
Submitted:
31 August 2023
Posted:
04 September 2023
You are already at the latest version
Abstract
Bacterial resistance to antibiotics is a public health menace that needs urgent efforts to mitigate and forestall their transmission in the general environment. Plasmid-mediated metallo beta-lactamase (MBL) gives bacteria an exceptional ability to transmit resistance genes. This study investigated plasmid-mediated resistance in MBL P. aeruginosa isolates from poultry food chain settings. Cloacal swab samples (n=120) were bacteriologically analyzed for the isolation of P. aeruginosa isolates. Susceptibility studies were performed by the CLSI guideline using selected antibiotics. MBL was phenotypically detected using modified Hodges test, and confirmed using PCR technique. Plasmid curing studies detected the presence of plasmids in the isolates. P. aeruginosa isolates (n=48) showed high levels of reduced susceptibility to cefoxitin (93.8%), imipenem (81.3%), gentamicin (85.4%), amikacin (85.4%), nitrofurantoin (83.3%), ofloxacin (72.9%) and cefotaxime (77.1%). MBL was phenotypically detected in 7 (14%) isolates of P. aeruginosa. The presence of blaIMP-1 and blaIMP-2 MBL genes in P. aeruginosa (n=5) isolates was confirmed by PCR. The PCR did not detect blaVIM-1 and blaVIM-2 MBL genes. The P. aeruginosa isolates were multidrug resistant, with about 50% of the isolates showing multidrug resistance (MDR) to the antibiotics. The genes for MDR in the P. aeruginosa isolates were borne on plasmids, indicating a high possibility of community transmission of MBL resistance genes. Finally, the P. aeruginosa isolates from poultry food chain milieus are multidrug resistant, produce MBL, and harbour their resistance genes on plasmids. Since many antibiotic resistance genes in bacteria are associated with mobile genetic elements (MGEs) like plasmids, it is important to step up actions to mitigate incessant use of antibiotics in poultry and other agricultural practices in Nigeria in order to diminish the evolution and spread of plasmid-mediated resistance which is capable of initiating community acquired infections that may prove difficult to treat in our hospitals with the available antibiotics.
Keywords:
Gram negative bacteria
; multidrug resistance
; plasmids
; carbapenemase
; carbapenems
; mobile genetic elements (MGEs)
1. Introduction
Antibiotic resistance is the unresponsiveness of pathogenic bacteria to the antimicrobial onslaughts of existing antimicrobials or antibiotics that is supposed to stop its growth or cause its death. It is a global problem that knows not the border of any nation, and has the potential to halt, and possibly roll back some of the notable progresses made in the field of antimicrobial therapy if nothing is done fast to mitigate it. Most problem of antibiotic resistance occurs in the hospital environment where antibiotics are usually used heavily for the treatment and management of infectious diseases [1,2,3,4]. Nonetheless, this trend is not usually the case – as some other cases of antibiotic resistance have been previously reported in the general environment including abattoirs, poultry, livestock and animal husbandry where antibiotics particularly those used in clinical medicine are also used as prophylaxis and for promoting animal growth [5,6,7,8]. Antibiotics are usually used at sub-inhibitory concentrations in non-clinical practices such as in poultry and livestock production. In such scenarios, the antibiotic only kills susceptible bacteria and allows other resistant strains (that survives its antimicrobial onslaught) to go on thriving and thus become resistant to the same antibiotic later. Antibiotic resistant bacteria can spread to humans from animals and poultry birds, as well as from human-to-human contact; and it can also spread from raw food, and from the environment into the human population and vice versa [3,5,7]. One of such resistance phenotype that have emerged as a result of the incessant use of antibiotics (as growth promoters or food supplements) in animal husbandry and livestock production, and which can spread from animals to humans through contact or consumption are metallo beta-lactamases (MBLs). MBLs are bacterial enzymes with exceptional ability to hydrolyze and confer resistance to carbapenems such as imipenem and meropenem [1,9]. MBLs are mostly found in non-lactose fermenters such as Pseudomonas and Acinetobacter species; and to a lesser extent in lactose-fermenting Enterobacteriaceae [9]. The presence of MBL-producing bacteria in the hospital or community milieus causes not only a therapeutic problem but also serious concern for infection control management in the health system. This is because bacteria producing MBLs are multidrug resistant, and can cause resistance to non-carbapenem agents [8]. MBL-producing bacteria could confer resistance to antibacterial agents from different classes, thereby limiting treatment options. Previous reports suggest that MBLs can efficiently hydrolyze all beta-lactam drugs except aztreonam (a monobactam) in vitro [9,10]. The prevalence of bacteria producing MBLs is gradually growing globally [11]. And bacterial strains such as Pseudomonas aeruginosa harbouring plasmids (extra-chromosomal DNA that allows transmission of genetic materials including resistance genes between bacteria species) that mediate MBL production could worsen the antibiotic crisis by allowing MBL genes to be easily transmitted between bacteria genera and species in a particular environment (e.g., poultry milieus). P. aeruginosa is a notable nosocomial pathogen that is intrinsically resistant to some commonly available antibiotics. Plasmid-mediated strains of MBL-producing P. aeruginosa from poultry milieus portend public health menace due to the transmissibility of their antibiotic resistance genes (ARGs) to other bacteria species through plasmids. Furthermore, strains of P. aeruginosa that produce MBLs are known to be multidrug resistant, and show reduced susceptibility to the carbapenems – which are critically important antibiotics used as a last-line of defense against bacterial infections caused by multidrug resistant bacteria including those producing extended spectrum β-lactamases (ESBLs) [8,9]. Thus, the mitigation of antibiotic resistance in the general environment, especially in the food chain sectors such as in poultry will help stop their evolution and spread, as well as help ensure a sustainable environment in terms of safer foods and water for all. The current study is a first report of plasmid curing studies of antibiotic resistance in Pseudomonas aeruginosa isolates from poultry milieus tested for metallo-β-lactamase (MBL) production and plasmid profile in Enugu, Nigeria.
2. Results
In this study, we investigated the antimicrobial susceptibility patterns and plasmid profile of metallo beta-lactamase-producing Pseudomonas aeruginosa isolates from poultry milieus in Enugu, Nigeria. Table 2 shows the frequency of isolation of P. aeruginosa isolates from the cloaca swab samples bacteriologically analyzed in this study.
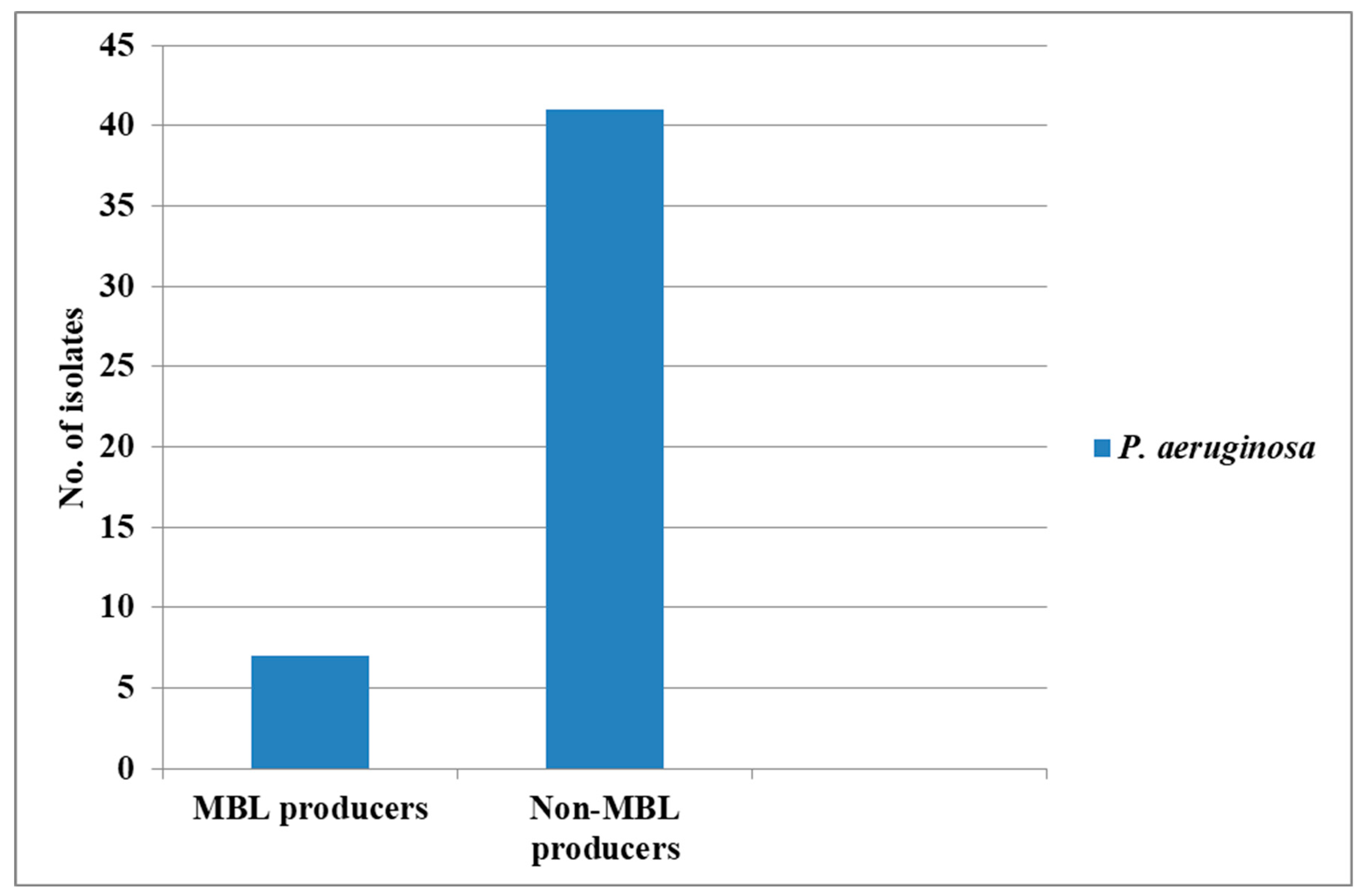
Out of the 48 isolates of P. aeruginosa investigated for metallo beta lactamase production using the modified Hodges test (MHT) protocol, only 14.6% (n=7) of the isolates were confirmed phenotypically to produce MBL – which is a high profile antibiotic degrading enzyme that renders carbapenems (e.g., imipenem) ineffective for treating bacterial-related infections (Figure 1)
Figure 1.
Number of P. aeruginosa isolates that produced metallo beta lactamase (MBL) by the MHT technique.
Figure 1.
Number of P. aeruginosa isolates that produced metallo beta lactamase (MBL) by the MHT technique.
The result of the antimicrobial susceptibility testing of the 48 isolates of P. aeruginosa in this study is shown in Figure 2. The P. aeruginosa isolates showed high levels of resistance to the tested antibiotics, particularly to cefoxitin (93.8%), imipenem (81.3%), gentamicin (85.4%), amikacin (85.4%), nitrofurantoin (83.3%), ofloxacin (72.9%) and cefotaxime (77.1%). On the other hand, least susceptibility was also recorded in the P. aeruginosa isolates against ceftriaxone (64.6%), ertapenem (68.8%), meropenem (58.3%), ciprofloxacin (54.2%), ceftazidime (50%) and aztreonam (37.5%). All the P. aeruginosa isolates were multiply resistant to the tested antibiotics as over 50% of the isolates showed multiple resistances to antibiotics in more than 3 classes. Multiple drug resistance was recorded in antibiotic classes of cephalosporins, carbapenems and aminoglycosides against the isolated P. aeruginosa isolates from poultry milieus investigated in this study.
Figure 2.
Percentage antibiotic susceptibility and resistance of P. aeruginosa isolated from cloaca swab samples (n=48). Key: CRO-ceftriaxone, FOX-cefoxitin, IPM-imipenem, CAZ-ceftazidime, ETP-ertapenem, OFX-ofloxacin, CN-gentamicin, AK-amikacin, CIP-ciprofloxacin, CTX-cefotaxime, MEM-meropenem, ATM-aztreonam, F-nitrofurantoin.
Figure 2.
Percentage antibiotic susceptibility and resistance of P. aeruginosa isolated from cloaca swab samples (n=48). Key: CRO-ceftriaxone, FOX-cefoxitin, IPM-imipenem, CAZ-ceftazidime, ETP-ertapenem, OFX-ofloxacin, CN-gentamicin, AK-amikacin, CIP-ciprofloxacin, CTX-cefotaxime, MEM-meropenem, ATM-aztreonam, F-nitrofurantoin.
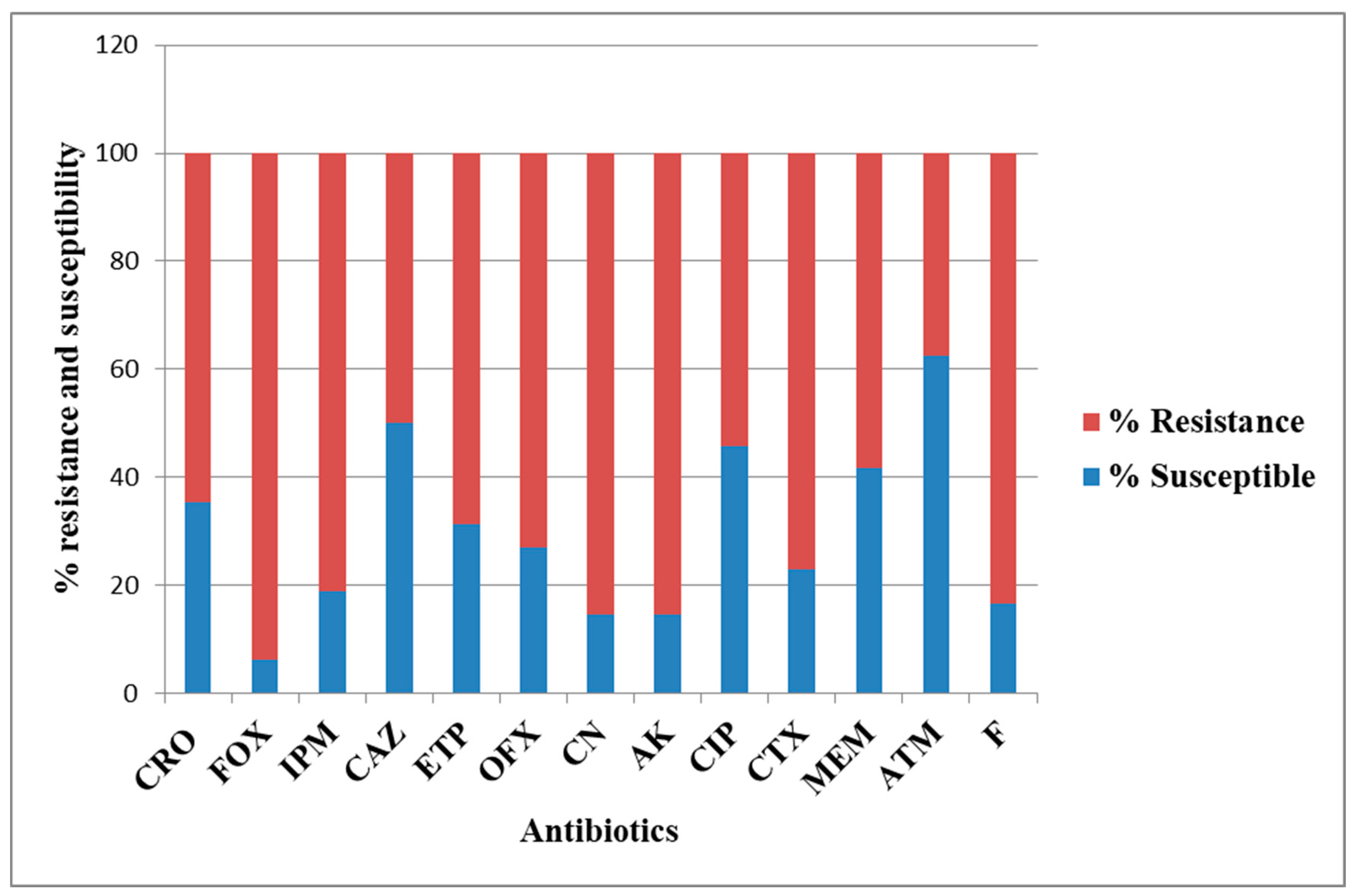
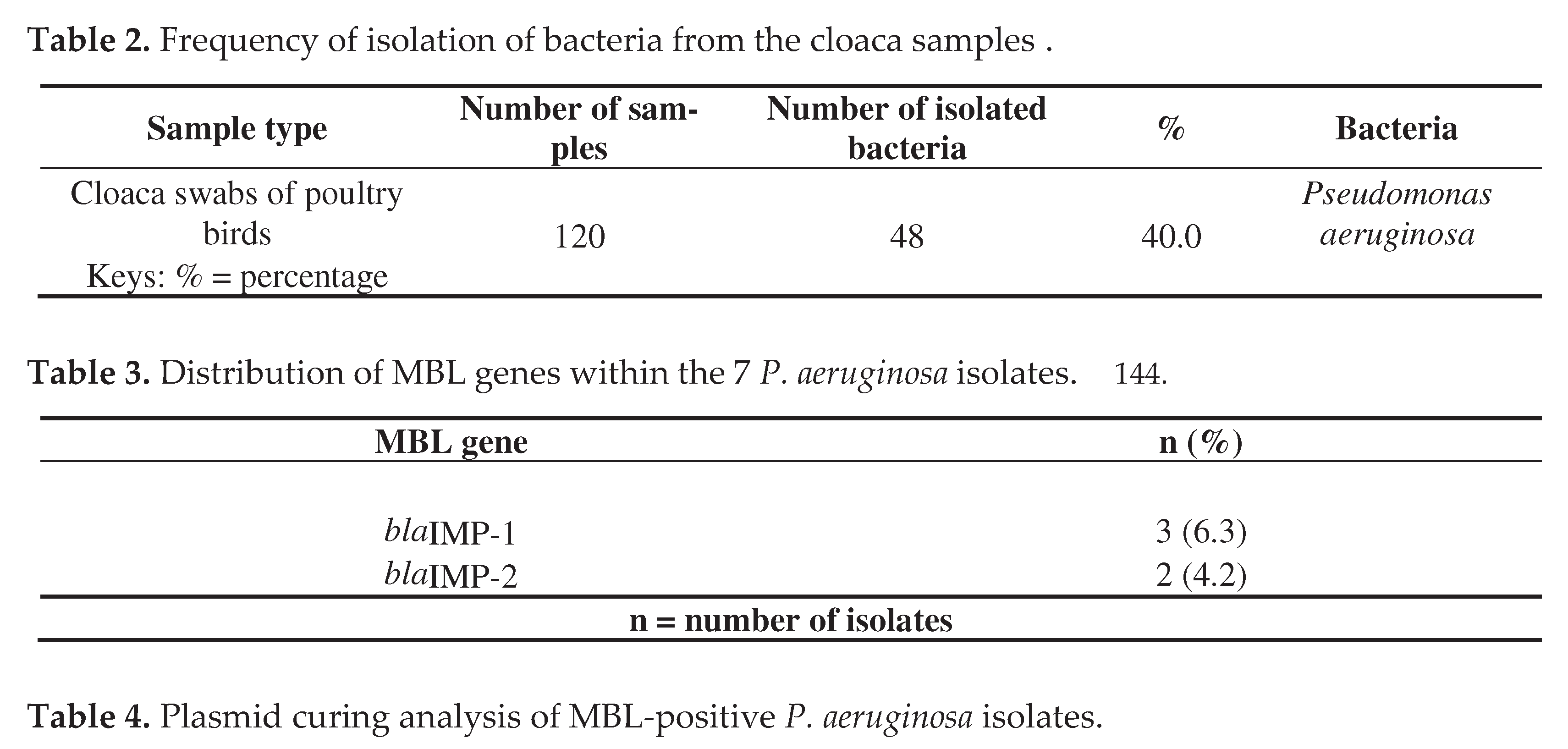
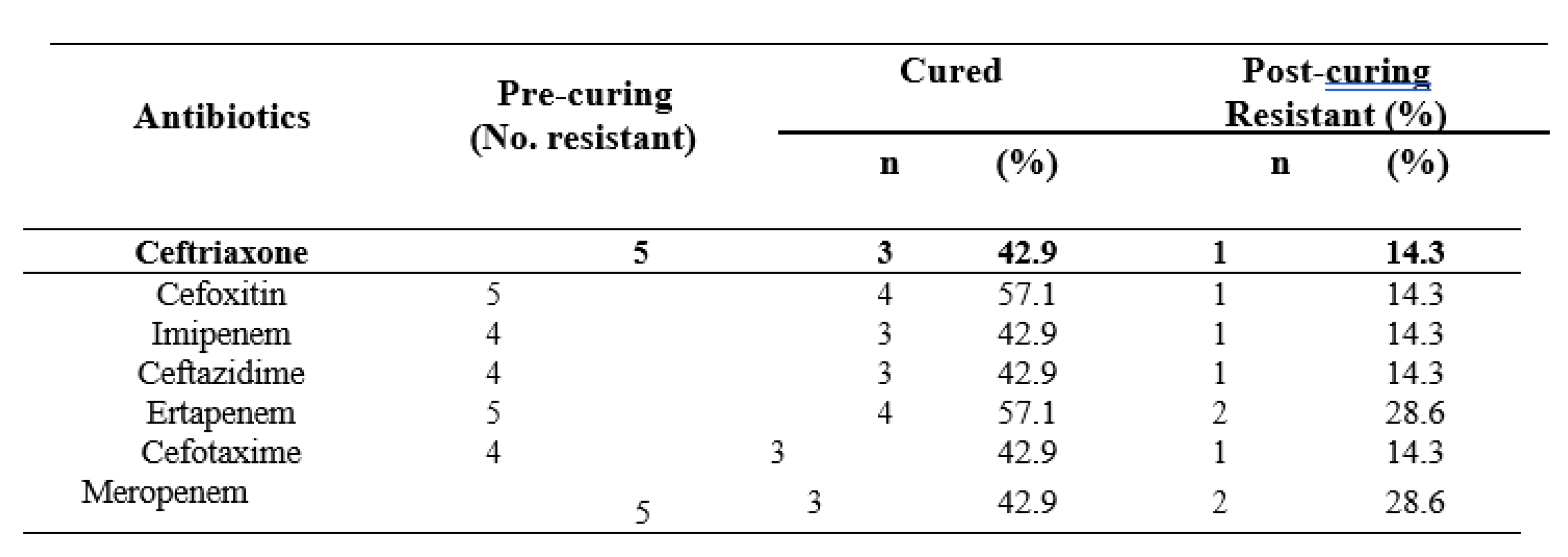
Table 3 shows the result of the molecular investigation to determine the genes responsible for metallo beta lactamase production in the P. aeruginosa isolates studied. From our results, the blaIMP-1 and blaIMP-2 MBL genes was detected by PCR in the P. aeruginosa isolates (n=7) investigated. While only 3 isolates of the P. aeruginosa isolates harboured the blaIMP-1 MBL genes; only 2 isolates of P. aeruginosa harboured the blaIMP-2 MBL genes (Table 3). The blaVIM-1 blaVIM-2 MBL genes was not detected in the P. aeruginosa isolates investigated in this study. The results of the plasmid studies carried out on the P. aeruginosa isolates (n=7) that produced metallo beta lactamase (MBL) in this study is shown in Table 4. From the results, it was observed that most of the antibiotic resistant P. aeruginosa isolates that produced MBL in this study were plasmid-mediated (Table 4).
3. Discussion
In our bid to sustainably contain the evolution and spread of antibiotic resistant bacteria and genes in the human population and the general environment, it is vital to strengthen environmental actions of antibiotic usage and resistance containment in the One-Health concept since the health of humans, animals and the environment are interconnected. The rate at which pathogenic bacteria from both the hospital and general environment are becoming resistant and even multiply-resistant to some current antibiotics treatment has significantly increased over the last decade according to studies [4,14,911]. This has also jeopardized our effort to effectively treat some common bacterial (microbial) infections, and also put more pressure on the available antibiotics since the rate at which microorganisms (bacteria) evolve resistance has surpassed the speed at which new antimicrobials that can withstand some multidrug resistant organisms are developed. In this work, we reported the antibiotic resistance and plasmid profile of Pseudomonas aeruginosa isolates from poultry milieus in Enugu, Nigeria. The P. aeruginosa isolates recovered in this study showed high levels of resistance to the tested antibiotics, particularly to drugs in the classes: carbapenems, cephalosporins, aminoglycosides and fluoroquinolones. In no particular order, the P. aeruginosa isolates were resistant to imipenem (81.3%), cefotaxime (77.1%), amikacin (85.4%), gentamicin (85.4%) and ofloxacin (72.9%). Moreover, the isolated P. aeruginosa isolates in this study also showed high levels of multidrug resistance to more than 50% of the tested antibiotics, particularly to agents in the carbapenems, aminoglycosides, cephalosporins and fluoroquinolone classes. A study in Egypt showed that about 50-90% of the P. aeruginosa isolates recovered from hospital samples showed high rates of resistance to antibiotics in the carbapenems, cephalosporins, and aminoglycosides and fluoroquinolones classes as reported in our study [18]. Another study in China corroborates our results of high levels of resistance, and also showed that P. aeruginosa isolates from Chinese poultry farms and chickens showed high levels of resistances to some clinically-relevant antibiotics including those in the classes: cephalosporins, aminoglycosides and fluoroquinolones [5,19]. To the best of our knowledge, it was observed from our study that out of the 48 isolates of P. aeruginosa recovered from the poultry samples, only 7 isolates representing about 15% of the total recovered isolates were phenotypically confirmed to produce metallo beta-lactamase (MBL). To further confirm MBL production in the P. aeruginosa isolates, the results of our PCR analysis confirmed MBL production in a total of 5 isolates. Several studies have shown that there is an increase in the evolution and spread of MBL-producing bacteria in the non-hospital environment as reported in our study [5,7,20]. Furthermore, the results from our study corroborates to previous studies which have reported the prevalence of MBL-producing bacteria in livestock and poultry [7,11]. In this study, we detected the blaIMP-1 MBL genes in 3 P. aeruginosa isolates (6.3%) while the blaIMP-2 MBL genes were detected in 2 isolates of P. aeruginosa (4.2%) from poultry milieus. The MBL genes detected in this current study (blaIMP-1 and blaIMP-2) have been previously reported in China, Egypt and Nigeria as important genes responsible for mediating MBL production in Gram negative bacteria including P. aeruginosa isolates [5,8,18,19]. The rate of MBL resistance genes detected in this study is lower than the rate detected in Egypt (11%), Algeria (16%), Iran (22%), and Turkey (29%) where high prevalence of MBL genes particularly the blaIMP-1 and blaIMP-2 genes was recorded from poultry samples and other samples from the general environment [18,21,22,23]. MBL is a high profile antibiotic degrading enzyme that have attracted a lot of public health attention globally in recent times due to its ability to render the carbapenems used for treating infections caused by bacteria producing extended spectrum beta-lactamases (ESBLs) ineffective. The abuse of antibiotics particularly in Nigeria, where there is no formal legislation or policy to control antibiotic use has made some bacteria species to become resistant to the carbapenems and other antibiotics of clinical relevance [17,24,25]. Plasmids are important mobile genetic elements (MGEs) for the transmission of antibiotic resistance genes amongst bacterial species and the general environment. The P. aeruginosa isolates tested for the location of their resistance (MBL) genes in this study shows that most of the isolates harboured their antibiotic resistance genes on their plasmids. This indicates that their antibiotic resistance genes are borne on a plasmid since about 40-50% of the P. aeruginosa isolates showed appreciable zones of inhibition (plasmids were cured) when tested against some selected antibiotics to which they previously showed reduced susceptibility. On the other hand, about 14-29% of the P. aeruginosa isolates did not show zones of inhibition (plasmids not cured) indicating that the isolates had their resistance genes borne on a chromosome. Acquired resistance which is mostly mediated by horizontal transfer of antibiotic resistance genes amongst bacterial species through plasmids and other MGEs is a critical public health issue which is driving the evolution and spread of antimicrobial resistance (AMR) in hospitals and the general environment. According to Zwanzig [26], the antibiotics used in agriculture and animal or fish farming pollute the environment and act as a selective pressure for resistances, potentially increasing the risk of transfer of resistance genes to human pathogens. P. aeruginosa is an important nosocomial pathogen that can also cause a handful of community acquired-infections. Thus, the prevalence of multidrug resistant (MDR) P. aeruginosa producing MBL in poultry milieus as reported in this study makes it an important MDR pathogen of public health significance that needs urgent attention to mitigate its evolution and spread in the general environment.
4. Materials and Methods
Ethical statement: This study received ethical approval from the Local Ethics and Research Committee of the Faculty of Pharmaceutical Sciences, Enugu State University of Science and Technology (ESUT), Nigeria [viz ref: ESUT/FPS/EA/01].
Sampling protocol: A total of 120 cloaca swab samples were aseptically collected with an informed (oral) consent of the poultry farmers, from poultry birds in various poultry farms in Enugu, Nigeria for this study. All samples were labeled and transported to the laboratory for bacteriological investigation using standard microbiology techniques.
Isolation of bacteria and identification: Each of the cloaca swab samples were plated on cetrimide selective agar (CSA) supplemented with 10% glycerol. All culture plates were incubated at 37oC for 18-24h [4,12]. Putative colonies of P. aeruginosa that produced bluish or greenish colonies (as a result of pigmentation) were recovered and further plated on freshly prepared CSA plates for the isolation of discrete (pure) colonies which were further subjected to biochemical tests including oxidase, indole, urease, catalase and citrate tests for the identification of P. aeruginosa strains [12].
Antibiotic susceptibility studies: This experiment was undertaken using our previous protocol on the determination of the antibiotic susceptibility of bacteria from the non-hospital environment [8]. All P. aeruginosa isolates were subjected to antibiotic susceptibility studies using single antibiotics disks (Oxoid, UK) comprising: ceftriaxone (CRO; 30μg), cefoxitin (FOX; 30μg), imipenem (IPM; 10μg), ceftazidime (CAZ; 30μg), ertapenem (ETP; 30μg), ofloxacin (OFX; 10μg), gentamicin (CN; 10μg), amikacin (AK; 10μg), ciprofloxacin (CIP; 10μg), cefotaxime (CTX; 30μg), meropenem (MEM; 10μg), aztreonam (ATM; 30μg), nitrofurantoin (F; 10μg). The susceptibility test disks were placed at a distance of 20mm apart on freshly prepared Mueller Hinton (MH) agar plates (Oxoid, UK) already inoculated with the test bacteria (adjusted to 0.5 McFarland’s turbidity standards). All susceptibility test plates were incubated at 37oC for 18-24h in the incubator. Inhibition zone diameter (IZD) was measured and reported as susceptible or resistance following the standard antibiotics breakpoints and guidelines of the Clinical Laboratory Standard Institute (CLSI) [13].
Detection of metallo-β-lactamase (MBL): MBL was phenotypically detected in the isolated P. aeruginosa isolates using the modified Hodges test (MHT) technique on MH agar plates as we previously described in our earlier study [4]. A standard strain of E. coli (ATCC 25922) was inoculated by spreading on a MH agar plate, and imipenem (IPM) disk (10µg) was applied at the center of the inoculated MH agar plate. Thereafter, a suspension of an overnight culture of the P. aeruginosa was streaked or inoculated onto the already inoculated MH agar plate using sterilized inoculating loop by making a streak of the test organism from the edge of the IPM disk (10µg) to the edge of the MH agar plate. All plates were incubated at 37oC for 18-24h in the incubator. The plates were observed for Cloverleaf effect at the intersection of the test bacterium and the E. coli ATCC 25922 standard organism, within the inhibition zone of the IPM disk (10µg). The growth of the P. aeruginosa towards the IPM disk (10µg) was inferred phenotypically for MBL production by the MHT protocol [4].
Molecular analysis: PCR analysis for the detection of specific MBL genes (blaIMP-1, blaIMP-2, blaVIM-1, blaVIM-2) in the P. aeruginosa isolates that showed reduced susceptibility to any of the tested carbapenems (imipenem and meropenem) was performed in accordance to a previously used protocol [11,14,15]. DNA was extracted from the P. aeruginosa isolates using GENESpin DNA isolation kit (Eurofins, Hamburg) according to the manufacturer’s guidelines. Extracted DNA was stored at 20oC for the PCR analysis. The PCR mix was a final volume of 26.5μl reaction mixture comprising: 0.2µl of Taq pol U/µl, 2.5µl of 10X PCR buffer, 2.5µl MgCl2, 1µl of 10pM from each of the forward and reverse primers (Table 1), 2.5µl of dNTPs MIX (2Mm), 3µl of DNA template, 14.8µl of nuclease-free water. The presence of MBL genes in the P. aeruginosa isolates was investigated by PCR amplification in a thermal cycler (Lumex instruments, Canada) using defined PCR conditions (Table 1). DNA amplicons were run in a 1.5% agarose gel electrophoresis for 60m at 120V while a UV transilluminator (Scientico, India) was used for the visualization of the amplified DNA products in the tested isolates.

Plasmid curing studies: Plasmid curing was carried out using acridine orange (as the plasmid curing agent) to determine the location (plasmid or chromosomal) of the antibiotic resistance phenotype of the isolated drug resistant (metallo beta-lactamase-producing) P. aeruginosa isolates. Two colonies of individual MBL-producing P. aeruginosa isolates were grown in 5ml nutrient broth supplemented with 0.1 mg/ml of acridine orange. Positive control (P. aeruginosa without acridine orange) and negative control (acridine orange without P. aeruginosa) tubes were run alongside the curing experiments. All tubes were incubated overnight at 37oC in the incubator. Tubes containing bacterial cells and the acridine orange were selected, and a loopful of the test organism in the tube(s) was streaked aseptically on MacConkey agar (MCA) plate(s) and CSA plates which served as the recovery medium [16]. The plates were incubated overnight at 37oC in the incubator, and resultant colonies were sub-cultured onto freshly prepared MAC and CSA plates to get pure strains of P. aeruginosa. Colonies of P. aeruginosa that emanated from the recovery plates were checked for the loss of antibiotic resistance determinants (genes) by investigating their susceptibility profile to any of the carbapenems (imipenem or meropenem) and their MBL production ability using the MHT technique. The carriage of the antibiotic resistant genes (ARGs) on the plasmid was confirmed in P. aeruginosa isolates that showed susceptibility to previously ineffective antibiotic (imipenem or meropenem or others) and were confirmed as non-MBL producers by the MHT technique.
Multiple antibiotic resistance index (MARI): The determination of the multiple antibiotics resistance index (MARI) of the resistant P. aeruginosa isolates was determined according to a previously used protocol using the MARI formular [17]. To calculate MARI for multidrug resistance profiling, the following formular was used: MARI= Number of antibiotics to which resistance occurred / total number of antibiotics to which the P. aeruginosa isolates were tested.
5. Conclusions
Despite the gains on antimicrobial stewardship across the globe, more still needs to be done to curb antimicrobial resistance (AMR) in the developing world like Nigeria where the healthcare system is still comatose and AMR is less debated at both institutional and national levels. In this current study, we report the prevalence of MDR P. aeruginosa isolates from poultry milieus, and which harbour their antibiotic resistance genes on a plasmid. Antibiotics misuse and faecal contamination in poultry and other non-hospital milieus may contribute to the evolution and spread of MBL-producing bacteria such as plasmid-mediated MBL-producing P. aeruginosa in the general environment. The prevalence of MBL-producing P. aeruginosa isolates in poultry milieus reported in this study suggests that the surveillance on multidrug resistance, particularly outside the hospital environment is needed urgently across the food chain in the various states in Nigeria.
Author Contributions
CE and MU: conceptualization, funding acquisition, writing - original draft preparation; formal analysis: TM, NU, CE and MA: methodology; EO and IE: investigation: EN and NO: data curation; CA and CD: visualization; IE, AM and TM: writing - review and editing; supervision: CE and MU. All authors performed sample collection and processing and revised the final draft of the manuscript with contributions from all authors.
Funding
This manuscript was drafted as a part of a deliverable of the research project “Functional Metagenomics to Reveal and Monitor Novel Antimicrobial Resistance Genes and Strains Circulating in the Food Chain in Nigeria”, funded by the Institute Mérieux (IMX), France (IMX-NIMR-Award 2021), in which Dr. Chika Ejikeugwu was awarded a Young Investigator Award on Antimicrobial Resistance (AMR) in 2021, in which all authors participated. The funders had no role in the collection of samples, management of the project, design and conduct of the study, analysis and interpretation of the data; and the preparation, review, and approval of the final draft of the manuscript.
Institutional Review Board Statement
This study received approval from the Local Ethics and Research Committee of the Faculty of Pharmaceutical Sciences, Enugu State University of Science and Technology (ESUT), Nigeria. (Approval code: ESUT/RES/FPS/001) Oral consent and approval was sought from the poultry farmers for sample collection.
Data Availability Statement
All data and protocol pertaining to the study are contained within the body of this manuscript. Additional datasets not contained in current manuscript will be supplied upon request from the corresponding author (Dr. Chika Ejikeugwu, chika.ejikeugwu@esut.edu.ng).
Acknowledgments
Authors thank Institute Mérieux (IMX), France and Nigerian Institute of Medical Research (NIMR) for providing the funding which supported this project. We are also grateful to the management of ESUT, Nigeria for other administrative support during the study. The technical and administrative support of staff of Faculty of Pharmaceutical Sciences is appreciated.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be interpreted as a potential conflict of interest.
References
- Abbas FM (2021). Metallo-b-lactamases: A review. Ann. Rom. Soc. Cell Biol. 25, 1308–1316.
- Hernando-Amado S, Coque TM, Baquero F, Martinez JL (2019). Defining and combating antibiotic resistance from One Health and global health perspectives. Nat. Microbiol. 4, 1432–1442. [CrossRef]
- Agarwal S, Durlabhji P, Gupta S, Mittal J, Dalela G (2017). Incidence of metallo-β-lactamase producing Pseudomonas aeruginosa isolates and their antimicrobial susceptibility pattern in clinical samples from a tertiary care hospital. International Journal of Research and Review, 4(1):92-98.
- Ejikeugwu C, Ugwu M, Iroha I, Gugu T, Duru C, Eze P, Esimone C (2013). Detection and antimicrobial susceptibility of some Gram negative bacteria producing carbapenemases and extended spectrum beta lactamases. International Journal of Microbiology and Immunology Research, 2(6):064-069.
- Zhang R, Liu Z, Li J, Lei L, Yin W, Li M, Wu C, Walsh TR, Wang Y, Wang S, Wu Y (2017). Presence of VIM-positive Pseudomonas species in chickens and their surrounding environment. Antimicrob Agents Chemother, 61:e00167-17. [CrossRef]
- Falodun OS and Musa IB (2020). Pseudomonas species from cattle dung producing extended spectrum and metallo beta-lactamases. European Journal of Biological Research, 10(1):1-10. [CrossRef]
- Woolhouse M, Ward M, Van Bunnik B, Farrar J (2015). Antimicrobial resistance in humans, livestock and the wider environment. Phil Trans Royal Soc B Lond Ser B, Biol Sci. 370: 1-7. [CrossRef]
- Ejikeugwu C, Okoro Nworie, Morteza Saki, Hussein O.M. Al-Dahmoshi, Noor S.K. Al-Khafaji, Chika Ezeador, Emmanuel Nwakaeze, Peter Eze, Eniola Oni, Chidiebere Obi, Ifeanyichukwu Iroha, Charles Esimone, Michael U. Adikwu (2021). Detection of metallo-β-lactamase and AmpC genes in Escherichia coli, Klebsiella species, and Pseudomonas aeruginosa isolates from abattoir and poultry origin in Nigeria. BMC Microbiology, 21(124):1-9. [CrossRef]
- Walsh T.R., Toleman M.A., Poirel L and Nordmann P (2005). Metallo β – Lactamases: the Quiet before the Storm? Clinical Microbiology Review, 18(2):306-325. [CrossRef]
- Saderi H., Karimi Z., Owlia P., Bahar M.A., Rad S.M (2008). Phenotypic Detection of Metallo – beta – lactamase producing Pseudomonas aeruginosa strains isolated from Burned Patients. Iranian Journal of Pathology, 3(1):20-24.
- Fang L, Liu Z, Lu Z, Huang R, Xiang R (2022). Identification and characterization of a novel metallo β-lactamase, SZM-1, in Shenzhen Bay, South China. Front. Microbiol. 13:996834. [CrossRef]
- Cheesbrough M (2006). District Laboratory Practice in Tropical Countries. Part 2, 2nd Ed. Cambridge University Press, 2006.
- Wayne PA (2006). Performance Standards for Antimicrobial Susceptibility Testing. In CLSI Approved Standard M100. (Clinical and Laboratory Standards Institute CLSI, 2019).
- Shibata N, Doi Y, Yamane K, Yagi T, Kurokawa H, Shibayama K, Kato H, Kai K and Arakawa Y (2003). PCR typing of genetic determinants for metallo-β-lactamases and integrases carried by Gram-negative bacteria isolated in Japan, with focus on the class 3 integron. Journal of Clinical Microbiology, 41(12):5407-5413. [CrossRef]
- Ejikeugwu C, Morteza S, Nwakaeze E, Eze P, Orinya C, Carissa D, Edeh C, Esimone C, Iroha I, Adikwu M (2019). Characterization of blaIMP-1 MBL genes among Klebsiella species from abattoir samples by multiplex PCR. Gene Reports, 16:100428.
- Onyeadi DJ and Agbagwa OE (2019). Plasmid curing in multidrug resistant hospital and community uropathogenic Escherichia coli. J. Appl. Sci. Environ. Manage. 23(1):29-34. [CrossRef]
- Ejikeugwu C, Onele S, Okonkwo E, Onu E, Afiukwa N, Nwakaeze E, Udu-Ibiam O, Iroha C, Edeh C, Iroha I (2022). Antimicrobial drug resistance in strains of Salmonella isolated from pig effluents in Abakaliki, Nigeria. Nigerian Journal of Microbiology, 36(2):6229-6235.
- Elmonir W, Abd El-Aziz NK, Tartor YH, Moustafa SM, Abo Remela EM, Eissa R, Saad HA, and Tawab AA (2021). Emergence of colistin and carbapenem resistance in extended-spectrum beta-lactamase producing Klebsiella pneumoniae isolated from chickens and humans in Egypt. Biology (Basel). 10:373. [CrossRef]
- Zhai R, Fu B, Shi X, Sun C, Liu Z, Wang S, Shen Z, Walsh TR, Cai C, Wang Y, and Wu C (2020). Contaminated in-house environment contributes to the persistence and transmission of NDM-producing bacteria in a Chinese poultry farm. Environ. Int.139:105715. [CrossRef]
- Kelly AM, Mathema B and Larson EL (2017). Carbapenem-resistant Enterobacteriaceae in the community: A scoping review. Int. J. Antimicrob. Agents, 50, 127–134. [CrossRef]
- Chaalal N, Touati A, Bakour S, Aissa MA, Sotto A, Lavigne JP, and Pantel A (2021). Spread of OXA-48 and NDM-1-Producing Klebsiella pneumoniae ST48 and ST101 in Chicken Meat in Western Algeria. Microb. Drug Resist, 27:492–500. [CrossRef]
- Sarhangi M, Motamedifar M, Sarvari J. Dissemination of Pseudomonas aeruginosa producing blaIMP1, blaVIM2, blaSIM1, blaSPM1 in Shiraz, Iran. Jundishapur J Microbiol, 2013; 6:e6920–5. [CrossRef]
- Toraman Z, Yakupogullari Yand Kizirgil A (2004). Detection of metallo β-lactamase production and antibiotic resistance with E-test method in Pseudomonas, Acinetobacter and Klebsiella strains, in Turkey. J Infect hemother, 10:257-261. [CrossRef]
- Mojica MF, Rossi MA, Vila AJ and Bonomo RA. (2022). The urgent need for metallo-beta-lactamase inhibitors: An unattended global threat. Lancet Infect. Dis, 22, e28–e34. [CrossRef]
- Pongchaikul P and Mongkolsuk P (2022). Comprehensive analysis of imipenemase (IMP)-type metallo-beta-lactamase: A global distribution threatening asia. Antibiotics, 11:236. [CrossRef]
- Zwanzig M (2021). The ecology of plasmid-coded antibiotic resistance: a basic framework for experimental research and modeling. Computational and Structural biotechnology Journal, 19:586-599. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



