
Submitted:
29 August 2023
Posted:
31 August 2023
You are already at the latest version
Abstract
Soybean seed sugars are among the most abundant beneficial compounds for human and animal
consumption in soybean seeds. Higher seed sugars such as sucrose are desirable as it contributes
to taste and flavor in soy-based food. Therefore, the objectives of this study were to use ‘Forrest’
by ‘Williams 82’ (F×W82) recombinant inbred line (RIL) soybean population (n=309) to identify
quantitative trait loci (QTL) and candidate genes that control seed sugar (sucrose, stachyose, and
raffinose) contents in two environments (North Carolina and Illinois) over two years (2018 and
2020). A total of 26 QTL that control seed sugars contents were identified and mapped on 16
soybean chromosomes (chrs.). Interestingly, five QTL regions were identified in both locations,
Illinois and North Carolina, in this study on chrs. 2, 5, 13, 17, and 20. Amongst 57 candidate genes
identified in this study, 16 were located within 10 Megabase (MB) of the identified QTL. Amongst
them a cluster of four genes involved in the sugars’ pathway was collocated within 6 MB with two
QTL that were detected in this study on chr. 17. Further functional validation of the identified
genes could be beneficial in breeding programs to produce soybean lines with high beneficial
sucrose and low raffinose family oligosaccharides.
Keywords:
soybean
; RIL
; Forrest
; Williams 82
; linkage map
; RFOs
; sucrose
; raffinose
; stachyose
; SNP
1. Introduction
Sugars including, sucrose, stachyose, glucose, raffinose, galactose, fructose, rhamnose, and starch, play a major role in seed and fruit development and seed desiccation tolerance (DT) [1,2,3,4]. Sucrose and raffinosaccharides (raffinose and stachyose), also called raffinose family oligosaccharides (RFOs), make up 5–7%, 1%, and 3–4% of total carbohydrates, respectively, of soybean seed dry weights [5]. RFOs are synthesized from sucrose by a series of additions of galactinol units and are involved in DT, freezing, stress tolerance, and seed longevity [6,7,8,9]. Galactinol synthase (GolS) is the key enzyme in the RFOs biosynthetic pathway converting galactinol and myo-inositol as the main precursors to form RFOs. Galactinol synthase (GolS) converts myo-inositol and UDP-galactose into galactinol, while sucrose and galactinol are converted into raffinose by the raffinose synthase [9,10]. In addition to being involved in stress tolerance, RFOs are reported to play a role in several signal transduction pathways [11], exports of specific mRNAs [12], and trafficking of certain vesical membranes [13].
Like most seed composition traits, seed sugars [4] are influenced by many factors, including abiotic and biotic stresses, and environmental factors, such as temperature, soil moisture, freezing, seed maturity, and growth conditions. [1,14,15,16,17,18,19]. It was shown that stachyose contents increased drastically in drying seeds but not in seeds kept at high humidity levels, which reveals the critical role of stachyose in DT [1]. The effect of water deficit (WD) on enzymes involved in sugar biosynthetic pathways in soybean nodules was investigated. Sucrose synthase activity declined drastically with increased WD while sucrose content increased [14]. Other studies showed that WD impacts negatively sucrose biosynthesis and translocation from sources to sinks more than other sugar (raffinose and stachyose) biosynthesis [16,19]. Investigating ‘Clark’ and ‘Harosoy’ near-isogenic lines (NILs) revealed that Clark’s sugars contents decreased with increased days of maturity for both cultivars while both positive and negative effects were observed concerning the effects of temperature in two different years (2004 and 2005) [15]. In 2004, seed sugars contents increased with temperature increase, while the contents decreased with increased temperatures in 2005 [15]. The effect of WD on several seed composition traits, including sugars on several Phomopsis susceptible and resistant soybean cultivars, was investigated. In fact, sugar (sucrose, raffinose, and stachyose) contents were higher in seeds of resistant maturity group III cultivars than their susceptible counterparts [16]. A recent study investigated the effect of soil moisture on seed sugars (sucrose, raffinose, stachyose) and starch contents among other compounds in two soybean cultivars in maturity group V (Asgrow, AG6332, and Progeny 5333RY) and showed that sucrose, stachyose, and raffinose contents in addition to the mineral nutrient (N, P, K, and Ca) contents decreased with increased soil moisture in both cultivars [17].
During the last decades, more than 53 QTL that control seed sucrose and RFOs, other sugar (glucose, galactose, fructose, fucose, rhamnose), and starch contents have been reported using different biparental and natural populations and mapping methods including single marker analysis, interval mapping (IM), composite interval mapping (CIM), and genome-wide association studies (GWAS) [18,20]. However, to our knowledge, only one of these studies identified candidate genes within these QTL regions [18,21]; The Glyma.01g127600 that encodes for a protein phosphatase on chr. 1, Glyma.03g019300 that encodes for a MADS-box protein, Glyma.03g064700 that encodes for a phosphatidylinositol monophosphate-5-kinase on chr. 3, and Glyma.06g194200 that encodes for a gibberellin-regulated protein on chr. 6 [18,21].
To improve seed quality, several attempts to manipulate seed sugars, phytic acid, and the content of other beneficial compounds have been conducted in recent years [22,23,24]. Monogastric animals (such as poultry and pigs) and humans do not produce α-galactosidase and cannot digest RFOs which reduces gastrointestinal performance, flatulence, and diarrhea. Therefore, reducing raffinose and stachyose and increasing sucrose in soybean seed content are desirable traits and the main goal in breeding programs [22,23,24,25,26,27]. The objective of this study was to genetically map QTL for seed sucrose, raffinose, and stachyose contents using the ‘Forrest’ by ‘Williams 82’ RIL population, in addition to identifying candidate genes involved in soybean seed sugars biosynthesis.
2. Materials and Methods
2.1. Plant Materials
The ‘Forrest’ × ‘Williams 82’ RIL population (F×W82, n=309) was previously studied and described in detail in our previous research [28,29]. The parents and RILs were evaluated in two locations: Spring Lake, NC (35.17° N, 78.97° W, 2018) and Carbondale, IL (37° N, 89° W, 2020). Details about growth conditions, crop management, and seed harvesting were carried out as described earlier [28,29].
2.2. Seed Sugars Quantification
RILs and parents (Forrest and Williams 82) and soybean germplasm seeds were harvested at maturity, and sugars (sucrose, raffinose, and stachyose) contents (%) were quantified using near-infrared reflectance (NIR) with an AD 7200 array feed analyzer (Perten, Springfield, IL) as described earlier [15,30].
2.3. DNA Isolation, SNP Genotyping, and Genetic Map Construction
Parents and RILs genomic DNA was extracted by cetyltrimethylammonium bromide (CTAB) method as previously described [31]. A NanoDrop spectrophotometer (NanoDrop Technologies Inc., Centreville, DE) was used to quantify DNA samples (50 ng/µl), and genotyping was done using the Illumina Infinium SoySNP6K BeadChips (Illumina, Inc. San Diego, CA) as described earlier [15] at the Soybean Genomics and Improvement Laboratory (USDA-ARS, Beltsville, MD 20705). The F×W82 genetic linkage map was constructed using JoinMap 4.0 [28,32] as previously described to detect QTL for seed isoflavones [28] and seed tocopherols contents [29].
2.4. Sugars QTL Detection
WinQTL Cartographer [33] Interval mapping (IM) and composite interval mapping (CIM) methods were used to identify QTL that control seed sugars (sucrose, stachyose, and raffinose) contents in this RIL population. The following parameters (Model 6, 1 cM step size, 10 cM window size, 5 control markers, and 1,000 permutations) have been used, and chromosomes were drawn using MapChart 2.2 [34].
2.5. Sugars Biosynthesis Candidate Genes Identification
The Glyma numbers of the sucrose and RFOs biosynthesis genes were obtained by reverse BLAST of the genes underlying the RFOs pathway in Arabidopsis using the available data at SoyBase. The sequences of the Arabidopsis genes were obtained from the Phytozome database (https://phytozome-next.jgi.doe.gov ; accessed on 08/15/2023). These sequences were used for Blast in SoyBase. The obtained genes that control the RFOs pathway were mapped to the identified sugars QTL.
2.6. Expression Analysis
The expression analysis of the identified candidate genes was performed using the publicly available data from SoyBase [20] to produce the expression profiles of these candidate genes in the soybean reference genome Williams 82 in the Glyma1.0 Gene Models version.
2.7. Comparison of the Williams 82 and Forrest Sequences
Sequences of Forrest and Williams 82 cv. were obtained from the variant calling and haplotyping analysis that was performed using the 108 soybean germplasm sequenced lines as described previously [35].
3. Results
3.1. Sugars Frequency Distribution
The frequency distributions among sucrose, raffinose, and stachyose contents were quite different in the F×W82 RIL population based on Shapiro–Wilk’s method for the normality test. Raffinose (2018), stachyose (2018), and sucrose (2020) were normally distributed. Only positive or negative skewness were identified in the RIL population, and all kurtosis values of these variables were positive (Table 1; Figure 1). In terms of coefficient of variation (CV), the value of sucrose 2018 showed the highest percentage of variation (62.86%) compared to other assessed traits, and the rest of the CVs appeared to be less varied within these two years. The histogram of sucrose 2018 was extremely skewed, and the other traits evaluated were normally distributed.
The broad-sense heritability (h2b) of seed sugar weight for sucrose, raffinose, and stachyose contents across two different environments appeared quite different. Stachyose had the highest heritability (92%), and the h2b for sucrose was 36.8% (Table 2). However, no negative h2b values for sugar contents were observed. The RILs-Year interactions still played a significant role in the molecular formation among three sugar contents in soybean seeds, The Sum Sq and Mean Sq to determine σG2 and σGE2 for each trait (Table 2) using type I sum of squares (ANOVA (model)) function in R program were implemented.
3.2. Sugars Contents QTL
IM and CIM have been used to identify QTL for seed sugar contents in this FxW82 RIL population; however, only QTL identified by CIM are presented here, although the QTL identified by the IM method were still reported in Tables S1 and S2. A total of 26 QTL that control seed sugar contents have been identified in both NC-2018 (19 QTL) and IL-2020 (7 QTL) by CIM (Table 3 and Table 4; Figure S1).
In Spring Lake, NC in 2018 (NC-2018), 12 QTL that control seed sucrose content (qSUC-1–qSUC-12) have been identified and mapped on Chrs. 1, 2, 3, 4, 5, 6, 9, 10, 13, 17, 18, and 19; 4 QTL that control seed stachyose content (qSTA-1–qSTA-4) have been identified and mapped on Chrs. 13 and 19; and 3 QTL that control seed raffinose content (qRAF-1–qRAF-3) have been identified and mapped on Chr. 9 and 12 (Table 3 and Table 5; Figure S1). Likewise, in Carbondale, IL in 2020 (IL-2020), 3 QTL that control seed sucrose content (qSUC-1–qSUC-3) have been identified and mapped on Chrs. 2, 5, and 8; and 4 QTL that control seed stachyose content (qSTA-1–qSTA-4) have been identified and mapped on Chrs. 13, 16, 17, and 20 (Table 4 and Table 6; Figure S1). No QTL that controls seed raffinose content have been identified in this location.
No QTL for seed sugar contents have been identified by other studies within the QTL regions on chr. 4 (qSUC-4-NC-2018, 6.5–16.5 cM), chr. 10 (qSUC-8-NC-2018, 214.1–216.1 cM), and chr. 18 (qSUC-11-NC-2018, 20.1–17.5 cM), which indicates they are novel QTL regions.
3.3. In silico Sucrose, Raffinose and Stachyose Biosynthetic Pathway Genes in Soybean
The sugars (sucrose, raffinose, and stachyose) biosynthetic pathway was studied in many plants, including the plant model Arabidopsis thaliana [36,37] and the leguminous model Medicago sativa L. [38]. A reverse BLAST of the genes identified in Arabidopsis thaliana was conducted using the SoyBase [20] to reconstruct the sugars (sucrose, raffinose, and stachyose) biosynthetic pathway in soybean (Figure 2).
A total of fifty-seven candidate genes were identified to underly the sugar (sucrose, raffinose, and stachyose) biosynthetic pathway (Figure 2). In this pathway, twelve candidate genes were identified for invertase including Glyma.05G185500, Glyma.20G177200, Glyma.08G043800, Glyma.10G214700, Glyma.08G143500, Glyma.05G236600, Glyma.17G037400, Glyma.10G145600, Glyma.20G095200, Glyma.07G236000, Glyma.02G016700, and Glyma.10G017300. Twelve candidate genes were identified for sucrose synthase including Glyma.02G240400, Glyma.03G216300, Glyma.09G073600, Glyma.09G167000, Glyma.13G114000, Glyma.14G209900, Glyma.15G151000, Glyma.16G217200, Glyma.17G045800, Glyma.19G212800, Glyma.11G212700, and Glyma.15G182600. Twelve candidate genes were identified for UDP-D-Glucose-4-Epimerase, Glyma.08G023100, Glyma.01G225800, Glyma.05G204700, Glyma.05G217100, Glyma.07G237700, Glyma.07G271200, Glyma.08G011800, Glyma.11G017100, Glyma.12G162600, Glyma.17G035800, Glyma.18G145700, and Glyma.18G211700. For the galactinol synthase, six candidate genes were identified, including Glyma.03G222000, Glyma.03G229800, Glyma.10G145300, Glyma.19G219100, Glyma.19G227800, and Glyma.20G094500. Fourteen candidate genes were identified for raffinose synthase, Glyma.03G137900, Glyma.04G145800, Glyma.19G140700, Glyma.04G190000, Glyma.02G303300, Glyma.05G003900, Glyma.06G175500, Glyma.09G016600, Glyma.13G160100, Glyma.14G010500, Glyma.17G111400, Glyma.19G004400, Glyma.05G040300, and Glyma.06G179200. For the stachyose synthase, only one candidate gene was identified Glyma.19G217700 (Figure 2).
3.4. Association between the Identified sugar (sucrose, raffinose, and stachyose) Biosynthetic Pathway Candidate Genes and Reported QTL
The identified genes for the sugar (sucrose, raffinose, and stachyose) biosynthesis in soybean have been mapped to the identified QTL. Amongst fifty-seven candidate genes, sixteen have been located less than 10 MB to the identified QTL on chrs. 2, 5, 6, 8, 9, 10, 17, and 19 (Table 3, Table 4, Table 5 and Table 6).
The sucrose synthase candidate gene Glyma.09G073600 and the raffinose synthase candidate gene Glyma.09G016600 are positioned close to the qSUC-7-IL-2018, qRAF-1-IL-2018, and qRAF-2-IL-2018 on Chr.9 (Table 3, Table 4, Table 5 and Table 6). The invertase candidate gene Glyma.02G016700 is located 3.6 and 0.2 MB apart from the qSUC-1-IL-2018 and qSUC-1-NC-2020, respectively, on Chr. 2 (Table 3, Table 4, Table 5 and Table 6). The raffinose synthase candidate genes Glyma.05G003900 and Glyma.05G040300 are located close to the qSUC-5-IL-2018 and qSUC-2-NC-2020 on Chr. 5 (Table 3, Table 4, Table 5 and Table 6). On chr. 6, the raffinose synthase candidate gene Glyma.06G175500 is located close to the qSUC-6-IL-2018 (Table 3, Table 4, Table 5 and Table 6). The invertase candidate genes Glyma.08G043800, and Glyma.08G143500; and the UDP-D-Glucose-4-Epimerase candidate genes Glyma.08G011800 and Glyma.08G023100 on chr. 8 are located close to the qSUC-3-NC-2020 (Table 3, Table 4, Table 5 and Table 6, Table S3 and S4). On chr. 10, the invertase candidate gene Glyma.10G017300 is located close to the qSUC-8-IL-2018 (Table 3, Table 4, Table 5 and Table 6). On Chr. 17, a cluster of four genes involved in the sugars’ pathway is collocated within 6 MB with two QTL (qSUC-10-NC-2018 and qSTA-3-IL-2020) that were identified in this study. These genes are the Glyma.17G037400 encoding for an invertase, Glyma.17G045800 encoding for sucrose synthase, Glyma.17G111400 encoding for raffinose synthase, and Glyma.17G035800 encoding for UDP-D-glucose-4-epimerase (Table 3, Table 4, Table 5 and Table 6, Figure S3.). The raffinose synthase candidate gene Glyma.19G004400 is positioned close to the qSTA-3-IL-2018 and the qSTA-4-IL-2018 (Table 3, Table 4, Table 5 and Table 6), as well as the qRAF-8-IL-2018 and qRAF-9-IL-2018 identified using the IM method (Table 3 and Table 4).
3.5. Association between the Identified Candidate Genes and the Previously Reported QTL
Several studies have identified and mapped QTL underlying seed sugar content using different populations and mapping methods [39,40,41,42], as summarized in [18].
The identified genes have been mapped to the previously reported QTL regions associated with the seed sugar content using the data from SoyBase [18,20,43] six candidate genes have been located within the identified seed sugars QTLs and 18 have been located < 9 MB apart from these regions (Table 7). Among them is the invertase candidate gene Glyma.08G143500 that is located within the seed sucrose 1-2 QTL on Chr. 8 [20,39]. Also, the galactinol-sucrose galactosyl-transferase 6-related candidate gene Glyma.13G160100 is situated within the seed sucrose 1-5 QTL [20,39](Table 7). Likewise, the raffinose synthase candidate gene Glyma.19G140700 is collocated within the seed sucrose 1-8 QTL [20,39], less than < 0.5 MB apart from seed sucrose 2-11 and seed sucrose 2-10 [20,41], and 1.9 MB from seed oligosaccharide 2-7 [20,40].
The sucrose synthase candidate gene Glyma.02G240400 was located close (< 1.5 MB) to two QTL controlling seed sugar contents, the seed sucrose 2-2 and seed oligosaccharide 1-1 [20,41]. Moreover, the raffinose synthase candidate gene Glyma.05G003900 is located less than < 4 MB apart from the seed sucrose 1-1 [20,39]. The raffinose synthase candidate gene Glyma.19G004400 is located less than 9 MB apart from four QTL controlling the sugar contents, namely seed sucrose 2-3, seed oligosaccharide 1-2, seed sucrose 2-6, and seed oligosaccharide 1-5 [20,41] (Table 7). On chr. 8, the seed sucrose 1-3 and seed sucrose 1-13 are located close to the invertase candidate genes Glyma.08G043800, and Glyma.08G143500; as well as UDP-D-glucose-4-epimerase candidate genes Glyma.08G011800 and Glyma.08G023100 [20,39] (Table 7). The sucrose synthase candidate gene Glyma.09G073600 and the raffinose candidate gene Glyma.09G016600 are positioned less than < 2 MB apart from the seed sucrose 4-2 [20,44] (Table 7). Interestingly, the sucrose synthase candidate genes Glyma.15G182600 and Glyma.15G151000 are located less than < 1.25 MB from the seed sucrose 3-3 and seed oligosaccharide 2-3 [20,40].
3.6. Organ-specific Expression of the Identified Candidate Genes
The expression pattern of the identified candidate genes was investigated in Williams 82 cv. using the RNA-seq data available at SoyBase [20]. The dataset includes several plant tissues, including leaves, nodules, roots, pods, and seeds (Figure 3A, 3B, and S2). Four of the fifty-seven identified candidate genes have no available RNA-seq data, including the sucrose synthase candidate genes Glyma.03G216300, Glyma.17G045800, and Glyma.19G212800, as well as the UDP-D-glucose-4-epimerase candidate genes Glyma.18G211700 (Figure S2). The raffinose synthase candidate gene Glyma.04G145800 is not expressed in any of the analyzed tissues, whilst the rest of the identified genes showed different expression patterns.
The sucrose synthase candidate genes Glyma.09G073600 and Glyma.13G114000 present a high expression profile in all the analyzed tissues except for the young leaves, while the raffinose synthase candidate gene Glyma.17G111400 is abundantly expressed in all the analyzed tissues except for the seeds and nodules. Interestingly, the sucrose synthase candidate gene Glyma.15G182600 is highly expressed in all the tissues excluding the young leaves and the nodules. The raffinose synthase candidate gene Glyma.03G137900 is abundantly expressed in flowers, nodules, and roots. The raffinose synthase candidate gene Glyma.14G010500 and the invertase candidate gene Glyma.05G236600 are highly expressed in the flowers and pods. Also, The UDP-D-glucose-4-epimerase candidate gene Glyma.05G204700 is abundantly expressed in the flowers and seeds. While the invertase candidate gene Glyma.20G177200 is highly expressed in nodules and roots, the raffinose synthase candidate gene Glyma.06G179200 was found to be highly expressed in seed (Figure 3A, Figure S2).
Seventeen of the identified candidate genes were situated less than 10 MB apart from the identified QTL regions. Glyma.09G073600 is highly expressed in seeds in Williams 82 cv., followed by Glyma.17G111400, Glyma.17G035800, and Glyma.08G043800 with moderated expression profile. The remaining genes have lower expression patterns, excluding the Glyma.02G016700, Glyma.06G175500, Glyma.09G016600, Glyma.10G017300, and Glyma.19G004400 genes that are not expressed in seeds in Williams 82 cv.
3.7. Comparison of the Williams 82 and Forrest Sequences
The sequences of the candidate genes that are located less than 10 MB from the identified QTL were compared. The results have shown that six of them have SNPs and InDels between Forrest and Williams 82 sequences, Glyma.09G073600, Glyma.08G143500, Glyma.05G003900, Glyma.17G035800, Glyma.17G111400, and Glyma.09G016600 ( S4, Figure 4).
The sucrose synthase Glyma.09G073600 has in total 28 SNPs and 7 InDels; three of these SNPs are located upstream the 5’UTR, two are downstream the 3’UTR, and seven are located in the exons (Table S4, Figure 4). For the invertase candidate gene Glyma.08G143500, there are 20 SNPs and 5 InDels. One of these SNPs is located in exon 7, causing a missense mutation, and two SNPs are located upstream the 5’UTR (Table S4, Figure 4). The raffinose synthase candidate gene Glyma.05G003900 has 9 SNPs and one InDel, four of those SNPs are in the exons, from which two SNPs resulted in missense mutations (Table S4, Figure 4). Likewise, the raffinose synthase candidate gene Glyma.09G016600 possesses 12 SNPs and 2 InDels. Amongst these SNPs, there are two located in exons that resulted in missense mutations in addition to the 2 InDels located in the exons (Table S4, Figure 4). For the raffinose candidate gene Glyma.17G111400, 8 SNPs were found from which one is located upstream the 5’ UTR, another one is downstream the 3’UTR, and the last six are in exons causing silent mutations (Table S4, Figure 4). Finally, the UDP-D-Glucose-4-Epimerase candidate gene Glyma.17G035800 has two SNPs that are positioned in introns (Table S4).
4. Discussion
Soybean seed sugars play a major role in seed and fruit development. Recently, soy products became very popular as a result of a growing demand for vegan diets [45]. The quality and taste of these products are determined by soybean seed sugar content [39]. These sugars include sucrose, raffinose, and stachyose that make up to 5–7%, 1%, and 3–4% of total carbohydrates, respectively [5]. However, the raffinose and stachyose fermentation by humans and monogastric animal intestines microbes leads to a reduced gastrointestinal performance, flatulence, and diarrhea. Thus, reducing raffinose and stachyose and increasing sucrose in soybean seed content are desirable[22,27].
Knowing the importance of soybean seed sucrose content in the quality of the soybean based products for food and feed, breeding programs are focused on developing soybean seeds with high sucrose content and low RFOs content [43,46]. Thus, soybean varieties with high sucrose content are valuable for soybean food and feed products [47].
The identification of QTL associated with sugars components using different types of molecular markers is one of the breeding process approaches that researches use to breed for a high sucrose soybean.
In the current study, all seed sugar (sucrose, raffinose, and stachyose) phenotypic data exhibited normal distributions in all environments studied (years and locations), showing that these traits are polygenic and complex as shown earlier [21,39,40,41,44,47,48,49,50,51,52,53].
The SNP-based genetic linkage map facilitated the identification of several QTL including QTL for seed isoflavone contents [28], seed tocopherol contents [29], and seed sugar (sucrose, stachyose, and raffinose) contents as reported in the current study.
A total of 26 QTL that control seed sugar contents have been identified in both IL-2018 and NC-2020 by CIM. Among these, three are novel QTL regions, including qSUC-4, qSUC-8, and qSUC-11 mapped on chrs. 4, 10, and 18, respectively. All the other sugar QTL reported in this study have been located within or very close to other sugar QTL previously reported [30,39,40,41,44] as summarized in [18]. Five other genomic regions on chrs. 2, 6, 12, 16, and 19 harboring sugar QTL either from this study or from other studies are of particular interest. On chr. 2, qSUC-2-NC-2018 may correspond to suc 1-1 identified previously [39]. This QTL region contains the Glyma.02G016700 candidate gene that encodes for invertase.
Interestingly, several QTL have been identified previously including a QTL that controls seed raffinose content within the qSUC-1-NC-2018 region (chr. 1) [30], two QTL (suc 2-2 and suc 3-2) that control seed sucrose content within the qSUC-2-NC-2018 region (chr. 2) [20,40,41], a QTL that controls seed sucrose content (suc-001) within the qSUC-3-NC-2018 region (chr. 3), [30]; 2 QTL that control seed sucrose (suc 1-1 and suc 4-1) content within the qSUC-5-NC-2018 region (chr. 5) [39,44]; a QTL that controls seed raffinose content (raf003 and raf004) within the qSUC-6-NC-2018 and qSUC-7-NC-2018 regions (chrs. 6 and 9), [30]; a QTL that controls seed sucrose (suc 1-5) content within the qSUC-9-NC-2018 region (chr. 13), [39]; and a QTL that controls seed sucrose (suc 1-4) content within the qSUC-12-NC-2018 region (chr. 20) [39].
Likewise, several other QTL have been identified previously a QTL that controls seed sucrose (suc 2-2, 3-2) content within the qSUC-1-IL-2020 region (chr. 2)[40,41]; QTL that control seed sucrose (suc 1-1, 4-1) content within the qSUC-2-IL-2020 region (chr. 5) [39,44]; and within qSUC-3-IL-2020 region on chr. 8, QTL that control seed sucrose (suc 1-2, 1-3, 1-13) content within the qSUC-3-IL-2020 region (chr. 8)[39]. Within the QTL regions that were found to control seed stachyose contents (qSTA-1-IL-2020, qSTA-2-IL-2020, and qSTA-4-IL-2020) reported in the current study on chrs. 13, 16, and 19, several QTL that control seed sucrose (suc 1-4, 1-5, 3-5, 3-6) and seed raffinose (raff007) contents have been identified previously [39,40,41].
On chr. 6, qSUC-6-NC-2018 most likely corresponds to suc 2-2 [41] and raffinose (raf003) QTL regions identified previously [30,39]. The QTL region contains Glyma.06G175500 candidate gene encoding for raffinose synthase. Interestingly, the genomic region on chr. 19 comprising a cluster of sucrose QTL (suc 1-6 to 1-8, 2-3 to 2-11) [39,41] also contains two stachyose QTL identified in this study (qSTA-3-NC-2018 and qSTA-4-NC-2018). The candidate gene Glyma.19G004400, that also encodes for raffinose synthase was identified within this QTL region.
No candidate genes have been identified on chrs. 12 (qRAF-3-NC-2018), 16 (qSTA-2-NC-2018), and 20 (qSTA-4-NC-2018).
Remarkably, within the novel QTL regions reported here on chrs. 4, 10, and 18, seven candidate genes have been identified; including the Glyma.18G145700 encoding for UDP-D-glucose-4-epimerase on chr. 18 (Table 5 and Table 6, and Figure 2).
Interestingly, five QTL regions were detected in both locations, IL and NC; The first QTL region contains qSUC-5-NC-2018 and qSUC-2-IL-2020 that were detected in the same location on chr. 5. Additionally, the qSUC-9-NC-2018, qSTA-1-NC-2018, and qSTA-2-NC-2018 were located only 1 MB apart from qSTA-1-IL-2020 on chr.13. Moreover, qSUC-12-NC-2018 was 1.3 MB away from qSTA-4-IL-2020 on chr. 20. Furthermore, qSUC-10-NC-2018 and qSTA-3-IL-2020 were positioned 3.1 MB apart from each other on chr. 17. Additionally, qSUC-2-NC-2018 and qSUC-1-IL-2020 were located ~4 MB apart on chr. 2. The QTL regions that were not detected in both locations may be affected by environmental conditions.
In a previous study [54], 31,245 SNPs and 323 soybean germplasm accessions grown in three different environments were used to identify 72 QTL associated with individual sugars and 14 associated with total sugar [54]. In addition, ten candidate genes that are within the 100 Kb flanking regions of the lead SNPs in six chromosomes were significantly associated with sugar content in soybean; eight of them are involved in the sugar metabolism in soybean [54]. Amongst these candidate genes, the raffinose synthase gene Glyma.05G003900 is also reported in this study.
A recent study used a RIL population from a cross of ZD27 by HF25 to identify 16 QTL controlling seed sucrose and soluble sugars content in soybean [43]. Amongst these QTL, qSU1701[43] with a LOD = 7.61 and phenotypic variation explained (PVE)= 16.8 % was identified on chr. 17 to be associated with the seed sucrose content. This QTL region is collocated with the qSUC-10-NC-2018 identified in this study for the same trait with a LOD = 33.2 and an R2= 20.5. On the same chr., qSS1701 [43] and qSS1702 identified to be associated with the seed soluble sugar content are collocated with the qSTA-3-IL-2020. These QTL are positioned within less than 8 MB with a cluster of four genes involved in the sugars’ pathway, including the Glyma.17G037400 encoding for invertase, Glyma.17G045800 encoding for sucrose synthase, Glyma.17G111400 encoding for raffinose synthase (showing 7 SNPs variations in exons) (Figure 4), and Glyma.17G035800 encoding for UDP-D-glucose-4-epimerase. Our results confirm that this region on chr. 17 is a major QTL associated with seed sugars content in soybean. In the same study [43], qSU2001 identified on chr. 20 with LOD=3.38 and PVE=5.6 % is collocated with the qSUC-12-NC-2018, and 0.3 MB apart from the qSTA-4-IL-2020. The invertase candidate gene Glyma.20G177200 is positioned within the qSU2002 [43] identified on chr. 20 with LOD=7.9 and PVE=14.4 %. These results confirm that this region on chr 20 is involved in soybean seed sugar contents. On chr. 3, qSS0301 was previously identified [43] to be associated with soluble sugar content in soybean with a LOD= 5.2 and PVE= 11.8. This QTL is located 1.4 MB apart from qSUC-3-NC-2018.
Although the major QTL qSU1901 reported in a previous study [43] on chr. 19 is ~40MB away from the qSTA-3-NC-2018 and qSTA-4-NC-2018, it could be that the gene(s) underlying this QTL are different or not due to chromosomal rearrangement that happened in ZD27 by HF25 population versus Forrest by Williams 82 population. Those QTL regions on chr. 19 were reported in several studies and could be subject to further high-density genetic mapping to isolate genes that underly sugar content in soybean seeds.
The sucrose synthase gene Glyma.09G073600 was highly expressed in seeds, followed by Glyma.17G111400, Glyma.17G035800, and Glyma.08G043800 with moderated expression patterns in seeds. Glyma.09G073600 and Glyma.09G016600 are located close to the qSUC-7-IL-2018, qRAF-1-IL-2018, and qRAF-2-IL-2018 on Chrs.9. Glyma.08G143500 is located close to the qSUC-3-NC-2020, and Glyma.05G003900 is positioned close to the qSUC-5-IL-2018 and qSUC-2-NC-2020 on Chr. 5. These genes could be useful in gene editing technology or breeding programs to develop soybean cultivars with reduced amounts of RFOs, and high amounts of sucrose which is beneficial for human consumption and animal feed.
Further studies are needed to characterize these genes, identify their enzymes and protein products, understand their roles in the sugar’s biosynthetic pathway in soybean.
5. Conclusions
In summary, we have identified 26 QTL associated with the seed sugars contents and 57 candidate genes involved in sucrose, raffinose, and stachyose biosynthetic pathway. Amongst these candidate genes, 16 were located less than 10 MB apart from the QTL regions identified in this study.
On chr. 17, a cluster of four genes controlling the sugar pathway is collocated within 6 MB with two QTL (qSUC-10-NC-2018 and qSTA-3-IL-2020) that were identified in this study. Moreover, the raffinose synthase candidate gene Glyma.06G175500 is 9.7MB apart from the qSUC-6-NC-2018 QTL on chr. 6. The invertase candidate gene Glyma.02G016700 is located 3.6 and 0.2 MB apart from qSUC-1-NC-2018 (R2=47.9) and qSUC-1-IL-2020 (R2=3.6) respectively, on chr. 2. Moreover, the sucrose synthase candidate gene Glyma.09G073600 and the raffinose synthase candidate gene Glyma.09G016600 were found close to the qSUC-7-IL-2018, qRAF-1-IL-2018, qRAF-2-IL-2018, and qRAF-1-IL-2018 on chr. 9.
Five QTL regions were commonly identified in the two environments, NC and IL, on chrs. 2, 5, 13, 17 and 20, ((qSUC-5-NC-2018 and qSUC-2-IL-2020), (qSUC-9-NC-2018 and qSTA-1-NC-2018, qSTA-1-IL-2020), (qSUC-12-NC-2018, qSTA-4-IL-2020), (qSUC-10-NC-2018 and qSTA-3-IL-2020), and (qSUC-2-NC-2018 and qSUC-1-IL-2020)).
Five genes (Glyma.09G073600, Glyma.08G143500, Glyma.17G111400, Glyma.05G003900, and Glyma.09G016600) have SNPs and InDels between Forrest and Williams 82 sequences. These SNPs could potentially explain the difference in sugar content between Forrest and Williams 82 cultivars.
Further studies are required to functionally characterize these genes understand and validate their roles in the sugar’s biosynthetic pathway in soybean, before being used in breeding programs to produce soybean lines with high beneficial sucrose and low RFOs.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, K.M. and M.A.K. and; methodology, D.K., J.Y., T.V., N.L., A.M., E.A., N.B. and M.E.; validation, M.A.K., K.M. and H.T.N.; formal analysis, D.K., J.Y. and N.B.; investigation, K.M., and M.A.K.; resources and data curation, K.M., M.A.K. and H.T.N.; writing—original draft preparation, D.K., M.A.K. and K.M.; review and editing, D.K., J.Y., N.B., N.L., T.V., M.A.K., K.M. and H.T.N.; supervision, M.A.K., K.M.; project administration, M.A.K., K.M., and H.T.N. All authors have read and agreed to the published version of the manuscript.
Acknowledgements
Technical support provided by Sandra Mosley is appreciated. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the United States Department of Agriculture (USDA). USDA is an equal opportunity provider and employer. This research was partially funded by the U.S. Department of Agriculture, Agricultural Research Service Project 6066-21220-014-000D. This project was partially funded by the United Soybean Board, project # 2220-152-0104 and Southern Illinois University at Carbondale.
References
- Blackman, S.A.; Obendorf, R.L.; Leopold, A.C. Maturation Proteins and Sugars in Desiccation Tolerance of Developing Soybean Seeds 1. Plant Physiology 1992, 100, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Hitz, W.D.; Carlson, T.J.; Kerr, P.S.; Sebastian, S.A. Biochemical and Molecular Characterization of a Mutation That Confers a Decreased Raffinosaccharide and Phytic Acid Phenotype on Soybean Seeds. Plant Physiology 2002, 128, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Koch, K. Sucrose metabolism: regulatory mechanisms and pivotal roles in sugar sensing and plant development. Current Opinion in Plant Biology 2004, 7, 235–246. [Google Scholar] [CrossRef]
- Redekar, N.R.; Glover, N.M.; Biyashev, R.M.; Ha, B.-K.; Raboy, V.; Maroof, M.A.S. Genetic interactions regulating seed phytate and oligosaccharides in soybean (Glycine max L.). PLOS ONE 2020, 15, e0235120. [Google Scholar] [CrossRef]
- Skoneczka, J.A.; Maroof, M.A.S.; Shang, C.; Buss, G.R. Identification of Candidate Gene Mutation Associated With Low Stachyose Phenotype in Soybean Line PI200508. Crop Science 2009, 49, 247–255. [Google Scholar] [CrossRef]
- Horbowicz, M.; Obendorf, R.L. Seed desiccation tolerance and storability: dependence on flatulence-producing oligosaccharides and cyclitols—review and survey. Seed Science Research 1994, 4, 385–405. [Google Scholar]
- Sprenger, R.; Schlagenhaufer, R.; Kerb, R.; Bruhn, C.; Brockmöller, J.; Roots, I.; Brinkmann, U. Characterization of the glutathione S-transferase GSTT1 deletion: discrimination of all genotypes by polymerase chain reaction indicates a trimodular genotype–phenotype correlation. Pharmacogenetics and Genomics 2000, 10, 557–565. [Google Scholar]
- Pennycooke, J.C.; Jones, M.L.; Stushnoff, C. Down-regulating α-galactosidase enhances freezing tolerance in transgenic petunia. Plant Physiology 2003, 133, 901–909. [Google Scholar]
- ElSayed, A.I.; Rafudeen, M.S.; Golldack, D. Physiological aspects of raffinose family oligosaccharides in plants: protection against abiotic stress. Plant Biology 2014, 16, 1–8. [Google Scholar]
- Keller, F.; Pharr, D.M. Metabolism of carbohydrates in sinks and sources: galactosyl-sucrose oligosaccharides. In Photoassimilate distribution in plants and crops; Routledge: 1996; pp. 157-183.
- Xue, H.; Chen, X.; Li, G. Involvement of phospholipid signaling in plant growth and hormone effects. Current opinion in plant biology 2007, 10, 483–489. [Google Scholar]
- Okada, M.; Ye, K. Nuclear phosphoinositide signaling regulates messenger RNA export. RNA biology 2009, 6, 12–16. [Google Scholar] [PubMed]
- Thole, J.M.; Nielsen, E. Phosphoinositides in plants: novel functions in membrane trafficking. Current opinion in plant biology 2008, 11, 620–631. [Google Scholar] [PubMed]
- González, E.M.; Gordon, A.J.; James, C.L.; Arrese-lgor, C. The role of sucrose synthase in the response of soybean nodules to drought. Journal of Experimental Botany 1995, 46, 1515–1523. [Google Scholar] [CrossRef]
- Bellaloui, N.; Smith, J.R.; Gillen, A.M.; Ray, J.D. Effect of Maturity on Seed Sugars as Measured on Near-Isogenic Soybean (<i>Glycine max</i> ) Lines. Crop Science 2010, 50, 1978–1987. [Google Scholar] [CrossRef]
- Bellaloui, N.; Mengistu, A.; Fisher, D.K.; Abel, C.A. Soybean Seed Composition Constituents as Affected by Drought and Phomopsis in Phomopsis Susceptible and Resistant Genotypes. Journal of Crop Improvement 2012, 26, 428–453. [Google Scholar] [CrossRef]
- Wijewardana, C.; Reddy, K.R.; Bellaloui, N. Soybean seed physiology, quality, and chemical composition under soil moisture stress. Food Chemistry 2019, 278, 92–100. [Google Scholar] [CrossRef]
- Kassem, M.A. Soybean Seed Composition: Protein, Oil, Fatty Acids, Amino Acids, Sugars, Mineral Nutrients, Tocopherols, and Isoflavones; Springer Nature: 2021.
- Taiz, L. Mineral nutrition, Plant Physiology ed.; Sinauer Associates Inc.: P.O. Box 407, Sunderland, MA 01375, USA, 1998. [Google Scholar]
- Brown, A.V.; Conners, S.I.; Huang, W.; Wilkey, A.P.; Grant, D.; Weeks, N.T.; Cannon, S.B.; Graham, M.A.; Nelson, R.T. A new decade and new data at SoyBase, the USDA-ARS soybean genetics and genomics database. Nucleic Acids Research 2021, 49, D1496–D1501. [Google Scholar] [CrossRef]
- Salari, M.W.; Ongom, P.O.; Thapa, R.; Nguyen, H.T.; Vuong, T.D.; Rainey, K.M. Mapping QTL controlling soybean seed sucrose and oligosaccharides in a single family of soybean nested association mapping (SoyNAM) population. Plant Breeding 2021, 140, 110–122. [Google Scholar] [CrossRef]
- Wang, T.L.; Domoney, C.; Hedley, C.L.; Casey, R.; Grusak, M.A. Can we improve the nutritional quality of legume seeds? Plant Physiology 2003, 131, 886–891. [Google Scholar]
- Arendt, E.K.; Zannini, E. Cereal grains for the food and beverage industries; Elsevier: 2013.
- Avilés-Gaxiola, S.; Chuck-Hernández, C.; Serna Saldívar, S.O. Inactivation methods of trypsin inhibitor in legumes: a review. Journal of Food Science 2018, 83, 17–29. [Google Scholar]
- Kerr, P.S.; Pearlstein, R.W.; Schweiger, B.J.; Becker-Manley, M.F.; Pierce, J.W. Nucleotide sequences of galactinol synthase from zucchini and soybean. 1998.
- Frías, J.; Bakhsh, A.; Jones, D.; Arthur, A.; Vidal-Valverde, C.; Rhodes, M.; Hedley, C.L. Genetic analysis of the raffinose oligosaccharide pathway in lentil seeds. Journal of Experimental Botany 1999, 50, 469–476. [Google Scholar] [CrossRef]
- Hedley, C.L. Carbohydrates in grain legume seeds: improving nutritional quality and agronomic characteristics; CABI: 2001.
- Knizia, D.; Yuan, J.; Bellaloui, N.; Vuong, T.; Usovsky, M.; Song, Q.; Betts, F.; Register, T.; Williams, E.; Lakhssassi, N. The Soybean High Density ‘Forrest’by ‘Williams 82’SNP-Based Genetic Linkage Map Identifies QTL and Candidate Genes for Seed Isoflavone Content. Plants 2021, 10, 2029. [Google Scholar] [PubMed]
- Knizia, D.; Yuan, J.; Lakhssassi, N.; El Baze, A.; Cullen, M.; Vuong, T.; Mazouz, H.; T. Nguyen, H.; Kassem, M.A.; Meksem, K. QTL and Candidate Genes for Seed Tocopherol Content in ‘Forrest’by ‘Williams 82’Recombinant Inbred Line (RIL) Population of Soybean. Plants 2022, 11, 1258. [Google Scholar] [CrossRef] [PubMed]
- Akond, M.; Liu, S.; Kantartzi, S.K.; Meksem, K.; Bellaloui, N.; Lightfoot, D.A.; Kassem, M.A. Quantitative trait loci underlying seed sugars content in “MD96-5722” by “Spencer” recombinant inbred line population of soybean. Food and Nutrition Sciences 2015, 6, 964. [Google Scholar]
- Allen, G.C.; Flores-Vergara, M.; Krasynanski, S.; Kumar, S.; Thompson, W. A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide. Nature protocols 2006, 1, 2320–2325. [Google Scholar]
- Wu, X.; Vuong, T.D.; Leroy, J.A.; Grover Shannon, J.; Sleper, D.A.; Nguyen, H.T. Selection of a core set of RILs from Forrest× Williams 82 to develop a framework map in soybean. Theoretical and applied genetics 2011, 122, 1179–1187. [Google Scholar]
- Wang, S.; Basten, C.; Zeng, Z. Windows QTL Cartographer 2.5 _011. North Carolina State University, Raleigh.
- Voorrips, R. MapChart: software for the graphical presentation of linkage maps and QTLs. Journal of heredity 2002, 93, 77–78. [Google Scholar] [CrossRef]
- Patil, G.B.; Lakhssassi, N.; Wan, J.; Song, L.; Zhou, Z.; Klepadlo, M.; Vuong, T.D.; Stec, A.O.; Kahil, S.S.; Colantonio, V.; et al. Whole-genome re-sequencing reveals the impact of the interaction of copy number variants of the rhg1 and Rhg4 genes on broad-based resistance to soybean cyst nematode. Plant Biotechnology Journal 2019, 17, 1595–1611. [Google Scholar] [CrossRef]
- Iftime, D.; Hannah, M.A.; Peterbauer, T.; Heyer, A.G. Stachyose in the cytosol does not influence freezing tolerance of transgenic Arabidopsis expressing stachyose synthase from adzuki bean. Plant Sci 2011, 180, 24–30. [Google Scholar] [CrossRef]
- González-Morales, S.I.; Chávez-Montes, R.A.; Hayano-Kanashiro, C.; Alejo-Jacuinde, G.; Rico-Cambron, T.Y.; de Folter, S.; Herrera-Estrella, L. Regulatory network analysis reveals novel regulators of seed desiccation tolerance in <i>Arabidopsis thaliana</i>. Proceedings of the National Academy of Sciences 2016, 113, E5232–E5241. [Google Scholar] [CrossRef]
- Blöchl, A.; March, G.G.-d.; Sourdioux, M.; Peterbauer, T.; Richter, A. Induction of raffinose oligosaccharide biosynthesis by abscisic acid in somatic embryos of alfalfa (Medicago sativa L.). Plant Science 2005, 168, 1075–1082. [Google Scholar] [CrossRef]
- Maughan, P.; Maroof, M.; Buss, G. Identification of quantitative trait loci controlling sucrose content in soybean (Glycine max). Molecular Breeding 2000, 6, 105–111. [Google Scholar]
- Kim, H.K.; Kang, S.T.; Oh, K.W. Mapping of putative quantitative trait loci controlling the total oligosaccharide and sucrose content of Glycine max seeds. Journal of Plant Research 2006, 119, 533–538. [Google Scholar]
- Kim, H.-K.; Kang, S.-T.; Cho, J.-H.; Choung, M.-G.; Suh, D.-Y. Quantitative trait loci associated with oligosaccharide and sucrose contents in soybean (Glycine max L.). Journal of Plant Biology 2005, 48, 106–112. [Google Scholar]
- Mainali, H.R.; Vadivel, A.K.A.; Li, X.; Gijzen, M.; Dhaubhadel, S. Soybean cyclophilin GmCYP1 interacts with an isoflavonoid regulator GmMYB176. Scientific reports 2017, 7, 1–12. [Google Scholar]
- Liu, C.; Chen, H.; Yu, Q.; Gu, H.; Li, Y.; Tu, B.; Zhang, H.; Zhang, Q.; Liu, X. Identification of quantitative trait loci (QTLs) and candidate genes for seed sucrose and soluble sugar concentrations in soybean. Crop Science 2023. [Google Scholar]
- Zeng, A.; Chen, P.; Shi, A.; Wang, D.; Zhang, B.; Orazaly, M.; Florez-Palacios, L.; Brye, K.; Song, Q.; Cregan, P. Identification of quantitative trait loci for sucrose content in soybean seed. Crop Science 2014, 54, 554–564. [Google Scholar]
- Cai, J.-S.; Feng, J.-Y.; Ni, Z.-J.; Ma, R.-H.; Thakur, K.; Wang, S.; Hu, F.; Zhang, J.-G.; Wei, Z.-J. An update on the nutritional, functional, sensory characteristics of soy products, and applications of new processing strategies. Trends in Food Science & Technology 2021, 112, 676–689. [Google Scholar]
- Sui, M.; Wang, Y.; Bao, Y.; Wang, X.; Li, R.; Lv, Y.; Yan, M.; Quan, C.; Li, C.; Teng, W. Genome-wide association analysis of sucrose concentration in soybean (Glycine max L.) seed based on high-throughput sequencing. The Plant Genome 2020, 13, e20059. [Google Scholar]
- Lee, J.S.; Kim, S.-M.; Kang, S. Fine mapping of quantitative trait loci for sucrose and oligosaccharide contents in soybean [Glycine max (L.) Merr.] using 180 K Axiom® SoyaSNP genotyping platform. Euphytica 2016, 208, 195–203. [Google Scholar]
- Stombaugh, S.; Orf, J.H.; Jung, H.; Chase, K.; Lark, K.; Somers, D. Quantitative trait loci associated with cell wall polysaccharides in soybean seed. Crop science 2004, 44, 2101–2106. [Google Scholar]
- Feng, C.; Morsy, M.; Giannoccaro, E.; Zhang, B.; Chen, P. Soybean seed sugar content and quantitative trait loci mapping. In Proceedings of the Plant nutrition for food security, human health and environmental protection; Fifteenth International Plant Nutrition Colloquium; 2005. [Google Scholar]
- Jaureguy, L.M. Identification of molecular markers associated with seed size, protein and sugar content in soybean; University of Arkansas: 2009.
- Wang, X.; Jiang, G.-L.; Green, M.; Scott, R.A.; Song, Q.; Hyten, D.L.; Cregan, P.B. Identification and validation of quantitative trait loci for seed yield, oil and protein contents in two recombinant inbred line populations of soybean. Molecular genetics and genomics 2014, 289, 935–949. [Google Scholar] [PubMed]
- Dhungana, S.K.; Kulkarni, K.P.; Park, C.W.; Jo, H.; Song, J.T.; Shin, D.H.; Lee, J.D. Mapping quantitative trait loci controlling soybean seed starch content in an interspecific cross of ‘Williams 82’(Glycine max) and ‘PI 366121’(Glycine soja). Plant Breeding 2017, 136, 379–385. [Google Scholar]
- Patil, G.; Vuong, T.D.; Kale, S.; Valliyodan, B.; Deshmukh, R.; Zhu, C.; Wu, X.; Bai, Y.; Yungbluth, D.; Lu, F. Dissecting genomic hotspots underlying seed protein, oil, and sucrose content in an interspecific mapping population of soybean using high-density linkage mapping. Plant Biotechnology Journal 2018, 16, 1939–1953. [Google Scholar] [PubMed]
- Hu, L.; Wang, X.; Zhang, J.; Florez-Palacios, L.; Song, Q.; Jiang, G.-L. Genome-Wide Detection of Quantitative Trait Loci and Prediction of Candidate Genes for Seed Sugar Composition in Early Mature Soybean. International Journal of Molecular Sciences 2023, 24, 3167. [Google Scholar]
Figure 1.
Frequency distribution of sugars (sucrose, raffinose, and stachyose) in the FXW82 RIL population grown in two environments over two years (Spring Lake, NC in 2018 and Carbondale, IL in 2020.
Figure 1.
Frequency distribution of sugars (sucrose, raffinose, and stachyose) in the FXW82 RIL population grown in two environments over two years (Spring Lake, NC in 2018 and Carbondale, IL in 2020.
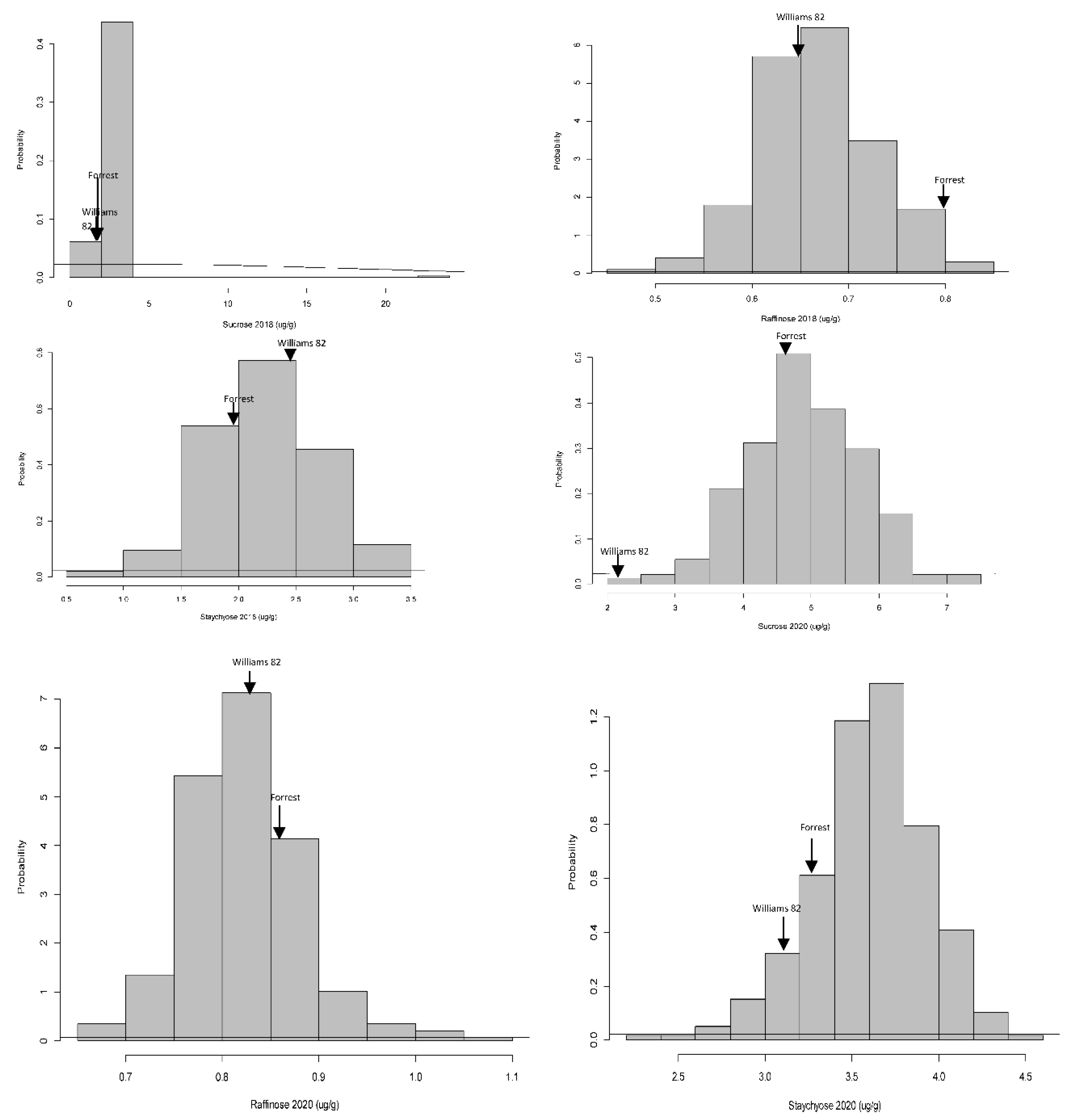
Figure 2.
The sugars (sucrose, raffinose, and stachyose) biosynthetic pathway with the identified candidate genes in soybean. The genes are in Wm82.a2.v1 annotation.
Figure 2.
The sugars (sucrose, raffinose, and stachyose) biosynthetic pathway with the identified candidate genes in soybean. The genes are in Wm82.a2.v1 annotation.
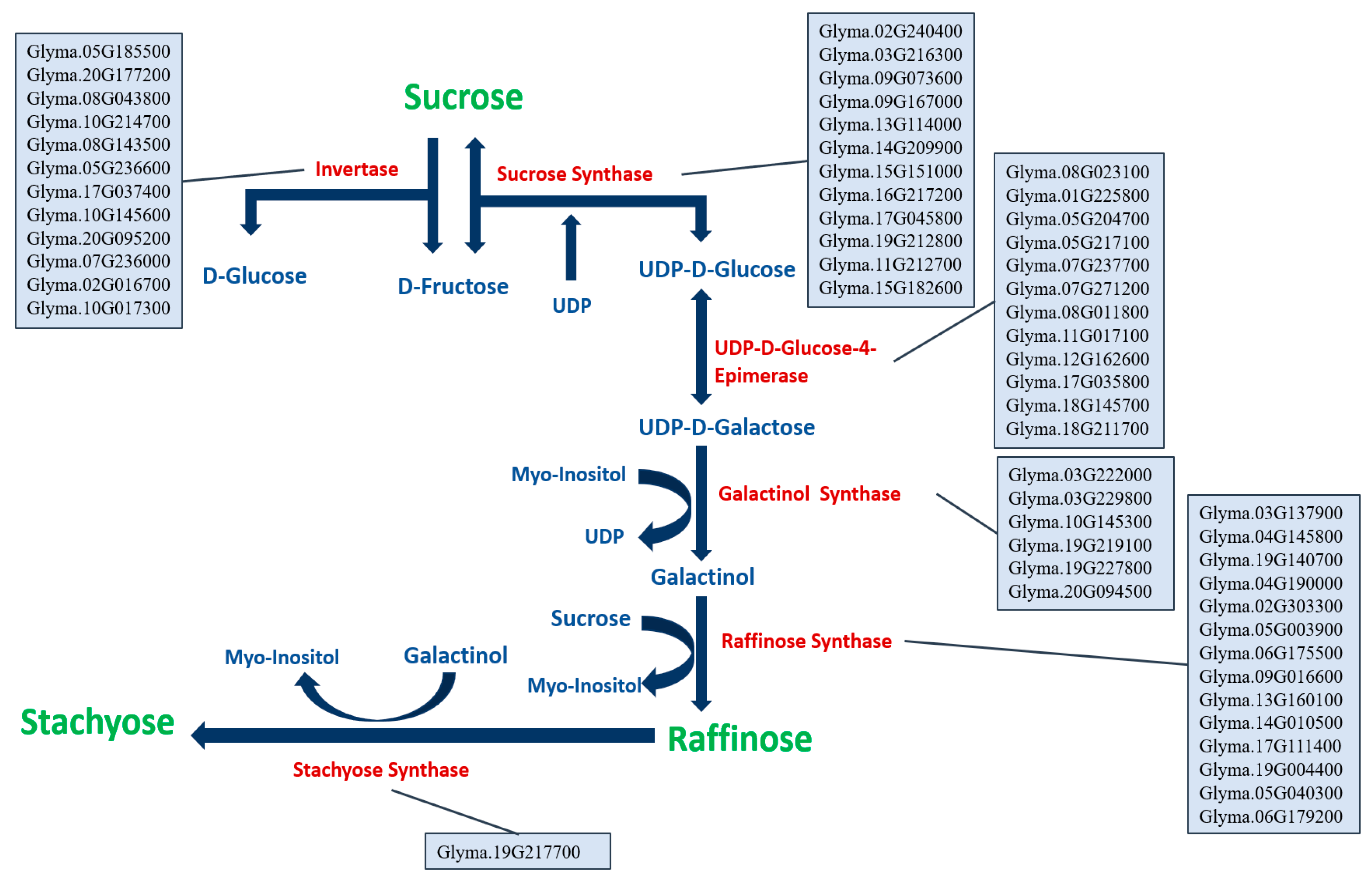
Figure 3.
A. Tissue specific expression of the identified sugars candidate genes. B. Expression HeatMap of the identified candidate genes located within 10 MB to the identified sugars QTL regions in Williams 82 (RPKM) were retrieved from publicly available RNA-seq data from Soybase database [20]. RNA-seq data is not available at Soybase for the Glyma.17G045800 candidate gene.
Figure 3.
A. Tissue specific expression of the identified sugars candidate genes. B. Expression HeatMap of the identified candidate genes located within 10 MB to the identified sugars QTL regions in Williams 82 (RPKM) were retrieved from publicly available RNA-seq data from Soybase database [20]. RNA-seq data is not available at Soybase for the Glyma.17G045800 candidate gene.
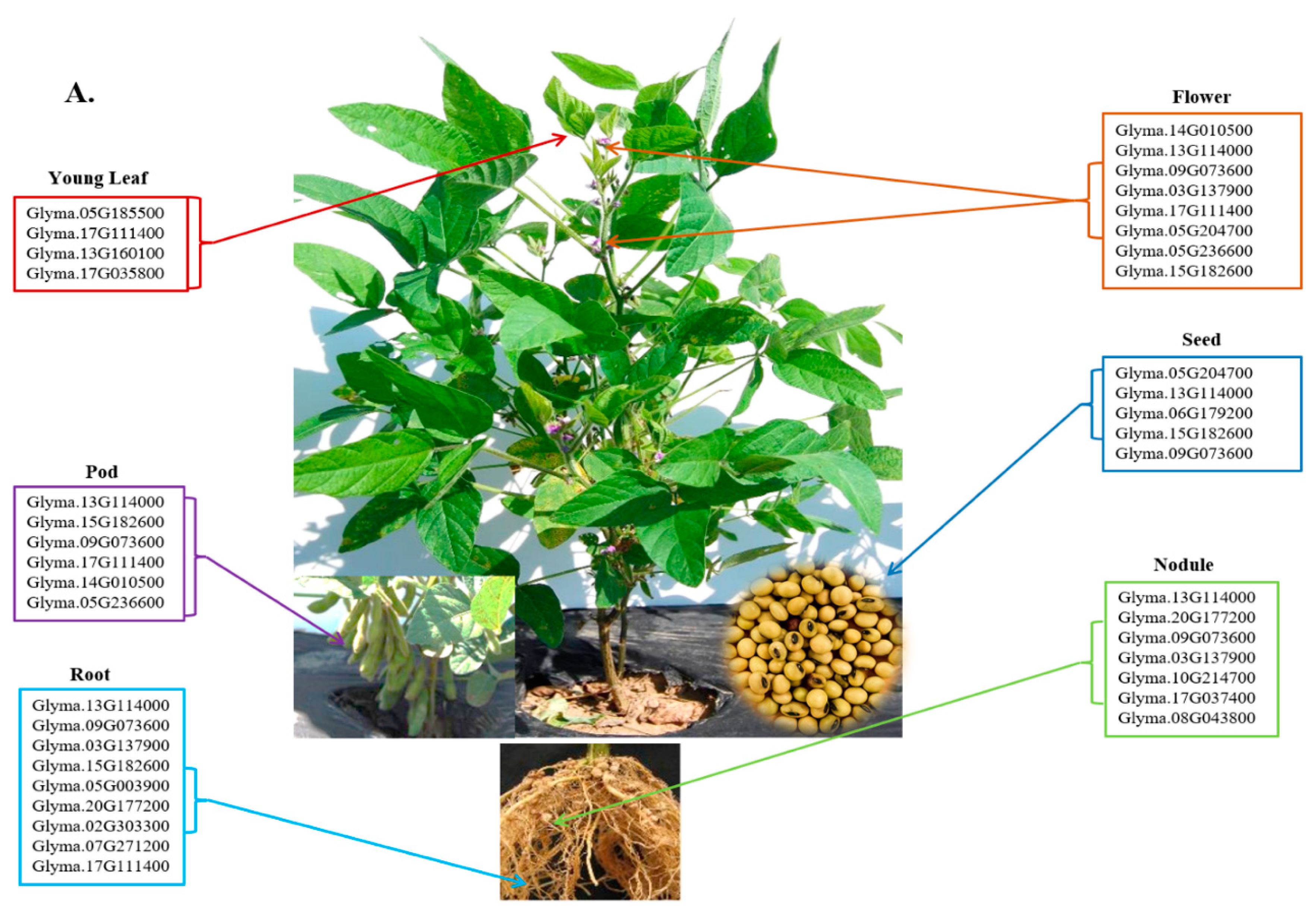
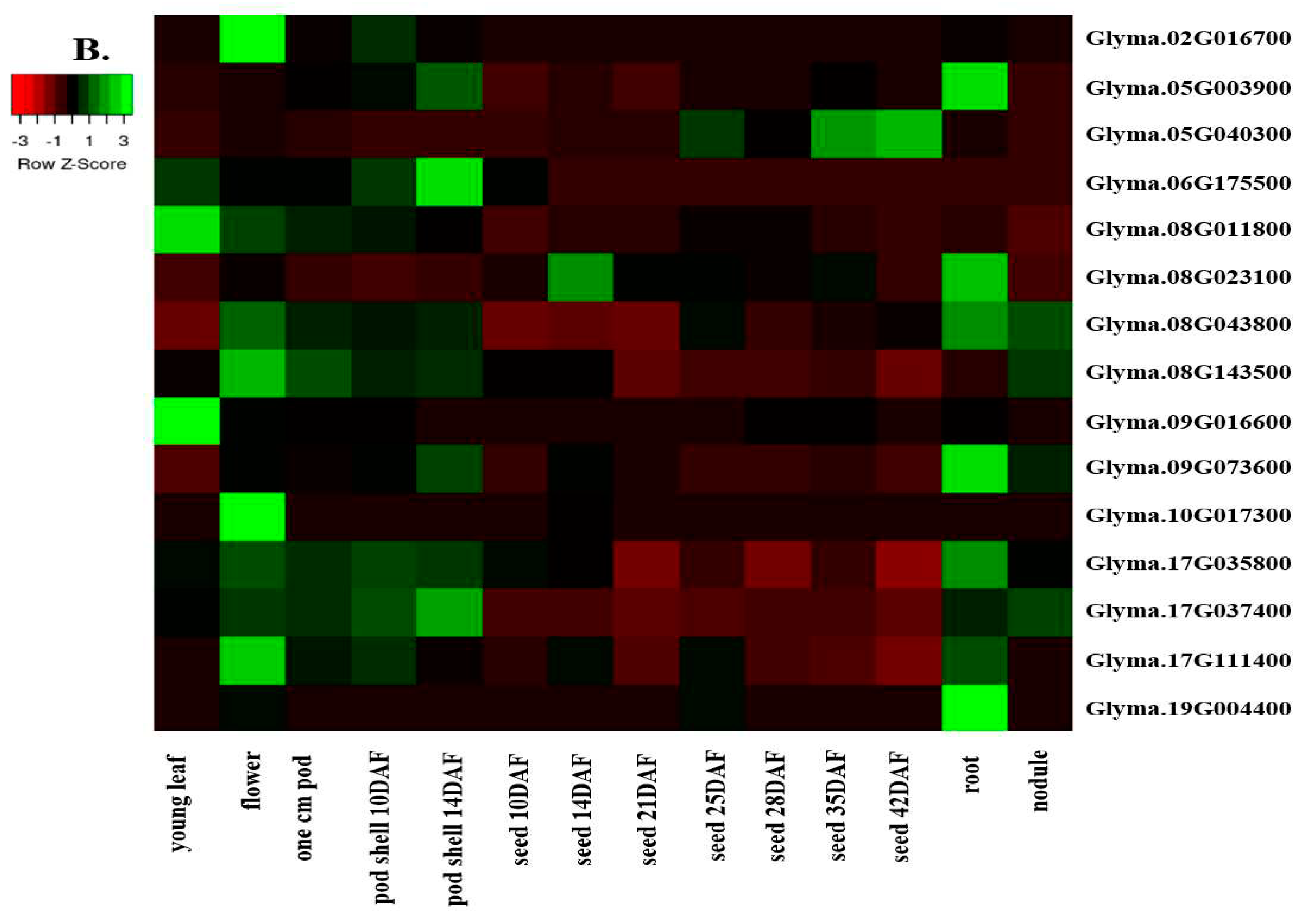
Figure 4.
Positions of SNPs between Forrest and Williams 82 cultivars in Glyma.09G073600, Glyma.08G143500, Glyma.05G003900, Glyma.17G111400, and Glyma.09G016600 coding sequences. In the gene model diagram, the light blue/light green boxes represent exons, blue/green bars represent introns, dark blue/dark green boxes represent 3′UTR or 5′UTR. SNPs were positioned relative to the genomic position in the genome version W82.a2.
Figure 4.
Positions of SNPs between Forrest and Williams 82 cultivars in Glyma.09G073600, Glyma.08G143500, Glyma.05G003900, Glyma.17G111400, and Glyma.09G016600 coding sequences. In the gene model diagram, the light blue/light green boxes represent exons, blue/green bars represent introns, dark blue/dark green boxes represent 3′UTR or 5′UTR. SNPs were positioned relative to the genomic position in the genome version W82.a2.
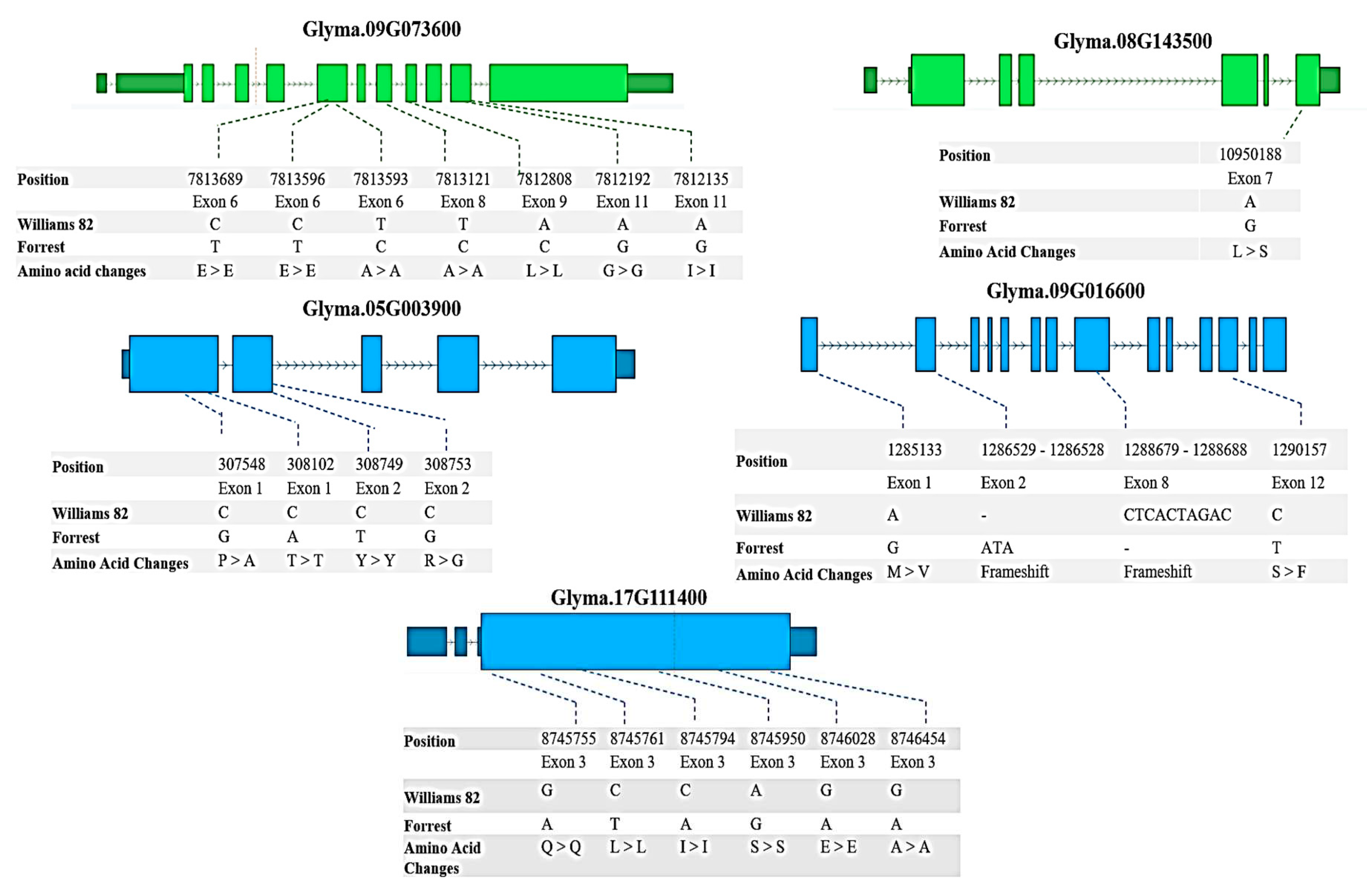

Table 1.
Seed sugars contents means, ranges, CVs, skewness, and kurtosis in the FxW82 RIL population evaluated in Spring Lake, NC (2018) and Carbondale, IL (2020). Mean and range values are expressed in μg/g of seed weight.
Table 1.
Seed sugars contents means, ranges, CVs, skewness, and kurtosis in the FxW82 RIL population evaluated in Spring Lake, NC (2018) and Carbondale, IL (2020). Mean and range values are expressed in μg/g of seed weight.
| Year | Trait | Mean | Range | CV (%) | SE | Skewness | Kurtosis | W value (P<0.05) |
|---|---|---|---|---|---|---|---|---|
| 2018 | Sucrose | 2.58 | 22.7 | 62.86 | 0.12 | 12.2 | 161.38 | 0.22*** |
| Raffinose | 0.67 | 0.26 | 9.16 | 0.01 | 0.18 | 3.26 | 0.99 | |
| Stachyose | 2.23 | 2.55 | 21.74 | 0.03 | -0.07 | 2.85 | 0.99 | |
| 2020 | Sucrose | 4.92 | 4.98 | 17.2 | 0.05 | -0.13 | 3.15 | 0.99 |
| Raffinose | 0.83 | 0.41 | 7.28 | 0.01 | 0.65 | 4.83 | 0.97*** | |
| Stachyose | 3.61 | 2.15 | 9.06 | 0.02 | -0.48 | 3.8 | 0.98** |
Table 2.
Two-way ANOVA of seed sugars (sucrose, stachyose, and raffinose) contents in the FxW82 RIL population evaluated in Spring Lake, NC (2018) and Carbondale, IL (2020).
Table 2.
Two-way ANOVA of seed sugars (sucrose, stachyose, and raffinose) contents in the FxW82 RIL population evaluated in Spring Lake, NC (2018) and Carbondale, IL (2020).
| Response: Sucrose | ||||
| Df | Sum Sq | Mean Seq | H2 | |
| Line | 369 | 1134.22 | 3.0738 | 0.378 |
| Year | 1 | 5.6 | 5.5975 | |
| Line × Year | 2 | 3.82 | 1.9108 | |
| Residuals | 0 | 0 | NA | |
| Response: Raffinose | ||||
| Df | Sum Sq | Mean Seq | H2 | |
| Line | 369 | 3.4552 | 0.0093891 | 0.739 |
| Year | 1 | 0.0253 | 0.0253139 | |
| Line × Year | 2 | 0.0048 | 0.0023972 | |
| Residuals | 0 | 0 | NA | |
| Response: Stachyose | ||||
| Df | Sum Sq | Mean Seq | H2 | |
| Line | 369 | 246.73 | 0.66865 | 0.92 |
| Year | 1 | 1.611 | 1.61115 | |
| Line × Year | 2 | 0.106 | 0.05307 | |
| Residuals | 0 | 0 | NA | |
Table 3.
Quantitative trait loci (QTL) that control sugars (sucrose, stachyose, and raffinose) contents in FxW82 RIL population in Spring Lake, NC in 2018. These QTL have been identified by CIM method. * Indicate novel QTL.
Table 3.
Quantitative trait loci (QTL) that control sugars (sucrose, stachyose, and raffinose) contents in FxW82 RIL population in Spring Lake, NC in 2018. These QTL have been identified by CIM method. * Indicate novel QTL.
| Trait | QTL | Chr. | Marker/Interval | Position (cM) | LOD | R2 | Add. Eff. |
|---|---|---|---|---|---|---|---|
| Sucrose | qSUC-1 | 1 | Gm01_3504836-Gm01_3466825 | 0.01-12.1 | 39.19 | 20.46 | -3.05 |
| qSUC-2 | 2 | Gm02_5155733-Gm02_9925870 | 128.5-142.2 | 42.77 | 47.90 | 4.42 | |
| qSUC-3 | 3 | Gm03_4595422-Gm03_4113546 | 39.2-39.8 | 32.62 | 20.50 | 3.05 | |
| qSUC-4* | 4 | Gm04_7672403 | 6.5-16.5 | 54.35 | 37.50 | 4.62 | |
| qSUC-5 | 5 | Gm05_3867435-Gm05_3273418 | 31.5-37.01 | 20.65 | 17.51 | 2.60 | |
| qSUC-6 | 6 | Gm06_1737718-Gm06_5014399 | 48.5-52.4 | 5.36 | 10.50 | -1.37 | |
| qSUC-7 | 9 | Gm09_1888876 | 173.9-178.1 | 32.62 | 20.50 | 3.05 | |
| qSUC-8* | 10 | Gm10_621706 | 214.01-216.01 | 34.25 | 19.10 | -4.48 | |
| qSUC-9 | 13 | Gm13_3891723-Gm13_3524828 | 0.2-58.2 | 19.12 | 17.51 | 2.60 | |
| qSUC-10 | 17 | Gm17_4967175-Gm17_5294475 | 0.4-1.0 | 33.22 | 20.50 | 3.05 | |
| qSUC-11* | 18 | Gm18_1620585-Gm18_2020823 | 94.7-96.5 | 20.10 | 17.51 | 2.60 | |
| qSUC-12 | 20 | Gm19_2552468 | 172.11 | 6.98 | 9.10 | 1.41 | |
| Stachyose | qSTA-1 | 13 | Gm13_3524828 | 96.2-98.2 | 2.52 | 14.8 | 0.19 |
| qSTA-2 | 13 | Gm13_3884070-Gm13_3803273 | 121.8-123.2 | 2.60 | 5.2 | 0.11 | |
| qSTA-3 | 19 | Gm19_3789399-Gm19_4362616 | 98.01-124.1 | 4.21 | 8.5 | -0.16 | |
| qSTA-4 | 19 | Gm19_4946208-Gm19_5032228 | 184.1-186.1 | 2.53 | 5.3 | 0.11 | |
| Raffinose | qRAF-1 | 9 | Gm09_4024436-Gm09_4082234 | 108.01-110.9 | 2.26 | 4.6 | -0.01 |
| qRAF-2 | 9 | Gm09_1888876 | 173.9-178.1 | 2.47 | 7.6 | 0.08 | |
| qRAF-3 | 12 | Gm12_6023395-Gm12_2379195 | 114.6-118.6 | 2.15 | 4.7 | -0.01 |
Table 4.
Quantitative trait loci (QTL) that control sugars (sucrose, stachyose, and raffinose) contents in FxW82 RIL population in Carbondale, IL in 2020. These QTL have been identified by CIM method. * Indicate novel QTL.
Table 4.
Quantitative trait loci (QTL) that control sugars (sucrose, stachyose, and raffinose) contents in FxW82 RIL population in Carbondale, IL in 2020. These QTL have been identified by CIM method. * Indicate novel QTL.
| Trait | QTL | Chr. | Marker | Position (cM) | LOD | R2 | Add. Eff. |
|---|---|---|---|---|---|---|---|
| Sucrose | qSUC-1 | 2 | Gm02_1199805-Gm02_1373746 | 196.4-205.6 | 2.63 | 3.60 | -0.16 |
| qSUC-2 | 5 | Gm05_3803682-Gm05_3748078 | 18.01-22.1 | 2.10 | 0.03 | -0.14 | |
| qSUC-3 | 8 | Gm08_5960619-Gm08_8268861 | 47.1-55.9 | 2.37 | 0.04 | 0.16 | |
| Stachyose | qSTA-1 | 13 | Gm13_2748576 | 0.5-4.5 | 2.03 | 0.09 | 0.21 |
| qSTA-2 | 16 | Gm16_3183754-Gm16_3010888 | 81.6-94.7 | 2.85 | 3.92 | 0.10 | |
| qSTA-3 | 17 | Gm17_8449684-Gm17_8352493 | 136.5-136.7 | 2.37 | 3.00 | -0.08 | |
| qSTA-4 | 20 | Gm20_294157-Gm20_1133712 | 145.4-148.5 | 3.59 | 4.50 | -0.12 |
Table 5.
QTL and candidate genes that control sugars (sucrose, stachyose, and raffinose) contents in FxW82 RIL population in Spring Lake, NC in 2018. These QTL have been identified by CIM method. Genes with (***) are apart from the identified QTL with less than 10 MB; Genes with (**) are apart from the identified QTL with less than 20 MB; Genes with (*) are apart from the identified QTL with more than 20 MB.
Table 5.
QTL and candidate genes that control sugars (sucrose, stachyose, and raffinose) contents in FxW82 RIL population in Spring Lake, NC in 2018. These QTL have been identified by CIM method. Genes with (***) are apart from the identified QTL with less than 10 MB; Genes with (**) are apart from the identified QTL with less than 20 MB; Genes with (*) are apart from the identified QTL with more than 20 MB.
| Trait | QTL | Marker/Interval | LOD | R2 | Wm82.a2.v1 | Start | End | Wm82.a1.v1.1 | Start | End | Dis. (MB) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sucrose | qSUC-1 | Gm01_3504836-Gm01_3466825 | 39.19 | 20.46 | Glyma.01G225800* | 55452580 | 55456886 | Glyma01g43540 | 54536305 | 54540597 | 51.03 |
| qSUC-2 | Gm02_5155733-Gm02_9925870 | 42.77 | 47.9 | Glyma.02G016700*** | 1490049 | 1491170 | Glyma02g02030 | 1475851 | 1476528 | 3.6 | |
| qSUC-3 | Gm03_4595422-Gm03_4113546 | 32.62 | 20.5 | Glyma.03G222000* | 43660855 | 43663317 | Glyma03g38080 | 44498027 | 44500613 | 39.9 | |
| Glyma.03G229800* | 43172456 | 43175687 | Glyma03g38910 | 45176126 | 45179418 | 40.5 | |||||
| Glyma.03G137900* | 35393011 | 35398758 | Glyma03g29440 | 37419739 | 37425659 | 32.8 | |||||
| Glyma.03G216300* | 42037913 | 42044153 | Glyma03g37441 | 44041487 | 44047783 | 39.4 | |||||
| qSUC-4 | Gm04_7672403 | 54.35 | 37.5 | Glyma.04G145800** | 27037731 | 27039621 | Glyma18g23060 | 26644665 | 26645606 | 18.97 | |
| Glyma.04G190000* | 46076888 | 46080907 | Glyma04g36410 | 42932203 | 42936043 | 35.2 | |||||
| qSUC-5 | Gm05_3867435-Gm05_3273418 | 20.65 | 17.51 | Glyma.05G040300*** | 3593378 | 3598821 | Glyma05g02510 | 1870330 | 1875692 | 1.3 | |
| Glyma.05G003900*** | 307460 | 312091 | Glyma05g08950 | 8806144 | 8810647 | 4.9 | |||||
| Glyma.05G217100* | 39735138 | 39739763 | Glyma05g36850 | 40599128 | 40603658 | 36.7 | |||||
| Glyma.05G185500* | 37243691 | 37249494 | Glyma05g31920 | 36953899 | 36959702 | 33.08 | |||||
| Glyma.05G236600* | 41293446 | 41294570 | Glyma05g34830 | 39054363 | 39055344 | 35.18 | |||||
| Glyma.05G204700* | 38804305 | 38807296 | Glyma05g38120 | 41530564 | 41533554 | 37.6 | |||||
| qSUC-6 | Gm06_1737718-Gm06_5014399 | 5.36 | 10.5 | Glyma.06G175500*** | 14845358 | 14849994 | Glyma06g18480 | 14802178 | 14807061 | 9.7 | |
| Glyma.06G179200** | 15217419 | 15223877 | Glyma06g18890 | 15175181 | 15181763 | 10.16 | |||||
| qSUC-7 | Gm09_1888876 | 32.62 | 20.5 | Glyma.09G073600*** | 7809852 | 7816248 | Glyma09g08550 | 7845409 | 7851685 | 5.9 | |
| Glyma.09G016600*** | 1285132 | 1290884 | Glyma09g01940 | 1270010 | 1276140 | 0.6 | |||||
| Glyma.09G167000* | 39103764 | 39109664 | Glyma09g29710 | 36530532 | 36536435 | 34.6 | |||||
| qSUC-8 | Gm10_621706 | 34.25 | 19.1 | Glyma.10G017300*** | 1523661 | 1524691 | Glyma10g02170 | 1519053 | 1519546 | 0.8 | |
| Glyma.10G214700* | 44674211 | 44679550 | Glyma10g35890 | 44094080 | 44098889 | 43.4 | |||||
| Glyma.10G145600* | 38035440 | 38039395 | Glyma10g28640 | 37509189 | 37513105 | 36.88 | |||||
| Glyma.10G145300* | 38014452 | 38016396 | Glyma10g28610 | 37488202 | 37490030 | 36.8 | |||||
| qSUC-9 | Gm13_3891723-Gm13_3524828 | 19.12 | 17.51 | Glyma.13G160100* | 27576191 | 27579282 | Glyma13g22890 | 26380083 | 26383137 | 22.48 | |
| Glyma.13G114000** | 22767704 | 22773231 | Glyma13g17420 | 21211880 | 21217237 | 17.3 | |||||
| qSUC-10 | Gm17_4967175-Gm17_5294475 | 33.22 | 20.5 | Glyma.17G037400*** | 2732048 | 2737399 | Glyma17g04160 | 2739794 | 2745132 | 2.2 | |
| Glyma.17G045800*** | 3404918 | 3410491 | Glyma17g05067 | 3412682 | 3418160 | 1.5 | |||||
| Glyma.17G035800*** | 2629011 | 2639005 | Glyma17g03990 | 2637080 | 2646732 | 2.3 | |||||
| Glyma.17G111400*** | 8744555 | 8747526 | Glyma17g11970 | 9015075 | 9018145 | 3.7 | |||||
| qSUC-11 | Gm18_1620585-Gm18_2020823 | 20.1 | 17.51 | Glyma.18G145700* | 24414069 | 24415225 | Glyma18g21870 | 24645144 | 24646447 | 22.6 | |
| qSUC-12 | Gm19_2552468 | 6.98 | 9.1 | Glyma.19G140700* | 40199041 | 40201038 | Glyma19g32250 | 40004601 | 40006724 | 37.4 | |
| Glyma.19G004400*** | 359933 | 363588 | Glyma19g00441 | 238429 | 242106 | 2.3 | |||||
| Glyma.19G217700* | 47033812 | 47037286 | Glyma19g40550 | 46915407 | 46918937 | 44.3 | |||||
| Glyma.19G212800* | 46633685 | 46639818 | Glyma19g40041 | 46515393 | 46521627 | 43.9 | |||||
| Glyma.19G219100* | 47148224 | 47150373 | Glyma19g40680 | 47029812 | 47032065 | 44.4 | |||||
| Glyma.19G227800* | 47911129 | 47914214 | Glyma19g41550 | 47789168 | 47792321 | 45.2 | |||||
| Stachyose | qSTA-1 | Gm13_3524828 | 2.52 | 14.8 | Glyma.13G160100* | 27576191 | 27579282 | Glyma13g22890 | 26380083 | 26383137 | 22.8 |
| Glyma.13G114000** | 22767704 | 22773231 | Glyma13g17420 | 21211880 | 21217237 | 17.6 | |||||
| qSTA-2 | Gm13_3884070-Gm13_3803273 | 2.6 | 5.2 | Glyma.13G160100* | 27576191 | 27579282 | Glyma13g22890 | 26380083 | 26383137 | 22.4 | |
| Glyma.13G114000** | 22767704 | 22773231 | Glyma13g17420 | 21211880 | 21217237 | 17.3 | |||||
| qSTA-3 | Gm19_3789399-Gm19_4362616 | 4.21 | 8.5 | Glyma.19G004400*** | 359933 | 363588 | Glyma19g00440 | 241366 | 241903 | 3.5 | |
| Glyma.19G140700* | 40199041 | 40201038 | Glyma19g32250 | 40004601 | 40006724 | 35.6 | |||||
| Glyma.19G217700* | 47033812 | 47037286 | Glyma19g40550 | 46915407 | 46918937 | 42.5 | |||||
| Glyma.19G212800* | 46633685 | 46639818 | Glyma19g40041 | 46515393 | 46521627 | 42.1 | |||||
| Glyma.19G219100* | 47148224 | 47150373 | Glyma19g40680 | 47029812 | 47032065 | 42.6 | |||||
| Glyma.19G227800* | 47911129 | 47914214 | Glyma19g41550 | 47789168 | 47792321 | 43.4 | |||||
| qSTA-4 | Gm19_4946208-Gm19_5032228 | 2.53 | 5.3 | Glyma.19G004400*** | 359933 | 363588 | Glyma19g00440 | 241366 | 241903 | 4.7 | |
| Glyma.19G140700* | 40199041 | 40201038 | Glyma19g32250 | 40004601 | 40006724 | 34.9 | |||||
| Glyma.19G217700* | 47033812 | 47037286 | Glyma19g40550 | 46915407 | 46918937 | 41.8 | |||||
| Glyma.19G212800* | 46633685 | 46639818 | Glyma19g40041 | 46515393 | 46521627 | 41.4 | |||||
| Glyma.19G219100* | 47148224 | 47150373 | Glyma19g40680 | 47029812 | 47032065 | 41.9 | |||||
| Glyma.19G227800* | 47911129 | 47914214 | Glyma19g41550 | 47789168 | 47792321 | 42.7 | |||||
| Raffinose | qRAF-1 | Gm09_4024436-Gm09_4082234 | 2.26 | 4.6 | Glyma.09G073600*** | 7809852 | 7816248 | Glyma09g08550 | 7845409 | 7851685 | 3.7 |
| Glyma.09G016600*** | 1285132 | 1290884 | Glyma09g01940 | 1270010 | 1276140 | 2.7 | |||||
| Glyma.09G167000*** | 39103764 | 39109664 | Glyma09g29710 | 36530532 | 36536435 | ||||||
| qRAF-2 | Gm09_1888876 | 2.47 | 7.6 | Glyma.09G073600*** | 7809852 | 7816248 | Glyma09g08550 | 7845409 | 7851685 | 5.9 | |
| Glyma.09G016600*** | 1285132 | 1290884 | Glyma09g01940 | 1270010 | 1276140 | 0.6 | |||||
| Glyma.09G167000* | 39103764 | 39109664 | Glyma09g29710 | 36530532 | 36536435 | 32.4 | |||||
| qRAF-3 | Gm12_6023395-Gm12_2379195 | 2.15 | 4.7 | Glyma.12G162600* | 30862398 | 30862873 | Glyma12g26693 | 30087270 | 30088386 | 24.06 |
Table 6.
QTL and candidate genes that control sugars (sucrose, stachyose, and raffinose) contents in FxW82 RIL population in Carbondale, IL in 2020. These QTL have been identified by CIM method. Genes with (***) are apart from the identified QTL with less than 10 MB; Genes with (**) are apart from the identified QTL with less than 20 MB; Genes with (*) are apart from the identified QTL with more than 20 MB.
Table 6.
QTL and candidate genes that control sugars (sucrose, stachyose, and raffinose) contents in FxW82 RIL population in Carbondale, IL in 2020. These QTL have been identified by CIM method. Genes with (***) are apart from the identified QTL with less than 10 MB; Genes with (**) are apart from the identified QTL with less than 20 MB; Genes with (*) are apart from the identified QTL with more than 20 MB.
| Trait | QTL | Marker | LOD | R2 | Wm82.a2.v1 | Start | End | Wm82.a1.v1.1 | Start | End | Dis. (MB) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sucrose | qSUC-1 | Gm02_1199805-Gm02_1373746 | 2.63 | 3.6 | Glyma.02G016700*** | 1490049 | 1491170 | Glyma02g02030 | 1475851 | 1476528 | 0.2 |
| qSUC-2 | Gm05_3803682-Gm05_3748078 | 2.1 | 0.03 | Glyma.05G040300*** | 3593378 | 3598821 | Glyma05g02510 | 1870330 | 1875692 | 1.8 | |
| Glyma.05G003900*** | 307460 | 312091 | Glyma05g08950 | 8806144 | 8810647 | 5.002 | |||||
| Glyma.05G217100* | 39735138 | 39739763 | Glyma05g36850 | 40599128 | 40603658 | 36.7 | |||||
| Glyma.05G185500* | 37243691 | 37249494 | Glyma05g31920 | 36953899 | 36959702 | 33.1 | |||||
| Glyma.05G236600* | 41293446 | 41294570 | Glyma05g34830 | 39054363 | 39055344 | 35.2 | |||||
| Glyma.05G204700* | 38804305 | 38807296 | Glyma05g38120 | 41530564 | 41533554 | 37.7 | |||||
| qSUC-3 | Gm08_5960619-Gm08_8268861 | 2.37 | 0.04 | Glyma.08G043800*** | 3450235 | 3451725 | Glyma08g04860 | 3446035 | 3447462 | 2.5 | |
| Glyma.08G143500*** | 10949673 | 10956219 | Glyma08g15220 | 11038816 | 11045375 | 2.7 | |||||
| Glyma.08G011800*** | 942037 | 944988 | Glyma08g01480 | 939512 | 942346 | 5.01 | |||||
| Glyma.08G023100*** | 1852651 | 1856671 | Glyma08g02690 | 1848105 | 1853380 | 4.1 | |||||
| Stachyose | qSTA-1 | Gm13_2748576 | 2.03 | 0.09 | Glyma.13G160100* | 27576191 | 27579282 | Glyma13g22890 | 26380083 | 26383137 | 23.6 |
| Glyma.13G114000** | 22767704 | 22773231 | Glyma13g17420 | 21211880 | 21217237 | 18.4 | |||||
| qSTA-2 | Gm16_3183754-Gm16_3010888 | 2.85 | 3.92 | Glyma.16G217200* | 37414228 | 37419838 | Glyma16g34290 | 36921346 | 36926746 | 33.7 | |
| qSTA-3 | Gm17_8449684-Gm17_8352493 | 2.37 | 3 | Glyma.17G037400*** | 2732048 | 2737399 | Glyma17g04160 | 2739794 | 2745132 | 5.6 | |
| Glyma.17G045800*** | 3404918 | 3410491 | Glyma17g05067 | 3412682 | 3418160 | 4.9 | |||||
| Glyma.17G035800*** | 2629011 | 2639005 | Glyma17g03990 | 2637080 | 2646732 | 5.8 | |||||
| Glyma.17G111400*** | 8744555 | 8747526 | Glyma17g11970 | 9015075 | 9018145 | 0.5 | |||||
| qSTA-4 | Gm20_294157-Gm20_1133712 | 3.59 | 4.5 | Glyma.20G177200* | 41446962 | 41451980 | Glyma20g31730 | 40330117 | 40334860 | 40.03 | |
| Glyma.20G095200* | 33827363 | 33831352 | Glyma20g22780 | 32686241 | 32690264 | 32.3 | |||||
| Glyma.20G094500* | 33759416 | 33761555 | Glyma20g22700 | 32618509 | 32620443 | 32.3 |
Table 7.
Candidate genes controlling sugars (sucrose, stachyose, and raffinose) contents associated with previously reported QTL.
Table 7.
Candidate genes controlling sugars (sucrose, stachyose, and raffinose) contents associated with previously reported QTL.
| Gene ID | Start | End | QTL | QTL Start | QTL End | Reference |
|---|---|---|---|---|---|---|
| Glyma.02G240400 | 42892680 | 42898279 | Seed sucrose 2-2 | 39547350 | 41441274 | [41] |
| Seed oligosaccharide 1-1 | 39547350 | 41441274 | [41] | |||
| Glyma.05G236600 | 41293446 | 41294570 | Seed sucrose 1-1 | 3924139 | 4279362 | [39] |
| Glyma.08G043800 | 3450235 | 3451725 | Seed sucrose 1-3 | 7892162 | 8937354 | [39] |
| Glyma.08G143500 | 10949673 | 10956219 | Seed sucrose 1-2 | 10865328 | 13126779 | [39] |
| Glyma.09G073600 | 7809852 | 7816248 | Seed sucrose 4-2 | 2973041 | 5901485 | [44] |
| Glyma.13G114000 | 22767704 | 22773231 | Seed sucrose 1-5 | 26196486 | 28912864 | [39] |
| Glyma.14G209900 | 47515899 | 47521687 | Seed sucrose 3-1 | 38859467 | 40060720 | [40] |
| Seed oligosaccharide 2-1 | 38859467 | 40060720 | [40] | |||
| Glyma.15G151000 | 12497113 | 12508050 | Seed sucrose 3-3 | 13755345 | 17021739 | [40] |
| Seed oligosaccharide 2-3 | 13755345 | 17021739 | [40] | |||
| Glyma.19G140700 | 40199041 | 40201038 | Seed sucrose 1-8 | 40205349 | 40265091 | [39] |
| Seed oligosaccharide 2-7 | 42119600 | 43329204 | [40] | |||
| Glyma.19G212800 | 46633685 | 46639818 | Seed oligosaccharide 2-7 | 42119600 | 43329204 | [40] |
| qSU1901 | 45311975 | 45464136 | [43] | |||
| Glyma.19G217700 | 47033812 | 47037286 | Seed oligosaccharide 2-7 | 42119600 | 43329204 | [40] |
| qSU1901 | 45311975 | 45464136 | [43] | |||
| Glyma.20G095200 | 33827363 | 33831352 | Seed sucrose 1-4 | 2716974 | 25498552 | [39] |
| Glyma.08G011800 | 942037 | 944988 | Seed sucrose 1-3 | 7892162 | 8937354 | [39] |
| Seed sucrose 1-13 | 8283676 | 9192408 | [39] | |||
| Glyma.08G023100 | 1852651 | 1856671 | Seed sucrose 1-3 | 7892162 | 8937354 | [39] |
| Seed sucrose 1-13 | 8283676 | 9192408 | [39] | |||
| Glyma.19G219100 | 47148224 | 47150373 | Seed sucrose 1-8 | 40205349 | 40265091 | [39] |
| Seed sucrose 2-10 | 40637071 | 41616190 | [41] | |||
| Seed sucrose 2-11 | 40637071 | 41616190 | [41] | |||
| Seed oligosaccharide 2-7 | 42119600 | 43329204 | [40] | |||
| Glyma.19G227800 | 47911129 | 47914214 | Seed sucrose 1-8 | 40205349 | 40265091 | [39] |
| Seed sucrose 2-10 | 40637071 | 41616190 | [41] | |||
| Seed sucrose 2-11 | 40637071 | 41616190 | [41] | |||
| Seed oligosaccharide 2-7 | 42119600 | 43329204 | [40] | |||
| Glyma.20G094500 | 33759416 | 33761555 | Seed sucrose 1-4 | 2716974 | 25498552 | [39] |
| Glyma.20G177200 | 41446962 | 41451980 | qSU2002 | 40523599 | 41882459 | [43] |
| Glyma.15G182600 | 17910130 | 17916426 | Seed sucrose 3-3 | 13755345 | 17021739 | [40] |
| Seed oligosaccharide 2-3 | 13755345 | 17021739 | [40] | |||
| Glyma.05G003900 | 307460 | 312091 | Seed sucrose 1-1 | 3924139 | 4279362 | [39] |
| Glyma.09G016600 | 1285132 | 1290884 | Seed sucrose 4-2 | 2973041 | 5901485 | [44] |
| Glyma.17G111400 | 8744555 | 8747526 | qSS1701 | 7470395 | 10014816 | [43] |
| qSS1702 | 7969537 | 10599548 | [43] | |||
| Glyma.13G160100 | 27576191 | 27579282 | Seed sucrose 1-5 | 26196486 | 28912864 | [39] |
| Glyma.19G004400 | 359933 | 363588 | Seed sucrose 2-3 | 4244065 | 12744826 | [41] |
| Seed oligosaccharide 1-2 | 4244065 | 12744826 | [41] | |||
| Seed sucrose 2-6 | 9284015 | 34059981 | [41] | |||
| Seed oligosaccharide 1-5 | 9284015 | 34059981 | [41] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



