
Submitted:
04 September 2023
Posted:
04 September 2023
You are already at the latest version
Abstract
Abstract
Forensic institutions throughout the country house patients with severe psychiatric illness and history of criminal violations. Improved medical care, hygiene, and nutrition led to unmatched longevity in this population which previously lived on average 15 to 20 years shorter than the public at large. On the other hand, longevity has contributed to increased prevalence of age-related diseases, including neurodegenerative disorders, which complicate clinical management.
Forensic institutions, originally intended for the treatment of younger individuals, are ill-equipped for the growing number of older offenders. Moreover, as antipsychotic drugs became available in 1950s and 1960s, we are observing the first generation of forensic detainees who had aged on dopamine blockers. Although the consequences of long-term treatment with these agents are unclear, schizophrenia-associated gray matter loss, may contribute to the development of early dementia.
Taken together, increased lifespan and subsequent cognitive deficit observed in long-term psychiatric institutions brought forth questions and dilemmas unencountered by the previous generations of clinicians, such as:
1. Does the presence of neurocognitive dysfunction justify antipsychotic dose reduction or discontinuation despite a lifelong history of schizophrenia and violent behavior?
2. Should neurolipidomic interventions become the standard of care in elderly individuals with lifelong schizophrenia and dementia?
3. Can patients with schizophrenia and dementia meet the Dusky standard and stand trial?
4. Should neurocognitive disorders in elderly with lifelong schizophrenia be treated differently than age-related neurodegeneration?
In this article, we describe strategies for potentially slowing the development of neurocognitive disorders in forensic patients with chronic mental illness by adopting a three-prong strategy:
1. Approaching lifelong psychosis from microbial and immunological perspective.
2. Identify modifiable risk factors for age-related diseases in forensic institutions.
3. Utilizing novel receptor-independent treatments for chronic psychosis.
Keywords:
Neurodegenerative disorders
; schizophremia
; PDK1 inhibitors
; Lipid replacement therapy
; Aryl hydrocarbon receptor antagonists
Long term psychiatric institutions remain in existence because the outcome of schizophrenia and schizophrenia-like disorders has been unsatisfactory. At present, criminal behavior cannot, without speculation, be accredited to specific brain areas or pathologies, but this may change in the future. Improved care has prolonged the life of patients with chronic psychosis and history of criminal behavior residing in state hospitals. Longevity in chronic psychiatric patients predisposes to the development of dementia, engendering a new pathology: neurodegeneration superimposed on chronic psychosis. Receptor-independent antipsychotic strategies revolve around gut barrier rehabilitation, averting microbial translocation, and restoring the homeostasis of neuronal membrane.
Introduction
The population worldwide is aging at a rapid pace and so are forensic detainees with severe psychiatric illnesses, including schizophrenia (SCZ) and schizophrenia-like disorders (SLDs). In forensic institutions, there are two subpopulations of elderly patients with neurocognitive disorders: career criminals with lifelong mental illness and elderly first offenders without previous psychiatric history. The latter category includes patients with frontotemporal dementia behavioral variant (bvFTD), a condition with clinical manifestations similar to those of schizophrenia (SCZ). As the incidence of bvFTD has increased dramatically in forensic population over the past two decades, screening all first offenders after the age of 50 could identify this disorder early to ensure adequate adjudication and placement, while and at the same time, decongesting the crowded State hospitals.
At the cellular level, bvFTD has been associated with the selective loss of von Economo neurons (VENs), large cells, located in the anterior cingulate cortex (ACC), insular cortex (IC), and frontopolar cortex (FPC). As VENs belong to the human neuro-moral network which plays a key role in amiability and empathy, loss of these cells is believed to engender a type of acquired sociopathy (1)(2)(3). Interestingly, dysfunctional ACC, IC and FPC have been associated not only with SCZ but also with impaired disease awareness or anosognosia, a common SCZ characteristic which increases the odds of criminal violations (4)(5)(6)(7). Indeed, several recent studies have suggested that anosognosia may be a better predictor of criminal behavior than focal brain lesions or clinical diagnoses (8)(9)(10).
Improved medical care, sanitation, and nutrition have contributed to increased longevity in both the population at large and institutionalized individuals with criminal history and chronic SCZ or SLDs (11)(12)(13). On the other hand, increased lifespan has engendered a novel pathology, late-life dementia in people living with SCZ (PLWS), a complex entity difficult to manage in forensic institutions (14)(15). The complexity is further compounded by the long-term treatment with dopamine (DA) blocking drugs, agents associated with serious adverse effects in patients with dementia (18)(19). This begets a clinical dilemma, as treatment of chronic psychosis can harm patients with dementia, while nontreatment may contribute to the reemergence of violent behaviors. For this reason, new therapeutic strategies are urgently needed to address this conundrum, strategies which may involve neurolipidomic approaches, restoration of gut barrier homeostasis and other receptor-independent antipsychotic treatments (RIATs)(20)(21).
Exposure to typical and atypical antipsychotic drugs and chronic DA depletion was demonstrated to disrupt synaptic plasticity, contributing to impaired learning of new information (22)(23)(24). When superimposed on late-life dementia, dysfunctional neuroplasticity leads to more pronounced cognitive deficits, consistent with a distinct pathological entity (25)(26). Indeed, under physiological circumstances, healthy aging was associated with increased synthesis of striatal DA, probably to compensate for the age-related downregulation of DA receptors (27)(28)(29). Loss of synaptic plasticity, a modifiable risk factor, may be reversed via neurolipidomic therapies and/or DA reuptake inhibitors (DRIs), suggested by some researchers and clinicians (30).
Antipsychotic drugs exert receptor-dependent and receptor-independent pharmacological actions, the former have been well-defined and frequently referred to, while the latter are rarely mentioned in the literature. Receptor-independent properties of antipsychotic drugs, include antimicrobial, antiproliferative, pro-autophagy, anti-endocytic as well as lysosomotropism and peroxidation of plasma membrane lipids (31)(32)(33)(34)(35). Indeed, several first and second-generation antipsychotic drugs were demonstrated to disrupt neuronal and non-neuronal cells by oxidation of membrane lipid bilayer. This in return, distorts the cell surface, altering receptor signaling as well as the transcellular transport. At the level of intestinal epithelia, antipsychotic drugs-mediated peroxidation of lipids in plasma membranes, increases gut permeability, facilitating microbial translocation into host tissues (34)(36)(37)(38)(39)(40)(Figure 1).
Another receptor-independent property of antipsychotic drugs is interference with iron metabolism (41)(42)(43). As DA functions as a “de facto” iron chelator, it averts lipid peroxidation by sequestrating iron; conversely antipsychotic drugs upregulate iron, predisposing to neuronal death by ferroptosis (44)(45). Indeed, induced ferroptosis, a new form of chemotherapy, has been utilized for eliminating neuroblastoma, a malignancy sensitive to iron overload (46)(47). Along this line, antipsychotic drugs-induced gray matter loss, reported by several neuroimaging studies, may be taking place by ferroptosis (48)(49)(50).
In this article, we discuss RIATs, including lipid replacement therapy (LRT), recombinant human IL-22, and aryl hydrocarbon receptor (AhR) antagonists. We also take a closer look at the dysfunctional interoceptive awareness and its role in bvFTD and recommend screening older first offenders for this disorder to identify it early and ensure proper sentencing and placement.
The neuroscience of criminal behavior
For the past two centuries, the relationship between neuroscience and criminal law has been an uneasy one. Due to the complexity of criminal behavior and overlapping brain areas driving behavioral responses, a crime cannot, without speculation, be directly corelated to a specific central nervous system (CNS) region or pathology, a situation which may change as the molecular underpinnings of cellular cognition are better defined (51). In addition, the disappointing results of schizophrenia (SCZ) outcome studies and the continued need for public institution to treat chronic mentally ill patients comprise proof of concept that a direct link between brain pathology and specific behaviors remains elusive (52)(53)(54). For example, the well-known actor, Robin Williams who died by suicide, had Lewy body disease (LBD), however, linking this pathology to suicide would not be accepted in a court of law as most people with this disease do not take their lives (55). Another example, Charles Whitman, an individual who committed mass murder in 1966, had a brain tumor involving amygdala, however, the extent to which this pathology contributed to his crime, if any, cannot be ascertained as individuals with similar conditions may not engage in violent acts (56).
Interoceptive awareness is the ability to perceive internal state of the body, such as the heart rate, respiration, position and belonging of limbs, in other words, human self-awareness or insight are dependent on this system. Interoceptive awareness can be impaired in various pathologies, including strokes of the right hemisphere. For example, stroke-related neurological deficits can give patients the false impression that the left limbs do not belong to them, linking left-sided neglect to the faulty right hemisphere (57)(58).
Like stroke, SCZ, bvFTD, and chronic traumatic encephalopathy (CTE) can be associated with anosognosia and neglect of body functions as specific brain areas may misprocess information, giving patients false impression or illusions which lead to erroneous beliefs known as delusions, suggesting that anosognosia begets psychosis, rather than the other way around (59)(60)(61). Since VENs have been associated with interoceptive awareness and a larger number of VENs reside in the right hemisphere compared to the left, it is reasonable to suggest that anosognosia is a function of the right IC, ACC, and FPC. As poor illness insight is found in up to 98% patients with SCZ, this disorder likely originates in the right hemisphere and probably disinhibits the speech areas in the left brain, accounting for auditory hallucinations (62). In addition, SCZ was associated with decreased number of VENs, cells of growing significance in this disorder, consistent with the idea that anosognosia leads to psychosis (63)(64)(65). Moreover, as anosognosia is a cognitive symptom, SCZ may be primarily a cognitive disorder in which psychosis is a secondary phenomenon, an observation endorsed by Emil Kraepelin (66).
Anosognosia became the focus of research during the COVID-19 pandemic which, like HIV, could lead to poor insight into cognitive impairment (67)(68)(69)(70). Discovered in 1925 by Constantin von Economo, VENs are unique, elongated, spindle shaped cells, documented in advanced mammals, which are present in higher number in humans compared to any other animal. In addition, VENs emerge after birth and their number increases gradually until the age of four, coinciding with the preoperational stage of development when children become capable of symbolical thinking, further linking VENs to insight (69(71)(72 )(73) Interestingly, the number of VENs does not decrease during healthy aging, showing that loss of these cells may contribute to dementia (73). Furthermore, VENs comprise the brain salience network (SN), a cortical neuronal hub associated with interoceptive awareness and insight which under pathological conditions, may lead to SCZ, bvFTD, and violent behavior (69)(74).
Schizophrenia as a segmental progeria
Segmental progeria, as SCZ is often called by researchers, is a syndrome of accelerated cellular aging marked by shortened telomeres which drive both neurodegeneration and all-cause mortality (75)(76). As a result, PLWS have an average lifespan of 15-20 years shorter compared to their healthy counterparts. However, due to improved care over the past three decades, the lifespan of this population has increased considerably, predisposing to neurodegenerative pathology (77)(78).
Cellular senescence is an anticancer program of proliferation arrest, resistance to apoptosis, and active metabolism marked by a proinflammatory secretome, named senescence-associated secretory phenotype (SASP) which can spread senescence to the neighboring healthy cells in a paracrine fashion (79)(80). Senescence may explain the low-grade inflammation which plays a major role in SCZ onset and maintenance. Indeed, SCZ-associated premature cellular senescence affects all tissues and organs, including the gut and blood-brain barrier (BBB), likely accounting for increased microbial translocation from the gastrointestinal (GI) tract into the host systemic circulation (81). In this regard, lipopolysaccharide (LPS), a cell wall component of Gram-negative bacteria, was detected postmortem in Alzheimer’s disease (AD) brains, suggesting that it migrates from the gut through a senescent intestinal barrier (82). This is significant as impaired intestinal permeability was documented in SCZ, explaining the high comorbidity of this disorder with inflammatory bowel disease (IBD)(81)(83).
Upon CNS entry, microbes, or their components, including LPS or mcfDNA, activate microglia, converting these brain macrophages into neurotoxic cells known to engage in the elimination of healthy neurons, probably including VENs (84). Furthermore, a recent study detected plasma antibodies against specific gut microbes, including Hafnei alvei, Pseudomonas aeruginosa, Morganella morganii, Pseudomonas putida, and Klebsiella pneumoniae in patients with negative SCZ symptoms, linking this condition to microbial translocation (85)(86).
Treatment with antipsychotic drugs affect neuronal cells via receptor-dependent and receptor-independent mechanisms. The latter include, among other changes, peroxidation of cell membrane lipids and intercalation of the drug in neuronal membrane’s lipid bilayer (37)(87)(88)(89)(Figure1). Moreover, antipsychotics-induced peroxidation of cell membrane lipids and iron upregulation may trigger neuronal loss by ferroptosis, a nonapoptotic cell death caused by excessive iron in the context of decreased antioxidants, such as glutathione peroxidase 4 (GPX-4)( 90)(91).
We hypothesize that, due to their large size, VENs are vulnerable to lipid peroxidation, explaining the preferential loss of these cells in bvFTD and probably SCZ (92). Moreover, mcfDNA, derived from the disintegration of translocated bacteria, induces Tau hyperphosphorylation and its accumulation in VENs, leading to the selective demise of these cells (3)(93). This may be significant as anti-ferroptotic agents as well as iron chelators may comprise viable treatment options for bvFTD and other dementias (94)(95)(96).
Schizophrenia outcome studies - Kraepelin was right!
Numerous studies have shown unfavorable longitudinal progress in SCZ, accounting for the continued need for long-term psychiatric facilities, such as State hospitals. For example, a century ago, there were large public institutions for tuberculosis, leprosy, and mental illness, while today only the latter remain in existence (52). This is proof of concept that sustained recovery in patients with SCZ or SLDs remains inadequate. Indeed, those of us who work routinely with SCZ patients, know that only a small number of individuals diagnosed with this disorder return to their premorbid level of functioning (97)(98)(99). For example, 33% of SCZ patients relapse during the first 12 months after an initial psychotic episode, 26% remain homeless at 2 years follow-up, while 5 years after the first psychotic outbreak, only 10% are employed (100)(101)(102). Overall, 13.5% of SCZ patients meet recovery criteria at any point in time after the first psychotic episode (102). These outcome studies are in line with a large metanalysis by Warner, R who looked at the entire 20th century and found that prognosis today is not different than in the first decades of the past century (103)(53) (Table 1).
Furthermore, contrary to the expectations of most clinicians, the recovery rate following the introduction of antipsychotic drugs was not very different compared to the pre-psychopharmacological era (53). Moreover, other studies have shown that employment of SCZ patients has decreased steadily over the past decades, a finding in line with neuroimaging studies which show lifelong gray matter loss, causing progressive disability (72)(104) (Figure 2).
Indeed, progressive cortical tinning which seems to occur regardless of antipsychotic treatment is in line with the natural progression of SCZ toward disability and cognitive deterioration (106)(107)(108)( Figure 2). In contrast to AD which is characterized by impaired recall, cognitive problems in schizophrenia stem primarily from dysfunctional learning of new information (15)(109). We surmise that defective VENs and the subsequent anosognosia are drivers of SCZ, while all the other symptoms may be secondary to this pathology.
Taken together, SCZ outcome studies suggest that the long-term evolution of chronic psychotic disorders leads to disability and cognitive deficits, regardless of treatment with DA-blocking agents. Moreover, anosognosia, the cardinal and driving SCZ mechanism, is rarely ameliorated by DA-blockers, suggesting that these drugs do not affect the root cause of the disease. Furthermore, novel data has corelated anosognosia with aggressive behaviors.
The molecular basis of SCZ and dementia: Tau protein loss of function
SCZ and dementia likely meet at the molecular level as both conditions have been associated with microtubular instability. Recent studies have demonstrated that SCZ is characterized by decreased microtubule-associated protein 2 (MAP2) or Tau which functions as a “molecular velcro”, holding axonal microtubules (MTs) together (110)(Figure 3).
MTs are components of cellular cytoskeleton which, aside from maintaining the cell shape, participate in awareness and insight, suggesting that at the molecular level, anosognosia in SCZ may be the result of unstable MT due to decreased Tau (111)(112)(113). Indeed, anosognosia in SCZ may be caused by dysfunctional MTs in VENs, cells with long axons which rely on MTs for intracellular communication. Along this line, the role of MTs in cognition, self-awareness and insight is further substantiated by anesthesia as anesthetics bind MTs at the Tau site, likely destabilizing these proteins. Removal of Tau, may account for both loss of awareness during anesthesia and postoperative cognitive dysfunction (PCD)(114)(115). Another example of Tau-mediated insight impairment, is “chemo-brain”, a phenomenon resulting from treatment of cancer patients with MT-targeting drugs, such as Taxol, which is often followed by changes in the cognitive and thought-process which may lower insight, linking further MTs to cognition and interoceptive awareness (116). Moreover, Tau and MTs are expressed in other tissues, including the heart which is known for possessing its own particular cognizance, distinct from the cortical awareness and memory (117)(118). For example, it has been established that following heart transplants, recipients often acquire not only select memories but also personality traits of the donor, further linking MTs to awareness, insight, and recall (119)(120)(121)(122). Indeed, local or cellular memory has been described in other tissues, including fascia, muscle, and gut microbiota, linking higher intellectual functions to MTs and Tau (123)(124). Furthermore, gut microbes contain primitive MTs and can transfer learned behavior from one cell to another, indicating that even rudimentary recall may be mediated by the MT and exported along with these molecules to other cells (125). Along this line, translocated microbes may ‘instruct” host cells in Tau aggregation as microbial DNA promotes pathological Tau (126). Viruses, including SARS-CoV-2 can spread Tau aggregates or neurofibrillary tangles (NFTs) from cell to cell in a prion-like manner, indicating that microorganisms may alter cognition, insight, and interoception (127)(Figure 3). Furthermore, upregulated intracellular iron in senescent cells can also promote Tau aggregation, predisposing to neurodegeneration as well as to loss of VENs by ferroptosis.
As opposed to dementia, including bvFTD, SCZ lacks NFTs, however, loss of Tau protein and its function can engender same results, unstable MTs in VENs, altering information processing (113) (126)(129). Another VENs vulnerability that may lead to their preferential loss in bvFTD and possibly SCZ, are lysosomes which appear to exhibit high affinity for NFTs, preferentially storing these aggregates (130).
Taken together, MTs have been hypothesized to play a major role in self-awareness, memory, and insight as they connect via integrins to extracellular matrix proteins, probably engendering CNS-wide molecular networks (131).
bvFTD: from insight to acquired psychopathy
BvFTD is a common, early-onset dementia and like other neurocognitive disorders is a clinical syndrome which can be easily confused with the primary psychiatric disorders, an error which in forensic setting can complicate patients’ legal status, placement, and sentencing (132)(133). The clinical characteristics of bvFTD include, early onset (compared to AD), criminal behavior (reported in 37–57% of patients), and neuropsychiatric symptoms, including apathy, disinhibition, and compulsions (134)(135). An interesting and at the same time confusing features of bvFTD consist of criminal violations without overt memory loss, at least during the early disease phases, a puzzling situation for forensic evaluators who may not suspect a neurodegenerative disorder (136).
Lack of specific neuroimaging and laboratory biomarkers, especially in the early disease stages, adds to the challenges of differentiating this syndrome from SCZ, bipolar disorder, or major depressive disorder (MDD) (137). Indeed, over half of bvFTD patients had psychiatric diagnoses prior to the emergence of clear neurodegenerative signs, an error interfering with adequate management and court proceedings (138).
The prevalence of bvFTD in forensic setting has increased dramatically over the past decade (measured by the surge of late-life first offenders), highlighting the importance of educating the clinicians in keeping a high level of suspicion in first offenders over the age of 50. It is also crucial for the State hospitals to capture data on the number of older first offenders.
The correct diagnosis in forensic setting has a significant bearing on the adjudication and sentencing as judicial approach to criminal violations in individuals with neurodegenerative disorders is usually different than the classical “insanity defense” (139).
We recommend that all first offenders 50 years of age or older be screened for bvFTD. Screening should be accomplished by neuropsychological profile which usually reveal executive impairments and relative sparing of memory and visuospatial functions. Microbial translocation component can be evaluated by mcfDNA Karius Test®® and IL-22 Singulex-Erenna®® (Table 2). In addition, a task force should be established at local or state level, comprised of neuropsychologists, psychiatrists, and medicine/neurology to screen for bvFTD and report to courts for appropriate action. Along this line, in 2015 at Patton State Hospital, California, we have initiated a program for educating our clinicians on bvFTD via presentations, case reports, grand rounds, and publications to help identify this pathology early, if possible, at admission (140). This idea was later adopted by other groups and clinician education in bvFTD continues to be implemented up to date in various institutions and jurisdictions (139).
Like bvFTD, SCZ is a syndrome which in the second half of life is characterized by a “stable phase” with negative and cognitive symptoms, resembling dementia (Figure 2). This complicates not only the differential diagnosis but also the psychopharmacological management of SCZ with overlapping dementia. Indeed, comorbidity of SCZ with bvFTD or other dementias poses special challenges to the clinician as antipsychotic drugs were associated with serious, even fatal, adverse effects when administered to dementia patients (141)(142). In this regard, clinicians may find themselves in a catch 22 as continuing antipsychotic treatment in PLWS and comorbid dementia may risk untoward effects, while discontinuing this therapy may lead to the reemergence of the dangerous behaviors. This dilemma calls for an interdisciplinary approach and involvement of hospital bioethics committee.
Neuropathological basis of bvFTD
VENs, numbering about 193,000 cells, are large, corkscrew neurons located in layer V of the IC, ACC, and FPC. These large, non-telencephalic cells project to various brain areas and participate in prosocial cognition, empathy, and emotional intelligence. As parts of the SN, VENs respond to endogenous or exogenous stimuli in the order of priority (69)(133).
SN dysfunction, documented in psychopathy and criminal behavior, connect this neuronal assembly to delinquency, however, depending on the jurisdiction, this evidence may not be allowed in a court of law (134). Interestingly, a recent study connected the SN with gut microbes Prevotella and Bifidobacterium, emphasizing that the microbiome can influence the function of VENs as well as the behavior, probably via MTs (144)(145). In addition, both Prevotella and VENs have been associated with suicidal behavior, further linking violence to dysfunctional gut permeability and microbial translocation outside the GI tract (146).
The homeostasis of intestinal barrier is maintained by interleukin 22 (IL-22), a cytokine regulated by aryl hydrocarbon receptor (AhR), a gut barrier protein responding to exogenous and endogenous ligands (63). In this regard, VENs-rich IC was demonstrated to keep the record of gut inflammations, emphasizing the tight link between the GI tract and insula (147)(148).
Dementia in PLWS, potential biomarkers
A novel marker of microbial translocation, mcfDNA, is likely to become a diagnostic tool for evaluating dementia in PLWS, while at the same time providing proof of concept that microbes can drive neuropathology (126)(149)(150). For example, the commercially available assay, mcfDNA Karius Test®® detects peripheral blood microbial DNA, a molecule demonstrated to drive Tau aggregation, predisposing to dementia (93)(126)(Table 2). Conversely, SCZ is characterized by downregulated Tau, indicating that this laboratory value could differentiate between SCZ with or without dementia (151)(152)(153)(154). For example, blood levels of phosphorylated tau (pTau) 217 and 181, elevated in dementia but normal in SCZ, can be helpful for differential diagnosis.
Another assay for gut barrier integrity available on the market, IL-22 Singulex-Erenna®®, measures IL-22 levels, a cytokine inversely corelated with gut barrier permeability and microbial translocation (Table 2). For example, low IL-22 with normal mcfDNA would reflect increased risk of microbial translocation without overt pathology. Conversely, low IL-22 and elevated mcfDNA would characterize frank bacterial translocation, requiring treatment. Interestingly, most antipsychotic drugs exert antibiotic properties, suggesting that blocking postsynaptic DA signaling may not be the only action mechanism of these agents. Moreover, IL-22 and mcfDNA could be useful for predicting the risk of relapse following discontinuation of antipsychotic drugs in forensic detainees.
Another marker that could differentiate SCZ from dementia, is ferritin (elevated in dementia and low in SCZ), implicating iron dysmetabolism in both syndromes (90)(157). Excessive intracellular iron associated with senescent cells, a predominant SCZ phenotype, was shown to induce Tau aggregation, implicating this biometal in tauopathies, including bvFTD (158)(159). Indeed, quantitative assessment of iron deposition is a new neuroimaging biomarker of bvFTD that can differentiate this phenotype from other FTD subtypes (160)(161). This suggests that in bvFTD, VENs may be lost through ferroptosis, a nonapoptotic cell death caused by iron-induced lipid peroxidation in the absence of glutathione-associated antioxidants. This is significant as intranasal deferoxamine was found effective for several neurodegenerative disorders, including bvFTD (162).
Chronic Traumatic Encephalopathy
A condition similar to bvFTD, chronic traumatic encephalopathy (CTE), is a pathology associated with repeated traumatic brain injuries (TBIs), manifested by behavioral changes, including irritability, apathy, memory loss, and intermittent disorientation (163)(164)(165). Upregulated iron and pTau may be the common denominators of bvFTD and CTE, emphasizing that neurodegeneration can be initiated by a localized or more generalized and diffuse pathology (166)(167). Along this line, the former NFL player, Phillip Adams, who killed six people, and committed suicide while serving his sentence was found at autopsy to have had CTE, a condition some clinicians believed should have been factored in sentencing.
Interventions: receptor-independent antipsychotic treatments (RIATs)
The currently available antipsychotic drugs are extremely effective for the treatment of acute psychotic disorders, however their efficacy in chronic psychosis is much less dramatic and may even predispose to neurocognitive disorders by the lipid peroxidation of cell membranes. Dysfunctional cell membranes may lead to increased permeability of gut barrier and BBB, enabling microbial translocation from the GI tract into the systemic circulation, eventually reaching the brain.
Neurolipidomics is a rapidly growing field made possible by the recent advances in mass spectrometry (MS), a technique capable of quickly processing biomolecules, identifying in seconds the lipid species involved in various CNS pathologies (168)(169). Neurolipidomics has facilitated the development of receptor-independent antipsychotic treatments (RIATs), including LRT.
Lipid replacement therapy (LRT) is a technique which utilizes healthy, natural glycerophospholipids to substitute oxidized components of the plasma membranes lipid bilayer, restoring the physiological fluidity as well as neurotransmitter signaling. This approach, based on the oral supplementation with natural phospholipids and antioxidants, was demonstrated to halt the dissemination of cellular senescence to the neighboring, healthy cells, probably by inhibiting SASP (170)(171)(172)(173)(174). Moreover, LRT was shown to supplant not only cell membranes but also the damaged mitochondrial inner and outer membranes with new and natural lipid species. Indeed, loss of lysophosphatidylethanolamine (LPE), phosphatidylglycerol (PG), and phosphatidylinositol (PI) in senescent mitochondria was shown to cause organelle demise; conversely replacement with healthy lipids promotes mitochondrial thriving (173).
Phosphoinositide-dependent kinase 1 (PDK-1) inhibitors
We surmise that combining LRT with inhibitors of phosphoinositide-dependent kinase 1 (PDK-1), such as Kaempferol, would produce superior results in SCZ compared to LRT alone. Indeed, PDK1 inhibitors reverse cellular senescence, a phenotype previously considered irreversible. Lowered SASP together with the healthy exogenous lipids decrease the risk of peroxidation and ferroptosis (174). Interestingly, Kaempferol is also an antagonist of AhR, a transcription factor and cellular senescence driver. On the other hand, PDK1 activation of protein kinase B (Akt) and glycogen synthase kinase 3 beta (GSK-3β) promotes SCZ pathogenesis (175)(Figure3). The point of contact between exogenously administered phospholipids and PDK1 is phosphatidylserine (PS) which binds PDK1 through its PH domain, blocking PS externalization and the initiation of cellular senescence. Moreover, PH-PS attachment inhibits PDK1 activation of downstream kinases, accounting for the antipsychotic actions of PDK1 inhibitors (176)(Figure 4).
Taken together, LRT inhibits PDK1 as well as Akt and GSK-3β, exerting antipsychotic effects by entirely natural means.
Recombinant human interleukin-22 (IL-22)
Hinting at the microbiome, Emil Kraepelin stated that SCZ was caused by toxins generated in various body regions that could eventually reach the brain, triggering psychosis (66)(177). In addition, the membrane phospholipid hypothesis of SCZ, developed in the 1990s, foreshadowed LRT by predicting that cell membranes can be rehabilitated by exogenous phospholipids (178).
Microbial translocation from the GI tract into the host circulatory system has been proposed over 100 years ago but completely forgotten after Kraepelin’s death. Along this line, the “Scientific American” from October 1896, Volume 75, Issue 15, ran the story “Is Insanity Due to a Microbe?”, highlighting bacteria-mediated neuropathology, the model supported by the scientists of that time (Figure 5).
During the HIV epidemic of the 1980s, microbial translocation, a common phenomenon encountered in this disease, was linked to virus-depleted IL-22, a cytokine considered “the guardian” of gut barrier (179). The same pathology was reported during the COVID-19 pandemic, emphasizing further that viruses and bacteria can trigger psychosis (180)(181). For this reason, we believe that supplementation with exogenous IL-22 can restore the integrity of gut barrier, ameliorating the symptoms of both SCZ and dementia. Recombinant human interleukin-22 (IL-22), currently being developed as a cancer therapy, is a cytokine known for protecting the biological barriers, iron downregulation, and stimulation of adult neurogenesis, suggesting a beneficial role in SCZ (181).
Aryl hydrocarbon receptor (AhR) antagonists
AhR is a cytosolic transcription factor, initially associated with dioxin toxicity, which responds to numerous endogenous and exogenous stimuli, including xenobiotics (183). In the cytoplasm, AhR is anchored by two molecules of heat shock protein 90 (HSP90) and upon detachment from these chaperones, it enters the nucleus to activate or silence the transcription of many genes. Aside from dioxin, AhR binds several ligands relevant for neuropsychiatry, including DA, serotonin, and clozapine, bringing this receptor into the psychopharmacological arena (184)(185)(Figure 6). In contrast, aripiprazole augments the HSP90 link, preventing AhR dissociation and nuclear entry, evidencing AhR antagonist effects (186).
Dietary AhR antagonists, such as Resveratrol, Luteolin, Kaempferol, Quercetin and the synthetic compounds CH223191, alpha-Naphthoflavone, 6-bromo-3'-nitroflavone, BAY2416964, HBU651, are beneficial by lowering cellular senescence, ameliorating psychosis, and cognition (187)(188). Furthermore, AhR exerts antagonistic pleiotropy (positive effects during the development, and detrimental ones later in life). Pleiotropy is responsible for the dioxin property of activating AhR in adulthood, inducing premature cellular senescence and cognitive impairment (189)(190)(191).
Conclusions
While acute psychosis responds very well to antipsychotic drugs, chronic psychotic illnesses are much more refractory to these agents. Over the past decades, psychopharmacology has focused excessively on the receptor-dependent actions of antipsychotic drugs and put much less emphasis on the receptor-independent ones, such as antimicrobial and anticancer actions. These are significant for the elimination of translocated microbes and de-escalation of immune responses directed at microbial molecules, such as LPS or mcfDNA. The neglect of noncanonical action mechanisms of antipsychotic drugs over the past decades, has severely limited the development of new models and therapies, leaving chronic psychotic illnesses dependent on the inefficient treatments for acute psychosis.
The long-term exposure to conventional antipsychotic drugs may have detrimental effects on cell membrane lipidome, including peroxidation and intercalation into the lipid bilayer. In return, this may lead to neuronal damage and gray matter loss documented by several neuroimaging studies. To avert neuronal loss by cell membrane damage, elderly with SCZ should receive LRT and PDK1 inhibitors in conjunction with conventional antipsychotics.
Exogenous human recombinant IL-22 protects intestinal epithelia, preventing the microbes or their components from translocating outside the GI tract. This in return averts aberrant immune system activation and the subsequent end-organ damage. Moreover, AhR inhibitors reverse the antagonistic pleiotropy by blocking late-life activation of this protein.
RIATs are likely beneficial for PLWS and comorbid dementia as these conditions are characterized by damaged lipidomes and cell membranes as well as increased intestinal permeability. Elderly forensic detainees in treatment with conventional antipsychotic drugs should receive lipid supplementation and PDK1 inhibitors to delay the onset of dementia. More studies are needed to evaluate the beneficial effects of these strategies as well as that of iron chelators for restoring cognitive capabilities in forensic detainees and ultimately their capacity to stand trial.
References
- Mendez, M.F. The Neurobiology of Moral Behavior: Review and Neuropsychiatric Implications. CNS Spectrums 2009, 14, 608–620. [Google Scholar] [CrossRef]
- Savopoulos, P.; Lindell, A.K. Born criminal? Differences in structural, functional and behavioural lateralization between criminals and noncriminals. Laterality 2018, 23, 738–760. [Google Scholar] [CrossRef]
- Shapira, J.S.; Saul, R.E.; Guimet, N.M.; Zapata-Restrepo, L.M.; Miller, B.L.; Mendez, M.F.; Moheb, N.; Charuworn, K.; Ashla, B.M.M.; Desarzant, B.R.; et al. The Spectrum of Sociopathy in Dementia. J. Neuropsychiatry 2011, 23, 132–140. [Google Scholar] [CrossRef]
- Lehrer, D.S.; Lorenz, J. Anosognosia in schizophrenia: hidden in plain sight. Innov. Clin. Neurosci. 2014, 11, 10–17. [Google Scholar]
- Räsänen, P.; Tähonen, J.; Isohanni, M.; Rantakallio, P.; Lehtonen, J.; Moring, J. Schizophrenia, Alcohol Abuse, and Violent Behavior: A 26-Year Followup Study of an Unselected Birth Cohort. Schizophr. Bull. 1998, 24, 437–441. [Google Scholar] [CrossRef]
- Polat, H.; Uğur, K.; Aslanoğlu, E.; Yıldız, S.; Yagin, F.H. The effect of functional remission and cognitive insight on criminal behavior in patients with schizophrenia. Arch. Psychiatr. Nurs. 2023, 45, 176–183. [Google Scholar] [CrossRef]
- Lien, Y.-J.; Chang, H.-A.; Kao, Y.-C.; Tzeng, N.-S.; Lu, C.-W.; Loh, C.-H. Insight, self-stigma and psychosocial outcomes in Schizophrenia: a structural equation modelling approach. Epidemiology Psychiatr. Sci. 2016, 27, 176–185. [Google Scholar] [CrossRef]
- Lincoln, T.M.; Hodgins, S. Is Lack of Insight Associated With Physically Aggressive Behavior Among People With Schizophrenia Living in the Community? J. Nerv. Ment. Dis. 2008, 196, 62–66. [Google Scholar] [CrossRef]
- Buckley, P.F.; Hrouda, D.R.; Friedman, L.; Noffsinger, S.G.; Resnick, P.J.; Camlin-Shingler, K. Insight and Its Relationship to Violent Behavior in Patients With Schizophrenia. Am. J. Psychiatry 2004, 161, 1712–1714. [Google Scholar] [CrossRef]
- Zago, S.; Scarpazza, C.; Difonzo, T.; Arighi, A.; Hajhajate, D.; Torrente, Y.; Sartori, G. Behavioral Variant of Frontotemporal Dementia and Homicide in a Historical Case. J Am Acad Psychiatry Law. 2021, 49, 219–227. [Google Scholar] [CrossRef]
- Wander, C. Schizophrenia: opportunities to improve outcomes and reduce economic burden through managed care. The American Journal of Managed Care 2020, 26, S62–S68. [Google Scholar] [CrossRef] [PubMed]
- Dregan, A.; McNeill, A.; Gaughran, F.; Jones, P.B.; Bazley, A.; Cross, S.; Lillywhite, K.; Armstrong, D.; Smith, S.; Osborn, D.P.J.; et al. Potential gains in life expectancy from reducing amenable mortality among people diagnosed with serious mental illness in the United Kingdom. PLOS ONE 2020, 15, e0230674. [Google Scholar] [CrossRef] [PubMed]
- Ilyas, A.; Chesney, E.; Patel, R. Improving life expectancy in people with serious mental illness: Should we place more emphasis on primary prevention? Br. J. Psychiatry 2017, 211, 194–197. [Google Scholar] [CrossRef] [PubMed]
- Stoliker, B.E.; Kerodal, A.G.; Jewell, L.M.; Brown, K.; Kent-Wilkinson, A.; Peacock, S.; O’connell, M.E.; Wormith, J.S. Older people in custody in a forensic psychiatric facility, prevalence of dementia, and community reintegration needs: an exploratory analysis. Health Justice 2022, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Huang, J. Schizophrenia and risk of dementia: a meta-analysis study. Neuropsychiatr. Dis. Treat. 2018, 14, 2047–2055. [Google Scholar] [CrossRef] [PubMed]
- Kales, H.C.; Valenstein, M.; Kim, H.M.; McCarthy, J.F.; Ganoczy, D.; Cunningham, F.; Blow, F.C. Mortality Risk in Patients With Dementia Treated With Antipsychotics Versus Other Psychiatric Medications. Am. J. Psychiatry 2007, 164, 1568–1576. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Bronskill, S.E.; Normand, S.-L.T.; Anderson, G.M.; Sykora, K.; Lam, K.; Bell, C.M.; Lee, P.E.; Fischer, H.D.; Herrmann, N.; et al. Antipsychotic Drug Use and Mortality in Older Adults with Dementia. Ann. Intern. Med. 2007, 146, 775–786. [Google Scholar] [CrossRef]
- Randle, J.M.; Heckman, G.; Oremus, M.; Ho, J. Intermittent antipsychotic medication and mortality in institutionalized older adults: A scoping review. Int. J. Geriatr. Psychiatry 2019, 34, 906–920. [Google Scholar] [CrossRef]
- Desai, V.C.; Heaton, P.C.; Kelton, C.M. Impact of the Food and Drug Administration's antipsychotic black box warning on psychotropic drug prescribing in elderly patients with dementia in outpatient and office-based settings. Alzheimer's Dement. 2012, 8, 453–457. [Google Scholar] [CrossRef]
- Tessier, C.; Sweers, K.; Frajerman, A.; Bergaoui, H.; Ferreri, F.; Delva, C.; Lapidus, N.; Lamaziere, A.; Roiser, J.P.; De Hert, M.; et al. Membrane lipidomics in schizophrenia patients: a correlational study with clinical and cognitive manifestations. Transl. Psychiatry 2016, 6, e906. [Google Scholar] [CrossRef]
- Maxwell, C.; Kanes, S.; Abel, T.; Siegel, S. Phosphodiesterase inhibitors: A novel mechanism for receptor-independent antipsychotic medications. Neuroscience 2004, 129, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Surmeier, D.J.; Shen, W.; Day, M.; Gertler, T.; Chan, S.; Tian, X.; Plotkin, J.L. The role of dopamine in modulating the structure and function of striatal circuits. Prog Brain Res. 2010, 183, 148–167. [Google Scholar] [CrossRef]
- Shen, W.; Flajolet, M.; Greengard, P.; Surmeier, D.J. Dichotomous Dopaminergic Control of Striatal Synaptic Plasticity. Science 2008, 321, 848–851. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Kai, L.; Day, M.; Ronesi, J.; Yin, H.H.; Ding, J.; Tkatch, T.; Lovinger, D.M.; Surmeier, D.J. Dopaminergic Control of Corticostriatal Long-Term Synaptic Depression in Medium Spiny Neurons Is Mediated by Cholinergic Interneurons. Neuron 2006, 50, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, A.; Voineskos, D.; Daskalakis, Z.J.; Rajji, T.K.; Blumberger, D.M. A Review of Impaired Neuroplasticity in Schizophrenia Investigated with Non-invasive Brain Stimulation. Front. Psychiatry 2016, 7, 45–45. [Google Scholar] [CrossRef] [PubMed]
- Goel, P.; Chakrabarti, S.; Goel, K.; Bhutani, K.; Chopra, T.; Bali, S. Neuronal cell death mechanisms in Alzheimer’s disease: An insight. Front. Mol. Neurosci. 2022, 15, 937133. [Google Scholar] [CrossRef]
- Berry, A.S.; Shah, V.D.; Baker, S.L.; Vogel, J.W.; O'Neil, J.P.; Janabi, M.; Schwimmer, H.D.; Marks, S.M.; Jagust, W.J. Aging Affects Dopaminergic Neural Mechanisms of Cognitive Flexibility. J. Neurosci. 2016, 36, 12559–12569. [Google Scholar] [CrossRef]
- Braskie, M.N.; Wilcox, C.E.; Landau, S.M.; O'Neil, J.P.; Baker, S.L.; Madison, C.M.; Kluth, J.T.; Jagust, W.J. Relationship of Striatal Dopamine Synthesis Capacity to Age and Cognition. J. Neurosci. 2008, 28, 14320–14328. [Google Scholar] [CrossRef]
- Huber, M.; Beyer, L.; Prix, C.; Schönecker, S.; Palleis, C.; Rauchmann, B.; Morbelli, S.; Chincarini, A.; Bruffaerts, R.; Vandenberghe, R.; et al. Metabolic Correlates of Dopaminergic Loss in Dementia with Lewy Bodies. Mov. Disord. 2019, 35, 595–605. [Google Scholar] [CrossRef]
- Lubec, J.; Kalaba, P.; Hussein, A.M.; Feyissa, D.D.; Kotob, M.H.; Mahmmoud, R.R.; Wieder, O.; Garon, A.; Sagheddu, C.; Ilic, M.; et al. Reinstatement of synaptic plasticity in the aging brain through specific dopamine transporter inhibition. Mol. Psychiatry 2021, 26, 7076–7090. [Google Scholar] [CrossRef]
- Canfrán-Duque, A.; Pastor, Ó.; García-Seisdedos, D.; Molina, Y.L.; Babiy, B.; Lerma, M.; Sánchez-Castellano, C.; Martínez-Botas, J.; Gómez-Coronado, D.; Lasunción, M.A.; et al. The Antipsychotic Risperidone Alters Dihydroceramide and Ceramide Composition and Plasma Membrane Function in Leukocytes In Vitro and In Vivo. Int. J. Mol. Sci. 2021, 22, 3919. [Google Scholar] [CrossRef] [PubMed]
- Vantaggiato, C.; Panzeri, E.; Citterio, A.; Orso, G.; Pozzi, M. Antipsychotics Promote Metabolic Disorders Disrupting Cellular Lipid Metabolism and Trafficking. Trends Endocrinol. Metab. 2019, 30, 189–210. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.; Au, E.; Agarwal, S.M.; Wright, D.C.; Hahn, M.K. Antipsychotic-Induced Alterations in Lipid Turnover. Endocrinology 2023, 164. [Google Scholar] [CrossRef] [PubMed]
- Dietrich-Muszalska, A.; Kolińska-Łukaszuk, J. Comparative effects of aripiprazole and selected antipsychotic drugs on lipid peroxidation in plasma. Psychiatry Clin. Neurosci. 2018, 72, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Kuzu, O.F.; Toprak, M.; Noory, M.A.; Robertson, G.P. Effect of lysosomotropic molecules on cellular homeostasis. Pharmacol. Res. 2017, 117, 177–184. [Google Scholar] [CrossRef]
- Róg, T.; Girych, M.; Bunker, A. Mechanistic Understanding from Molecular Dynamics in Pharmaceutical Research 2: Lipid Membrane in Drug Design. Pharmaceuticals 2021, 14, 1062. [Google Scholar] [CrossRef]
- Elbaradei, A.; Wang, Z.; Malmstadt, N. Oxidation of Membrane Lipids Alters the Activity of the Human Serotonin 1A Receptor. Langmuir 2022, 38, 6798–6807. [Google Scholar] [CrossRef]
- Li, M.; Gao, Y.; Wang, D.; Hu, X.; Jiang, J.; Qing, Y.; Yang, X.; Cui, G.; Wang, P.; Zhang, J.; et al. Impaired Membrane Lipid Homeostasis in Schizophrenia. Schizophr. Bull. 2022, 48, 1125–1135. [Google Scholar] [CrossRef]
- Pillai, A.; Parikh, V.; Terry, A.V.; Mahadik, S.P. Long-term antipsychotic treatments and crossover studies in rats: Differential effects of typical and atypical agents on the expression of antioxidant enzymes and membrane lipid peroxidation in rat brain. J. Psychiatr. Res. 2007, 41, 372–386. [Google Scholar] [CrossRef]
- Oruch, R.; Lund, A.; Pryme, I.F.; Holmsen, H. An intercalation mechanism as a mode of action exerted by psychotropic drugs: results of altered phospholipid substrate availabilities in membranes? J. Chem. Biol. 2010, 3, 67–88. [Google Scholar] [CrossRef]
- Schoretsanitis, G.; Nikolakopoulou, A.; Guinart, D.; Correll, C.U.; Kane, J.M. Iron homeostasis alterations and risk for akathisia in patients treated with antipsychotics: A systematic review and meta-analysis of cross-sectional studies. Eur. Neuropsychopharmacol. 2020, 35, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chong, S.-A.; Mythily; Remington, G. Tardive Dyskinesia and Iron Status. J. Clin. Psychopharmacol. 2004, 24, 235–236. [Google Scholar] [CrossRef] [PubMed]
- Calarge, C.A.; Ziegler, E.E.; Del Castillo, N.; Aman, M.; McDougle, C.J.; Scahill, L.; McCracken, J.T.; Arnold, L.E. Iron homeostasis during risperidone treatment in children and adolescents. J. Clin. Psychiatry 2015, 76, 1500–1505. [Google Scholar] [CrossRef] [PubMed]
- Dichtl, S.; Demetz, E.; Haschka, D.; Tymoszuk, P.; Petzer, V.; Nairz, M.; Seifert, M.; Hoffmann, A.; Brigo, N.; Würzner, R.; et al. Dopamine Is a Siderophore-Like Iron Chelator That Promotes Salmonella enterica Serovar Typhimurium Virulence in Mice. Mbio 2019, 10, e02624-18. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Gan, M.; Yen, S.-H.C. Dopamine prevents lipid peroxidation-induced accumulation of toxic α-synuclein oligomers by preserving autophagy-lysosomal function. Front. Cell. Neurosci. 2013, 7, 81. [Google Scholar] [CrossRef] [PubMed]
- Tung, M.-C.; Lin, Y.-W.; Lee, W.-J.; Wen, Y.-C.; Liu, Y.-C.; Chen, J.-Q.; Hsiao, M.; Yang, Y.-C.; Chien, M.-H. Targeting DRD2 by the antipsychotic drug, penfluridol, retards growth of renal cell carcinoma via inducing stemness inhibition and autophagy-mediated apoptosis. Cell Death Dis. 2022, 13, 400. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Chen, H.; Chen, K.; Zhong, C.; Song, C.; Huang, Y.; Wang, T.; Chen, L.; Li, C.; Huang, A.; et al. The DRD2 Antagonist Haloperidol Mediates Autophagy-Induced Ferroptosis to Increase Temozolomide Sensitivity by Promoting Endoplasmic Reticulum Stress in Glioblastoma. Clin. Cancer Res. 2023, OF1–OF17. [Google Scholar] [CrossRef] [PubMed]
- Vita, A.; De Peri, L.; Deste, G.; Barlati, S.; Sacchetti, E. The Effect of Antipsychotic Treatment on Cortical Gray Matter Changes in Schizophrenia: Does the Class Matter? A Meta-analysis and Meta-regression of Longitudinal Magnetic Resonance Imaging Studies. Biol. Psychiatry 2015, 78, 403–412. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Liao, J.; Jiang, S.; Yan, J.; Yue, W.; Zhang, D.; Yan, H. Progressive Grey Matter Volume Changes in Patients with Schizophrenia over 6 Weeks of Antipsychotic Treatment and Their Relationship to Clinical Improvement. Neurosci. Bull. 2018, 34, 816–826. [Google Scholar] [CrossRef]
- Chen, Y.; Womer, F.Y.; Feng, R.; Zhang, X.; Zhang, Y.; Duan, J.; Chang, M.; Yin, Z.; Jiang, X.; Wei, S.; et al. A Real-World Observation of Antipsychotic Effects on Brain Volumes and Intrinsic Brain Activity in Schizophrenia. Front. Neurosci. 2022, 15, 749316. [Google Scholar] [CrossRef]
- Pustilnik, Amanda C., "Violence on the Brain: A Critique of Neuroscience in Criminal Law" (2009). Faculty Scholarship. 1035. Available online: https://digitalcommons.law.umaryland.edu/fac_pubs/1035.
- Insel, T.R. Rethinking schizophrenia. Nature 2010, 468, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Warner, R. Recovery from schizophrenia and the recovery model. Curr. Opin. Psychiatry 2009, 22, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Sfera, A. Six Decades of Dopamine Hypothesis: Is Aryl Hydrocarbon Receptor the New D2? Reports 2023, 6, 36. [Google Scholar] [CrossRef]
- Williams, S.S. The terrorist inside my husband's brain. Neurology 2016, 87, 1308–1311. [Google Scholar] [CrossRef] [PubMed]
- Fields, R.D. THE ROOTS OF HUMAN AGGRESSION: Experiments in humans and animals have started to identify how violent behaviors begin in the brain. Sci Am. 2019, 320, 65–71. [Google Scholar] [PubMed]
- Parton, A.; Malhotra, P.; Husain, M. Hemispatial neglect. J. Neurol. Neurosurg. Psychiatry. 2004, 75, 13–21. [Google Scholar] [PubMed]
- Grattan, E.S.; Skidmore, E.R.; Woodbury, M.L. Examining Anosognosia of Neglect. OTJR: Occup. Particip. Heal. 2017, 38, 113–120. [Google Scholar] [CrossRef]
- Koreki, A.; Funayama, M.; Terasawa, Y.; Onaya, M.; Mimura, M. Aberrant Interoceptive Accuracy in Patients With Schizophrenia Performing a Heartbeat Counting Task. Schizophr. Bull. Open 2020, 2, sgaa067. [Google Scholar] [CrossRef]
- Torregrossa, L.J.; Amedy, A.; Roig, J.; Prada, A.; Park, S. Interoceptive functioning in schizophrenia and schizotypy. Schizophr. Res. 2021, 239, 151–159. [Google Scholar] [CrossRef]
- Yao, B.; Thakkar, K. Interoception abnormalities in schizophrenia: A review of preliminary evidence and an integration with Bayesian accounts of psychosis. Neurosci. Biobehav. Rev. 2021, 132, 757–773. [Google Scholar] [CrossRef]
- Ribolsi, M.; Daskalakis, Z.J.; Siracusano, A.; Koch, G. Abnormal Asymmetry of Brain Connectivity in Schizophrenia. Front. Hum. Neurosci. 2014, 8, 1010. [Google Scholar] [CrossRef] [PubMed]
- Brüne, M.; Schöbel, A.; Karau, R.; Benali, A.; Faustmann, P.M.; Juckel, G.; Petrasch-Parwez, E. Von Economo neuron density in the anterior cingulate cortex is reduced in early onset schizophrenia. Acta Neuropathol. 2010, 119, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Brasso, C.; Stanziano, M.; Bosco, F.M.; Morese, R.; Valentini, M.C.; Vercelli, A.; Rocca, P. Alteration of the Functional Connectivity of the Cortical Areas Characterized by the Presence of Von Economo Neurons in Schizophrenia, a Pilot Study. J. Clin. Med. 2023, 12, 1377. [Google Scholar] [CrossRef] [PubMed]
- López-Ojeda, W.; Hurley, R.A. Von Economo Neuron Involvement in Social Cognitive and Emotional Impairments in Neuropsychiatric Disorders. J. Neuropsychiatry 2022, 34, 302–306. [Google Scholar] [CrossRef] [PubMed]
- Kraepelin, E. (1990) Psychiatry. A Textbook for Students and Physicians, 2 vols, translated by H. Metoui and S. Ayed (Canton, MA: Science History Publications); originally publishedas Psychiatrie. Ein Lehrbuch fur Studirende und Aerzte (6th edn, 1899).
- Voruz, P.; Cionca, A.; de Alcântara, I.J.; Nuber-Champier, A.; Allali, G.; Benzakour, L.; Thomasson, M.; Lalive, P.H.; Lövblad, K.-O.; Braillard, O.; et al. Functional connectivity underlying cognitive and psychiatric symptoms in post-COVID-19 syndrome: is anosognosia a key determinant? Brain Commun. 2022, 4, fcac057. [Google Scholar] [CrossRef]
- Juengst, S.; Skidmore, E.; Pramuka, M.; McCue, M.; Becker, J. Factors contributing to impaired self-awareness of cognitive functioning in an HIV positive and at-risk population. Disabil. Rehabilitation 2011, 34, 19–25. [Google Scholar] [CrossRef]
- Allman, J.M.; Tetreault, N.A.; Hakeem, A.Y.; Park, S. The von economo neurons in apes and humans. Am. J. Hum. Biol. 2010, 23, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Faissner, A. Low-density lipoprotein receptor-related protein-1 (LRP1) in the glial lineage modulates neuronal excitability. Front. Netw. Physiol. 2023, 3, 1190240. [Google Scholar] [CrossRef]
- Pillmann, F.; Rohde, A.; Ullrich, S.; Draba, S.; Sannemüller, U.; Marneros, A. Violence, Criminal Behavior, and the EEG. J. Neuropsychiatry 1999, 11, 454–457. [Google Scholar] [CrossRef]
- Marwaha, S. Prevalence of Principles of Piaget’s Theory Among 4-7-year-old Children and their Correlation with IQ. J. Clin. Diagn. Res. 2017, 11, ZC111–ZC115. [Google Scholar] [CrossRef]
- Gefen, T.; Papastefan, S.T.; Rezvanian, A.; Bigio, E.H.; Weintraub, S.; Rogalski, E.; Mesulam, M.-M.; Geula, C. Von Economo neurons of the anterior cingulate across the lifespan and in Alzheimer's disease. Cortex 2017, 99, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Cauda, F.; Nani, A.; Costa, T.; Palermo, S.; Tatu, K.; Manuello, J.; Duca, S.; Fox, P.T.; Keller, R. The morphometric co-atrophy networking of schizophrenia, autistic and obsessive spectrum disorders. Hum. Brain Mapp. 2018, 39, 1898–1928. [Google Scholar] [CrossRef] [PubMed]
- Papanastasiou, E.; Gaughran, F.; Smith, S. Schizophrenia as segmental progeria. J. R. Soc. Med. 2011, 104, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.Y.; Chang, H.W.; Lin, C.H.; Cho, C.L. Short telomeres in patients with chronic schizophrenia who show a poor response to treatment. J Psychiatry Neurosci. Erratum in: J Psychiatry Neurosci. 2008, 33, 343. 2008, 33, 244–247. [Google Scholar] [PubMed]
- Laursen, T.M. Life expectancy among persons with schizophrenia or bipolar affective disorder. Schizophr. Res. 2011, 131, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Peritogiannis, V.; Ninou, A.; Samakouri, M. Mortality in Schizophrenia-Spectrum Disorders: Recent Advances in Understanding and Management. Healthcare 2022, 10, 2366. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Shen, J.; He, J.; Lu, M. Schizophrenia and cell senescence candidate genes screening, machine learning, diagnostic models, and drug prediction. Front. Psychiatry 2023, 14, 1105987. [Google Scholar] [CrossRef]
- Acosta, J.C.; Banito, A.; Wuestefeld, T.; Georgilis, A.; Janich, P.; Morton, J.P.; Athineos, D.; Kang, T.-W.; Lasitschka, F.; Andrulis, M.; et al. A complex secretory program orchestrated by the inflammasome controls paracrine senescence. Nature 2013, 15, 978–990. [Google Scholar] [CrossRef]
- Ishida, I.; Ogura, J.; Aizawa, E.; Ota, M.; Hidese, S.; Yomogida, Y.; Matsuo, J.; Yoshida, S.; Kunugi, H. Gut permeability and its clinical relevance in schizophrenia. Neuropsychopharmacol. Rep. 2022, 42, 70–76. [Google Scholar] [CrossRef]
- Zhao, Y.; Cong, L.; Lukiw, W.J. Lipopolysaccharide (LPS) Accumulates in Neocortical Neurons of Alzheimer’s Disease (AD) Brain and Impairs Transcription in Human Neuronal-Glial Primary Co-cultures. Front. Aging Neurosci. 2017, 9, 407. [Google Scholar] [CrossRef]
- Sung, K.; Zhang, B.; Wang, H.E.; Bai, Y.; Tsai, S.; Su, T.; Chen, T.; Hou, M.; Lu, C.; Wang, Y.; et al. Schizophrenia and risk of new-onset inflammatory bowel disease: a nationwide longitudinal study. Aliment. Pharmacol. Ther. 2022, 55, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Yanuck, S.F. Microglial Phagocytosis of Neurons: Diminishing Neuronal Loss in Traumatic, Infectious, Inflammatory, and Autoimmune CNS Disorders. Front. Psychiatry 2019, 10, 712. [Google Scholar] [CrossRef] [PubMed]
- Secher, T.; Samba-Louaka, A.; Oswald, E.; Nougayrède, J.-P. Escherichia coli Producing Colibactin Triggers Premature and Transmissible Senescence in Mammalian Cells. PLOS ONE 2013, 8, e77157. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Gao, F.; Zhou, L.; Fan, Y.; Zhao, B.; Xi, W.; Wang, C.; Zhu, F.; Ma, X.; Wang, W.; et al. Characterizing serum amino acids in schizophrenic patients: Correlations with gut microbes. J. Psychiatr. Res. 2022, 153, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Dietrich-Muszalska, A.; Kolodziejczyk-Czepas, J.; Nowak, P. Comparative Study of the Effects of Atypical Antipsychotic Drugs on Plasma and Urine Biomarkers of Oxidative Stress in Schizophrenic Patients. Neuropsychiatr. Dis. Treat. 2021, 17, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Pillai, A.; Parikh, V.; Terry, A.V.; Mahadik, S.P. Long-term antipsychotic treatments and crossover studies in rats: Differential effects of typical and atypical agents on the expression of antioxidant enzymes and membrane lipid peroxidation in rat brain. J. Psychiatr. Res. 2007, 41, 372–386. [Google Scholar] [CrossRef]
- Li, M.; Gao, Y.; Wang, D.; Hu, X.; Jiang, J.; Qing, Y.; Yang, X.; Cui, G.; Wang, P.; Zhang, J.; et al. Impaired Membrane Lipid Homeostasis in Schizophrenia. Schizophr. Bull. 2022, 48, 1125–1135. [Google Scholar] [CrossRef]
- Lotan, A.; Luza, S.; Opazo, C.M.; Ayton, S.; Lane, D.J.R.; Mancuso, S.; Pereira, A.; Sundram, S.; Weickert, C.S.; Bousman, C.; et al. Perturbed iron biology in the prefrontal cortex of people with schizophrenia. Mol. Psychiatry 2023. [Google Scholar] [CrossRef]
- Killilea, D.W.; Atamna, H.; Liao, C.; Ames, B.N. Iron Accumulation During Cellular Senescence in Human FibroblastsIn Vitro. Antioxidants Redox Signal. 2003, 5, 507–516. [Google Scholar] [CrossRef]
- Santillo, A.F.; Nilsson, C.; Englund, E. von E conomo neurones are selectively targeted in frontotemporal dementia. Neuropathol. Appl. Neurobiol. 2013, 39, 572–579. [Google Scholar] [CrossRef]
- Lin, L.-C.; Nana, A.L.; Hepker, M.; Hwang, J.-H.L.; Gaus, S.E.; Spina, S.; Cosme, C.G.; Gan, L.; Grinberg, L.T.; Geschwind, D.H.; et al. Preferential tau aggregation in von Economo neurons and fork cells in frontotemporal lobar degeneration with specific MAPT variants. Acta Neuropathol. Commun. 2019, 7, 159. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Guo, Z. Recent progress in ferroptosis: inducers and inhibitors. Cell Death Discov. 2022, 8, 501. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Dong, Y.; Chu, Y.; Guo, Y.; Li, L. The mechanisms of ferroptosis and its role in alzheimer’s disease. Front. Mol. Biosci. 2022, 9, 965064. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, M.T.; Chana-Cuevas, P. New Perspectives in Iron Chelation Therapy for the Treatment of Neurodegenerative Diseases. Pharmaceuticals 2018, 11, 109. [Google Scholar] [CrossRef] [PubMed]
- Huxley, P.; Krayer, A.; Poole, R.; Prendergast, L.; Aryal, S.; Warner, R. Schizophrenia outcomes in the 21st century: A systematic review. Brain Behav. 2021, 11, e02172. [Google Scholar] [CrossRef]
- Robinson, D.G.; Woerner, M.G.; McMeniman, M.; Mendelowitz, A.; Bilder, R.M.; Velthorst, E.; Fett, A.-K.J.; Reichenberg, A.; Perlman, G.; van Os, J.; et al. Symptomatic and Functional Recovery From a First Episode of Schizophrenia or Schizoaffective Disorder. Am. J. Psychiatry 2004, 161, 473–479. [Google Scholar] [CrossRef]
- Harrison, G.; Hopper, K.; Craig, T.; Laska, E.; Siegel, C.; Wanderling, J.; Dube, K.C.; Ganev, K.; Giel, R.; Der Heiden, W.A.; et al. Recovery from psychotic illness: A 15- and 25-year international follow-up study. Br. J. Psychiatry 2001, 178, 506–517. [Google Scholar] [CrossRef]
- Holm, M.; Taipale, H.; Tanskanen, A.; Tiihonen, J.; Mitterdorfer-Rutz, E. Employment among people with schizophrenia or bipolar disorder: A population-based study using nationwide registers. Acta Psychiatr. Scand. 2020, 143, 61–71. [Google Scholar] [CrossRef]
- Lévesque, I.S.; Abdel-Baki, A. Homeless youth with first-episode psychosis: A 2-year outcome study. Schizophr. Res. 2019, 216, 460–469. [Google Scholar] [CrossRef]
- Jääskeläinen, E.; Juola, P.; Hirvonen, N.; McGrath, J.J.; Saha, S.; Isohanni, M.; Veijola, J.; Miettunen, J. A Systematic Review and Meta-Analysis of Recovery in Schizophrenia. Schizophr. Bull. 2012, 39, 1296–1306. [Google Scholar] [CrossRef]
- Kotov, R.; Fochtmann, L.; Li, K.; Tanenberg-Karant, M.; Constantino, E.A.; Rubinstein, J.; Perlman, G.; Velthorst, E.; Fett, A.-K.J.; Carlson, G.; et al. One hundred years of schizophrenia: a meta-analysis of the outcome literature. Am. J. Psychiatry 1994, 151, 1409–1416. [Google Scholar] [CrossRef]
- Vita, A.; De Peri, L.; Deste, G.; Sacchetti, E. Progressive loss of cortical gray matter in schizophrenia: a meta-analysis and meta-regression of longitudinal MRI studies. Transl Psychiatry 2012, 2, e190. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, J.A. Neurobiology and the natural history of schizophrenia. J. Clin. Psychiatry 2006, 67, e14. [Google Scholar] [CrossRef] [PubMed]
- Fusar-Poli, P.; Smieskova, R.; Kempton, M.; Ho, B.; Andreasen, N.; Borgwardt, S. Progressive brain changes in schizophrenia related to antipsychotic treatment? A meta-analysis of longitudinal MRI studies. Neurosci. Biobehav. Rev. 2013, 37, 1680–1691. [Google Scholar] [CrossRef] [PubMed]
- Ho, B.C.; Andreasen, N.C.; Ziebell, S.; Pierson, R.; Magnotta, V. Long-term antipsychotic treatment and brain volumes: A longitudinal study of first-episode schizophrenia. Arch. Gen. Psychiatry 2011, 68, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Cahn, W.; Pol HE, H.; Lems, E.B.; van Haren, N.E.; Schnack, H.G.; van der Linden, J.A.; Schothorst, P.F.; van Engeland, H.; Kahn, R.S. Brain volume changes in first-episode schizophrenia: A 1-year follow-up study. Arch. Gen. Psychiatry 2002, 59, 1002–1010. [Google Scholar] [CrossRef]
- Ting, C.; Rajji, T.K.; Ismail, Z.; Tang-Wai, D.F.; Apanasiewicz, N.; Miranda, D.; Mamo, D.; Mulsant, B.H. Differentiating the Cognitive Profile of Schizophrenia from That of Alzheimer Disease and Depression in Late Life. PLOS ONE 2010, 5, e10151. [Google Scholar] [CrossRef]
- Dehmelt, L.; Halpain, S. The MAP2/Tau family of microtubule-associated proteins. Genome Biol. 2004, 6, 204. [Google Scholar] [CrossRef]
- Hameroff, S. Consciousness, Cognition and the Neuronal Cytoskeleton – A New Paradigm Needed in Neuroscience. Front. Mol. Neurosci. 2022, 15, 869935. [Google Scholar] [CrossRef]
- Tonello, L.; Cocchi, M.; Gabrielli, F.; Tuszynski, J.A. On the possible quantum role of serotonin in consciousness. J. Integr. Neurosci. 2015, 14, 295–308. [Google Scholar] [CrossRef]
- Craddock, T.J.A.; Priel, A.; Tuszynski, J.A. Keeping time: Could quantum beating in microtubules be the basis for the neural synchrony related to consciousness? J. Integr. Neurosci. 2014, 13, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Planel, E.; Krishnamurthy, P.; Miyasaka, T.; Liu, L.; Herman, M.; Kumar, A.; Bretteville, A.; Figueroa, H.Y.; Yu, W.H.; Whittington, R.A.; et al. Anesthesia-Induced Hyperphosphorylation Detaches 3-Repeat Tau from Microtubules without Affecting Their StabilityIn Vivo. J. Neurosci. 2008, 28, 12798–12807. [Google Scholar] [CrossRef] [PubMed]
- Run, X.; Liang, Z.; Zhang, L.; Iqbal, K.; Grundke-Iqbal, I.; Gong, C.-X. Anesthesia Induces Phosphorylation of Tau. J. Alzheimer's Dis. 2009, 16, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Rowinsky, E.K.; Chaudhry, V.; Cornblath, D.R.; Donehower, R.C. Neurotoxicity of Taxol. J Natl Cancer Inst Monogr. 1993, 107–115. [Google Scholar] [PubMed]
- Luciani, M.; Montalbano, M.; Troncone, L.; Bacchin, C.; Uchida, K.; Daniele, G.; Wolf, B.J.; Butler, H.M.; Kiel, J.; Berto, S.; et al. Big tau aggregation disrupts microtubule tyrosination and causes myocardial diastolic dysfunction: from discovery to therapy. Eur. Hear. J. 2023, 44, 1560–1570. [Google Scholar] [CrossRef]
- Bunzel, B.; Schmidl-Mohl, B.; Wollenek, G.; Grundböck, A. Does changing the heart mean changing personality? A retrospective inquiry on 47 heart transplant patients. Qual. Life Res. 1992, 1, 251–256. [Google Scholar] [CrossRef]
- Pearsall, P.; Schwartz, G.E.; Russek, L.G. Changes in heart transplant recipients that parallel the personalities of their donors. Integr. Med. 2000, 2, 65–72. [Google Scholar] [CrossRef]
- Prinzen, F.W.; Vernooy, K.; Cornelussen, R.N. Cardiac Memory and Cortical Memory. Circulation 2004, 109, e226. [Google Scholar] [CrossRef]
- Liester, M.B. Personality changes following heart transplantation: The role of cellular memory. Med Hypotheses 2019, 135, 109468. [Google Scholar] [CrossRef]
- Bahramnezhad, F.; Asgari, P.; Jackson, A.C. Adjustment to a new heart: Concept analysis using a hybrid model. Iran. J. Nurs. Midwifery Res. 2021, 26, 89–96. [Google Scholar] [CrossRef]
- Letourneau, J.; Holmes, Z.C.; Dallow, E.P.; Durand, H.K.; Jiang, S.; Carrion, V.M.; Gupta, S.K.; Mincey, A.C.; Muehlbauer, M.J.; Bain, J.R.; et al. Ecological memory of prior nutrient exposure in the human gut microbiome. ISME J. 2022, 16, 2479–2490. [Google Scholar] [CrossRef] [PubMed]
- Snijders, T.; Aussieker, T.; Holwerda, A.; Parise, G.; Loon, L.J.C.; Verdijk, L.B. The concept of skeletal muscle memory: Evidence from animal and human studies. Acta Physiol. 2020, 229, e13465. [Google Scholar] [CrossRef] [PubMed]
- Vogel, D.; Dussutour, A. Direct transfer of learned behaviour via cell fusion in non-neural organisms. Proc. R. Soc. B: Boil. Sci. 2016, 283, 20162382. [Google Scholar] [CrossRef] [PubMed]
- Tetz, G.; Pinho, M.; Pritzkow, S.; Mendez, N.; Soto, C.; Tetz, V. Bacterial DNA promotes Tau aggregation. Sci. Rep. 2020, 10, 2369. [Google Scholar] [CrossRef] [PubMed]
- Seneff, S.; Kyriakopoulos, A.M.; Nigh, G.; A McCullough, P.; McCullough, P.A. A Potential Role of the Spike Protein in Neurodegenerative Diseases: A Narrative Review. Cureus 2023, 15, e34872. [Google Scholar] [CrossRef]
- Demirel. F.; Cetin, I.; Turan,.; Yıldız, N.; Sağlam, T.; Duran, A. Total Tau and Phosphorylated Tau Protein Serum Levels in Patients with Schizophrenia Compared with Controls. Psychiatr. Q. 2017, 88, 921–928. [Google Scholar] [CrossRef]
- Grubisha, M.J.; Sun, X.; MacDonald, M.L.; Garver, M.; Sun, Z.; Paris, K.A.; Patel, D.S.; DeGiosio, R.A.; Lewis, D.A.; Yates, N.A.; et al. MAP2 is differentially phosphorylated in schizophrenia, altering its function. Mol. Psychiatry 2021, 26, 5371–5388. [Google Scholar] [CrossRef]
- Krause, M.; Theiss, C.; Brüne, M. Ultrastructural Alterations of Von Economo Neurons in the Anterior Cingulate Cortex in Schizophrenia. Anat. Rec. 2017, 300, 2017–2024. [Google Scholar] [CrossRef]
- Agnati, L.F.; Zunarelli, E.; Genedani, S.; Fuxe, K. On the Existence of a Global Molecular Network Enmeshing the Whole Central Nervous System: Physiological and Pathological Implications. Curr. Protein Pept. Sci. 2006, 7, 3–15. [Google Scholar] [CrossRef]
- Boeve, B.F. Behavioral Variant Frontotemporal Dementia. Contin. Lifelong Learn. Neurol. 2022, 28, 702–725. [Google Scholar] [CrossRef]
- Seeley, W.W. Behavioral Variant Frontotemporal Dementia. Contin. Lifelong Learn. Neurol. 2019, 25, 76–100. [Google Scholar] [CrossRef] [PubMed]
- Dols, A.; van Liempt, S.; Gossink, F.; Krudop, W.A.; Sikkes, S.; Pijnenburg, Y.A.L.; Stek, M.L. Identifying Specific Clinical Symptoms of Behavioral Variant Frontotemporal Dementia Versus Differential Psychiatric Disorders in Patients Presenting with a Late-Onset Frontal Lobe Syndrome. J. Clin. Psychiatry 2016, 77, 1391–1395. [Google Scholar] [CrossRef] [PubMed]
- Cipriani, G.; Danti, S.; Nuti, A.; Di Fiorino, M.; Cammisuli, D.M. Is that schizophrenia or frontotemporal dementia? Supporting clinicians in making the right diagnosis. Acta Neurol. Belg. 2020, 120, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Bott, N.T.; Radke, A.; Stephens, M.L.; Kramer, J.H. Frontotemporal dementia: diagnosis, deficits and management. Neurodegener. Dis. Manag. 2014, 4, 439–454. [Google Scholar] [CrossRef] [PubMed]
- Ducharme, S.; Dols, A.; Laforce, R.; Devenney, E.; Kumfor, F.; Stock, J.v.D.; Dallaire-Théroux, C.; Seelaar, H.; Gossink, F.; Vijverberg, E.; et al. Recommendations to distinguish behavioural variant frontotemporal dementia from psychiatric disorders. Brain 2020, 143, 1632–1650. [Google Scholar] [CrossRef] [PubMed]
- Woolley, J.D.; Khan, B.K.; Murthy, N.K.; Miller, B.L.; Rankin, K.P. The diagnostic challenge of psychiatric symptoms in neurodegenerative disease: rates of and risk factors for prior psychiatric diagnosis in patients with early neurodegenerative disease. J Clin Psychiatry 2011, 72, 126–133. [Google Scholar] [CrossRef]
- Liljegren, M.; Naasan, G.; Temlett, J.; Perry, D.C.; Rankin, K.P.; Merrilees, J.; Grinberg, L.T.; Seeley, W.W.; Englund, E.; Miller, B.L. Criminal Behavior in Frontotemporal Dementia and Alzheimer Disease. JAMA Neurol. 2015, 72, 295–300. [Google Scholar] [CrossRef]
- Sfera, A.; Osorio, C.; Gradini, R.; Price, A. Neurodegeneration Behind Bars: from Molecules to Jurisprudence. Front. Psychiatry 2014, 5, 115. [Google Scholar] [CrossRef]
- Meeks, T.W.; Jeste, D.V. Beyond the Black Box: What is The Role for Antipsychotics in Dementia? Curr. Psychiatr. 2008, 7, 50–65. [Google Scholar]
- Desai, V.C.; Heaton, P.C.; Kelton, C.M. Impact of the Food and Drug Administration's antipsychotic black box warning on psychotropic drug prescribing in elderly patients with dementia in outpatient and office-based settings. Alzheimer's Dement. 2012, 8, 453–457. [Google Scholar] [CrossRef]
- Deming, P.; Cook, C.J.; Meyerand, M.E.; Kiehl, K.A.; Kosson, D.S.; Koenigs, M. Impaired salience network switching in psychopathy. Behav. Brain Res. 2023, 452, 114570. [Google Scholar] [CrossRef] [PubMed]
- Kohn, N.; Szopinska-Tokov, J.; Arenas, A.L.; Beckmann, C.; Arias-Vasquez, A.; Aarts, E. Multivariate associative patterns between the gut microbiota and large-scale brain network connectivity. Gut Microbes 2021, 13, 2006586. [Google Scholar] [CrossRef] [PubMed]
- Mulder, D.; Aarts, E.; Vasquez, A.A.; Bloemendaal, M. A systematic review exploring the association between the human gut microbiota and brain connectivity in health and disease. Mol. Psychiatry 2023. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, A.P.; Sanchez-Padilla, D.E.; Drew, J.C.; Oli, M.W.; Roesch, L.F.W.; Triplett, E.W. Saliva microbiome, dietary, and genetic markers are associated with suicidal ideation in university students. Sci. Rep. 2022, 12, 14306. [Google Scholar] [CrossRef] [PubMed]
- Koren, T.; Yifa, R.; Amer, M.; Krot, M.; Boshnak, N.; Ben-Shaanan, T.L.; Azulay-Debby, H.; Zalayat, I.; Avishai, E.; Hajjo, H.; et al. Insular cortex neurons encode and retrieve specific immune responses. Cell 2021, 184, 5902–5915. [Google Scholar] [CrossRef]
- Rolls, A. Immunoception: the insular cortex perspective. Cell. Mol. Immunol. 2023. [Google Scholar] [CrossRef]
- Bodea, L.; Eckert, A.; Ittner, L.M.; Piguet, O.; Götz, J. Tau physiology and pathomechanisms in frontotemporal lobar degeneration. J. Neurochem. 2016, 138, 71–94. [Google Scholar] [CrossRef]
- Rademakers, R.; Cruts, M.; van Broeckhoven, C. The role of tau (MAPT) in frontotemporal dementia and related tauopathies. Hum. Mutat. 2004, 24, 277–295. [Google Scholar] [CrossRef]
- Marchisella, F.; Coffey, E.T.; Hollos, P. Microtubule and microtubule associated protein anomalies in psychiatric disease. Cytoskeleton 2016, 73, 596–611. [Google Scholar] [CrossRef]
- Arnold, S.E.; Lee, V.M.; Gur, R.E.; Trojanowski, J.Q. Abnormal expression of two microtubule-associated proteins (MAP2 and MAP5) in specific subfields of the hippocampal formation in schizophrenia. Proc. Natl. Acad. Sci. USA 1991, 88, 10850–10854. [Google Scholar] [CrossRef]
- Jones, L.B.; Johnson, N.; Byne, W. Alterations in MAP2 immunocytochemistry in areas 9 and 32 of schizophrenic prefrontal cortex. Psychiatry Res. Neuroimaging 2002, 114, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Andreou, D.; Jørgensen, K.N.; Nerland, S.; Smelror, R.E.; Wedervang-Resell, K.; Johannessen, C.H.; Myhre, A.M.; Andreassen, O.A.; Blennow, K.; Zetterberg, H.; et al. Lower plasma total tau in adolescent psychosis: Involvement of the orbitofrontal cortex. J. Psychiatr. Res. 2021, 144, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Shukla, R.; Santoro, J.; Bender, F.C.; Laterza, O.F. Quantitative determination of human interleukin 22 (IL-22) in serum using Singulex-Erenna® Technology. J. Immunol. Methods 2013, 390, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Eichenberger, E.M.; Degner, N.; Scott, E.R.; Ruffin, F.; Franzone, J.; Sharma-Kuinkel, B.; Shah, P.; Hong, D.; Dalai, S.C.; Blair, L.; et al. Microbial Cell-Free DNA Identifies the Causative Pathogen in Infective Endocarditis and Remains Detectable Longer Than Conventional Blood Culture in Patients with Prior Antibiotic Therapy. Clin. Infect. Dis. 2022, 76, e1492–e1500. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-W.; Stewart, R.; Park, W.-Y.; Jhon, M.; Lee, J.-Y.; Kim, S.-Y.; Kim, J.-M.; Amminger, P.; Chung, Y.-C.; Yoon, J.-S. Latent Iron Deficiency as a Marker of Negative Symptoms in Patients with First-Episode Schizophrenia Spectrum Disorder. Nutrients 2018, 10, 1707. [Google Scholar] [CrossRef] [PubMed]
- Wan, W.; Cao, L.; Kalionis, B.; Murthi, P.; Xia, S.; Guan, Y. Iron Deposition Leads to Hyperphosphorylation of Tau and Disruption of Insulin Signaling. Front. Neurol. 2019, 10, 607. [Google Scholar] [CrossRef]
- Rao, S.S.; Adlard, P.A. Untangling Tau and Iron: Exploring the Interaction Between Iron and Tau in Neurodegeneration. Front. Mol. Neurosci. 2018, 11, 276. [Google Scholar] [CrossRef]
- Sheelakumari, R.; Kesavadas, C.; Varghese, T.; Sreedharan, R.; Thomas, B.; Verghese, J.; Mathuranath, P. Assessment of Iron Deposition in the Brain in Frontotemporal Dementia and Its Correlation with Behavioral Traits. Am. J. Neuroradiol. 2017, 38, 1953–1958. [Google Scholar] [CrossRef]
- Tisdall, M.D.; Ohm, D.T.; Lobrovich, R.; Das, S.R.; Mizsei, G.; Prabhakaran, K.; Ittyerah, R.; Lim, S.; McMillan, C.T.; Wolk, D.A.; et al. Ex vivo MRI and histopathology detect novel iron-rich cortical inflammation in frontotemporal lobar degeneration with tau versus TDP-43 pathology. NeuroImage: Clin. 2022, 33, 102913. [Google Scholar] [CrossRef]
- Kosyakovsky, J.; Fine, J.M.; Frey, W.H.; Hanson, L.R. Mechanisms of Intranasal Deferoxamine in Neurodegenerative and Neurovascular Disease. Pharmaceuticals 2021, 14, 95. [Google Scholar] [CrossRef]
- Aaronson, A.L.; Bordelon, S.D.; Brakel, S.J.; Morrison, H. A Review of the Role of Chronic Traumatic Encephalopathy in Criminal Court. J. Am. Acad. Psychiatry Law 2020, 49, 60–65. [Google Scholar] [PubMed]
- Byard, R.; Tiemensma, M.; Buckland, M.E.; Vink, R. Chronic traumatic encephalopathy (CTE)—features and forensic considerations. Forensic Sci. Med. Pathol. 2023. [Google Scholar] [CrossRef] [PubMed]
- McKee, A.C.; Stein, T.D.; Kiernan, P.T.; Alvarez, V.E. The Neuropathology of Chronic Traumatic Encephalopathy. Brain Pathol. 2015, 25, 350–364. [Google Scholar] [CrossRef] [PubMed]
- Katsumoto, A.; Takeuchi, H.; Tanaka, F. Tau Pathology in Chronic Traumatic Encephalopathy and Alzheimer's Disease: Similarities and Differences. Front. Neurol. 2019, 10, 980. [Google Scholar] [CrossRef]
- Daglas, M.; Adlard, P.A. The Involvement of Iron in Traumatic Brain Injury and Neurodegenerative Disease. Front. Neurosci. 2018, 12, 981. [Google Scholar] [CrossRef] [PubMed]
- Han, X. Neurolipidomics: challenges and developments. Front. Biosci. 2007, 12, 2601–2615. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Seo, Y.; Jo, Y.S.; Lee, S.; Cho, E.; Cazenave-Gassiot, A.; Shin, Y.-S.; Moon, M.H.; An, H.J.; Wenk, M.R.; et al. Brain lipidomics: From functional landscape to clinical significance. Sci. Adv. 2022, 8, eadc9317. [Google Scholar] [CrossRef]
- Nicolson, G.L.; Ash, M.E. Lipid Replacement Therapy: A natural medicine approach to replacing damaged lipids in cellular membranes and organelles and restoring function. Biochim. et Biophys. Acta (BBA) - Biomembr. 2014, 1838, 1657–1679. [Google Scholar] [CrossRef] [PubMed]
- Hamsanathan, S.; Gurkar, A.U. Lipids as Regulators of Cellular Senescence. Front. Physiol. 2022, 13, 796850. [Google Scholar] [CrossRef] [PubMed]
- Horn, A.; Jaiswal, J.K. Structural and signaling role of lipids in plasma membrane repair. Curr. Top. Membr. 2019, 84, 67–98. [Google Scholar] [CrossRef]
- Cadenas, C.; Vosbeck, S.; Hein, E.-M.; Hellwig, B.; Langer, A.; Hayen, H.; Franckenstein, D.; Büttner, B.; Hammad, S.; Marchan, R.; et al. Glycerophospholipid profile in oncogene-induced senescence. Biochim. et Biophys. Acta (BBA) - Mol. Cell Biol. Lipids 2012, 1821, 1256–1268. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, H.-S.; Choi, D.-H.; Choi, J.; Cho, S.Y.; Kim, S.-H.; Baek, H.-S.; Yoon, K.D.; Son, S.W.; Son, E.D.; et al. Kaempferol tetrasaccharides restore skin atrophy via PDK1 inhibition in human skin cells and tissues: Bench and clinical studies. BioMedicine 2022, 156, 113864. [Google Scholar] [CrossRef] [PubMed]
- Emamian, E.S. AKT/GSK3 signaling pathway and schizophrenia. Front. Mol. Neurosci. 2012, 5, 33. [Google Scholar] [CrossRef] [PubMed]
- Lucas, N.; Cho, W. Phosphatidylserine Binding Is Essential for Plasma Membrane Recruitment and Signaling Function of 3-Phosphoinositide-dependent Kinase-1. J. Biol. Chem. 2011, 286, 41265–41272. [Google Scholar] [CrossRef] [PubMed]
- Noll, R. Kraepelin's `lost biological psychiatry'? Autointoxication, organotherapy and surgery for dementia praecox. Hist. Psychiatry 2007, 18, 301–320. [Google Scholar] [CrossRef] [PubMed]
- Horrobin, D.F. The membrane phospholipid hypothesis as a biochemical basis for the neurodevelopmental concept of schizophrenia. Schizophr. Res. 1998, 30, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.J.; Nazli, A.; Rojas, O.L.; Chege, D.; Alidina, Z.; Huibner, S.; Mujib, S.; Benko, E.; Kovacs, C.; Shin, L.Y.Y.; et al. A role for mucosal IL-22 production and Th22 cells in HIV-associated mucosal immunopathogenesis. Mucosal Immunol. 2012, 5, 670–680. [Google Scholar] [CrossRef]
- Tsounis, E.P.; Triantos, C.; Konstantakis, C.; Marangos, M.; Assimakopoulos, S.F. Intestinal barrier dysfunction as a key driver of severe COVID-19. World J. Virol. 2023, 12, 68–90. [Google Scholar] [CrossRef]
- Fang, S.; Ju, D.; Lin, Y.; Chen, W. The role of interleukin-22 in lung health and its therapeutic potential for COVID-19. Front. Immunol. 2022, 13, 951107. [Google Scholar] [CrossRef]
- Coronas, V.; Arnault, P.; Jégou, J.-F.; Cousin, L.; Rabeony, H.; Clarhaut, S.; Harnois, T.; Lecron, J.-C.; Morel, F. IL-22 Promotes Neural Stem Cell Self-Renewal in the Adult Brain. STEM CELLS 2023, 41, 252–259. [Google Scholar] [CrossRef]
- Rothhammer, V.; Quintana, F.J. The aryl hydrocarbon receptor: an environmental sensor integrating immune responses in health and disease. Nat. Rev. Immunol. 2019, 19, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Jin, U.-H.; Karki, K.; Jayaraman, A.; Allred, C.; Michelhaugh, S.K.; Mittal, S.; Chapkin, R.S.; Safe, S. Dopamine is an aryl hydrocarbon receptor agonist. Biochem. J. 2020, 477, 3899–3910. [Google Scholar] [CrossRef] [PubMed]
- Fehsel, K.; Schwanke, K.; Kappel, B.; Fahimi, E.; Meisenzahl-Lechner, E.; Esser, C.; Hemmrich, K.; Haarmann-Stemmann, T.; Kojda, G.; Lange-Asschenfeldt, C. Activation of the aryl hydrocarbon receptor by clozapine induces preadipocyte differentiation and contributes to endothelial dysfunction. J. Psychopharmacol. 2022, 36, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Ishima, T.; Iyo, M.; Hashimoto, K. Neurite outgrowth mediated by the heat shock protein Hsp90α: a novel target for the antipsychotic drug aripiprazole. Transl. Psychiatry 2012, 2, e170. [Google Scholar] [CrossRef] [PubMed]
- Sun, L. Recent advances in the development of AHR antagonists in immuno-oncology. RSC Med. Chem. 2021, 12, 902–914. [Google Scholar] [CrossRef] [PubMed]
- De Juan, A.; Segura, E. Modulation of Immune Responses by Nutritional Ligands of Aryl Hydrocarbon Receptor. Front. Immunol. 2021, 12, 645168. [Google Scholar] [CrossRef]
- Salminen, A. Aryl hydrocarbon receptor (AhR) reveals evidence of antagonistic pleiotropy in the regulation of the aging process. Cell. Mol. Life Sci. 2022, 79, 489. [Google Scholar] [CrossRef]
- Ojo, E.S.; Tischkau, S.A. The Role of AhR in the Hallmarks of Brain Aging: Friend and Foe. Cells 2021, 10, 2729. [Google Scholar] [CrossRef]
- Eckers, A.; Jakob, S.; Heiss, C.; Haarmann-Stemmann, T.; Goy, C.; Brinkmann, V.; Cortese-Krott, M.M.; Sansone, R.; Esser, C.; Ale-Agha, N.; et al. The aryl hydrocarbon receptor promotes aging phenotypes across species. Sci. Rep. 2016, 6, 19618–19618. [Google Scholar] [CrossRef]
Figure 1.
Neuronal membranes are comprised of a lipid bilayer which anchors numerous neurotransmitter receptors. Antipsychotic drugs and excessive iron trigger peroxidation of the lipid bilayer, altering membrane permeability. Intracellular iron activates the downstream SCZ-related phosphatase, including Akt and PI3K contributing to the pathogenesis of this disease. Psychotropic drugs, especially those belonging to the thiazine group, intercalate themselves into the lipid bilayer, changing the biophysical properties of plasma membranes which in turn, disrupts neurotransmission.
Figure 1.
Neuronal membranes are comprised of a lipid bilayer which anchors numerous neurotransmitter receptors. Antipsychotic drugs and excessive iron trigger peroxidation of the lipid bilayer, altering membrane permeability. Intracellular iron activates the downstream SCZ-related phosphatase, including Akt and PI3K contributing to the pathogenesis of this disease. Psychotropic drugs, especially those belonging to the thiazine group, intercalate themselves into the lipid bilayer, changing the biophysical properties of plasma membranes which in turn, disrupts neurotransmission.
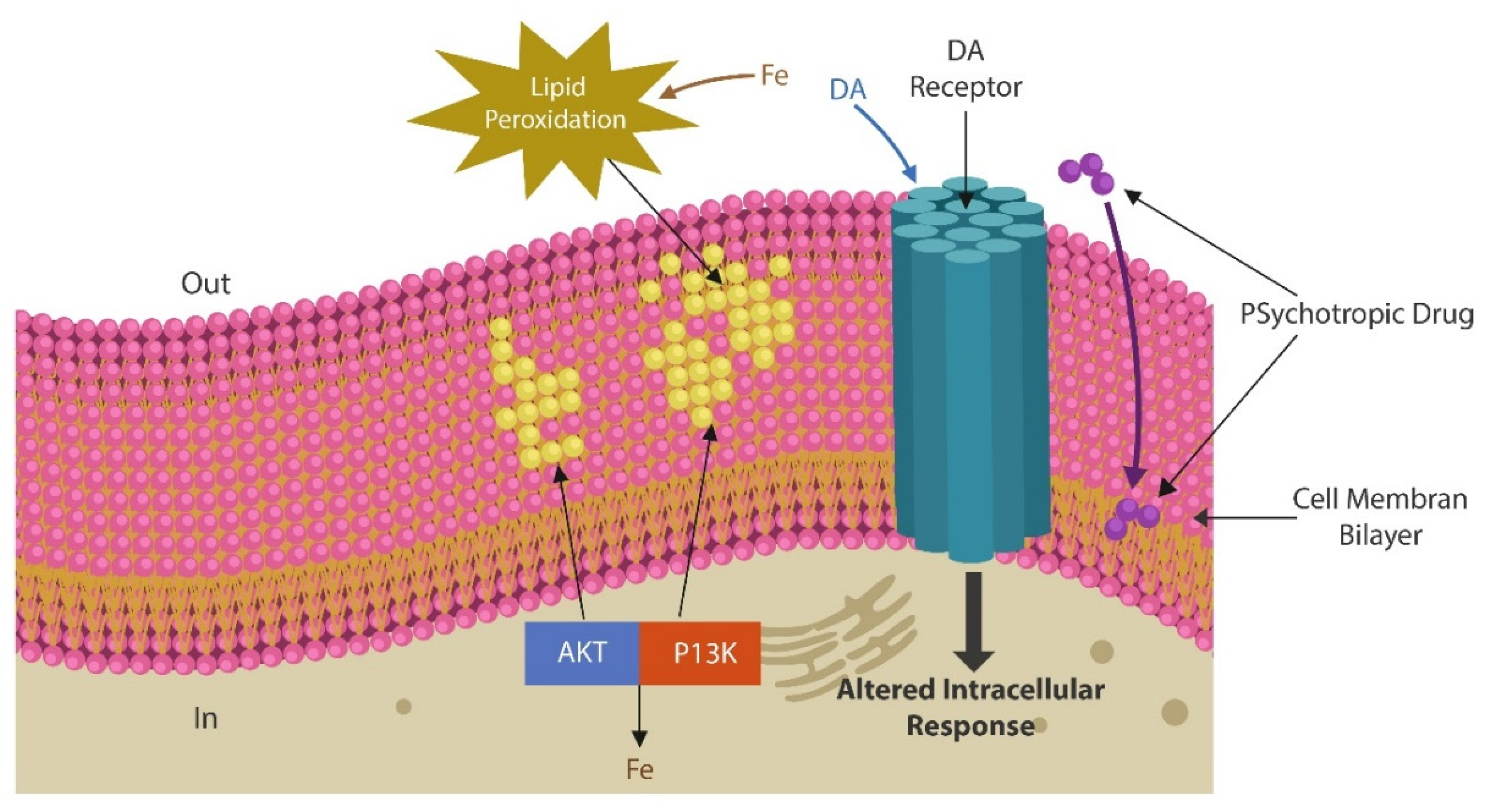
Figure 2.
Natural course of SCZ starts in early childhood with a premorbid phase with no symptoms or very mild signs. Prodromal phase is marked by mild symptoms, such as social isolation, anxiety, and insomnia. This phase blends gradually into the psychotic phase during which patients are usually hospitalized numerous times for exhibiting positive symptoms. Around midlife the positive symptoms gradually subside and are replaced by disabling negative, and cognitive manifestations. (Table adapted from ref. 105).
Figure 2.
Natural course of SCZ starts in early childhood with a premorbid phase with no symptoms or very mild signs. Prodromal phase is marked by mild symptoms, such as social isolation, anxiety, and insomnia. This phase blends gradually into the psychotic phase during which patients are usually hospitalized numerous times for exhibiting positive symptoms. Around midlife the positive symptoms gradually subside and are replaced by disabling negative, and cognitive manifestations. (Table adapted from ref. 105).
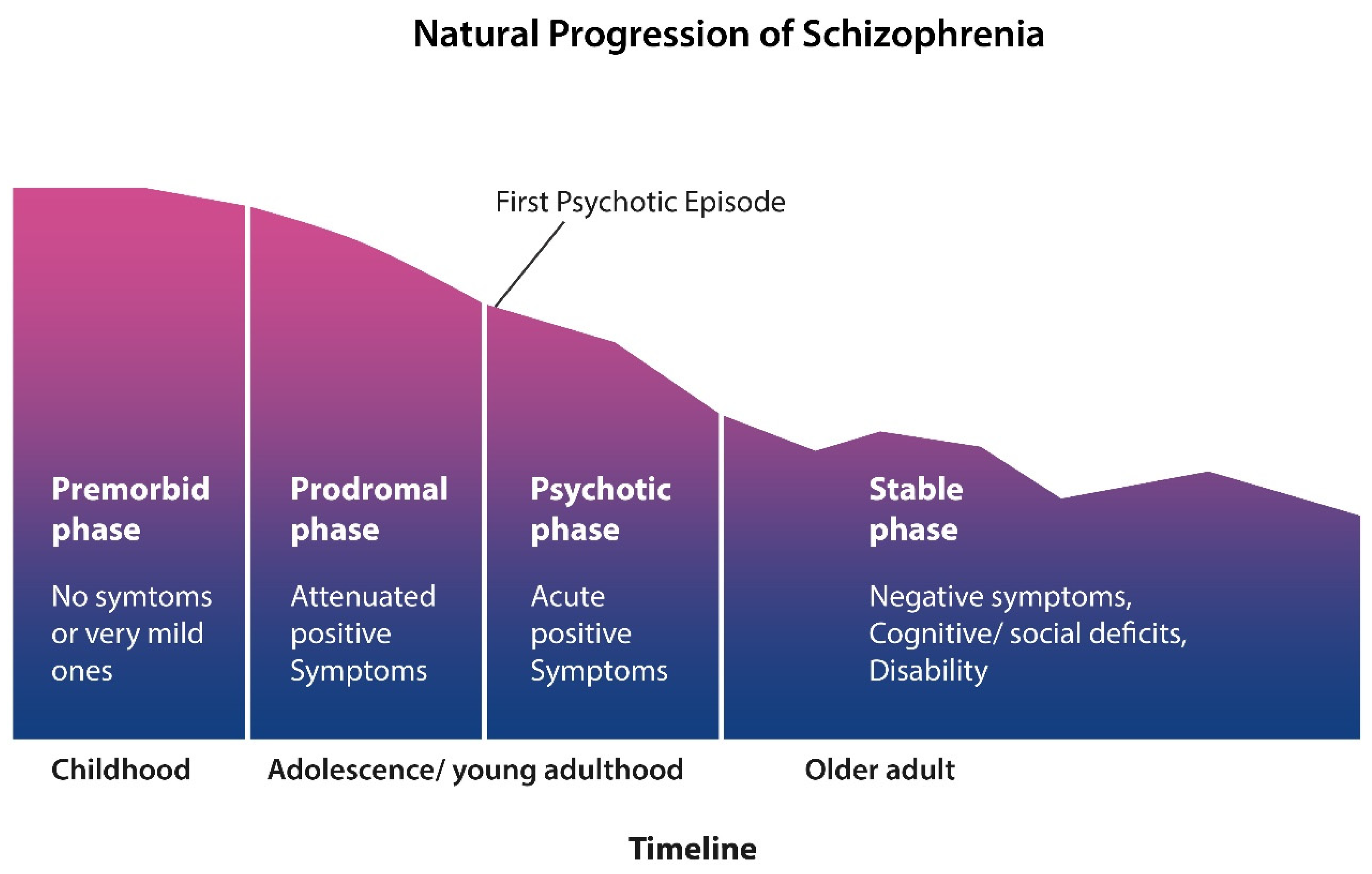
Figure 3.
MTs are stabilized by MAP2 or Tau protein. Excessive Tau phosphorylation detaches this protein, destabilizing the MTs. Hyperphosphorylated Tau can aggregate, forming neurofibrillary tangles (NFTs), which accumulate, triggering neuronal death. Both iron and mcfDNA promote Tau aggregation, while pathogens, including COVID disseminate pathological Tau to neighboring healthy cells, facilitating the pathogenesis of dementia.
Figure 3.
MTs are stabilized by MAP2 or Tau protein. Excessive Tau phosphorylation detaches this protein, destabilizing the MTs. Hyperphosphorylated Tau can aggregate, forming neurofibrillary tangles (NFTs), which accumulate, triggering neuronal death. Both iron and mcfDNA promote Tau aggregation, while pathogens, including COVID disseminate pathological Tau to neighboring healthy cells, facilitating the pathogenesis of dementia.
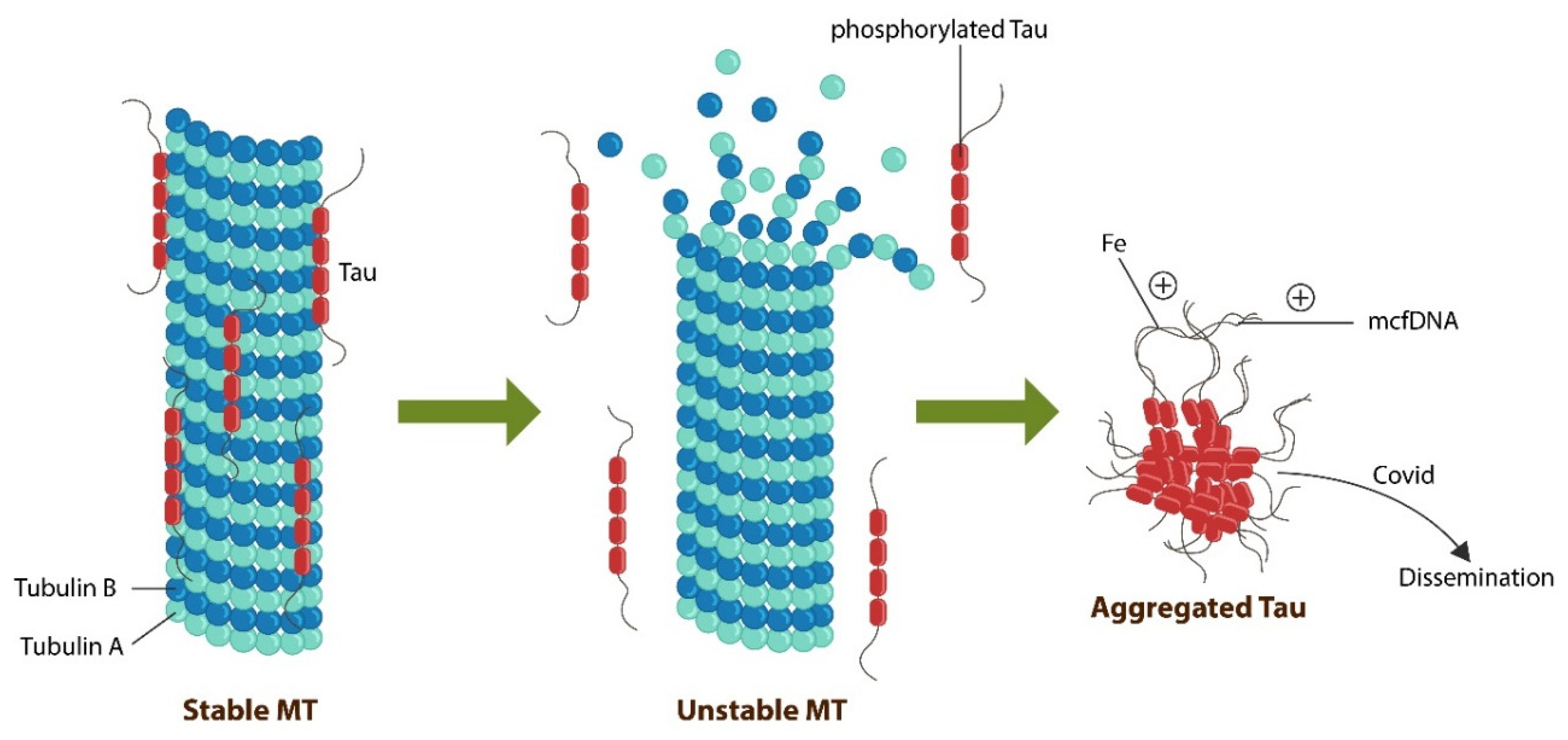
Figure 4.
LRT inhibits PDK1 as exogenous phosphatidylserine (PS) binds the PH domain of PDK1, inactivating the downstream kinases, protein kinase B (Akt), and glycogen synthase kinase 3 beta (GSK-3β). LRT and PDK1 inhibitors act as natural antipsychotic agents The novel PDK1 inhibitor, OSU 03012 readily crosses the BBB, enhancing the antipsychotic properties of LRT.
Figure 4.
LRT inhibits PDK1 as exogenous phosphatidylserine (PS) binds the PH domain of PDK1, inactivating the downstream kinases, protein kinase B (Akt), and glycogen synthase kinase 3 beta (GSK-3β). LRT and PDK1 inhibitors act as natural antipsychotic agents The novel PDK1 inhibitor, OSU 03012 readily crosses the BBB, enhancing the antipsychotic properties of LRT.
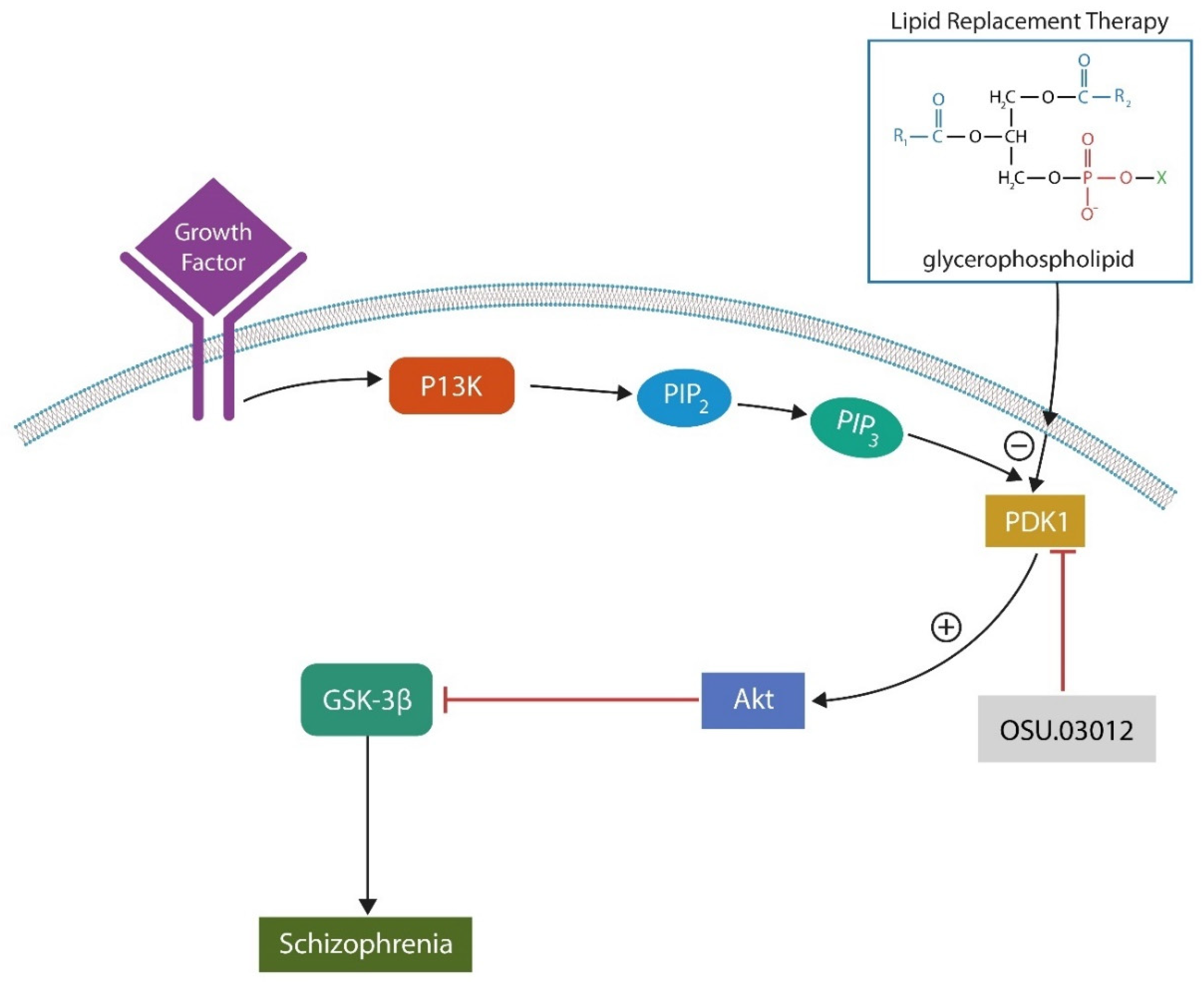
Figure 5.
Endorsed by Emil Kraepelin and other 19th century researchers, microbial hypothesis of. SCZ was forgotten until the arrival of HIV and COVID-19.
Figure 5.
Endorsed by Emil Kraepelin and other 19th century researchers, microbial hypothesis of. SCZ was forgotten until the arrival of HIV and COVID-19.

Figure 6.
AhR, an upstream receptor for several neurotransmitters, including DA, serotonin, melatonin, vitamin D3, as well as clozapine, carbidopa, and microbial phenazines (natural phenothiazines). Aripiprazole enhances HSP90 attachment to AhR, blocking its entry into the nucleus and the activation of SCZ vulnerability genes.
Figure 6.
AhR, an upstream receptor for several neurotransmitters, including DA, serotonin, melatonin, vitamin D3, as well as clozapine, carbidopa, and microbial phenazines (natural phenothiazines). Aripiprazole enhances HSP90 attachment to AhR, blocking its entry into the nucleus and the activation of SCZ vulnerability genes.
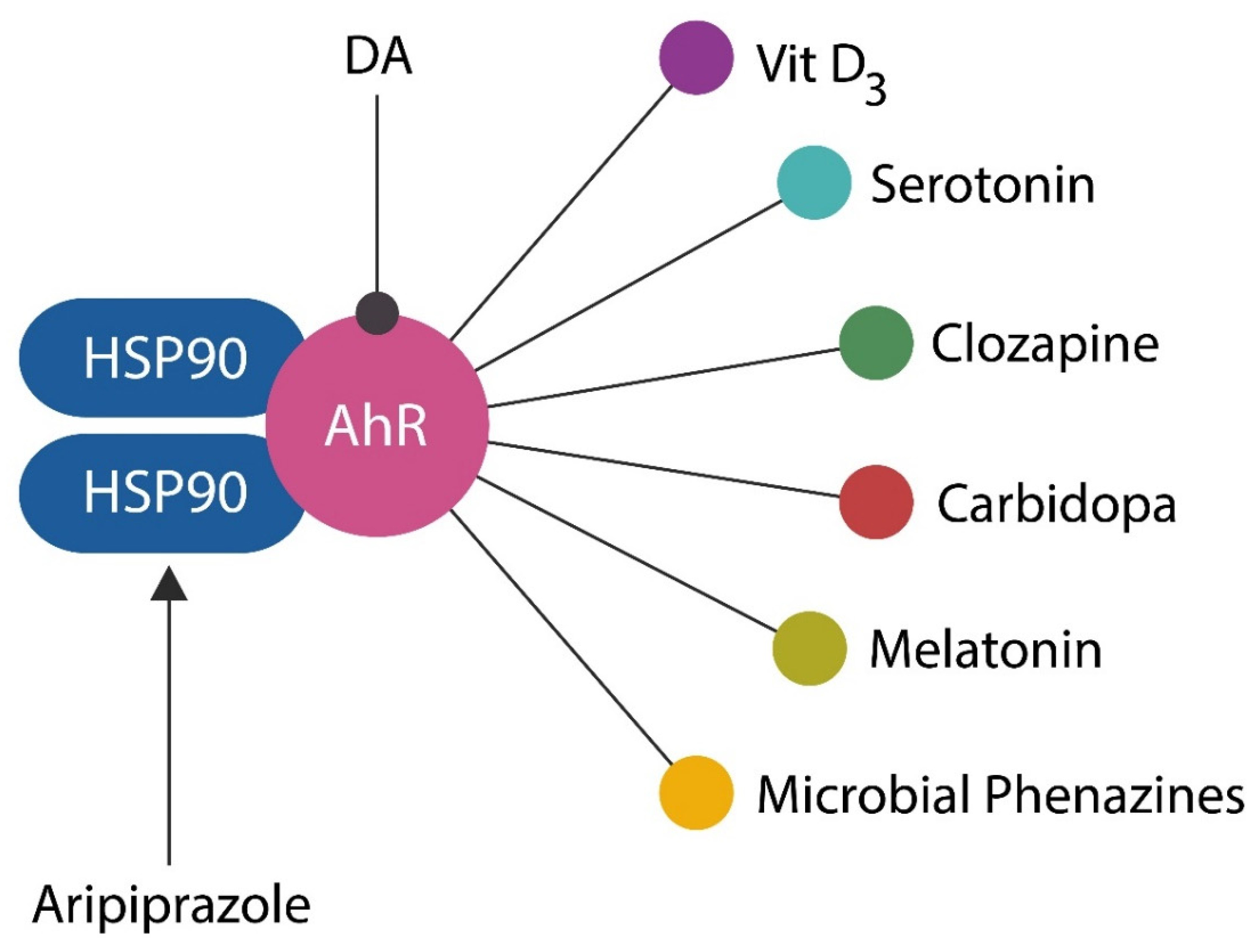

Table 1.
Sustained recovery of SCZ patients during the 20th century, Including employment (Table adapted from Warner R data).
Table 1.
Sustained recovery of SCZ patients during the 20th century, Including employment (Table adapted from Warner R data).
| INTERVAL | SUSTAINED RECOVERY | EMPLOYED |
|---|---|---|
| 1901- 1920 | 20% | 4.7% |
| 1921- 1940 | 12% | 11.9% |
| 1941- 1955 | 23% | 4.1% |
| 1956- 1975 | 20% | 5.1% |
| 1976- 1995 | 20% | 6.9% |
Table 2.
Standardized tests for dysfunctional gut barrier and translocation.
| Marker Type | Marker Assay | References |
|---|---|---|
| Integrity of gut barrier | IL-22 Singulex-Erenna®® | 155 |
| Translocated microbes | mcfDNA Karius Test®® | 156 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



