
Submitted:
31 August 2023
Posted:
04 September 2023
You are already at the latest version
Abstract
Hydroxamic acid (HA) derivates displayed antibacterial and antifungal activities. HA with various numbers of carbon atoms (C2, C6, C8, C10, C12 and C17), complexed to different metal ions Fe(II/III), Ni(II), Cu(II) and Zn(II) were evaluated for their antimycobacterial activities and their anti-biofilm activities. Some derivates, among others HA12Fe2, inhibited the development of Mycobacterium tuberculosis, Mycobacterium bovis BCG and Mycobacterium marinum biofilms and could even attack pre-formed Pseudomonas aeruginosa biofilm. Proteomic profiles showed that the potential targets of HA10FeCl were mainly related to mycobacteria stress adaptation, involving cell wall lipid biosynthesis, drug resistance and tolerance, siderophore metabolism. This study provides new insights regarding the antimycobacterial activities of the HA and their complexes, especially about their potential antibiofilm activities.
Keywords:
Hydroxamic acids
; Antibacterial
; Anti-biofilm
; Mycobacterium tuberculosis
1. Introduction
Tuberculosis (TB) is a communicable disease caused by the pathogenic bacillus Mycobacterium tuberculosis. According to the WHO Global Tuberculosis Report 2022, globally, in 2021, an estimated 10.6 million people developed an active tuberculosis and 1.58 million died from tuberculosis [1]. In addition, occurrence of increasing numbers of multi drug-resistant (MDR) and extensively drug-resistant (XDR) Mycobacterium tuberculosis strains is of great concern [1]. Mycobacterium tuberculosis cell wall contains from the inside to the outside, an arabinogalactan layer covalently attached to the peptidoglycan layer and mycolic acids, covalently attached to the arabinogalactan, in which various unique long carbon chain lipids can be further embedded. The complex outer membrane is further wrapped up by capsular and biofilm extracellular compounds [2]. Some of the long and complex lipids non-covalently attached to the outer membrane, only present in pathogenic mycobacteria, such as the phthiocerol dimycocerosate (PDIM), have been shown to play important roles, not only in virulence, i.e., for camouflage in macrophages, but also in antibiotic intrinsic resistance [3,4]. Furthermore, pathogenic mycobacteria show slow growth, tending to enter in a non-replicative state when facing in vivo stress conditions, eventually leading to dormancy. To circumvent this metabolic characteristic, long (more than 6 months) and complex (more than 4 drugs) must be administrated to the patients in order to eradicate the infection. Despite the efficacy of the current anti-TB treatments, several factors, such as poor patient compliance, toxicity of drugs and irrational prescribing practices increase the risk of drug-resistant strain selection. This leads to even further difficult and more expensive treatment [1]. Thus, novel and efficient anti-tuberculosis drugs are urgently need to fight TB.
Since the first hydroxamic acid (HA) discovery in 1869, HA and its related analogs have been intensively studied. This chemical component class shows diverse applications, in biology, industry and medicine. Interestingly, some of the HA derivates displayed antimicrobial properties with anti-bacterial, anti-virulence or anti-fungal activities. For instance, the benzoylacetone and HA complexes could inhibit the A. hydrophila, S. aureus, E. aerogenes, S. sonnei, P. aeruginosa, S. typhimurium, S. epidermidis, and M. luteus [5]. Cyclic hydroxamic acid (N-hydroxylactam) able to chelated iron ions inhibited iron-containing enzyme lipoxygenase in Gram-negative P. aeruginosa [6]. Heterocyclic methylsulfone hydroxamates analogous inhibited the LpxC enzyme activity, involved in lipopolysaccharide (LPS) production of Gram-negative bacteria [7]. Due to the potential values of HA analogues in the field of medicine development, we previously synthesized various HA derivates, including complexes with varying numbers of carbon atoms (C2, C6, C8, C10, C12 and C17) and their corresponding Fe(II/III), Ni(II), Cu(II) and Zn(II) complexes and we observed that the C6 to C12 HA derivates complexed with Fe(II/III) had antibacterial activities against gram positive and negative bacteria while the C12 HA had a larger spectrum activities, on bacteria and yeast [8]. The C12 HA compounds complexed or not to the Fe(II/III) even showed an antimycobacterial activity on M. smegmatis [8].
Furthermore, HA analogs also displayed antimycobacterial activity against mycobacteria, like M. tuberculosis, M. abscessus, M. marinum, M. smegmatis [9,10,11]. The complexes of benzohydroxamate associated with transition metallic ions (Cu2+ and Co2+) could inhibit M. tuberculosis growth by interaction with the urease in the nitrogen metabolism [9]. The compound of para-nitrobenzohydroxamic acid demonstrated a minimum inhibitory concentration (MIC) of 0.71 μM in glycerol-alanine salt medium or a MIC of 7.79 μM in 7H12 medium on M. tuberculosis [12]. The pentacyanoferrate moiety in Fe(II) coordination hydroxamic complexes benefited of the release of HNO from HA, improving pyrazinamide and delamanid efficiency against M. tuberculosis [13]. The suberoylanilide hydroxamic acid (SAHA) also had adjunctive potential to enhance the effects of first-line anti-TB drugs (isoniazid and rifampicin) against intracellular M. tuberculosis [14].
The aims of this study was to evaluate the potential and selective antimycobacterial and anti-biofilm activities (on M. tuberculosis, M. bovis BCG and M. marinum) of 47 HA derivates, containing various numbers of carbon atoms (C2, C6, C8, C10, C12 and C17) and eventually complexed with Fe(II/III), Ni(II), Cu(II) and Zn(II), to further assess their potentials as anti-TB or anti-biofilm drug candidates.
2. Material and methods
2.1. Materials
The mycobacteria strains, including M. bovis BCG (Pasteur 1173P2), M. tuberculosis H37Ra, M. marinum (M strain) were obtained from Gene Optimal (Shanghai, China). The P. aeruginosa (LMG 6395) was purchased from Belgian Coordinated Collection of Microorganisms (BCCM), University of Gent. The 7H9 broth and OADC medium were purchased form BD (BD BBLTM, USA). The Mueller Hinton broth (MHB) was purchased from Sigma- Aldrich (Saint Louis, USA). HA derivate stocks at 10 mM to 200 mM were prepared in DMSO or methanol.
2.2. Methods
2.2.1. Antimycobacterial drug susceptibility assay
Mycobacteria precultures were grown in 25 cm2 flasks at 37°C without shaking in 7H9 medium (BD) supplemented with 10% (v/v) OADC (BD) to an OD600 of 0.7-0.9. The macrodilution method was performed in 10 mL screw tubes in 7H9 medium (BD) containing 0.05% glycerol and 10% OADC. Five hundred μL inoculum diluted in the supplemented 7H9 medium to reach an optical density at 600 nm (OD600) of 0.01 were added to 500 μL serial drug dilutions in the same 7H9 medium. Tubes were placed without shaking at 37°C. Growth or absence of growth were recorded on the day that the growth of the 100-fold diluted drug-free inoculum control became visible in order to assess the minimal inhibitory concentration of the drugs (MIC, being the lowest drug concentration inhibiting more than 99% of mycobacteria growth)[15]. This experiment was performed three times to check the reproducibility of the MIC determination.
The combined effect of vancomycin and chemical component was also performed using microdilution method. The fractional inhibitory concentration index (FICI) was calculated according to the checkerboard method. FICI=FICa+FICb=MICab/MICa+MICba/MICb. MICa or MICb is the MIC of the chemical component or vancomycin alone, MICab is the MIC of the chemical component in combination with a fixed vancomycin concentration, MICba is the MIC of the vancomycin in combination with a fixed chemical component concentration were obtained. The fixed concentrations were 4 and 20-fold lower than the MICa or MICb, respectively. In agreement with the checkerboard method, synergy is reached when the FICI is < 0.5 [16].
2.2.2. Biofilm growth assay
For mycobacteria, precultures were grown in 7H9 medium with 10% (v/v) OADC to an optical density at 600 nm (OD600) of 0.8-0.9. The biofilm growth inhibition assay was performed in the 6-well plate or 24-well plate in the Sauton’s medium[17,18]. All the compound samples were first diluted within Sauton’s medium to various concentrations. The mycobacteria precultures were inoculated as a 100-fold dilution (corresponding approximately to 9 ×105 CFU/mL inoculum) in the plates, giving a final culture volume of 4 mL or 2 mL, in the 6-well plate or 24-well plate, respectively. The plates were covered with two layers of Parafilm® “M” and incubated at 37 °C. Biofilm formation was visually assessed after 3-4 weeks of culture.
In contrast to the mycobacteria biofilm inhibition test, the P. aeruginosa biofilm inhibition test was performed on pre-formed biofilms[19]. P. aeruginosa pre-formed biofilms were obtained on the TSP (solid phase transfer) covers (Nunc) of the 96-well from a 106 CFU/mL starting inoculum incubated for 24 h in MHB. After 24 h, covers with pre-formed biofilms were immersed and incubated at 37 °C for twelve days in 100 µL of MHB containing compound samples at different concentrations ranging from between 20-2500 µM. Cetrimide was used as a control (concentrations between 78 ng/mL to 1 µg/mL). Biofilm control wells developed in the presence of 5% (v/v), DMSO or MeOH were present on each plate. The TSP coverslips containing the previously formed biofilm were used to cover the supports of the plates containing the test compound solutions. These plates were incubated at 37 °C for twelve days [19]. During the incubation period, observation of the plates was performed daily to ensure the presence of the MHB medium. The experiment was performed twice in triplicate for each solution of tested compounds.
2.3. Proteomic analysis
In order to identify potential targets of the HA complexes, we performed proteomic analysis. The M. bovis BCG (Pasteur 1173P2) strain was treated with HA10FeCl for 7 days and mycobacteria without HA10FeCl were used as control. The bacterial pellets were collected by centrifugation with 3000 g for 15 min and washed twice with cold PBS. Bacteria were resuspended and lysed in the SDT lysis buffer (4% m/v, SDS,100 mM Tris-HCl pH7.6,1 mM DTT). The extracted proteins were quantified with the BCA Protein Assay Kit (Bio-Rad, USA). Protein digestion by trypsin was performed according to filter-aided sample preparation (FASP) procedure [20]. The digested peptides of each sample were desalted on C18 Cartridges (Empore SPE cartridges, Sigma), concentrated by vacuum centrifugation and reconstituted in 40 µl of 0.1% (v/v) formic acid.
LC-MS/MS analysis was performed on a timsTOF Pro mass spectrometer (Bruker) that was coupled to Nanoelute (Bruker Daltonics) for 60/120/240 min. The peptides were loaded onto a reverse phase trap column (Thermo Scientific) connected to the C18-reversed phase analytical column (Thermo Scientific Easy Column, 10 cm long, 75 μm inner diameter, 3 μm resin) in buffer A (0.1% formic acid) and separated with a linear gradient of buffer B (84% acetonitrile and 0.1% formic acid) at a flow rate of 300 nl/min. The mass spectrometer was performed in a positive ion mode. The mass spectrometer collected ion mobility MS spectra over a mass range of m/z 100-1700 and 1/k0 of 0.6 to 1.6, and then performed 10 cycles of PASEF MS/MS with a target intensity of 1.5 k and a threshold of 2500. Active exclusion was enabled with a release time of 0.4 minutes.
MS raw data were combined and searched using the MaxQuant software (version: 1.5.3.17) for identification and quantitation analysis and a UniProt Mycobacterium bovis (strain BCG/Pasteur 1173P2) database (https://www.uniprot.org/). The protein hits with a p value < 0.05 (t test) and a fold change <0.8 or >1.2 were further analysed.
3. Results
3.1. HA compounds present anti-mycobacterial activities
In order to identify whether the HA derivatives could have antimycobacterial activities, we assessed those activities in planktonic condition and in stress condition with high glycerol concentration leading to biofilm development. Biofilm conditions are indeed known to better mimic the stressful in vivo condition [21,22,23]. These antimycobacterial activities were assessed using three strains, harboring various characteristics, among others, the presence of absence of the phthiocerol dimycocerosate (PDIM) lipids in their cell wall: the H37Ra M. tuberculosis strain is PDIM negative and is a slow growing strain, the M. marinum is PDIM positive and a slow growing mycobacteria (however in planktonic culture it grows faster than M. bovis BCG or H37Ra M. tuberculosis strains) and the M. bovis BCG is PDIM positive and a slow growing mycobacteria. The drug susceptibility assays performed under planktonic growth condition eventually allowed to identify the minimal inhibitory concentration (MIC), while those performed under biofilm growth development allowed to eventually identify the minimal biofilm-formation inhibitory concentration (MBIC).
In planktonic conditions, various HA and complexes displayed anti-mycobacterial activities, especially on M. tuberculosis H37Ra strain (Table 1, Table 2 and Table 3). The active compounds had generally a MIC ≥ 200 µM on the three species. An inhibitory activity was less often observed on M. marinum, with only the HA10 and the HA8FeCl and HA8Fe3 complexes showing an anti-mycobacterial activity. On the M. bovis BCG, 8 compounds showed an inhibitory activity, however with relatively high MIC (≥ 200 µM), except for HA10FeCl showing a MIC of 100-200 µM.
In stressful condition, mycobacteria were grown into biofilm. Biofilm formation was visible from 10-12 days for M. bovis BCG and M. tuberculosis H37Ra, from 15-20 days for M. marinum and mature biofilms were obtained in 20-24 days for M. bovis BCG, in 28-30 days for H37Ra M. tuberculosis and in 35-40 days for M. marinum. The biofilm was thinner for M. marinum, compared to those of M. bovis BCG and H37Ra M. tuberculosis. Again, more compounds had biofilm inhibitory activities on the H37Ra M. tuberculosis strain. The HA10 was active on the three biofilms with minimal biofilm-formation inhibitory concentration (MBIC) ranging between the 100 and 250 µM, depending on the bacteria strain. For most active compounds on the M. marinum biofilm, the MBIC were ≥ 200 µM, except for HA8FeCl, HA10FeCl, HA10Fe, HA8Ni2 with MBIC of 62.5 µM and for HA12Ni2 and HA12Cu2 with MBIC of 125 µM. For M. bovis BCG, the active compounds had a MBIC generally ≥ 100 µM, except for HA10Fe2 (20 µM), HA10FeCl (20-100 µM), HA10Fe3 (20-40 µM), HA10Ni2 (20 µM) and HA10Cu2 (20 µM) (Supplementary figure 1) (Table 1, Table 2 and Table 3). For the H37Ra M. tuberculosis strain, the active compounds had generally a MBIC ≥ 125 µM, except for HA6Ni2, HA8Ni2, HA10FeCl and HA8Fe3 with a MIC of 31.25 µM, HA10Fe3 with a MIC of 31.25-62.5 µM, HA17Fe3 with a MIC of 62.5 µM (Table 1, Table 2 and Table 3).
In order to assess whether some of the HA compounds could target PDIM biosynthesis in pathogenic mycobacteria, we investigated the susceptibility M. bovis BCG to vancomycin in the presence of HA10FeCl. Indeed, we previously showed that the vancomycin, usually used to treat Gram positive bacteria and inactive on pathogen mycobacteria, can target those ones when they are lacking PDIM in their cell wall [4]. Drug targeting the compounds involved in PDIM biosynthesis can thus synergize with vancomycin in the drug susceptibility assay. Interestingly, in the present study the complex HA10FeCl increased by more than 4-fold the susceptibility of M. bovis BCG to vancomycin in drug susceptibility assay. To investigate whether this growth inhibition results from a synergy between the HA10FeCl and the vancomycin, the Checkerboard method was also used to calculate the FICI (Table 4). The MIC of complexes HA10FeCl dropped from 46.34-92.7μg/mL to 11.59 μg/mL in the presence of vancomycin (50 μg/mL), and the MIC of vancomycin dropped from 750 µg/mL to 125 µg/mL in the presence of complexes HA10FeCl (46.35 µg/mL), suggesting that the combination can inhibit M. bovis BCG growth in synergy (FICI=0.292-0.417).
3.2. HA10Fe2, HA12Fe2 and HA12FeCl can also reduce pre-formed P. aeruginosa biofilm.
Furthermore, in view of the large spectrum of compounds able to inhibit mycobacterial biofilm development, we investigated the anti-biofilm activity of the compounds on pre-formed P. aeruginosa biofilm, a well-known biofilm difficult to eradicate. Most of the compounds were totally inactive in this assay, such as HA10Ni2, HA10Cu2, HA8Cu2, CuCl2, NiCl2 and FeCl2 even at 2.5 mM. Interestingly, the iron-complexes HA10Fe2, HA12Fe2 and HA12FeCl could inhibited the P. aeruginosa biofilm formation with a MBIC of 625 µM, 312.5 µM and 312.5 µM, respectively (data not shown).
3.3. Proteomic profile of the HA10FeCl-treated bacilli
In order to better understand the mode of the action of the drug candidate, we also compared the proteomics of the HA10FeCl-treated M. bovis BCG cells with untreated cells (Figure 1 and Figure 2). A total 41 proteins were screened and identified with significant difference (p<0.05) by t test and with a fold change (FC) >1.1 and <0.9, including 32 up-regulated and 9 down-regulated proteins in the HA10FeCl-treated bacilli, compared with the control without treatment (Table S1).
Generally, the hydrolase proteins encoded by the genes (pstC2, bglS, BCG_1059c, BCG_0364, moaC3, gltD, lipG, BCG_2973, BCG_0099, truA, BCG_1935c, mltG, topA, pks12) were up-regulated. The possible membrane proteins encoded by the genes BCG_1478, BCG_3932, BCG_3545c, mmpS4, BCG_1127c were also up-regulated in the HA10FeCl-treated bacilli. The Phosphate transport system permease protein encoded by the gene pstC2 with a FC of 2.19 and the Rubredoxin encoded by the BCG_3279c with a FC of 2.19 were also up-regulated. The proteins encoded by the genes crgA, BCG_3932, BCG_1708, BCG_1384c, BCG_0352, BCG_2826, BCG_3492c, gltD, lipG, BCG_2813c, BCG_0099, BCG_1268, truA, mutT2, BCG_1935c, mltG, topA, BCG_3927, BCG_1127c, rnpA_2 were also up-regulated in HA treated M. bovis BCG. In addition, the proteins encoded by pks12 (BCG_2067c), BCG_2973 involved in PDIM biosynthesis and the proteins mmpS4 involved in siderophore export was also up-regulated in HA treated M. bovis BCG. By contrast, the proteins encoded by the genes thiG, BCG_2384c and BCG_2177c were down-regulated in the HA10FeCl-treated M. bovis BCG.
Furthermore, we also observed some up-regulated proteins, encoded BCG_1100c, narK2, BCG_0259c, BCG_3092 in HA10FeCl-treated M. bovis BCG and down-regulated proteins, encoded by genes PE15, mmpL9a (BCG_2361) in the control were not identified from the proteomic profiles (data not shown).
4. Discussion
In the present study, the anti-mycobacterial activities and anti-biofilm activities were assessed using three mycobacteria strains (H37Ra M. tuberculosis, M. bovis BCG and M. marinum) and P. aeruginosa. Globally, the obtained results showed that HA antimycobacterial activity depended on the carbon chain length and the metal ions in the complexes. HA with C10 and C12 carbon chain (HA10 and HA12, respectively) displaying higher activity than C2 and C6 carbon chain HA (HA2 and HA6). Among the complexes, those with at C8, C10, and C12 length carbon chain, for instance, HA10FeCl, HA12FeCl, HA8Fe3, HA10Fe3 showed interesting antimycobacterial activities with MIC of 125 µM and MBIC of 31.25-62.5 µM on M. tuberculosis, with MBIC similar to Ni-complexes HA6Ni2 and HA8Ni2. The iron-complexes HA10FeCl, HA10Fe2, HA10Fe3 displayed higher inhibitory on M. bovis BCG biofilm with a MBIC of 20-100 µM, compared to the other iron-complexes. By contrast, these iron-complexes showed less activity on M. bovis BCG growth in planktonic conditions. Indeed, the iron-complexes inhibitory effect on mycobacteria biofilm formation is globally better than in planktonic growth conditions.
In this study our compounds were evaluated to target three mycobacterial strains, M. bovis BCG (PDIM+/PGL+), M. marinum (PDIM+/PGL+) and M. tuberculosis H37Ra (PDIM-). As expected from a PDIM negative strain, the M. tuberculosis H37Ra strain was more susceptible to a larger panel of compounds, compared with other two mycobacterial strains. This was probably due to the impaired cell wall impermeability. As well known, the mycobacterial cell wall lipids, like trehalose monomycolate and dimycolate (TMM, TDM), PDIM, sulpholipid-1 (SL-1), diacyl trehalose (DAT), and pentacyl trehalose (PAT) are known to play an important role in pathogenesis. The proteins involved in the cell lipids biosynthesis were also considered as the potential virulence factors, like transport proteins. In M. tuberculosis, the mycobacterial membrane protein MmpL and MmpS family mediating transport of important cell wall lipids across the mycobacterial membrane, together with their interactor play important roles in the synthesis and export of mycobacterial outer membrane lipids [24], such as drug efflux (MmpL5 and MmpL7), siderophore export (MmpL4/MmpS4 and MmpL5/MmpS5), and heme uptake (MmpL3 and MmpL11) [25,26]. The iron–sulfur proteins [3Fe-4S] and [4Fe-4S] ferredoxins and [1Fe-0S] rubredoxins play the important role of iron-containing proteins in maintaining redox homeostasis [27]. The rubredoxin are involved in the electron transfer processes and was considered to be part of an evolutionary chain between ferredoxins and flavodoxins, might be catalyzed by cytochrome P450 (CYP) proteins [27]. It was reported that the rubredoxins transfer metabolic reducing equivalents to oxygen or reactive oxygen species and act as electron carriers in oxidative stress responses, often accompanied by inhibition of ferredoxin expression. Our results also indicate that the HA10FeCl participate the ferredoxins and flavodoxins balance in M. bovis BCG, by decrease the CYP124 production and up-regulated the rubredoxin to adapt the growth condition.
As well known, the virulence mycobacterial cell wall lipids PDIM and PGL, were considered as the potential targets for antituberculosis drug development. The phenolphthiocerol (phthiocerol) moiety biosynthesis of PGL (PDIM) contains several genes ppsA-E and pks15/1, encoding type I polyketide synthases (PKS), and they are highly conserved in PDIM/PGL producing mycobacteria strains, e.g. M. tuberculosis H37Rv, M. bovis BCG (Pasteur 1173P2), M. marinum (M strain).The pks1 expression is also correlated with other genes, like fadD22, Rv2949c, lppX, fadD29, pks6 and pks12 [28]. The largest open reading frame (pks12) in the genome of M. tuberculosis H37Rv encodes probable polyketide synthase needed to produce fatty acids, probably involved in the synthesis of phthiocerol, the diol required for DIM synthesis [29]. Indeed, the phthiotriol/phenolphthiotriol dimycocerosates methyltransferase encoded by the gene BCG_2973 (Rv2952 in M. tuberculosis) encoded could catalyze the reduction of phthiodiolone and phenolphthiodiolone to yield phthiotriol and phenolphthiotriol in M. tuberculosis [30]. The proteins from the proteomic profiles encoded by the gene pks12 and BCG_2973 were significantly up-regulated also indicated that the HA10FeCl could affect the virulent PDIM/PGL biosynthesis, further verify the sensibility of HA10FeCl to PDIM negative M. tuberculosis in the experimental conditions.
Additionally, the two-component regulatory system (2CRS) SenX3-RegX3 is required for M. tuberculosis virulence. During phosphate depletion and nutrient starvation, the phosphate-specific transport operon pstS3-pstC2-pstA1 was induced and dependent on the (2CRS) SenX3-RegX3[29]. The disruption of the 2CRS induced the phosphate-specific transport gene pstC2 downregulated and the phosphate-starved bacilli became phenotypically tolerant to isoniazid, further indication the role of PstC2 in stressful condition [29,30]. The up-regulated PstC2 in HA10FeCl-treated bacilli suggest that the HA10FeCl could increase the susceptibility to isoniazid and their combination could fight against the isoniazid resistant M. tuberculosis.
Furthermore, the proteins PE15 and MmpL9a in HA10FeCl-treated M. bovis BCG were not identified, probably due to the lower expression, compared with the control. As well known, the M. tuberculosis is surrounded by a highly impermeable outer cell wall that is composed primarily of the complex PDIM that form an outer membrane. Due to the absence of typical porins, like the β-barrel porins in Gram-negative bacteria, transports small molecules such as nutrients, metabolites, Ca2+ across the outer membrane need the special transporters in M. tuberculosis. The PE/PPE proteins were essential for Ca2+ efficient uptake as a specific channel[31]. Here, we observed that the PE15 was absolutely decrease in the HA10FeCl-treated bacilli from the proteomic profiles, suggesting that the PE15 could be one of the targets. The lower expression of MmpL9a (BCG_2361) in the HA10FeCl-treated bacilli also suggested that it could be related to the linezolid resistance of the complex HA10FeCl on M. bovis BCG.
5. Conclusion
In summary, out of the HA and their Fe(II/III), Ni(II), Cu(II) and Zn(II) complexes evaluated for their antimycobacterial activity, the most promising growth inhibitors were some Fe(III), Cu(II) and Zn(II) complexes (HA10FeCl, HA10Fe3, HA8Fe3, HA10Zn2, HA12Cu2) which exhibited the highest antibacterial activity against pathogen mycobacteria. Their mechanisms of action showed from the proteomic profiles could be involved the cell wall lipids biosynthesis, drug resistance and tolerance, as well as siderophore metabolism. Further investigation should focus on the verification the optimal targets based on the proteomic profiles.
Author Contributions
Conceptualization: D.Y; Methodology: D.Y., I.S.S, Y.F. Z., V.F.; Resources: D.Y., R.S.L., V.F., and F. D.; Investigations: D.Y., I.S.S, N.E.M, Y.F. Z., L.-N.D., H.P.L. and G.X.C.; Formal analysis: D.Y., I.S.S, Y.F. Z.; Funding acquisition: D.Y., V.F. and R.S.L.; Supervision: V.F., F.D., M.G. and R.S.L.; Writing-original draft: D.Y. and I.S.S.; Writing-review & editing: D.Y., I.S.S. R.S.L., M.G., V.F., and F.D. All authors have given approval to the final version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research in the biological section was funded by Natural Science Foundation of Shanxi Province(20210302123356).
Conflicts of Interest
The authors declare no conflict of interest.
References
- W.H. Organization, Global tuberculosis report 2022, (2022).
- Daffé, M.; Marrakchi, H. Unraveling the Structure of the Mycobacterial Envelope. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Astarie-Dequeker, C.; Le Guyader, L.; Malaga, W.; Seaphanh, F.-K.; Chalut, C.; Lopez, A.; Guilhot, C. Phthiocerol Dimycocerosates of M. tuberculosis Participate in Macrophage Invasion by Inducing Changes in the Organization of Plasma Membrane Lipids. PLOS Pathog. 2009, 5, e1000289–e1000289. [Google Scholar] [CrossRef]
- Soetaert, K.; Rens, C.; Wang, X.-M.; De Bruyn, J.; Lanéelle, M.-A.; Laval, F.; Lemassu, A.; Daffé, M.; Bifani, P.; Fontaine, V.; et al. Increased Vancomycin Susceptibility in Mycobacteria: a New Approach To Identify Synergistic Activity against Multidrug-Resistant Mycobacteria. Antimicrob. Agents Chemother. 2015, 59, 5057–5060. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, R.; Thakur, S.; Nehra, K. ct-DNA Binding and Antibacterial Activity of Octahedral Titanium (IV) Heteroleptic (Benzoylacetone and Hydroxamic Acids) Complexes. Int. J. Med. Chem. 2016, 2016, 1–11. [Google Scholar] [CrossRef]
- Bakulina, O.; Bannykh, A.; Levashova, E.; Krasavin, M. Conjugates of Iron-Transporting N-Hydroxylactams with Ciprofloxacin. Molecules 2022, 27, 3910. [Google Scholar] [CrossRef] [PubMed]
- L.A. McAllister, J.I. L.A. McAllister, J.I. Montgomery, J.A. Abramite, U. Reilly, M.F. Brown, J.M. Chen, R.A. Barham, Y. Che, S.W. Chung, C.A. Menard, M. Mitton-Fry, L.M. Mullins, M.C. Noe, J.P. O'Donnell, R.M. Oliver, 3rd, J.B. Penzien, M. Plummer, L.M. Price, V. Shanmugasundaram, A.P. Tomaras, D.P. Uccello, Heterocyclic methylsulfone hydroxamic acid LpxC inhibitors as Gram-negative antibacterial agents, Bioorg Med Chem Lett 22(22) (2012) 6832-8.
- Sow, I.S.; Gelbcke, M.; Meyer, F.; Vandeput, M.; Marloye, M.; Basov, S.; Van Bael, M.J.; Berger, G.; Robeyns, K.; Hermans, S.; et al. Synthesis and biological activity of iron(II), iron(III), nickel(II), copper(II) and zinc(II) complexes of aliphatic hydroxamic acids. J. Co-ord. Chem. 2023, 76, 76–105. [Google Scholar] [CrossRef]
- Coelho, T.S.; Halicki, P.C.B.; Silva, L.; Vicenti, J.R.d.M.; Gonçalves, B.L.; da Silva, P.E.A.; Ramos, D.F. Metal-based antimicrobial strategies against intramacrophageMycobacterium tuberculosis. Lett. Appl. Microbiol. 2020, 71, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, P.; Crumbliss, A.L.; Miller, M.J.; Möllmann, U. Synthesis and biological activity of saccharide based lipophilic siderophore mimetics as potential growth promoters for mycobacteria. BioMetals 2007, 21, 41–51. [Google Scholar] [CrossRef]
- Mavrikaki, V.; Pagonis, A.; Poncin, I.; Mallick, I.; Canaan, S.; Magrioti, V.; Cavalier, J.-F. Design, synthesis and antibacterial activity against pathogenic mycobacteria of conjugated hydroxamic acids, hydrazides and O-alkyl/O-acyl protected hydroxamic derivatives. Bioorganic Med. Chem. Lett. 2022, 64, 128692. [Google Scholar] [CrossRef]
- Majewski, M.W.; Cho, S.; Miller, P.A.; Franzblau, S.G.; Miller, M.J. Syntheses and evaluation of substituted aromatic hydroxamates and hydroxamic acids that target Mycobacterium tuberculosis. Bioorganic Med. Chem. Lett. 2015, 25, 4933–4936. [Google Scholar] [CrossRef]
- Carvalho, E.M.; Paulo, T.d.F.; Saquet, A.S.; Abbadi, B.L.; Macchi, F.S.; Bizarro, C.V.; Campos, R.d.M.; Ferreira, T.L.A.; Nascimento, N.R.F.D.; Lopes, L.G.F.; et al. Pentacyanoferrate(II) complex of pyridine-4- and pyrazine-2-hydroxamic acid as source of HNO: investigation of anti-tubercular and vasodilation activities. JBIC J. Biol. Inorg. Chem. 2020, 25, 887–901. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.; Valentini, D.; Zumla, A.; Maeurer, M. Evaluation of the efficacy of valproic acid and suberoylanilide hydroxamic acid (vorinostat) in enhancing the effects of first-line tuberculosis drugs against intracellular Mycobacterium tuberculosis. Int. J. Infect. Dis. 2018, 69, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Rens, C.; Laval, F.; Daffé, M.; Denis, O.; Frita, R.; Baulard, A.; Wattiez, R.; Lefèvre, P.; Fontaine, V. Effects of Lipid-Lowering Drugs on Vancomycin Susceptibility of Mycobacteria. Antimicrob. Agents Chemother. 2016, 60, 6193–6199. [Google Scholar] [CrossRef] [PubMed]
- D. Yang, G. D. Yang, G. Vandenbussche, D. Vertommen, D. Evrard, R. Abskharon, J.F. Cavalier, G. Berger, S. Canaan, M.S. Khan, S. Zeng, A. Wohlkonig, M. Prevost, P. Soumillion, V. Fontaine, Methyl arachidonyl fluorophosphonate inhibits Mycobacterium tuberculosis thioesterase TesA and globally affects vancomycin susceptibility, FEBS Lett 594(1) (2020) 79-93.
- Yang, D.; Klebl, D.P.; Zeng, S.; Sobott, F.; Prévost, M.; Soumillion, P.; Vandenbussche, G.; Fontaine, V. Interplays between copper and Mycobacterium tuberculosis GroEL1. Metallomics 2020, 12, 1267–1277. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Constant, P.; Yang, D.; Baulard, A.; Lefèvre, P.; Daffé, M.; Wattiez, R.; Fontaine, V. Cpn60.1 (GroEL1) Contributes to Mycobacterial Crabtree Effect: Implications for Biofilm Formation. Front. Microbiol. 2019, 10, 1149. [Google Scholar] [CrossRef] [PubMed]
- M. Tre-Hardy, C. M. Tre-Hardy, C. Nagant, N. El Manssouri, F. Vanderbist, H. Traore, M. Vaneechoutte, J.P. Dehaye, Efficacy of the combination of tobramycin and a macrolide in an in vitro Pseudomonas aeruginosa mature biofilm model, Antimicrob Agents Chemother 54(10) (2010) 4409-15.
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Boopathi, S.; Ramasamy, S.; Haridevamuthu, B.; Murugan, R.; Veerabadhran, M.; Jia, A.-Q.; Arockiaraj, J. Intercellular communication and social behaviors in mycobacteria. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef]
- Niño-Padilla, E.I.; Velazquez, C.; Garibay-Escobar, A. Mycobacterial biofilms as players in human infections: a review. Biofouling 2021, 37, 410–432. [Google Scholar] [CrossRef] [PubMed]
- Viljoen, A.; Dufrêne, Y.F.; Nigou, J. Mycobacterial Adhesion: From Hydrophobic to Receptor-Ligand Interactions. Microorganisms 2022, 10, 454. [Google Scholar] [CrossRef]
- Belardinelli, J.M.; Stevens, C.M.; Li, W.; Tan, Y.Z.; Jones, V.; Mancia, F.; Zgurskaya, H.I.; Jackson, M. The MmpL3 interactome reveals a complex crosstalk between cell envelope biosynthesis and cell elongation and division in mycobacteria. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Bailo, R.; Bhatt, A.; Aínsa, J.A. Lipid transport in Mycobacterium tuberculosis and its implications in virulence and drug development. Biochem. Pharmacol. 2015, 96, 159–167. [Google Scholar] [CrossRef]
- Kurthkoti, K.; Amin, H.; Marakalala, M.J.; Ghanny, S.; Subbian, S.; Sakatos, A.; Livny, J.; Fortune, S.M.; Berney, M.; Rodriguez, G.M. The Capacity of Mycobacterium tuberculosis To Survive Iron Starvation Might Enable It To Persist in Iron-Deprived Microenvironments of Human Granulomas. Mbio 2017, 8. [Google Scholar] [CrossRef]
- Sushko, T.; Kavaleuski, A.; Grabovec, I.; Kavaleuskaya, A.; Vakhrameev, D.; Bukhdruker, S.; Marin, E.; Kuzikov, A.; Masamrekh, R.; Shumyantseva, V.; et al. A new twist of rubredoxin function in M. tuberculosis. Bioorganic Chem. 2021, 109, 104721. [Google Scholar] [CrossRef]
- B. Ramos, S.V. B. Ramos, S.V. Gordon, M.V. Cunha, Revisiting the expression signature of pks15/1 unveils regulatory patterns controlling phenolphtiocerol and phenolglycolipid production in pathogenic mycobacteria, PLoS One 15(5) (2020) e0229700.
- Sirakova, T.D.; Dubey, V.S.; Kim, H.-J.; Cynamon, M.H.; Kolattukudy, P.E. The Largest Open Reading Frame ( pks12 ) in the Mycobacterium tuberculosis Genome Is Involved in Pathogenesis and Dimycocerosyl Phthiocerol Synthesis. Infect. Immun. 2003, 71, 3794–3801. [Google Scholar] [CrossRef] [PubMed]
- Siméone, R.; Constant, P.; Malaga, W.; Guilhot, C.; Daffé, M.; Chalut, C. Molecular dissection of the biosynthetic relationship between phthiocerol and phthiodiolone dimycocerosates and their critical role in the virulence and permeability of Mycobacterium tuberculosis. FEBS J. 2007, 274, 1957–1969. [Google Scholar] [CrossRef] [PubMed]
- V. Boradia, A. V. Boradia, A. Frando, C. Grundner, The Mycobacterium tuberculosis PE15/PPE20 complex transports calcium across the outer membrane, PLoS Biol 20(11) (2022) e3001906.
Figure 1.
Proteomic analysis of the HA10FeCl-treated M. bovis BCG versus untreated M. bovis BCG control. The volcano plot of proteomic data in the HA10FeCl-treated M. bovis BCG compared to control. Three independent samples for each group were analysed for proteomic analysis.
Figure 1.
Proteomic analysis of the HA10FeCl-treated M. bovis BCG versus untreated M. bovis BCG control. The volcano plot of proteomic data in the HA10FeCl-treated M. bovis BCG compared to control. Three independent samples for each group were analysed for proteomic analysis.
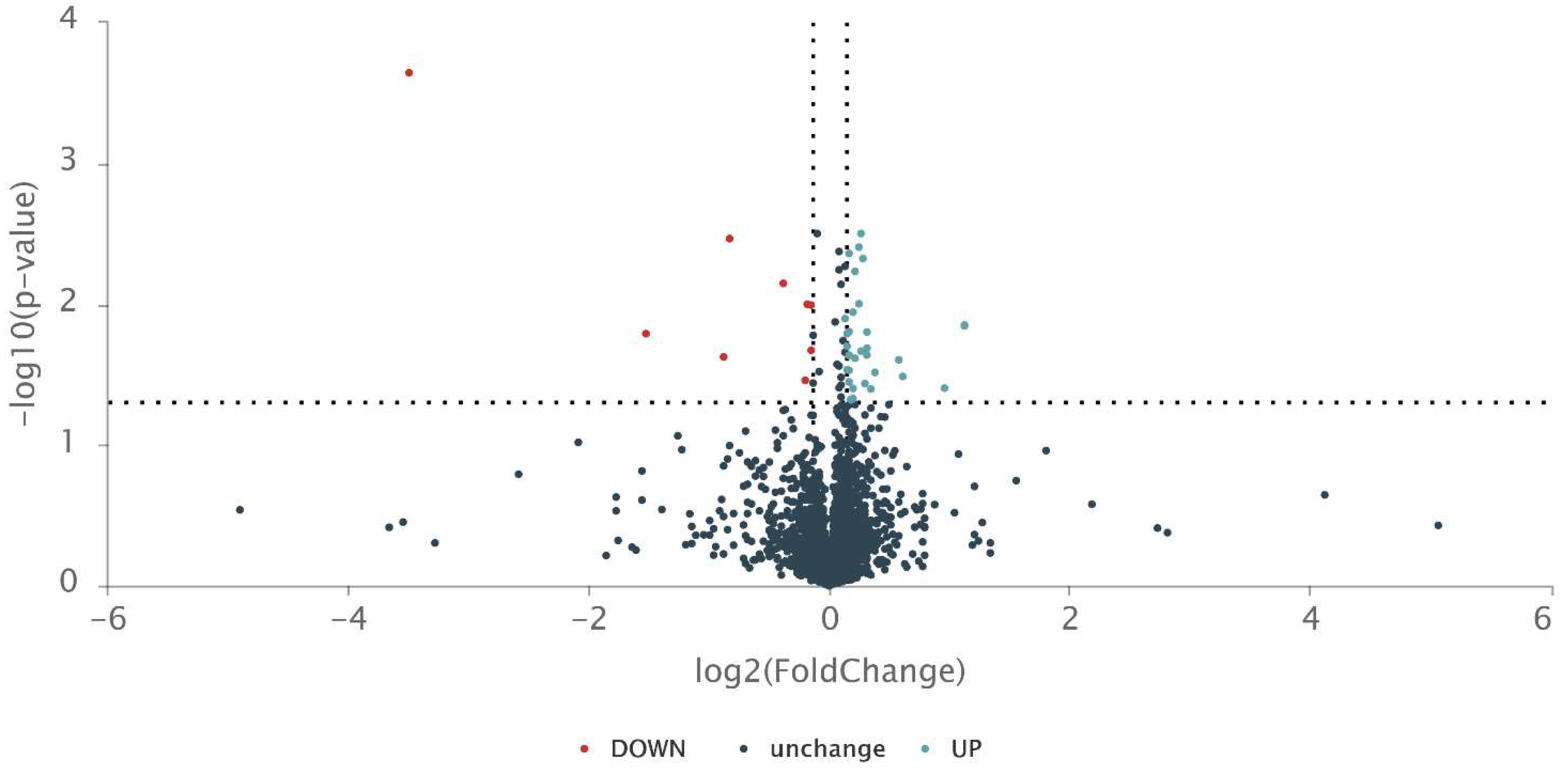
Figure 2.
Proteomic analysis of the HA10FeCl-treated M. bovis BCG versus untreated M. bovis BCG control. The heatmap showing differentially expressed proteins in HA10FeCl-treated M. bovis BCG and control. Colours changing from blue to red indicate increased levels of proteins. Gray color indicates protein not detected in proteomic profiles. Three independent samples for each group were analysed for proteomic analysis.
Figure 2.
Proteomic analysis of the HA10FeCl-treated M. bovis BCG versus untreated M. bovis BCG control. The heatmap showing differentially expressed proteins in HA10FeCl-treated M. bovis BCG and control. Colours changing from blue to red indicate increased levels of proteins. Gray color indicates protein not detected in proteomic profiles. Three independent samples for each group were analysed for proteomic analysis.
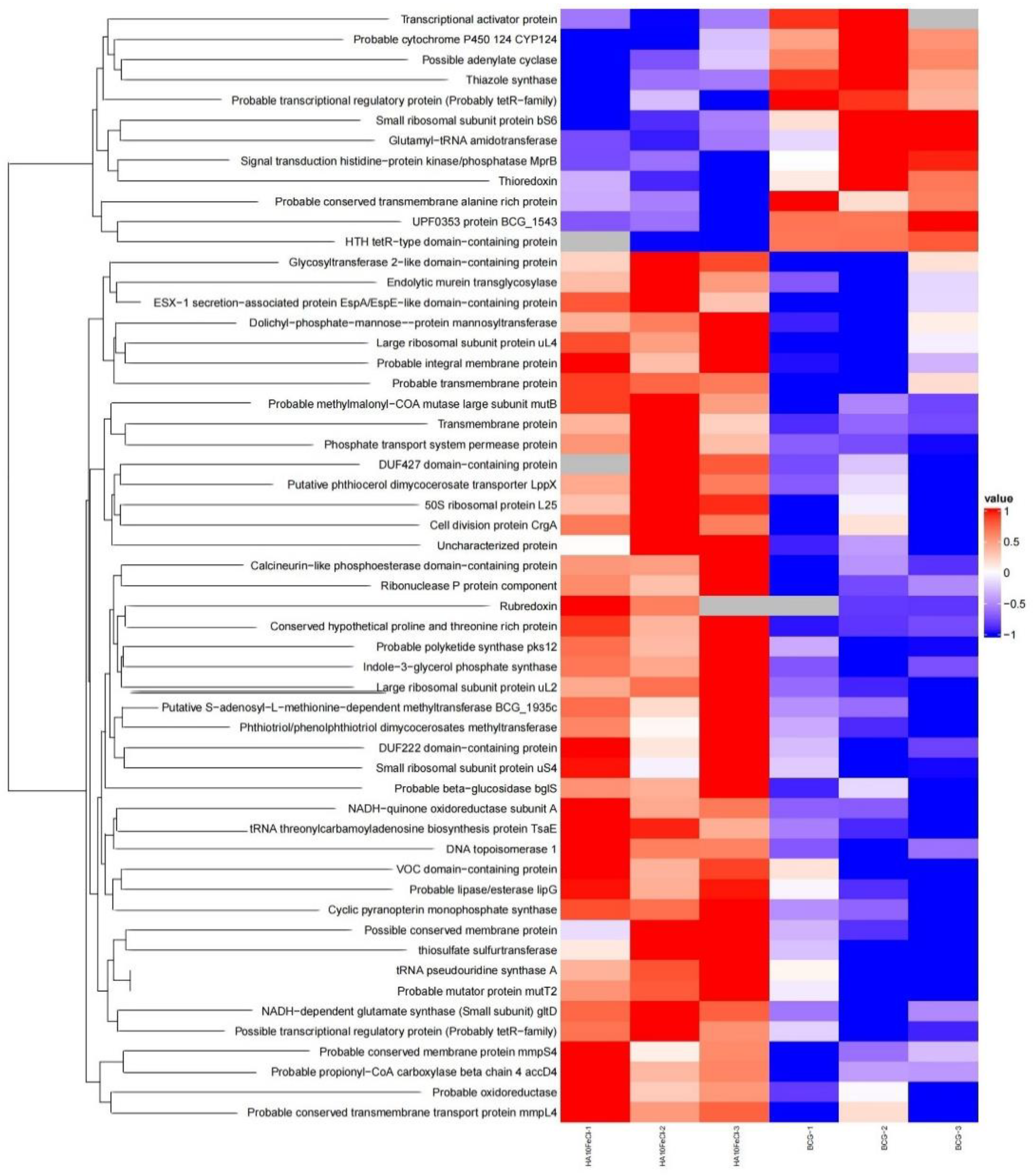

Table 1.
MIC and MBIC values (µM) of HA iron complexes against M. bovis, M. marinum and M. tuberculosis.
Table 1.
MIC and MBIC values (µM) of HA iron complexes against M. bovis, M. marinum and M. tuberculosis.
| Compound | M. bovis BCG | M. marinum | M. tuberculosis H37Ra | |||
|---|---|---|---|---|---|---|
| MIC | MBIC | MIC | MBIC | MIC | MBIC | |
| µM | µM | µM | µM | µM | µM | |
| HA2FeCl | > 500 | 500 | > 500 | 250 | > 500 | 250 |
| HA6FeCl | > 500 | 300 | > 500 | ≥ 250 | 500 | 125 |
| HA8FeCl | ≥ 500 | 100 | 200 | 62.5 | 500 | 250 |
| HA10FeCl | 100-200 | 20-100 | > 200 | 62.5 | 125 | 31.25 |
| HA12FeCl | > 200 | 100 | > 200 | 500 | 125 | 125 |
| HA17FeCl | > 500 | > 500 | > 500 | 500 | 250 | > 500 |
| HA2Fe2 | > 500 | > 500 | > 500 | > 500 | > 500 | > 500 |
| HA6Fe2 | > 500 | 100-500 | > 500 | 500 | 500 | 250 |
| HA8Fe2 | > 500 | 100-200 | > 500 | 500 | 500 | 250 |
| HA10Fe2 | > 200 | 20 | > 200 | > 500 | 250 | > 500 |
| HA12Fe2 | > 500 | 100 | > 500 | > 500 | 500 | 500 |
| HA17Fe2 | 250 | > 500 | > 500 | 500 | 125 | 250 |
| HA2Fe3 | 250-500 | 300-500 | > 500 | 250 | > 500 | 125 |
| HA6Fe3 | 500 | 100-500 | > 500 | 250 | ≥ 500 | 62.5-125 |
| HA8Fe3 | 500 | 100 | 250-500 | ≥ 125 | 125 | 31.25 |
| HA10Fe3 | ≥ 200 | 20-40 | > 200 | 62.5 | 125 | 31.25-62.5 |
| HA12Fe3 | ≥ 200 | 100 | > 200 | 200-500 | 125-250 | 125 |
| HA17Fe3 | > 200 | > 500 | > 500 | 200-500 | 125 | 62.5 |
HA= hydroxamic acid, HAnFe2= iron(II) complexes, HAnFeCl= iron(III) complexes with n=2, 6, 8, 10, 12 and 17 (carbon chain).
Table 2.
MIC and MBIC values (µM) of zinc, nickel(II) and copper(II) complexes against M. bovis BCG, M. marinum and M. tuberculosis.
Table 2.
MIC and MBIC values (µM) of zinc, nickel(II) and copper(II) complexes against M. bovis BCG, M. marinum and M. tuberculosis.
| Compound | M. bovis BCG | M. marinum | M. tuberculosis H37Ra | |||
|---|---|---|---|---|---|---|
| MIC | MBIC | MIC | MBIC | MIC | MBIC | |
| µM | µM | µM | µM | µM | µM | |
| HA2Zn2 | > 500 | > 500 | > 500 | 500 | > 500 | 250 |
| HA6Zn2 | > 500 | 200 | > 500 | 500 | 125 | 125 |
| HA8Zn2 | > 500 | 100-200 | > 500 | 500 | 250 | 250 |
| HA10Zn2 | 250 | 100 | 250 | 250-500 | 250 | 250 |
| HA12Zn2 | > 500 | 100 | > 500 | 250 | 250 | 125 |
| HA17Zn2 | > 500 | > 500 | > 500 | 500 | > 500 | 250 |
| HA2Ni2 | > 500 | 300-500 | > 500 | > 500 | > 500 | 62.5-125 |
| HA6Ni2 | > 500 | 200 | > 500 | 500 | > 500 | 31.25 |
| HA8Ni2 | > 500 | 100 | > 500 | 62.5 | > 500 | 31.25 |
| HA10Ni2 | > 200 | 20 | > 200 | 500 | 500 | 125 |
| HA12Ni2 | > 500 | 100 | > 500 | 125 | > 500 | 250-500 |
| HA17Ni2 | > 500 | > 500 | > 500 | 250 | > 500 | 250 |
| HA2Cu2 | 250-500 | 300 | > 500 | ≥ 250 | ≥ 500 | 250 |
| HA6Cu2 | ≥ 500 | 100 | > 500 | 500 | > 500 | 500 |
| HA8Cu2 | > 200 | 100-200 | > 200 | 250 | > 500 | 250 |
| HA10Cu2 | > 200 | 20 | > 200 | 250 | 250 | 250 |
| HA12Cu2 | > 500 | 100 | > 500 | 125 | > 500 | 250 |
| HA17Cu2 | > 500 | ≥ 500 | > 500 | ≥ 500 | > 500 | 250 |
HAZn2=zinc complexes, HAnNi2= nickel(II) complexes, HAnCu2= copper(II) complexes with n=2, 6, 8, 10, 12 and 17 (carbon chain).
Table 3.
MIC and MBIC values (µM) of HA complexes, iron(II)-, iron(III)-, nickel(II)-, copper(II)-, zinc(II)- chloride against M. bovis, M. marinum and M. tuberculosis.
Table 3.
MIC and MBIC values (µM) of HA complexes, iron(II)-, iron(III)-, nickel(II)-, copper(II)-, zinc(II)- chloride against M. bovis, M. marinum and M. tuberculosis.
| Compound | M. bovis BCG | M. marinum | M. tuberculosis H37Ra | |||
|---|---|---|---|---|---|---|
| MIC | MBIC | MIC | MBIC | MIC | MBIC | |
| µM | µM | µM | µM | µM | µM | |
| HA2 | > 500 | > 500 | > 500 | 500 | > 500 | 250 |
| HA6 | > 500 | ~ 100 | > 500 | 500 | 500 | 125 |
| HA8 | > 500 | 100 | > 500 | 500 | 500 | 125 |
| HA10 | 250-500 | 100 | 250-500 | 250 | > 500 | 125 |
| HA12 | > 500 | 100-500 | > 500 | > 500 | 125 | 250 |
| HA17 | > 500 | > 500 | > 500 | ≥ 500 | ≥ 500 | > 500 |
| FeCl2 | > 500 | > 500 | > 500 | 500 | > 500 | > 500 |
| FeCl3 | > 500 | > 500 | > 500 | 500 | > 500 | > 500 |
| NiCl2 | > 200 | > 500 | 500 | ≥ 500 | > 500 | 250 |
| CuCl2 | > 200 | 200 | > 200 | 250 | > 500 | 500 |
| ZnCl2 | > 500 | > 500 | > 500 | 250 | > 500 | 125-250 |
HA complexes with n=2, 6, 8, 10, 12 and 17 (carbon chain).
Table 4.
Vancomycin susceptibility assay obtained in macrodilution series and analysed by the checkerboard method for M. bovis BCG strain. .
Table 4.
Vancomycin susceptibility assay obtained in macrodilution series and analysed by the checkerboard method for M. bovis BCG strain. .
| Compounds | MIC (μg/mL)/FICI |
|---|---|
| Vancomycin | 750/- |
| HA10FeCl | 48.18-96.37/- |
| HA10FeCl (46.35 μg/mL) + Vancomycin | 125/0.176 |
| Vancomycin (50 μg/mL) + HA10FeCl | 11.59/0.125-0.25 |
-: means without calculation (it is not possible to get the FICI in the presence of only one drug in the experiment according to the equation 1).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



