
Submitted:
01 September 2023
Posted:
01 September 2023
You are already at the latest version
Abstract
Cardiovascular disease (CVD) is one of the leading causes of death in Puerto Rico, where clopidogrel is commonly prescribed to prevent ischemic events. Genetic contributors to both a poor clopidogrel response and the severity of CVD have been identified mainly in Europeans. However, the non-random enrichment of single nucleotide polymorphisms (SNPs) associated with clopidogrel resistance within risk loci linked to underlying CVDs, and the role of admixture, has yet to be tested. This study aimed to assess possible interaction between genetic biomarkers linked to CVDs and those associated with clopidogrel resistance among admixed Caribbean Hispanics. We identified 50 SNPs significantly associated with CVDs in previous genome-wide association studies (GWAS). These SNPs were combined with another ten SNPs related to clopidogrel resistance in Caribbean Hispanics. We developed Python scripts to determine whether SNPs related to CVDs are in close proximity to those associated with clopidogrel response. Average and individual local ancestry (LAI) within each locus were inferred, and 60 random SNPs with their corresponding LAI were generated for enrichment estimation purposes. Our results showed no CVD-linked SNPs in close proximity to those associated with clopidogrel response among Caribbean Hispanics. Consequently, no genetic loci with a dual predictive role for risk of CVD severity and clopidogrel resistance were found in this population. Native American ancestry was the most enriched within risk loci linked to CVDs in this population. Non-random enrichment of disease susceptibility loci with drug response SNPs is a new frontier in Precision Medicine that needs further attention.
Keywords:
Clopidogrel
; Caribbean Hispanics
; Biomarkers
; Cardiovascular Diseases
; Pharmacogenomics
; Ancestry
1. Introduction
Caribbean Hispanics are underrepresented in genetic studies, limiting knowledge of genetic risk factors for both CVD severity and poor response to clopidogrel [1]. Most of the characterized SNPs associated with CVDs such as acute coronary syndrome (ACS), coronary artery disease (CAD), and peripheral artery disease (PAD), have been identified in patients of predominantly European ancestry [2,3]. Cardiovascular risk factors to inform disease severity (e.g., type-2 diabetes, left ventricular ejection fraction (LVEF)< 30%, adjoined length of stent(s) ≥30mm, smoking) are also predictive biomarkers of poor response to clopidogrel [57,58,59,60]. However, there is no evidence of significant associations between these markers at the genomic level. We have hypothesized that genetic loci linked to these risk factors and response to clopidogrel are inherited together or associated with each other.
Unlike Mendelian-type monogenic conditions with large effect sizes, complex traits affected by many genes with moderate to small effect sizes each (e.g., CVDs) are less likely to transfer across populations. Given their diverse evolutionary histories, resulting in differential allele frequency distributions and varying effect sizes across ethnicities of distinct ancestry and degree of admixture, the role of unique genomic architectures on transferability is critical and must be accounted for. Indeed, the poor transferability of findings from European-centered genomic associations to Hispanics influences how well true causal variants are captured by SNPs identified in Europeans and ultimately exacerbate health inequities. To mitigate the impact of population-specific attributes and the resulting poor transferability of genomic results on health equity, it is imperative to take ancestry into account and expand current studies to admixed populations where the trait of interest is highly prevalent. Caribbean Hispanics are an admixed population with high genetic heterogeneity that gives rise to phenotypic diversity. By accounting for the local ancestry (LA) effect we may also uncover novel variants associated with clopidogrel response. Therefore, inferring genetic ancestry while assessing the relationship between ACS/CAD- and PAD-associated SNPs and those associated with clopidogrel resistance is important for reducing health disparities in this underrepresented population.
Bioinformatic computational tools have been developed to facilitate the study of global ancestry composition in individuals. This has permitted us to understand the evolutionary and migration patterns of populations [4,5]. Since SNP frequency can change from one population to another, it could serve as a genetic signature or landmark in population genetic studies. In addition, advancement in local ancestry inference (LAI) tools has continued, helping elucidate disease risk and optimal drug therapy in populations [6,7]. LAI is described as the ancestry in a particular location of the chromosome of an individual. LAI works by matching a segment of the reference population to a segment of the chromosome of an admixed individual and assigning the respective ancestry. In the case of Caribbean Hispanics, the reference populations include European, African, and Native American.
LAI results will also depend on the applied method. Various methods for LAI have been developed, including the Local Ancestry in adMixed Populations (LAMP-LD) [8], the RFMix [9], the Efficient Local Ancestry Inference (ELAI) [10], and MOSAIC [11] software. LAMP-LD is a software package for the inference of locus-specific ancestry in recently admixed populations [8]. RFMix is a powerful discriminative modeling approach for rapid and robust local-ancestry inference [9]. ELAI performs local ancestry inference for admixed individuals [10]; whereas MOSAIC infers segments of ancestry and characterizes admixture events involving an arbitrary number of genetically distinct groups putting together [11]. However, a study showed that the most accurate method is RFMix [12]. Also, a study comparing LAMP-LD and RFMix showed that RFMix had a shorter run time [13].
Studies inferring local ancestry by using RFMix have been carried out in Hispanics, but not in a cohort of Caribbean Hispanic patients with ACS, CAD, and PAD receiving clopidogrel treatment [14,15]. In this study we ascertained ancestry enrichments at relevant CV loci, as well as non-random associations between clopidogrel-related genomic markers and those linked to the severity of CV diseases, in admixed Caribbean Hispanics.
2. Materials and Methods
2.1. Study Population
Caribbean Hispanic patients, self-identified as Cuban, Dominican, or Puerto Rican, on treatment with clopidogrel (75mg/day) were included in the study. The participants were recruited at five medical facilities across the Commonwealth of Puerto Rico: The Cardiovascular Center of Puerto Rico and the Caribbean, San Francisco Hospital, Dr. Hilton Franqui Clinics, Pavia Hospital, and UPR-Hospital Dr. Federico Trilla. All participants met inclusion/exclusion criteria (Table S1) and signed an informed consent approved by the Institutional Review Board (IRB-protocol number A4070417). Relevant demographical and clinical data were collected along with a blood sample in two 3.0 ml 3.2% sodium citrate tubes. Medication adherence was assessed by self-report and record reviews, using the highly validated and reliable four-item Morisky Medication Adherence Scale (MMAS-4), as described elsewhere [56].
2.2. Identification of Variants
A literature assessment was performed to identify 50 SNPs that were related to CVDs such as ACS, CAD, and PAD. No date restrictions were employed. We used the NHGRI-EBI GWAS Catalog [16] (https://www.ebi.ac.uk/gwas/) as search engine for SNPs associated to each of the three CVDs. First, we searched using the key term “acute coronary syndrome” and we arranged the SNPs in order of p-value significance, followed by the selection of the 20 most significant SNPs. Then we repeated the same procedure using the key term “coronary artery disease”. Some SNPs for CAD were already included for ACS, therefore, we evaluated and selected the SNP with the highest p-value for the corresponding condition. For PAD, we used the key term “peripheral artery disease” and we selected only the first 10 most significant SNPs, since the minority of the patients are on treatment with clopidogrel for PAD. The SNPs positions were acquired as the id for further analysis steps. The positions were based on GRCh37 as the reference genome build. In addition, 10 SNPs corresponding to the top 10 risk loci signals from a GWAS of high on-treatment platelet reactivity (HTPR) performed in Caribbean Hispanics patients on clopidogrel were included (unpublished data). We evaluated if any of these SNPs associated with either CVDs or clopidogrel resistance could also be expression quantitative trait loci (eQTLs). To this purpose, we employed the Gtex Portal (https://gtexportal.org/home/), an online tool used to evaluate possible eQTLs from our variants of interest.
2.3. Local Ancestry Inference
Genotyped data were used to estimate haplotypes using Shapeit v2.17 software. RFMix software v1 was performed to infer local ancestry by using the code available at: https://github.com/indraniel/rfmix/blob/master/RunRFMix.py. For reference population, we used genotyped data from 107 individuals of the Iberian population in Spain (IBS) to represent European ancestry, 61 individuals of Yoruba in Ibadan, Nigeria (YRI) to represent African ancestry, and 103 Continental Native American individuals from the America population of Human Genome Diversity Project (HGDP) to represent Native American (NA) ancestry. The IBS and YRI population came from the NHGRI Sample Repository for Human Genetic Research of the Coriell Institute (https://www.internationalgenome.org/data-portal/population). The America population were defined as Native American, and the samples came from Surui and Karitiana in Brazil, Piapoco and Colombian in Colombia, and Maya and Pima in Mexico (https://www.internationalgenome.org/data-portal/data-collection/hgdp).
2.4. Python Scripts
Since computational tools have allowed us to analyze high-throughput data, we assessed the non-random enrichment in the association between clopidogrel resistance and the severity of underlying ACS, CAD, and PAD by determining close proximity (±0.5 cM) between SNPs. Three python scripts were developed fittingly for the purpose of each objective: 1) to estimate the local ancestral proportions by variant, 2) to determine if variants of interest are found in close proximity (± 0.5 cM), and 3) to acquire random variants from an existing dataset. The scripts can be found at https://github.com/mariangelimonero/Non-random_enrichment. The open-source Python v3.10.2 application was used for this purpose.
2.4.1. Ancestry Search
Ancestry data from each individual was obtained by RFMix software (https://github.com/indraniel/rfmix) [13]. The python script was developed to determine the ancestry within each locus genotyped or imputed across the whole genome of the 510 samples and available in the dataset. As input file we used the information about the location files in cM and the forward-backward files containing ancestry information for each variant. The script consisted of opening the location files and forward-backward files of each chromosome and appending them to a list. This information was used to gather ancestry data for the variants and was stored as “all_chr.anc” file extension for each of the samples. A text file with the samples to be included in the analysis was set on the script. Ancestries were matched with the given sample names by appending them to a list. The rs-number and the chromosome number were obtained and appended to their respective lists. The two lists were then transformed into a data frame, respectively, and then concatenated into a single data frame. Also, the list containing the information about position and cM was transformed into a data frame and concatenated with the data frame of rs-number and chromosome creating a single data frame. A column with the calculation of the average ancestry by variant was performed. Finally, the previously created data frames were concatenated, creating a file with the columns rs-number, chromosome, position, cM, average ancestry, and subsequent samples.
2.4.2. Random Search
A python script was developed to randomly find and select another 60 variants from the dataset. A text file was provided as input, with the positions of rs-numbers of interest that we did not want to include as random variants. The ancestry file developed in the previous ancestry script was used to provide the information about ancestry from the 60 random signals. The script was developed to search for n random signals. It is designed to append all rs-numbers in a list, excluding the ones of interest. The function shuffle was applied to the list. The output was given with the quantity of variants desired and the columns rsnumber, chromosome, position, cM, average ancestry, and subsequent samples.
2.4.3. Bracket Search
A python script was developed to find two variants of interest in close proximity (i.e., located within ± 0.5 cM of each other). A list of variants of interest and ancestry file developed in previous ancestry search step were used as input file. The variants of interest and the ancestry file were added to individual lists. The intersection function was used to determine the variants of interest that were available in the ancestry list. If the intersection was true, then the matching variants were written in the output file. The output file was then converted into a data frame, and we determined if the variants were + 0.5 or -0.5 cM far apart from each other. Repeated matching pair variants were eliminated. This created a csv file with only the position and cM of the variants in proximity. The data from ancestry and matching variants were converted into a csv file with columns of rsnumber, chromosome, position, cM, and subsequent samples.
3. Results
3.1. Data Cohort
This is a secondary analysis of a dataset from a multicenter clinical study protocol (NCT03419325) [56]. To this purpose, a subcohort of 510 Caribbean Hispanic patients on treatment with clopidogrel and with full genetics and clinical data available was used. Patients were diagnosed with CVDs such as ACS, CAD, and/or PAD. All diagnoses were obtained from the patient’s electronic medical records at each participating medical facility using the corresponding ICD-9/10-CM codes, which were subsequently confirmed by the study cardiologists. Patients receiving either a 600mg or 300 mg loading dose of clopidogrel followed by a maintenance dose of 75 mg daily or started with a 75mg daily maintenance dose (i.e., alone or as a component of dual anti-platelet therapy (DAPT) in mainly post-PCI patients with ACS or stable CAD with elective stenting) were enrolled in the study between January 2018 and June 2020 from five different medical facilities across the island. Enrollment criteria (Table S1) were minimal with the goal of gathering an ‘all comers’ population representative of real-world clinical practice. We observed high adherence scores (>85%) during the study period. Participants with a significant lack of adherence to therapy were removed from subsequent analyses. The cohort was genotyped for approximately 1.4 million of SNPs with the InfiniumTM Multi-Ethnic AMR/AFR Genotyping Array (MEGA) BeadChip by Illumina (San Diego, CA).
Table 1 describes the baseline characteristics of participants (n=510). The average age of all participants in this study was 68 years-old, with 55% identified as male. Overall, 78.3% aged 60 years old or above and 18.6% were within the 45-59 years-old range. Largely, most were middle-aged males, with high prevalence of conventional risk factors (i.e., overweight (28.4 kg/m2); hypertension (83.9%); hypercholesterolemia/ dyslipidemias (71.9%) and type-2 diabetes (54.8%), among others). Furthermore, 20% of participants were on proton pump inhibitors (PPI, mainly pantoprazole); whereas statins and calcium channel blockers (CCB) were prescribed in 79.2% and 26.8% of patients, respectively. Patients who were taking aspirin administered as part of DAPT represented 63.3% of the total cohort. In 75.9% of participants, clopidogrel was given for stable CAD/ACS indication. No differences in baseline characteristics (e.g., comorbidities) existed among patients after stratifying by carrier status or indication.
3.2. Expression Quantitative Trait Loci (eQTLs) SNPs
The intronic variant rs55791371 (Table 2 and Table 3 and Table S10) is an eQTL that resides within the gene encoding the BRG1 protein. The rs9349379 SNP is another identified eQTL variant located at an intronic position in the phosphatase and actin regulator 1 (PHACTR1) gene (Table 3 and Table S4). Moreover, the rs10455872, rs118039278, and rs140570886 SNPs are also eQTL variants associated with thrombus formation (Table 2 and Table 3 and Table S4). In addition, the rs55791371 variant (Table 2 and Table 3 and Table S10) is an eQTL for the SLC44A2 gene that encodes a choline transporter protein.
3.3. Non-Random Enrichment of SNPs
After a literature review, we identified 20 SNPs associated with ACS, 20 SNPs associated with CAD, and 10 SNPs associated with PAD (Table 2). Although some SNPs were repeated in two different CVDs, we kept the SNP for the condition with the strongest evidence. Additionally, we obtained the ten top signals from a GWAS of clopidogrel resistance among Caribbean Hispanics patients in our laboratory (Table 2). The total of these 60 SNPs were analyzed with the python script developed for bracket search.
To determine if the SNPs associated with poor clopidogrel response in our cohort (i.e., top 10 GWAS hits) are occurring close to or within the same genetic loci of SNPs linked to CVDs, our python script was used. Not a single SNP linked to the severity of CVDs was found either upstream (+0.5 cM) or downstream (-0.5 cM) the corresponding loci of any SNP associated with clopidogrel resistance. Additional functional information of these 60 SNPs was assessed and reported in Tables S2–S12 (Supplemental material).
3.4. Local Ancestry Inferences (LAI)
The developed script for ancestry search was able to identify the locus-specific enrichment of ancestries related to European (Iberians), African (Yoruba), and Native Americans contributions within each of the selected SNPs in each individual. However, we were only able to infer the ancestry for 10 out of the 50 SNPs associated with CVDs (Table 3; Figure 1). It is because available databases from reference parental population lacked the necessary information on these other 40 SNPs genotyped in our cohort of Caribbean Hispanics, which is required by RFMix software to infer LAIs. As control group, we also estimated the ancestry proportions at 60 random SNPs for the purpose of ancestry enrichment comparison. Our results showed ≥50% enrichment of Native American, African, and European ancestries in 25, 20 and 2 of these 60 variants, respectively (Table 4; Figure 2).
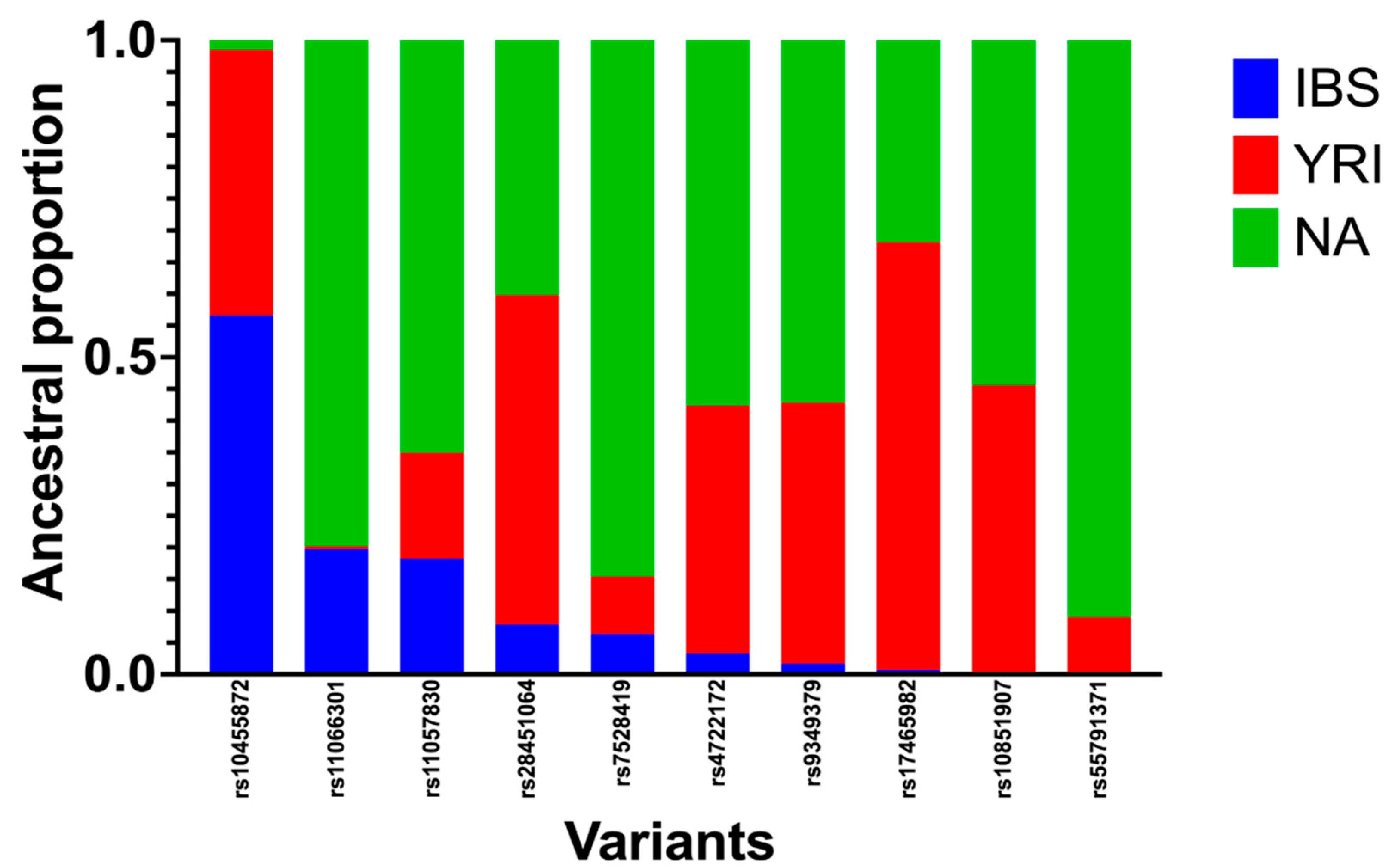
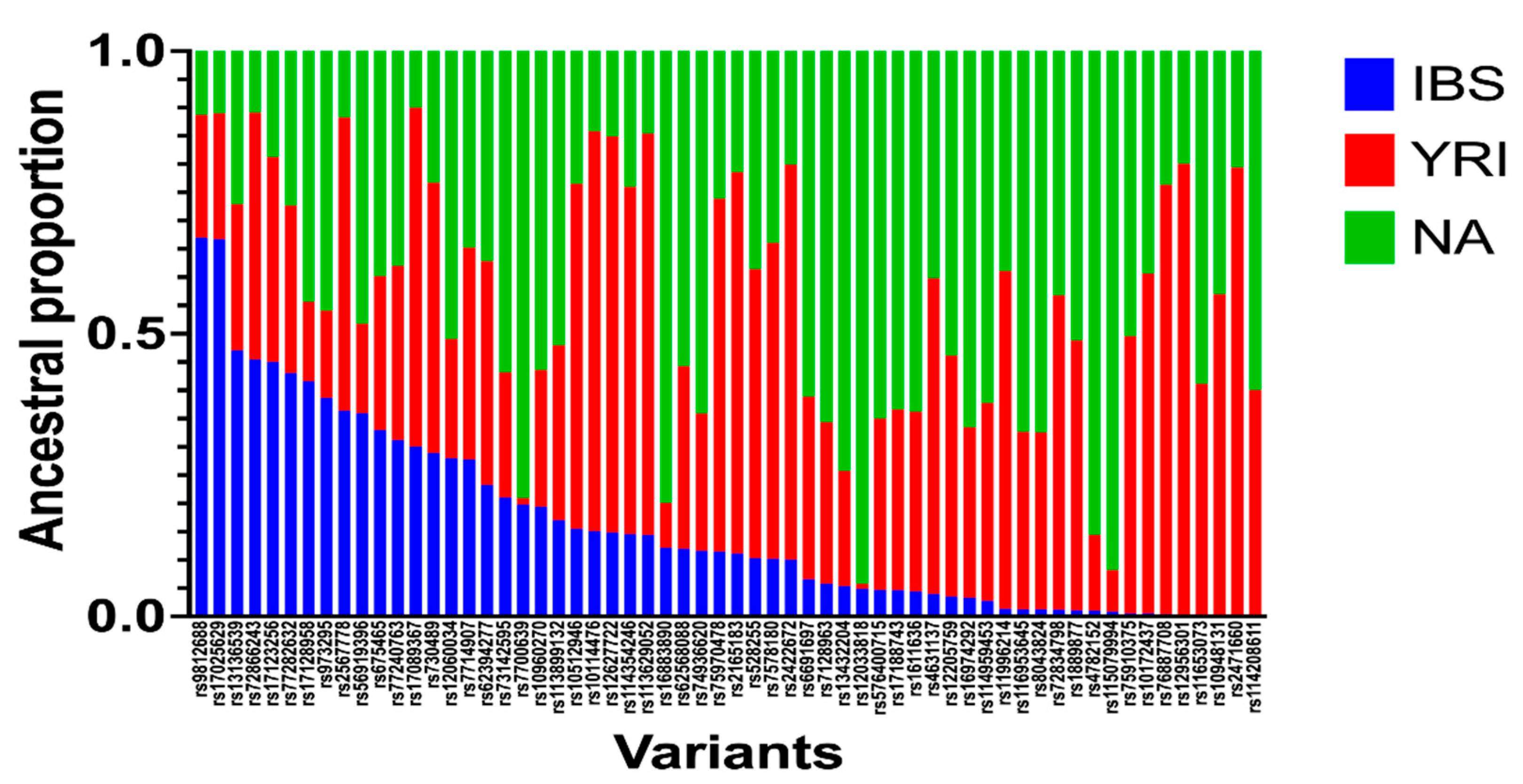
Figure 1.
Ancestry proportions around the 10 SNPs associated with CVDs (ACS, CAD, and PAD) using the python script for ancestry search. Genotyping data of 510 Caribbean Hispanics were obtained by Infinium Multi-Ethnic AMR/AFR Genotyping Array (MEGA) BeadChip by Illumina®. RFMix software was used to estimate locus specific ancestries by using reference populations from Iberians, Yoruba, and Native Americans. A python script was developed to infer ancestry proportions from the list of SNP of interest.
Figure 1.
Ancestry proportions around the 10 SNPs associated with CVDs (ACS, CAD, and PAD) using the python script for ancestry search. Genotyping data of 510 Caribbean Hispanics were obtained by Infinium Multi-Ethnic AMR/AFR Genotyping Array (MEGA) BeadChip by Illumina®. RFMix software was used to estimate locus specific ancestries by using reference populations from Iberians, Yoruba, and Native Americans. A python script was developed to infer ancestry proportions from the list of SNP of interest.
Figure 2.
Ancestral proportions of 60 SNPs generated from ancestry search python script. A total of 510 Caribbean Hispanic patients were genotyped by the Illumina MEGA chip microarray. Genotyped data was phased using Shapeit software. Local ancestry inference was obtained by RFMix software. A python script was designed to identify ancestral proportions for available variants. Ancestries are represented as Iberian population (IBS), Yoruba population (YRI), and Native population from America (NA).
Figure 2.
Ancestral proportions of 60 SNPs generated from ancestry search python script. A total of 510 Caribbean Hispanic patients were genotyped by the Illumina MEGA chip microarray. Genotyped data was phased using Shapeit software. Local ancestry inference was obtained by RFMix software. A python script was designed to identify ancestral proportions for available variants. Ancestries are represented as Iberian population (IBS), Yoruba population (YRI), and Native population from America (NA).
4. Discussion
Understanding the genetic architecture of an admixed population is key to the prevention, diagnosis, and treatment of CVDs. Previous GWAS have identified SNPs associated with the risk of CVDs [18,39,40]. Although these studies are informative enough about potential risk markers, admixed populations are neglected because most of the participants are of European descent.
The top ten signals with the lowest p-values in our GWAS of HTPR in CV patients on clopidogrel were used for testing their non-random enrichment with risk variants of CVDs. Seven of these signals (Table 2) are located closed to a cluster of cytochromes P450 genes on chromosome 10q23.33, which is where the highly polymorphic CYP2C19 gene is mapped to (reference GRCh38.p14 assembly: 94,762,681 to 94,855,547). The bio-activation of clopidogrel is mediated by the CYP2C19 metabolic pathway and, therefore, CYP2C19 genotypes are predictive of clopidogrel resistance [55]. Particularly, the CYP2C19*2 variant (rs4244285) has been associated with clopidogrel resistance in previous studies in Europeans [41,42]. However, rs4244285 was not among the top 10 signals in our study with Caribbean Hispanics. The rs7916697 SNP, an ATHO7 intronic variant encoding a transcription factor for the regulation of retinal ganglion cells and optic nerve, was among the 10 most significant variants selected instead. We also selected the intronic variants rs4646743, rs4745950, and rs1900005, located within the region encoding a lncRNA called LINC02640; while rs1900003 and rs1900002 variants located ~2kb upstream the LINC02640 were also considered. In addition, the intronic rs3796692 variant that is related to the ncRNA called LOC105377582 was included. Finally, the single nucleotide variant rs4021557, located ~7kb upstream the MAP2K3 gene involved in the regulation of the MAPK kinase pathway and MAPK kinase-mediated signaling cascades, was considered.
Reports of numerous SNPs associated with CVDs are found in the available literature [32,43,44]. We have identified 50 of these genetic variants significantly associated with the severity of CVDs (e.g., ACS, CAD, and/or PAD). These variants (Table 2) were associated with inflammatory pathways, cholesterol transport, and regulation and signaling cascades in previous GWAS [32,43,44]. Although most of these CVDs-linked variants are occurring within intronic regions, they can either enhance or reduce gene expressions [45]. The eQTLs are genetic loci that alter gene expression and trait variation quantitatively [46]. These eQTL SNPs can change a quantitative trait through different mechanisms such as epigenetic modifications, transcription, pre-mRNA processing, and post-transcriptional processing (mRNA stability & translation rate) [46].
BRG1 is part of the SWI/SNF complex, with ATPase and helicase activity, that participates in chromatin remodeling. Although the underlying pathway related to BRG1 is not yet fully elucidated, a recent human in vitro study demonstrated that BRG1 is highly expressed in hypertrophic cardiomyopathy and can alter myosin expression [47]. The rs9349379 SNP in the PHACTR1 gene is an eQTL that has been found to be associated with CAD risk. PHACTR1 protein regulates remodeling of the heart after myocardial infarction (MI), increasing its expression and the total protein levels after the event [48]. MI is commonly caused by coronary artery occlusion due to a thrombus; therefore, eQTL variants associated with thrombus formation (e.g., rs10455872, rs118039278, and rs140570886 in Table 2 and Table 3 and Table S4) are expected to be identified as signals of GWAS significance. These three SNPs have been correlated to one another for affecting the trait expression for lipoprotein A, a low-density lipoprotein (LDL) with apolipoprotein A, and have been associated with CAD and PAD [30,31,49,50,51]. On the other hand, the relevance of rs55791371 (Table 2 and Table 3 and Table S10) as an eQTL for the SLC44A2 gene encoding the choline transporter was demonstrated in studies in vitro. Authors suggested that ATP will be available for platelet aggregation by regulating choline transport into the mitochondria through SLC44A2, thus resulting in a higher risk for thrombus formation [52].
In this study, we found that relevant genetic risk loci associated with the severity of CVDs in Europeans are not enriched with SNPs linked to clopidogrel resistance among Caribbean Hispanic patients. Therefore, we did not identify a possible genetic marker with a dual role of predicting the severity of the CVDs and the response to clopidogrel in this target population. A possible explanation would be the limited number of SNPs to consider from reference parental populations, narrowing the pool of possible variants to be studied. Databases containing genetic information from worldwide populations need to expand their variant catalog diversity, especially to account for relevant SNPs within the Native American populations.
Due to the emergence of novel bioinformatic tools, the genetically-inferred ancestral composition of Caribbean Hispanics can now be better understood. To determine if the enrichment was different from that observed in Caribbean Hispanics, the resulting ancestral enrichments at each of the 60 random SNPs were compared to those within the variant loci previously found to be associated with CVDs in Europeans. We found that the ancestral composition in the vicinity of the ten CVDs-linked variants tested was similar to that around the 60 random variants. We speculate that certain locus-specific ancestry enrichment (e.g., African) at these risk loci can explain to some extent a possible breakup of overlapping between SNPs related to clopidogrel resistance and the severity of CVDs or lack thereof in Caribbean Hispanics. The occurrence of novel haplotype blocks as a results of altered linkage disequilibrium (LD) patterns and the substantial admixture in Caribbean Hispanics might interfere with expected associations between both types of SNPs.
Our study is unique in that we carried out a targeted approach using bioinformatic tools and newly developed scripts to identify such ancestry enrichments and non-random associations in a quite heterogeneous and admixed population of Caribbean Hispanic patients. Significant enrichments of African and Native American ancestry, but not of clopidogrel-associated genomic markers, were found within these 10 CVD-specific risk loci. We firmly believe that ancestry enrichments observed at these 10 disease-specific relevant loci linked to the severity of CVDs will help predict the prevalence of significantly different inter-ethnic phenotypes even for highly admixed populations like Caribbean Hispanics. Consequently, such information could be of help for future admixture mapping studies in this population in order to refine discovery of novel predictive signals by stratifying individuals regarding their ancestral enrichment at the locus of interest. That is, genetic loci showing high enrichment of ancestry related to the ethnic group with the highest prevalence of the surveyed phenotype will be targeted for more in-depth scrutiny, as long as the phenotype shows differential risk by ancestry.
Recently admixed populations, like Caribbean Hispanics, have a genome that is reminiscent of a mosaic, with sections inherited from Africa and others inherited from Europe or native populations. Consequently, genetically-inferred global and local ancestry is an important consideration in drug response specifically in admixed population at the gene level. Hispanic population is highly admixed, but their ancestral proportions differ across different ethno-geographic locations due to historical context and other determinants. This admixture highlights challenges for the current CPIC guideline for clopidogrel selection, which does not discriminate between patients with different ethnic-geographic backgrounds. For example, should any amount of enrichment with European ancestry or certain CVD-linked risk loci require an individual to obtain CYP2C19 genotyping prior to benefiting from PGx-guided antiplatelet therapy? Therefore, our results could also help to delineate optimal strategies for the individualization of antiplatelet therapies based on the ancestral origin and non-random CVD-related enrichment of patients at risk.
Study limitations include a relatively modest sample size in the surveyed cohort when compared to the usual larger number of participants in similar studies within other continental populations (i.e., 2,750 Europeans in the International Clopidogrel Pharmacogenomics Consortium (ICPC) study) [53]. However, our patients represent a more diverse group than that of the ICPC study. Ancestral proportions, both local and global, are inherently biased due to the European-centered nature of reference genomes used for their inferences [54]. Although of minimal impact, collection biases due to the potential for patient noncompliance and the exclusion of stroke patients are additional limitations of this analysis. Finally, our work did not include data regarding the use of alternative antiplatelet drugs (e.g., prasugrel, ticagrelor), which may be preferred in many clinical scenarios.
5. Conclusions
In conclusion, even though we did not find a non-random enrichment of CVD-associated variants within sites nearby those associated with clopidogrel resistance, there are still possible novel ethnic-specific variants to be discovered and added to reference panels for future inquiries. Our data brings insightful information about locus specific ancestry in Caribbean Hispanics, being Native American ancestry the most enriched within risk loci for variants linked to CVDs. Non-random enrichment of disease susceptibility loci with SNPs related to drug responsiveness needs further attention to improve experimental designs of studies seeking adoption of the Precision Medicine paradigm.
The use of valid bioinformatic tools to identify novel or overlooked eQTLs linked to critical biological pathways in this admixed population may further unravel relevant biomarkers of CVD severity, as well as clopidogrel resistance. We strongly believe that the integration of information about ancestry and genetic risk variants can expedite the identification of underlying disease-related mechanisms in Caribbean Hispanics. Since most of the published studies identifying SNPs linked to CVDs (i.e., ACS, CAD, and PAD) were conducted in mainly Europeans, the assessment of a potential non-random enrichment of SNPs associated with clopidogrel resistance within risk loci linked to CVD severity in a cohort of Caribbean Hispanics, and the comparison of LAIs at 10 CVD risk loci and 60 additional randomly selected SNPs will also contribute to further address the existing health disparity among Caribbean Hispanics.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1. Inclusion and Exclusion Criteria. Table S2. eQTL’s from SNP’s on chromosome 1 associated to cardiovascular conditions. Table S3. eQTL’s from SNP’s on chromosome 2 associated to cardiovascular conditions. Table S4. eQTL’s from SNP’s on chromosome 6 associated to cardiovascular conditions. Table S5. eQTL’s from SNP’s on chromosome 7 associated to cardiovascular conditions. Table S6. eQTL’s from SNP’s on chromosome 10 associated to cardiovascular conditions and resistance to clopidogrel in Caribbean Hispanics. Table S7. eQTL’s from SNP’s on chromosome 11 associated to cardiovascular conditions. Table S8. eQTL’s from SNP’s on chromosome 12 associated to cardiovascular conditions. Table S9. eQTL’s from SNP’s on chromosome 15 associated to cardiovascular conditions. Table S10. eQTL’s from SNP’s on chromosome 19 associated to cardiovascular conditions. Table S11. eQTL’s from SNP’s on chromosome 20 associated to cardiovascular conditions. Table S12. eQTL’s from SNP’s on chromosome 21 associated to cardiovascular conditions.
Author Contributions
MMP wrote the first draft of the manuscript, collected and performed data analyses; RFM developed the python scripts; KCC and PG performed computational data analyses; JD developed the original research idea, edited and prepared the final version of the manuscript; IR and ARL provided the infrastructure, supervised, and participated in editing the manuscript. All authors have read and approved this manuscript.
Funding
This study was financially supported by grant 2 U54 MD007600-31 from the National Institute on Minority Health and Health Disparities (NIMHD) of the National Institutes of Health and by the National Institute of General Medical Sciences (NIGMS)-Research Training Initiative for Student Enhancement (RISE) Program grant R25 GM061838.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and was approved by the Institutional Review Board (IRB-protocol number A4070417) from the University of Puerto Rico, Medical Sciences Campus. All subjects participated voluntarily and signed an informed consent.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study. Written informed consent has been obtained from the patient(s) to publish this paper.
Data Availability Statement
The data used as reference population are Surui and Karitiana in Brazil, Piapoco and Colombian in Colombia, and Maya and Pima in Mexico found at https://www.internationalgenome.org/data-portal/data-collection/hgdp. IBS and YRI population are from the NHGRI Sample Repository for Human Genetic Research of the Coriell Institute found at https://www.internationalgenome.org/data-portal/population. The datasets from Caribbean Hispanics analyzed during the current study are available from the corresponding author upon request.
Acknowledgments
We thank Ms. Brenda Nieves for her assistance and support of this research project.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A
- List of abbreviations:
- ACS = acute coronary syndrome
- CAD = coronary artery disease
- CV = cardiovascular
- CHR = chromosome
- cM = centimorgan
- DAPT = dual anti-platelet therapy
- eQTL = expression quantitative trait loci
- GWAS = genome-wide association study
- Hct = hematocrit
- HGDP = Human Genome Diversity Project
- HTPR = high on-treatment platelet reactivity
- IBS = Iberian
- LA = local ancestry
- LAI = local ancestry inference
- LDL = low-density lipoprotein
- MEGA = Multi-Ethnic AMR/AFR Genotyping Array
- MI = myocardial infarction
- NA = Native American
- PAD = peripheral artery disease
- PHACTR1= phosphatase and actin regulator 1
- POS = position
- SNP = single nucleotide polymorphism
- TIA = transient ischemic attack
- YRI = Yoruba
References
- Sirugo, G.; Williams, S.M.; Tishkoff, S.A. The Missing Diversity in Human Genetic Studies. Cell 2019, 177, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Evangelou, E.; Program, T.M.V.; Warren, H.R.; Mosen-Ansorena, D.; Mifsud, B.; Pazoki, R.; Gao, H.; Ntritsos, G.; Dimou, N.; Cabrera, C.P.; et al. Genetic analysis of over 1 million people identifies 535 new loci associated with blood pressure traits. Nat. Genet. 2018, 50, 1412–1425. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.D.; Yanek, L.R.; Chen, M.-H.; Faraday, N.; Larson, M.G.; Tofler, G.; Lin, S.J.; Kraja, A.T.; A Province, M.; Yang, Q.; et al. Genome-wide meta-analyses identifies seven loci associated with platelet aggregation in response to agonists. Nat. Genet. 2010, 42, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Via M, Gignoux CR, Roth LA, Fejerman L, Galanter J, Choudhry S, et al. History Shaped the Geographic Distribution of GenomicAdmixture on the Island of Puerto Rico. PLoS One. 2011;6(1).
- Hernández, C.L.; Pita, G.; Cavadas, B.; López, S.; Sánchez-Martínez, L.J.; Dugoujon, J.-M.; Novelletto, A.; Cuesta, P.; Pereira, L.; Calderón, R. Human Genomic Diversity Where the Mediterranean Joins the Atlantic. Mol. Biol. Evol. 2019, 37, 1041–1055. [Google Scholar] [CrossRef]
- Shriner, D.; Adeyemo, A.; Ramos, E.; Chen, G.; Rotimi, C.N. Mapping of disease-associated variants in admixed populations. Genome Biol. 2011, 12, 223–223. [Google Scholar] [CrossRef]
- Dien et al. 2013. Case-control admixture mapping in Latino populations enriches for known asthma-associated genes. Bone. 2008;23(1):1–7. [CrossRef]
- Baran, Y.; Pasaniuc, B.; Sankararaman, S.; Torgerson, D.G.; Gignoux, C.; Eng, C.; Rodriguez-Cintron, W.; Chapela, R.; Ford, J.G.; Avila, P.C.; et al. Fast and accurate inference of local ancestry in Latino populations. Bioinformatics 2012, 28, 1359–1367. [Google Scholar] [CrossRef]
- Uren, C.; Hoal, E.G.; Möller, M. Putting RFMix and ADMIXTURE to the test in a complex admixed population. BMC Genet. 2020, 21, 1–8. [Google Scholar] [CrossRef]
- Guan, Y. Detecting Structure of Haplotypes and Local Ancestry. Genetics 2014, 196, 625–642. [Google Scholar] [CrossRef]
- Salter-Townshend M, Myers S. Fine-scale inference of ancestry segments without prior knowledge of admixing groups. Genetics. 2019 Jul 1;212(3):869–89. [CrossRef]
- Schubert, R.; Andaleon, A.; Wheeler, H.E. Comparing local ancestry inference models in populations of two- and three-way admixture. PeerJ 2020, 8, e10090. [Google Scholar] [CrossRef] [PubMed]
- Maples, B.K.; Gravel, S.; Kenny, E.E.; Bustamante, C.D. RFMix: A Discriminative Modeling Approach for Rapid and Robust Local-Ancestry Inference. Am. J. Hum. Genet. 2013, 93, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Browning SR, Grinde K, Plantinga A, Gogarten SM, Stilp AM, Kaplan RC, et al. Local ancestry inference in a large US-based Hispanic/Latino study: Hispanic community health study/study of Latinos (HCHS/SOL). G3: Genes, Genomes, Genetics. 2016;6(6):1525–34. [CrossRef]
- Sofer T, Baier LJ, Browning SR, Thornton TA, Talavera GA, Wassertheil-Smoller S, et al. Admixture mapping in the Hispanic Community Health Study/Study of Latinos reveals regions of genetic associations with blood pressure traits. PLoS One. 2017;12(11). [CrossRef]
- Buniello, A.; MacArthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E.; et al. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef]
- Yeo, A.; Li, L.; Warren, L.; Aponte, J.; Fraser, D.; King, K.; Johansson, K.; Barnes, A.; MacPhee, C.; Davies, R.; et al. Pharmacogenetic meta-analysis of baseline risk factors, pharmacodynamic, efficacy and tolerability endpoints from two large global cardiovascular outcomes trials for darapladib. PLOS ONE 2017, 12, e0182115–e0182115. [Google Scholar] [CrossRef]
- Åkerblom, A.; Eriksson, N.; Wallentin, L.; Siegbahn, A.; Barratt, B.J.; Becker, R.C.; Budaj, A.; Himmelmann, A.; Husted, S.; Storey, R.F.; et al. Polymorphism of the cystatin C gene in patients with acute coronary syndromes: Results from the PLATelet inhibition and patient Outcomes study. Am. Hear. J. 2014, 168, 96–102. [Google Scholar] [CrossRef]
- Varenhorst, C.; Eriksson, N.; Johansson. ; Barratt, B.J.; Hagström, E.; Åkerblom, A.; Syvänen, A.-C.; Becker, R.C.; James, S.K.; Katus, H.A.; et al. Effect of genetic variations on ticagrelor plasma levels and clinical outcomes. Eur. Hear. J. 2015, 36, 1901–1912. [Google Scholar] [CrossRef]
- Klarin D, Zhu QM, Emdin CA, Chaffin M, Horner S, McMillan BJ, et al. Genetic analysis in UK Biobank links insulin resistance and transendothelial migration pathways to coronary artery disease. Nat Genet. 2017 Sep 1;49(9):1392–7. [CrossRef]
- van der Harst, P.; Verweij, N. Identification of 64 Novel Genetic Loci Provides an Expanded View on the Genetic Architecture of Coronary Artery Disease. Circ. Res. 2018, 122, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Mehta, N.N.; Y, D.; C, G.; Z, L.; L, W.; B, L.; F, H.; T, Z.; Y, W.; X, W.; et al. A Genome-Wide Association Study in Europeans and South Asians Identifies 5 New Loci for Coronary Artery Disease. Circ. Cardiovasc. Genet. 2011, 4, 465–466. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.-P.; Wu, H.; Chen, J.-Y.; Li, X.-X.; Lin, H.-M.; Zhang, B.; Zhang, Z.-W.; Ma, D.-L.; Sun, S.; Li, H.-P.; et al. Genomewide Association Study Identifies Novel Genetic Loci That Modify Antiplatelet Effects and Pharmacokinetics of Clopidogrel. Clin. Pharmacol. Ther. 2017, 101, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Saw, J.; Yang, M.-L.; Trinder, M.; Tcheandjieu, C.; Xu, C.; Starovoytov, A.; Birt, I.; Mathis, M.R.; Hunker, K.L.; Schmidt, E.M.; et al. Chromosome 1q21.2 and additional loci influence risk of spontaneous coronary artery dissection and myocardial infarction. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Koyama, S.; Ito, K.; Terao, C.; Akiyama, M.; Horikoshi, M.; Momozawa, Y.; Matsunaga, H.; Ieki, H.; Ozaki, K.; Onouchi, Y.; et al. Population-specific and trans-ancestry genome-wide analyses identify distinct and shared genetic risk loci for coronary artery disease. Nat. Genet. 2020, 52, 1–9. [Google Scholar] [CrossRef]
- Hager, J.; Kamatani, Y.; Cazier, J.-B.; Youhanna, S.; Ghassibe-Sabbagh, M.; Platt, D.E.; Abchee, A.B.; Romanos, J.; Khazen, G.; Othman, R.; et al. Genome-Wide Association Study in a Lebanese Cohort Confirms PHACTR1 as a Major Determinant of Coronary Artery Stenosis. PLOS ONE 2012, 7, e38663. [Google Scholar] [CrossRef]
- Lu, X.; The Coronary ARtery DIsease Genome-Wide Replication And Meta-Analysis (CARDIoGRAM) Consortium; Wang, L.; Chen, S.; He, L.; Yang, X.; Shi, Y.; Cheng, J.; Zhang, L.; Gu, C.C.; et al. Genome-wide association study in Han Chinese identifies four new susceptibility loci for coronary artery disease. Nat. Genet. 2012, 44, 890–894. [CrossRef]
- Nelson, C.P.; Goel, A.; Butterworth, A.S.; Kanoni, S.; Webb, T.R.; Marouli, E.; Zeng, L.; Ntalla, I.; Lai, F.Y.; Hopewell, J.C.; et al. Association analyses based on false discovery rate implicate new loci for coronary artery disease. Nat. Genet. 2017, 49, 1385–1391. [Google Scholar] [CrossRef]
- Nikpay M, Goel A, Won HH, Hall LM. A comprehensive 1000 Genomes-based genome-wide association meta-analysis of coronary artery disease. Nat Genet [Internet]. 2015;47(10):1121–30. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4589895/pdf/emss-64693.pdf.
- Dichgans M, Rainer M, König IR. Shared Genetic Susceptibility to Ischemic Stroke and Coronary Artery Disease: A Genome-Wide Analysis of Common Variants. In: Stroke. Lippincott Williams and Wilkins; 2013.
- Temprano-Sagrera, G.; Sitlani, C.M.; Bone, W.P.; Martin-Bornez, M.; Voight, B.F.; Morrison, A.C.; Damrauer, S.M.; de Vries, P.S.; Smith, N.L.; Sabater-Lleal, M.; et al. Multi-phenotype analyses of hemostatic traits with cardiovascular events reveal novel genetic associations. J. Thromb. Haemost. 2022, 20, 1331–1349. [Google Scholar] [CrossRef] [PubMed]
- Vujkovic, M.; Keaton, J.M.; Lynch, J.A.; Miller, D.R.; Zhou, J.; Tcheandjieu, C.; Huffman, J.E.; Assimes, T.L.; Lorenz, K.; Zhu, X.; et al. Discovery of 318 new risk loci for type 2 diabetes and related vascular outcomes among 1.4 million participants in a multi-ancestry meta-analysis. Nat. Genet. 2020, 52, 680–691. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Nielsen, J.B.; Fritsche, L.G.; Dey, R.; Gabrielsen, M.E.; Wolford, B.N.; LeFaive, J.; VandeHaar, P.; Gagliano, S.A.; Gifford, A.; et al. Efficiently controlling for case-control imbalance and sample relatedness in large-scale genetic association studies. Nat. Genet. 2018, 50, 1335–1341. [Google Scholar] [CrossRef]
- Matsunaga, H.; Ito, K.; Akiyama, M.; Takahashi, A.; Koyama, S.; Nomura, S.; Ieki, H.; Ozaki, K.; Onouchi, Y.; Sakaue, S.; et al. Transethnic Meta-Analysis of Genome-Wide Association Studies Identifies Three New Loci and Characterizes Population-Specific Differences for Coronary Artery Disease. Circ. Genom. Precis. Med. 2020, 13, e002670. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ma, H.; Zhu, Q.; Zhang, B.; Yan, H.; Li, H.; Meng, J.; Lai, W.; Li, L.; Yu, D.; et al. A genome-wide association study on lipoprotein (a) levels and coronary artery disease severity in a Chinese population. J. Lipid Res. 2019, 60, 1440–1448. [Google Scholar] [CrossRef] [PubMed]
- Schunkert, H.; König, I.R.; Kathiresan, S.; Reilly, M.P.; Assimes, T.L.; Holm, H.; Preuss, M.; Stewart, A.F.; Barbalic, M.; Gieger, C.; et al. Large-scale association analysis identifies 13 new susceptibility loci for coronary artery disease. Nat. Genet. 2011, 43, 333–338. [Google Scholar] [CrossRef]
- van Zuydam NR, Stiby A, Abdalla M, Austin E, Dahlström EH, McLachlan S, et al. Genome-Wide Association Study of Peripheral Artery Disease. Circ Genom Precis Med. 2021 Oct 1;14(5):e002862. [CrossRef]
- Ishigaki, K.; Akiyama, M.; Kanai, M.; Takahashi, A.; Kawakami, E.; Sugishita, H.; Sakaue, S.; Matoba, N.; Low, S.-K.; Okada, Y.; et al. Large-scale genome-wide association study in a Japanese population identifies novel susceptibility loci across different diseases. Nat. Genet. 2020, 52, 669–679. [Google Scholar] [CrossRef]
- Fall, T.; Gustafsson, S.; Orho-Melander, M.; Ingelsson, E. Genome-wide association study of coronary artery disease among individuals with diabetes: the UK Biobank. Diabetologia 2018, 61, 2174–2179. [Google Scholar] [CrossRef]
- Ward-Caviness CK, Neas LM, Blach C, Haynes CS, LaRocque-Abramson K, Grass E, et al. Genetic variants in the bone morphogenic protein gene family modify the association between residential exposure to traffic and peripheral arterial disease. PLoS One. 2016 Apr 1;11(4). [CrossRef]
- Kazui, M.; Nishiya, Y.; Ishizuka, T.; Hagihara, K.; Farid, N.A.; Okazaki, O.; Ikeda, T.; Kurihara, A. Identification of the Human Cytochrome P450 Enzymes Involved in the Two Oxidative Steps in the Bioactivation of Clopidogrel to Its Pharmacologically Active Metabolite. Drug Metab. Dispos. 2009, 38, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Bura A, Villard E, Azizi M, Goyenvalle C, Aiach M, Lechat P, et al. Cytochrome P450 2C19 loss-of-function polymorphism is a major determinant of clopidogrel responsiveness in healthy subjects. Blood. 2006;108(7):2244–7. [CrossRef]
- Yamada Y, Yasukochi Y, Kato K, Oguri M, Horibe H, Fujimaki T, et al. Identification of 26 novel loci that confer susceptibility to early-onset coronary artery disease in a Japanese population. Biomed Rep. 2018 Nov 1;9(5):383–404. [CrossRef]
- Liu X, Xu H, Xu H, Geng Q, Mak WH, Ling F, et al. New genetic variants associated with major adverse cardiovascular events in patients with acute coronary syndromes and treated with clopidogrel and aspirin. Pharmacogenomics Journal. 2021 Dec 1;21(6):664–72. [CrossRef]
- Shaul O. How introns enhance gene expression. Vol. 91, International Journal of Biochemistry and Cell Biology. Elsevier Ltd; 2017. p. 145–55. [CrossRef]
- Pai, A.A.; Pritchard, J.K.; Gilad, Y. The Genetic and Mechanistic Basis for Variation in Gene Regulation. PLOS Genet. 2015, 11, e1004857. [Google Scholar] [CrossRef]
- Scherba, J.C.; Halushka, M.K.; Andersen, N.D.; Maleszewski, J.J.; Landstrom, A.P.; Bursac, N.; Glass, C. BRG1 is a biomarker of hypertrophic cardiomyopathy in human heart specimens. Sci. Rep. 2022, 12, 1–7. [Google Scholar] [CrossRef]
- Kelloniemi, A.; Szabo, Z.; Serpi, R.; Näpänkangas, J.; Ohukainen, P.; Tenhunen, O.; Kaikkonen, L.; Koivisto, E.; Bagyura, Z.; Kerkelä, R.; et al. The Early-Onset Myocardial Infarction Associated PHACTR1 Gene Regulates Skeletal and Cardiac Alpha-Actin Gene Expression. PLOS ONE 2015, 10, e0130502. [Google Scholar] [CrossRef]
- Zhuang, Z.; Yao, M.; Wong, J.Y.Y.; Liu, Z.; Huang, T. Shared genetic etiology and causality between body fat percentage and cardiovascular diseases: a large-scale genome-wide cross-trait analysis. BMC Med. 2021, 19, 1–15. [Google Scholar] [CrossRef]
- Klarin, D.; Program, V.M.V.; Lynch, J.; Aragam, K.; Chaffin, M.; Assimes, T.L.; Huang, J.; Lee, K.M.; Shao, Q.; Huffman, J.E.; et al. Genome-wide association study of peripheral artery disease in the Million Veteran Program. Nat. Med. 2019, 25, 1274–1279. [Google Scholar] [CrossRef]
- Shah S, Henry A, Roselli C, Lin H, Sveinbjörnsson G, Fatemifar G, et al. Genome-wide association and Mendelian randomisation analysis provide insights into the pathogenesis of heart failure. Nat Commun. 2020 Dec 1;11(1). [CrossRef]
- Bennett, J.A.; Mastrangelo, M.A.; Ture, S.K.; Smith, C.O.; Loelius, S.G.; Berg, R.A.; Shi, X.; Burke, R.M.; Spinelli, S.L.; Cameron, S.J.; et al. The choline transporter Slc44a2 controls platelet activation and thrombosis by regulating mitochondrial function. Nat. Commun. 2020, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.S.; Bergmeijer, T.O.; Gong, L.; Reny, J.; Lewis, J.P.; Mitchell, B.D.; Alexopoulos, D.; Aradi, D.; Altman, R.B.; Bliden, K.; et al. Genomewide Association Study of Platelet Reactivity and Cardiovascular Response in Patients Treated With Clopidogrel: A Study by the International Clopidogrel Pharmacogenomics Consortium. Clin. Pharmacol. Ther. 2020, 108, 1067–1077. [Google Scholar] [CrossRef] [PubMed]
- Levy-Sakin, M.; Pastor, S.; Mostovoy, Y.; Li, L.; Leung, A.K.Y.; McCaffrey, J.; Young, E.; Lam, E.T.; Hastie, A.R.; Wong, K.H.Y.; et al. Genome maps across 26 human populations reveal population-specific patterns of structural variation. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Lee, C.R.; Luzum, J.A.; Sangkuhl, K.; Gammal, R.S.; Sabatine, M.S.; Stein, C.M.; Kisor, D.F.; Limdi, N.A.; Lee, Y.M.; Scott, S.A.; et al. Clinical Pharmacogenetics Implementation Consortium Guideline for CYP2C19 Genotype and Clopidogrel Therapy: 2022 Update. Clin. Pharmacol. Ther. 2022, 112, 959–967. [Google Scholar] [CrossRef]
- Duconge, J.; Santiago, E.; Hernandez-Suarez, D.F.; Moneró, M.; López-Reyes, A.; Rosario, M.; Renta, J.Y.; González, P.; Fernández-Morales, L.I.; Vélez-Figueroa, L.A.; et al. Pharmacogenomic polygenic risk score for clopidogrel responsiveness among Caribbean Hispanics: A candidate gene approach. Clin. Transl. Sci. 2021, 14, 2254–2266. [Google Scholar] [CrossRef]
- Hernandez-Suarez, D.F.; Botton, M.R.; Scott, S.; Tomey, M.; Garcia, M.J.; Wiley, J.; Villablanca, P.; Melin, K.; Lopez-Candales, A.; Renta, J.Y.; et al. Pharmacogenetic association study on clopidogrel response in Puerto Rican Hispanics with cardiovascular disease: a novel characterization of a Caribbean population. Pharmacogenomics Pers. Med. 2018, ume 11, 95–106. [Google Scholar] [CrossRef]
- Hernandez-Suarez, D.F.; Scott, S.A.; Tomey, M.I.; Melin, K.; Lopez-Candales, A.; Buckley, C.E.; Duconge, J. Clinical determinants of clopidogrel responsiveness in a heterogeneous cohort of Puerto Rican Hispanics. Ther. Adv. Cardiovasc. Dis. 2017, 11, 235–241. [Google Scholar] [CrossRef]
- Tcheng JE, Lim IH, Srinivasan S, et al. Stent parameters predict major adverse clinical events and the response to platelet glycoprotein IIb/IIIa blockade: findings of the ESPRIT trial. Circ Cardiovasc Interv 2009;2:43–51. [CrossRef]
- Janssen, P.W.A.; Bergmeijer, T.O.; Vos, G.-J.A.; Kelder, J.C.; Qaderdan, K.; Godschalk, T.C.; Breet, N.J.; Deneer, V.H.M.; Hackeng, C.M.; Berg, J.M.T. Tailored P2Y12 inhibitor treatment in patients undergoing non-urgent PCI—the POPular Risk Score study. Eur. J. Clin. Pharmacol. 2019, 75, 1201–1210. [Google Scholar] [CrossRef]

Table 1.
Characteristics of participants in the study cohort.
| Variable | mean | SD | SEM | Min | Max | median | ||
|---|---|---|---|---|---|---|---|---|
| Age (years) | 68.01 | 10.95 | 0.51 | 27.00 | 94.00 | 69.00 | ||
| BMI (kg/m2) | 28.40 | 5.71 | 0.27 | 11.48 | 52.67 | 27.67 | ||
| Variable | n | % | ||||||
| Gender (Males) | 282 | 55.29 | ||||||
| Diabetes Mellitus | 280 | 54.79 | ||||||
| Hypertension | 428 | 83.92 | ||||||
| Dyslipidemias | 367 | 71.96 | ||||||
| Smoking | 69 | 13.53 | ||||||
| MACE¶ | 42 | 13.77 | ||||||
| MI¶ | 19 | 6.23 | ||||||
| Stent Thrombosis¶ | 15 | 4.92 | ||||||
| Deaths¶ | 4 | 1.31 | ||||||
| Bleedings events* | 83 | 16.24 | ||||||
| Aspirin use | 323 | 63.32 | ||||||
| Statins use | 404 | 79.21 | ||||||
| CCB | 137 | 26.86 | ||||||
| PPI | 102 | 19.96 | ||||||
| LVEF ≤40% | 42 | 8.23 | ||||||
| ACS & stable CAD | 387 | 75.88 | ||||||
| Coronary artery stenting# | 191 | 37.45 | ||||||
| PAD | 123 | 24.12 | ||||||
Note. Due to rounding errors, percentages may not equal 100%. ¶MACE reports available only in a subset of 305 patients. *Bleeding events is a combination of major and minor events. #All patients with ACS and some with CAD underwent PCI (i.e., percutaneous coronary interventions) or coronary artery stenting. MI: myocardial infarctions (i.e., STEMI and NSTEMI). CCB: calcium channel blockers. PPI: proton pump inhibitors. BMI means body mass index. PAD stands for peripheral artery disease.
Table 2.
Identified SNPs associated with ACS, CAD, and PAD in previous literature and SNPs related to clopidogrel resistance in Caribbean Hispanics.
Table 2.
Identified SNPs associated with ACS, CAD, and PAD in previous literature and SNPs related to clopidogrel resistance in Caribbean Hispanics.
| Chr | POS | SNP | Relation | Reference |
|---|---|---|---|---|
| 19 | 45412079 | rs7412 | ACS | [17] |
| 6 | 46677098 | rs76863441 | ACS | [17] |
| 1 | 109817590 | rs12740374 | ACS | [17] |
| 20 | 23616469 | rs35610040 | ACS | [18] |
| 7 | 99421085 | rs62471956 | ACS | [19] |
| 6 | 161010118 | rs10455872 | ACS | [17] |
| 15 | 44408401 | rs2733201 | ACS | [17] |
| 15 | 44293137 | rs11638352 | ACS | [17] |
| 6 | 161111700 | rs186696265 | ACS | [17] |
| 7 | 99286639 | rs188845491 | ACS | [19] |
| 7 | 98932759 | rs147642358 | ACS | [19] |
| 7 | 100103523 | rs140607780 | ACS | [19] |
| 6 | 160751531 | rs9295128 | ACS | [17] |
| 7 | 99543627 | rs140104968 | ACS | [19] |
| 14 | 84804488 | rs117714106 | ACS | [17] |
| 7 | 99841354 | rs117038461 | ACS | [19] |
| 15 | 44564692 | rs144972973 | ACS | [17] |
| 1 | 172995643 | rs201052613 | ACS | [17] |
| 12 | 125307053 | rs11057830 | ACS | [17] |
| 9 | 107396924 | rs189889864 | ACS | [17] |
| 6 | 161013013 | rs140570886 | CAD | [20,21] |
| 6 | 12903957 | rs9349379 | CAD | [20,21,22,23,24,25,26,27,28,29] |
| 1 | 109821511 | rs602633 | CAD | [21,30,31,32] |
| 19 | 11202306 | rs6511720 | CAD | [17,21,28,31] |
| 19 | 11188153 | rs55791371 | CAD | [21,25,30] |
| 15 | 79141784 | rs7173743 | CAD | [21,25,30] |
| 21 | 35593827 | rs28451064 | CAD | [20,21,24,28,29,31,33] |
| 2 | 203873743 | rs6728861 | CAD | [21] |
| 2 | 203968973 | rs72934535 | CAD | [21,31,33] |
| 2 | 203893999 | rs115654617 | CAD | [21] |
| 6 | 160911596 | rs147555597 | CAD | [21] |
| 6 | 134209837 | rs2327429 | CAD | [21,25,34] |
| 6 | 161018174 | rs7770628 | CAD | [35] |
| 12 | 111884608 | rs3184504 | CAD | [21,29,30,31,36] |
| 11 | 103660567 | rs974819 | CAD | [21,22] |
| 6 | 134159622 | rs1966248 | CAD | [21] |
| 1 | 222829550 | rs35158675 | CAD | [21,25] |
| 1 | 56966350 | rs17114046 | CAD | [21,22,33] |
| 1 | 222837939 | rs17465982 | CAD | [21] |
| 12 | 111932800 | rs7137828 | CAD | [21,31] |
| 6 | 160985526 | rs118039278 | PAD | [20,37] |
| 9 | 22103183 | rs1537372 | PAD | [20,37] |
| 15 | 78915864 | rs10851907 | PAD | [20,37] |
| 1 | 169519049 | rs6025 | PAD | [20,37] |
| 7 | 19049388 | rs2107595 | PAD | [20,37,38] |
| 1 | 109817192 | rs7528419 | PAD | [20,37] |
| 12 | 112871372 | rs11066301 | PAD | [20,37] |
| 7 | 22786532 | rs4722172 | PAD | [20,37] |
| 10 | 114758349 | rs7903146 | PAD | [20,37] |
| 9 | 136149229 | rs505922 | PAD | [20,37] |
| 10 | 69996292 | rs4746743 | Clopidogrel | unpublished data |
| 10 | 69998055 | rs1900005 | Clopidogrel | unpublished data |
| 10 | 69999026 | rs12098677 | Clopidogrel | unpublished data |
| 21 | 39485558 | rs9980291 | Clopidogrel | unpublished data |
| 10 | 70004551 | rs1900003 | Clopidogrel | unpublished data |
| 10 | 70004552 | rs1900002 | Clopidogrel | unpublished data |
| 10 | 69996455 | rs4745950 | Clopidogrel | unpublished data |
| 4 | 185205210 | rs3796692 | Clopidogrel | unpublished data |
| 17 | 21225519 | rs4021557 | Clopidogrel | unpublished data |
| 10 | 69991853 | rs7916697 | Clopidogrel | unpublished data |
Chr (chromosome); pos (position in GRCh37); SNP (single nucleotide polymorphism). Position is based on human assembly GRCh37/h19 genome reference build by the Genome Reference Consortium.
Table 3.
Ancestry search for SNPs associated with cardiovascular conditions in previous literature.
| rsnumber | chr | POS | cM | IBS | YRI | NA |
|---|---|---|---|---|---|---|
| rs7528419 | 1 | 109817192 | 138.712247 | 0.064 | 0.09053 | 0.84549 |
| rs17465982 | 1 | 222837939 | 246.478996 | 0.0074 | 0.67445 | 0.3182 |
| rs9349379 | 6 | 12903957 | 28.8898785 | 0.0172 | 0.41137 | 0.57144 |
| rs10455872 | 6 | 161010118 | 177.557204 | 0.5659 | 0.41854 | 0.01556 |
| rs4722172 | 7 | 22786532 | 40.0928349 | 0.0329 | 0.39048 | 0.57665 |
| rs11066301 | 12 | 112871372 | 130.305862 | 0.1981 | 0.00386 | 0.79809 |
| rs11057830 | 12 | 125307053 | 149.857655 | 0.1827 | 0.16692 | 0.65043 |
| rs10851907 | 15 | 78915864 | 102.193041 | 0.0029 | 0.45321 | 0.54385 |
| rs55791371 | 19 | 11188153 | 31.8736735 | 0.0027 | 0.08713 | 0.91016 |
| rs28451064 | 21 | 35593827 | 36.2865085 | 0.0789 | 0.51911 | 0.40204 |
Table 4.
Ancestry enrichment for 60 random SNPs generated by computational tools.
| rsnumber | chr | POS | cM | IBS | YRI | NA |
|---|---|---|---|---|---|---|
| rs62394277 | 5 | 169924749 | 185.487416 | 0.23 | 0.39 | 0.37 |
| rs12033818 | 1 | 106368717 | 134.610925 | 0.05 | 0.01 | 0.94 |
| rs10960270 | 9 | 11818115 | 25.717268 | 0.19 | 0.24 | 0.56 |
| rs2471660 | 12 | 75534099 | 89.727097 | 0.00 | 0.79 | 0.21 |
| rs12627722 | 21 | 17918366 | 7.52177123 | 0.15 | 0.70 | 0.15 |
| rs11653073 | 17 | 55687365 | 83.7730559 | 0.00 | 0.41 | 0.59 |
| rs17025629 | 3 | 88354232 | 109.781656 | 0.67 | 0.22 | 0.11 |
| rs7578180 | 2 | 158772058 | 176.901137 | 0.10 | 0.56 | 0.34 |
| rs75910375 | 17 | 11281457 | 29.8450381 | 0.01 | 0.49 | 0.50 |
| rs74936620 | 1 | 55832969 | 81.3389065 | 0.12 | 0.24 | 0.64 |
| rs77240763 | 5 | 145752501 | 153.573126 | 0.31 | 0.31 | 0.38 |
| rs75970478 | 3 | 138843097 | 150.937494 | 0.11 | 0.62 | 0.26 |
| rs12956301 | 18 | 71834423 | 105.245876 | 0.00 | 0.80 | 0.20 |
| rs10172437 | 2 | 174701949 | 193.753831 | 0.01 | 0.60 | 0.39 |
| rs56919396 | 14 | 95350520 | 95.3439121 | 0.36 | 0.16 | 0.48 |
| rs2567778 | 13 | 103742001 | 103.442192 | 0.36 | 0.52 | 0.12 |
| rs17128958 | 14 | 93708224 | 91.6536648 | 0.42 | 0.14 | 0.44 |
| rs576400715 | 2 | 89571430 | 120.624 | 0.05 | 0.30 | 0.65 |
| rs17089367 | 13 | 73011013 | 70.4386922 | 0.30 | 0.60 | 0.10 |
| rs10948131 | 6 | 44291641 | 69.8263172 | 0.00 | 0.57 | 0.43 |
| rs114354246 | 3 | 137295824 | 149.759253 | 0.15 | 0.61 | 0.24 |
| rs7128963 | 11 | 33599446 | 50.7219531 | 0.06 | 0.29 | 0.66 |
| rs6691697 | 1 | 42534375 | 70.0178338 | 0.07 | 0.32 | 0.61 |
| rs12060034 | 1 | 198224252 | 215.353224 | 0.28 | 0.21 | 0.51 |
| rs12205759 | 6 | 122752471 | 127.395537 | 0.04 | 0.43 | 0.54 |
| rs1611636 | 6 | 29836703 | 50.3216201 | 0.04 | 0.32 | 0.64 |
| rs10512946 | 3 | 134916937 | 148.665958 | 0.15 | 0.61 | 0.23 |
| rs72866243 | 18 | 2094235 | 5.55281054 | 0.45 | 0.44 | 0.11 |
| rs114208611 | 11 | 11100541 | 21.4527621 | 0.00 | 0.40 | 0.60 |
| rs13136539 | 4 | 174513213 | 183.196957 | 0.47 | 0.26 | 0.27 |
| rs77282632 | 3 | 121384210 | 133.9296 | 0.43 | 0.30 | 0.27 |
| rs730489 | 6 | 151399891 | 162.916228 | 0.29 | 0.48 | 0.23 |
| rs528255 | 8 | 12878637 | 33.8526902 | 0.10 | 0.51 | 0.39 |
| rs72834798 | 17 | 38217299 | 64.1399895 | 0.01 | 0.56 | 0.43 |
| rs16883890 | 5 | 9950908 | 22.6794291 | 0.12 | 0.08 | 0.80 |
| rs11996214 | 8 | 135211378 | 159.947948 | 0.01 | 0.60 | 0.39 |
| rs4782152 | 16 | 9326011 | 22.425679 | 0.01 | 0.13 | 0.86 |
| rs13432204 | 2 | 107262432 | 128.94 | 0.05 | 0.20 | 0.74 |
| rs16974292 | 16 | 84652644 | 114.995507 | 0.03 | 0.30 | 0.67 |
| rs116953645 | 19 | 46613411 | 72.3867019 | 0.01 | 0.31 | 0.67 |
| rs2422672 | 20 | 1990286 | 7.8190193 | 0.10 | 0.70 | 0.20 |
| rs17188743 | 6 | 30385111 | 50.6059471 | 0.05 | 0.32 | 0.63 |
| rs973295 | 14 | 94372768 | 92.5873731 | 0.39 | 0.15 | 0.46 |
| rs1889877 | 6 | 69729678 | 84.7215414 | 0.01 | 0.48 | 0.51 |
| rs10114476 | 9 | 96631459 | 113.576597 | 0.15 | 0.71 | 0.14 |
| rs113629052 | 9 | 95358140 | 112.692987 | 0.14 | 0.71 | 0.15 |
| rs62568088 | 9 | 6277534 | 15.8022578 | 0.12 | 0.32 | 0.56 |
| rs4631137 | 5 | 52420445 | 63.8860104 | 0.04 | 0.56 | 0.40 |
| rs2165183 | 2 | 4830190 | 9.36070063 | 0.11 | 0.67 | 0.21 |
| rs675465 | 4 | 20543817 | 36.4528078 | 0.33 | 0.27 | 0.40 |
| rs76887708 | 5 | 98125372 | 109.316657 | 0.00 | 0.76 | 0.24 |
| rs8043824 | 16 | 79847142 | 101.522556 | 0.01 | 0.31 | 0.68 |
| rs17123256 | 14 | 87745773 | 81.7304956 | 0.45 | 0.36 | 0.19 |
| rs115079994 | 22 | 45016212 | 55.6235323 | 0.01 | 0.07 | 0.92 |
| rs113899132 | 5 | 156284101 | 166.222937 | 0.17 | 0.31 | 0.52 |
| rs73142595 | 20 | 54320992 | 85.8174786 | 0.21 | 0.22 | 0.57 |
| rs7700639 | 5 | 6606807 | 16.6711274 | 0.20 | 0.01 | 0.79 |
| rs7714907 | 5 | 142145386 | 149.584982 | 0.28 | 0.37 | 0.35 |
| rs114959453 | 6 | 24423918 | 47.4334264 | 0.03 | 0.35 | 0.62 |
| rs9812688 | 3 | 86878962 | 109.260905 | 0.67 | 0.22 | 0.11 |
chr (chromosome); POS (position in GRCh37); cM (centimorgan). Ancestry data is presented as the average from IBS (Iberian population in Spain), YRI (Yoruba in Ibadan, Nigeria), and NA (America population) of 510 Caribbean Hispanic patients. Position is based on the human assembly GrCh37/h19 genome reference build by the Genome Reference Consortium.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



