
Submitted:
01 September 2023
Posted:
04 September 2023
You are already at the latest version
Abstract
Ants have a very sophisticated olfactory system; their communication is based on the interpretation of chemical trails known as semiochemicals. The escamolera ant (Liometopum apiculatum), has behaviors and physiological responses is induced by semiochemicals, without however, these have not been studied. Using an electronic nose analysis (e-nose) and chromatographic techniques, semiochemicals of L. apiculatum in nests, foraging trails and dissected the gasters were evaluated. Data were analyzed with multivariate statistics and a logistic regression model based on predictors (ant counts) and the semiochemicals identified in pre-season (PRE), season (TEM) and post-season (POS) of larval collecting. From the general pattern of volatiles with a natural separation in POS and a partial distribution between PRE and TEM 32 semiochemicals were identified inside the nests, those who can induce the behavioral responses of trail-following, alarm, aggression, and nestmate recognition. Trail counts indicated that ant traffic was higher in POS and that the presence of certain semiochemicals (response variable) have good fit in the model regression. These findings are intended to provide useful information and support decision-making for the conservation and sustainable use of L. apiculatum in central Mexico.
Keywords:
escamoles
; life cycle
; social insects
; chemical communication
; electronic nose
1. Introduction
Ants are insects with a wide geographic distribution, they are characterized by colonizing a great variety of terrestrial ecosystems [1,2,3,4,5]. The success of their survival is due to their sophisticated communication and work organization by castes, which is why ants are considered as one of the engineers of the ecosystem, since they recycle soil nutrients, improve the flow of energy in the edaphic component and biogeochemical cycles, they also collected and disperse seeds favoring plant dispersal [6,7].
Liometopum apiculatum (Mayr, 1870) (Hymenoptera: Fomicidae: Dolichoderinae) is a species of ant that is distributed in the Nearctic and subtropical zone of Mexico [8,9], this arthropod is known as the escamolera ant, a popular term for its reproductive caste larvae. Escamoles derives from the Nahuatl language "azcatlmolli" (Axcatl = ant and molli = stew) meaning ant stew [10]. L. apiculatum nests in the soil and in various substrates, e.g., at the base of Agave spp., decaying stems of Yucca spp., Opuntia spp., dead logs and under stones [11]. Collaborative work (castes) is divided into workers and breeders (queen, and winged males and females) [12]. Mating occurs in the spring (May to June), and mating between breeders takes place in the air, then the fertilized female lands and colonizes a new nest, ovipositing the first generation of workers (about 400 to 600 eggs) [13,14]. Escamoles are extracted from the nests in March-April and marketed for culinary preparations. They are appreciated for their delicate flavor and nutritional value, as they contain amino acids, fatty acids, minerals and vitamins [15,16].
The diet of L. apiculatum is omnivorous; however, it prefers a liquid diet, which it obtains mainly through trophobiosis, a behavior that consists of collecting honeydew excreted from other insects [11,14,17,18]. The workers of L. apiculatum are in charge of foraging and harvesting honeydew from different aphids and scale insects [17], they are observed walking 24.3 m long paths, in an area of 1 565 m2 in a schedule from 8:00 to 19:00 h, with a peak of activity at 17:00 h [14,19,20].
Mutualistic relationships as trophobiosis of L. apiculatum [17], and other ant species of the order Hemiptera, is moderated by semiochemicals involved in the search and detection of partners that provide them with the best and highest honeydew production [21,22]. The chemical communication of ants is involved in their entire life cycle, regulating the reproduction, nest mate recognition (cuticular hydrocarbons), queen and brood care, alarm systems, aggressiveness and pheromone tracking to locate food [23,24,25,26,27,28]. Trail-following to the food source has already been documented for three species of the genus Liometopum in North America: L. apiculatum Mayr, L. luctuosum Wheeler and L. accidentale; however, their composition has not yet been characterized [29], so it has only been reported that the odor on foraging trails appears to be similar to butyric acid. This behavior is suspected to be due to a trail pheromone, which may function as guidance and recruitment to feeding resources [18].
Integrated management and monitoring of insects includes various strategies for their control, of which semiochemicals such as pheromones and allelochemicals (allomones, kairomones, synomones, antimones and apneumones) that intervene in the intraspecific and interspecific communication of different groups of insects are used [30]. For example, Oliver et al. [31] reported different semiochemicals emitted by ants (Lasius niger) to control aphid dispersal (Acyrthosiphon pisum Harris and Aphis fabae Scopoli). While Appiah et al. [32] did fruit fly (Bactrocera invadens) control, using the weaver ant Oecophylla longinoda to expose its semiochemicals to a natural enemy of the fly (the endoparasite Fopuis arisanus) and significantly increase the percentage of parasitism in pupae of B. invadens. Another case is the findings of Xu et al. [21] who documented an association between ants-hemiptera (Solenopsis invicta and Aphis gossypii), reporting E-β-farnesene (EBF) as a semiochemical that stimulates a mutualistic relationship, where the ant controls the reproduction and dispersal of apterous individuals of A. gossypii.
In the last decade, trail pheromones have been synthesized on the foraging trails of several species of ants. The study of these chemical compounds is aimed at the creation of lethal baits (added with trail pheromones and insecticides) for ants considered a pest for humans, since they affect in adversely to ecosystems, agricultural systems and buildings [33,34,35,36]. The technology for the creation of synthetic pheromonal compounds is through extracts from various body parts (head, thorax, mesosome, etc.), it is based on chromatographic techniques, electroantennography [34] and electronic nose. The latter tool is considered a disruptive technology in the detection of volatile organic compounds (VOCs) for biological matrices [37,38].
Currently, the unplanned and unregulated extractive exploitation of escamoles is based on traditional practices as extractive management-filling-capping [39]; which, if well managed, managed to maintain stable populations of L. apiculatum. However, market supply and demand has led to excessive exploitation of their larvae, putting the survival of the species at risk [40,41]. An alternative to contribute to the sustainable management of L. apiculatum is to characterize its chemical odor signature and explain the physiological or behavioral responses that VOCs can cause. Therefore, the objective of the present investigation was to evaluate the semiochemical compounds in nests and on trails of L. apiculatum in pre-season, season and post-season the collection of larval, and to test if the chemical signature inside the nests is different by season. This study presents diverse ecological and biological reasoning on how semiochemicals compounds can influence the life cycle of L. apiculatum, a species of ecological and economic importance in central Mexico.
2. Materials and Methods
The present study was carried out in Environmental Management Unit (Unidad de Manejo Ambiental, UMA El Milagro) with an area of 328.4 ha, located in Villa González Ortega, Zacatecas (22° 37' 46.41" N, 101° 56' 25.36" W) at 2208 m of altitude (Figure 1). The climate type is arid temperate (BS0Kw), with a mean annual temperature between 12 and 18 °C [42] and an annual precipitation of 300 to 400 mm [43]. The dominant vegetation type is crassicule scrub (98%) and the rest is shrub secondary vegetation derived from desert microphilous scrub [44], with cacti (Opuntia spp., Opuntia imbricata, Agave spp.), and shrubs (Larrea tridentata, Flourensia cernua, Dalea bicolor, Parthenium incanum and Mimosa biuncifera) being the best represented plant species [45].
In the study area, were identified 15 nests (sampling units) of L. apiculatum subject to management, i.e., that the escamoles could be extracted. The choice of sampling units consisted of identifying the first nest and then, evaluating the nearest nest [45], selecting five anthills 70 m apart (Figure 1).
2.1. Air sampling inside the nests
Sampling was scheduled in the pre-season (PRE), season (TEM) and post-season (POS) of escamoles larvae collection (last week of March to first week of April 2022). These were conducted in PRE (03/03/2022), TEM (03/22/2022) and POS (04/04/2022).
An air sampling port was installed in the brood chamber of each nest, using a PVC pipe line (Ө = 15") coupled to a shut-off valve and a cast iron screw cap at the end. On the outside of each nest, the foraging trail with the highest traffic of ants was identified and marked by iron stake on the side of the trail. In each nest, air was sucked for one hour with a BOYU® pump at a flow rate of 4 l/min, connected with a hose to the sampling port, while at the outlet of the pump another hose was attached to direct the air flow to a metal balloon with capacity of 1.0 liter. The samples were transported in coolers at 4 °C to the Coordinación para la Innovación y Aplicación de la Ciencia y la Tecnología (CIACyT), Universidad Autónoma de San Luis Potosí. In each season, one balloon per nest and a duplicate, and two environmental air balloons were obtained as a control (n = 36 for the whole study).
2.2. Sampling on foraging trails and ant counts
Artificial trails were installed in the trails marked with stakes, fastened with 2.54 cm nails. These were constructed with 3 mm thick MDF (Medium Density Fibreboard) panels 20 cm wide × 15 cm long, and a walkway 5 cm wide × 10.5 cm long; the surface was lined with printer paper. Activated carbon cloth was attached to the walkways with tacks. The walkways were placed at a distance of approximately 5 m from the nests, making the center line of the structure (25.5 cm) coincide with the natural path of the nests.
To similar the trail-following behavior on the MDF trails, a 0.5 M sugar solution scented with rosemary essential oil [46] was prepared and applied in a straight line with a syringe (3 ml) onto the paper and activated charcoal cloth. Ants attracted to the sugar solution, at both ends of the artificial trail, consumed it and turned back to the foraging trails, emitting a trail of semiochemicals. This behavior was repeated until the arthropods reached the center of the path, stabilizing the free transit of ants in two directions with respect to the nest [47,48].
The MDF trails were left for 24 hours, the next day the activated carbon cloths were removed and introduced in 20 ml glass vials, which were capped with septa and aluminum seal. The vials were transported to the CIACyT in a cooler at 4 °C. At the end of the sampling seasons, 15 activated carbon cloths were obtained. To count ants, videos were taken with a digital camera LSR Nikon D5300 (Nikon Corp., Tokyo, Japan) on a tripod and an objective lens (18 ‒ 155 mm) focused on the rectangular surface of the artificial paths (avoiding the activated carbon walkways), four videos of 5 min duration and 15 min between each video. A total of 20 videos were obtained for all seasons in the five nests, which captured the movement of ants to track them in two directions [49].
2.3. Ant trapping
Live ants foraging on the surface of agaves, or on the trunks of yuccas at a height of 1.30 cm, were captured outside the nests. The collection was carried out with the individuals climbing into an entomological clamp and then shaking inside a Ziploc bag (16 × 16 cm), which was immediately closed to prevent the escape of the arthropods. Another strategy consisted of placing the open bag, held with both hands, on the bark of Yucca spp. and waiting for the ants to enter, achieving the capture by closing the bag. For each nest, 20 to 50 ants were captured (n= 150 to 250 individuals per season). The ants were transported in coolers at room temperature and transported to CIACyT.
2.4. Laboratory analysis
2.4.1. Determination of semiochemicals with the electronic nose (e-nose)
Fingerprint detection of semiochemicals from live ants was performed with a Cyranose 320 e-nose. This device is equipped with 32 chemical nanosensors that measure the intensities of VOCs with electrical resistance in mΩ, through a flow of nitrogen. For the analysis of the samples, four to six ants were separated from each nest in glass vials of 20 ml, which were capped with septa and aluminum seal, obtaining a total of five replicates per colony for each season (a total of n= 15 samples). Then, each vial was injected into the electronic nose with a constant flow of 120 ml/min of ultrapure hydrogen during 20 sec of recording, obtaining a specific odor impression for each stimulus. Before injection, each vial was left capped for 3 to 5 min, so that the ants would produce analytes (odor) in vapor phase, entrained by the e-nose suction system [38,50].
2.4.2. Identification of semiochemicals by gas chromatography coupled to mass spectrometry (GC-MS) on activated carbon cloths
Activated carbon cloths were incubated at 32 °C during 30 min and taken for analysis to a gas chromatograph (GC) (Agilent 6890) coupled to a mass spectrometry (MS) detector (Agilent 5975) with electron impact ionization (EI) sample ionization. Port injection was in splitless mode at a temperature of 250 °C. Helium was used as a carrier gas at a constant flow rate of 1.0 ml/min (total) and a pressure of 6.55 psi. An Agilent HP-5MS column (30 m × 0.25 mm x 0.25 μm) was used. The initial oven temperature was 40 °C/1 min, with programming at 2.5 °C/min up to 50 °C, increasing 10 °C/min up to 100 °C, then an increase of 8 °C/min up to 260 °C and held until minute 30. The emission tuning parameters were 34.6 μA, energy 69.9. SCAN mode (50 ‒ 230 m/z) was used to identify the fingerprint of VOCs emitted by ants transiting the activated carbon cloths.
The compounds were identified through the NIST 14 library. The results were obtained and processed using Chemstation software (Agilent®).
2.4.3. Identification of semiochemicals by GC-MS inside nests
Air samples from the nests were incubated at 32 °C during 60 min, then Headspace solid-phase microextraction (HS-SPME) was performed for manual GC-MS injection. Prior to use, the fiber was exposed in the GC injector for 30 min to 65 °C [38]. SPME was performed by exposing the fiber (silica fused with 100 μm polydimethylsiloxane) inside the metal balloons, and then manually introduced into the GC-MS injection port at 200 °C for 5 min. Samples were analyzed with a gas chromatograph (Agilent 6890) coupled to a mass spectrometry detector (Agilent 5975) in ionization mode (EI). Port injection was in splitless mode (220 °C). Helium was used as a carrier gas at a constant flow rate of 1.0 ml/min and a pressure of 8.67 psi. An Agilent HP-5MS column (30 m × 0.25 mm x 0.25 μm) was used. The initial oven temperature was 70 °C, with programming of 22 °C/min up to 180 °C, increasing 5 °C/min up to 200 °C and held constant until 15 min. The emission tuning parameters were 34.6 μA and energy 69.9. SCAN mode (50 ‒ 230 m/z) was used to fingerprint the semiochemicals emitted by ants inside the nests. The compounds were identified through the NIST 14 library. The results were obtained and processed using Chemstation software (Agilent®) [51].
2.4.4. Identification of semiochemicals by GC-MS of ant gasters
Sample preparation consisted of dissecting 50 ant gasters in 300 μl of hexane. Then, two aliquots were prepared by adding 10 ml of hexane in test tubes, which were placed in a sonicator for 1 min; the supernatant was transferred to two tubes (approximately 9 ml) and weighed on a granatary balance until the weights were equalized. Samples were centrifuged at 3500 rpm/4 min and subsequently evaporated at 37 °C and 40 psi pressure for 10 min. Samples were injected into the GC-MS (Agilent 6890, Agilent 5975) in electron impact ionization mode; operated in splitless mode at a temperature of 250 °C, using helium as a carrier gas, at a pressure of 9.80 psi and a constant flow rate of 1.0 ml/min. Separation of the components was performed with a HP 5MS column (30 m × 0.25 mm x 0.25 μm) (Agilent). The oven temperature ramp was 90 °C/2 min, with programming of 20 °C/min up to 250 °C, with hold at 5 min, and an increment of 300 °C (10° C/ min). Emission tuning parameters were 34.6 μA, energy 69.9; SCAN mode (50 ‒ 350 m/z) was used to identify the fingerprint of volatiles from ant gasters. Compounds were identified through the NIST 14 library and integrated using Chemstation software (Agilent®).
2.5. Statistical analysis
2.5.1. Analysis of data obtained from the e-nose
A multivariate analysis was performed to evaluate the responses of the electronic nose sensors and the volatiles emitted by the ants captured from each nest. The data recorded were the response changes of the resistances of the 32 nanosensors (Table S1). To reduce the effects of magnitude of the sensor responses (mΩ), automatic scaling was performed by subtracting the mean of the individual samples and dividing their standard deviation [50]. To predict the membership of each group, a Partial Least Squares-Discriminant Analysis (PLS-DA) and Variable Importance in Projection (VIP) [38]. This statistical method extracts the independent variables (e-nose sensor responses) by linear combination, and predicts the membership of each class (PRE, TEP and POS) to facilitate data interpretation. The analyses were performed with the freely distributed MetaboAnalyst 5.0 statistical software [52].
2.5.2. Analysis of data obtained with CG-SM on nests
With the semiochemicals identified by GC-SM and the NIST 14 library, a database and the CAS code of each compound was created. Then an a priori search was performed in Pherobase [53] and various scientific articles [30,35,54,55,56,57,58,59] for volatiles emitted by ants. With this information, semiochemicals that regulate some type of behavior (e.g. path-following) in ants and/or other associated insect groups (trophobiotic relationships) were refined and identified. With the cleaned database and the individual peak-area percentages obtained by GC-MS of each compound, a Principal Component Analysis (PCA) was performed; this method provides a summary of all the data by searching for correlations between variables, and reducing the number of components. The PCA expressed a qualitative global visual pattern, highlighting the similarities and differences between and within the sample, for which the percentage of variance explained was estimated. Subsequently a PLS-DA/VIP was performed to obtain the similarities and differences between the groups (PRE, TEP and POS) with MetaboAnalyst 5.0 statistical software.
2.5.3. Ant counts, effect of semiochemicals on observed frequencies
The videos taken on the artificial trails were analyzed with AntCounter, which is a freely distributed software that automatically detects the movement of ants (in two opposite directions) to identify their position, track them and count them over a period of time [49].
To analyze the effect of semiochemicals identified in the nests with the proportion of ants transiting the trails and by season, a nominal logistic regression was performed using JMP 17 (SAS Institute, Cary, NC). Prior to analysis, the adjusted AntCounter data (number of individuals to and from the nest) were taken, averaged and transformed to a
nominal polytomous variable with three categories (98 ‒ 229, 230 ‒ 362 and 363 ‒ 494) which was considered as the dependent variable of the model, while the predictors were the semiochemicals identified inside the nests, which were entered into the logit model as a binary variable (1, 0); where 1 represented the presence of semiochemical(s) and 0 the absence of semiochemical(s).
3. Results
3.1. E-nose
The PLS-DA of the semiochemicals emitted by the ants explained 61.9% of the variability of the data (Component 1 and 3) (Figure 2A). A natural separation was shown in the patterns of volatiles from post-season (POS) samples, a partial discrimination between pre-season (PRE) and season (TEM) is observed (Figure 2A). The variable of importance in the projection indicates that for the PRE group the sensors S31, S24, S5, S23, S13, S16, S22, S17 and S10 have the greatest contribution in the separation of the group. For POS they were S7, S28, S2, S4 and S8; while in TEM S11, S7, S28, S24, S13, S16 and S2 were observed in lower intensity (Figure 2B).
3.2. Gas chromatograph
Figure 3 shows a representative GC-MS chromatogram of the semiochemicals obtained from each season. 205 VOCs in PRE, 172 in TEM, and 271 in POS were identified, respectively. After data searching (Pherobase and scientific articles), 47 semiochemicals in PRE, 15 in TEM, and 40 in POS, were behavior ants associated. Final data (peak-area percentaje) (Table S2) were analyzed by multivariate statistical tests.
The PCA of the semiochemicals inside the nests explained 79.2% of the variability of the data in five components, while PC1 and PC2 explained 37.8% (PC3 =18.2%, PC4 = 18% and PC5 = 5.2%) (Figure 4).
The PLS-DA showed a clear separation between the patterns of the volatiles of each sample (Figure 5A), it was possible to visualize the discrimination between PRE, TEM and POS. The variable of importance in the projection indicates that for the PRE group, compounds 14 (5-methylundecane), 12 (eicosane), 29 (4-methyloctane), 28 (1- hexadecene) and 16 (1-tetradecene) present a greater contribution in the separation of the group. While for TEM the projection grouped compounds 19 (7- methyltridecane), 11 (2,6-dimethyl-7-octen-2-ol), 8 (undecane) and 4 (dodecane). Finally, in POS 7 (tridecane) and 6 (heptadecane) were the most important (Figure 5B).
Thirty-two semiochemicals were identified: nine (28%) in PRE, nine (28%) in TEM and 14 in POS (44%). The functional chemical groups identified in PRE were three alkanes, two alkenes and one cycloalkane. In TEM seven alkanes, one monoterpenoid (tertiary alcohol) and one aliphatic fluorinated compound; while in POS only alkanes were identified. The shortest chain compound was 4-methyloctane (C9H20) and eicosane (C20H42) with the highest number of carbons, both of which were only identified in PRE. In PRE and TEM, 1-hexadecene was identified, while TEM and POS matched dodecane and undecane. The most frequently occurring compounds in the three seasons were dodecane, undecane and 4-methyloctane.
3.3. Ant counts, effect of semiochemicals on observed frequencies
The videos analyzed with AntCounter, adjusted averages of the number of ants going to and from the nests (5 min/video), indicated that the season with the highest ant traffic on the artificial paths was in POS (40.8%, Figure 6A), with 1256 individuals counted for that period. The anthill with the highest incidence of ants in PRE was nest 5 (494 ± 180.5 individuals, Figure 6B). During TEM, nest 1 was the most transited (228 ± 53.3 individuals, Figure 6C), while in POS the highest transit of ants was observed in the same trail (430 ± 126.4 individuals, Figure 6D).
Figure 7 presents the summary of the effects of semiochemicals on ant transit, with a probability level of 0.05 (logistic regression ꭕ2 = 24.886, g.l. = 8, P = 0.001). Therefore, the statistical hypothesis that semiochemicals have a significant effect on the number of ants traversing the foraging paths in two directions is tested.
4. Discussion
Semiochemicals identified in L. apiculatum nests can induce behavioral or physiological responses. For example, 5-methylundecane (PRE), 7- methyltridecane and tridecane (POS) are volatiles that have been identified in virgin females and queens of Formica polyctena with some intraspecific function [60], perhaps this is a mixture of oviposition deterrents. In the study area the mating flight of L. apiculatum occurs in summer (first rain of June), the reproductive (females and males) are still in the pupal stages of development, so these compounds may be emitted exclusively by the queen.
In current research that has characterized the mixture of cuticular hydrocarbons (CHCs) and their functions, eicosane in Odontomachus brunneus is a volatile that stimulates recognition among colonies of its species [61]. Another formicide that emits eicosane is Oecophylla longinoda [62], which is suspected to induce the alarm reaction, and the source of emission is in the abdominal glands, including Dufour's gland. In the case of the subfamily Dolichoderinae, to which L. apiculatum belongs, studies reporting CHCs have possibly not been conducted. However, it is recorded that one of the sources of semiochemical emission in dolichoderines is the Pavan gland, in which volatiles with the same number of carbons as eicosane (eicosanal and eicosenal) are produced, which are a mixture of semiochemicals that induce trails-following [63].
The alkane 4-methyloctane, apparently not reported in ants, is likely to be present in L. apiculatum transfer in the trophic chain at different levels. It has been reported that the escamolera ant has trophobiotic relationships (ant-insect relationship) with hymenopterans of the families Pseudococcidae, Coccidae, Diaspididae, Eriococcidae, Ortheziidae, Aphididae and Membracidae [17], from which it obtains honeydew and these in turn feed on the phloem of Agave spp. and Yucca spp. (insect-plant relationship). It has also been documented that 4-methyloctane induces reactions in another group of insects. It has been demonstrated in Taxoptera aurantii (Boyer), that 4-methyloctane is a cuticular component of the aphid (in conjunction with other semiochemicals), which encourages the recognition of individuals in its population, but it is also one of the volatiles that attracts its predators (Chrysopa synica, Coccinella septempunctata and Aphidius sp.) [64].
The alkene 1-tetradecene was identified in PRE and TEM. For example, in the ant Pogonomyrmex barbatus it is one of the volatiles that emits with greater abundance, it has been suggested that it is one of the trunk routes markers leading to the food source [54]. In other social arthropods 1-tetradecene is a substance that functions as an antimicrobial agent [65], it is possible that L. apiculatum produces this compound as a CHC, which acts as a defense to different pathogens.
The compound 2,6-dimethyl-7-octen-2-ol (dihydromyrcenol) is an unsaturated non-cyclic alcohol used as a citrus-type fragrance ingredient [66,67], which is apparently not found in nature [68]. In contrast, dihydromyrcenol has been identified in bee honey in Brazil [69] and Türkiye [70], in study areas with different floral varieties, mixtures and characterized have been with different analytical techniques for VOCs. It cannot be inferred why this alcohol is present in honey produced by bees, but this has a more direct relationship with ants, since 2,6-dimethyl-7-octen-2-ol can be a precursor of formic acid, it is known that L. apiculatum emits formic acid [12], as well as in most of the ants in the family Formicidae [71], however; the metabolic pathway of 2,6- dimethyl-7-octen-2-ol has not been studied in depth [72].
Undecane was a volatile observed in TEM and POS in nests, although it is not possible to directly infer what behavioral or physiological reactions this volatile stimulates in L. apiculatum in our work, it possibly affects intraspecific and interspecific communication in the ant. Such is the case in Linepithema humile, where undecane induces aggressive responses against Pogonomyrmex spp. [25]. It has also been reported as an alarm pheromone of Camponotus obscuripes [73], a marker of foraging routes in Pogonomyrmex barbatus and P. rugosus [54], as well as a trace semiochemical in trails trodden by Solenopsis geminata [56].
Undecane is an important volatile for Formica argentea and Camponotus modoc, and is a major component of the alarm pheromone for with Dufour's gland many ant species [35,74]. Another of their physiological responses is related to mating, which occurs in the form of swarming [60]. The identification of undecane using SPME and extracts with hexane has already been documented for Oecophylla longinoda and Camponotus japonicus, in the case of L. apiculatum the same analysis technique and solvent was used for the formulation of extracts with gasters. However, undecane was not identified in these chemical aggregates, it is possible that this difference is the source of emission, since for O. longinoda Dufour glands were dissected and in C. japonicus Dufour glands plus poison gland were dissected for analysis [62,75].
In an analyzed sample of gasters (PRE), iridomyrmecin was detected, a compound identified in the Dolichoderinae family, which is a volatile secretion of the anal gland that causes alarm behaviors in Liometopum occidentale and Iridomyrmex humilis [76].
Dodecane was another semiochemical identified in TEM. This volatile is found abundantly in the Dufour's gland of Pogonomyrmex rugosus and P. barbatus, which stimulates aggression behaviors, but it is also speculated that, in conjunction with other pheromone mixtures, it may be used as a colony return guidance system or even for defense and alarm in interspecific relationships [25]. Dodecane has also been identified in Pogonomyrmex salinus and Messor lobognathus, along with other compounds, is a cuticular hydrocarbon that may be related to trail-following and interspecific recognition [77].
The interspecific relationships of L. apiculatum with other insects or other ants, as well as the semiochemicals that stimulate behavioral responses have not been studied. However, the presence of the red mite Trombidium holosericeum Linnaeus [12], which possibly mimics its odor to avoid being attacked by ants, has been detected inside the nests and trails of L. apiculatum.
It has been documented that different mixtures of volatile chemicals, used as a guidance system to food sources and recruitment, are used by ants from different colonies (eavesdrop) [78]. This behavior has not been investigated with L. apiculatum, but in the study area it shares habitat with Atta mexicana [9], a leaf-cutting species, which has been observed transiting several sections of the paths of L. apiculatum when it is sheltered during the hours of highest solar incidence (12:00 to 15:00 h) [14]. The feeding regime of both species is completely different, but it is possible that eavesdrop by A. mexicana; this behavior perhaps allows it to find plant matter within the forage range of L. apiculatum, which is 1,565 ± 535.63 m2 [20].
Heptadecane was identified in POS, it is a hydrocarbon with different functions, for example, in Solenopsis invicta and S. richteri it is a component of trail pheromones, which has been reported in worker ants and winged females using HS-SPME and GC-MS as analysis technique [79]. In the weaver ant Oecophylla smaragdina is one of the CHCs, which can function as nest mate recognition [80]. Heptadecane has also been identified in a hydrocarbon mixture in Pachycondyla sennaarensis in Dufour glands, which is considered the main source of trail pheromones for the species [81]. In our research, no trace semiochemicals were identified in activated carbon cloths. In this regard, Attygalle et al. [63] analyzed trail pheromones from ants of the subfamily Dolichederinae, reported that (Z)-9-hexadecenal, (Z)-9-octadecenal and octadecenal stimulated this behavior; they used GC-MS and subsequent derivatizations (dimethyl disulfide), with molecular ion at m/z 86, 173 and 360. The experiments with L. apiculatum used non-targeted metabolomics with at m/z ratio 50-230, it is likely, that the chemical analytes in the samples did not match the detection level of the equipment, or the volatile constituents degraded and/or escaped to the environment before being injected.
The results of ant count on the artificial trails and their association with volatilomas indicated that dodecane (P = 0.00063) and undecane (P = 0.01797) are hydrocarbons that are related to the number of ants traversing the trails. These are likely part of trail semiochemicals or trail are chemical signals that maintain social cohesion among nest mates of L. apiculatum [78].
Tridecane (P = 0.03855), has been identified in the reproductive caste of several species of ants with Dufour's gland, but its biological function is still unclear [60,75]. However, this alkane has been reported most frequently in fertile males and females, and in lower proportions in worker ants of Pachycondyla sennaarensis, Messor meridionalis and M. foreli mixed with other hydrocarbons [81]. The presence of tridecane in L. apiculatum nests may be due to the queen emitting the volatile, or even young (more developed) princesses in POS.
The identification of semiochemicals associated with the transit of L. apiculatum ants does not have a precedent or research that reports, much less in its natural distribution. However, there are records that L. apiculatum workers increase their foraging intensity in the dry season (March-April), period in which male and female larvae are developing [12,20]. In our work, in POS the greatest traffic of ants was observed on the trails. It is likely that the workers circle the food resources more to increase food consumption and transport, providing it to the reproductive caste, as a strategy to recover individuals that have been extracted from the brood chamber during the escamoles larvae collection season, an activity that ant’s worker may continue carry out until males and females emerge for mating in the first rain of June [12].
The study of semiochemicals from L. apiculatum can be useful for the production of synthetic pheromonal compounds with different functions, such as marking trails that guide ants to specific food resources, and even dispersing volatiles that incite them to reproductive flight. In our research we identified metabolites with a non-targeted (qualitative) approach; future research should perform targeted metabolomics, synthesize the semiochemicals and prove that they are inciters of certain behavior and/or physiological response through bioassays with L. apiculatum nests.
5. Conclusions
The global analysis of the semiochemical (e-nose) patterns of L. apiculatum confirms that the ants emitted different volatiles seasonally. Inside the nests, 32 semiochemicals were identified, where the most prominent functional group was alkanes of different chain lengths (C11:C20). We did not identify semiochemicals in the artificial trails; however, the count of ants on the trails was a good predictor to associate with the presence of volatiles. All identified compounds are likely to be behavioral regulators that modulate physiological responses, intraspecific communication, and social cohesion in L. apiculatum nests. The findings of this study strengthen the current knowledge of the escamolera ant, and is the starting point for the creation of artificial nests and to promote sustainable collection of escamoles.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Response of electronic nose sensors, Table S2: Individual percentages of peak area obtained by GC-MS (L. apiculatum nests).
Author Contributions
Conceptualization, C.L.J.D and T.A.L.A.; methodology, C.L.J.D. and F.R.R.; formal analysis, C.L.J.D, F.R.R. and U.L.S.; investigation, C.L.J.J.D. and V.P.M.R.; supervision, V.P.M.R.; writing-original draft preparation, C.L.J.D, M.M.J.F. and S.L.R.; writing-review and editing, C.L.J.D., V.P.M.R. and T.A.L.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research did not receive external funding.
Institutional Review Board Statement
The animal subjects used in this study were ant species Liometopum apiculatum, which are invertebrates and exempt from this requirement. No specific permits were required for the collection from the field and for the maintenance in laboratory. This study did not involve any endangered or protected species, or any protected areas.
Data Availability Statement
All data are contained in the article and the Supplementary Materials.
Acknowledgments
To Consejo Nacional de Humanidades, Ciencias y Tecnologías, Programa de Becas de Estancia Posdoctoral Académica (I1200/224/2021-l1200/320/2022) for awarding a grant for the financial support of the first author.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Escárraga, M.; Guerrero, R. Hormigas. Un Mundo de Meñiques Gigantes. INFOZOA Boletín de Zoología 2014, 4, 1–16. [Google Scholar]
- Guénard, B.; Weiser, M.D.; Gómez, K.; Narula, N.; Economo, E.P. The Global Ant Biodiversity Informatics (GABI) Database: Synthesizing Data on the Geographic Distribution of Ant Species (Hymenoptera: Formicidae). Myrmecological News 2017, 24, 83–89. [Google Scholar] [CrossRef]
- Stork, N.E. How Many Species of Insects and Other Terrestrial Arthropods Are There on Earth? Annual Review of Entomology 2018, 63, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Hakala, S.M.; Seppä, P.; Helanterä, H. Evolution of Dispersal in Ants (Hymenoptera: Formicidae): A Review on the Dispersal Strategies of Sessile Superorganisms. Myrmecological News 2019, 29, 35–55. [Google Scholar] [CrossRef]
- Lessard, J.P. Ant Community Response to Disturbance: A Global Synthesis. Journal of Animal Ecology 2019, 88, 346–349. [Google Scholar] [CrossRef]
- Fernandez-Bou, A.S.; Dierick, D.; Swanson, A.C.; Allen, M.F.; Alvarado, A.G.F.; Artavia-León, A.; Carrasquillo-Quintana, O.; Lachman, D.A.; Oberbauer, S.; Pinto-Tomás, A.A.; et al. The Role of the Ecosystem Engineer, the Leaf-Cutter Ant Atta Cephalotes, on Soil CO2 Dynamics in a Wet Tropical Rainforest. Journal of Geophysical Research: Biogeosciences 2019, 124, 260–273. [Google Scholar] [CrossRef]
- De Almeida, T.; Mesléard, F.; Santonja, M.; Gros, R.; Dutoit, T.; Blight, O. Above- and below-Ground Effects of an Ecosystem Engineer Ant in Mediterranean Dry Grasslands. Proceedings of the Royal Society B: Biological Sciences 2020, 287, 20201840. [Google Scholar] [CrossRef]
- Del Toro, I.; Pacheco, J.A.; Mackay, W.P. Revision of the Ant Genus Liometopum (Hymenoptera: Formicidae). Sociobiology 2009, 53, 309–316. [Google Scholar]
- Dáttilo, W.; Vásquez-Bolaños, M.; Ahuatzin, D.A.; Antoniazzi, R.; Chávez-González, E.; Corro, E.; Luna, P.; Guevara, R.; Villalobos, F.; Madrigal-Chavero, R.; et al. Mexico Ants: Incidence and Abundance along the Nearctic–Neotropical Interface. Ecology 2020, 101, e02944–1. [Google Scholar] [CrossRef]
- Ramos Rostro, B.; Salazar, B.Q.; Ramos-Elorduy, J.; Moreno, J.M.P.; Campos, S.C.A.; Pérez, A.G.; García, V.D.B. Análisis Químico y Nutricional de Tres Insectos Comestibles de Interés Comercial en la Zona Arqueologica del Municipio de San Juan Teotihuacan y en Otumba, en el Estado de México. Interciencia 2012, 37, 914–920. [Google Scholar]
- Miller, T.E.X. Does Having Multiple Partners Weaken the Benefits of Facultative Mutualism? A Test with Cacti and Cactus-Tending Ants. Oikos 2007, 116, 500–512. [Google Scholar] [CrossRef]
- Lara, P.; Aguirre, J.R.; Castillo, P.; Reyes, J.A. Biología y Aprovechamiento de la Hormiga de Escamoles, Liometopum Apiculatum Mayr (Hymenoptera: Formicidae). Acta zool. mexicana 2015, 31, 251–264. [Google Scholar]
- Ramos-Elorduy, J.; Delage Darchen, B.; Cuadriello Aguilar, J.I.; Galindo Miranda, N.; Pino Moreno, J.M. Ciclo de Vida y Fundación de las Sociedades de Liometopum apiculatum M. (Hymenoptera, Formicidae). Anales del Instituto de Biología. UNAM. Ser Zool. 184, 54, 161–176. [Google Scholar]
- Ramos-Elorduy, J.; Delage-Darchen, B.; Miranda, N.G.; Moreno, J.M.P. Observaciones Bioecotologicas de Liometopum Apiculatum M. y Liometopum Occidentale Var. Luctuosum W. (Hymenoptera-Formicidae). An. Inst. Biol. Univ. Nac. Auton. Mexico Ser. Zool. 1988, 58, 341–354. [Google Scholar]
- Ángeles Tovar, N.; Estrada Yescas, M.T.; Sandoval García, M. del P.; Vega Serrano, F.V.; Onofre Sánchez, J.E. Recolección y Temporalidad de Liometopum apiculatum M. (Escamoles) En El Municipio de Nantzha, Hidalgo México. Punto de vista 2022, 12, 72–87. [Google Scholar] [CrossRef]
- Escamilla, F.; Ariza, J. Nutrient and Oil Profile of Escamol, an Edible Larva of Ants (Liometopum Apiculatum Mayr). Future of Food: Journal on Food, Agriculture and Society 2021, 9, 1–9. [Google Scholar] [CrossRef]
- Velasco, C.; Corona-Vargas, M.; Peña-Martinez, R. Liometopum apiculatum (Formicidae: Dolichoderinae) y su Relacion Trofobiotica con Hemiptera Sternorrhyncha en Tlaxco, Tlaxcala, México. Acta Zoológica Mexicana (n.s) 2007, 23, 31–42. [Google Scholar] [CrossRef]
- Hoey-Chamberlain, R.; Rust, M.K.; Klotz, J.H. A Review of the Biology, Ecology and Behavior of Velvety Tree Ants of North America. Sociobiology 2013, 60, 1–10. [Google Scholar] [CrossRef]
- Mackay, W.; Mackay, E. The Ants of New Mexico (Hymenoptera: Fromicidae), 1st ed.; The Edwin Mellen Press: Lewiston, Ny, 2002; pp. 231–234. [Google Scholar]
- Rafael-Valdez, J.; Tarango-Arámbula, L.A.; Ugalde-Lezama, S.; Cruz-Labana, J.D.; Clemente-Sánchez, F.; Cadena-Iñiguez, J. Foraging Amplitude of the Escamolera Ant (Liometopum Apiculatum Mayr, Hymenoptera: Formicidae) in a Semi-Arid Area of the Zacatecan Highlands. Revista Chapingo Serie Zonas Áridas 2019, 18, 5–19. [Google Scholar] [CrossRef]
- Xu, T.; Xu, M.; Lu, Y.; Zhang, W.; Sun, J.; Zeng, R.; Turlings, T.C.J.; Chen, L. A Trail Pheromone Mediates the Mutualism between Ants and Aphids. Current Biology 2021, 31, 4738–4747. [Google Scholar] [CrossRef]
- Xu, T.; Chen, L. Chemical Communication in Ant-Hemipteran Mutualism: Potential Implications for Ant Invasions. Current Opinion in Insect Science 2021, 45, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Kleeberg, I.; Menzel, F.; Foitzik, S. The Influence of Slavemaking Lifestyle, Caste and Sex on Chemical Profiles in Temnothorax Ants: Insights into the Evolution of Cuticular Hydrocarbons. Proceedings of the Royal Society B: Biological Sciences 2017, 284, 20162249. [Google Scholar] [CrossRef]
- Wyatt, T.D. Pheromones. Current Biology 2017, 27, R739–R743. [Google Scholar] [CrossRef] [PubMed]
- Welzel, K.F.; Lee, S.H.; Dossey, A.T.; Chauhan, K.R.; Choe, D.H. Verification of Argentine Ant Defensive Compounds and Their Behavioral Effects on Heterospecific Competitors and Conspecific Nestmates. Scientific Reports 2018, 8, 1477. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Grodowitz, M.J.; Chen, J. Electrophysiological Responses of Eighteen Species of Insects to Fire Ant Alarm Pheromone. Insects 2019, 10, 403. [Google Scholar] [CrossRef] [PubMed]
- Billen, J.; Morgan, E.D. Pheromone Communication in Social Insects: Sources and Secretions. In Pheromone Communication in Social Insects: Ants, Wasps, Bees, and Termites, 1st ed; Vander Meer, R.K., Breed, M.D., Winston, M., Espelie, K.E, Eds.; Taylor & Francis: New York, NY, 2019; Volume 2, pp. 1–31. [Google Scholar]
- Ge, J.; Ge, Z.; Zhu, D.; Wang, X. Pheromonal Regulation of the Reproductive Division of Labor in Social Insects. Frontiers in Cell and Developmental Biology 2020, 8. [Google Scholar] [CrossRef]
- Pekár, S. Ant-Mimicking Spider Actively Selects Its Mimetic Model (Araneae: Gnaphosidae; Hymenoptera: Formicidae). Myrmecological News 2020, 30, 131–137. [Google Scholar] [CrossRef]
- Abd El-Ghany, N.M. Semiochemicals for Controlling Insect Pests. Journal of Plant Protection Research 2019, 59, 1–11. [Google Scholar] [CrossRef]
- Oliver, T.H.; Mashanova, A.; Leather, S.R.; Cook, J.M.; Jansen, V.A.A. Ant Semiochemicals Limit Apterous Aphid Dispersal. Proceedings of the Royal Society B: Biological Sciences 2007, 274, 3127–3131. [Google Scholar] [CrossRef]
- Appiah, E.F.; Ekesi, S.; Afreh-Nuamah, K.; Obeng-Ofori, D.; Mohamed, S.A. African Weaver Ant-Produced Semiochemicals Impact on Foraging Behaviour and Parasitism by the Opiine Parasitoid, Fopius Arisanus on Bactrocera Invadens (Diptera: Tephritidae). Biological Control 2014, 79, 49–57. [Google Scholar] [CrossRef]
- Welzel, K.F.; Choe, D.H. Development of a Pheromone-Assisted Baiting Technique for Argentine Ants (Hymenoptera: Formicidae). Journal of Economic Entomology 2016, 109, 1303–1309. [Google Scholar] [CrossRef] [PubMed]
- Chalissery, J.M.; Renyard, A.; Gries, R.; Hoefele, D.; Alamsetti, S.K.; Gries, G. Ants Sense, and Follow, Trail Pheromones of Ant Community Members. Insects 2019, 10, 383. [Google Scholar] [CrossRef]
- Renyard, A.; Alamsetti, S.K.; Gries, R.; Munoz, A.; Gries, G. Identification of the Trail Pheromone of the Carpenter Ant Camponotus Modoc. Journal of Chemical Ecology 2019, 45, 901–913. [Google Scholar] [CrossRef] [PubMed]
- Angulo, E.; Hoffmann, B.D.; Ballesteros-Mejia, L.; Taheri, A.; Balzani, P.; Bang, A.; Renault, D.; Cordonnier, M.; Bellard, C.; Diagne, C.; et al. Economic Costs of Invasive Alien Ants Worldwide. Biological Invasions 2022, 24, 2041–2060. [Google Scholar] [CrossRef]
- Diamond, A.; Schmuker, M.; Berna, A.Z.; Trowell, S.; Nowotny, T. Classifying Continuous, Real-Time e-Nose Sensor Data Using a Bio-Inspired Spiking Network Modelled on the Insect Olfactory System. Bioinspiration & Biomimetics 2016, 11, 26002. [Google Scholar] [CrossRef]
- Díaz de León-Martínez, L.; Flores-Ramírez, R.; López-Mendoza, C.M.; Rodríguez-Aguilar, M.; Metha, G.; Zúñiga-Martínez, L.; Ornelas-Rebolledo, O.; Alcántara-Quintana, L.E. Identification of Volatile Organic Compounds in the Urine of Patients with Cervical Cancer. Test Concept for Timely Screening. Clinica Chimica Acta 2021, 522, 132–140. [Google Scholar] [CrossRef]
- Dinwiddie, M.L.; Jones, R.W.; Roitman-Genoud, P.; Tarango-Arámbula, L.A.; Malda-Barrera, G.X. Estudio Etnoentomologico de la Hormiga Escamolera (Liometopum Apiculatum) en dos Localidades del estado de Querétaro. AGROProductividad 2013, 6, 27–34. [Google Scholar]
- Berumen Jiménez, M.; Valdez Cepeda, R.D.; Méndez Gallegos, S. de J.; Cadena Íñiguez, J.; Esparza Orozco, A.; Tarango Arámbula, L.A. Determination of the Conservation Status of the “Escamolera” Ant (Liometopum apiculatum Mayr) in Mexico by the Species Risk Assessement Methodology – MER. Agrociencia 2021, 55, 539–555. [Google Scholar] [CrossRef]
- Briones-Santoyo, J.A.; Tarango-Arámbula, L.A.; Velázquez-Martínez, A.; Reyes-Hernández, V.J.; Salazar-Borunda, M.A. Edible Insect Harvest in Pinos, Zacatecas, Mexico. Agro Productividad 2022, 15, 37–49. [Google Scholar] [CrossRef]
- García, E.; Comisión Nacional Para El Conocimiento y Uso de la Biodiversidad (CONABIO, 2001). “Climas” (Clasificación de Koppen, Modificado Por García). Available online: http://www.conabio.gob.mx/informacion/gis/?vns=gis_root/clima/climas/clima1mgw (accessed on 11 April 2023).
- García, E.; Comisión Nacional Para El Conocimiento y Uso de La Biodiversidad (CONABIO, 2008). Climatología, Precipitación. Available online: http://www.conabio.gob.mx/informacion/gis/?vns=gis_root/clima/precip/isoyt1mgw (accessed on 11 April 2023).
- INEGI Conjunto de Datos Vectoriales de Uso de Suelo y Vegetación. Escala 1:250 000, Serie VII. Conjunto Nacional.’, Escala: 1:250 000. Edición: 1. Instituto Nacional de Estadística y Geografía, México Available online:. Available online: http://www.conabio.gob.mx/informacion/gis/?vns=gis_root/clima/precip/isoyt1mgw (accessed on 11 April 2023).
- Hernández-Roldan, E.; Tarango-Arámbula, L.A.; Ugalde-Lezama, S.; Hernández-Juárez, A.; Cortez-Romero, C.; Cruz-Miranda, Y.; Morales-Flores, F.J. Hábitat y Densidad de Nidos de la Hormiga Escamolera (Liometopum apiculatum Mayr) En Una UMA de Zacatecas, México. Agroproductividad 2017, 10, 10–17. [Google Scholar]
- Oberhauser, F.B.; Schlemm, A.; Wendt, S.; Czaczkes, T.J. Private Information Conflict: Lasius niger Ants Prefer Olfactory Cues to Route Memory. Animal Cognition 2019, 22, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Oberhauser, F.B.; Wendt, S.; Czaczkes, T.J. Trail Pheromone Does Not Modulate Subjective Reward Evaluation in Lasius niger Ants. Ants. Front. Psychol. 2020, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Nishisue, K.; Koyama, S.; Satoh, T. Identification of the Argentine Ant Linepithema humile (Hymenoptera: Formicidae) Using an Artificially Synthesized Trail Pheromone and Its Effects on Native Japanese Ants. Applied Entomology and Zoology 2020, 55, 141–147. [Google Scholar] [CrossRef]
- Bustamante, S.; Amarillo-Suárez, A.R. AntCounter Software: Counting Leaf-Cutting Ants Was Never so Precise, Fast and Easy. Journal of Insect Behavior 2016, 29, 262–272. [Google Scholar] [CrossRef]
- Rodríguez-Aguilar, M.; Díaz de León-Martínez, L.; Gorocica-Rosete, P.; Pérez-Padilla, R.; Domínguez-Reyes, C.A.; Tenorio-Torres, J.A.; Ornelas-Rebolledo, O.; Mehta, G.; Zamora-Mendoza, B.N.; Flores-Ramírez, R. Application of Chemoresistive Gas Sensors and Chemometric Analysis to Differentiate the Fingerprints of Global Volatile Organic Compounds from Diseases. Preliminary Results of COPD, Lung Cancer and Breast Cancer. Clinica Chimica Acta 2021, 518, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Lee-Rangel, H.A.; Mendoza-Martinez, G.D.; Díaz de León-Martínez, L.; Relling, A.E.; Vazquez-Valladolid, A.; Palacios-Martínez, M.; Hernández-García, P.A.; Chay-Canul, A.J.; Flores-Ramirez, R.; Roque-Jiménez, J.A. Application of an Electronic Nose and HS-SPME/GC-MS to Determine Volatile Organic Compounds in Fresh Mexican Cheese. Foods 2022, 11, 1887. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; Morais, D.A. de L.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the Gap between Raw Spectra and Functional Insights. Nucleic Acids Research 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Becher, P.G.; Hagman, A.; Verschut, V.; Chakraborty, A.; Rozpędowska, E.; Lebreton, S.; Bengtsson, M.; Flick, G.; Witzgall, P.; Piškur, J. Chemical Signaling and Insect Attraction Is a Conserved Trait in Yeasts. Ecology and Evolution 2018, 8, 2962–2974. [Google Scholar] [CrossRef]
- Hölldobler, B.; Oldham, N.J.; Liebig, J.; Liu, Y.; David Morgan, E. Dufour Gland Secretion in the Harvester Ant Genus Pogonomyrmex. Chemoecology 2004, 14, 101–106. [Google Scholar] [CrossRef]
- Morgan, D.E. Trail Pheromones of Ants. Physiological Entomology 2009, 34, 1–17. [Google Scholar] [CrossRef]
- Igwe, O.U.; Offiong, S. Chemistry of Semiochemicals Used as Trail Pheromones in Tropical Fire Ant (Solenopsis geminata) 2015, 7, 34–40.
- Thoenen, M. The Effect of Iridomyrmecin on Native Ant Foraging Behavior. Spring 2017, 1–15. [Google Scholar]
- Hamilton, N.; Jones, T.H.; Shik, J.Z.; Wall, B.; Schultz, T.R.; Blair, H.A.; Adams, R.M.M. Context Is Everything: Mapping Cyphomyrmex-Derived Compounds to the Fungus-Growing Ant Phylogeny. Chemoecology 2018, 28, 137–144. [Google Scholar] [CrossRef]
- Chalissery, J.M.; Renyard, A.; Gries, R.; Hoefele, D.; Alamsetti, S.K.; Gries, G. Ants Sense, and Follow, Trail Pheromones of Ant Community Members. Insects 2019, 10, 383. [Google Scholar] [CrossRef]
- Lofqvist, J.; Bergstrom, G. Volatile Communication Substances in Dufour’s Gland of Virgin Females and Old Queens of the Ant Formica Polyctena. Journal of Chemical Ecology 1980, 6, 309–320. [Google Scholar] [CrossRef]
- Santos Junior, L.C.; Michelutti, K.B.; Bernardi, R.C.; Silva, E.P.; Cardoso, C.A.L.; Antonialli-Junior, W.F. You Smell Different! Temperature Interferes with Intracolonial Recognition in Odontomachus brunneus. Sociobiology 2022, 69, e6235. [Google Scholar] [CrossRef]
- Mekonnen, B.; Cheseto, X.; Pirk, C.; Yusuf, A.; Ekesi, S.; Deletre, E.; Torto, B. Re-Analysis of Abdominal Gland Volatilome Secretions of the African Weaver Ant, Oecophylla longinoda (Hymenoptera: Formicidae). Molecules 2021, 26, 871. [Google Scholar] [CrossRef]
- Attygalle, A.B.; Mutti, A.; Rohe, W.; Maschwitz, U.; Garbe, W.; Bestmann, H.J. Trail Pheromone from the Pavan Gland of the Ant Dolichoderus thoracicus (Smith) Pheromones, 108 [1]. Naturwissenschaften 1998, 85, 275–277. [Google Scholar] [CrossRef]
- Han, B.; Chen, Z. Behavioral and Electrophysiological Responses of Natural Enemies to Synomones from Tea Shoots and Kairomones from Tea Aphids, Toxoptera aurantii. Journal of Chemical Ecology 2002, 28, 2203–2219. [Google Scholar] [CrossRef] [PubMed]
- Lammers, A.; Zweers, H.; Sandfeld, T.; Bilde, T.; Garbeva, P.; Schramm, A.; Lalk, M. Antimicrobial Compounds in the Volatilome of Social Spider Communities. Frontiers in Microbiology 2021, 12, 700693. [Google Scholar] [CrossRef]
- Ham, J.E.; Raymond Wells, J. Surface Chemistry of Dihydromyrcenol (2,6-Dimethyl-7-Octen-2-Ol) with Ozone on Silanized Glass, Glass, and Vinyl Flooring Tiles. Atmospheric Environment 2009, 43, 4023–4032. [Google Scholar] [CrossRef]
- Politano, V.T.; Lewis, E.M.; Hoberman, A.M.; Christian, M.S.; Diener, R.M.; Api, A.M. Evaluation of the Developmental Toxicity of Dihydromyrcenol in Rats. International Journal of Toxicology 2009, 28, 80–87. [Google Scholar] [CrossRef]
- McGinty, D.; Letizia, C.S.; Api, A.M. Fragrance Material Review on Dihydromyrcenol. Food and Chemical Toxicology 2010, 48, S70–S75. [Google Scholar] [CrossRef] [PubMed]
- Nunes Moreira, F.I.; Lucena de Medeiros, L.; Moreira de Carvalho, L.; Souza Olegario, L.; Sousa Galvão, M.; Alves Monteiro da Franca, S.; Alencar Bezerra, T.K.; Dos Santos Lima, M.; Suely Madruga, M. Quality of Brazilian Stingless Bee Honeys: Cephalotrigona capitata/Mombucão and Melipona scutellaris Latrelle/Uruçu. Food Chemistry 2023, 404, 134306. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, O.; Gurkan, H.; Sahingil, D.; Degirmenci, A.; Er Kemal, M.; Kolayli, S.; Hayaloglu, A.A. Floral Authentication of Some Monofloral Honeys Based on Volatile Composition and Physicochemical Parameters. European Food Research and Technology 2022, 248, 2145–2155. [Google Scholar] [CrossRef]
- Peerzada, N.; Pakkiyaretnam, T.; Renaud, S. Volatile Constitutents of the Green Ant Oecophylla smaragdina. Agricultural and Biological Chemistry 1990, 54, 3335–3336. [Google Scholar] [CrossRef]
- Belsito, D.; Bickers, D.; Bruze, M.; Calow, P.; Greim, H.; Hanifin, J.M.; Rogers, A.E.; Saurat, J.H.; Sipes, I.G.; Tagami, H. A Safety Assessment of Non-Cyclic Alcohols with Unsaturated Branched Chain When Used as Fragrance Ingredients. Food and Chemical Toxicology 2010, 48, S1–S42. [Google Scholar] [CrossRef]
- Fujiwara-Tsujii, N.; Yamagata, N.; Takeda, T.; Mizunami, M.; Yamaoka, R. Behavioral Responses to the Alarm Pheromone of the Ant Camponotus obscuripes (Hymenoptera: Formicidae). Zoological Science 2006, 23, 353–358. [Google Scholar] [CrossRef]
- Lenz, E.L.; Krasnec, M.O.; Breed, M.D. Identification of Undecane as an Alarm Pheromone of the Ant Formica Argentea. Journal of Insect Behavior 2013, 26, 101–108. [Google Scholar] [CrossRef]
- Xu, W.; Zhao, M.; Tang, L.; Ma, R.; He, H. The Chemical Components of Dufour’s and Venom Gland of Camponotus japonicus (Hymenoptera, Formicidae). Research Square 2022, 1–12. [Google Scholar] [CrossRef]
- Wilson, E.O.; Pavan, M. Glandular Sources and Specificity of Some Chemical Releasers of Social Behavior in Dolichoderine Ants. Psyche: A Journal of Entomology 1959, 66, 70–76. [Google Scholar] [CrossRef]
- Do Nascimento, R.R.; Jackson, B.D.; Morgan, E.D.; Clark, W.H.; Blom, P.E. Chemical Secretions of Two Sympatric Harvester Ants, Pogonomyrmex salinus and Messor lobognathus. Journal of Chemical Ecology 1993, 19, 1993–2005. [Google Scholar] [CrossRef]
- Adams, R.M.M.; Wells, R.L.; Yanoviak, S.P.; Frost, C.J.; Fox, E.G.P. Interspecific Eavesdropping on Ant Chemical Communication. Frontiers in Ecology and Evolution 2020, 8. [Google Scholar] [CrossRef]
- Chen, J. Freeze–Thaw Sample Preparation Method Improves Detection of Volatile Compounds in Insects Using Headspace Solid-Phase Microextraction. Analytical Chemistry 2017, 89, 8366–8371. [Google Scholar] [CrossRef] [PubMed]
- Azhagu Raj, R.; Sathish, R.; Prakasam, A.; Krishnamoorthy, D.; Balachandar, M.; Majesh, T. Extraction and Analysis of Cuticular Hydrocarbons in the Weaver Ant Oecophylla smaragdina (Fabricius) (Hymenoptera: Formicidae). Int. J. Fauna Biol. Stud. 2017, 4, 102–107. [Google Scholar]
- Mashaly, A.M.A.; Ahmed, A.M.; Nunes, T.M.; Morgan, E.D. Secretions of Dufour’s Gland in Some Ants (Hymenoptera: Formicidae). African Entomology 2014, 22, 779–782. [Google Scholar] [CrossRef]
Figure 1.
This is a figure. Schemes follow the same formatting.
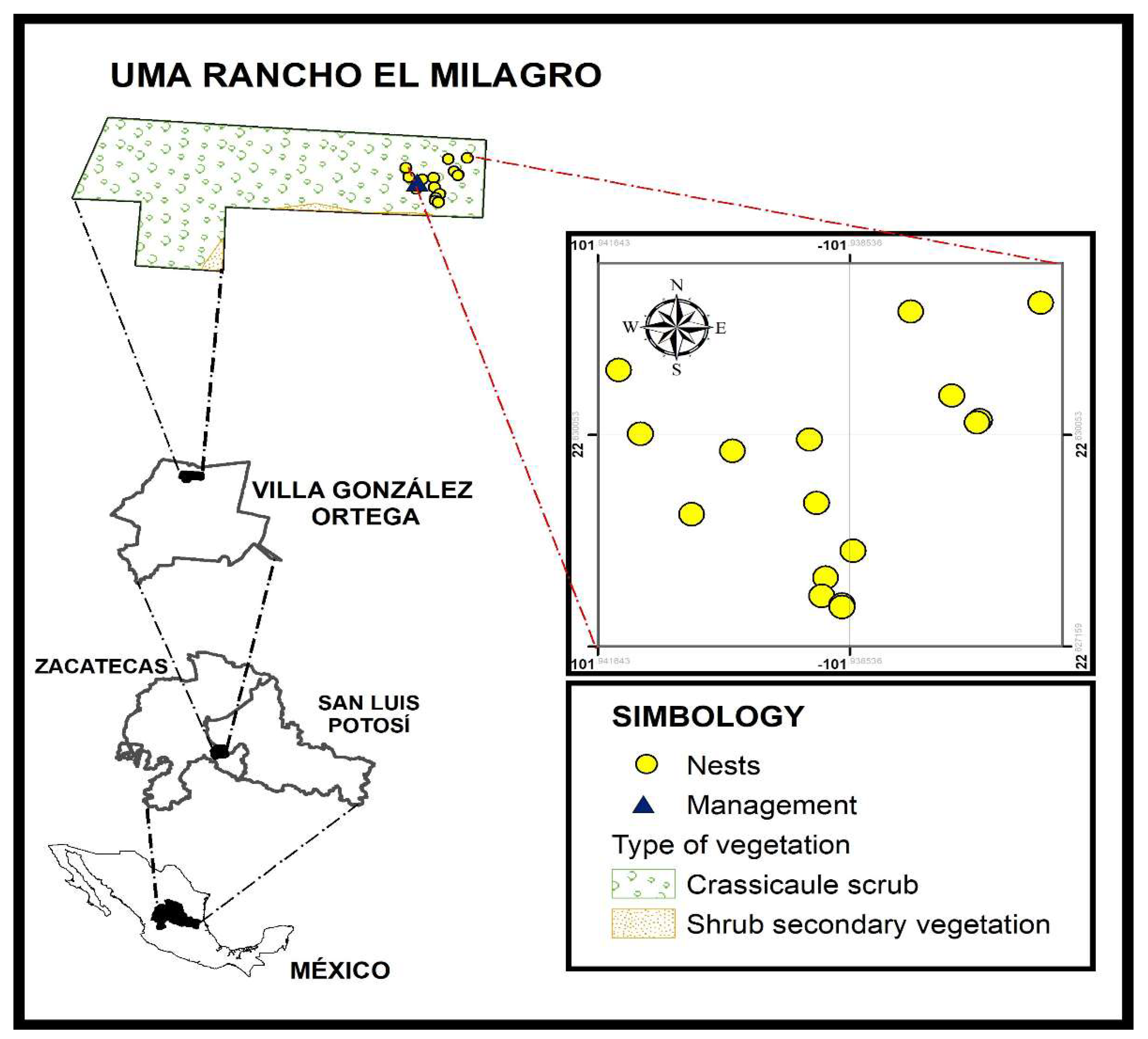
Figure 2.
Partial Least Squares-Discriminant Analysis (PLS-DA) (A); Plot of Variable Importance in Projection (VIP) (B).
Figure 2.
Partial Least Squares-Discriminant Analysis (PLS-DA) (A); Plot of Variable Importance in Projection (VIP) (B).
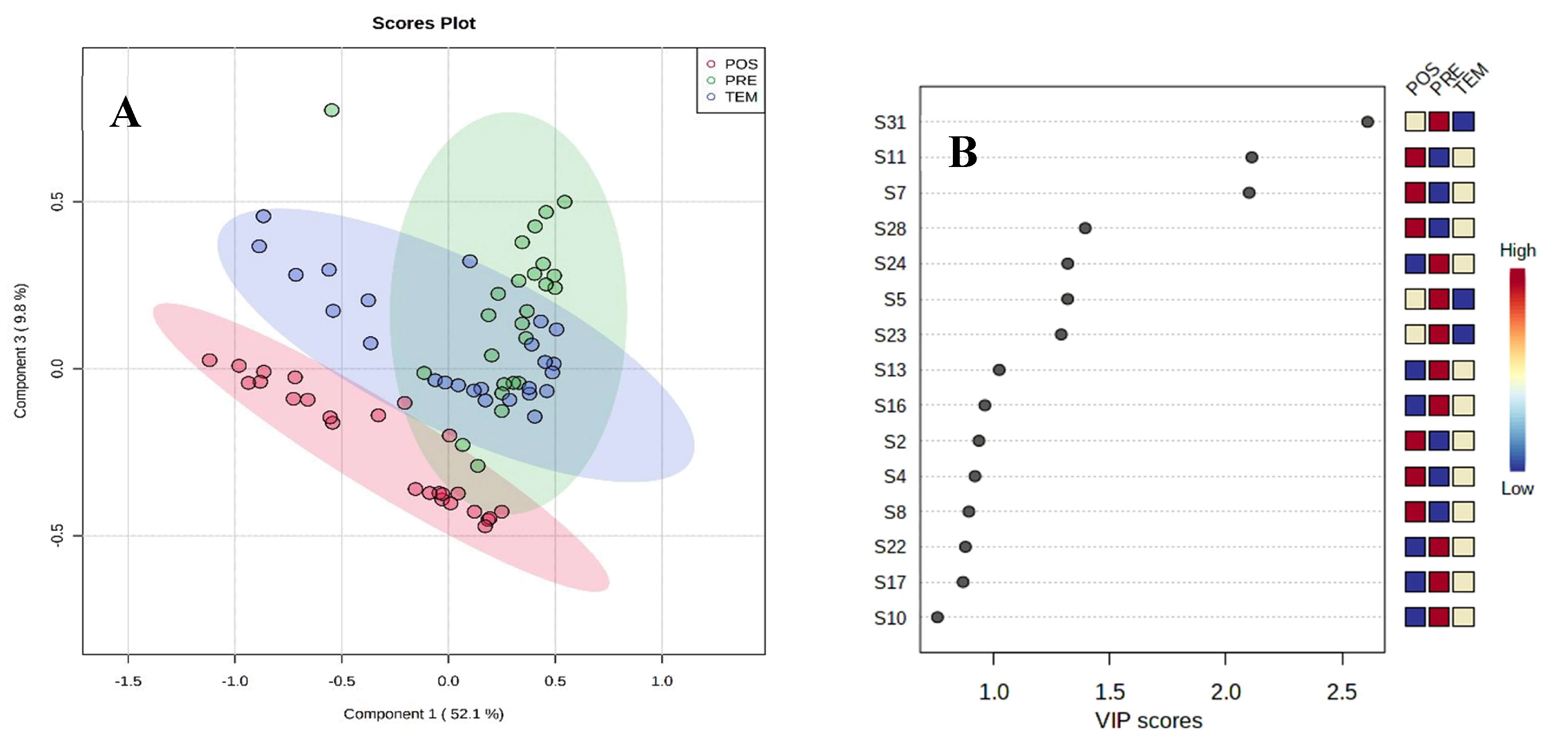
Figure 3.
Chromatograms in nests of L. apiculatum analyzed by HS-SPME-GC–MS. Five nests in PRE (A); Five nests in TEM (B); Five nests in POS (C).
Figure 3.
Chromatograms in nests of L. apiculatum analyzed by HS-SPME-GC–MS. Five nests in PRE (A); Five nests in TEM (B); Five nests in POS (C).
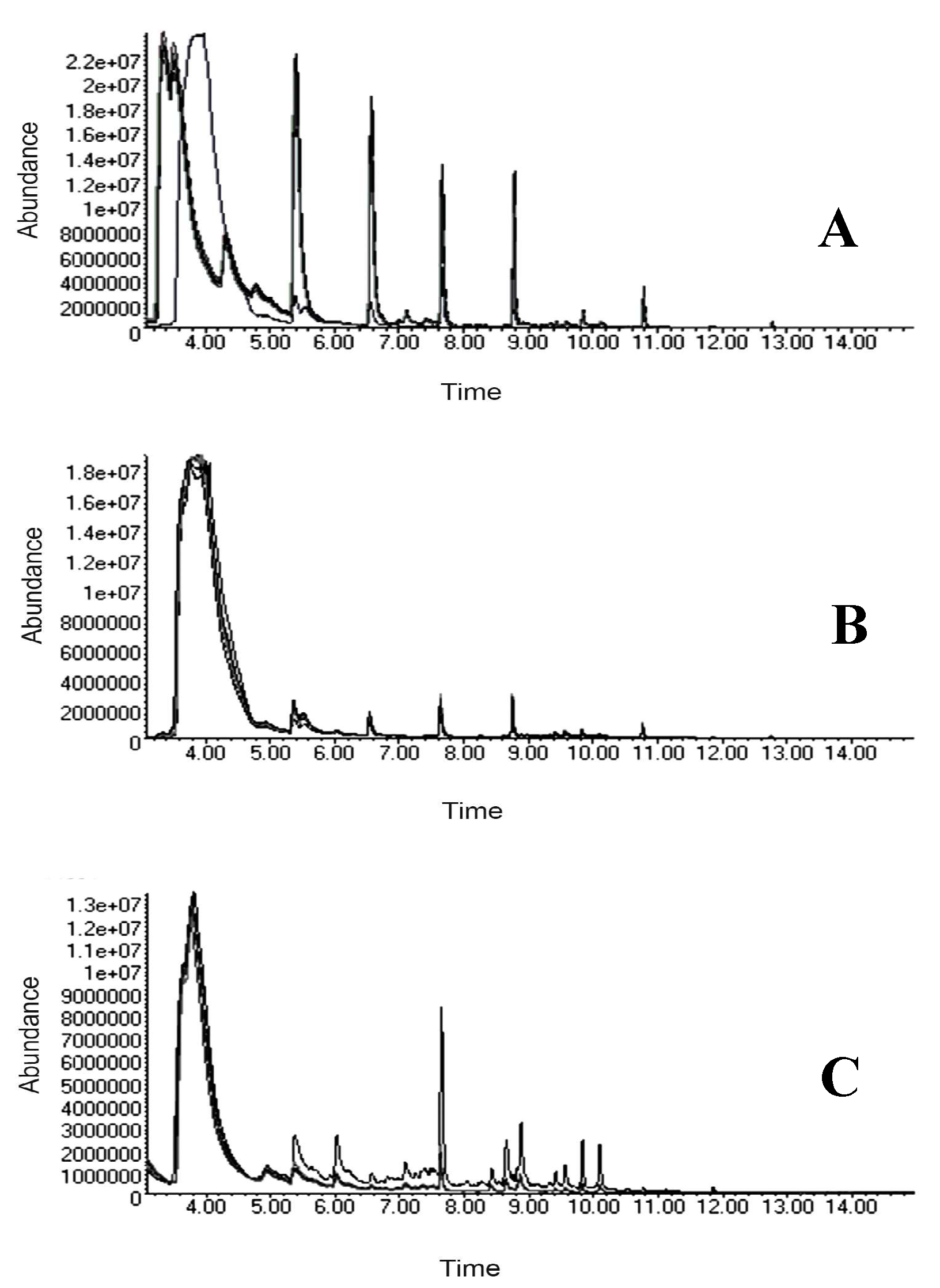
Figure 4.
Principal Component Analysis (PCA) of the semiochemicals by season.
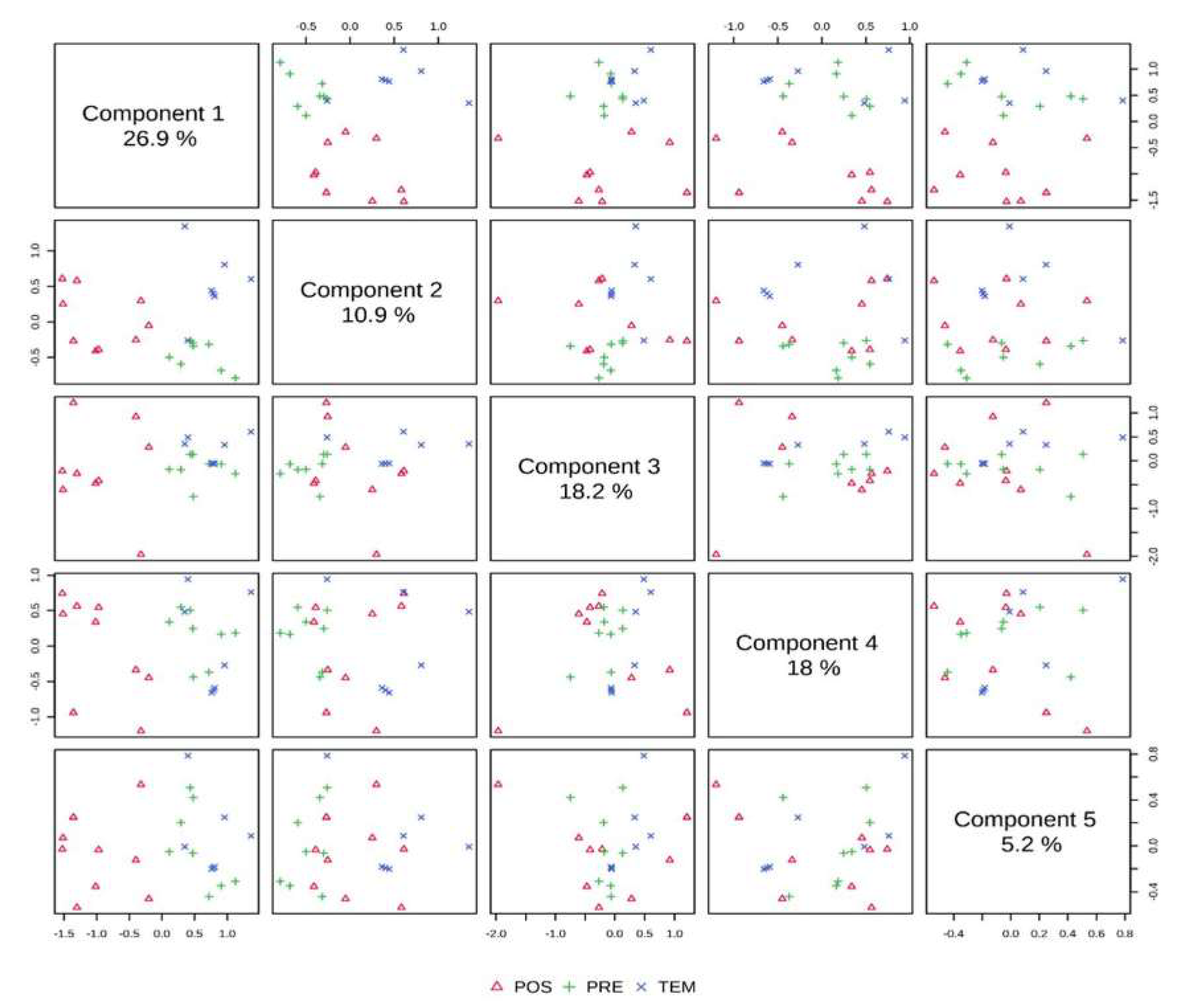
Figure 5.
Partial Least Squares-Discriminant Analysis (PLS-DA) (A); Plot of Variable Importance in Projection (B).
Figure 5.
Partial Least Squares-Discriminant Analysis (PLS-DA) (A); Plot of Variable Importance in Projection (B).
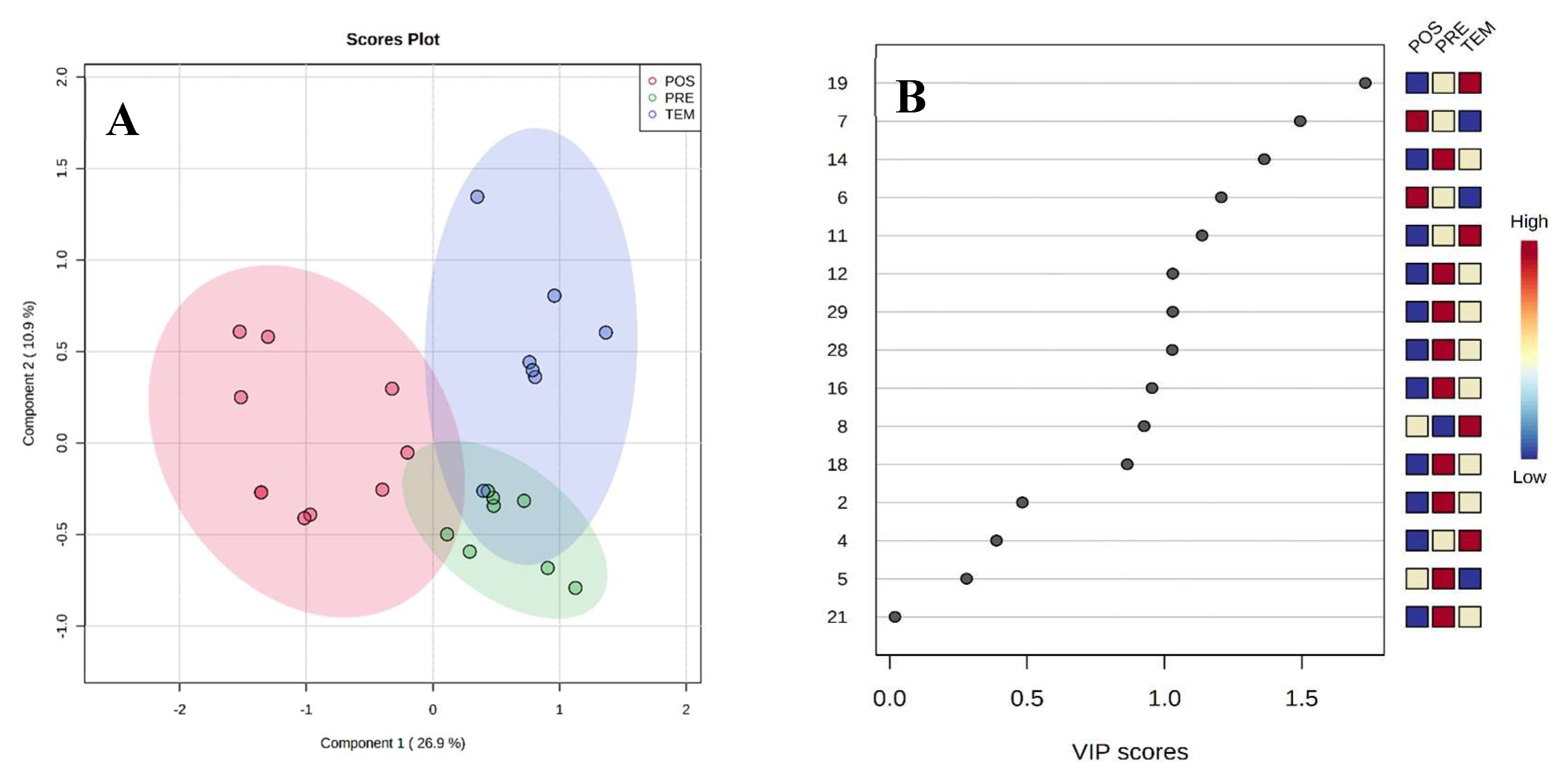
Figure 6.
Transit of ants on the artificial trails by nests and seasons.
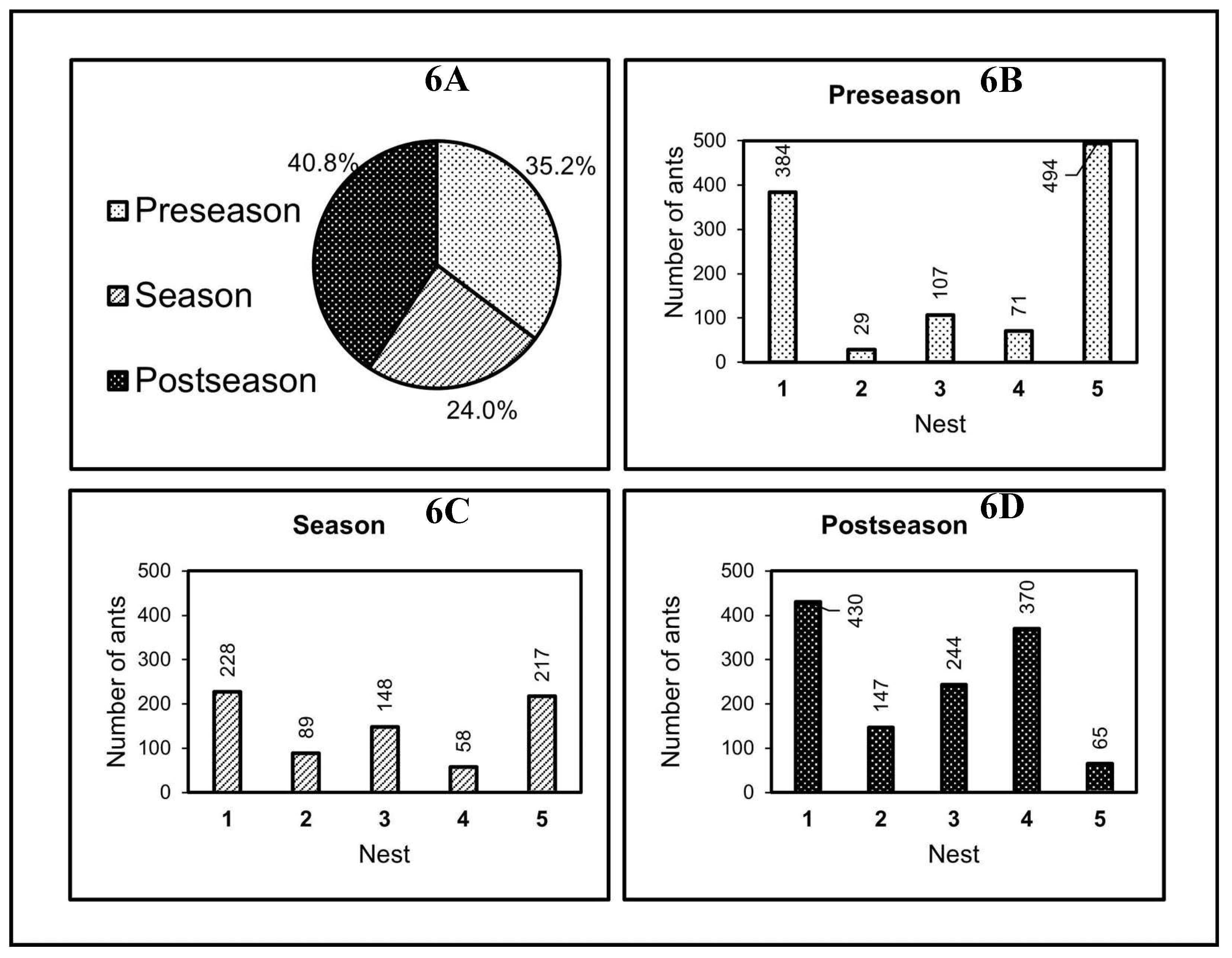
Figure 7.
Summary of semiochemical effects and test of the complete model.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



