
Submitted:
02 September 2023
Posted:
04 September 2023
You are already at the latest version
Abstract
Dioscorea remotiflora, a perennial climbing herbaceous plant wild and native to México, bears tubers of important ethnobotanical and nutritional value. However, the information for its production and use is precarious. The objective of this research was to characterize the adaptation habitats of D. remotiflora in order to determine its ecological descriptors and current distribution, as well as to model its potential distribution. A comprehensive database encompassing 481 geo-referenced accessions was assembled. Using the Agroclimatic Information System for México and Central America (SIAMEXCA), 42 environmental variables were formulated. The MaxEnt model within the Kuenm R-package was employed to predict the species distribution. The findings reveal a greater presence of D. remotiflora in harsh environments, characterized by arid to semi-arid conditions, poor soils, and hot climates with long dry periods. The plant's nutritional and medicinal attributes, combined with its ecological adaptability, suggest its viability within evolving regional cropping systems under the influence of climate change. Niche modeling revealed that seven key variables determine the geographical distribution of D. remotiflora: precipitation of the warmest quarter, precipitation of the driest month, minimum temperature of the coldest month, November-April solar radiation, annual mean relative humidity, annual moisture availability index, and May-October mean temperature. Favorable regions for D. remotiflora coincide with its current presence sites, while other suitable areas, such as the Yucatan Peninsula, Northeast region, and Gulf of México, offer potential expansion opportunities for the species distribution. The characterization obtained for D. remotiflora, together with the description of its edaphic and climatic adaptation habitats, will enable an efficient design of strategies for its use and conservation in the near future.
Keywords:
Dioscorea remotiflora
; Mexican endemic species
; Niche modeling
; Ecological descriptors
; Climatic adaptation
1. Introduction
Dioscorea remotiflora is a monocotyledonous, perennial, and climbing plant, with heart-shaped leaves, dioecious flowers, and seeds in axillary clusters. It belongs to the Dioscoreaceae family [1] and is one of the native species of México from the genus Dioscorea [2,3]; currently, it is considered an endemic species [4]. Although D. remotiflora is not a cultivated plant, its roots, tubers, and rhizomes are collected since prehistoric times to be used as food [5], they contain 85% carbohydrates, 7.35% proteins, 3.76% lipids, and 3.68% minerals (K, Fe, Na, and Mg), therefore, it is used as a healthy snack or even as a gourmet dish [6,7]. The tubers contain secondary metabolites such as steroids, saponins, and diosgenin, though in low concentration [7], thus, they are not used for the extraction of diosgenin. Currently, D. remotiflora is considered an underutilized species; being a factor that contributes to this, the lack of research on this species; Up to now, the reports regarding the description of the ecology, climatic adaptation, and potential distribution of D. remotiflora are bare and insufficient; this hinders the design of strategies for its conservation and optimal use [8,9]. Therefore, the objective of this research was to use the occurrence data of D. remotiflora, in its natural distribution areas, to conduct an eco-geographical analysis in order to elucidate the contribution of several ecological descriptors in determine its current distribution, identify its adaptation patterns, and develop optimal Maxent models of potential geographic distribution, through the use of the Kuenm R-package, which automates the creation, calibration, and evaluation of ecological niche models [10].
Plant growth and distribution are the results of the species’ response to the environmental complex that prevails in the occurrence sites [11,12], where aspects of climate, soil, vegetation, and others concur. As a species moves from its center of origin to other geographic regions, it finds it necessary to adapt to different environmental conditions. Whether the adaptation process is successful, then the species will have colonized new territories, extending its distribution, and thus, its adaptation environmental scope, which usually triggers the increase of its tolerance to abiotic and biotic stress [13]; this aspect may not be manifested uniformly in all the eco-geographic populations of the species [14], and, moreover, the novel traits may be the result of the action of genetic or epigenetic adaptation processes [15,16] with the possibility of simultaneous and complementary action of both mechanisms [16].
D. remotiflora is distributed mainly in the central, southern, and western regions of México [17], generally in dry deciduous tropical forests [18,19], which indicates its adaptive capacity to diverse environments [20]. This is why characterizing the ecogeography of a plant species considering their all accessions and populations is a key aspect.
With the presence of climate change in agricultural areas, and the increasing environmental stress for crops, food production must face the challenge with productive plant species that are resistant to drought and heat (main types of abiotic stress in México; [21]. However, in various regions of México, the crop pattern is reduced due to a short growing season with a low availability of rainwater [22]. In these cases, it is necessary to explore new crop species, for which a characterization study is required such as the one addressed in the present investigation for D. remotiflora.
2. Results
2.1. Selection of environmental variables
2.2. Current distribution, climatic adaptation and ecological descriptors
Figure 1 shows the current distribution of D. remotiflora in the agroclimatic regions of México, as shown, this species is found predominantly in the areas near the Mexican Pacific coast, mainly in the central and southern portions. A greater presence of D. remotiflora is observed in the agroclimatic regions: Humid-subhumid semi-warm (98 accessions), Dry-subhumid semi-warm (88 accessions), Humid-subhumid warm (67 accessions, and Dry-subhumid warm (66 accessions). However, D. remotiflora is present in 15 of the 29 agroclimatic regions of México (Table 3), which enables this species to develop in thermal zones from semi-cold to very warm and in hydric zones from semi-arid to humid.
Table 4 shows the FAO soil units and the textural classes in which D. remotiflora is distributed. As can be seen, most of the occurrence sites of this species are distributed in the following soil types: Lithosol (144 accessions), Calcaric Regosol (98 accessions), Eutric Regosol (99 accessions) and Haplic Faozem (45 accessions).
Figure 2.
Current distribution of D. remotiflora in soil units of México.
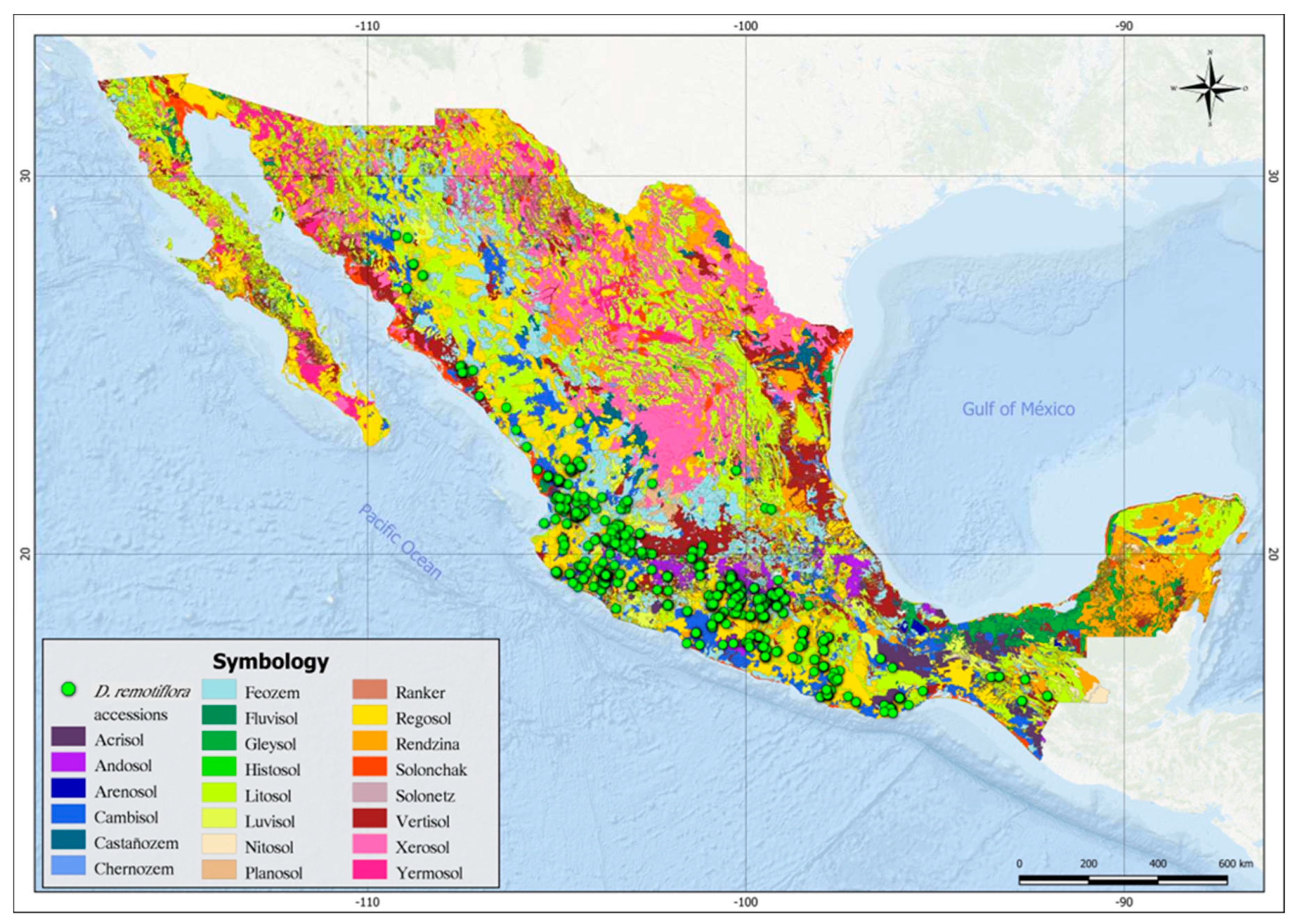
Table 5 shows the ecological descriptors of D. remotiflora, which contain the environmental adaptation ranges (RAA) of the species (minimum and maximum values of environmental variables that allow the presence of D. remotiflora, and the optimal environmental range (RAO), which makes possible the higher frequency of occurrence sites of this plant. According to the information in Table 5, D. remotiflora is distributed in environments with an annual moisture availability index (MAI) ranging from 0.27 to 2.32, with an optimal range of 0.40 to 0.99, which corresponds to an RAA and RAO of an annual precipitation interval of 444 to 2886 and 700 to 1299 mm, respectively. This plant prefers areas where precipitation in the wettest quarter ranges from 400 to 884 mm, although it tolerates conditions from 240 to 1204 mm. This species can tolerate a long season (November-April) with low precipitation, even with an accumulation of only 23 mm in those six months (Table 5). The optimum growing season goes from 120 to 150 days, although it grows in regions with a growing season as long as 120-190 days.
Altitudinally, D. remotiflora is present from 6 to 4295 masl, but most of the presence sites occur between 200 and 800 masl. This encourages this species to develop in areas with an average annual temperature between 14.7 and 28.5°C, with an optimum of 19 to 27°C, an extreme monthly average maximum temperature of 41.2°C (Maximum maximorum temperature) and an extreme monthly average minimum temperature of 1.7°C (Minimum minimorum temperature). Both in the seasonal periods May-October and November-April, the interval in which the greatest number of accessions occurs is from 19 to 26°C, very similar to the optimum average annual temperature. Regarding soil properties, D. remotiflora is present in coarse, medium and fine textured soils, but prefers its presence is more abundant in coarse-textured soils, the number of occurrence sites decreases markedly in medium and fine-textured soils, indicating a clear preference of the species for soils with excellent drainage.
2.3. Modeling distribution niches of D. remotiflora
Due to Maxent not being exempt from the effects of collinearity and the fact that these can intervene in the estimation of the factors, as well as inferring the uncertainties when the models are transferred spatially and temporally [25], and with the purpose of obtaining an accurate model with a reduced number of variables, eliminating the possibility of obtaining an over-fit or over-parameterized model[10]; the Kuenm R-package allowed, through ecological niche modeling, optimizing the number of environmental variables, leaving only 7, due to their greater contribution to the presence and distribution of D. remotiflora. The results of the Jackknife statistical test of factorial importance reported that the most determining environmental variables in the presence and distribution of D. remotiflora are: precipitation of the warmest quarter (42.4%), mean precipitation of the driest month (17.5%), minimum temperature of the coldest month (15%), November-April mean solar radiation (10%), annual mean relative humidity (8.5%), annual moisture availability index (5.7%), and May-October mean temperature (0.9%) (Table 6).
Based on the results obtained through the Jackknife test, three sets of cases were analyzed: "only with variable", "without variable" and "with all variables" [26], thus revealing the effects of environmental variables in the appropriate range of D. remotiflora (Figure 3). Among these environmental variables, it was found that solar radiation from November to April was the most relevant, with a regularized training gain greater than 0.8. In addition, other variables of significant importance were identified, such as the precipitation of the warmest quarter, the annual humidity index, relative humidity, the minimum temperature of the coldest month, the precipitation of the driest month, and the average temperature from May to October. In all these cases, the regularized training gains exceeded 0.6.
Kuenm R-package enabled to obtain 372 models (Figure 4), all of them significant, however, only two models positively met the Akaike criterion (AIC=0) and a maximum omission rate of 5%. The Kuenm R-package implemented with MaxEnt determined that 3.0 was the optimum regularization multiplier. The model finally selected to depict the distribution of D. remotiflora was judged excellent since the AUC of the ROC curve accounted for 0.935.
Figure 5 shows the current and potential distribution of D. remotiflora based on thresholding the environmental suitability with the “Balance training omission, predicted area, and threshold value” criterion. The map in Figure 4 shows four types of areas: areas not suitable (white color), areas with low environmental suitability (light green), areas with medium suitability (dark green), and areas highly suitable for D. remotiflora presence (dark grey), which matches the largest number of occurrence sites. The total potential distribution area for D. remotiflora accounted 428,747.68 km2, distributed as follows: The areas with low suitability account for 150,547.104 Km2 and are located in Sonora, north Sinaloa, Guanajuato, Querétaro, México State, coasts of Guerrero, center Oaxaca, Chiapas, Campeche, San Luis Potosí, south Tamaulipas, and Veracruz. The areas with medium suitability cover 127,817,096 km2 and locate in southern Sonora, southern Chihuahua, northern Sinaloa, the coasts of Nayarit, Jalisco, Michoacán, Guerrero, Oaxaca, and Chiapas. The highly suitable areas resulted in 150,383.48 km2 (dark gray), and are located in Sinaloa, Nayarit, Jalisco, Colima, Michoacán, Morelos, little portions of Puebla and Tamaulipas, northern Guerrero, and the coasts of Oaxaca. As can be seen in Figure 4, there are potential areas in regions where the presence of D. remotiflora has not yet been reported, which indicates territories that could be the target of future exploration of new populations. This is the case of the Peninsula of Yucatán, center Veracruz, southeastern San Luis Potosí, southern Tamaulipas, Guanajuato, northeastern Jalisco, northern Sinaloa, northern Hidalgo, and western México state.
3. Discussion
3.1. Current distribution, climate adaptation and ecological descriptors
Most of the D. remotiflora occurrence sites are concentrated amongst the dry-subhumid and humid-subhumid agroclimatic regions with the warm and semiwarm variants, with an annual mean temperature between 19 and 27°C and an annual moisture availability index (MAI) between 0.4 and 0.99 (Table 4). These results agree with the ones reported by [2] who indicate that D. remotiflora is distributed in areas with semi-warm, warm and sub-humid climates, ranging from the northern portion to the central region of México. However, an interesting aspect is the climatic extremes in which D. remotiflora can adapt, meaning that even when most of the occurrence sites of this species are located between 700 and 1299 mm of annual rainfall, there are populations that subsist with 444 mm per year, and on the other extreme, other populations do it in sites with 2886 mm of precipitation per year. When combined with annual potential evapotranspiration data, these precipitation values translate into MAI values from 0.27 to 2.32, which according to arid zones scheme from UNEP (Middleton & Thomas, 1997; adapted by Ruiz et al., 2004); match with semiarid to very wet lands. Regarding this, [28] mentions that some species of Dioscorea adapt to dry periods and can survive under conditions of water deficit better than many other species and crops. This seems to be the case of D. remotiflora, according to the ecological indicators obtained. The presence of populations of D. remotiflora in extremely humid sites (2886 annual mm and MAI > 2) could be explained in sites with excellent soil drainage or to consider those data as an indicator of the ability of this species to colonize habitats typically unsuitable for its development [18].
Regarding the variable precipitation of the warmest quarter, which was the most significant for the presence of D. remotiflora in the MaxEnt modelling, Table 4 indicates a range of 240 to 1,204 mm, with an optimal range of 400-884 mm. Considering then that D. remotiflora is present in sites with a minimum annual rainfall of 444 mm, and that, from those millimeters at least 240 must occur in the warmest quarter [29](Avalo and García, 2021), we can conclude that this plant has the ability to adapt and develop in environments with an irregular distribution of precipitation, which may also represent a comparative advantage of D. remotiflora in relation to other species. This also leads to conclude that D. remotiflora is a species that adapts more to moderate to low humidity conditions, thus, the very humid orarid environments are not conducive to high productivity of this species. The results obtained also coincide with previous reports related to the adaptation of the genus Dioscorea to tropical and subtropical zones, with tolerance of some species to conditions of water deficit [28,30].
The environmental characterization of the presence sites of D. remotiflora shows that there is a greater number of accessions of this plant in Lithosol, calcaric Regosol and eutric Regosol soils, which do not offer the best conditions for the development of vegetation and crops [31]. According to [32], Leptosols (Litosols and Rendzinas) represent 28.3% of the Mexican territory and are characterized by very thin, stony and poorly developed soils, that can contain a large amount of calcareous material, which immobilizes mineral nutrients. They are common in mountainous areas and on shallow limestone plains. Their agricultural potential is limited by their shallow depth and high compaction, which makes them difficult to work on. On the other hand, Regosols are considered very young soils that develop on unconsolidated material, light in color and poor in organic matter. They are common in arid, semi-arid (including the dry tropics) and montaneous regions, they can be found associated to Leptosols and with rock or tepetate outcrops. From the above, it can be deduced that D. remotiflora has a great adaptive capacity to poor soils, with non-optimal agroclimatic conditions for the rest of the species, which may be an attribute that makes this species an alternative for cultivation in regions where climate change is deteriorating the environmental conditions of agricultural production systems [33].
Regarding soil texture, according to Table 4, D. remotiflora prefers coarse-textured soils, typical of Lithosol and Regosol soils, which do not store a large amount of water. Unlike other species of the Dioscorea genus, D. remotiflora is susceptible to tuber putrescence [18], therefore it requires well-drained soils. The presence of D. remotiflora detected in medium-textured soils is related to the occurrence of lower annual precipitation levels (735.6 mm on average) than those of sites with coarse-textured soils (1034 mm on average), which ensures that even in soils that store more moisture, it is possible for this species to adapt, as long as the volumes of precipitation are not high; this compensatory effect has been previously reported for diverse crop species [34].
According to table 4, D. remotiflora can tolerate an extreme monthly average minimum temperature of 1.7°C and, on the other hand, it can survive an extreme monthly average maximum temperature of 41.7°C, with annual thermal oscillations ranging from 10 to 20° C and with an optimal annual thermal range of 13 to 16°C, fact that shows the wide range of thermal conditions in which this plant can survive, and this includes temperature regimes that are classified as very extreme [35]. Other species of the genus Dioscorea have shown tolerance to extreme thermal environments, such as D. divaricata, which can tolerate temperatures as low as -18°C. The occurrence of these extreme minimum temperatures, however, causes delayed full maturity and reached until 3 to 4 years after [36].
On the other hand, extreme temperatures are generally considered a source of dormancy in postharvest tubers and seeds [37]. [28] mentions that the dormancy of the tubers of the Dioscorea species lasts 120 days, which limits their agricultural production; this indicates an aspect that should be worked on in the immediate future to make D. remotiflora a more promising agricultural species. [37] report that tuberization is induced by environmental cues such as short days, low temperatures, and higher soil moisture content.
Another important aspect in the development of this species is photoperiod since it intervenes in the formation and growth of leaves and tubers; there are differences among Dioscorea species in relation to their response to photoperiod. In the case of D. remotiflora, in the long-day season, which is from May to October, foliar growth is favored, and in short days, the growth and swelling of the tuber are stimulated, promoting the production and storage of starches [38]. In Table 4, it can be observed that the optimal range of photoperiod for exhibiting adequate leaf growth is from 12.6 to 12.9 h, and for good tuber development the optimal range is from 11.10 to 11.39 h.
3.2. Modeling of distribution niches of D. remotiflora
The Kuenm R-package implemented with MaxEnt, allowed to obtain an optimal niche model to appropriately depict the D. remotiflora distribution in México; based on the requirements of statistical significance, the optimal regularization multiplier, the feature classes, and the omission rate established, the analysis process with Kuenm produced two possible models (Figure 4). One of them was selected, which can be considered a good decision since it fulfills the requirements established [39], and the AUC of the ROC curve accounted for a value greater than 0.93 [40]. The AUC value is an important tool to assess model performance, the higher the AUC value (closer to 1) the better the model performance is [41,42]. The Jackknife test identified the most determining variables for the presence of D. remotiflora, the precipitation of the warmest quarter, precipitation of the driest month, minimum temperature of the coldest month, November-April solar radiation, annual mean relative humidity, annual moisture availability index, May-October mean temperature. However, the Jackknife test indicates that the most important variable is solar radiation from November to April (Figure 3). These results agree with what was reported by [43], who mentions that the production of diosgenin, corticosteroids, carbohydrates, and other compounds produced by these species are subject to solar radiation. On the other hand, these results agree with those reported by Viruel et al. (2014) who pointed out that the environmental variables of major influence for the presence of Dioscorea humilis are the precipitation of the wettest quarter, the precipitation of the warmest quarter, and the precipitation of the coldest quarter. On the other hand, [43] report that for 10 species of the Dioscorea genus, the variables that are most important and intervene in the production of secondary metabolites are annual precipitation and average annual radiation.
The capabilities observed in the present research by D. remotiflora, to develop in adverse environments, could be explained through the ability to adjust its metabolism and modify its morphological characteristics, such as the size and thickness of the leaf, allowing it to adapt to extreme conditions, where other species fail to thrive [44,45].
The current distribution of D. remotiflora obtained in the present research (Figure 4), agrees with that reported by [46], who mentions that this species is distributed in Chiapas, Chihuahua, Colima, Durango, Guanajuato, Jalisco, Oaxaca, Guerrero, Michoacán, Guerrero, Nayarit, Puebla, Tamaulipas, Tabasco, and Zacatecas; which represents areas along the foot of the Sierra Madre Occidental, the Sierra Madre del Sur and its confluence with the Transversal Neovolcanic Axis, where coniferous and oak forests are found, these two, are home to herbaceous and forest communities with presence of endemism [47]. However, the distribution, diversity, and structure of populations are strongly influenced by historical, geographical, and climatic events [48], which also trigger speciation processes [49,50]. These arguments explain the number of species of the genus Dioscorea and the differences between them in terms of environmental ranges. On the other hand, the areas of high environmental suitability are located on the Mexican Pacific coast; this location suggests that optimal conditions of agroclimatic variables exist in these areas for the development of this plant. In contrast, in the areas of medium and low environmental suitability, it is possible that the species is adapting to agroclimatic ranges that go from very arid to very humid, moving to the central part of México. These areas, therefore, have the potential to establish crops of the species. However, it is important to note that the classification of environmental suitability areas may be modified by the effects of climate change [51], thus, a complementary research regarding potential distribution áreas under climate change scenarios would be required.
3.3. D. remotiflora cultivation prospects
Currently, D. remotiflora is a plant that is exploited for human consumption, but basically through the collection of tubers in its natural habitat (Figure 5). However, it presents favorable characteristics to integrate into regional crop patterns, given its comparative advantages regarding other plant species, such as its adaptation and production in poor, shallow, and infertile soils, as well as its tolerance to drought, since it does not have a high water requirement for its development. Besides, some characteristics inherent to the species should be taken into account before considering it, as a cultivation option, such as the fact that sexual propagation is not considered a viable alternative up to now, due to the fact that the seeds it produces are attacked by pests, damaging 60 to 80% of them; thus, the health of the seeds should be ensured through pest control or opting for asexual propagation through the sprouting of tubers. However, the tubers have a prolonged dormancy of up to 120 days, which is a limitation to program their propagation. This leads to the need of exploring some of the available techniques to reduce the duration of dormancy and accelerate the germination process.
4. Materials and Methods
4.1. Occurrence data
1,030 geo-referenced accessions from herbaria, floristic inventories, scientific articles and databases were identified, later on, they were reviewed to eliminate repeated records, records with poor geographic coordinates, and those out of the study area [52]. These strategies were used to avoid considering accessions that correspond to introductions outside the natural areas of distribution [53]. Finally, 480 records were selected (Table 1).
4.2. Climatic data
Monthly, quarterly, seasonal, and annual rasters of precipitation, temperature, solar radiation and relative humidity were used to determine potential distribution areas of D. remotiflora. These climatic data were obtained from the Agroclimatic Information System for México-Central America (SIAMEXCA) [21]. The raster images have a resolution of 30” of arc and correspond to the period 1961-2010. From the SIAMEXCA rasters, other additional variables were generated adding a total of 42 environmental variables (Table 2).
4.3. Environmental characterization of the occurrence sites
Based on the geographic coordinates of the D. remotiflora occurrence sites, an environmental characterization of these sites was carried out using the 42 variables mentioned in Table 2. For this, data extraction procedures were done, out from the raster images, using the ArcMap software version 10.8 [54]. With data extracted from the 42 variables, an environmental data matrix (EDM) was built in Microsoft Excel.
4.4. Selection of environmental variables
Prior to the execution of the statistical analysis, the Shapiro-Wilk test was applied to verify the data normality, not finding normality (P<0.05) for any of the data series within the 42 environmental variables included in the EDM.
Diverse studies have shown that multicollinearity is a problem that can cause high correlations among independent variables, a fact that can lead to unreliable and unstable estimations of the regression coefficients [55]. For determining the presence of multicollinearity, a Spearman correlation coefficient r > 0.9 was established as a threshold value [39]. In this way, the correlated variables with an r <0.9 coefficient were selected, and among the variables with collinearity, the one considered the most relevant for the presence of the species was selected. Data from the EDM were used to perform the correlation analyses; this statistical analysis was carried out with programs developed in the R software version 4.05 [56] and with normalized data. The result of these analyses reported 20 useful variables, later Preliminary analyses were carried out in MaxEnt individually and in conjunction with the Kuenm R-package in order to use the Jackknife test to select the most relevant variables in the distribution of D. remotiflora and thus carry out the final modeling of the ecological niche.
4.5. Characterization of the adaptive capacity of D. remotiflora.
Based on the geographic location of the D. remotiflora occurrence sites, the agroclimatic regions, soil units, and vegetation types where the species is currently distributed were characterized. For this purpose, the map of agroclimatic regions of México and Central America was used [13], as well as the soil units map and the vegetation types map for México [57]. This made possible the elaboration of a list of the edapho-climatic conditions to which D. remotiflora currently adapts and its preference for certain hábitats [58.
In addition, the ecological descriptors for D. remotiflora were determined by using the EDM information, but only taking into account the 20 environmental variables that were selected after the correlation-colinearity analysis. The ecological descriptors were established in terms of environmental ranges for D. remotiflora adaptation, and optimal environmental ranges for D. remotiflora presence, which corresponded to the highest frequency of occurrence sites.
4.6. Ecological niche modeling
Ecological niche modeling was done by using Maxent model, which uses the principle of maximum entropy with species presence and environmental data to create a correlative model that relates the ecological requirements of a species with the regional environmental availabilities to predict the relative habitat suitability, also it allows deriving specific descriptors for enriching the ecological characterization of the territories [59,60,61]. We used the Kuenm R-package [10,56,62] to automate and optimize the Ecological Niche Modeling (ENM) process. Kuenm's Kuenm_ceval function creates preliminary models [63] with occurrence data and environmental predictors, and also evaluates the efficiency of these models through their statistical significance with the cal_eval function, in addition, the relative quality of the model is evaluated by using the Akaike Information Criterion (AIC), corrected for small sample sizes (AICc) [10]. Kuenm also enables to assess diverse regularization muliplier (RM) factors, combinations of feature classes (FC) and different groups of environmental predictors, as well as to establish the allowable omission rate (OR). For each parameter setting, two models are created: one based on the complete set of occurrences, and the other based only on the training data. For this research, models were tested using a sequential order of the FC (L, LQ, H, LQH, LQHP, LQHPT) and RM values of 0.1 to 5 with 0.1 increases, a maximum omission rate of 5%, and run 50 k-fold replicates of each configuration; 500 iterations were used [64]. The D. remotiflora occurrence sites and the ASCII files for the 20 variables selected after correlation-collinearity analysis were used as inputs in the ENM process with Kuenm R-package.
5. Conclusions
D. remotiflora is an endemic species from México, whose tubers are collected in their natural habitat for human consumption. D. remotiflora is mostly adapted to semi-warm to warm environments and to semi-arid to sub-humid climates with a long dry season. However, the whole range of environmental adaptation of D. remotiflora also includes extremely humid, temperate, and very warm habitats, which indicates a high adaptive capacity of this species. In addition, D. remotiflora has a greater presence in regions where very thin, stony, poorly developed, and low-fertility soils predominate. All these characteristics make D. remotiflora a species with comparative advantages to develop in edapho-climatic environments that are adverse for most crop species. However, the greater presence of D. remotiflora occurrence sites indicates optimal environmental conditions, under which the development and productivity of the species would be expected to be maximum. In this way, populations of D. remotiflora proliferate in soils of coarse texture, in low to midlands, with annual precipitation of 700 to 1300 mm, and annual mean temperature of 19 to 27°C.
D. remotiflora is currently distributed in the western portion of México, along regions bordering the Mexican Pacific, which hints at its center of geographic origin. Niche modeling identified precipitation of the warmest quarter, precipitation of the driest month, mean minimum temperature of the coldest month, November-April mean solar radiation, Annual mean relative humidity, Annual moisture availability index, and May-October mean temperature as the variables with the greatest contribution to explain the presence of D. remotiflora. Furthermore, the Kuenm R package enabled the selection of a niche model that optimized the depiction of the potential distribution areas for this species. Thus, potential areas with high environmental suitability for D. remotiflora were located in the Mexican States where this species is already present, and potential areas with low to mid environmental suitability were identified in regions with the current presence of D. remotiflora as well as regions where it has not yet been reported, as in the Yucatan Peninsula, Northeast region, and the Gulf of México.
The cultivation perspectives of D. remotiflora are favorable considering its capacity to adapt to harsh environments, and that its nutritional and medicinal properties are valuable, however, the prolonged dormancy of its tubers is one of the intrinsic aspects of this plant that limits its offer as a cultivation option.
The information generated in this research will make it possible to learn more about the distribution, adaptation, and ecology of this species, which can also contribute to the design of conservation strategies for D. remotiflora and to the planning and selection of potential areas for its cultivation.
Currently, plant genetic resources worldwide are facing the pressure of overexploitation and environmental change, resulting in habitat fragmentation and biodiversity threats. To ensure the permanence of these natural resources, it is essential to maintain or increase the resilience of these systems against these pressures (Ten Broeke et al., 2019; Gülçin, 2021). Therefore, an efficient conservation management model must be adopted to address these changes and adequately preserve this plant genetic resources.
References
- Hussein, I.; Mengs, B.; Matiwos, T. Effect of Plant Growth Regulators on in Vitro Propagation of Yam Landraces (Dioscorea Species) Using Nodal Segments. Journal of Biology Agriculture and Healthcare 2018, 8, 13–23, https://core.ac.uk/ download/pdf/234662712.pdf. [Google Scholar]
- Rodríguez, R.R.; Téllez, V.O. Las Dioscoreas (Dioscoreaceae) del Estado de Morelos, México. Anales del Instituto de Biología. Serie Botánica 1992, 63, 67–99, https://www.redalyc.org/pdf/400/40063104.pdf. [Google Scholar]
- Castañeda, NJJ; Santacruz, RF; de Jesús SGJ; Parra, JR; De la Cruz, LL; Barba, R Shading and Container Effects on the Weight of the Dioscorea sparsiflora Tuber. Agronomy Journal 2017, 109, 33–38. [CrossRef]
- Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). Available online: https://enciclovida.mx/especies/192190-dioscorea-remotiflora: (accessed on 9 September 2022).
- Obidiegwu, JE; Akpabio, EM The geography of yam cultivation in southern Nigeria: Exploring its social meanings and cultural functions. Journal of Ethnic Foods 2017, 4, 28–35. [CrossRef]
- Akissoé, N; Hounhouigan, J; Mestres, C; Nago, M How blanching and drying affect the color and functional characteristics of yam (Dioscorea cayenensis-rotundata) flour. Food Chemistry 2003, 82, 257–264. [CrossRef]
- Montañez, SJL; Venegas, GJ; Bernardno, NA; González, CL; Yañez, FJ Chemical Characterization and Nutritional Evaluation of Mountain’s yam (Dioscorea remotiflora Kunth). Tubers 2014, 5, 153–160.
- Bernabe, AA; Santacruz, RF; Cruz, SF Effect of plant growth regulators on plant regeneration of Dioscorea remotiflora (Kunth) through nodal explants. Plant Growth Regulation 2012, 68, 293–301. [CrossRef]
- Obidiegwu, JE; Lyons, JB; Chilaka, CA The Dioscorea Genus (Yam) An Appraisal of Nutritional and Therapeutic Potentials. Foods 2020, 9, 1304. [CrossRef]
- Cobos, ME; Peterson, AT; Osorio, OL; Jiménez GD; An exhaustive analysis of heuristic methods for variable selection in ecological niche modeling and species distribution modeling. Ecological Informatics 2019, 53, 100983. [CrossRef]
- Chandra, A; Naithani, HB; Verma, PK; Saxena, J; & Prajapati, S. Plant diversity assessment of selected forest sites of Gaya district of Bihar, India. Journal of Applied and Natural Science 2021, 13, 424-432. [CrossRef]
- Ghorbani, A; Bahrami, B The influence environmental factors on the distribution of plant species in the southeast rangelands of Sabalan. Watershed Management Research Journal 2017, 30, 15–29. [CrossRef]
- Sánchez, GJDJ; Ruiz, CJA; García, GM; Ojeda, GR; Larios, LDLC; Holland, JB; García, RGE Ecogeography of teosinte. PLoS One 2018, 13, e0192676. /: https. [CrossRef]
- Aburto-Cansino, GN; Ruiz-Corral, JA; Sánchez González, JDJ; & González Eguiarte, DR Temperaturas cardinales de desarrollo del teocintle (Zea spp.). Revista mexicana de ciencias agrícolas 2018 9, 1269-1281. [CrossRef]
- Grativol, C; Silva HA; and Gomes, FP. Genetic and epigenetic regulation of stress responses in natural plant populations. Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms 2012, 1819, 176-185. [CrossRef]
- Ashapkin, VV; Kutueva, LI; Aleksandrushkina, NI; & Vanyushin, BF Epigenetic mechanisms of plant adaptation to biotic and abiotic stresses. International journal of molecular sciences 2020, 21, 7457. [CrossRef]
- Sosa, V; Nova, J Linajes de angiospermas endémicas en México: zonas de alto endemismo para la conservación. Acta Botánica Mexicana 2012, 100, 293–315, https://abm.ojs.inecol.mx/index.php/abm/article/view/38.
- Santacruz, RF; Casas, SJF; Pérez, PR; Rodríguez, GE; Torres, MMI; Castillo, HC; Iturbe SI; Conservación, manejo y aprovechamiento del camote de cerro (Dioscorea spp.) en el estado de Jalisco, México. Avances de la Investigación Científica en el CUCBA. XVI: 2005, 179-183. http://www.floradejalisco.cucba.udg.mx /sites/default/files/publicaciones1/avances/avances_2005/Agronomia/SantacruzRuvalcabaFernando/SantacruzRuvalcabaFernando.pdf.
- Miranda, AG; Soto, JLM; Ruiz, IG. Parcial caracterización de nuevos almidones obtenidos del tubérculo de camote del cerro (Dioscorea spp). Revista Iberoamericana de Tecnología Postcosecha 2008, 9, 81–88, https://www.redalyc.org /pdf/813/81311226011.pdf.
- González, VME El ñame (Dioscorea spp.) Características, usos y valor medicinal. Aspectos de importancia en el desarrollo de su cultivo. Cultivos Tropicales 2012, 33, 5–15, http://scielo.sld.cu/scielo.php?pid=S0258-59362012000400001& script=sci_ arttext&tlng=en.
- Ruiz, CJA; Medina, GG; García, RGE Sistema de información agroclimática para México-Centroamérica. Revista Mexicana de Ciencias Agrícolas 2018, 9, 1–10. [CrossRef]
- Barrera-Sánchez, CF; Ruiz-Corral, JA; Zarazúa-Villaseñor, P; Lépiz-Ildefonso, R; & González-Eguiarte, DR Cambio climático y distribución potencial de frijol lima en Mesoamérica y Aridoamérica. Revista Mexicana Ciencias Agrícolas 2020 11, 1361-1375. [CrossRef]
- Dormann, CF; McPherson, JM; Araujo, MB; Bivand, R; Bolliger, J; Carl, G Methods to account for spatial autocorrelation in the analysis of species distributional data: a review. Ecography 2007, 30, 609–628. [CrossRef]
- Peterson, AT; Nakazawa, Y Environmental data sets matter in ecological niche modelling: an example with Solenopsis invicta and Solenopsis richteri. Global Ecology and Biogeography 2008, 17, 135–144. [CrossRef]
- Feng, X. , Park, DS; Liang, Y; Pandey, R; Papeş, M Collinearity in ecological niche modeling: Confusions and challenges. Ecology and evolution 2019, 9, 10365–10376. [Google Scholar] [CrossRef]
- Xu, Y; Zhu, R; Gao, L; Huang, D; Fan, Y; Liu, C; Chen, J. Predicting the current and future distributions of Pennisetum alopecuroides (L.) in China under climate change based on the MaxEnt model. Plos one 2023, e0281254. [CrossRef]
- UNEP (United Nations Environmental Program). World Atlas of Desertification 2 ed.; Middleton N. and Thomas D. S. G: London, UK,1997. pp 1-182.
- Rodríguez, W Botánica, domesticación y fisiología del cultivo de ñame (Dioscorea alata). Agronomía Mesoamericana 2000, 11, 133–152, https://www.redalyc.org/pdf/437/ 43711221.pdf.
- Avalo, DMR; García, MB Producción y diversificación sostenible del cultivo de ñame (Dioscorea spp.) en condiciones de sequía agrícola en el municipio de Jiguaní. Agrisost 2021, 27, 1–7, https://revistas.reduc.edu.cu/index.php/agrisost/article/view/ e10333-1.
- Velázquez, HJM; Durán, PN; Ruiz, CJA; González, EDR; Santacruz, RF; Gallegos, RA. Distribución geográfica y usos de especies del género Dioscorea. E-cucba 2022, 19, 141–150. [CrossRef]
- Mueller, L; Schindler, U; Mirschel, W; Shepherd, TG; Ball, BC; Helming, K; Rogasik, J; Eulenstein, F; Wiggering, H Assessing the productivity function of soils. A review. Sustainable Agriculture, 2010; 30, 601–614. [CrossRef]
- INEGI Suelos. Instituto Nacional de Estadística y Geografía. D. F México 2007. pp 36 https://apps1. semarnat.gob.mx:8443/dgeia/informe_12/pdf/Cap3_suelos.pdf.
- López, FAJ; Hernández, CD Cambio climático y agricultura: una revisión de la literatura con énfasis en América Latina. El trimestre económico. 2016, 83, 459–496. [CrossRef]
- Ruiz, CJA.; Medina, GG. González, AIJ. Flores, LHE. Ramírez, OG. Ortiz, TC. Byerly, MKF. Requerimientos agroecológicos de cultivos, 3a ed.; INIFAP-Prometeo Editores: Jalisco, México, 2013; pp. 564. https://www.researchgate.net/publication/343047223_REQUERIMIENTOS_AGROECOLOGICOS_DE_CULTIVOS_2da_Edicion.
- García, E. Modificaciones al Sistema de Clasificación Climática de Köppen (Para adaptarlo a las condiciones de la República Mexicana), 5ª Edición.; Instituto de Geografía-UNAM: Ciudad de México, México, 2004; pp. 90. http://www.publicaciones.igg.unam.mx/index.php/ig/catalog/book/83.
- Thompson, AK; Oduro, I. Yams: Botany, Production and Uses, 1aed.; CABI: Boston, USA, 2021; pp. 296. [CrossRef]
- Patricio, VEG; Rodrigo, ARJ; Isaías, MAR; Cristina, LVI; del Pilar, PMN; Santiago, EVJ; Inhibición de la brotación del tubérculo de papa: una revisión de los métodos empleados Inhibition of potato tuber sprouting: a review of the methods employed. Journal of the Selva Andina Biosphere 2018, 6, 55–64, http://scielo.org.bo/pdf/jsab/v6n2/v6n2_a04.pdf.
- Coursey, D. Yams: Dioscorea spp, 2nd Edition.; In: Wiley: London, UK, 1967; pp. 70–74. https://agris.fao.org/ agrissearch/search.do?recordID=US201303063121.
- Yang, J; Huang, Y; Jiang, X; Chen, H; Liu, M; Wang, R Potential geographical distribution of the endangered plant Isoetes under human activities using MaxEnt and GARP. Global Ecology and Conservation 2022, e02186. [CrossRef]
- Abdelaal, M; Fois, M; Fenu, G., Bacchetta, G Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. in Egypt. Ecological informatics 2019, 50, 68–75. [CrossRef]
- Muscarella, R; Galante, PJ; Soley, GM; Boria, RA; Kass, JM; Uriarte, M; Anderson, RP ENMeval: an R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods in Ecology and Evolution 2014, 5, 1198–1205. [CrossRef]
- Fois, MA; Cuena, LG; Fenu, G Bacchetta Using species distribution models at local scale to guide the search of poorly known species: review, methodological issues and future directions. Ecol. Model. 2018, 124–132. [CrossRef]
- Shen, L; Xu, J; Luo, L; Hu, H; Meng, X; Li, X.; Chen, S Predicting the potential global distribution of diosgenin-contained Dioscorea species. Chinese medicine 2018, 13, 58. [CrossRef] [PubMed]
- Yin, DS; Chen, F; Chen, Z; Guan WF Morpho- anatomical and physiological responses of two Dendranthema species to waterlogging. Environmental and Experimental Botany. 2010, 68, 122–130, https://doi.org/10.1016/j.envexpbot.2009.11.008. [CrossRef]
- Jiménez, JDLC; Moreno, LP; Magnitskiy, S Respuesta de las plantas a estrés por inundación. Una revisión. Revista Colombiana de Ciencias Hortícolas 2012, 6, 96–109, https://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S2011-21732012000 100010.
- Jiménez, JDLC; Moreno, LP; Magnitskiy, S Respuesta de las plantas a estrés por inundación. Una revisión. Revista Colombiana de Ciencias Hortícolas 2012, 6, 96–109, https://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S2011-21732012000 100010.
- Medina, GC; Giménez, deJ; Velázquez, MA Las comunidades vegetales del bosque de coníferas altimontano en el macizo del Tancítaro, Michoacán, México. Acta botánica mexicana 2020, p.127. [CrossRef]
- Schaal, BA; Hayworth, DA; Olsen, KM; Rauscher, JT; Smith, WA Phylogeographic studies in plants: problems and perspectives. Molecular Ecology 1998, 465–474. https://www.researchgate.net/profile/Kenneth-Olsen-4/publication/227339 790_Phylogeographic_studies_in_plants_Problems_and_prospects/links/5dfd016da6fdcc2837318eee/Phylogeographic-studies-in-plants-Problems-and-prospects.pdf. /: https.
- Dillon, MO; Tu, T; Xie, L; Quipuscoa, SV; Wen, J Biogeographic diversification in Nolana (Solanaceae), a ubiquitous member of the Atacama and Peruvian Deserts along the western coast of South America. Journal of Systematics and Evolution. 2009, 47, 457–476. [CrossRef]
- Schmidt, JR; Sytsma, KJ Phylogenetics of Puya (Bromeliaceae): placement, major lineages, and evolution of Chilean species. American Journal of Botany 2010, 97, 337–356. [CrossRef]
- Gao, X; Liu, J; Huang, Z The impact of climate change on the distribution of rare and endangered tree Firmiana kwangsiensis using the Maxent modeling. Ecology and Evolution 2022, 12, e9165. [CrossRef]
- Zhao, Y; Deng, X; Xiang, W; Chen, L; Ouyang, S Predicting potential suitable habitats of Chinese fir under current and future climatic scenarios based on Maxent model. Ecological Informatics. 2021, 64, 101393. [CrossRef]
- Ramírez, OG; Peralta, IE; Rodríguez, GE; Sahagún, CJ; Chávez, SJL; Medina, HTC; Rodríguez, PJE Edaphoclimatic Descriptors of Wild Tomato Species (Solanum Sect. Lycopersicon) and Closely Related Species (Solanum Sect. Juglandifolia and Sect. Lycopersicoides) in South America. Frontiers in genetics 2021, 12. [CrossRef]
- vironmental Systems Research Institute (ESRI) ArcGIS Desktop: Release 10. Environmental Systems Research Institute. Redlands, CA, USA. 2010, 1 p5. https://earthobservations.org/about_geo.shtml.
- O’brien, RM Caution Regarding Rules of Thumb for Variance Inflation Factors. Qual Quant 2007, 41, 673–690. [CrossRef]
- Core, T.R. R.A., Language and environment for statistical computing. R Foundation for Statistical Computing, Vienna.Austria. (Versión 4.2.1). 2021. https://www.R-project.org/.
- INEGI., 2017. Conjunto de datos vectoriales de edafología serie 4, escala 1:250,000. Aguascalientes: Instituto Nacional de Estadística y Geografía. https://www. inegi.org.mx/temas/edafologia/.
- Thouverai, E; Marcantonio, M; Lenoir, J; Galfré, M; Marchetto, E; Bacaro, G; Rocchini, D Integrals of life: Tracking ecosystem spatial heterogeneity from space through the area under the curve of the parametric Rao’s Q index. Ecological Complexity 2023, 52, 101029. [CrossRef]
- Phillips, SJ; Anderson, RP; Schapire, RE Maximum entropy modeling of species geographic distributions. Ecological Modelling 2006, 190, 231–259. [CrossRef]
- Elith, J. , Phillips, SJ; Hastie, T; Dudík, M; Chee, YE; Yates, CJA Statistical explanation of MaxEnt for ecologists. Diversity and Distributions 2011, 171, 43–57. [Google Scholar] [CrossRef]
- Warren, DL; Seifert, SN Ecological niche modeling in Maxent: the importance of model complexity and the performance of model selection criteria. Ecological applications 2011, 21, 335–342. [CrossRef]
- Mazzolari, AC; Millán, EN; Bringa, EM; Vázquez, DP Modeling habitat suitability and spread dynamics of two invasive rose species in protected areas of Mendoza, Argentina. Ecological Complexity 2020, 44, 100868. [CrossRef]
- Gull E; Fareen, A; Mahmood, T; Bodlah, I; Rashid, A; Khalid, A; Mahmood, S. Modeling potential distribution of newly recorded ant, Brachyponera nigrita using Maxent under climate change in Pothwar region, Pakistan. PLoS One 2022, 17, e0262451. [CrossRef]
- Cao, Y; DeWalt, RE; Robinson, JL; Tweddale, T; Hinz, L; Pessino, M Using Maxent to model the historic distributions of stonefly species in Illinois streams: The effects of regularization and threshold selections. Ecological Modelling 2013, 259, 30–39. [CrossRef]
Figure 1.
The current distribution of D. remotiflora in the agroclimatic regions of México.
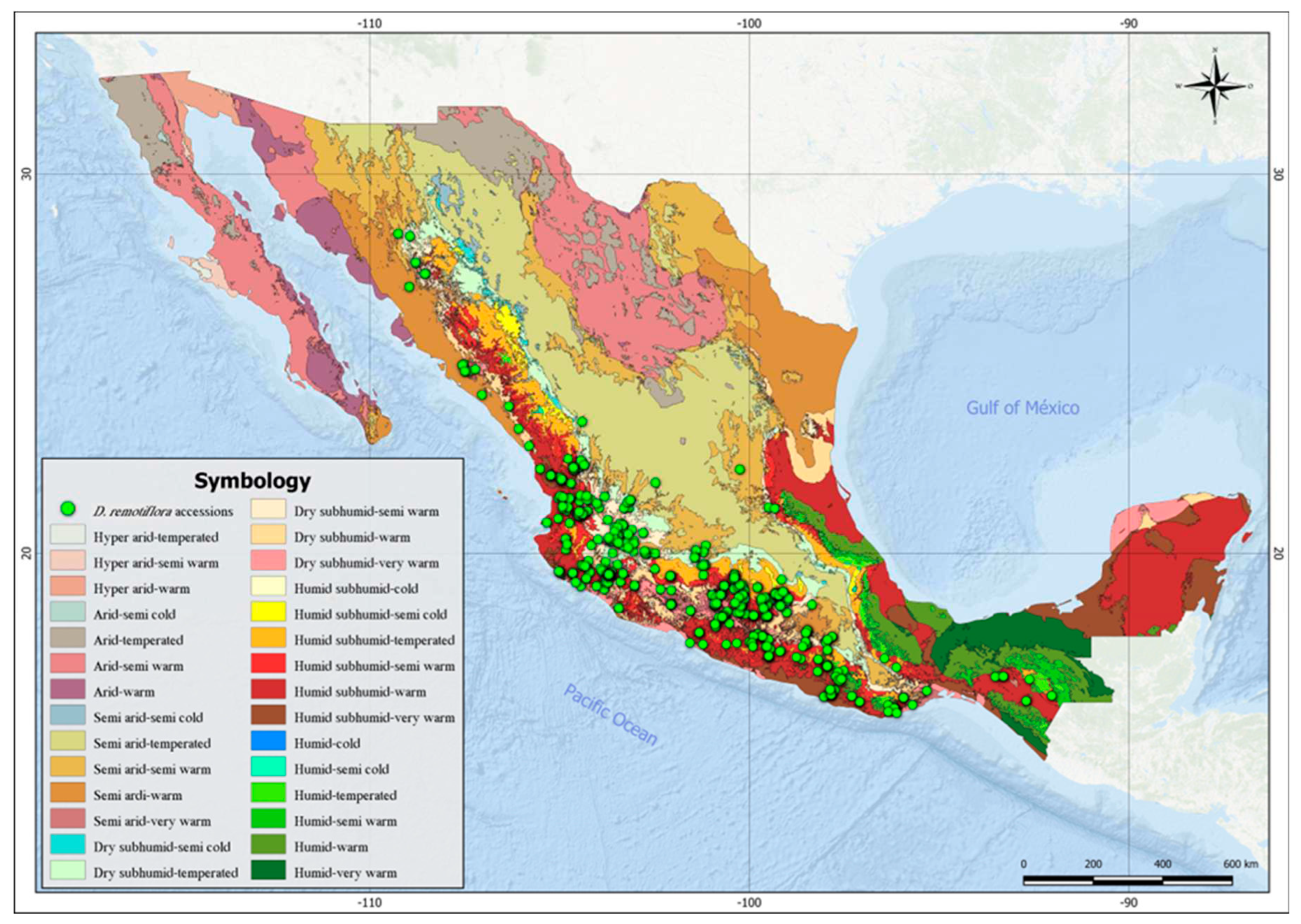
Figure 3.
Results of jackknife test of the relative importance of predictor environmental variables in MaxEnt model for D. remotiflora in México.
Figure 3.
Results of jackknife test of the relative importance of predictor environmental variables in MaxEnt model for D. remotiflora in México.
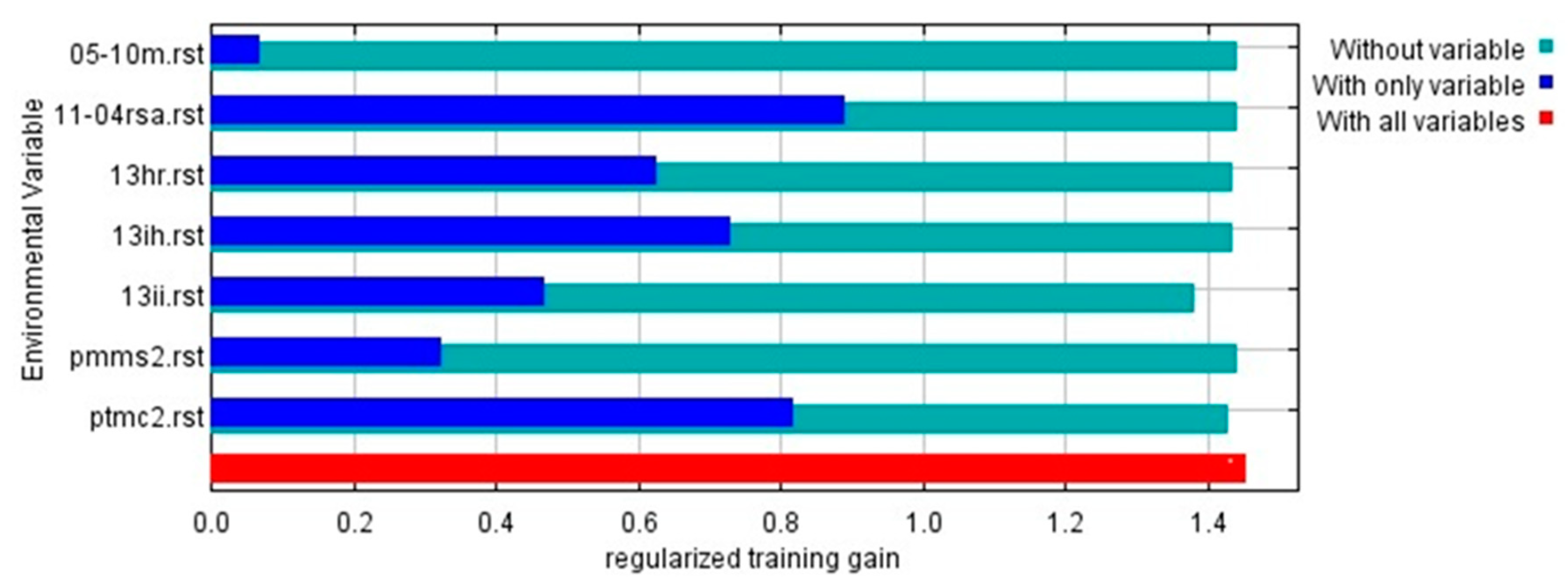
Figure 4.
Models obtained and evaluated by Kuenm R-Package for D. remotiflora.
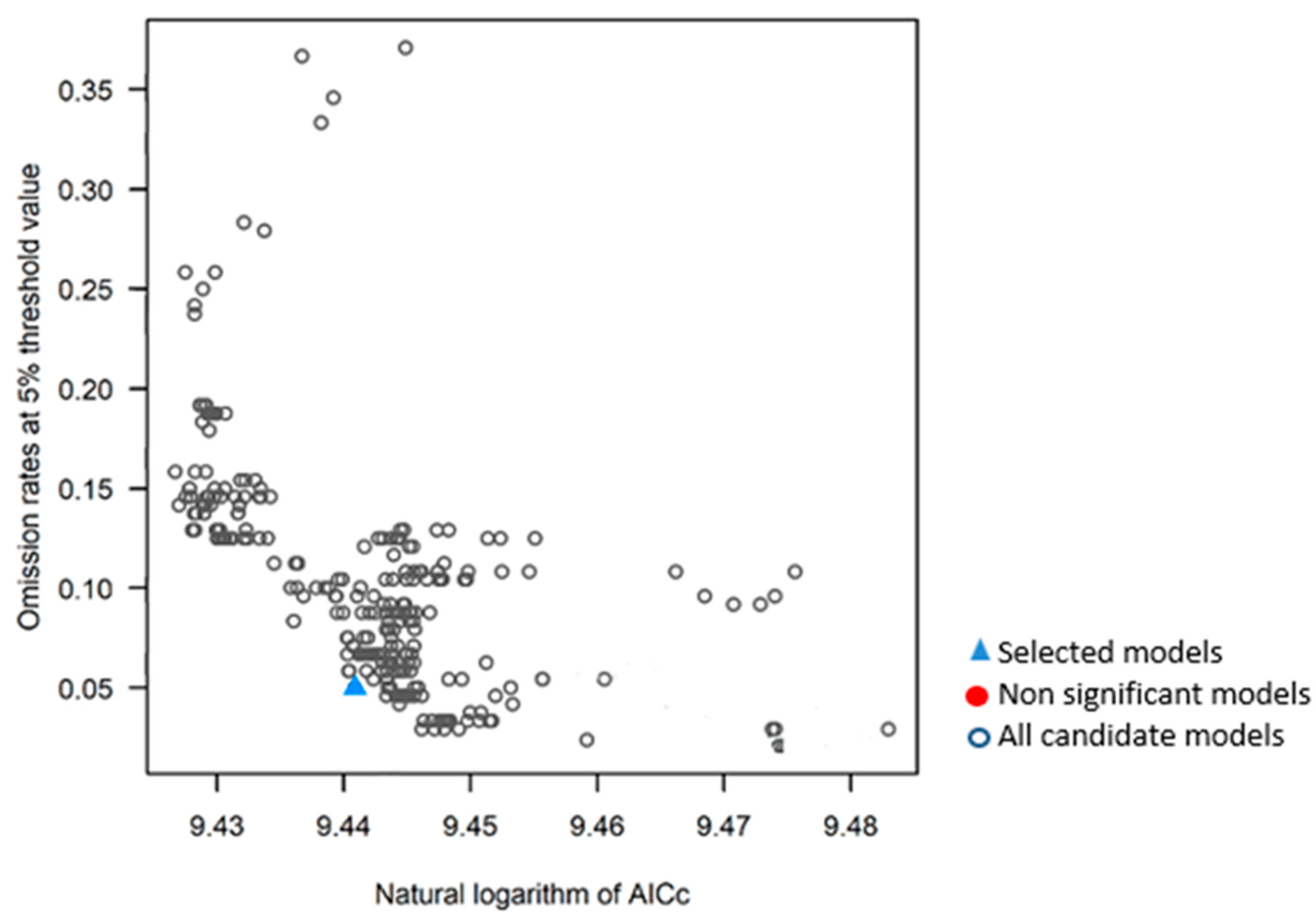
Figure 5.
Areas with environmental suitability for D. remotiflora in México.
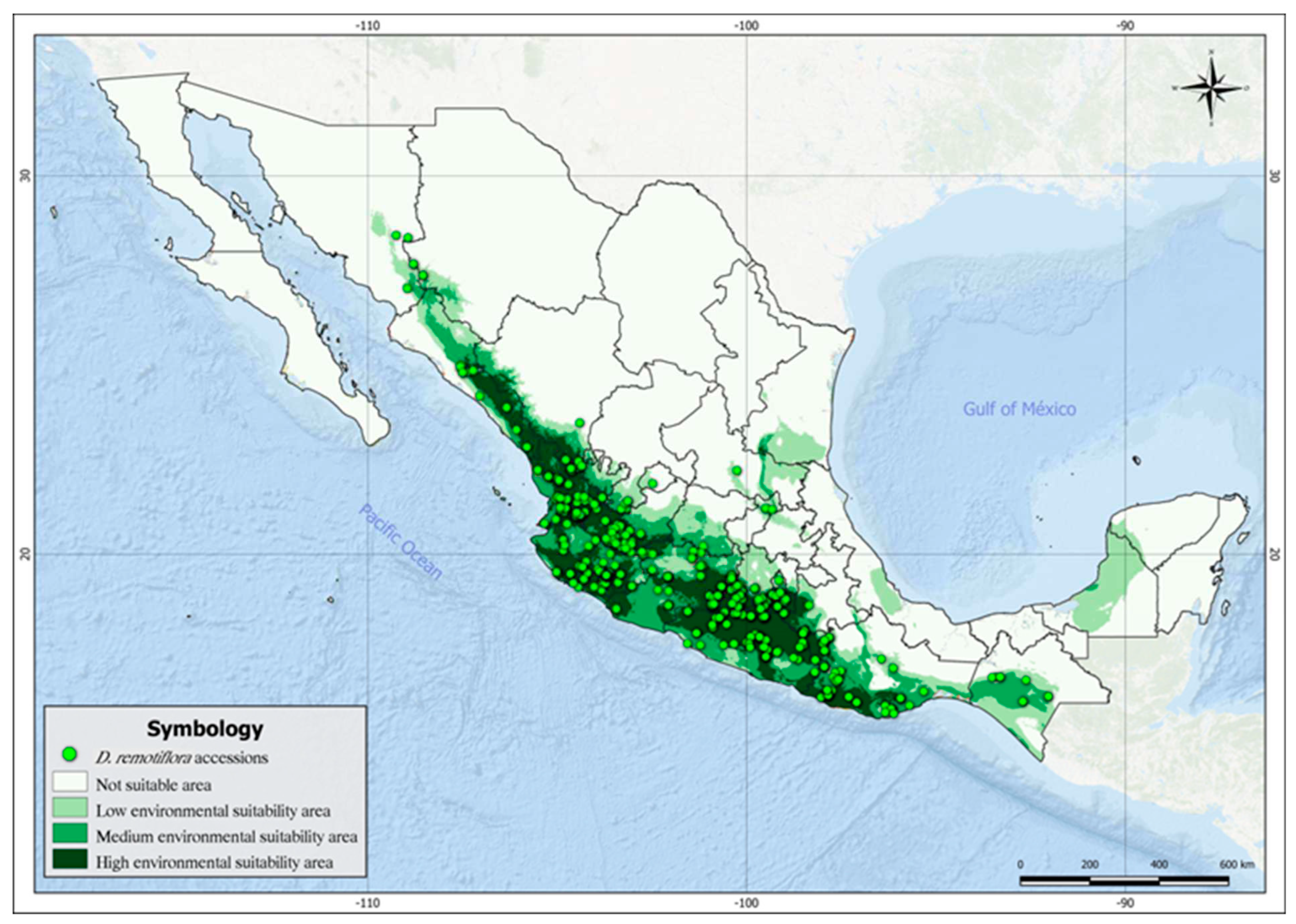

Table 3.
Annual mean temperature and annual moisture availability index intervals for 17 agroclimatic regions with the presence of D. remotiflora in México.
Table 3.
Annual mean temperature and annual moisture availability index intervals for 17 agroclimatic regions with the presence of D. remotiflora in México.
| Agroclimatic Region | Annual moisture availability index | Annual mean temperature (°C) | Total Accessions |
|---|---|---|---|
| Semiarid very warm | 0.2 - 0.5 | >26 | 17 |
| Semiarid warm | 0.2 - 0.5 | 22 - 26 | 22 |
| Semiarid semi-warm | 0.2 - 0.5 | 18 a 22 | 14 |
| Semiarid temperate | 0.2 - 0.5 | 12 a 18 | 3 |
| Dry-subhumid very warm | 0.5 - 0.65 | > 26 | 20 |
| Dry-subhumid warm | 0.5 - 0.65 | 22 – 26 | 66 |
| Dry-subhumid semi-warm | 0.5 - 0.65 | 18 – 22 | 88 |
| Dry-subhumid temperate | 0.5 - 0.65 | 12 – 18 | 7 |
| Humid-subhumid very warm | 0.5 - 0.65 | >26 | 19 |
| Humid-subhumid warm | 0.65 - 1.0 | 22 - 26 | 67 |
| Humid-subhumid semi-warm | 0.65 - 1.0 | 18 – 22 | 98 |
| Humid-subhumid temperate | 0.65 - 1.0 | 12 - 18 | 11 |
| Humid very warm | >1.0 | >26 | 2 |
| Humid warm | >1.0 | 22 - 26 | 26 |
| Humid semi-warm | >1.0 | 18 - 22 | 12 |
| Humid temperate | >1.0 | 12 - 18 | 6 |
| Humid semi-cold | >1.0 | 5 - 12 | 3 |
Table 4.
Soil units and soil texture classes with the presence of D. remotiflora.
| FAO Soil Unit | Soil Texture | Total Accessions |
|---|---|---|
| Lithosol | Coarse | 108 |
| Regosol calcaric | Coarse | 57 |
| Regosol eutric | Coarse | 209 |
| Faozem haplic | Coarse | 34 |
| Vertisol cromic | Fine | 39 |
| Solonchak ortic | Fine | 22 |
| Fluvisol eutric | Medium | 10 |
| Fluvisol calcaric | Coarse | 1 |
Table 5.
Ecological descriptors for D. remotiflora.
| Environmental variables | Min | Max | Optimum |
|---|---|---|---|
| 1.Precipitation of the warmest quarter (mm) | 240 | 1,204 | 400-884 |
| 2.Precipitation of the driest month (mm) | 1 | 73 | 1-7 |
| 3.Annual mean precipitation (mm) | 444 | 2,886 | 700-1299 |
| 4.May-October mean precipitation (mm) | 344 | 1,943 | 700-1199 |
| 5.November-April mean precipitation | 23 | 863 | 30-100 |
| 6.Annual moisture availability index | 0.27 | 2.32 | 0.40-0.99 |
| 7.November-April availability index | 0.026 | 1.83 | 0.030-1,300 |
| 8.May-October availability index | 0.005 | 1.47 | 0.009-1.4 |
| 9.Maximum maximorum temperature (°C) | 24.61 | 41.17 | 29-37 |
| 10.Minimum minimorum temperature (°C) | 1.7 | 18.2 | 5-15 |
| 11.Annual mean temperature (°C) | 14.66 | 28.51 | 19-27 |
| 12.May-October mean temperature | 9.13 | 29.88 | 19-26 |
| 13.November-April mean temperature | 7.95 | 27.83 | 19-26 |
| 14.Annual thermal oscillation (°C) | 10.42 | 19.54 | 13.16 |
| 15.Annual temperature range (°C) | 1.54 | 14.52 | 3-7 |
| 16.Soil texture | Arenoso | Fina | Media |
| 17.May-October mean photoperiod (h) | 12.5 | 12.9 | 12.6-12.9 |
| 18.November-April mean photoperiod (h) | 10.97 | 11.47 | 11.10-11.39 |
| 19.Growing season | 120-190 | ||
| 20.Altitude (mm) | 6 | 4,295 | 200-1800 |
Min= Minimum value; Max= Maximum value; Optimum= Optimal range.
Table 6.
Contribution of seven environmental variables to depict the presence and distribution of D. remotiflora.
Table 6.
Contribution of seven environmental variables to depict the presence and distribution of D. remotiflora.
| Environmental variables | Contribution (%) | Permutation importance (%) |
|---|---|---|
| Precipitation of the warmest quarter (mm) | 42.4 | 49 |
| Pprecipitation of the driest month (mm) | 17.5 | 2.8 |
| Minimum temperature of the coldest month (°C) | 15 | 31 |
| November-April mean solar radiation (w/m2) | 10 | 0.8 |
| Annual mean relative humidity (%) | 8.5 | 3.6 |
| Annual moisture availability index | 5.7 | 7.4 |
| May-October mean temperature (°C) | 0.9 | 5.3 |
Table 1.
Occurrence data sources for D. remotiflora.
| Institution/source | Institution/Department | Accessions |
|---|---|---|
| Universidad Nacional Autónoma de México (UNAM). | Instituto de Biología | 169 |
| Instituto de Ecología (INECOL). | Xalapa Veracruz | 30 |
| Universidad Autónoma de Querétaro (UAQ). | Facultad de Ciencias Naturales | 3 |
| Instituto Nacional de Estadística y Geografía (INEGI). | Departamento de Botánica | 2 |
| Universidad Autónoma de Aguascalientes (UAA). | Centro de Ciencias Básicas | 2 |
| Universidad Autónoma de Veracruz (UPAV) (CIB). | Instituto de Investigaciones Biológicas | 1 |
| Universidad Autónoma de San Luis Potosí (UASLP). | Instituto de Investigación de Zonas Desérticas | 3 |
| Colegio de la Frontera Sur (ECO SUR). | Herbario San Cristóbal | 3 |
| Universidad de Guadalajara (CUCBA, CUC SUR). | Herbario IBUG, Herbario ZEA | 6 |
| Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Michoacán. | Herbario Facultad de Biología Universidad Michoacana de San Nicolás de Hidalgo | 6 |
| Artículos científicos/Inventarios florísticos de los estados de Oaxaca, Chiapas, Veracruz, Tabasco, Guerrero, Puebla, Jstor Plant Science. | 20 | |
| Universidad Autónoma de Nuevo León (UNL). | Facultad de Ciencias Biológicas | 1 |
| La Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). | Herbario digital de CONABIO | 3 |
| Trópicos.org. | 3 | |
| Red de Herbarios del Noroeste de México. | 13 | |
| GBIF | 215 | |
| Total | 480 |
Table 2.
Environmental variables considered in this research.
| Variable | Description | Temporal scale |
|---|---|---|
| BIO01 | (Annual mean temperature) | Annual |
| BIO02 | Mean diurnal range | Variation |
| BIO03 | Isothermality | Variation |
| BIO04 | Temperature seasonality | Variation |
| BIO05 | (Maximum temperature of the warmest month) | Month |
| BIO06 | (Minimum temperature of the coldest month) | Month |
| BIO07 | Temperature annual range | Annual |
| BIO08 | (Mean temperature of the wettest quarter) | Quarter |
| BIO09 | (Mean temperature of the driest quarter) | Quarter |
| BIO10 | (Mean temperature of the warmest quarter) | Quarter |
| BIO11 | (Mean temperature of the coldest quarter) | Quarter |
| BIO12 | Annual precipitation | Annual |
| BIO13 | Precipitation of the wettest month | Month |
| BIO14 | Precipitation of the driest month | Month |
| BIO15 | Precipitation seasonality | Variation |
| BIO16 | Precipitation of the wettest quarter | Quarter |
| BIO17 | Precipitation of the driest quarter | Quarter |
| BIO18 | Precipitation of the warmest quarter | Quarter |
| BIO19 | Precipitation of the coldest quarter | Quarter |
| N-AMT | November-April mean temperature | Seasonal |
| M-OMT | May-October mean temperature | Seasonal |
| M-OXT | Maximum temperature May-October | Seasonal |
| N-AXT | November-April maximum temperature | Seasonal |
| AXT | Annual maximum temperature | Annual |
| M-OIT | May-October minimum temperature | Seasonal |
| N-AIT | November-April minimum temperature | Seasonal |
| AIT | Annual minimum temperature | Annual |
| ATO | Annual thermal oscillation | Annual |
| M-OP | May-October precipitation | Seasonal |
| N-AP | November-April Precipitation | Seasonal |
| M-OPH | May-October photoperiod | Seasonal |
| N-APH | November-April photoperiod | Seasonal |
| AMI | Annual moisture index | Annual |
| M-OMI | May-October mean moisture index | Seasonal |
| N-AMI | November-April mean moisture index | Seasonal |
| ASR | Annual mean solar radiation | Annual |
| M-OSR | May-October mean solar radiation | Seasonal |
| N-ASR | November-April solar radiation | Seasonal |
| ARH | Annual relative humidity | Annual |
| M-ORH | May-October relative humidity | Seasonal |
| N-ARH | November-April relative humidity | Seasonal |
| GSL | Growing season length | Seasonal |
BIO = Bioclimatic variable.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



