Xiao, H. Inhibitory Effects of Peptide Lunasin in Colorectal Cancer Hct-116 Cells and Their Tumorsphere- Derived Subpopulation. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef]
- Dia, V.P.; De Mejia, E.G. Lunasin Induces Apoptosis and Modifies the Expression of Genes Associated with Extracellular Matrix and Cell Adhesion in Human Metastatic Colon Cancer Cells. Mol. Nutr. Food Res. 2011, 55, 623–634. [Google Scholar] [CrossRef]
- Jiang, Q.; Pan, Y.; Cheng, Y.; Li, H.; Liu, D.; Li, H. Lunasin Suppresses the Migration and Invasion of Breast Cancer Cells by Inhibiting Matrix Metalloproteinase-2/-9 via the FAK/Akt/ERK and NF-ΚB Signaling Pathways. Oncol. Rep. 2016, 36, 253–262. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Gehlot, P. Inflammation and Cancer: How Friendly Is the Relationship for Cancer Patients? Curr. Opin. Pharmacol. 2009, 9, 351–369. [Google Scholar] [CrossRef]
- Wan, X.; Liu, H.; Sun, Y.; Zhang, J.; Chen, X.; Chen, N. Lunasin: A Promising Polypeptide for the Prevention and Treatment of Cancer. Oncol. Lett. 2017, 13, 3997–4001. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; Hsieh, C.C.; de Lumen, B.O. Antioxidant and Anti-Inflammatory Properties of Cancer Preventive Peptide Lunasin in RAW 264.7 Macrophages. Biochem. Biophys. Res. Commun. 2009, 390, 803–808. [Google Scholar] [CrossRef]
- Vuyyuri, S.B.; Shidal, C.; Davis, K.R. Development of the Plant-Derived Peptide Lunasin as an Anticancer Agent. Curr. Opin. Pharmacol. 2018, 41, 27–33. [Google Scholar] [CrossRef] [PubMed]
- McConnell, E.J.; Devapatla, B.; Yaddanapudi, K.; Davis, K.R. The Soybean-Derived Peptide Lunasin Inhibits Non-Small Cell Lung Cancer Cell Proliferation by Suppressing Phosphorylation of the Retinoblastoma Protein. Oncotarget 2015, 6, 4649–4662. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.C.; Hernández-Ledesma, B.; de Lumen, B.O. Lunasin, a Novel Seed Peptide, Sensitizes Human Breast Cancer MDA-MB-231 Cells to Aspirin-Arrested Cell Cycle and Induced Apoptosis. Chem. Biol. Interact. 2010, 186, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Youle, R.J.; Huang, A.H.C. OCCURRENCE OF LOW MOLECULAR WEIGHT AND HIGH CYSTEINE CONTAINING ALBUMIN STORAGE PROTEINS IN OILSEEDS OF DIVERSE SPECIES. Am. J. Bot. 1981, 68, 44–48. [Google Scholar] [CrossRef]
- Madhusudhan, K.T.; Singh, N. Isolation and Characterization of a Small Molecular Weight Protein of Linseed Meal. Phytochemistry 1985, 24, 2507–2509. [Google Scholar] [CrossRef]
- Dev, D.K.; Sienkiewicz, T. Isolation and Subunit Composition of 11 S Globulin of Linseed (Linum Usitatissimum L.). Food / Nahrung 1987, 31, 767–769. [Google Scholar] [CrossRef]
- El-Saadany, A.S.; Hanafy, M.M.; Elkomy, A.E. Flaxseed and Agnus-Castuson Vitex as a Source of Phytoestrogens and Their Impact on Productive Performance, Some Blood Constituents, and Blood Oestradiol Profile of Aged Laying Hens. Ital. J. Anim. Sci. 2022, 21, 821–830. [Google Scholar] [CrossRef]
- Singh, A.; Meena, M.; Kumar, D.; Dubey, A.K.; Hassan, M.I. Structural and Functional Analysis of Various Globulin Proteins from Soy Seed. Crit. Rev. Food Sci. Nutr. 2015, 55, 1491–1502. [Google Scholar] [CrossRef]
- Paul, M.K.; Mukhopadhyay, A.K. Tyrosine Kinase – Role and Significance in Cancer. Int. J. Med. Sci. 2012, 1, 101–115. [Google Scholar] [CrossRef]
- Bochnak-Niedźwiecka, J.; Szymanowska, U.; Kapusta, I.; Świeca, M. Antioxidant Content and Antioxidant Capacity of the Protein-Rich Powdered Beverages Enriched with Flax Seeds Gum. Antioxidants 2022, 11, 582. [Google Scholar] [CrossRef]
- Yousif, A.N. Effect of Flaxseed on Some Hormonal Profile and Genomic DNA Concentration in Karadi Lambs. In Proceedings of the IOP Conference Series: Earth and Environmental Science; IOP Publishing, November 1 2019; Vol. 388; p. 012035. [Google Scholar]
- Nowak, D.A.; Snyder, D.C.; Brown, A.J.; Demark-Wahnefried, W. The Effect of Flaxseed Supplementation on Hormonal Levels Associated with Polycystic Ovarian Syndrome: A Case Study. Curr. Top. Nutraceutical Res. 2007, 5, 177–181. [Google Scholar] [PubMed]
- Chang, V.C.; Cotterchio, M.; Boucher, B.A.; Jenkins, D.J.A.; Mirea, L.; McCann, S.E.; Thompson, L.U. Effect of Dietary Flaxseed Intake on Circulating Sex Hormone Levels among Postmenopausal Women: A Randomized Controlled Intervention Trial. Nutr. Cancer 2019, 71, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Ajibola, C.F.; Aluko, R.E. Physicochemical and Functional Properties of 2S, 7S, and 11S Enriched Hemp Seed Protein Fractions. Molecules 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Adachi, M.; Kanamori, J.; Masuda, T.; Yagasaki, K.; Kitamura, K.; Mikami, B.; Utsumi, S. Crystal Structure of Soybean 11S Globulin: Glycinin A3B4 Homohexamer. Proc. Natl. Acad. Sci. U. S. A. 2003, 100, 7395–7400. [Google Scholar] [CrossRef]
- Quiroga, A. V.; Barrio, D.A.; Añón, M.C. Amaranth Lectin Presents Potential Antitumor Properties. LWT 2015, 60, 478–485. [Google Scholar] [CrossRef]
- Mora-Escobedo, R.; Robles-Ramírez, M. del C.; Ramón-Gallegos, E.; Reza-Alemán, R. Effect of Protein Hydrolysates from Germinated Soybean on Cancerous Cells of the Human Cervix: An In Vitro Study. Plant Foods Hum. Nutr. 2009, 64, 271–278. [Google Scholar] [CrossRef]
- Wang, L.; Sun, Z.; Xie, W.; Peng, C.; Ding, H.; Li, Y.; Feng, S.; Wang, X.; Zhao, C.; Wu, J. 11S Glycinin Up-Regulated NLRP-3-Induced Pyroptosis by Triggering Reactive Oxygen Species in Porcine Intestinal Epithelial Cells. Front. Vet. Sci. 2022, 9, 677. [Google Scholar] [CrossRef]
- Silva-Sánchez, C.; Barba De La Rosa, A.P.; León-Galván, M.F.; De Lumen, B.O.; De León-Rodríguez, A.; González De Mejía, E. Bioactive Peptides in Amaranth (Amaranthus Hypochondriacus) Seed. J. Agric. Food Chem. 2008, 56, 1233–1240. [Google Scholar] [CrossRef]
- Zou, X.G.; Hu, J.N.; Li, J.; Yang, J.Y.; Du, Y.X.; Yu, Y.F.; Deng, Z.Y. ICellular Uptake of [1–9-NαC]-Linusorb B2 and [1–9-NαC]-Linusorb B3 Isolated from Flaxseed, and Their Antitumor Activities in Human Gastric SGC-7901 Cells. J. Funct. Foods 2018, 48, 692–703. [Google Scholar] [CrossRef]
- Zou, X.G.; Hu, J.N.; Wang, M.; Du, Y.X.; Li, J.; Mai, Q.Y.; Deng, Z.Y. [1–9-NαC]-Linusorb B2 and [1–9-NαC]-Linusorb B3 Isolated from Flaxseed Induce G1 Cell Cycle Arrest on SGC-7901 Cells by Modulating the AKT/JNK Signaling Pathway. J. Funct. Foods 2019, 52, 332–339. [Google Scholar] [CrossRef]
- Okinyo-Owiti, D.P.; Dong, Q.; Ling, B.; Jadhav, P.D.; Bauer, R.; Maley, J.M.; Reaney, M.J.T.; Yang, J.; Sammynaiken, R. Evaluating the Cytotoxicity of Flaxseed Orbitides for Potential Cancer Treatment. Toxicol. Reports 2015, 2, 1014–1018. [Google Scholar] [CrossRef]
- Badley, R.A.; Atkinson, D.; Hauser, H.; Oldani, D.; Green, J.P.; Stubbs, J.M. The Structure, Physical and Chemical Properties of the Soy Bean Protein Glycinin. BBA - Protein Struct. 1975, 412, 214–228. [Google Scholar] [CrossRef]
- Gavage, M.; Van Vlierberghe, K.; Van Poucke, C.; De Loose, M.; Gevaert, K.; Dieu, M.; Renard, P.; Arnould, T.; Gillard, N. High-Resolution Mass Spectrometry-Based Selection of Peanut Peptide Biomarkers Considering Food Processing and Market Type Variation. Food Chem. 2020, 304, 125428. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Jin, Y.; Cerny, R.; Ballmer-Weber, B.; Goodman, R.E. Combining 2-DE Immunoblots and Mass Spectrometry to Identify Putative Soybean (Glycine Max) Allergens. Food Chem. Toxicol. 2018, 116, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Magni, C.; Scarafoni, A.; Herndl, A.; Sessa, F.; Prinsi, B.; Espen, L.; Duranti, M. Combined 2D Electrophoretic Approaches for the Study of White Lupin Mature Seed Storage Proteome. Phytochemistry 2007, 68, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Duranti, M.; Horstmann, C.; Gilroy, J.; Croy, R.R.D. The Molecular Basis for N-Glycosylation in the 11S Globulin (Legumin) of Lupin Seed. J. Protein Chem. 1995, 14, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Romero-Rodríguez, M.C.; Maldonado-Alconada, A.M.; Valledor, L.; Jorrin-Novo, J. V. Back to Osborne. Sequential Protein Extraction and Lc-Ms Analysis for the Characterization of the Holm Oak Seed Proteome. Methods Mol. Biol. 2014, 1072, 379–389. [Google Scholar] [CrossRef]
- Ribeiro, A.C.; Teixeira, A.R.; Ferreira, R.B. Characterization of Globulins from Common Vetch (Vicia Sativa L.). J. Agric. Food Chem. 2004, 52, 4913–4920. [Google Scholar] [CrossRef]
- Amponsah, A.; Nayak, B. Evaluation of the Efficiency of Three Extraction Conditions for the Immunochemical Detection of Allergenic Soy Proteins in Different Food Matrices. J. Sci. Food Agric. 2018, 98, 2378–2384. [Google Scholar] [CrossRef]
- Vaintraub, I.A.; Bassüner, R.; Shutov, A.D. The Action of Trypsin and Chymotrypsin on the Reserve Proteins of Some Leguminous Seeds. Food / Nahrung 1976, 20, 763–771. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Adebiyi, A.P.; Doyen, A.; Li, H.; Bazinet, L.; Aluko, R.E. Low Molecular Weight Flaxseed Protein-Derived Arginine-Containing Peptides Reduced Blood Pressure of Spontaneously Hypertensive Rats Faster than Amino Acid Form of Arginine and Native Flaxseed Protein. Food Chem. 2012, 132, 468–475. [Google Scholar] [CrossRef]
- Boschin, G.; Scigliuolo, G.M.; Resta, D.; Arnoldi, A. ACE-Inhibitory Activity of Enzymatic Protein Hydrolysates from Lupin and Other Legumes. Food Chem. 2014, 145, 34–40. [Google Scholar] [CrossRef]
- Ingkaninan, K.; De Best, C.M.; Van Der Heijden, R.; Hofte, A.J.P.; Karabatak, B.; Irth, H.; Tjaden, U.R.; Van Der Greef, J.; Verpoorte, R. High-Performance Liquid Chromatography with on-Line Coupled UV, Mass Spectrometric and Biochemical Detection for Identification of Acetylcholinesterase Inhibitors from Natural Products. J. Chromatogr. A 2000, 872, 61–73. [Google Scholar] [CrossRef]
- Dutta, S.; Ray, S.; Nagarajan, K. Glutamic Acid as Anticancer Agent: An Overview. Saudi Pharm. J. 2013, 21, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lu, Y.; Li, Y.; Xu, Y.; Song, W. Poly(Glutamic Acid)-Engineered Nanoplatforms for Enhanced Cancer Phototherapy. Curr. Drug Deliv. 2023, 20. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Yamamoto, K.; Sato, Y.; Inoue, S.; Morinaga, T.; Hirano, E. Combination of Aspartic Acid and Glutamic Acid Inhibits Tumor Cell Proliferation. Biomed. Res. 2016, 37, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Shen, M.; Xu, W. Arginine, Glycine, Aspartic Acid Peptide-Modified Paclitaxel and Curcumin Co-Loaded Liposome for the Treatment of Lung Cancer: In Vitro/Vivo Evaluation. Int. J. Nanomedicine 2018, 13, 2561–2569. [Google Scholar] [CrossRef]
- Park, K.G.M.; Hayes, P.D.; Eremin, O.; Sewell, H.; Park, K.G.M.; Garlick, P.J. Stimulation of Lymphocyte Natural Cytotoxicity by L-Arginine. Lancet 1991, 337, 645–646. [Google Scholar] [CrossRef]
- Futaki, S.; Suzuki, T.; Ohashi, W.; Yagami, T.; Tanaka, S.; Ueda, K.; Sugiura, Y. Arginine-Rich Peptides. An Abundant Source of Membrane-Permeable Peptides Having Potential as Carriers for Intracellular Protein Delivery. J. Biol. Chem. 2001, 276, 5836–5840. [Google Scholar] [CrossRef]
- Schmidt, N.; Mishra, A.; Lai, G.H.; Wong, G.C.L. Arginine-Rich Cell-Penetrating Peptides. FEBS Lett. 2010, 584, 1806–1813. [Google Scholar] [CrossRef]
- Wang, H.; Chen, S.; Kang, W.; Ding, B.; Cui, S.; Zhou, L.; Zhang, N.; Luo, H.; Wang, M.; Zhang, F.; et al. High Dose Isoleucine Stabilizes Nuclear PTEN to Suppress the Proliferation of Lung Cancer. Discov. Oncol. 2023, 14, 25. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Li, L.; Yu, X.; Deng, Q.; Xiang, Q.; Zhu, Y. Comparative Composition Structure and Selected Techno-Functional Elucidation of Flaxseed Protein Fractions. Foods 2022, 11. [Google Scholar] [CrossRef]
- Mueed, A.; Shibli, S.; Korma, S.A.; Madjirebaye, P.; Esatbeyoglu, T.; Deng, Z. Flaxseed Bioactive Compounds: Chemical Composition, Functional Properties, Food Applications and Health Benefits-Related Gut Microbes. Foods 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.W.Y.; Lei, B.; Li-Chan, E.C.Y. Isolation and Structural Characterization of the Major Protein Fraction from NorMan Flaxseed (Linum Usitatissimum L.). Food Chem. 2005, 90, 271–279. [Google Scholar] [CrossRef]
- Bhatty, R.S.; Cherdkiatgumchai, P. Compositional Analysis of Laboratory-Prepared and Commercial Samples of Linseed Meal and of Hull Isolated from Flax. J. Am. Oil Chem. Soc. 1990, 67, 79–84. [Google Scholar] [CrossRef]
- Lee, K.H.; Qi, G.H.; Sim, J.S. Metabolizable Energy and Amino Acid Availability of Full-Fat Seeds, Meals, and Oils of Flax and Canola. Poult. Sci. 1995, 74, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Porokhovinova, E.A.; Shelenga, T. V.; Kerv, Y.A.; Khoreva, V.I.; Konarev, A. V.; Yakusheva, T. V.; Pavlov, A. V.; Slobodkina, A.A.; Brutch, N.B. Features of Profiles of Biologically Active Compounds of Primary and Secondary Metabolism of Lines from VIR Flax Genetic Collection, Contrasting in Size and Color of Seeds. Plants 2022, 11, 750. [Google Scholar] [CrossRef] [PubMed]
- Shim, Y.Y.; Gui, B.; Arnison, P.G.; Wang, Y.; Reaney, M.J.T. Flaxseed (Linum Usitatissimum L.) Bioactive Compounds and Peptide Nomenclature: Areview. Trends Food Sci. Technol. 2014, 38, 5–20. [Google Scholar] [CrossRef]
- Kolodziejczyk, P.; Ozimek, L.; Kozłowska, J. The Application of Flax and Hemp Seeds in Food, Animal Feed and Cosmetics Production. In Handbook of Natural Fibres; Woodhead Publishing, 2012; pp. 329–366. [Google Scholar]
- Wu, S.; Wang, X.; Qi, W.; Guo, Q. Bioactive Protein/Peptides of Flaxseed: A Review. Trends Food Sci. Technol. 2019, 92, 184–193. [Google Scholar] [CrossRef]
- Pirmohammadi, A.; Khalaji, S.; Yari, M. Effects of Linseed Expansion on Its Dietary Molecular Structures, and on Broiler Chicks Digestive Enzymes Activity, Serum Metabolites, and Ileal Morphology. J. Appl. Poult. Res. 2019, 28, 997–1012. [Google Scholar] [CrossRef]
- Ahmed Abdullah Bakhashwain, A.A.B. Evaluation of Different Flax Cultivars in Their Oil, Fatty Acids Protein AndAmino Acids and Correlations. J. King Abdulaziz Univ. - Meteorol. Environ. Arid L. Agric. Sci. 2017, 27, 51–58. [Google Scholar] [CrossRef]
- Subba, P.; Narayana Kotimoole, C.; Prasad, T.S.K. Plant Proteome Databases and Bioinformatic Tools: An Expert Review and Comparative Insights. Omi. A J. Integr. Biol. 2019, 23, 190–206. [Google Scholar] [CrossRef] [PubMed]
- Barvkar, V.T.; Pardeshi, V.C.; Kale, S.M.; Kadoo, N.Y.; Giri, A.P.; Gupta, V.S. Proteome Profiling of Flax (Linum Usitatissimum) Seed: Characterization of Functional Metabolic Pathways Operating during Seed Development. J. Proteome Res. 2012, 11, 6264–6276. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, R.; Raina, P.; Shinde, K.; Mansara, P.; Karandikar, M.; Kaul-Ghanekar, R. Flax Seed Oil Reduced Tumor Growth, Modulated Immune Responses and Decreased HPV E6 and E7 Oncoprotein Expression in a Murine Model of Ectopic Cervical Cancer. Prostaglandins Other Lipid Mediat. 2019, 143, 106332. [Google Scholar] [CrossRef]
- Sung, N.Y.; Jeong, D.; Shim, Y.Y.; Ratan, Z.A.; Jang, Y.J.; Reaney, M.J.T.; Lee, S.; Lee, B.H.; Kim, J.H.; Yi, Y.S.; et al. The Anti-Cancer E_ect of Linusorb B3 from Flaxseed Oil through the Promotion of Apoptosis, Inhibition of Actin Polymerization, and Suppression of Src Activity in Glioblastoma Cells. Molecules 2020, 25, 5881. [Google Scholar] [CrossRef]
- Lee, J.; Cho, K. Flaxseed Sprouts Induce Apoptosis and Inhibit Growth in MCF-7 and MDA-MB-231 Human Breast Cancer Cells. Vitr. Cell. Dev. Biol. - Anim. 2012, 48, 244–250. [Google Scholar] [CrossRef]
- De Silva, S.F.; Alcorn, J. Flaxseed Lignans as Important Dietary Polyphenols for Cancer Prevention and Treatment: Chemistry, Pharmacokinetics, and Molecular Targets. Pharmaceuticals 2019, 12. [Google Scholar] [CrossRef]
- de Mey, S.; Dufait, I.; De Ridder, M. Radioresistance of Human Cancers: Clinical Implications of Genetic Expression Signatures. Front. Oncol. 2021, 11. [Google Scholar] [CrossRef]
- Fukui, R.; Saga, R.; Matsuya, Y.; Tomita, K.; Kuwahara, Y.; Ohuchi, K.; Sato, T.; Okumura, K.; Date, H.; Fukumoto, M.; et al. Tumor Radioresistance Caused by Radiation-Induced Changes of Stem-like Cell Content and Sub-Lethal Damage Repair Capability. Sci. Reports 2022 121 2022, 12, 1–12. [Google Scholar] [CrossRef]
- Engel, A.L.; Lorenz, N.I.; Klann, K.; Münch, C.; Depner, C.; Steinbach, J.P.; Ronellenfitsch, M.W.; Luger, A.L. Serine-Dependent Redox Homeostasis Regulates Glioblastoma Cell Survival. Br. J. Cancer 2020, 122, 1391–1398. [Google Scholar] [CrossRef]
- Tang, L.; Wei, F.; Wu, Y.; He, Y.; Shi, L.; Xiong, F.; Gong, Z.; Guo, C.; Li, X.; Deng, H.; et al. Role of Metabolism in Cancer Cell Radioresistance and Radiosensitization Methods. J. Exp. Clin. Cancer Res. 2018 371 2018, 37, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Okunieff, P.; Swarts, S.; Keng, P.; Sun, W.; Wang, W.; Kim, J.; Yang, S.; Zhang, H.; Liu, C.; Williams, J.P.; et al. ANTIOXIDANTS REDUCE CONSEQUENCES OF RADIATION EXPOSURE. Adv. Exp. Med. Biol. 2008, 614, 165. [Google Scholar] [CrossRef]
- Di Maggio, F.M.; Minafra, L.; Forte, G.I.; Cammarata, F.P.; Lio, D.; Messa, C.; Gilardi, M.C.; Bravatà, V. Portrait of Inflammatory Response to Ionizing Radiation Treatment. J. Inflamm. (United Kingdom) 2015, 12, 1–11. [Google Scholar] [CrossRef]
- Ravasco, P. Nutrition in Cancer Patients. J. Clin. Med. 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, S.M.J.; Mosleh-Shirazi, M.A.; Tavassoli, A.R.; Taheri, M.; Mehdizadeh, A.R.; Namazi, S.A.S.; Jamali, A.; Ghalandari, R.; Bonyadi, S.; Haghani, M.; et al. Increased Radioresistance to Lethal Doses of Gamma Rays in Mice and Rats after Exposure to Microwave Radiation Emitted by a GSM Mobile Phone Simulator. Dose. Response. 2012, 11, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Taibi, A.; Thompson, L.; Comelli, E. Flaxseed Alters Gut Microbiota-Mammary Gland MicroRNA Relationships Differently Than Its Oil and Lignan Components. Curr. Dev. Nutr. 2022, 6, 342. [Google Scholar] [CrossRef]
- Christofidou-Solomidou, M.; Pietrofesa, R.; Arguiri, E.; McAlexander, M.A.; Witwer, K.W. Dietary Flaxseed Modulates the MiRNA Profile in Irradiated and Non-Irradiated Murine Lungs: A Novel Mechanism of Tissue Radioprotection by Flaxseed. Cancer Biol. Ther. 2014, 15, 930–937. [Google Scholar] [CrossRef]
- Lee, J.C.; Krochak, R.; Blouin, A.; Kanterakis, S.; Chatterjee, S.; Arguiri, E.; Vachani, A.; Solomides, C.C.; Cengel, K.A.; Christofidou-Solomidou, M. Dietary Flaxseed Prevents Radiation-Induced Oxidative Lung Damage, Inflammation and Fibrosis in a Mouse Model of Thoracic Radiation Injury. Cancer Biol. Ther. 2009, 8, 47–53. [Google Scholar] [CrossRef]
- Christofidou-Solomidou, M.; Tyagi, S.; Tan, K.S.; Hagan, S.; Pietrofesa, R.; Dukes, F.; Arguiri, E.; Heitjan, D.F.; Solomides, C.C.; Cengel, K.A. Dietary Flaxseed Administered Post Thoracic Radiation Treatment Improves Survival and Mitigates Radiation-Induced Pneumonopathy in Mice. BMC Cancer 2011, 11, 1–15. [Google Scholar] [CrossRef]
- Yau, A.; Lee, J.; Chen, Y. Nanomaterials for Protein Delivery in Anticancer Applications. Pharmaceutics 2021, 13, 1–23. [Google Scholar] [CrossRef]
- Lila, A.S.A.; Ishida, T. Liposomal Delivery Systems: Design Optimization and Current Applications. Biol. Pharm. Bull. 2017, 40, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Muscaritoli, M.; Corsaro, E.; Molfino, A. Awareness of Cancer-Related Malnutrition and Its Management: Analysis of the Results From a Survey Conducted Among Medical Oncologists. Front. Oncol. 2021, 11, 1669. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H. Nutritional Issues in Patients with Cancer. Intest. Res. 2019, 17, 455–462. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Flaxseed proteins composition. Note: percentages may vary depending on the source and processing of the flaxseed.
Figure 1.
Flaxseed proteins composition. Note: percentages may vary depending on the source and processing of the flaxseed.
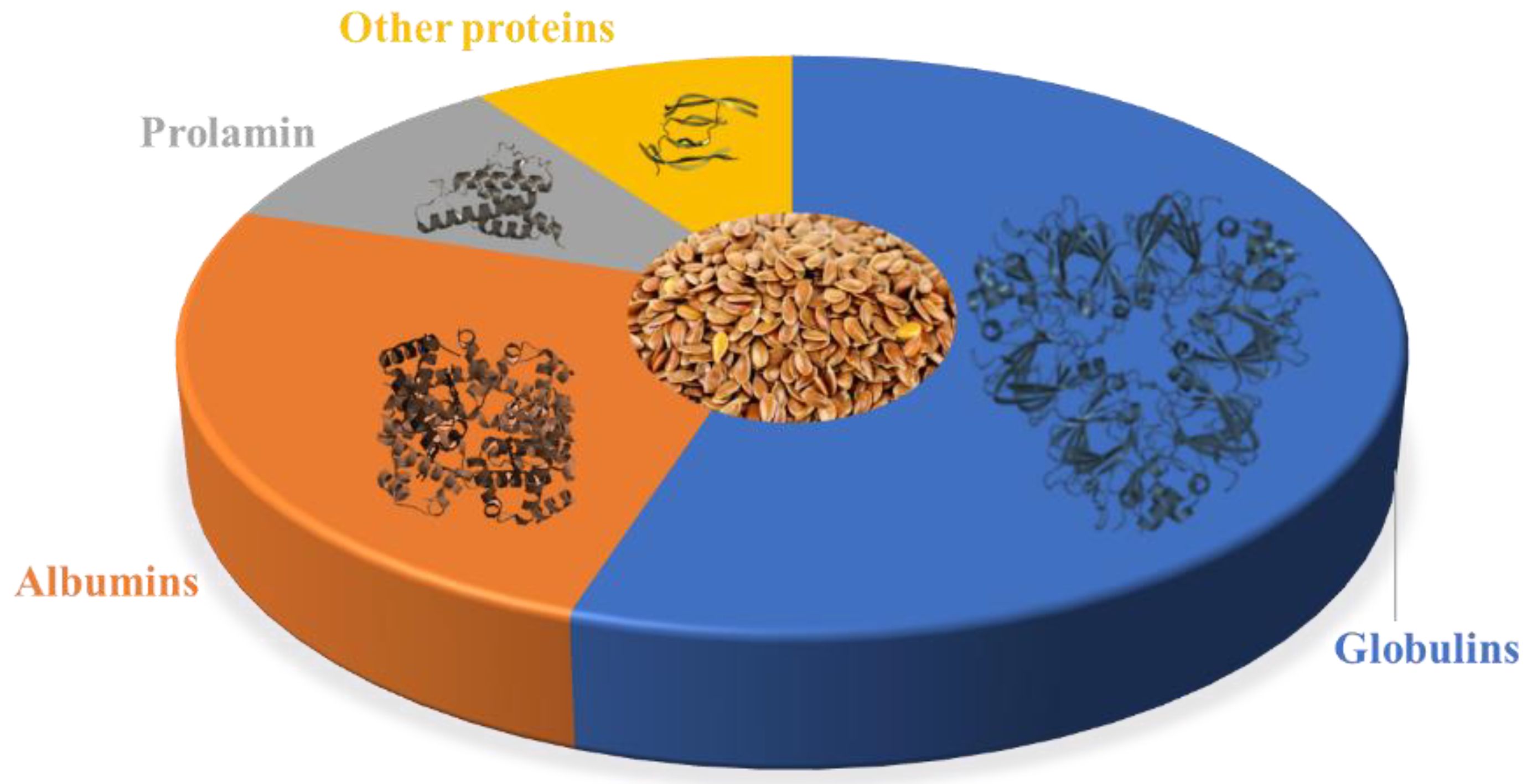
Figure 2.
Isolating and characterizing amino acids from flaxseed proteins.
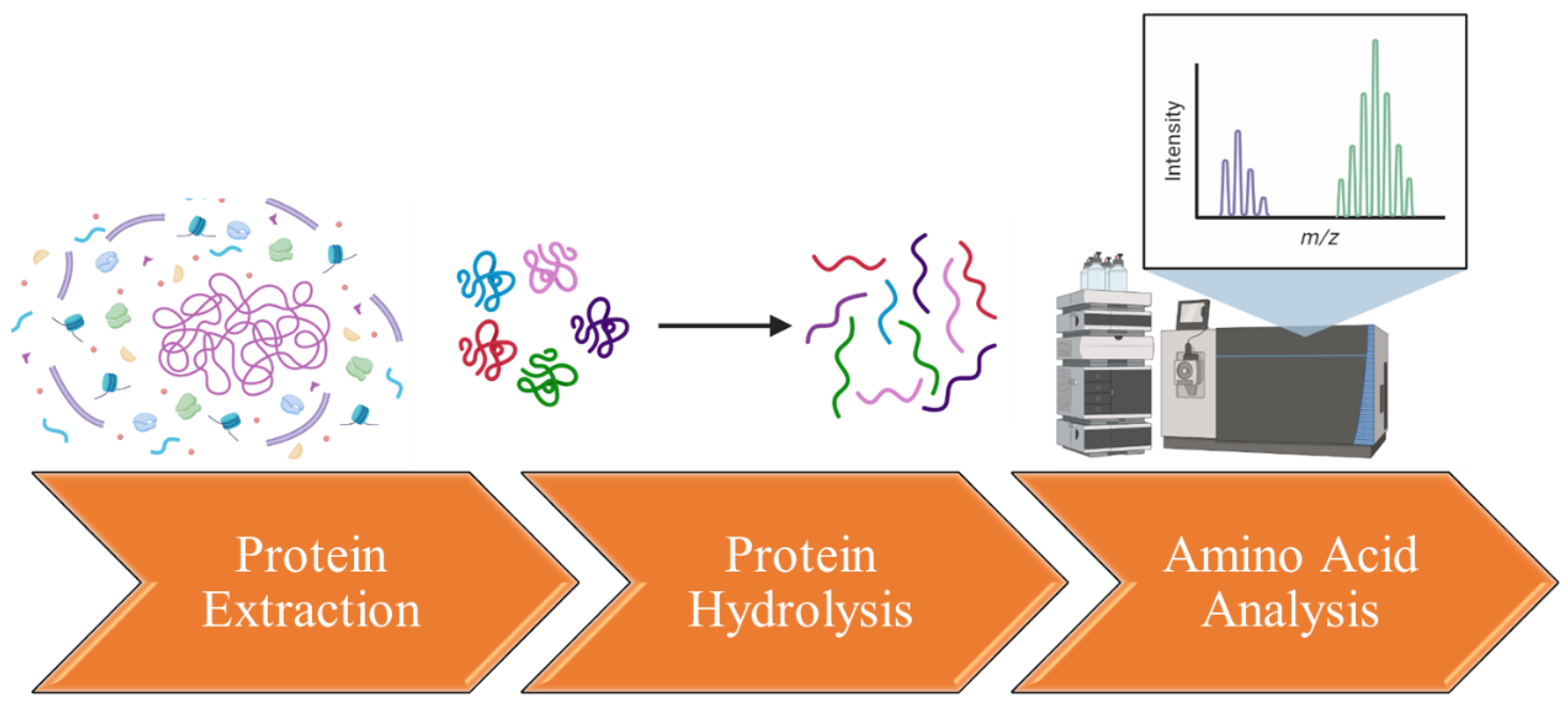
Figure 3.
The main flaxseed proteins profiling and sequencing. Sequence comparison and alignment was performed using Basic Local Alignment Search Tool (BLAST). A and B respectively – alignment of 2S albumins profiles and 11S globulins profiles from various plant sources. Alignment columns with no gaps are colored in blue or red. The red color indicates highly conserved columns and blue indicates less conserved ones. C and D – sequences of 2S albumins and 11S globulins respectively.
Figure 3.
The main flaxseed proteins profiling and sequencing. Sequence comparison and alignment was performed using Basic Local Alignment Search Tool (BLAST). A and B respectively – alignment of 2S albumins profiles and 11S globulins profiles from various plant sources. Alignment columns with no gaps are colored in blue or red. The red color indicates highly conserved columns and blue indicates less conserved ones. C and D – sequences of 2S albumins and 11S globulins respectively.
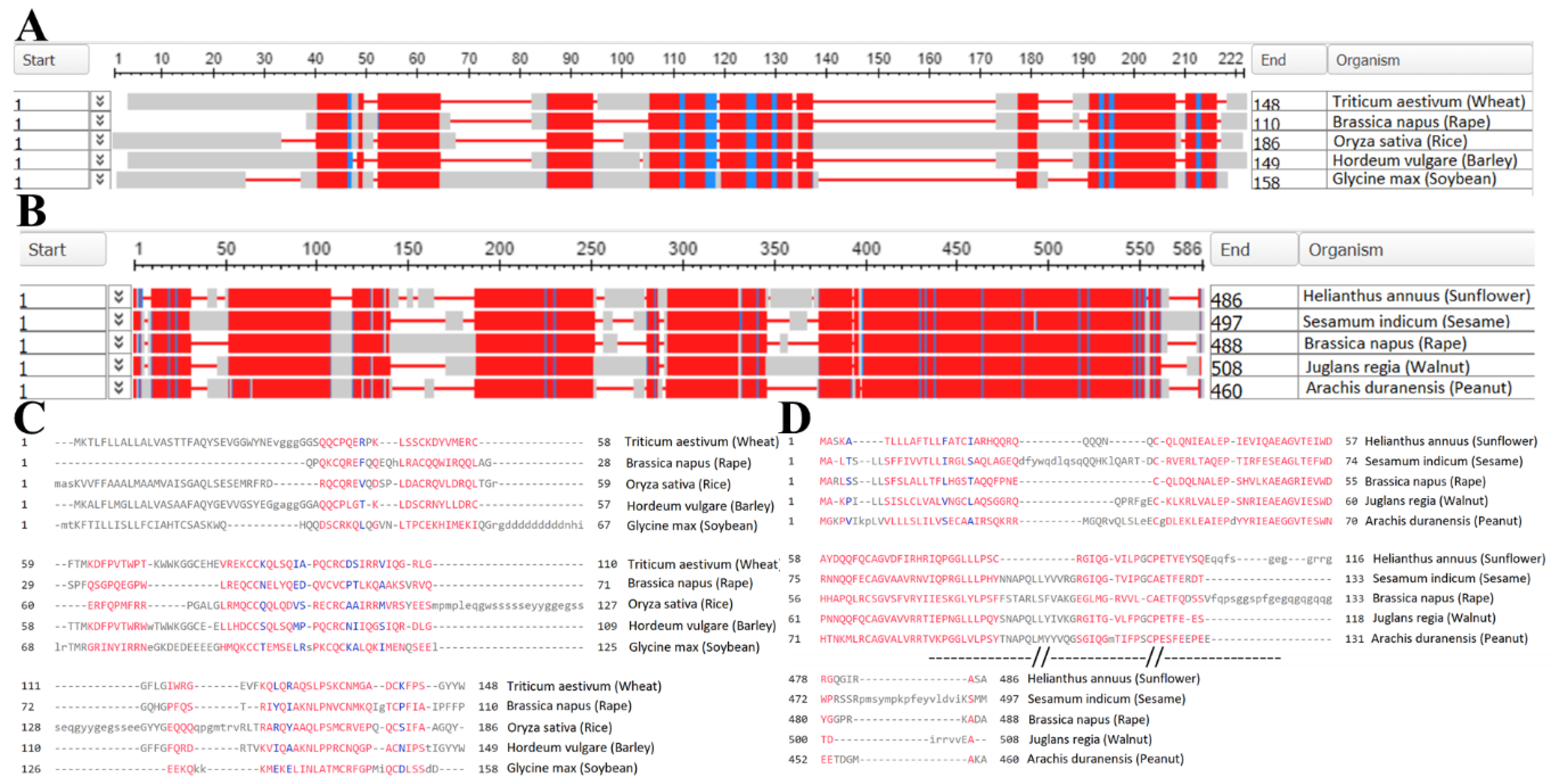
Figure 4.
Anti-cancer action of main flaxseed proteins.
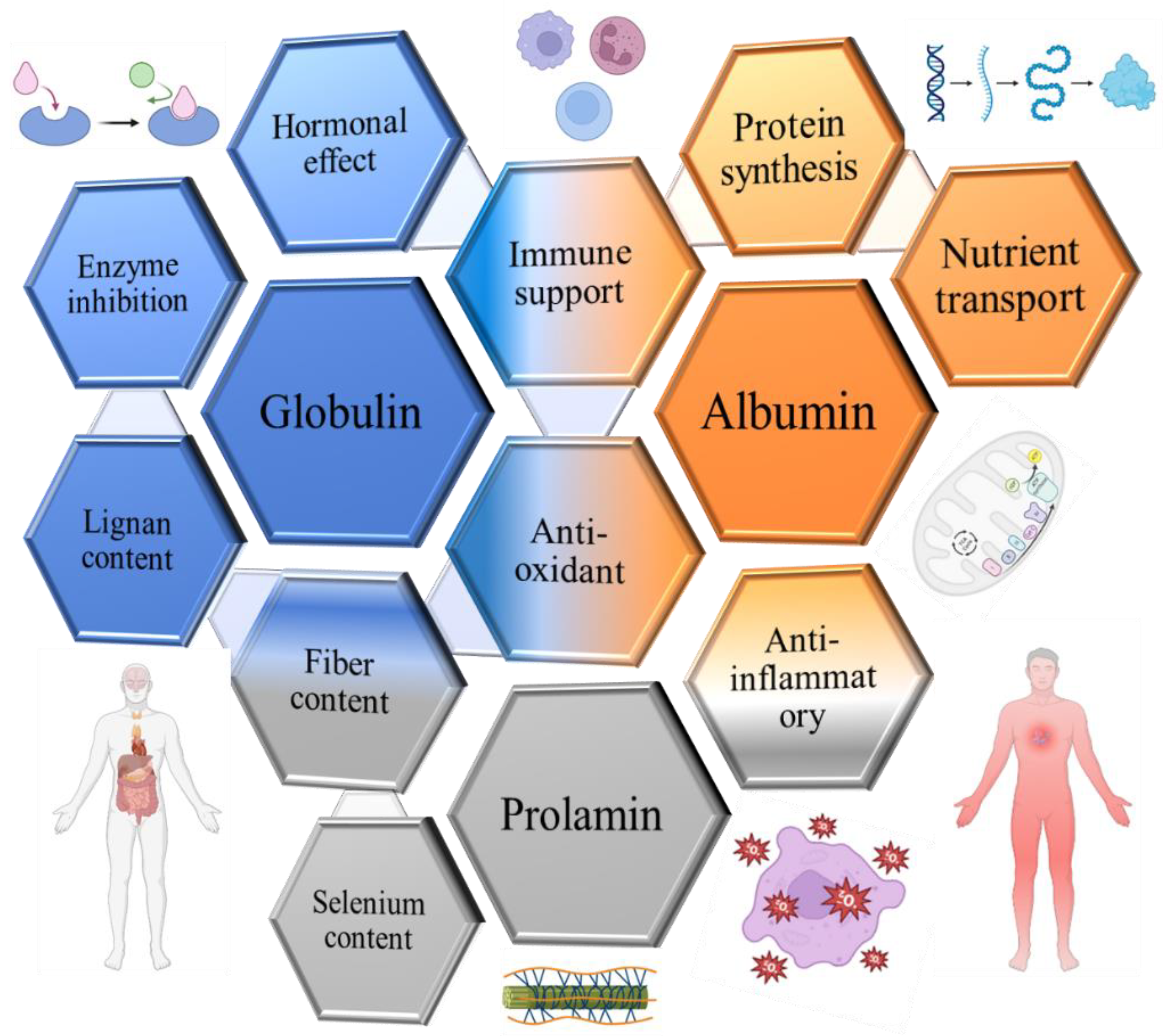
Figure 5.
Further research is required in order to establish cancer treatment with flaxseed proteins.
Figure 5.
Further research is required in order to establish cancer treatment with flaxseed proteins.
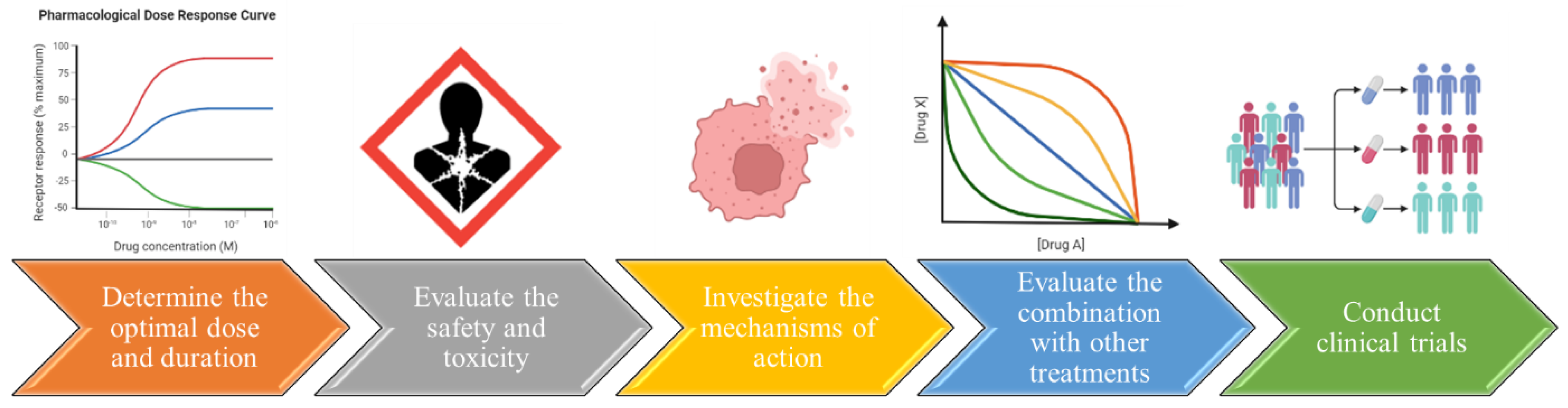
Table 1.
Proteome components found in flaxseed, and their potential heals benefits. Prepared using RCSB Protein Data Bank software.
Table 1.
Proteome components found in flaxseed, and their potential heals benefits. Prepared using RCSB Protein Data Bank software.
| Proteom Component | Structure | Percentage of Total Protein | Potential Health Benefits |
| 11S Globulin |  |
43-48% | Contains anti-cancer peptides [31,32,33]; may have anti-inflammatory and antioxidant effects [34,35] |
| 2S Аlbumin |  |
16-18% | Contains peptides with potential anti-cancer and anti-inflammatory effects [33,36,37]; may have cholesterol-lowering effects [38] |
| 7S Vicilin-Like Globulin | 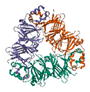 |
11-13% | May have antioxidant and anti-inflammatory effects [35,39]; contains peptides with potential anti-cancer effects [40] |
| Prolamin |  |
4-6% | Contains peptides with potential anti-cancer effects [41,42] |
| Glutelin-Like Protein |  |
2-3% | May have antioxidant and anti-inflammatory effects [43,44] |
| Other Proteins | 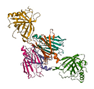 |
10-15% | May include lignans, which have antioxidant and anti-inflammatory and thus potential anti-cancer effects [45,46] |
Table 2.
Anti-metastatic action of 2S albumin component - Lunasin.
| Action | Description | Evidence |
| Inhibits cancer proliferation and induce apoptosis | Lunasin is critical for cellular internalization and activity against cancer-stem cells through interaction with the αvβ3 integrin via the FAK/ERK/NF-κB signaling pathway [64,65] | Lunasin inhibited the proliferation and the tumorsphere-forming capacity of HCT-116 cells. Lunasin induced apoptosis through activation of caspase-3 and cleavage of PARP, and could modulate cell cycle progress through the cyclin-dependent kinase inhibitor p21 [66,67] |
| Inhibits cancer cell invasion and migration | Lunasin can interact with cellular receptors, such as integrins and EGFR, and disrupt the activity of key signaling proteins, such as matrix metalloproteinase-2/-9 via the FAK/Akt/ERK and NF-κB signaling pathways that promote cancer cell invasion and spread [68] | Lunasin caused an increase in the expression of the inhibitor of kappa B alpha (IκB-α), a decrease in nuclear p50 NF-κB and a reduction in the migration of HCT-116 and KM12L4 colon cancer cells [65]. It was demonstrated that Lunasin effectively inhibited the migration and invasion activity and expression of matrix metalloproteinase (MMP)-2/-9 in MDA-MB-231 and MCF-7 breast cancer cell lines [68] |
| Reduces inflammation and oxidation | Lunacin has anti-inflammatory properties that may help to reduce inflammation, which is known to contribute to cancer development and progression [69,70] | The anti-inflammatory effects were demonstrated in mouse macrophage cells (LPS-stimulated RAW 264.7), by reducing the production of certain inflammatory markers (ROS, TNF-α and IL-6) [71]. |
| Reduces cancer cells colonization | Lunasin reduced activating phosphorylation of the intracellular kinases FAK and AKT as well as reduced histone acetylation of lysine residues in H3 and H4 histones [62] | Mice receiving Lunasin treatment had significantly reduced pulmonary colonization after injection of highly metastatic B16-F10 melanoma cells compared to mice in the control group [62,72] |
| Arrest cell cycle | Lunasin arrested the cell cycle at the G2/M and G1/S phase in various cancer types. Lunasin altered the expression of the G1 specific cyclin-dependent kinase complex components, increased levels of p27Kip1, reduced levels of phosphorylated Akt [67,73] | NSCLC H661 cells showed that Lunasin inhibited cell cycle progression at the G1/S phase [73]. Lunasin increased twice the amount of colon cancer cells KM12L4 undergoing apoptosis by arresting G2/M phase [67]. Lunasin treatment of MDA-MB-231 breast cancer cells resulted in a notable increase of RB1 level, which lead to arrest of G1 phase [74] |
Table 3.
Anti-tumor action of 11S globulin components.
| Mechanism of Action | Description | Evidence |
| Inhibits cancer cell growth and proliferation | 11S-globulin peptides in the more active fraction (MPI-h(V)) also presented antiproliferative activity [87]. Acid fraction of glycinin, composed of low molecular weight peptides is able to inhibit cancer cells growth [88]. | Studies have shown that 11S globulin inhibits the proliferation of UMR106 rat osteosarcoma-derived cells [87] and inhibited the growth of HeLa cells in a dose-dependent manner [88]. |
| Antioxidant effect | Small (~1kDa) peptides generated from 11S globulin demonstrated peroxyl radical scavenging activity [32] | The peptides were able to inhibit the formation of hydroxyl radicals by reaction of H2O2 and Co+2 in human adenocarcinoma cell line, Caco-2 [32] |
| Induces cancer cell death (apoptosis) | 11S globulin (glycinin) has been shown to induce cell death, or apoptosis, in cancer epithelial cells [89]. Flaxseed orbitides have been shown to be involved in apoptosis process [23]. | Glutelin extracts digested with trypsin, showed the induction of apoptosis against HeLa cells [90]. Human gastric SGC-7901 cell apoptosis was induced by flaxseed orbitides by a mitochondrial- and death receptor-mediated intrinsic and extrinsic pathways [91] |
| Arresting cell cycle | Cyclolinopeptides/linusorbs are capable to arrest cell cycle and thus reduce metastasis spreading. | flaxseed LOs arresting the cell cycle in SGC-7901 cells at the G1 phase by downregulation of CDK2/4 and overexpression of p21WAF1/CIP1 and p27KIP1 genes [92] |
| Cytotoxic effect on cancer cells | Cyclolinopeptides/linusorbs (LOs), which are cyclic peptides with 8–10 amino acid residues have strong cytotoxic effect over cancer cells [23]. 11S globulin was shown to have the cytotoxic effect at high concentration on a variety of human cancer cell lines [33]. | LOs from flaxseed globulin are cytotoxic to human melanoma cells A375 and breast cancer cells Sk-Br-3 and MCF7 at high concentration [93] |
Table 5.
Coverage of main flaxseed proteins from plant sources by currently existent databases. Protein`s synonyms are listed below the table.
Table 5.
Coverage of main flaxseed proteins from plant sources by currently existent databases. Protein`s synonyms are listed below the table.
|
Proteins Plants |
11S Globulin | 2S Albumin | 7S Vicilin-like protein | Prolamin |
| Linum Usitatissimum (Flax) | 0 | 12 | 0 | 0 |
| Triticum Aestivum (Wheat) | 3397 | 3944 | 22 | 2925 |
| Brassica Napus (Rape) | 136 | 2252 | 57 | 998 |
| Oryza Sativa (Rice) | 73 | 1920 | 20 | 215 |
| Hordeum Vulgare (Barley) | 286 | 539 | 2 | 284 |
| Glycine Max (Soybean) | 19 | 574 | 70 | 62 |
| Arachis Hypogaea (Peanut) | 15 | 342 | 13 | 29 |
| Avena Sativa (Oat) | 182 | 210 | 0 | 174 |
| Helianthus Annuus (Sunflower) | 34 | 181 | 2 | 8 |
| Sesamum Indicum (Sesame) | 9 | 105 | 13 | 4 |
| Juglans Regia (Walnut) | 8 | 102 | 4 | 3 |
| Carya Illinoinensis (Pecan) | 4 | 92 | 5 | 18 |
11S Globulin - Cruciferin; Glycinin; 12S seed storage globulin; 13S globulin seed storage protein; Legumin; Glutelin; Glutelin-like protein; Gliadin; Glutenin. 2S Albumin - 2S seed storage albumin protein; Conglutin; Conlinin; Glutenin; Gliadin; Puroindoline; Phytepsin; Hordoindoline; Hordein; Avenin; Vromindoline; Avenoindoline; Tryptophanin; Napin; Non-specific lipid-transfer protein. 7S Vicilin-like protein - Basic 7S globulin; Vicilin; Cupincin; Globulin-1 S; Beta-conglycinin; Leginsulin; PA1b; Plant functional proteins/peptides; γ-Conglutin; Albumin-1. Prolamin - Hordoindoline; Hordein; Gliadin; Avenin; Avenoindoline; Egg cell-secreted protein; Puroindoline; Serpin; Vromindoline; Tryptophanin.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



