
Submitted:
30 August 2023
Posted:
31 August 2023
You are already at the latest version
Abstract
This study investigates the synergistic impact of fermenting kale juice with Limosilactobacillus strains on its antioxidant and anti-inflammatory properties. Kale's rich nutrient profile, especially its flavonoids, offers potential health benefits. Probiotic lactic acid bacteria are employed in kale fermentation to enhance nutrient bioavailability and generate bioactive compounds. Kale juices fermented with L. reuteri EFEL6901 or L. fermentum EFEL6800 exhibited superior microbial growth. Free sugars and amino acids were converted to alcohols and organic acids, affecting the organoleptic and health-related properties of product. In addition, fermentation increased quercetin and kaempferol content, indicating improved availability. Furthermore, the fermented juice exhibited notable antioxidant activity and suppressed NO production, revealing anti-inflammatory potential. Gene expression analysis confirmed reduced pro-inflammatory markers and elevated anti-inflammatory cytokines. This research highlights the promising potential of fermented kale juice, enriched with Limosilactobacillus strains, as a functional food with combined antioxidant and anti-inflammatory benefits.
Keywords:
kale juice
; probiotics
; Limosilactobacillus reuteri
; Limosilactobacillus fermentum
; Anti-inflammatory
1. Introduction
The development of novel nondairy-fermented products utilizing vegetables, fruits, legumes, or cereals has garnered significant attention in recent times [1]. These products not only exhibit probiotic properties but also serve as abundant sources of phytochemicals and phytonutrients [2,3]. Simultaneously, there has been a growing interest in fermented juices made from fruits or vegetables among health-conscious consumers and researchers, driven by the increasing popularity of veganism [4]. Fermented vegetable juices have emerged as a promising category within the functional foods market, offering not only nutritional benefits but also the potential for innovative flavors [5]. This trend aligns with the rising consumer demand for healthier and more sustainable food choices, while also addressing the need for convenient and versatile products suited for today's fast-paced lifestyle [6].
Kale is a vegetable belonging to the genus Brassica, species Brassica oleracea, group acephala, appears frequently in the 'healthiest foods' or 'superfoods' [7]. This leafy green vegetable is highly regarded for its abundant nutritional value, encompassing flavonoids, antioxidant enzymes, and other trace compounds [8]. Notably, kale stands out for its rich content of flavonoids, specifically quercetin and kaempferol, which surpass the levels found in other vegetables such as collard greens, mustard greens, sweet potato greens, and green onion [9]. These components exhibit significant potential, not only for disease prevention but also as therapeutic agents. Previous studies suggested that kale may contribute significantly to the management of chronic conditions such as obesity, cholesterol reduction, and inflammatory bowel disease [10].
The concept of a starter culture involves the addition of one or more microorganisms to raw materials, facilitating and controlling the fermentation process in production of fermented foods [11]. Particularly, lactic acid bacteria (LAB) are widely employed as starter culture for vegetable fermentation such as kimchi, sauerkraut, and tempeh [12]. LAB can enhance the bioavailability and digestibility of nutrients present in the vegetables, making them more easily absorbed by the body, and produce bioactive metabolites that exhibit antioxidant properties [13,14]. Moreover, LAB can contribute to sensory properties by breaking down various food components in vegetable into flavor precursors and converting them into aromatic compounds including acetoin and diacetyl [15].
Inflammation is a crucial process that safeguards the host against external threats, such as bacteria, viruses, and toxins, by eliminating pathogens and promoting tissue recovery [16]. However, prolonged inflammation can disrupt immune tolerance, cause significant physiological changes in tissues, organs, and normal cells, and increase the risk of various non-communicable diseases [17]. Kale contains phytochemicals, sulfur-containing indolic glucosinolates, and aliphatic glucosinolates that have demonstrated anti-inflammatory activity [18]. Phenolic compounds such as kaempferol and quercetin have been shown to possess anti-inflammatory effects by inhibiting the expression of PGE2, COX-2, and mPGES-1, which are upregulated by lipopolysaccharide (LPS)-induced inflammation [19]. Moreover, probiotics are widely used for the management of chronic diseases due to their ability to modulate the immune system and elicit anti-inflammatory responses [20,21]. Probiotic strains have shown anti-inflammatory effects by reducing the expression of pro-inflammatory cytokines, primarily through toll-like receptor (TLR)-mediated mechanisms [22]. Previous studies have reported that fermentation of kale juice using L. acidophilus IFO 3025 led to improvements in nutrient content, including calcium, phosphorus, and magnesium [6]. Fermentation of kale juice by various LAB has also been found to enhance its antioxidant activity and exhibit antibacterial effects, particularly against Gram-negative bacteria, during the fermentation process, thereby enhancing its physiological properties [23].
This study aimed to develop an enhanced antioxidative and anti-inflammatory product by fermenting kale juice with selected probiotic strains and assessing their health-promoting effects. Specifically, we utilized Limosilactobacillus reuteri EFEL6901 [24], L. fermentum EFEL6800 [24], L. fermentum MG7011 [25], and Leuconostoc citreum EFEL2061 [26], which have previously demonstrated probiotic properties with antioxidant and anti-inflammatory activities in relevant research. The fermentation characteristics of these strains were analysed in kale juice and the metabolites and flavonoid contents, such as kaempferol and quercetin, were measured. Moreover, the health functionality of the fermented kale juice was evaluated including its anti-inflammatory and antioxidant activities.
2. Materials and methods
2.1. Bacterial strains and growth conditions
The bacterial strains used in this study are presented in Table 1. For the assessment of probiotic activity, Lactiplantibacillus plantarum WCFS1 (WCFS1) and Lacticaseibacillus rhamnosus GG (LGG) were used as positive controls. All strains were preserved in a deep freezer at -80 ℃ in 50% glycerol (30% v/v). The strains were cultured in MRS broth (BD Difco, Franklin Lakes, NJ, USA) for 24 h under optimal anaerobic conditions. To initiate the juice fermentation process, the cultured strains were harvested through centrifugation (10,000 × g for 1 minute), washed twice with sterile 0.85% (w/v) NaCl, and used as inoculums.
2.2. Characteristics of fermented kale juices
Kale (Brassica oleraceae L. var. acephala DC.) was obtained from a local organic market in Cheongju, Korea. The leaves were trimmed to remove damaged portions and washed with tap water. A total of 600 g of kale was then ground using a grinder and mixed with 1800 mL of distilled water. The mixture was pasteurized at 70 ℃ for 30 minutes and inoculated with a 4% starter culture consisting of L. reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Le. citreum EFEL2061 at a final concentration of 106 CFU/mL. The inoculated samples were incubated at 37 ℃ for 72 h. Viable cell counts were determined by serial dilution with 0.85% NaCl and plating on MRS agar medium. The plates were incubated at 37 ℃ for 48 h, and the viable cell counts were expressed as a colony forming unit (CFU)/mL. The pH of the kale juices was measured using a pH meter (Orion Star A211, Thermo Fisher Scientific Inc., MA, USA), and the titratable acidity was determined by titrating with 0.01 N NaOH, expressed as the percentage of lactic acid in the samples. Color measurements of kale juices were performed using a colorimeter (Minolta CR-300, Minolta, Osaka, Japan) with calibration using white and black standard tiles. The samples were placed in transparent cuvettes for color detection, and the chromatic values L*, a*, and b* were recorded. Higher values of L*, a*, and b* indicate higher brightness, redness, and yellowness, respectively. The total color difference (ΔE*) of the samples was calculated using the following formula: . All measurements and analyses were performed in triplicate to ensure accuracy and reproducibility.
2.3. DPPH radical scavenging activity assay
Fermented kale juices were centrifuged at 11,000 × g for 10 min at 4 ℃, and the supernatants were collected. The DPPH (2,2-diphenyl-1-picrylhydrazyl) radical scavenging activities were measured according to the method of Hur [27]. A 200 μL volume of 0.2 mM DPPH solution (in methanol) was added to 50 μL of the fermented kale juice sample, followed by incubation in the dark for 30 min. The absorbance was measured at 517 nm. The various concentrations of ascorbic acid (1 mM, 0.5 mM, and 0.25 mM) were used as positive controls. The DPPH radical scavenging activities were calculated according to the following equation: DPPH scavenging activity (%) where, A sample: absorbance of the sample added group; A control: absorbance of the distilled water added group.
2.4. ABTS radical scavenging activity assay
The ABTS+ scavenging activities of samples were measured by modifying the method of Biglar [28]. ABTS+ radical cations were generated by reacting 7 mM 2,2'-azobis (2-amidinopropane) dihydrochloride and 2.45 mM potassium persulfate in distilled water at room temperature for 24 h in the dark. Before analysis, the ABTS+ solution was diluted with distilled water to adjust an absorbance of 1.00 ± 0.02 at 734 nm. Consecutively, 10 μL of fermented kale juices and 200 μL of ABTS+ solution was mixed, and the absorbances were measured at 734 nm after reaction for 6 min. The various concentrations of ascorbic acid (1 mM, 0.5 mM, and 0.25 mM) were used as positive controls. The ABTS+ radical scavenging activities were calculated according to the following equation:
ABTS radical scavenging activity (%) where, A sample: absorbance of the sample added group; A control: absorbance of the distilled water added group.
2.5. Quercetin and kaempferol analysis
For analysis of quercetin and kaempferol, the lyophilized fermented kale juice sample was extracted with 50% aqueous methanol and 1.5 M HCl (37.5 mL of 66.7% aqueous methanol and 12.5 mL of 6 M HCl) for twice. And then, refluxed at 90 ℃ for 2 h and the extract was filtered through a 0.45 μm PTFE filter for HPLC analysis [29]. HPLC analysis of the flavonoids was conducted using an Infinity 1260 HPLC (Agilent Technologies, Santa Clara, CA, USA) consisting of autosampler, the Phoroshell C18 column (4.6 × 150 mm, 120 Å, 2.7 µm) (Agilent technologies), and UV detector. The measurement was performed using the method of Rha [30]. In detail, the flow rate was set at 0.8 mL/min, and the gradient elution was made with mobile phase A (0.1% formic acid in water) and mobile phase B (0.1% formic acid in acetonitrile). The elution gradient was as follows: 0-8% B (0-2 min), 8-12% B (2-3 min), 12-16% B (3-4 min), 16% B (4-12 min), 16-20% B (12-15 min), 20% B (15-18 min), 20-24% B (18-21 min). 24-30% B (21-22 min), 30% B (22-26 min), 30-50% B (26-28 min), 50% B (28-30 min), 50-80% B (30-32 min), 80% B (32-33min), 80-8% B (33-34 min), and 8% B (34-35 min). The UV chromatograms recorded at 365 nm were used for quantification of quercetin (≥95%, Sigma-Aldrich), and kaempferol (≥90%, Sigma-Aldrich), based on external standard calibration curves.
2.61. H-NMR analysis
To analyse the metabolites of fermented kale juices, nuclear magnetic resonance (NMR) spectroscopy analysis was conducted. For the preparation of samples, fermented kale juices were centrifuged (11,000 × g, 10 min), then the supernatants were collected. The pH of supernatants was adjusted to 6.0 ± 0.2 and mixed with 350 μL of 1 mM sodium 2,2-dimethyl-2-silapentane-5-sulfonate (DSS; Sigma-Aldrich) as an internal standard in 10% deuterium oxide (D2O; Sigma-Aldrich). The mixtures (700 μL) were transferred to NMR tubes and subjected to 1H-NMR analysis. The 1H-NMR spectra were recorded on an Avance 500-MHz spectrometer (Bruker BioSpin, Karlsruhe, Germany). The metabolites peaks in the NMR spectra were identified, and its concentrations were calculated using Chenomx NMR suite 8.4 library software (Chenomx Inc, Edmonton, AB, Canada).
2.7. Cytotoxicity of fermented kale juice
The cytotoxicity was evaluated by the 3-(4, 5-dimethylthiazol-2-yl)-2, 5- diphenyltetrazolium bromide (MTT) assay using RAW 264.7 cell lines. The RAW 264.7 cell lines were obtained from the Korean Cell Line Bank (Seoul, Korea). Briefly, RAW 264.7 cells were seeded at 2.0×105 cells per well with DMEM containing 10% FBS in 96-well plates and cultured at 37 ℃ for 18–20 h. The cells were treated with 6.25-100 μg/mL of fermented kale juice extracted with DMSO. All samples were treated simultaneously with LPS (1 μg/mL)(Sigma-Aldrich). After incubating for 24 h, the medium was removed, and cells were washed twice with PBS. And then, MTT solution (0.25 mg/mL) (HiMedia, Mumbai, India) was added followed by incubation in the dark at 37 ℃ for 1 h. Finally, the formazan crystals were dissolved by adding 150 μL of dimethylsulfoxide, and the absorbance of each well was measured at 570 nm using a microplate spectrophotometer (BioTek, Santa Barbara, CA, USA). The percent cell viability was calculated according to the following equation: Cell viability (%) = () 100.
2.8. Measurement of nitric oxide (NO) production
The effect of kale juices fermented with different starters on the production of NO was determined in LPS-induced RAW 264.7 cell lines using Griess reaction [31]. RAW 264.7 cells (2×105 cells/well) were seeded in 96-well plates and incubated for 18-20 h. RAW 264.7 cells were treated with fermented kale juice at the above-mentioned concentrations followed by stimulation with LPS (1 μg/mL). Methyl arginine was used as the positive control. After 24 h of incubation, 100 μL of culture supernatants were transferred to 96-well plates and mixed with an equal volume of Griess reagent (Sigma-Aldrich) and placed in the dark for 15 min at room temperature. The absorbance of each mixture at 540 nm was measured using a microplate spectrophotometer (BioTek). Nitrite concentration was calculated using dilutions of sodium nitrite as standards, and fresh medium was used as the blank control.
2.9. mRNA expression level of nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2), and cytokine
The effect of fermented kale juice on the pro-inflammatory transcriptome was assessed in LPS-induced RAW 264.7 cells using an RT-qPCR. RAW 264.7 cells (1×106 cells/well) were seeded in 6-well plates and incubated for 24 h. Cells were treated with LPS (1 μg/mL) with kale juice fermented with different strains for 24 h. After incubation, cells were washed twice with sterile PBS, and total RNA was extracted with the with the Trizol RNA isolation reagent (Invitrogen, Carlsbad, USA) according to the manufacturer’s protocol. An adequate amount of RNA was reverse transcribed into cDNA using LeGene Express 1st Strand cDNA Synthesis System Kit (LeGene Biosciences, San Diego, CA, USA). Real-time PCR was conducted using Exicycler 96 Real-Time Quantitative Thermal Block (Exicycler 96; Bioneer, Daejeon, Korea). The mixtures containing synthesized cDNA, 10 pmol of specific primers, and PCR Master Mix containing SYBR Green Mix were amplified as follows: 95 ℃ for 5 min followed by 45 cycles at 95 ℃ for 15 sec, 59 ℃ for 30 sec with a final extension at 59 ℃ for 30 sec. The result was analysed after normalization with GAPDH as a reference gene. Relative expression levels of target genes were calculated with the delta-delta Ct (ΔΔCt) method. The specific primer sequences used in this study are listed in Table 2.
2.10. Enzyme and related genes
Genetic analysis was conducted to identify the genes related to utilization of phytochemical by a sequence similarity search using BLASTP. The sequences were obtained from the NCBI database (https://www.ncbi.nlm.nih.gov/).
2.11. Statistical analysis
Each experiment was conducted in triplicate, and the data were presented as the mean value ± standard deviation (SD). Statistical analysis was performed using IBM SPSS software version 22 (SPSS Inc., USA). Independent t-tests were used to analyse differences between two groups, and one-way analysis of variance (ANOVA) with Tukey’s method was used to analyse differences between multiple groups. Different letters and symbols on the error bars indicate significant differences. Statistical significance was basically set at p < 0.05.
3. Results
3.1. Quality characteristics of fermented kale juice
3.1.1. Microbial growth in kale juice
To evaluate the growth characteristics of different bacterial strains in fermented kale juice, four experimental groups were prepared: kale juice fermented with L. reuteri EFEL6901 (KJ-EFEL6901), L. fermentum EFEL6800 (KJ-EFEL6800), L. fermentum MG7011 (KJ-MG7011), and L. citreum EFEL2061 (KJ-EFEL2061). Four strains were inoculated at a concentration of 106 CFU/mL in pasteurized kale juice, and their growth rates were monitored under anaerobic conditions at 37 ℃ for 72 h. As shown in Figure 1A, all strains exhibited robust growth in kale juice. Notably, EFEL6901 and EFEL6800 demonstrated the most significant increase in cell count, reaching up to 9.0 Log CFU/mL, which corresponded to a 200-fold increment. The pH of the kale juice decreased progressively as the fermentation time advanced, as illustrated in Figure 1B. After 72 h of fermentation, the pH levels ranged from 4.5 to 4.8, depending on the strain. The total acidity of the kale juice also increased during fermentation, surpassing 0.10% in the case of EFEL6901, as shown in Figure 1C. These findings indicate that four bacterial strains are capable of propagating in kale juice, particularly EFEL6901 and EFEL6800 exhibiting more adapted characteristics in kale.
3.1.2. Chromaticity analysis
To investigate the color changes during the fermentation, kale juice samples were analysed using a colorimeter, and the results were indicated as lightness (L*), redness (a*), yellowness (b*), and total color difference (ΔE*) (Table 3). After the pasteurization process, the L*, a*, and b* values exhibited changes, showing an increase in lightness and yellowness. After the fermentation process, those values exhibited variable changes depending on the starters. The L* value remained relatively constant (34.61), except for the KJ-EFEL6901 sample (34.02), indicating minimal variation in lightness. The redness (a*) value showed a significant increase after fermentation. In contrast, there were no significant changes observed in yellowness (b*). The color differences (ΔE*) between pre- and post-fermentation samples demonstrated significant variations, ranging from 5.67 to 6.80. These results indicate that both pasteurization and fermentation can influence the color of kale juice with the extent of change dependent on the specific strain used.
3.1.3. Quantification of quercetin and kaempferol
To investigate the change in the flavonoid contents after kale juice fermentation, samples were extracted by reflux and analysed by HPLC (Figure 2). As a result, the initial concentrations of quercetin and kaempferol in kale juice were measured as 7.87 μM (45.03 mg/100 g) and 54.46 μM (311.78 mg/100 g), respectively, and these levels remained stable even after pasteurization. Following 72 h of fermentation, the flavonoid contents in four samples showed an increase. In quercetin content, KJ-EFEL6800 exhibited a significant increase, reaching 40.06 μM, which was five folds higher than the pre-fermentation level. KJ-EFEL6901 showed a three-fold increase in quercetin level (20.82 μM), while KJ-MG7011 and KJ-EFLE206 had slight increases of 11.42 and 11.64 μM, respectively. In kaempferol content, KJ-EFEL6901 and KJ-EFEL6800 exhibited about two-fold increases, reaching 97.82 and 101.59 μM, respectively, compared to the pre-fermentation level. KJ-EFEL2061 also demonstrated increased kaempferol content (78.62 μM). Overall, the flavonoid levels in kale juice remained stable after pasteurization, and their contents were significantly enhanced by the fermentation process, particularly in samples inoculated with EFEL6901 and EFEL6800.
3.1.41. H-NMR analysis
1H-NMR analysis was conducted to analyse the metabolite changes such as carbohydrates, alcohols, organic acids, and amino acids in fermented kale juice. As shown in Figure 3, during the fermentation, carbohydrates and amino acids were consumed as microbial cells grew, whereas alcohols and organic acids were produced with the microbial reduction reaction of nutrients. The detailed contents are presented in Table 4. First, the carbohydrate and amino acid contents in the pasteurized kale juice demonstrated rich and balanced nutrient composition in kale, which is essential as a microbial growth medium. This result is consistent with the robust growth of LAB observed in this study (Figure 1). In case of carbohydrates, glucose and fructose were identified as the major free sugars, and their concentrations decreased after fermentation in all kale juice samples. The reduction in glucose content was consistent across all strains, while the decrease in fructose varied, suggesting differences in fructose metabolic capability among the strains. The results revealed that mannitol, ethanol, acetate, and lactate were produced in all samples after fermentation, indicating hetero-lactic metabolic characteristics of LAB tested in this study. Glucose was mainly converted to ethanol, acetate, and lactate, and fructose was converted to mannitol. Regarding ethanol, the concentration was relatively high in KJ-EFEL6800 and KJ-MG7011, but low in KJ-EFEL6901 and KJ-EFEL2061, revealing the superior ethanol productivity of L. fermentum strains. In terms of organic acids, acetate, lactate, butyrate, propionate, and succinate were detected as the main metabolites, and their concentrations increased after fermentation. Among them, acetate was produced in the highest amount, followed by lactate, with their highest levels observed in KJ-EFEL6800 and KJ-EFEL6901. Succinate, propionate, and butyrate were also synthesized in all samples, while citrate decreased. Regarding amino acids, aspartate, alanine, arginine, glutamate, serine, and threonine were identified as major amino acids. Most amino acids showed a decrease after fermentation, except for cysteine and glycine, which increased in all kale juice samples. Particularly, the content of glutamate, which contributes to the umami taste when combined with sodium salt, significantly increased after fermentation in KJ-EFEL6901. Overall, these results indicate that, during fermentation, free sugars and amino acids in kale juice were converted to various metabolites such as alcohols (mannitol and ethanol) and organic acids (acetate, lactate, succinate, propionate, and butyrate), affecting the organoleptic and health-related properties of fermented products.
3.1.5. Enzyme and genes
The genetic analysis of EFEL6901 and EFEL6800 unveiled the presence of several enzymes, including glycoside hydrolase and glucosidase (Table 5), suggesting their potential involvement in the metabolism of kale's abundant glycosides. The breakdown of glycosidic bonds by these enzymes would release sugars, which could serve as a carbon source for the growth and proliferation of the microorganisms during fermentation. Furthermore, the increased levels of quercetin and kaempferol, observed after fermentation, suggest that the enzymatic activities of EFEL6901 and EFEL6800 played a significant role in enhancing the bioavailability of these bioactive compounds.
3.2. Health-related characteristics of fermented kale juice
3.2.1. Cytotoxicity
To assess the cytotoxicity of fermented kale juice, an MTT assay was conducted using RAW 264.7 cells. Figure 4 shows the cytotoxicity results of tested samples. KJ-EFEL6901 and KJ-EFEL6800 showed no apparent cytotoxicity (>95% cell viability) across all concentrations tested. Meanwhile, KJ-MG7011 and KJ-EFEL2061 exhibited toxicity within 6.25-25 μg/mL and 6.25-12.5 μg/mL, respectively. This result indicates that KJ-EFEL6901 and KJ-EFEL6800 are safe with no cytotoxicity to RAW 264.7 cells within the range of 6.25-100 μg/mL.
3.2.2. Antioxidant activities
To evaluate the antioxidative activity of fermented kale juice, DPPH and ABTS+ scavenging capacities were measured (Figure 5). In the experiment using DPPH (Figure 3A), four kale juices, KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061, exhibited significantly increased activities (57.9, 58.6, 55.6, and 55.1% respectively) compared to kale juice (26.9%) or pasteurized kale juice (29.9%). Their levels were comparable to the DPPH scavenging activity of 0.5 mM ascorbic acid (62.9%). As same, in the experiment using ABTS (Figure 3B), four kale juice showed significant increases in ABTS+ scavenging activity compared to the pre-fermentation samples. Their ABTS+ scavenging activities were in the order of KJ-EFEL6800, KJ-EFEL6901, KJ-MG7011, and KJ-EFEL2061. Their levels were also superior or comparable to the ABTS+ scavenging activity of 0.5 mM ascorbic acid. Overall, the fermentation of kale juice with the selected LAB strains resulted in a significant increase in antioxidant activity, and particularly it was superior in KJ-EFEL6901 and KJ-EFEL6800.
3.2.3. NO production
To assess the inhibitory activity of fermented kale juice on NO production in LPS-induced RAW 264.7 cells, the levels of NO in the cellular supernatants were measured (Figure 6). Treatment with LPS (1 μg/mL) significantly increased NO production compared to the control group (p < 0.001), while treatment with methyl arginine resulted in a dose-dependent decrease in NO production. When it comes to the fermented kale juices, their treatment significantly decreased NO production and exhibited higher inhibitory activity compared to the control juice. Among the fermented kale juices, KJ-EFEL6901 showed the highest inhibitory activity in reducing NO production (p < 0.01) compared to the other juices. It is noteworthy that KJ-MG7011 and KJ-EFEL2061 did not induce NO production at high concentrations (50-100 μg/mL) owing to their high cytotoxicity on RAW 264.7 cells.
3.2.4. mRNA levels of iNOS and COX-2
To analyse the anti-inflammatory effect of fermented kale juice, the transcription levels of iNOS and COX-2 in RAW 264.7 cells, which are related to the synthesis of NO or prostaglandin, were analysed using RT-qPCR (Figure 7). The results demonstrated that treatment with LPS (1 μg/mL) significantly increased the mRNA levels of iNOS and COX-2 compared to the control group. When it comes to the fermented kale juices, their treatment significantly reduced the mRNA levels of iNOS and COX-2 in a dose-dependent manner compared to PKJ. Among the tested juices, KJ-EFEL6901 and KJ-EFEL6800 exhibited superior inhibitory activities on the mRNA transcription of iNOS and COX-2 at 6.25 and 12.5 μg/mL concentrations.
3.2.5. mRNA levels of cytokines
To assess the anti-inflammatory effects of fermented kale juices, the transcription levels of IL-6 and IL-1ß as pro-inflammatory cytokines and IL-10 as an anti-inflammatory cytokine were measured in LPS-treated RAW 264.7 cells (Figure 7). The results demonstrated that treatment with LPS (1 μg/mL) significantly increased the mRNA levels of IL-6 and IL-1ß compared to the control group. In the case of fermented kale juices, their treatment significantly reduced the mRNA levels of IL-6 and IL-1ß in a dose-dependent manner, compared to PKJ. At the same time, their treatment except KJ-EFEL2061 significantly increased the transcription levels of IL-10 in a dose-dependent manner compared to PKJ, revealing their superior anti-inflammatory potential. Overall, these results indicate that the kale juices fermented with selected strains exhibited anti-inflammatory activities not only by down-regulating the mRNA levels of IL-6 and IL-1ß but also by up-regulating the mRNA levels of IL-10. Notably, KJ-EFEL6901 and KJ-EFEL6800 exhibited superior anti-inflammatory activities compared to PKJ, indicating the synergistic effects of kale juice and LAB starter. These findings highlight the potential of fermented kale juice in modulating the expression levels of pro-inflammatory and anti-inflammatory cytokines, suggesting its anti-inflammatory properties.
4. Discussion
Fermented foods are good carriers for delivering probiotics while having health-promoting effects [32]. In this study, EFEL6901 and EFEL6800 strains were selected as starters for kale juice fermentation. The two strains exhibited substantial cell growths in kale juice, reaching cell counts of > 9.0 log CFU/mL (a 200-fold increment), pH of 4.5~4.7 after 72 h of fermentation (Figure 1). Comparing these values with the growth of Streptococcus thermophilus, a commonly used starter culture for milk yogurt, which reached a cell count of 8.78 log CFU/mL and a pH of 4.3 [33], the fermentation characteristics of EFEL6901 and EFEL6800 in kale juice were comparable to the commercial product. The suitability of kale juice as a culture medium for selected LAB may be attributed to its rich nutrient composition. The pasteurized kale juice before fermentation contained various carbon sources such as glucose, fructose, mannose, sucrose, acetate, and citrate and different nitrogen sources of 18 amino acids with large amounts of aspartate and glutamate (Table 3). This result indicates that kale juice can serve not only as a suitable medium for the cell growth of the selected LAB strains but also as an excellent carrier for delivering probiotics.
Kale has abundant flavonoid content, specifically quercetin and kaempferol [29]. In this study, the initial concentrations of quercetin and kaempferol in kale juice were 7.87 μM (45.03 mg/100 g) and 54.46 μM (311.78 mg/100 g), respectively. Those levels are significantly higher than that of collard greens (12.4 and 43.3 mg/100g, respectively), mustard greens (8.8 and 38.3 mg/100g, respectively), sweet potato greens (27.9 and 5.0 mg/100g, respectively), okra (11.1 mg/100g quercetin and non-detectable kaempferol), green onion (non-detectable quercetin and 4.8 mg/100g kaempferol) [9]. Furthermore, this study shows that, after fermentation of kale juice with KJ-EFEL6901 and KJ-EFEL6800, the quercetin contents were increased three- and five-folds, respectively, and the kaempferol contents were increased about two-folds in both samples (Figure 2). Consequently, the higher level of flavonol contents in the kale juice was remarkably enhanced to the superior levels by a simple fermentation process using selected LAB.
The phenolic compounds found in plants can be divided into two categories: free phenolic compounds, which are located in the vacuoles of plant cells, and bound phenolic compounds that are covalently linked to structural components of the cell wall [34]. Fermentation has emerged as a favorable method for obtaining phenolic extracts of high quality and activity from different plant sources, utilizing techniques that are both economically and environmentally friendly [35]. Changes in phenolic compounds by fermentation process are associated with several enzymatic reactions: hydrolysis of ester bonds which link phenolic compounds to the cell wall matrix with esterase [36], the oxidative degradation of lignin with laccase or peroxidase [37], or deglycosylation of flavonoids to produce aglycone with β-glucosidase [38]. Several enzymes produced by microorganism can hydrolyse the glycosidic bonds in alkyl and aryl-β-D-glucoside, and β-glucosidase is a representative enzyme that produces aglycone [39]. For example, in soybean fermentation, L. plantarum WCFS1 could convert soybean isoflavones from glucosides to aglycones by β-glucosidase [40]. Also, L. plantarum CECT 748T had the ability to convert arylglucosides into their bioactive aglycone, increasing the antioxidant activity of plant foods during fermentation [41]. Likewise, the genetic analysis of EFEL6901 and EFEL6800 revealed the presence of multiple enzymes, such as glycoside hydrolase or glucosidase (Table 5), indicating their potential involvement in the utilization of kale's abundant glycosides. These enzymes likely played a significant role in facilitating the growth of microorganisms by releasing sugars and potentially contributed to the increased levels of quercetin and kaempferol after fermentation. These findings suggest that the efficiency of bio-conversion can be influenced by the strain's adaptability to kale juice and its ability to produce hydrolytic enzymes [42].
Antioxidants are major contributors to the functionality of the vegetable product [43]. Overall, antioxidant activity was significantly increased in fermented kale juice, and this result might be associated with the increased flavonol contents after fermentation (Figure 3). Notably, flavonol aglycones, quercetin and kaempferol, in plants are potent antioxidants that serve to protect the plant from reactive oxygen species (ROS) [44].
The anti-inflammatory effects of quercetin and kaempferol have been demonstrated through the inhibition of PGE2, COX-2, and mPGES-1 expression, which are upregulated during LPS-induced inflammation [19]. Additionally, probiotic strains have been shown to reduce the expression of pro-inflammatory cytokines, primarily TLR-mediated mechanisms, thus exerting their anti-inflammatory effects [22]. In this study, KJ-EFEL6901 and KJ-EFEL6800 resulted in a strong anti-inflammatory effect on LPS-induced RAW 264.7 cells by inhibiting NO production via down-regulating pro-inflammatory cytokines and up-regulating anti-inflammatory cytokine (Figure 4 and Figure 5). LPS activation in the mouse macrophage cell line RAW 264.7 induces an inflammatory response, producing inflammatory cytokines such as reactive oxygen species, IL-6, and TNF-α [45]. IL-6 and IL-1β are pro-inflammatory cytokines, and IL-1β induces several genes involved with inflammation and tissue destruction, such as Th1 cell responses [46]. And IL-6 acts as a chronic stressor that can accelerate the risk of age-related diseases by promoting the differentiation and inflammation of immune cells such as T and B cells [47]. Meanwhile, IL-10 plays an anti-inflammatory role in returning the immune system to a dormant state by inhibiting activated macrophage. Notably, the results of this study suggest that the anti-inflammatory effect of LBRE6901-KJ and LBF6800-KJ was improved compared to that of kale juice, possibly due to LAB themselves and/or their metabolites synthesized during fermentation. In our previous studies, strong antioxidative and anti-inflammatory activities of L. reuteri EFEL6901 and L. fermentum EFEL6800 have been demonstrated in in vitro and in vivo experiments [24,25]. Therefore, the superior levels of antioxidant and an-inflammatory activities observed in KJ-EFEL6901 and KJ-EFEL6800 were attributed to the dual sources that are kale, a flavonoid-rich vegetable, and L. reuteri EFEL6901 and L. fermentum EFEL6800, probiotic starters.
5. Conclusion
In this study, EFEL6901 and EFEL6800 strains exhibited superior microbial growth (> 9.0 log CFU/mL) and proper acidity change (pH4.5~4.7) in kale juice, indicating their suitability as microbial starters for kale fermentation. After fermentation, the quercetin and kaempferol contents were dramatically increased. The superior levels of antioxidant and anti-inflammatory activities observed in KJ-EFEL6901 and KJ-EFEL6800 were attributed to both kale and LAB strains in a synergistic manner. This study indicates that the kale juices fermented with EFEL6901 and EFEL6800 strains can be a superfood serving abundant nutrients like quercetin and kaempferol as well as carrying health-promoting probiotics.
Author Contributions
Conceptualization, Kim G. Y. and Han NS;Methodology, Seong H. Kong S. Y. and Bae J.-H.;Investigation, Kim G. Y. and Kim S.-A.;Original Draft Preparation, Kim G. Y. ;Writing – review & editing, Kim S.-A. and Han N. S; Supervision, Han N. S.
Funding
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (IPET) through Technology Commercialization Support Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA) (122040-02-1-SB010).
Acknowledgments
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (IPET) through Technology Commercialization Support Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA) (122040-02-1-SB010). This work was based on the thesis submitted by the first author Kim GY for Master’s Thesis, Chungbuk National University, 2022.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Marrero, S. C.; Martínez-Rodríguez, A.; Pérez, S. E. M.; Moya, S. P. New Trends and Applications in Fermented Beverages. Fermented Beverages, Elsevier, 2019, pp 31-66. [CrossRef]
- Septembre-Malaterre, A.; Remize, F.; Poucheret, P. Fruits and Vegetables, as a Source of Nutritional Compounds and Phytochemicals: Changes in Bioactive Compounds during Lactic Fermentation. Food Res Int. 2018, 104, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Michalak, M.; Kubik-Komar, A.; Waśko, A.; Polak-Berecka, M. Starter Culture for Curly Kale Juice Fermentation Selected using Principal Component Analysis. Food Biosci. 2020, 35, 100602. [Google Scholar] [CrossRef]
- Szutowska, J. Functional Properties of Lactic Acid Bacteria in Fermented Fruit and Vegetable Juices: A Systematic Literature Review. Eur Food Res Technol. 2020, 246, 357–372. [Google Scholar] [CrossRef]
- Chung, H.-J.; Lee, H.; Na, G.; Jung, H.; Kim, D.-G.; Shin, S.-I.; Jung, S.-E.; Choi, I.-d.; Lee, J.-H.; Sim, J.-H. Metabolic and Lipidomic Profiling of Vegetable Juices Fermented with Various Probiotics. Biomolecules. 2020, 10, 725. [Google Scholar] [CrossRef]
- Kim, S. Y. Production of Fermented Kale Juices with Lactobacillus Strains and Nutritional Composition. Prev Nutr Food Sci. 2017, 22, 231. [Google Scholar] [CrossRef]
- Šamec, D.; Urlić, B.; Salopek-Sondi, B. Kale (Brassica oleracea var. acephala) as a Superfood: Review of the Scientific Evidence behind the Statement. Crit Rev food Sci Nutr. 2019, 59, 2411–2422. [Google Scholar] [CrossRef]
- Han, J.-H.; Lee, H.-J.; Kim, T.-S.; Kang, M.-H. The Effect of Glutathione S-Transferase M1 and T1 Polymorphisms on Blood Pressure, Blood Glucose, and Lipid Profiles following the Supplementation of Kale (Brassica oleracea acephala) Juice in South Korean Subclinical Hypertensive Patients. Nutr Res Pract. 2015, 9, 49–56. [Google Scholar] [CrossRef]
- Huang, Z.; Wang, B.; Eaves, D. H.; Shikany, J. M.; Pace, R. D. Phenolic Compound Profile of Selected Vegetables Frequently Consumed by African Americans in the Southeast United States. Food Chem. 2007, 103, 1395–1402. [Google Scholar] [CrossRef]
- Zielińska, M.; Lewandowska, U.; Podsędek, A.; Cygankiewicz, A. I.; Jacenik, D.; Sałaga, M.; Kordek, R.; Krajewska, W. M.; Fichna, J. Orally Available Extract from Brassica Oleracea var. Capitata Rubra Attenuates Experimental Colitis in Mouse Models of Inflammatory Bowel Diseases. J Funct Foods. 2015, 17, 587–599. [Google Scholar] [CrossRef]
- Leroy, F.; De Vuyst, L. Lactic acid Bacteria as Functional Starter Cultures for the Food Fermentation Industry. Trends Food Sci Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Tamang, J. P.; Cotter, P. D.; Endo, A.; Han, N. S.; Kort, R.; Liu, S. Q.; Mayo, B.; Westerik, N.; Hutkins, R. Fermented Foods in a Global Age: East Meets West. Compr Rev Food Sci Food Saf 2020, 19, 184–217. [Google Scholar] [CrossRef]
- Hess, J. M. Role of Lactic Acid Bacteria in Impacting Nutrient Bioavailability. Lactic Acid Bacteria: A Functional Approach. 2020, 3, 35–57. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, X.; Xu, H.; Sun, Y.; Zhang, Y.; Wang, Y. Improvement of the Nutritional, Antioxidant and Bioavailability Properties of Corn Gluten-Wheat Bran Mixture Fermented with Lactic Acid Bacteria and Acid Protease. LWT 2021, 144, 111161. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, L.; Wen, R.; Chen, Q.; Kong, B. Role of Lactic Acid Bacteria in Flavor Development in Traditional Chinese Fermented Foods: A Review. Crit Rev Food Sci Nutr. 2022, 62, 2741–2755. [Google Scholar] [CrossRef] [PubMed]
- Khatami, M. Unresolved Inflammation:‘Immune Tsunami’or Erosion of Integrity in Immune-Privileged and Immune-Responsive Tissues and Acute and Chronic Inflammatory Diseases or Cancer. Expert Opin Biol Ther. 2011, 11, 1419–1432. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D. W.; Fasano, A.; Miller, G. W. Chronic Inflammation in the Etiology of Disease across the Life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Shahinozzaman, M.; Raychaudhuri, S.; Fan, S.; Obanda, D. N. Kale Attenuates Inflammation and Modulates Gut Microbial Composition and Function in C57BL/6J Mice with Diet-Induced Obesity. Microorganisms. 2021, 9, 238. [Google Scholar] [CrossRef]
- Hämäläinen, M.; Nieminen, R.; Asmawi, M. Z.; Vuorela, P.; Vapaatalo, H.; Moilanen, E. Effects of Flavonoids on Prostaglandin E2 Production and on COX-2 and mPGES-1 Expressions in Activated Macrophages. Planta Medica 2011, 77, 1504–1511. [Google Scholar] [CrossRef]
- De Marco, S.; Sichetti, M.; Muradyan, D.; Piccioni, M.; Traina, G.; Pagiotti, R.; Pietrella, D. Probiotic Cell-Free Supernatants Exhibited Anti-Inflammatory and Antioxidant Activity on Human Gut Epithelial Cells and Macrophages Stimulated with LPS. Evid based Complement Altern Med. 2018. [Google Scholar] [CrossRef]
- Cristofori, F.; Dargenio, V. N.; Dargenio, C.; Miniello, V. L.; Barone, M.; Francavilla, R. Anti-Inflammatory and Immunomodulatory Effects of Probiotics in Gut Inflammation: a Door to the Body. Front. Immunol. 2021, 12, 578386. [Google Scholar] [CrossRef]
- Plaza-Díaz, J.; Ruiz-Ojeda, F. J.; Vilchez-Padial, L. M.; Gil, A. Evidence of the Anti-Inflammatory Effects of Probiotics and Synbiotics in Intestinal Chronic Diseases. Nutrients. 2017, 9, 555. [Google Scholar] [CrossRef]
- Michalak, M.; Szwajgier, D.; Paduch, R.; Kukula-Koch, W.; Waśko, A.; Polak-Berecka, M. Fermented Curly Kale as a New Source of Gentisic and Salicylic Acids with Antitumor Potential. J Func Foods. 2020, 67, 103866. [Google Scholar] [CrossRef]
- Seo, H.; Seong, H.; Kim, G. Y.; Jo, Y. M.; Cheon, S. W.; Song, Y.; Ryu, B. H.; Kang, H.; Han, N. S. Development of Anti-Inflammatory Probiotic Limosilactobacillus reuteri EFEL 6901 as Kimchi Starter: In vitro and In vivo Evidence. Front Microb. 2021, 12, 760476. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y. M.; Kim, G. Y.; Kim, S.-A.; Cheon, S. W.; Kang, C.-H.; Han, N. S. Limosilactobacillus fermentum MG7011: an Amylase and Phytase Producing Starter for the Preparation of Rice-based Probiotic Beverages. Front Microb. 2021, 12, 745952. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Moon, J. S.; Lee, M.-G.; Han, N. S. Immunomodulatory Effects of Leuconostoc citreum EFEL2061 isolated from Kimchi, a Traditional Korean Food, on the Th2 Type-Dominant Immune Response in vitro and in vivo. J Func Foods. 2016, 20, 79–87. [Google Scholar] [CrossRef]
- Hur, S. J.; Lee, S. Y.; Kim, Y.-C.; Choi, I.; Kim, G.-B. Effect of Fermentation on the Antioxidant Activity in Plant-based Foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Biglari, F.; AlKarkhi, A. F.; Easa, A. M. Antioxidant activity and Phenolic Content of Various Date Palm (Phoenix dactylifera) Fruits from Iran. Food Chem. 2008, 107, 1636–1641. [Google Scholar] [CrossRef]
- Hagen, S. F.; Borge, G. I. A.; Solhaug, K. A.; Bengtsson, G. B. Effect of Cold Storage and Harvest Date on Bioactive Compounds in Curly Kale (Brassica oleracea L. var. acephala). Postharvest Biol Technol. 2009, 51, 36–42. [Google Scholar] [CrossRef]
- Rha, C.-S.; Seong, H.; Jung, Y. S.; Jang, D.; Kwak, J.-G.; Kim, D.-O.; Han, N. S. Stability and Fermentability of Green Tea Flavonols in in-vitro-Simulated Gastrointestinal Digestion and Human Fecal Fermentation. Int J Mol Sci. 2019, 20, 5890. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.-S.; Lee, N.-K.; Choi, A.-J.; Choe, J.-S.; Bae, C. H.; Paik, H.-D. Anti-Inflammatory Potential of Probiotic Strain Weissella cibaria JW15 Isolated from Kimchi through Regulation of NF-κB and MAPKs Pathways in LPS-Induced RAW 264.7 cells. J Microbiol Biotechnol. 2019. [Google Scholar] [CrossRef]
- Tangyu, M.; Muller, J.; Bolten, C. J.; Wittmann, C. Fermentation of Plant-based Milk Alternatives for Improved Flavour and Nutritional Value. Appl Microbiol Biotechnol. 2019, 103, 9263–9275. [Google Scholar] [CrossRef] [PubMed]
- Dave, R. I.; Shah, N. P. Viability of Yoghurt and Probiotic Bacteria in Yoghurts Made from Commercial Starter Cultures. Int Dairy J. 1997, 7, 31–41. [Google Scholar] [CrossRef]
- Pinelo, M.; Arnous, A.; Meyer, A. S. Upgrading of Grape Skins: Significance of Plant Cell-Wall Structural Components and Extraction Techniques for Phenol Release. Trends Food Sci Technol. 2006, 17, 579–590. [Google Scholar] [CrossRef]
- Martins, S.; Mussatto, S. I.; Martínez-Avila, G.; Montañez-Saenz, J.; Aguilar, C. N.; Teixeira, J. A. Bioactive Phenolic Compounds: Production and Extraction by Solid-State Fermentation. A Review. Biotechnol Adv. 2011, 29, 365–373. [Google Scholar] [CrossRef]
- Acosta-Estrada, B. A.; Gutiérrez-Uribe, J. A.; Serna-Saldívar, S. O. Bound Phenolics in Foods, a Review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef]
- Wang, X.; Yao, B.; Su, X. Linking Enzymatic Oxidative Degradation of Lignin to Organics Detoxification. Int J Mol Sci. 2018, 19, 3373. [Google Scholar] [CrossRef]
- Park, J. S.; Rho, H. S.; Kim, D. H.; Chang, I. S. Enzymatic Preparation of Kaempferol from Green Tea Seed and its Antioxidant Activity. J Agric Food Chem. 2006, 54, 2951–2956. [Google Scholar] [CrossRef]
- Nguyen Thai, H.; Van Camp, J.; Smagghe, G.; Raes, K. Improved Release and Metabolism of Flavonoids by Steered Fermentation Processes: a Review. Int J Mol Sci. 2014, 15, 19369–19388. [Google Scholar] [CrossRef]
- Gan, R. Y.; Shah, N. P.; Wang, M. F.; Lui, W. Y.; Corke, H. Lactobacillus plantarum WCFS1 Fermentation Differentially Affects Antioxidant Capacity and Polyphenol Content in Mung Bean (Vigna radiata) and Soya Bean (Glycine max) milks. J Food Process Preserv. 2017, 41, e12944. [Google Scholar] [CrossRef]
- Landete, J. M.; Curiel, J. A.; Rodríguez, H.; de las Rivas, B.; Munoz, R. Aryl Glycosidases from Lactobacillus plantarum Increase Antioxidant Activity of Phenolic Compounds. J Func Foods. 2014, 7, 322–329. [Google Scholar] [CrossRef]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M. T.; Wu, M.; Sackey, A. S.; Xiao, L.; Tahir, H. E. Effect of Lactobacillus Strains on Phenolic Profile, Color Attributes and Antioxidant Activities of Lactic-Acid-Fermented Mulberry Juice. Food Chem. 2018, 250, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Stübler, A.-S.; Lesmes, U.; Heinz, V.; Rauh, C.; Shpigelman, A., Aganovic, K. Digestibility, Antioxidative Activity and Stability of Plant Protein-Rich Products after Processing and Formulation with Polyphenol-Rich Juices: Kale and Kale–Strawberry as a Model. Eur Food Res Technol, 2019, 245, 2499-2514. [CrossRef]
- Jeganathan, B.; Punyasiri, P.; Kottawa-Arachchi, J. D.; Ranatunga, M. A.; Abeysinghe, I. S. B.; Gunasekare, M.; Bandara, B. Genetic Variation of Flavonols Quercetin, Myricetin, and Kaempferol in the Sri Lankan Tea (Camellia sinensis L.) and Their Health-Promoting Aspects. I J Food Sci. 2016. [CrossRef]
- Lampiasi, N.; Montana, G. The Molecular Events behind Ferulic Acid Mediated Modulation of IL-6 Expression in LPS-Activated Raw 264.7 Cells. Immunobiology. 2016, 221, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Blonska, M.; Czuba, Z.; Krol, W. Effect of Flavone Derivatives on Interleukin-1β (IL-1β) mRNA Expression and IL-1β Protein Synthesis in Stimulated RAW 264.7 Macrophages. Scandinavian J Immunol. 2003, 57, 162–166. [Google Scholar] [CrossRef]
- Saiki, P.; Nakajima, Y.; Van Griensven, L. J.; Miyazaki, K. Real-Time Monitoring of IL-6 and IL-10 Reporter Expression for Anti-Inflammation Activity in Live RAW 264.7 Cells. Biochem Biophys Res Commun, 2018, 505, 885–890. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Cell growth (A), pH changes (B), titratable acidity (C) of kale juices fermented at 37 ℃. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juices fermented with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively. Results are expressed as means ± standard deviations (n=3).
Figure 1.
Cell growth (A), pH changes (B), titratable acidity (C) of kale juices fermented at 37 ℃. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juices fermented with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively. Results are expressed as means ± standard deviations (n=3).
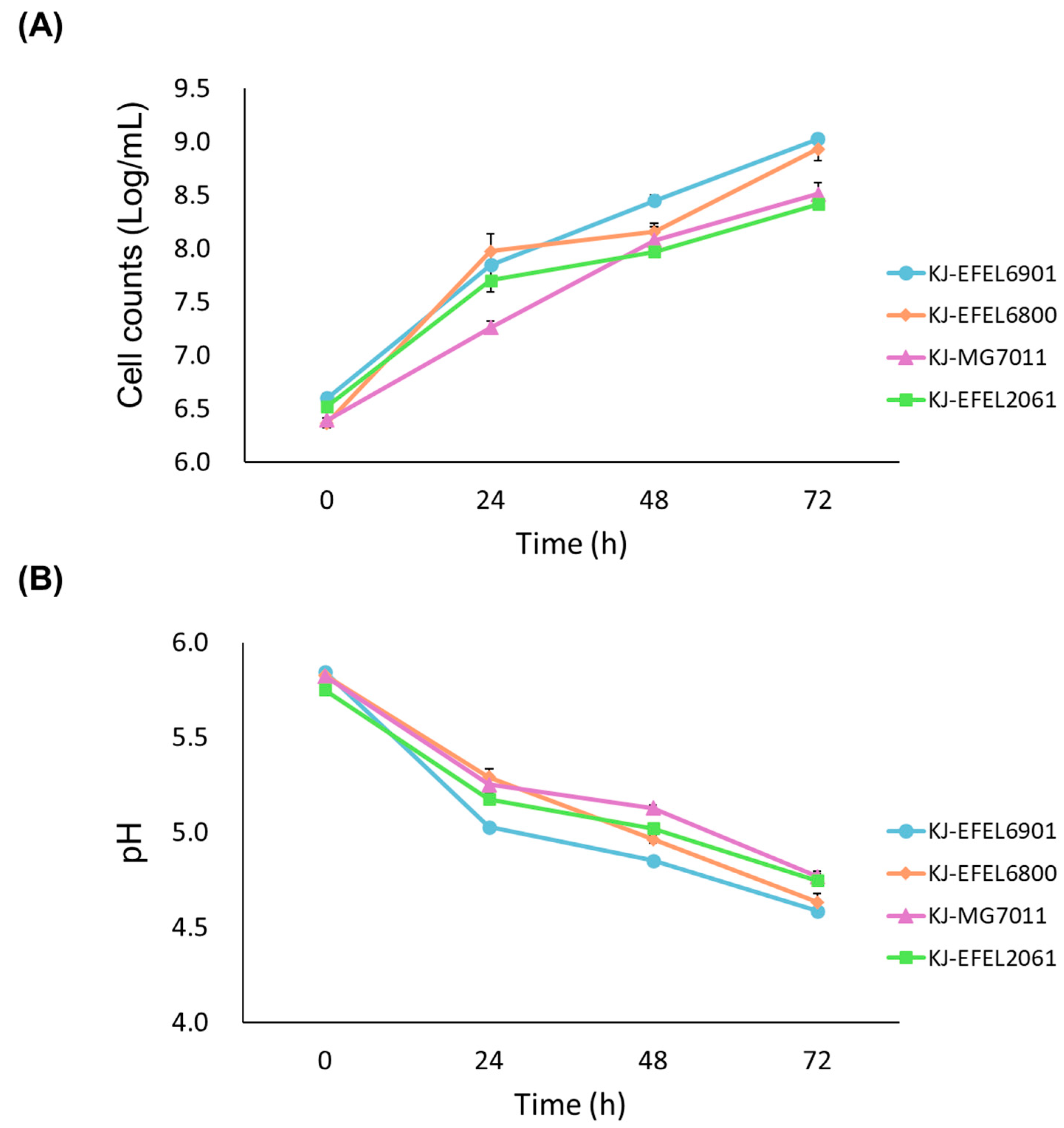
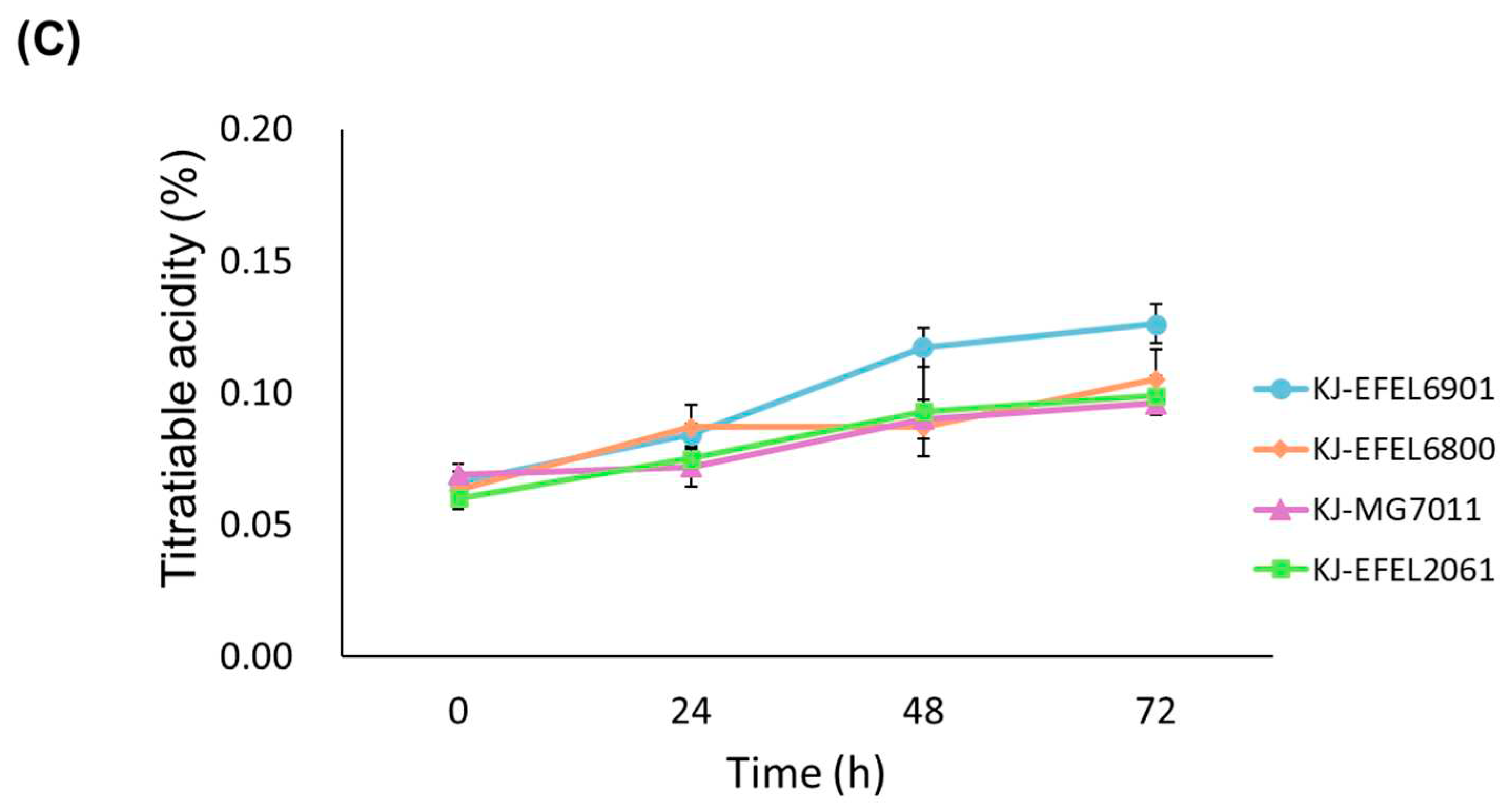
Figure 2.
Quantification of quercetin (A) and kaempferol (B) in kale juices fermented at 37 ℃. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juices fermented with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively. Quercetin and kaempferol contents were measured by HPLC analysis. Results are expressed as means ± SD (n = 3). PKJ, pasteurized kale juice before fermentation. The statistical analysis was performed using the independent T-test compared to PKJ (*p<0.001).
Figure 2.
Quantification of quercetin (A) and kaempferol (B) in kale juices fermented at 37 ℃. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juices fermented with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively. Quercetin and kaempferol contents were measured by HPLC analysis. Results are expressed as means ± SD (n = 3). PKJ, pasteurized kale juice before fermentation. The statistical analysis was performed using the independent T-test compared to PKJ (*p<0.001).
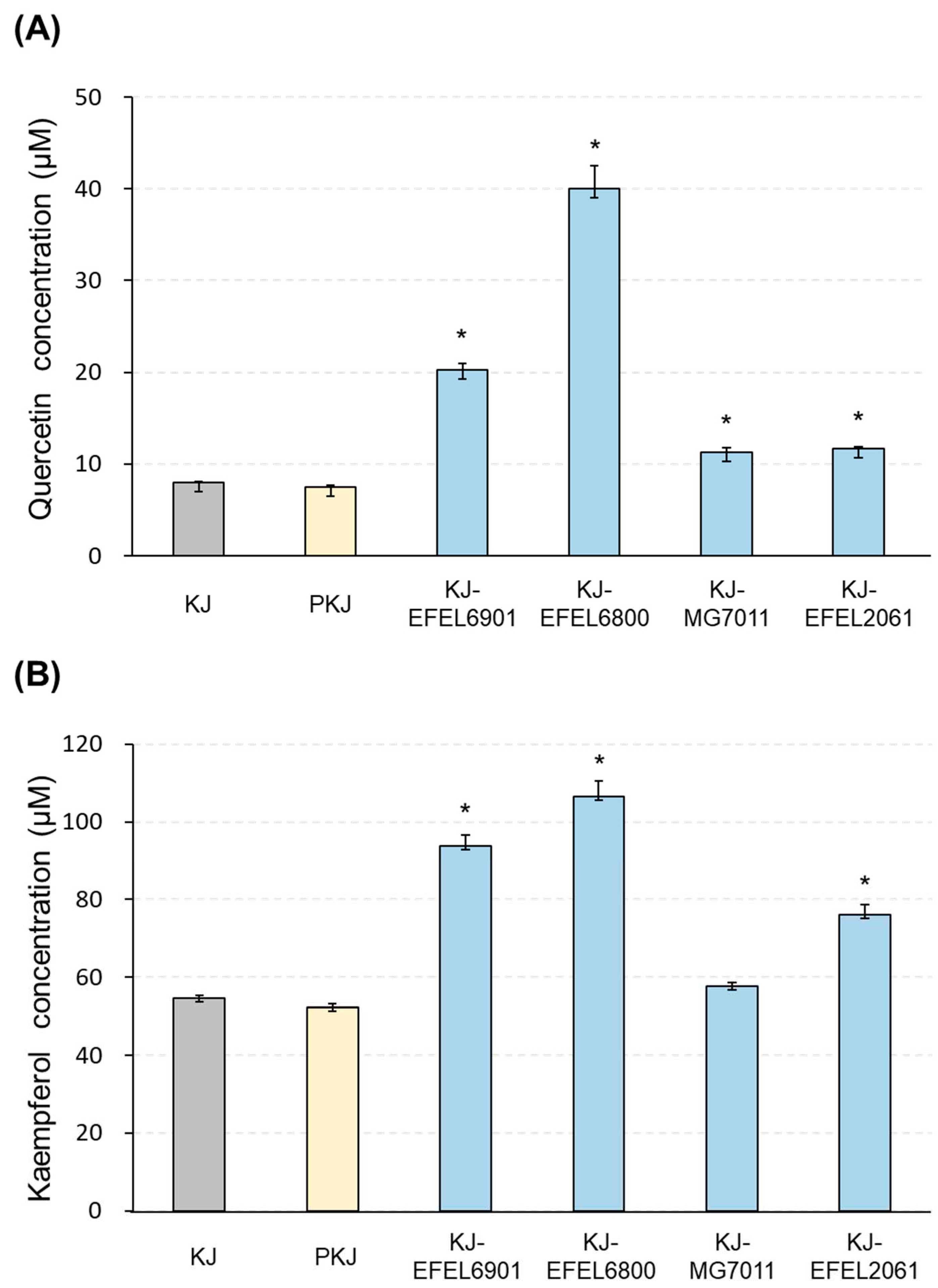
Figure 3.
Changes in metabolite concentrations of kale juices fermented at 37 ℃. Metabolites were analyzed by 1H-NMR. PKJ, pasteurized kale juice before fermentation. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juices fermented with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively. Results are expressed as means ± standard deviations (n=3). The statistical analysis was performed using the independent T-test compared to PKJ (*p<0.05). Different letters indicate a significant difference in the same group at p<0.05 according to Tukey’s multiple range test.
Figure 3.
Changes in metabolite concentrations of kale juices fermented at 37 ℃. Metabolites were analyzed by 1H-NMR. PKJ, pasteurized kale juice before fermentation. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juices fermented with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively. Results are expressed as means ± standard deviations (n=3). The statistical analysis was performed using the independent T-test compared to PKJ (*p<0.05). Different letters indicate a significant difference in the same group at p<0.05 according to Tukey’s multiple range test.
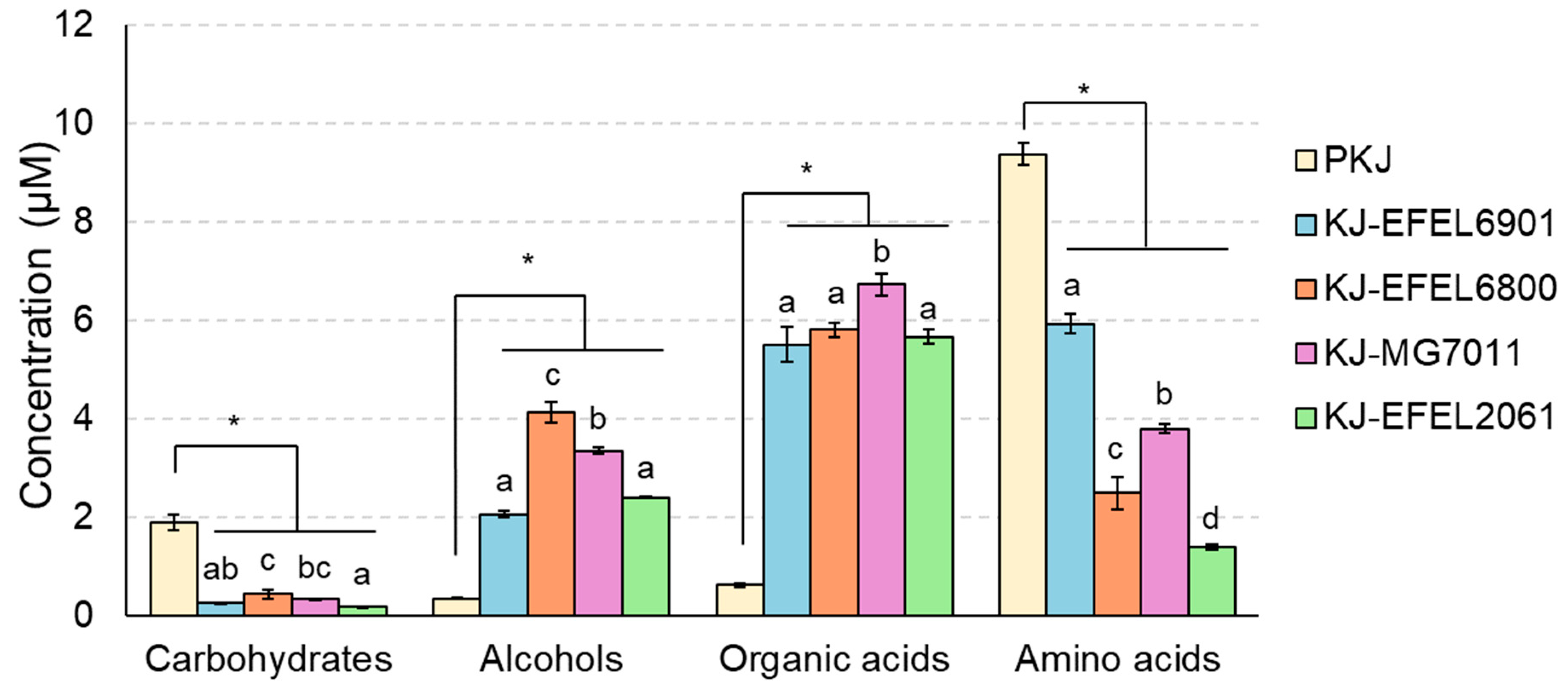
Figure 4.
Effects of fermented kale juice on cell viability of RAW 264.7 cells. RAW 264.7 cells (2×105 cells/well) were treated with 6.25, 12.5, 25, 50, and 100 μg/mL of fermented kale juice powder with 1 μg/mL LPS for 24 h, and cell viability was determined by the MTT assay. Dimethylsulfoxide was used as a vehicle. H2O2 was used as a negative control. LPS, lipopolysaccharide; PKJ, pasteurized kale juice before fermentation. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juices fermented at 37 ℃ for 72 h with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively. Data are the mean ± SD (n = 3). The statistical analysis was performed using the independent T-test compared to LPS (*p<0.05).
Figure 4.
Effects of fermented kale juice on cell viability of RAW 264.7 cells. RAW 264.7 cells (2×105 cells/well) were treated with 6.25, 12.5, 25, 50, and 100 μg/mL of fermented kale juice powder with 1 μg/mL LPS for 24 h, and cell viability was determined by the MTT assay. Dimethylsulfoxide was used as a vehicle. H2O2 was used as a negative control. LPS, lipopolysaccharide; PKJ, pasteurized kale juice before fermentation. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juices fermented at 37 ℃ for 72 h with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively. Data are the mean ± SD (n = 3). The statistical analysis was performed using the independent T-test compared to LPS (*p<0.05).
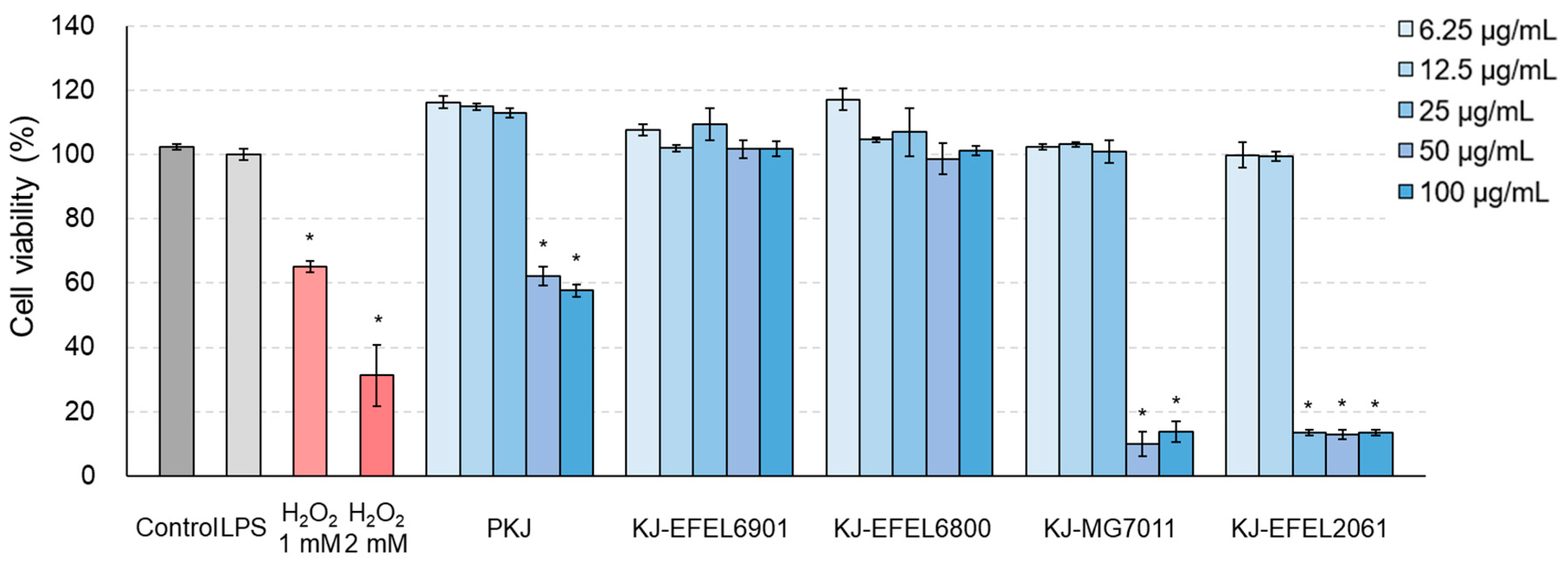
Figure 5.
DPPH free radical scavenging activity (A) and ABTS+ free radical scavenging activity (B) of fermented kale juices. KJ, kale juice; PKJ, pasteurized kale juice before fermentation. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juices fermented at 37 ℃ for 72 h with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively. Different letters indicate a significant difference at p<0.05 according to Tukey’s multiple range test.
Figure 5.
DPPH free radical scavenging activity (A) and ABTS+ free radical scavenging activity (B) of fermented kale juices. KJ, kale juice; PKJ, pasteurized kale juice before fermentation. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juices fermented at 37 ℃ for 72 h with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively. Different letters indicate a significant difference at p<0.05 according to Tukey’s multiple range test.
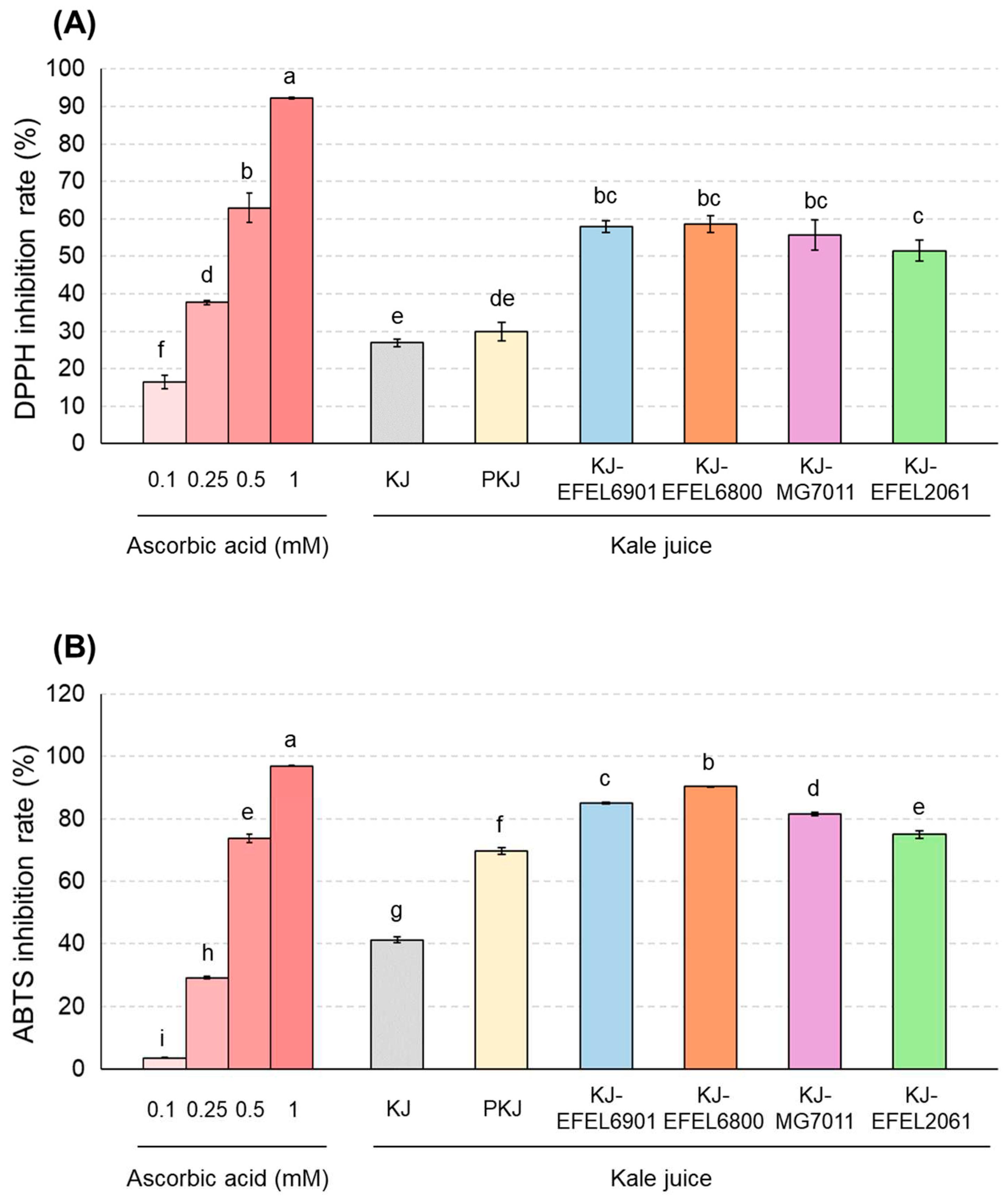
Figure 6.
Effects of fermented kale juice on the production of NO in LPS-induced RAW 264.7 cells. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juices fermented at 37 ℃ for 72 h with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively. RAW 264.7 cells were incubated with 6.25, 12.5, 25, 50, and 100 μg/mL of fermented kale juice powder with 1 μg/mL LPS for 24 h. Dimethylsulfoxide was used as a vehicle. NO was measured according to the Griess reaction. LPS, lipopolysaccharide; MA, methyl arginine (positive control); PKJ, pasteurized kale juice before fermentation. Data are the mean ± SD (n = 3). The statistical analysis was performed using the independent T-test compared to LPS (* p < 0.05, ** p < 0.01, *** p < 0.001).
Figure 6.
Effects of fermented kale juice on the production of NO in LPS-induced RAW 264.7 cells. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juices fermented at 37 ℃ for 72 h with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively. RAW 264.7 cells were incubated with 6.25, 12.5, 25, 50, and 100 μg/mL of fermented kale juice powder with 1 μg/mL LPS for 24 h. Dimethylsulfoxide was used as a vehicle. NO was measured according to the Griess reaction. LPS, lipopolysaccharide; MA, methyl arginine (positive control); PKJ, pasteurized kale juice before fermentation. Data are the mean ± SD (n = 3). The statistical analysis was performed using the independent T-test compared to LPS (* p < 0.05, ** p < 0.01, *** p < 0.001).
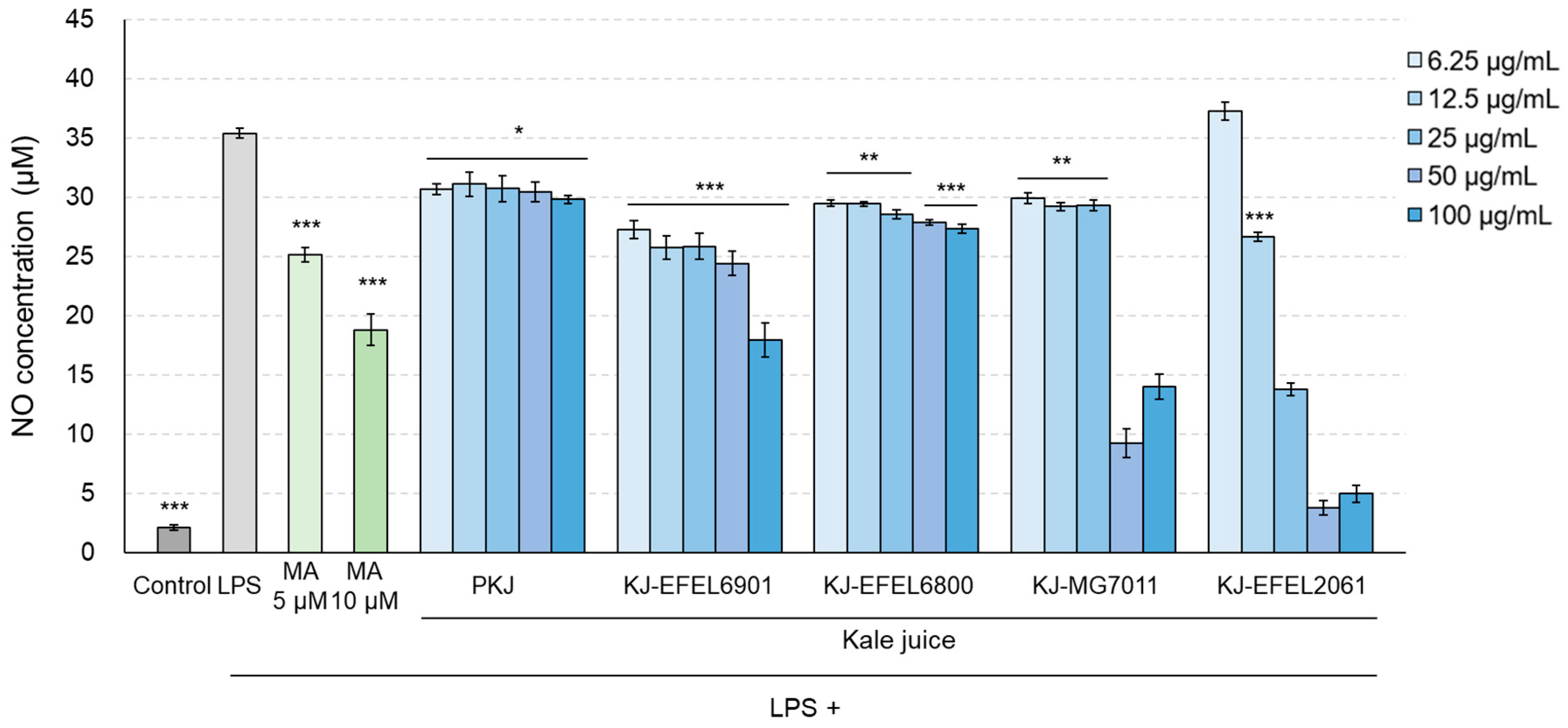
Figure 7.
Effects of fermented kale juice on mRNA expression of iNOS (A), COX-2 (B), IL-6 (C), IL-1ß (D), and IL-10 (E) in LPS-induced RAW 264.7 cells. RAW 264.7 cells were incubated with 6.25 and 12.5 μg/mL of fermented kale juice powder with 1 μg/mL LPS for 24 h. Dimethylsulfoxide was used as a vehicle. The mRNA expression levels of iNOS and COX-2 were determined by real-time PCR. LPS, lipopolysaccharide; PKJ, pasteurized kale juice before fermentation. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juices fermented at 37 ℃ for 72 h with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively. Data are the mean ± SD (n = 3). The statistical analysis was performed using the independent T-test compared to 6 μg/mL of PKJ (#p<0.05, ##p<0.01) or 12.5 μg/mL of PKJ (* p < 0.05, ** p < 0.01). Different letters indicate a significant difference at p<0.05 according to Tukey’s multiple range test.
Figure 7.
Effects of fermented kale juice on mRNA expression of iNOS (A), COX-2 (B), IL-6 (C), IL-1ß (D), and IL-10 (E) in LPS-induced RAW 264.7 cells. RAW 264.7 cells were incubated with 6.25 and 12.5 μg/mL of fermented kale juice powder with 1 μg/mL LPS for 24 h. Dimethylsulfoxide was used as a vehicle. The mRNA expression levels of iNOS and COX-2 were determined by real-time PCR. LPS, lipopolysaccharide; PKJ, pasteurized kale juice before fermentation. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juices fermented at 37 ℃ for 72 h with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively. Data are the mean ± SD (n = 3). The statistical analysis was performed using the independent T-test compared to 6 μg/mL of PKJ (#p<0.05, ##p<0.01) or 12.5 μg/mL of PKJ (* p < 0.05, ** p < 0.01). Different letters indicate a significant difference at p<0.05 according to Tukey’s multiple range test.
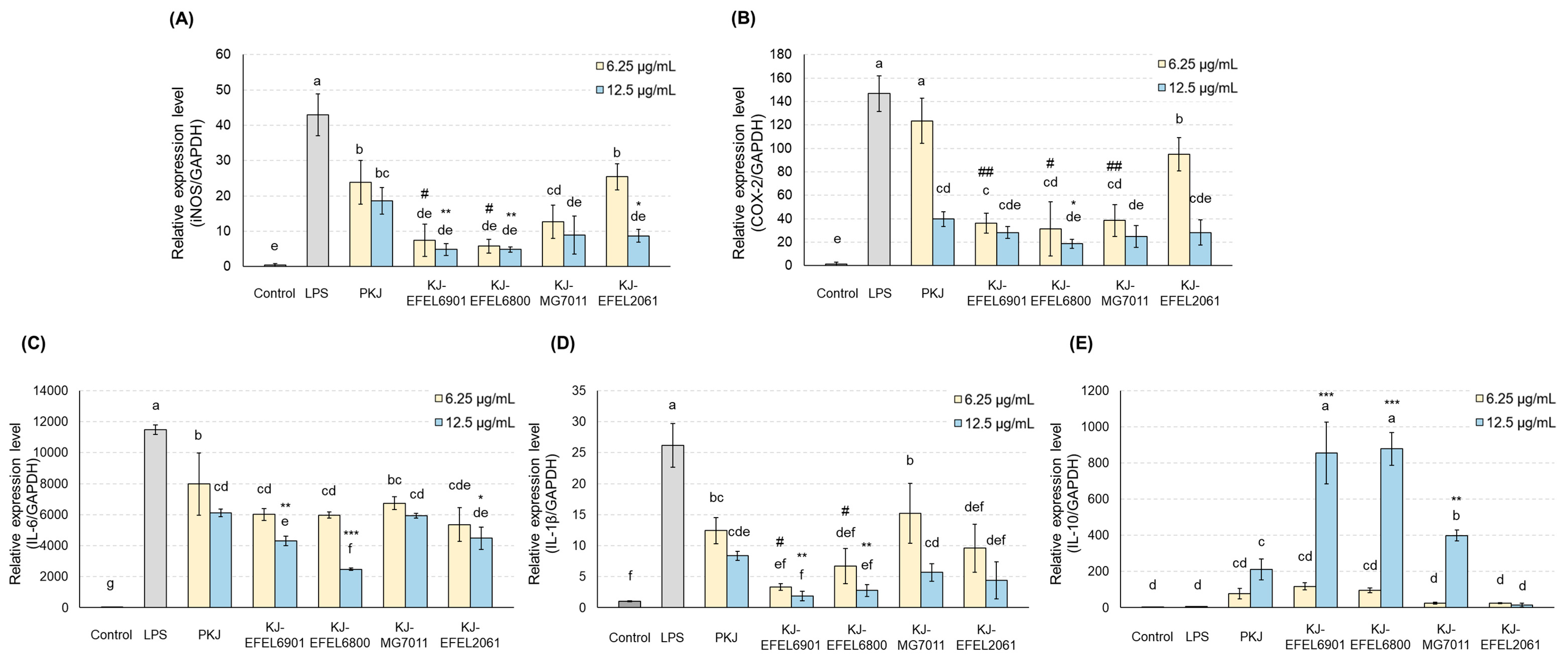

Table 1.
List of strains used in this study.
| Species | Collection | Abbreviation | Culture condition |
|---|---|---|---|
| Limosilactobacillus reuteri EFEL6901 | KACC 81105BP | EFEL6901 | 37 ℃, MRS |
| L. fermentum EFEL6800 | KACC 81106BP | EFEL6800 | 37 ℃, MRS |
| L. fermentum MG7011 | KACC 81147BP | MG7011 | 37 ℃, MRS |
| Leuconostoc citreum EFEL2061 | KACC 92070P | EFEL2601 | 37 ℃, MRS |
| Lactiplantibacillus plantarum WCFS1 | ATCC BAA-793 | WCFS1 | 37 ℃, MRS |
| Lacticaseibacillus rhamnosus GG | KCTC 5033 | LGG | 37 ℃, MRS |
Table 2.
Primer sequences for mouse-specific PCR.
| Gene | Forward primer (5’-3’) | Reverse primer (5’-3’) |
|---|---|---|
| GAPDH | TTGTCTCCTGCGACTTCAACA | GCTGTAGCCGTATTCATTGTCATA |
| iNOS | ACCATGGAGCATCCCAAGTA | CCATGTACCAACCATTGAAGG |
| COX-2 | AGCATTCATTCCTCTACATAAGC | GTAACAACACTCACATATTCATACAT |
| TNF-α | ATGATCCGCGACGTGGAA | ACCGCCTGGAGTTCTGGA |
| IL-1β | GTTGACGGACCCCAAAAGAT | CACACACCAGCAGGTTATCA |
| IL-10 | GGACAACATACTGCTAACCGACTC | AAAATCACTCTTCACCTGCTCCAC |
Table 3.
Colormetric properties of fermented kale juice.
| Sample | L* | a* | b* | ΔE* |
|---|---|---|---|---|
| Kale juice | 29.95±0.02 | -6.56±0.02 | 10.91±0.02 | - |
| PKJ | 34.61±0.00 a | -6.80±0.01 e | 15.14±0.03a | 6.30±0.04 |
| KJ-EFEL6901 | 34.02±0.01b | -0.01±0.01a | 13.73±0.02a | 6.80±0.02a |
| KJ-EFEL6800 | 34.59±0.03a | -1.36±0.01d | 15.25±0.09a | 5.67±0.04b |
| KJ-MG7011 | 34.61±0.04a | -0.76±0.04b | 15.70±1.44a | 6.49±0.47a |
| KJ-EFEL2061 | 34.64±0.02a | -0.85±0.01c | 14.65±0.01a | 6.07±0.04ab |
L* - lightness, a* - redness, b* - yellowness, ΔE - color difference. Kale juice was fermented at 37 ℃ for 72 h. PKJ, pasteurized kale juice before fermentation. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juice fermented with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively. The measurements were made in triplicate are represented as mean values (±SD). Different letters indicate a significant difference at p<0.05 according to Tukey’s multiple range test. For ΔE*, pasteurized kale juice was compared to kale juice, and the other samples were compared to pasteurized kale juice.
Table 4.
Metabolite concentrations in fermented kale juice analyzed by 1H-NMR.
| Group | Metabolites | PKJ | KJ-EFEL6901 | KJ-EFEL6800 | KJ-MG7011 | KJ-EFEL2061 | |||
|---|---|---|---|---|---|---|---|---|---|
| Carbohydrates | Fructose | 0.65±0.12 | 0.08±0.01c* | 0.20±0.03a* | 0.14±0.00b* | 0.04±0.00d* | |||
| Glucose | 0.90±0.04 | 0.06±0.01* | 0.06±0.01* | 0.05±0.00* | 0.04±0.00* | ||||
| Mannose | 0.16±0.05 | 0.09±0.01* | 0.13±0.04 | 0.09±0.00* | 0.07±0.00* | ||||
| Sucrose | 0.14±0.01 | 0.02±0.00* | 0.05±0.04* | 0.05±0.00* | 0.02±0.00* | ||||
| Alcohols | Mannitol | 0.10±0.01 | 1.23±0.03a* | 1.14±0.05ab* | 1.10±0.02b* | 0.98±0.02c* | |||
| Ethanol | 0.24±0.01 | 1.74±0.06c* | 2.98±0.24a* | 2.25±0.05b* | 1.42±0.01c* | ||||
| Organic acids | Acetate | 0.19±0.01 | 3.24±0.33* | 3.54±0.11b* | 2.63±0.13b* | 1.76±0.25b* | |||
| Succinate | 0.09±0.01 | 0.85±0.04b* | 0.30±0.01c* | 1.30±0.17a* | 1.13±0.03 a* | ||||
| Lactate | 0.09±0.01 | 1.69±0.21a* | 1.86±0.03a* | 0.97±0.01b* | 0.49±0.02c* | ||||
| Propionate | 0.03±0.01 | 0.54±0.00bc* | 0.71±0.06b* | 0.63±0.04bc* | 0.84±0.03a* | ||||
| Pyruvate | 0.07±0.01 | 0.08±0.04 | 0.07±0.01 | 0.06±0.01* | 0.04±0.00* | ||||
| Butyrate | 0.04±0.00 | 0.30±0.01b* | 0.18±0.01b* | 0.23±0.40b* | 0.19±0.01a* | ||||
| Citrate | 0.12±0.02 | 0.02±0.01ab* | 0.04±0.01a* | 0.02±0.00ab* | 0.01±0.00b* | ||||
| Amino acids | Aspartate | 1.40±0.04 | 1.08±0.08a* | 0.04±0.01b* | 0.03±0.01b* | 0.06±0.01b* | |||
| Glutamate | 1.13±0.11 | 1.35±0.07a* | 0.33±0.02b* | 0.27±0.01b* | 0.06±0.00c* | ||||
| Cysteine | 0.06±0.01 | 0.17±0.01a* | 0.12±0.03b* | 0.12±0.00b* | 0.07±0.00bc | ||||
| Glycine | 0.08±0.03 | 0.13±0.01ab* | 0.16±0.02a* | 0.18±0.02a* | 0.06±0.00b | ||||
| Histidine | 0.01±0.00 | 0.01±0.01 | 0.01±0.00 | 0.00±0.00 | 0.01±0.00 | ||||
| Alanine | 0.91±0.03 | 0.58±0.01a* | 0.62±0.13a* | 0.48±0.03a* | 0.09±0.01b* | ||||
| Serine | 1.18±0.21 | 0.09±0.01b* | 0.12±0.01a* | 0.12±0.01ab* | 0.05±0.01c* | ||||
| Threonine | 0.30±0.03 | 0.02±0.00b* | 0.08±0.04b* | 0.18±0.03a* | 0.03±0.00b* | ||||
| Arginine | 0.78±0.04 | 0.05±0.01ab* | 0.03±0.01bc* | 0.02±0.00c* | 0.07±0.00a* | ||||
| Proline | 0.44±0.15 | 0.37±0.10 | 0.26±0.03 | 0.30±0.04 | 0.17±0.00 | ||||
| Tyrosine | 0.19±0.01 | 0.15±0.00b* | 0.03±0.00d* | 0.19±0.01a* | 0.06±0.00c* | ||||
| Valine | 0.53±0.21 | 0.47±0.08a* | 0.07±0.01b* | 0.40±0.01a* | 0.20±0.01ab* | ||||
| Methionine | 0.08±0.01 | 0.06±0.01 | 0.02±0.01a* | 0.01±0.00a* | 0.02±0.00a* | ||||
| Isoleucine | 0.43±0.05 | 0.29±0.02a* | 0.05±0.04b* | 0.11±0.01b* | 0.05±0.01b* | ||||
| Leucine | 0.48±0.02 | 0.31±0.07* | 0.14±0.00* | 0.16±0.00* | 0.10±0.01* | ||||
| Phenylalanine | 0.56±0.05 | 0.16±0.01b* | 0.05±0.03c* | 0.38±0.01a* | 0.10±0.01bc* | ||||
| Tryptophan | 0.23±0.02 | 0.21±0.01a* | 0.07±0.00c* | 0.14±0.01b* | 0.07±0.00c* | ||||
| Lysine | 0.63±0.03 | 0.43±0.05b* | 0.30±0.02c* | 0.70±0.01a | 0.09±0.00d* | ||||
PKJ, pasteurized kale juice before fermentation. KJ-EFEL6901, KJ-EFEL6800, KJ-MG7011, and KJ-EFEL2061 corresponded to the kale juice fermented with Limosilactobacillus reuteri EFEL6901, L. fermentum EFEL6800, L. fermentum MG7011, and Leuconostoc citreum EFEL2061, respectively, at 37 ℃ for 72 hours. Results are expressed as means ± standard deviations (n=3). Different letters indicate a significant difference at p<0.05 according to Tukey’s multiple range test. The statistical analysis was performed using the independent T-test compared to the PKJ (PKJ; *p<0.05).
Table 5.
Genetic analysis results of EFEL6901 and EFEL6800 revealing the presence of glycoside hydrolase or glucosidase enzymes.
Table 5.
Genetic analysis results of EFEL6901 and EFEL6800 revealing the presence of glycoside hydrolase or glucosidase enzymes.
| Related enzyme | EFEL6901 | EFEL6800 | ||
|---|---|---|---|---|
| Gene annotation | Protein ID | Gene annotation | Protein ID | |
| Glycoside hydrolase | Glycoside hydrolase family 13 protein | WP_003668994.1 | Glycoside hydrolase family 13 protein | WP_023466491.1 |
| Glycoside hydrolase family 65 protein | WP_003669571.1 | Glycoside hydrolase family 65 protein | WP_023465690.1 | |
| Glycoside hydrolase family 70 protein | WP_229392148.1 | |||
| Glycoside hydrolase family 73 protein | WP_003674860.1WP_003669289.1 | Glycoside hydrolase family 73 protein | WP_023466547.1WP_003681382.1WP_004562972.1 | |
| Glycoside hydrolase family 2 TIM barrel-domain containing protein | WP_003667314.1 | Glycoside hydrolase family 2 TIM barrel-domain containing protein | WP_282347715.1WP_282348494.1 | |
| Glycoside hydrolase family 3 N-terminal domain-containing protein | WP_075667436.1WP_240824081.1 | |||
| Family 78 glycoside hydrolase catalytic domain | WP_282348481.1 | |||
| Glucosidase | Alpha-glucosidase | WP_003669620.1 | Alpha-glucosidase | WP_023466314.1 |
| Glycosyl hydrolase 53 family protein | WP_003668597.1 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



