
Submitted:
31 August 2023
Posted:
04 September 2023
You are already at the latest version
Abstract
Sporothrix brasiliensis is the most pathogenic species, responsible for the Brazilian cat-transmitted sporotrichosis hyperendemics. In this scenario, investigation of pathogen-host interaction can provide relevant information for future treatment strategies. To this end, the invertebrate Galleria mellonella has proven to be a suitable alternative to evaluate the virulence of pathogenic fungi since the insect immune system is similar to the mammalian innate immune response. The aim of this work was to investigate phenotypic and molecular aspects of the immune response of G. mellonella throughout the S. brasiliensis infection. Hemocyte density and the evolution of the fungal load were evaluated. In parallel, RT- qPCR expression analysis of genes encoding antimicrobial peptides (Gallerimycin and Galiomycin) and stress management genes (C7 Contig 15362 and C8 Contig 1910) was conducted. Fungal load and hemocyte densities were proportionally increasing simultaneously to the deleterious morphological events and larvae mortality. Gallerimycin, C7 Contig 15362, and C8 Contig 19101 genes were positively regulated (p<0.05) at distinct moments of S. brasiliensis infection, characterizing a time-dependent and alternately modulated profile. Galiomycin gene expression remained unchanged. Our results contribute to the future proposal of potential alternative pathways to treat and, consequently, control S. brasiliensis zoonosis, a major public Health in Latin America.
Keywords:
Sporotrichosis
; Gene Expression
; Invertebrates
; Antimicrobial Peptides
; Zoonosis
; Immune response
1. Introduction
Sporotrichosis is an affection of great importance to public health since human infection can occur through direct contact with contaminated soil organic matter (sapronosis) or, more frequently, by the zoonotic route through the domestic feline [1,2]. In this last scenario, the incriminated fungus is Sporothrix brasiliensis, the highest virulent and pathogenic species of the Sporothrix genus, with strong evidence of drug resistance [3,4]. The S. brasiliensis-zoonotic transmitted sporotrichosis began as an outbreak in the late 1990s in Rio de Janeiro, Brazil, and currently is considered a nationwide hyperendemic [5,6] with reports of recent geographical expansion to other Latin American countries [7,8]. Recently, the first three cases of domestic feline-transmitted S. brasiliensis outside South America were described in the United Kingdom, Europe [9].
Itraconazole is the drug of choice for human and feline sporotrichosis [10,11]. However, the cat’s treatment remains a challenge for the veterinarian, given the limited number of antifungal agents, their high cost and adverse effects, and their commercial presentations. Moreover, little is known about the immune response triggered by the infected hosts, especially the domestic feline [1,11].
In this direction, the expansion of studies that aim to expand information related to the pathogen-host interaction is necessary. To this end, Galleria mellonella stands out as an invertebrate model for the investigation of the immune response triggered by distinct fungal pathogens such as yeast of the Candida genus [12,13] and Cryptococcus neoformans [14]. Among filamentous fungi, Aspergillus niger, Rhizopus spp. and Rhizomucor spp. have also been challenged against the invertebrate biomodel’s defense [15,16,17], as have also the dimorphic Paracoccidioides lutzii and Histoplasma capsulatum [18,19]. However, few studies were dedicated to the interaction between G. mellonella and fungi of the Sporothrix genus, all focusing on phenotypic aspects to investigate the host-pathogen interaction [20,21,22] or in vivo drug-response [23].
The G. mellonella immune system has close both structural and functional similarities to the mammalian innate immune system [24,25], and is divided into two strongly interconnected pathways: the cell-mediated pathway and the humoral pathway [26]. The cellular response of insects is mediated by hemocytes, phagocytic cells in the hemolymph. These have the function of phagocytosing foreign bodies, capturing and encapsulating them in multicellular structures called nodules or capsules [26,27,28]; playing roles analogous to those of human macrophages and neutrophils [24,25]. In parallel, the humoral response of insects includes antimicrobial peptides (AMPs), which act directly against invaders. Among the G. mellonella AMPs, gallerimycin and galiomycin are the most well-described in the literature [29], with evidence of positive regulation on gene expression in fungal infections, such as Candida spp. and Aspergillus niger by this insect [17,30,31].
In a complementary investigation, other parameters such as the role of stress-managing genes can be evaluated during the immune response of the larvae against pathogens [30]. Among the best-characterized ones are those responsible for the regulation of cytokines (C7 Contig 15362), those linked to the phagocytosis and related to protein binding (C8 Contig 19101) and those involved in inflammatory responses (C3- Contig 15265, C4- Contig 290595, C5- Contig 21310, C6- Contig 1327) [30,32,33].
Data about the cellular and humoral insect defenses during the infectious process can provide new information on the host-pathogen interaction. Furthermore, elucidating mechanisms of S. brasiliensis infection in the invertebrate model may contribute to the future detection of new therapeutic targets as well as to the description of promising molecules to control this mycosis. Thus, the objective of the present study was to monitor G. mellonella's phenotypic and genotypic responses against S. brasiliensis. To the best of our knowledge, the present study is the first to evaluate the defense-related gene expression of G. mellonella in response to infection by a dimorphic fungus.
2. Materials and Methods
2.1. Strains and Culture Conditions
The reference strain of Sporothrix brasiliensis used in this work was the American Type Culture Collection isolate, ATCC MYA 4823. The yeast cells were maintained by cryopreservation at -80 °C in the mycotheque of the Center for Microorganisms' Investigation (CIM-UFF), until their reactivation for the experiments. For this purpose, the cryotubes were defrosted with subsequent replicas in YPD medium (Yeast Extract Peptone Dextrose), with incubation under shaking at 37°C for 5 days. The microbial growth in the broth was then centrifuged at 2000 xg for 5 minutes, washed with sterile PBS (Phosphate Buffered Saline; Fujifilm Irvine Scientific), and suspended in the same buffer. After these processes, standardized suspensions of 1x107 yeast/larvae were prepared using a hemocytometer [20,21].
2.2. Galleria mellonella Survival Assays
The invertebrate biomodel investigated was kindly provided from a well-established colony by the Biofilms and Microbial Diversity Laboratory of the Federal University of Rio Grande do Sul. Survival curves were performed with larvae in the final larval stage of development, weighing between 0.2g and 0.3g, with uniform coloration, without spots or signs of melanization [34]. Inoculation of the fungus was subcutaneous, with 10 μL of the inoculum applied in PBS with a Hamilton syringe (701N, Caliber 26, Hamilton Company Reno, USA) in the last left proleg, as recommended [21,34]. Infected larvae were grouped by experimental condition, kept at 37º C in 14 cm Petri dishes, and monitored daily for survival. Lack of movement and extensive melanization of the body were both taken as indicators of animal death. As a control, two groups of animals were selected, one group with only the physical injury of the needle (naive) and the other injected with PBS, to assess mortality from animal handling and mechanical injury. Each group, including the control, consisted of 30 larvae.
2.3. Determination of fungal burden during the survival curve
To monitor the fungal load during the different phases of infection, an experiment with ten (10) inoculated larvae was conducted with the same inoculum (1x107 yeast/larvae) and under the same temperature conditions, in triplicate, as detailed earlier in the survival curve. Daily, one larva was randomly selected from the pool, which was previously cleaned with ethanol 70% and then the hemolymph was collected. For this purpose, the larvae were sectioned with a sterile scalpel on the lower part and 30 μL of hemolymph were collected from each individual, placed in an Eppendorf® tube containing physiological saline buffer for insects (IPS: 150 mM sodium chloride, 5 mM potassium chloride, 10 mM Tris-HCl, pH 6.9, 10 mM EDTA and 30 mM sodium citrate) in a 1:10 proportion (35). Soon after, serial dilutions were made on Sabouraud agar plates plus Chloramphenicol (Sigma-Aldrich, St. Louis, Missouri, USA), incubated for seven days at 25ºC. After this period, one of the colonies was chosen for observation of confirmatory micromorphology of the genus Sporothrix using methyl blue staining. Once the genus was detected, all colony forming units (CFUs) were counted as described by Clavijo-Giraldo and colleagues [21].
2.4. Determination of hemocyte density in hemolymph
Similar to the methodology used for hemolymph collection in the previous item, hemocyte density was estimated from 30 μL of hemolymph diluted in IPS in a 1:10 proportion. The number of hemocytes was counted using a hemocytometer [36]. The experiment was performed in triplicate and the results were expressed in hemocytes/mL. In parallel to the quantitative analysis, qualitative notes of phenomena such as nodule formation as well as yeast adhered to the hemocytes were made.
2.5. Analysis of gene expression
2.5.1. RNA extraction and cDNA sythesis
For the RNA extraction procedure, 20 control larvae with only the physical injury and 20 larvae inoculated with 1x107 S. brasiliensis/larvae were selected. The assays to investigate the dynamics of gene expression were performed at three moments of the survival curve: on days one, five, and eight (D1, D5, and D8) after infection by S. brasiliensis. For each incubation time (D), six (06) larvae from each group were randomly selected, cooled on dry ice, and conditioned in microtubes for total RNA extraction from the tissue. For this purpose, the larvae were macerated using a sterile pestle, and the extraction was performed with SV Total RNA Isolation System kit (Promega Corporation, Madison, WI, USA), following the manufacturer's recommendations. The total RNA was transcribed into complementary DNA (cDNA) using GoScript Reverse Transcription System Kit (Promega Corporation, Madison, WI, USA), according to the manufacturer's instructions. In the cDNA synthesis, besides the reverse transcriptase, the DNase enzyme was also used to eliminate possible contaminating DNA in the sample. The cDNA was then quantified in the Quantus Fluorometer (Promega Corporation, Madison, WI, USA) using the QuantiFluor One dsDNA System kit (Promega Corporation, Madison, WI, USA). Finally, samples were diluted to the final use concentration of 2.5 ng/μl.
2.5.2. Reverse Transcription-quantitative Polymerase Chain Reaction (RT-qPCR)
Quantification of gene expression was performed by RT-qPCR using the Power SYBR™ Green PCR Master Mix (Thermo Fisher Scientific, Carlsbad, CA). The gene expression levels related to the production of Gallerimycin and Galiomycin AMPs in addition to C7 Contig 15362 and C8 Contig 19101 genes were evaluated at the three time points of infection. The gene β-actin was used as a normalizing reference control to monitor the amount of a housekeeping gene not involved in immune response, using the same cDNA preparations. The primer sequences used are specified in Table 1. The group of non-inoculated larvae was used as a control, which was compared with the group inoculated with the fungal pathogen. The samples and controls were tested in triplicate and all reactions were performed in 96-well plates with a final volume of 15 μl, being: 7.5 μl of Power SYBR™ Green PCR Master Mix, 300 nM of each primer, 7.5 ng of cDNA and DEPEC water (InvitrogenTM, Carlsbad, CA, USA) to complete the volume.
The reaction was performed in the 7500 Fast Real-Time PCR System (Applied Biosystems, Framingham, MA, USA) with the following conditions: initial heating at 50ºC for 2 minutes, denaturation at 95ºC for 2 minutes, followed by 40 cycles of denaturation at 95ºC for 15 seconds, annealing and extension at temperatures ranging from 55º-60ºC for 1 minute, depending on each primer (Table 1). In the end, a melting curve was performed to verify the specificity of the reaction - 95ºC for 15 seconds, 60º for 1 minute, followed by 30 seconds at 95ºC. The mean of the threshold cycle (cT) values, measured in triplicate, were used to calculate the expression of the target genes. The results were obtained as relative gene expression values (based on the formula: 2-ΔΔCT) compared to the reference gene expression, the β-actin, resulting in a value equal to 1.
2.6. Statistical Analyses
Survival analyses of G. mellonella infected with fungal pathogens were estimated using the Kaplan-Meier survival curve, in which the log-rank test was used to compare the groups in terms of survival. The evolution of fungal load over days and the relationship between hemocyte count and CFU were analyzed using simple linear regression models. Adherence to the Normal distribution of the variables was done using the Shapiro-Wilks test. Kruskal Wallis test and Dunn's post hoc test were used to check the dynamics of gene expression as a function of day for each experimental group. The Mann-Whitney test was used to compare the relative quantification of genes on days D1, D5, and D8, as a function of the non-inoculated and inoculated groups. The significance level adopted was 5%, and the software used was R, version 4.1.2.
3. Results
3.1. Galleria mellonella Survival Assays
The survival curve showed that larvae inoculated with 1x107 yeast of S. brasiliensis started to die on day 4, while the majority of all naive larvae and those inoculated with PBS only remained unaltered (p < 0.001; Kaplan-Meier), reaching the pupal stage in the last days of the curve. The results are represented in Figure 1A.
3.2. Fungal load during the infection
Parallel to the survival curve, daily collection followed by hemolymph serial dilutions and subsequent plating were performed to obtain CFUs. Colonies whose macromorphology was compatible with the genus Sporothrix were further evaluated by micromorphological analyses, evidencing thin, hyaline, septate, branched hyphae, and conidia arranged in flower-like structures. Figure 1B shows the evolution of the fungal load along the survival curve, with increasing values from 6x103 up to 4x105 CFU/larva (x̅ = 1,01 x105; σ ±1.4x105). A direct correlation between the increase of S. brasiliensis yeasts over the days was observed (p= 0.0157).
3.3. Sporothrix brasiliensis-Galleria mellonella Interaction
Daily hemocyte counting allowed the detection of increasing insect cellular defense generated by the fungus, verified by values from 6.4x106 up to 3.0 x107 hemocytes/mL along the curve (x̅ = 1,28 x107; σ ± 7.6 x106). The hemocyte count of the larvae from control groups (naive and PBS) was performed until D5 since afterward, both began to develop into pupae. In contrast, larvae infected with S. brasiliensis showed slower development in the applied experimental conditions (Figure 1C).
The hemocyte count associated with the determination of the fungal load along the survival curve allowed the evaluation of the Sporothrix brasiliensis-Galleria mellonella interaction dynamics. Progression of infection by the fungal pathogen triggered significant, proportional growth in insect hemocyte density (p = 0.0019; r2 = 0.823; Figure 1D).
3.4. Gene Expression
3.4.1. Antimicrobial peptides
For the gene encoding the AMP Gallerimycin dynamics, an early higher expression was observed, since the first day after infection (D1; Figure 2A), compared to naive larvae (p<0.0001; Mann-Whitney test). The higher expression in S. brasiliensis infected larvae remained on the subsequent days D5 (p> 0.05) and D8 (p = 0.0011; Mann-Whitney test). In parallel, comparing the intragroup Gallerimycin gene expression, a significant reduction was observed in the infected larvae (p = 0.0055; Kruskal-Wallis test), contrarily from the observed for the control group (p = 0.9968; Figure S2A; Supplementary).
Differently, the quantification of the expression of the gene encoding the Galiomycin AMP revealed no significant differences between the two experimental groups, as well as after the intragroup analysis (p>0.05; Mann-Whitney test; Figure 2B and Figure S2B).
3.4.2. Stress managing genes
The C7 Contig 15362 mRNA expression analysis differs between the investigated groups at D5 post-inoculation (p<0.0001; Mann-Whitney test). After this period, it decreases and at D8 shows its lowest values (Figure 2C). Analysis of the intragroup gene expression showed a significant difference between the three evaluated periods (p = 0.0024; Kruskal-Wallis test; Figure S2C), with higher levels of mRNA production at D5. This difference was not observed in non-infected larvae between the evaluated days (p> 0.05; Supplementary).
The expression of C8 Contig 19101 showed an initial, non-significant increase in mRNA expression, since the first day after S. brasiliensis inoculation (D1; p>0.05). However, at D5 an important increase in C8 Contig 19101 gene expression was detected compared to naive larvae (Figure 2D). In the intragroup comparison, C8 Contig 19101 mRNA expression did not vary significantly between D1 x D5 x D8 (p>0.05; Kruskall-Wallis test; Figure S2D).
4. Discussion
Although sporotrichosis is a disease of great importance in public health, numerous aspects of the pathogen-host relationship are yet to be elucidated, mainly for the main host of this zoonosis, the domestic feline [1,39]. As a matter of fact, such an investigation requires an appropriate experimental model. For decades, the murine model has been used as the in vivo gold standard model for pathogenicity mechanisms of distinct microorganisms [40], including Sporothrix spp [41]. However, recently, the scientific community has been showing ethical and social concerns, applied to the rationalization of animal model use [29,42,43].
Therefore, aiming at obtaining an alternative approach to mammalian models, several invertebrate models of infection have been studied, with emphasis on Galleria mellonella [44,45]. In spite of the previous use of this insect for the investigation of different fungal and bacterial infections [19,46], studies concerning the Sporothrix genus pathogenicity are still limited. As a matter of fact, only four previous studies investigated S. brasiliensis-G. mellonella binomial, based on the description of phenotypic aspects [20,21,22,23]. Therefore, this is the first study to draw a parallel between G. mellonella inate immunity-related gene expression and deleterious events during Sporothrix infection.
The results obtained on the survival curve of S. brasiliensis infection in G. mellonella standardization showed that the ideal yeast inoculum (1x107 yeast/larva) and temperature (37°C) were partially similar to those described by Clavijo-Giraldo and collaborators [21]. For more efficient killing by the fungal pathogen, these authors concluded that temperatures closer to the natural mammalian host are required to maintain all virulence attributes expressed by the yeast morphology. However, a less concentrated inoculum of 1x105 yeast/larvae was sufficient for these authors to obtain an appropriate survival curve. It is believed that this difference may be related mainly to the lineage of the larva used or even uncontrolled environmental conditions. On the other hand, Freitas and co-authors [20] used the same fungal load of 1x107 yeast/larva, since when testing lower concentrations (1x104 yeast/larva and 1x106 yeast/larva), they did not observe mortality in the survival curve. In addition, the temperature of 37°C was also recommended for the experiment, corroborating in both aspects with the results obtained here.
Daily monitoring of S. brasiliensis load during the infection curve provided progressive increasing values ranging from 1.2x104 to 4x105 CFU per animal. An average fungal load ranging from 1.8x105 to 2.4x105 CFU per animal was previously described [22] after 24 hours of infection. Gandra and colleagues [31] observed that by inoculating 1x107 Candida albicans/larvae, CFUs ranged from 1.0x108 to 1.0x1012 at 6, 24, and 48 hours. These data reinforce the suitability of G. mellonella as a model of yeast infection since it differentially mirrors Candida and Sporothrix fungal loads increase, such as previously described for mammalian hosts [47,48].
It is noteworthy to mention the proportional increase of hemocytes in response to S. brasiliensis's higher fungal loads, beginning on the fifth day of the curve. As a matter of fact, on the fifth day the major phenotypic event registered was the beginning of larvae death. Thus, it is possible to hypothesize that larvae morbidity is a result of higher fungal loads as the immune system, although proportionally stimulated reaching greater hemocyte recruitment, fails [26,49].
Lozoya-Pérez and collaborators [22] described a similar S. brasiliensis-G. mellonella approach, at a single-specific point of the survival curve. CFU and hemocyte counting from the insect's hemolymph were obtained 24 hours after yeast inoculation, previously grown in distinct culture media. CFU values between 1.8x105 and 2.4x105 were found and 3.9 and 9.2 ×106 hemocytes/mL, depending on the culture medium used. Such values were lower than those found in the present study (6.4x106 to 3.0x107 hemocytes/mL). This difference may be related to the 100x higher S. brasiliensis inoculated in the present study protocol.
In spite of the increasing amount of literature involving the use of G. mellonella as an invertebrate biomodel, little is known about its defense strategies against S. brasiliensis. Even though the present study was able to establish a comparison with the previous work concerning the phenotypic events of the Sporothrix-G. mellonella interaction [21,22,50] we are limited to few previous publications approaching the molecular aspects of the S. schenckii infection [50]. Actually, gene expression data in such experimental scenario is limited to the genus Candida [31,44].
To the best of our knowledge this is the first study to investigate G. mellonella phenotypic and molecular events triggered by the infection with the most virulent Sporothrix species, S. brasiliensis. In contrast, a considerable number of published works is available on the G. mellonella humoral response to Candida species. These quantify the expression of AMPs promoter genes with distinct purposes, among them the description of the larval immune response profiles under different infection protocols and the evaluation of potential antimicrobial drugs. By respecting the differences inherent to each fungal species, these findings were used as parameters in order to discuss the present study findings.
Given the previous works highlighting the importance of the G. mellonella AMPs encoding genes Gallerimycin and Galiomycin, during both filamentous and yeast fungal infections [29,31] this study seeked to quantify these genes expression after the infection by the dimorphic fungus Sporothrix brasiliensis. While Gallerimycin presented an expressive increase in gene expression along the survival curve, Galiomycin remained inaltered. These results suggest that the AMP Gallerimycin plays a relevant role in the insect’s response to S. brasiliensis infection, as previously described during Candida spp. infection [31,51]. Contrarily, Dekkerová-Chupáčová and co-authors [12] observed that the inoculation of 2x105 yeast/larvae of C. albicans and C. dubliniensis into G. mellonella larvae triggered an expressive increase in AMP coding genes, especially Galiomycin.
The differential relevance of the G. mellonella Galiomycin gene expression during Candida albicans and dubliniensis versus S. brasiliensis infections may be explained by the fact that the first is a well know microbiota yeast member interacting with vertebrates during millions of years of evolution while Sporothrix species evolved from saprophytic mycelial fungi recently adapted to parasitism by termodimorphism. Therefore, Sporothrix species may be in the process of gene expression adapting to the animal organic matter [2,52].
In addition to positive regulation, time-dependent regulation was observed, and Gallerimycin evidenced a significant increase on days D1 and D8. Such a result partially corroborates with that found by Dekkerová-Chupáčová and co-authors [12]. They noted that the maximum up-regulation of both genes was shown at 24 h post-infection, but as early as one hour into the infection, a positive regulation of expression was already observed. More specifically, the gene encoding Gallerimycin was up to 1.3-fold higher (1 h post-infection) and 3.2-fold higher (24 h post-infection) in larvae infected with C. albicans compared to C. dubliniensis. Whereas for Galiomycin we observed up to 3.7-fold (1 h post-infection) and 7.1-fold (24 h post-infection) relative up-regulation in C. albicans-infected larvae. The up-regulation of both genes decreased at 48 h post-infection by these two species.
In this regard, the difference in gene expression analysis intervals was based on the metabolic characteristics of the fungal species. Candida spp. demonstrates considerably faster growth both in vitro and in vivo, in the invertebrate model itself, compared to Sporothrix spp. [13]. Moreover, given the scarcity of information regarding the gene expression of G. mellonella against Sporothrix spp., the choice of day (D) for gene expression analysis was defined according to the defense phenomena of the larvae observed in response to the fungus, such as melanization, mobility changes, and the onset of mortality.
Concerning the results observed in the expression of stress management genes, C7 Contig 15362 and C8 Contig 19101 showed a significant difference in gene expression between the experimental groups on the fifth day (D5) of infection. Drawing a parallel with the study conducted by Melo and collaborators [30] evaluating G. mellonella gene expression after C. albicans infection versus different antifungal therapies, in 24 and 48 hours, it was also possible to observe a positive regulation of both genes in the infected, untreated larvae. A relevant point is that in this case C8 Contig 19101, was about 4 times overexpressed than C7 Contig 15362 gene.
In the present study, it was possible to observe that the expression of the genes studied is not only time-dependent but also alternately modulated. Since, when comparing the genes responsible for encoding AMPs with those of stress management, this significant increase D-day varied. C7 Contig 15362 and C8 Contig 19101 are considered stress management genes and showed a significant increase in expression on day 5 post-infection. Interestingly, this moment coincides with the onset of mortality and deleterious phenotypic changes observed during the survival curve, as well as with the beginning of the expressive increase in defense cells and fungal load. In contrast, genes encoding AMPs, especially Gallerimycin, showed an immediate response verified by the intense mRNA encoding as early as the first post-infection day. This finding is compatible with the participation of the AMPs, previously characterized as an early element of humoral immunity against the infectious process [53,54]. For this same fact, different authors determine evaluation protocols in hours, both in the investigation of the infectious process by Candida spp. [12,31,51] and by Aspergillus niger [17].
This work is the first to evaluate the gene expression related to the defense of the insect G. mellonella in response to infection by a dimorphic fungus. Most authors have dedicated themselves to the investigation of yeasts and, among these, of the genus Candida [12,30,31]. Among the filamentous fungi, previous investigation considered Aspergillus niger [17] and Fusarium oxysporum [55].
The possibility that filamentations occurred during the infectious process in G. mellonella cannot be excluded. Thus, the comparative discussion of some of the parameters evaluated here is limited. It is important to add the fact that previously the presence of hyphae of S. brasiliensis has been reported, both in vitro in feline phagocytes [56] and either during human or animal parasitism (manuscript in preparation). It is known that the fungal cell wall undergoes profound transformation altering, during the dimorphic transition, β-1-3-glucans to α-glucans posing an immediate challenge to the host immune response (PAMPs) [17,57]. However, only studies that consider distinct periods within the survival curve will be able to answer whether the verified gene expression was exclusively yeast-triggered and or yeast-targeted.
Another acknowledged limitation is the absence of data regarding gene expression in the intervals between the defined days. However, because this is a pioneer investigation in the area, new perspectives and methodologies can be generated and improved in order to overcome the scarcity of data in the literature and elucidate the aspects of this relevant pathogen-host interaction. The present work opens new doors of investigation on the pathogenesis by Sporothrix spp. as well as offers a better characterized model for the research of potential antifungal drugs effective for the control of this important zoonosis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, N.F.R, R.L.D.M, H.M.M-M. and A.R.S.B; Methodology, N.F.R, M.C.S.J, J.A.C.R. and L.C.S.V.S; Software, J.R.S.S and M.C.S.J.; Validation, N.F.R, L.M.A and H.M.M-M.; Formal Analysis, P.A.P., C.S.V, H.M.M-M., R.L.D.M.; Investigation, N.F.R., M.C.S.J., J.A.C.R. and S.C.P.B; Resources, A.R.S.B, P.A.P. and R.L.D.M.; Data Curation, N.F.R, M.C.S.J and A.R.S.B; Writing – Original Draft Preparation, N.F.R, M.C.S.J. and A.R.S.B; Writing – Review & Editing, A.R.S.B, B.A.P and H.M.M-M.; Visualization, H.M.M-M.; Supervision, A.R.S.B, B.A.P, P.A.P and C.S.V; Project Administration, A.R.S.B and R.L.D.M; Funding Acquisition, A.R.S.B, B.A.P and R.L.D.M. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by grants from the Brazilian Agencies: Conselho Nacional de Desenvolvimento Científico e Tecnológico (PIBIC-CNPq-UFF, Brazil); Fundação de Amparo à Pesquisa no Estado do Rio de Janeiro, Brazil (FAPERJ E-26/211.502/2021), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brazil (CAPES)—Financial Code 001, Fopesq/Proppi-UFF and Rede Micologia RJ. R.L.D.M. and A.R.S.B. are research fellows of CNPq (PQ-CNPq).
Acknowledgments
The authors wish to express their gratitude to Professor Alexandre Macedo of the Federal University of Rio Grande do Sul for donating Galleria mellonella eggs and to PhD Laura Nunes, for technical advice. The authors would also like to thank Centro Integrado de Tradução e Escrita (CITE/UFF) for assistance with English language translation and developmental editing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gremião, I. D. F.; Miranda, L. H. M.; Reis, E. G.; Rodrigues, A. M.; Pereira, S. A. Zoonotic Epidemic of Sporotrichosis: Cat to Human Transmission. PLoS Pathog 2017, 13, e1006077. [Google Scholar] [CrossRef]
- Rodrigues, A. M.; Della Terra, P. P.; Gremião, I. D.; Pereira, S. A.; Orofino-Costa, R.; de Camargo, Z. P. The Threat of Emerging and Re-Emerging Pathogenic Sporothrix Species. Mycopathologia 2020, 185, 813–842. [Google Scholar] [CrossRef]
- Della Terra, P. P.; Rodrigues, A. M.; Fernandes, G. F.; Nishikaku, A. S.; Burger, E.; de Camargo, Z. P. Exploring Virulence and Immunogenicity in the Emerging Pathogen Sporothrix brasiliensis. PLoS Negl Trop Dis 2017, 11, e0005903. [Google Scholar] [CrossRef]
- Nakasu, C. C. T.; Waller, S. B.; Ripoll, M. K.; Ferreira, M. R. A.; Conceição, F. R.; Gomes, A. dos R.; Osório, L. da G.; de Faria, R. O.; Cleff, M. B. Feline Sporotrichosis: A Case Series of Itraconazole-Resistant Sporothrix brasiliensis Infection. Braz J Microbiol 2021, 52, 163–171. [Google Scholar] [CrossRef]
- Gremião, I. D. F.; Oliveira, M. M. E.; Monteiro de Miranda, L. H.; Saraiva Freitas, D. F.; Pereira, S. A. Geographic Expansion of Sporotrichosis, Brazil. Emerg. Infect. Dis. 2020, 26, 621–624. [Google Scholar] [CrossRef] [PubMed]
- Schechtman, R. C.; Falcão, E. M. M.; Carard, M.; García, M. S. C.; Mercado, D. S.; Hay, R. J. Sporotrichosis: Hyperendemic by Zoonotic Transmission, with Atypical Presentations, Hypersensitivity Reactions and Greater Severity. Anais Brasileiros de Dermatologia 2022, 97, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Etchecopaz, A.; Toscanini, M. A.; Gisbert, A.; Mas, J.; Scarpa, M.; Iovannitti, C. A.; Bendezú, K.; Nusblat, A. D.; Iachini, R.; Cuestas, M. L. Sporothrix brasiliensis: A Review of an Emerging South American Fungal Pathogen, Its Related Disease, Presentation and Spread in Argentina. JoF 2021, 7, 170. [Google Scholar] [CrossRef]
- Thomson, P.; González, C.; Blank, O.; Ramírez, V.; Río, C. del; Santibáñez, S.; Pena, P. Sporotrichosis Outbreak Due to Sporothrix brasiliensis in Domestic Cats in Magallanes, Chile: A One-Health-Approach Study. JoF 2023, 9, 226. [Google Scholar] [CrossRef] [PubMed]
- Barnacle, J. R.; Chow, Y. J.; Borman, A. M.; Wyllie, S.; Dominguez, V.; Russell, K.; Roberts, H.; Armstrong-James, D.; Whittington, A. M. The First Three Reported Cases of Sporothrix brasiliensis Cat-Transmitted Sporotrichosis Outside South America. Medical Mycology Case Reports 2023, 39, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Orofino-Costa, R.; Macedo, P. M. de; Rodrigues, A. M.; Bernardes-Engemann, A. R. Sporotrichosis: An Update on Epidemiology, Etiopathogenesis, Laboratory and Clinical Therapeutics. An. Bras. Dermatol. 2017, 92, 606–620. [Google Scholar] [CrossRef]
- Gremião, I. D. F.; Martins da Silva da Rocha, E.; Montenegro, H.; Carneiro, A. J. B.; Xavier, M. O.; de Farias, M. R.; Monti, F.; Mansho, W.; de Macedo Assunção Pereira, R. H.; Pereira, S. A.; Lopes-Bezerra, L. M. Guideline for the Management of Feline Sporotrichosis Caused by Sporothrix brasiliensis and Literature Revision. Braz J Microbiol 2021, 52, 107–124. [Google Scholar] [CrossRef]
- Dekkerová-Chupáčová, J.; Borghi, E.; Morace, G.; Bujdáková, H. Up-Regulation of Antimicrobial Peptides Gallerimycin and Galiomicin in Galleria mellonella Infected with Candida Yeasts Displaying Different Virulence Traits. Mycopathologia 2018, 183, 935–940. [Google Scholar] [CrossRef] [PubMed]
- García-Carnero, L. C.; Clavijo-Giraldo, D. M.; Gómez-Gaviria, M.; Lozoya-Pérez, N. E.; Tamez-Castrellón, A. K.; López-Ramírez, L. A.; Mora-Montes, H. M. Early Virulence Predictors during the Candida Species–Galleria mellonella Interaction. JoF 2020, 6, 152. [Google Scholar] [CrossRef]
- Trevijano-Contador, N.; Herrero-Fernández, I.; García-Barbazán, I.; Scorzoni, L.; Rueda, C.; Rossi, S. A.; García-Rodas, R.; Zaragoza, O. Cryptococcus neoformans Induces Antimicrobial Responses and Behaves as a Facultative Intracellular Pathogen in the Non Mammalian Model Galleria mellonella. Virulence 2015, 6, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, I. Animal Models to Study Mucormycosis. JoF 2019, 5, 27. [Google Scholar] [CrossRef]
- Maurer, E.; Hörtnagl, C.; Lackner, M.; Grässle, D.; Naschberger, V.; Moser, P.; Segal, E.; Semis, M.; Lass-Flörl, C.; Binder, U. Galleria mellonella as a Model System to Study Virulence Potential of Mucormycetes and Evaluation of Antifungal Treatment. Medical Mycology 2019, 57, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Stączek, S.; Zdybicka-Barabas, A.; Wojda, I.; Wiater, A.; Mak, P.; Suder, P.; Skrzypiec, K.; Cytryńska, M. Fungal α-1,3-Glucan as a New Pathogen-Associated Molecular Pattern in the Insect Model Host Galleria mellonella. Molecules 2021, 26, 5097. [Google Scholar] [CrossRef] [PubMed]
- Thomaz, L.; García-Rodas, R.; Guimarães, A. J.; Taborda, C. P.; Zaragoza, O.; Nosanchuk, J. D. Galleria mellonella as a Model Host to Study Paracoccidioides lutzii and Histoplasma capsulatum. Virulence 2013, 4, 139–146. [Google Scholar] [CrossRef]
- Singulani, J. L.; Scorzoni, L.; de Oliveira, H. C.; Marcos, C. M.; Assato, P. A.; Fusco-Almeida, A.; Mendes-Giannini, M. Applications of Invertebrate Animal Models to Dimorphic Fungal Infections. JoF 2018, 4, 118. [Google Scholar] [CrossRef]
- Freitas, D. F.; Santos, S. S.; Almeida-Paes, R.; de Oliveira, M. M.; do Valle, A. C.; Gutierrez-Galhardo, M. C.; Zancopé-Oliveira, R. M.; Nosanchuk, J. d. Increase in Virulence of Sporothrix brasiliensis over Five Years in a Patient with Chronic Disseminated Sporotrichosis. Virulence 2015, 6, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Clavijo-Giraldo, D. M.; Matínez-Alvarez, J. A.; Lopes-Bezerra, L. M.; Ponce-Noyola, P.; Franco, B.; Almeida, R. S.; Mora-Montes, H. M. Analysis of Sporothrix schenckii sensu stricto and Sporothrix brasiliensis Virulence in Galleria mellonella. Journal of Microbiological Methods 2016, 122, 73–77. [Google Scholar] [CrossRef]
- Lozoya-Pérez, N. E.; Casas-Flores, S.; Martínez-Álvarez, J. A.; López-Ramírez, L. A.; Lopes-Bezerra, L. M.; Franco, B.; Mora-Montes, H. M. Generation of Sporothrix schenckii Mutants Expressing the Green Fluorescent Protein Suitable for the Study of Host-Fungus Interactions. Fungal Biology 2018, 122, 1023–1030. [Google Scholar] [CrossRef]
- Borba-Santos, L. P.; Barreto, T. L.; Vila, T.; Chi, K. D.; dos Santos Monti, F.; de Farias, M. R.; Alviano, D. S.; Alviano, C. S.; Futuro, D. O.; Ferreira, V.; de Souza, W.; Ishida, K.; Rozental, S. In Vitro and In Vivo Antifungal Activity of Buparvaquone against Sporothrix brasiliensis. Antimicrob Agents Chemother 2021, 65, e00699–21. [Google Scholar] [CrossRef]
- Browne, N.; Heelan, M.; Kavanagh, K. An Analysis of the Structural and Functional Similarities of Insect Hemocytes and Mammalian Phagocytes. Virulence 2013, 4, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, G.; Garvey, A.; Croke, M.; Kavanagh, K. Innate Humoral Immune Defences in Mammals and Insects: The Same, with Differences ? Virulence 2018, 9, 1625–1639. [Google Scholar] [CrossRef]
- Wojda, I. Immunity of the Greater Wax Moth Galleria mellonella: Galleria mellonella Immunity. Insect Science 2017, 24, 342–357. [Google Scholar] [CrossRef]
- Ratcliffe, N. A.; Gagen, S. J. Studies on the in Vivo Cellular Reactions of Insects: An Ultrastructural Analysis of Nodule Formation in Galleria mellonella. Tissue and Cell 1977, 9, 73–85. [Google Scholar] [CrossRef]
- Lavine, M. D.; Strand, M. R. Insect Hemocytes and Their Role in Immunity. Insect Biochemistry and Molecular Biology 2002, 32, 1295–1309. [Google Scholar] [CrossRef]
- Pereira, T.; de Barros, P.; Fugisaki, L.; Rossoni, R.; Ribeiro, F.; de Menezes, R.; Junqueira, J.; Scorzoni, L. Recent Advances in the Use of Galleria mellonella Model to Study Immune Responses against Human Pathogens. JoF 2018, 4, 128. [Google Scholar] [CrossRef] [PubMed]
- Melo, N. R. de; Abdrahman, A.; Greig, C.; Mukherjee, K.; Thornton, C.; Ratcliffe, N. A.; Vilcinskas, A.; Butt, T. M. Myriocin Significantly Increases the Mortality of a Non-Mammalian Model Host during Candida Pathogenesis. PLoS ONE 2013, 8, e78905. [Google Scholar] [CrossRef] [PubMed]
- Gandra, R. M.; McCarron, P.; Viganor, L.; Fernandes, M. F.; Kavanagh, K.; McCann, M.; Branquinha, M. H.; Santos, A. L. S.; Howe, O.; Devereux, M. In Vivo Activity of Copper(II), Manganese(II), and Silver(I) 1,10-Phenanthroline Chelates Against Candida Haemulonii Using the Galleria mellonella Model. Front. Microbiol. 2020, 11, 470. [Google Scholar] [CrossRef] [PubMed]
- Vogel, H.; Altincicek, B.; Glöckner, G.; Vilcinskas, A. A Comprehensive Transcriptome and Immune-Gene Repertoire of the Lepidopteran Model Host Galleria mellonella. BMC Genomics 2011, 12, 308. [Google Scholar] [CrossRef]
- Nale, J. Y.; Chutia, M.; Cheng, J. K. J.; Clokie, M. R. J. Refining the Galleria mellonella Model by Using Stress Marker Genes to Assess Clostridioides difficile Infection and Recuperation during Phage Therapy. Microorganisms 2020, 8, 1306. [Google Scholar] [CrossRef]
- Champion, O.; Titball, R.; Bates, S. Standardization of G. mellonella Larvae to Provide Reliable and Reproducible Results in the Study of Fungal Pathogens. JoF 2018, 4, 108. [Google Scholar] [CrossRef]
- Senior, N. J.; Titball, R. W. Isolation and Primary Culture of Galleria mellonella Hemocytes for Infection Studies. F1000Res 2021, 9, 1392. [Google Scholar] [CrossRef] [PubMed]
- Scorzoni, L.; de Lucas, M. P.; Mesa-Arango, A. C.; Fusco-Almeida, A. M.; Lozano, E.; Cuenca-Estrella, M.; Mendes-Giannini, M. J.; Zaragoza, O. Antifungal Efficacy during Candida krusei Infection in Non-Conventional Models Correlates with the Yeast In Vitro Susceptibility Profile. PLoS ONE 2013, 8, e60047. [Google Scholar] [CrossRef] [PubMed]
- Brown, S. E.; Howard, A.; Kasprzak, A. B.; Gordon, K. H.; East, P. D. A Peptidomics Study Reveals the Impressive Antimicrobial Peptide Arsenal of the Wax Moth Galleria mellonella. Insect Biochemistry and Molecular Biology 2009, 39, 792–800. [Google Scholar] [CrossRef]
- Zhao, H.-X.; Xiao, W.-Y.; Ji, C.-H.; Ren, Q.; Xia, X.-S.; Zhang, X.-F.; Huang, W.-Z. Candidate Chemosensory Genes Identified from the Greater Wax Moth, Galleria mellonella, through a Transcriptomic Analysis. Sci Rep 2019, 9, 10032. [Google Scholar] [CrossRef]
- Miranda, L. H. M.; Santiago, M. de A.; Schubach, T. M. P.; Morgado, F. N.; Pereira, S. A.; Oliveira, R. de V. C. de; Conceição-Silva, F. Severe Feline Sporotrichosis Associated with an Increased Population of CD8 low Cells and a Decrease in CD4 + Cells. Med. Myco. 2015, myv079. [Google Scholar] [CrossRef]
- Legrand, N.; Weijer, K.; Spits, H. Experimental Models to Study Development and Function of the Human Immune System In Vivo. The Journal of Immunology 2006, 176, 2053–2058. [Google Scholar] [CrossRef]
- Arrillaga-Moncrieff, I.; Capilla, J.; Mayayo, E.; Marimon, R.; Marine, M.; Genis, J.; Cano, J.; Guarro, J. Different Virulence Levels of the Species of Sporothrix in a Murine Model. Clinical Microbiology and Infection 2009, 15, 651–655. [Google Scholar] [CrossRef]
- Borman, A. M. Of Mice and Men and Larvae: Galleria mellonella to Model the Early Host-Pathogen Interactions after Fungal Infection. Virulence 2018, 9, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gaviria, M.; Martínez-Álvarez, J. A.; Mora-Montes, H. M. Current Progress in Sporothrix brasiliensis Basic Aspects. JoF 2023, 9, 533. [Google Scholar] [CrossRef] [PubMed]
- Piatek, M.; Sheehan, G.; Kavanagh, K. Utilising Galleria mellonella Larvae for Studying in vivo Activity of Conventional and Novel Antimicrobial Agents. Pathogens and Disease 2020, 78, ftaa059. [Google Scholar] [CrossRef] [PubMed]
- Dinh, H.; Semenec, L.; Kumar, S. S.; Short, F. L.; Cain, A. K. Microbiology’s next Top Model: Galleria in the Molecular Age. Pathogens and Disease 2021, ftab006. [Google Scholar] [CrossRef] [PubMed]
- Binder, U.; Maurer, E.; Lass-Flörl, C. Galleria mellonella: An Invertebrate Model to Study Pathogenicity in Correctly Defined Fungal Species. Fungal Biology 2016, 120, 288–295. [Google Scholar] [CrossRef]
- Fakhim, H.; Vaezi, A.; Dannaoui, E.; Chowdhary, A.; Nasiry, D.; Faeli, L.; Meis, J. F.; Badali, H. Comparative Virulence of Candida Auris with Candida haemulonii, Candida glabrata and Candida albicans in a Murine Model. Mycoses 2018, 61, 377–382. [Google Scholar] [CrossRef]
- Corrêa-Moreira, D.; Menezes, R. C.; Romeo, O.; Borba, C. M.; Oliveira, M. M. E. Clinical and Anatomopathological Evaluation of BALB/c Murine Models Infected with Isolates of Seven Pathogenic Sporothrix Species. Pathogens 2021, 10, 1647. [Google Scholar] [CrossRef]
- Trevijano-Contador, N.; Zaragoza, O. Immune Response of Galleria mellonella against Human Fungal Pathogens. JoF 2018, 5, 3. [Google Scholar] [CrossRef]
- Tamez-Castrellón, A. K.; van der Beek, S. L.; López-Ramírez, L. A.; Martínez-Duncker, I.; Lozoya-Pérez, N. E.; van Sorge, N. M.; Mora-Montes, H. M. Disruption of Protein Rhamnosylation Affects the Sporothrix schenckii-Host Interaction. The Cell Surface 2021, 7, 100058. [Google Scholar] [CrossRef]
- Rossoni, R. D.; Fuchs, B. B.; de Barros, P. P.; Velloso, M. dos S.; Jorge, A. O. C.; Junqueira, J. C.; Mylonakis, E. Lactobacillus Paracasei Modulates the Immune System of Galleria mellonella and Protects against Candida albicans Infection. PLoS ONE 2017, 12, e0173332. [Google Scholar] [CrossRef]
- de Beer, Z. W.; Duong, T. A.; Wingfield, M. J. The Divorce of Sporothrix and Ophiostoma : Solution to a Problematic Relationship. Studies in Mycology 2016, 83, 165–191. [Google Scholar] [CrossRef] [PubMed]
- Meister, M.; Lemaitre, B.; Hoffmann, J. A. Antimicrobial Peptide Defense In Drosophila. Bioessays 1997, 19, 1019–1026. [Google Scholar] [CrossRef]
- Bulet, P.; Hetru, C.; Dimarcq, J.-L.; Hoffmann, D. Antimicrobial Peptides in Insects; Structure and Function. Developmental & Comparative Immunology 1999, 23, 329–344. [Google Scholar] [CrossRef]
- Lee, Y. S.; Yun, E. K.; Jang, W. S.; Kim, I.; Lee, J. H.; Park, S. Y.; Ryu, K. S.; Seo, S. J.; Kim, C. H.; Lee, I. H. Purification, CDNA Cloning and Expression of an Insect Defensin from the Great Wax Moth, Galleria mellonella. Insect Mol Biol 2004, 13, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Gabriele Barros Mothé. Estudo Da Interação Entre Fagócitos Do Felis Catus (LINNAEUS,1758) e os Principais Agentes Etiológicos Da Esporotricose. Tese de Doutorado, Universidade Federal Fluminense, Niterói, Rio de Janeiro, Brasil, 2021. https://app.uff.br/riuff/handle/1/24482.
- Złotko, K.; Wiater, A.; Waśko, A.; Pleszczyńska, M.; Paduch, R.; Jaroszuk-Ściseł, J.; Bieganowski, A. A Report on Fungal (1→3)-α-d-Glucans: Properties, Functions and Application. Molecules 2019, 24, 3972. [Google Scholar] [CrossRef]
Figure 1.
Phenotypic investigation of the infection dynamics in the invertebrate host Galleria mellonella against the fungal pathogen Sporothix brasiliensis. (A) Survival curve performed up to the tenth day of infection. (B) Monitoring the fungal load from the insect's hemolymph during the infection process. (C) Hemocyte count performed from insects in the control groups and the group infected with S. brasiliensis. (D) Parallel drawn between insect cellular immune response versus fungal load along the survival curve.
Figure 1.
Phenotypic investigation of the infection dynamics in the invertebrate host Galleria mellonella against the fungal pathogen Sporothix brasiliensis. (A) Survival curve performed up to the tenth day of infection. (B) Monitoring the fungal load from the insect's hemolymph during the infection process. (C) Hemocyte count performed from insects in the control groups and the group infected with S. brasiliensis. (D) Parallel drawn between insect cellular immune response versus fungal load along the survival curve.
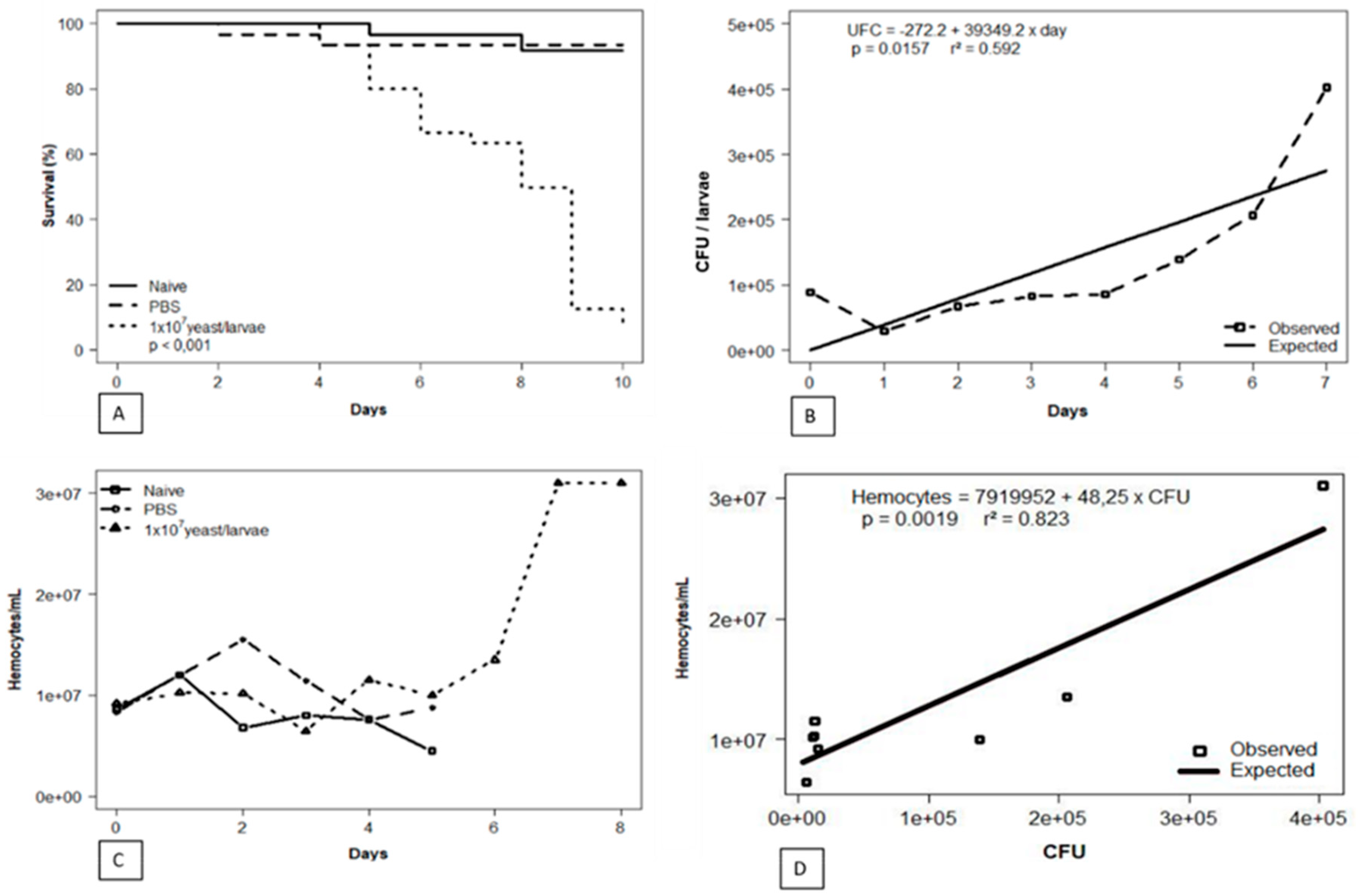
Figure 2.
Investigation of the gene expression profile of the invertebrate host Galleria mellonella in infection by the pathogenic fungus Sporothix brasiliensis. Relative quantification of genes encoding antimicrobial peptides (A) Gallerimycin, (B) Galliomycin. And the stress manager genes (C) C7 Contig 15362 and (D) C8 Contig 19101. The units on the Y axis were calculated based onthe 2-ΔΔCT method and are expressed as mean. Each gene was normalized and compared to the expression of control (naive) insects using the β-actin reference gene. The Mann-Whitney test was used to compare the relative quantification of genes and a p ≤0.05 value was considered significant.
Figure 2.
Investigation of the gene expression profile of the invertebrate host Galleria mellonella in infection by the pathogenic fungus Sporothix brasiliensis. Relative quantification of genes encoding antimicrobial peptides (A) Gallerimycin, (B) Galliomycin. And the stress manager genes (C) C7 Contig 15362 and (D) C8 Contig 19101. The units on the Y axis were calculated based onthe 2-ΔΔCT method and are expressed as mean. Each gene was normalized and compared to the expression of control (naive) insects using the β-actin reference gene. The Mann-Whitney test was used to compare the relative quantification of genes and a p ≤0.05 value was considered significant.
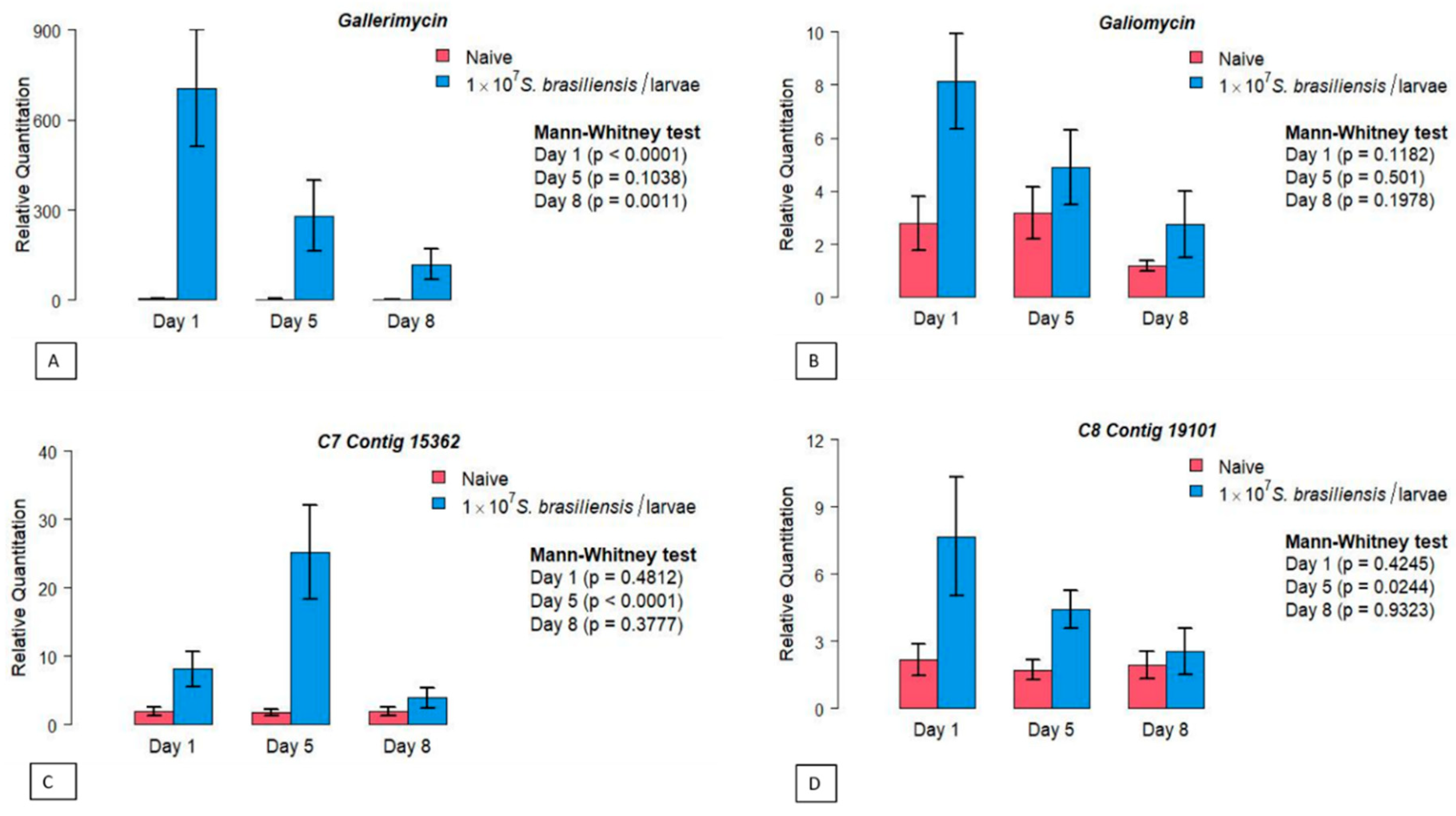

Table 1.
Genes quantified during the fungal infection process of Galleria mellonella by Sporothrix brasiliensis. The references used as well as the primer sequence.
Table 1.
Genes quantified during the fungal infection process of Galleria mellonella by Sporothrix brasiliensis. The references used as well as the primer sequence.
| Gene | NCBI Genbank References |
Sequence (5’ – 3’) |
Annealing temperature |
|---|---|---|---|
| Galiomycin | AY528421.1 | F- TCCAGTCCGTTTTGTTGTTG | 60ºC |
| [37] | R- CAGAGGTGTAATTCGTCGCA | ||
| Gallerimycin | AF453824.1 | F- GAAGATCGCTTTCATAGTCGC | 60ºC |
| [37] | R- TACTCCTGCAGTTAGCAATGC | ||
| C7 Contig 15362 | Contig 15362 | F- CGAGCTAAAGACAGGCGATT | 58ºC |
| [30] | R- TCACCTGCGGTTGAATCATA | ||
| C8 Contig 19101 | Contig 19101 | F- ATTGCTAGCCAGGTTCAGGA | 60ºC |
| [30] | R- AGCTATTTGGCGGAAACTCA | ||
| β-actin | [38] |
F- GGACTTGTACGCCAACACAG R- CCACATCTGCTGGAATGTCG |
55ºC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



