
Submitted:
04 September 2023
Posted:
05 September 2023
You are already at the latest version
Abstract
Enzymatic hydrolysis of bovine and human hemoglobin yields a diversity of bioactive peptides, some of which are recognized for their antimicrobial properties. Certain small antimicrobial and anticancer peptides target protein synthesis inhibition. This study aims to produce bioactive peptides through enzymatic hemoglobin hydrolysis and evaluate their effects on cell growth inhibition, potentially serving as candidates for biological anticancer drugs. Rapid screening of candidate anticancer peptides derived from enzymatic hydrolysis of bovine and human hemoglobin was conducted using the Lepidium sativum radicle growth technique. The results demonstrate that: 1) the degree of hydrolysis (DH) significantly influences the production of bioactive peptides. DH levels of 3-10% yield considerably stronger radicle growth inhibition compared to DH 0 (native hemoglobin form), exhibiting three to four times greater intensity. 2) Certain peptide fractions from bovine hemoglobin exhibit higher activity than those from human hemoglobin. 3) The structural characteristics of peptides play a crucial role in the observed biological effects. 4) The α137-141 peptide is the most active among the fractions obtained from bovine (IC50 = 29 ± 1 µg/mL) and human hemoglobin (IC50 = 48 ± 2 µg/mL), standing out 10 to 15 times more potent than other hemoglobin fractions. In conclusion, these findings affirm the bioactivity of peptide fractions obtained from bovine and human hemoglobin. The study highlights how enzymatic hydrolysis of bovine and human hemoglobin can yield peptides that inhibit protein synthesis. Ultimately, these results suggest the feasibility of obtaining candidate peptides for biological anticancer drugs from both human and bovine hemoglobin sources. These scientific advances show new hope in the fight against cancer which affects a large number of people around the world.
Keywords:
human hemoglobin
; bovine hemoglobin bovine
; enzymatic hydrolysis
; bioactive peptides
; medicine candidate anticancer
; Screening
1. Introduction
According to the most recent data from the World Health Organization (WHO), cancer is a major cause of mortality, with an estimated 19.3 million new cases and 10 million related deaths in 2020. Forecasts for 2030 also indicate an alarming increase, with almost 26 million new cases and 17 million deaths per year [1]. The term "cancer" is commonly used to describe malignant tumors or neoplasms, which are a group of different conditions affecting various parts of the human body. These tumors result from the rapid multiplication of abnormal cells that escape the normal mechanisms regulating cell growth [2]. Current treatment options for cancer include methods such as immunotherapy, hormone therapy, stem cell transplantation, biomarker testing, and radiotherapy, with surgery and chemotherapy being the most promising options. Chemotherapy is used to disrupt this uncontrolled process of cell division [3,4]. However, many anti-cancer drugs are generally not specific to cancer cells, which means they can also damage healthy cells that are rapidly multiplying, leading to toxic side effects [5,6]. Furthermore, tumor cells can develop resistance to chemotherapy, limiting its effectiveness [7,8]. So it would be advantageous to find other classes of drugs that can specifically target cancer cells without harming normal cells. These drugs should have a lower propensity to cause the development of resistance than conventional chemotherapeutic treatments. Antimicrobial peptides (AMPs) represent a promising and under-exploited alternative. AMPs can specifically target cancer cells while exhibiting reduced toxicity for rapidly proliferating healthy cells. In addition, MPAs have been shown to have a low probability of inducing the development of resistance by their target [9,10,11]. In this respect, hemoglobin, an agri-food protein rich in active peptides, is of particular interest due to its predominance in peptide activity databases [12]. In particular, the enzymatic hydrolysis of bovine hemoglobin generates various peptides with biological activities such as opioid [13,14], hematopoietic [15], or antihypertensive [16]. However, antimicrobial activity is the one most frequently observed and reported in scientific studies [16,17,18,19]. Human hemoglobin, like bovine hemoglobin, is an ideal substrate for proteolysis and the generation of bioactive peptides [20,21,22]. As previously mentioned, antimicrobial peptides, in addition to their role in defending against infection, have been shown to have potent toxicity towards cancer cells [2,11]. This study aimed to produce bioactive peptides from the enzymatic hydrolysis of bovine and human hemoglobin, to assess their anti-cancer potential. The target of this study was the α137-141 peptide, known as neokyotorphin (NKT), a peptide derived from bovine and human hemoglobin, whose antimicrobial and antioxidant effects characterize it as a potential candidate for incorporation into anticancer agents [19,22]. The main aim of this study was to analyze the anticancer activity of bovine and human hemoglobin hydrolysates and their peptide fractions, using two different approaches: The first approach consists of using the screening process for drug candidates that inhibit protein synthesis and can be used as antibacterial and anticancer agents. This method, developed by the TBC laboratories, uses inhibition of the growth of the rootlets of watercress (Lepidium sativum) to rapidly screen drug candidates and anticancer, antibacterial and antiparasitic drugs used or not used clinically [23,24]. In our second approach to studying the in vitro anti-cancer activity of peptidic hydrolysates of human and bovine hemoglobin, we initiated a study specifically targeting the eL42 protein. This protein is of major interest because it is over-expressed in all cancer cells, making it a particularly attractive potential target for anti-cancer molecules. Recent research has highlighted its role in the catalytic activity of the elongation step of translation, suggesting its involvement in cancer cell proliferation [25,26]. During translation in eukaryotes, the ribosomal protein eL42 binds to the CCA arm of tRNA at the 80S ribosome [27]. With this in mind, we undertook a crucial preliminary step by carrying out an experiment to label formylase (formyl-methionyl-tRNA transformylase - FMTS) with tRNAox. This approach was chosen because of the similarities between the interaction of formylase and the eL42 protein with tRNA oxidized by its CCA arm, independently of their state (free or bound to the ribosome) [28]. The aim was to determine whether potential competition would occur, resulting in the inability of the tRNA substrate to bind to the ribosomal protein in the presence of chemotherapeutic molecules. This step enabled us to gain a better understanding of how bioactive peptides could potentially interfere with tRNA and formylase, opening up new perspectives in our search for targeted anti-cancer molecules.
Finally, a comparative analysis of the peptide populations in the peptide hydrolysate fractions was carried out by UPLC-MS/MS.
2. Results
2.1. Effects of bovine and human hemoglobin hydrolysates and their peptide fractions on the inhibition of protein biosynthesis.
2.1.1. Study of the influence of initial Bovine and human hemoglobin concentration on Protein biosynthesis
Figure 3A,B show the results of the effect of the initial concentration of bovine and human hemoglobin (following a hydrolysis period of 3 hours, corresponding to a degree of hydrolysis (DH)=10%) on the inhibition of LS rootlet growth. These results are expressed in concentration-response curves. In this experimental model, it was observed that the initial hemoglobin concentration, whether bovine or human, did not influence the inhibition of LS rootlet growth. An analysis of the IC50s for bovine hemoglobin showed broadly equivalent values, indicating that varying the initial concentration (1%, 2%, 8%, or 10%) did not make a significant difference in its impact on inhibition of LS rootlet growth. Similarly, increasing the initial concentration of human hemoglobin did not appear to influence the inhibition of LS rootlet growth, with very similar IC50s. A comparison of the IC50 of the two species did not reveal any significant differences either (Table 1).
2.1.2. Study of the inhibition of rootlet growth activity by bovine and human hemoglobin hydrolysates according to their degree of hydrolysis
Studies carried out on the hydrolysis of human hemoglobin revealed that the enzymatic process involved the same reaction mechanism as that observed in bovine hemoglobin, generally referred to as the 'zipper' mechanism. This mechanism generates a considerable diversity of peptides [22]. It is also important to note that different sets of peptides are produced at different degrees of hydrolysis (DH) [19]. Consequently, for the remainder of the study, it was essential to select several hydrolysates with varied peptide populations. Eight hydrolysates were chosen, with DHs of 0, 3, 4, 5, 6, 8, 10 and 18%, corresponding to hydrolysis times of 0, 5, 15 and 30 minutes, as well as 1, 2, 3 and 24 hours, respectively.
With this in mind, the effect of the influence of the degree of hydrolysis (DH) of bovine and human hemoglobin on the inhibition of LS rootlet growth was analyzed and presented in the form of dose-response curves in Figure 4A–D. The IC50 values obtained are recorded in Table 2. These results show that for some DH, bovine and human hemoglobin hydrolysates show a more marked effect in inhibiting rootlet growth than for others. For example, when examining the degree of hydrolysis (DH) of 0, the IC50 values were found to be 5.06 ± 2.00 mg/mL for bovine hemoglobin and 3.97 ± 0.60 mg/mL for human hemoglobin. In these circumstances, the inhibitory effects were less marked for both species compared with the other DHs. This was followed by DH of 3, with IC50 of 2.42 ± 0.96 mg/mL for bovine and 2.68 ± 0.67 mg/mL for human, and DH of 18, with IC50 of 2.25 ± 0.84 mg/mL for bovine and 2.25 ± 0.84 mg/mL for human. It should be emphasized that these results, although still significantly better than those observed for non-hydrolyzed hemoglobins at a DH of 0, demonstrate improvements of up to twice the efficacy. The other DHs showed very interesting inhibitory effects on rootlet growth, three to four times more intense than for the DH of 0. For example, DHs of 4, 5, 6, 8 and particularly 10% showed significantly greater inhibitory effects.
2.1.3. Study of the inhibition of rootlet growth activity of bovine and human hemoglobin hydrolysate fractions and peptidomics approach
In the light of previous results which revealed marked inhibitory effects on rootlet growth, particularly at a degree of hydrolysis (DH) of 10% (equivalent to a hydrolysis time of 3 hours), the fractionation of these hydrolysates was also carried out after a 3-hour peptic hydrolysis. The first fraction (Fraction 1) corresponds to the NKT peptide (α137-141, neokyotorphin), followed by fractions collected at 5-minute intervals (Figure 5A). Subsequent analyses were performed to assess their inhibitory effects on rootlet germination.
The results of this study made it possible to identify the specific bovine (B) and human (H) hemoglobin fractions with the strongest inhibitory effects on rootlet growth (Figure 5B–E). Analysis of the IC50 values revealed several fractions that stood out for their robust inhibitory activity (Table 3). By comparing the performance of fractions from the two types of hemoglobin, similarities as well as nuances in their bioactive potential were highlighted. Of the fractions analyzed, Fraction 1 stood out for its remarkable inhibitory efficacy. The IC50 values for this fraction are 29 ± 1 µg/mL for bovine hemoglobin and 45 ± 2µg/mL for human hemoglobin. These results indicate a significant inhibitory effect for both species. In the case of Fraction 2, there was a marked contrast between the two types of hemoglobin. The IC50 values of Fraction 2 from bovine hemoglobin were approximately 7 to 8 times better than those obtained from human hemoglobin. This observation clearly suggests the superiority of bovine hemoglobin for this specific fraction. Fractions 3, 4, 5, 6 and 9 stand out for their significant inhibitory capacity towards rootlet growth. At the same time, Fraction 8, whether of bovine or human origin, also has a lesser inhibitory effect than the fractions mentioned above. What links them is that despite their distinct peptide profiles from the two hemoglobin sources, they demonstrate an ability to inhibit LS rootlet growth. This uniformity in inhibitory performance between the two types of hemoglobin suggests a degree of convergence in their bioactive properties. Fraction 7 was also found to have inhibitory activity for both species, although this fraction exhibited relatively less inhibitory activity than the other fractions.
Next, triple RP-HPLC-MS/MS analyses were carried out following a 3-hour peptide hydrolysis on the two types of hemoglobin. A comparative analysis of the peptide populations in the fractions was carried out. To illustrate the results, a histogram (Figure 5F) showing the number of unique peptide sequences identified was used, to allow precise visualization of the disparities and similarities between the peptide populations in the fractions. With cross-referencing the results, it was found that the fractions with LS rootlet inhibitory properties in both species, particularly fractions 3, 4, 5, 6 and 7, had the highest number of peptide sequences identified. An exception is seen in fraction 1, corresponding to the NKT peptide (α137-141, neokyotorphin, which stands out as a unique and pure fraction.
2.1.4. Study of the inhibition of rootlet growth activity of pure antimicrobial peptides α137-141 and α1-32 with different structural characteristics.
To further our investigation, this section of the study focused on analyzing the inhibitory effects on LS rootlet growth of standard α137-141 and α1-32 antimicrobial peptides (Figure 6). These two peptides have distinct structural characteristics. The NKT peptide, or α137-141, is characterized by a 'random-coil' structure and a restricted number of amino acids (5), with a monoisotopic molecular weight of 654 Da. In contrast, the α1-32 peptide has an alpha-helix structure (monoisotopic molecular weight: 3327 Da) and a higher number of amino acids (32) [29,30]. These antimicrobial peptides differ in their IC50 values, reflecting the variety of their inhibitory effects. The standard peptide α137-141 shows an IC50 of 53 µg/mL, indicating strong inhibitory activity. This value is consistent with the IC50 obtained for NKT peptides derived from bovine (29±1 µg/mL) and human hemoglobin (45±2 µg/mL), respectively. In contrast, the α1-32 peptide exhibited less inhibitory activity with an IC50 of 280 µg/mL, representing a 5-fold lower value than NKT (Table 4). However, its inhibitory activity on LS rootlet growth remains positive.
2.2. Study of the inhibition of rootlet growth activity of bovine and human hemoglobin hydrolysates and their fractions: Covalent labeling of proteins using tRNAox.
2.2.1. Covalent labeling of Formylase by tRNAox
Affinity labeling with tRNAox was developed to identify amino acid residues located at the binding site of the CCA arm of tRNA on enzymes purified from the translational apparatus. These enzymes include the aminoacyl-tRNA synthetase family, which esterifies amino acids at the 3'-OH end of tRNAs, and formyl-methionyl-tRNAtransformylase (FMTS or formylase). These studies led to the discovery of the consensus sequence 332Lys-Met-Ser-Lys-Ser336 as the catalytic signature of all class 1 aminoacyl-tRNA synthetases. The Lys-335 residue of this tRNAox-labelled motif was shown to be a catalytic residue that stabilizes the transition state preceding the formation of the aminoacyl-tRNA product [26]. To develop tRNAox labeling of the human ribosomal eL42 protein, we reproduced a tRNAox labeling experiment for formylase (FMTS). Since any molecule that binds FMTs by preventing the FMTs-tRNAox complex interacts with the eL42 protein of the human 60S ribosome (a protein found overexpressed in a large number of human cancers), it is a good candidate for antitumor molecule research. Figure 7 shows formyl-methionyl-tRNAtransformylase (FMTS or formylase) in the presence and absence of tRNAox and 4-phenyl-3-thiosemicarbasone piperitone (UCK-36) analyzed by electrophoresis on a 10% polyacrylamide gel under denaturing conditions (SDS-PAGE), revealed by Coomassie blue staining. In well 1, we deposited free FMTS; in well 2, FMTS and tRNAox; in well 3, FMTS and tRNAox in the presence of 4-phenyl-3-thiosemicarbasone piperitone (UCK-36), an inhibitor molecule that is already known to make a covalent bond with the Lysine residue concerned instead of tRNAox.
In the presence of tRNAox (well 2), the 32 kDa control band corresponding to free FMTS (well 1) was transformed into a 57 kDa band corresponding to the covalent labeling of an FMTS molecule (32 kDa) by a tRNAox molecule (25 kDa) (well 2). Well, 3 shows that this molecule masks the side chain of the Lysine residue to prevent it from being labeled by tRNAox. It is therefore more competitive than tRNAox.
2.2.2. Testing of bovine and human hemoglobin peptide hydrolysates and their peptide fractions
Figure 8 shows the characterization of formylase (FMTS) in the presence and absence of tRNAox as well as bovine (8A) and human hemoglobin (8B) hydrolysates and their fractions, performed by 10% polyacrylamide gel electrophoresis under denaturing conditions (SDS-PAGE), followed by Coomassie blue staining. The samples were arranged as follows: well 2 contained free FMTS, well 3 contained FMTS in the presence of tRNAox, and wells 4 to 8 contained FMTS bound to tRNAox in the presence of the products to be evaluated. Observation of the results revealed that in well 2 (Figure 8A), a protein band of approximately 32 kDa was present, indicating the presence of formylase. Bands at around 32 kDa (corresponding to free FMTS) and at around 57 kDa (resulting from the covalent binding of a 32 kDa FMTS molecule to a 25 kDa tRNAox molecule) were visible in wells 3, 5, 6 and 7. However, in wells 4 and 8, the 57 kDa band was less intense. This observation was made for samples containing fraction 1 (representing bovine NKT) in well 4, as well as for the total hydrolysate in well 8. These results suggest that these samples could influence the interaction between FMTS and tRNAox, by modulating the formation of the covalent bond. Similar observations were made for human hemoglobin (Figure 8B). In well 4 (containing fraction 1 representing human NKT) and well 8 (total human hydrolysate), the 57 kDa band was also attenuated. This suggests that these samples may also affect the interaction between HTSF and tRNAox. The results indicate that fractions 1 and total hydrolysate of human and bovine hemoglobins may modulate the interaction between FMTS and tRNAox. This observation suggests that these peptide fractions could influence molecular binding mechanisms, potentially hindering efficient binding of the tRNA substrate to the ribosomal protein in the presence of chemotherapeutic compounds. In short, this experimental phase enriches our understanding of the competitive interactions between bioactive peptides and tRNA, as well as their possible interference with formylase.
3. Discussion
In the search for new therapeutic approaches in the fight against cancer, antimicrobial peptides (AMPs) are emerging as a promising and under-exploited avenue. Possessing the ability to selectively target cancer cells while minimizing their toxicity to rapidly proliferating healthy cells, AMPs also exhibit a low tendency to elicit resistance in their target [9,10,11]. In particular, the enzymatic hydrolysis of bovine hemoglobin generates a diverse range of peptides with numerous biological activities. However, antimicrobial activity remains the most frequently documented and studied in the scientific literature [17,18,19,29]. Human hemoglobin, like bovine hemoglobin, represents an optimal substrate for proteolysis and the production of bioactive peptides [20,21,22]. With this in mind, the main aim of this study was to generate bioactive peptides using enzymatic hydrolysis of bovine and human hemoglobin, followed by analysis of their potential anti-cancer properties using two distinct approaches. The first innovative approach has paved the way for a promising methodology for assessing the anti-cancer activity of various molecules, including our bovine and human hemoglobin hydrolysates and their peptide fractions.
Initial investigations focused on the influence of initial concentrations of bovine and human hemoglobin. Contrary to expectations, no significant differences were observed between the two species. The results showed that the inhibition of LS rootlet growth was not significantly influenced by the initial concentration of hemoglobin of any kind. This finding suggests that antimitotic properties are not directly correlated with initial hemoglobin concentration. Continuing the analysis, the effect of the degree of hydrolysis (DH) on antimitotic activity was explored. The results showed that bovine and human hemoglobin hydrolysates exhibited more pronounced inhibitory effects at certain DHs than at others. In particular, DHs of 3, 4, 5, 6, 8 and especially 10% showed significantly greater inhibitory effects than those observed at a DH of 0, and these effects were three to four times more intense. This suggests that the hydrolysis process generates bioactive compounds that are favorable for inhibiting rootlet growth, and it should be noted that different sets of peptides are generated depending on the degree of hydrolysis (DH) applied. Overall, advancing the degree of hydrolysis led to an increase in the number of peptide sequences identified, particularly for peptides with masses between 0 and 1000 Da [19]. A comparative analysis of the IC50 fractions of the two types of hemoglobin was also carried out. This revealed that despite the distinct peptide profiles from the two hemoglobin sources, fractions 3, 4, 5, 6 and 9 showed comparable inhibitory effects between cattle and humans. However, for Fraction 2, the IC50 values obtained from bovine hemoglobin were significantly better (around 7 to 8 times) than those obtained from human hemoglobin. This suggests the superiority of bovine hemoglobin for this specific fraction. As for our peptide of interest, NKT, identified in Fraction 1, it stands out for its particularly robust inhibition, as shown by its considerably low IC50 values, demonstrating a significant capacity to restrict rootlet growth. IC50 values for bovine hemoglobin are 29µg/mL and for human hemoglobin are 45µg/mL, which is 10 to 15 times higher than other fractions. This observation could be attributed to the pure nature of this fraction, combined with the presence of a peptide with exceptionally potent antimicrobial activity. At the same time, the results obtained with standard NKT confirm this trend, showing particularly interesting IC50 values. In addition, it is essential to highlight another relevant criterion. These results highlight the diversity of inhibitory effects of bioactive peptides and underline the major importance of their specific structural feature in the regulation of rootlet growth. Specifically, the α137-141 peptide has a 'random-coil' structure and a molecular weight of 654 Da, with an IC50 of 53 µg/mL. In comparison, the α1-32 peptide, an alpha helix with a molecular weight of 3327 Da, shows an IC50 of 280µg/mL, 5 times higher. However, its inhibitory capacity remains significant [29,30]. These observations highlight the importance of several parameters in the hydrolysis process and their impact on anticancer activity. They also underline the fact that hydrolysates and their fractions, in particular the NKT fraction, specifically target the host cell's protein synthesis mechanism, a crucial aspect of curbing tumor proliferation. Recent advances in cancer therapy underline the importance of targeting multiple translation elements and signaling pathways. For example, the synergistic approach of targeting both HSP 90/70 and the 26S proteasome is emerging as a promising strategy for treating cancer, highlighting the crucial role of multifactorial approaches [31]. Various compounds have been identified as potential cancer therapies, targeting different translation factors [32]. For example, some antibiotics such as puromycin and sparsomycin act by mimicking aminoacyl-tRNA or blocking peptide synthetase to inhibit protein synthesis [33]. Cycloheximide is another example of a protein synthesis inhibitor, disrupting peptidyl transferase in eukaryotic ribosomes [34]. Doxorubicin (DOX) inhibits eukaryotic ribosome formation [35], while erythromycin (ERY) targets protein synthesis in prokaryotic and eukaryotic cells [36]. Doxycycline (DOC) directly disrupts protease-activated receptor 1 to slow tumor progression [37]. Monensin (MON) targets the EGFR signaling pathway, inhibiting cell proliferation and growth in chemotherapy-resistant pancreatic cancer cells [38]. Amikacin (AMK) binds to the A site of 16S RNA similarly to kanamycin A, with specific interactions between the L-(-)-γ-amino-α-hydroxybutyryl group and RNA [39]. What is particularly remarkable is that the combination of these various compounds exhibited notable anticancer potential by inducing significant inhibition of rootlet growth [23].
Subsequently, a striking aspect emerges from the results of the second method of assessing anti-cancer activity, namely the preliminary study centered on the in vitro analysis of the anti-cancer activity of hydrolysates and their peptide fractions, focusing on the eL42 protein. These findings not only reinforce the findings of the first approach but also support the essential hypothesis that inhibition of protein synthesis is central to the development of carcinoma. Indeed, the results suggest that samples containing the 1(NKT) fractions and the total hydrolysate for both types of hemoglobin may exert modulatory effects on the interaction between HTSF and tRNAox. This finding highlights the possibility that these peptide fractions may influence molecular binding mechanisms. In other words, there could be potential competition leading to the inability of the tRNA substrate to bind efficiently to the ribosomal protein in their presence. This observation is particularly striking for NKT, whether derived from human or bovine hemoglobin, as it is perfectly consistent with its remarkable efficacy in inhibiting LS seed germination.
These observations suggest that NKT could be one of the most promising pathways for inhibitory activity. In addition, its significant antimicrobial and antioxidant properties could qualify it as a potential candidate for inclusion in the class of anticancer agents.
In short, this experimental phase will enable us to gain a better understanding of the possible competitive interactions between bioactive peptides and tRNA, as well as their possible interference with formylase. This approach opens up new research prospects for the targeted discovery of anti-cancer molecules. However, further investigations are crucial to detail the mechanisms underlying these inhibitory effects and to better understand the influence of our bioactive peptides on potential competition with transfer RNA (tRNA) for binding to the eL42 protein within the human 80S ribosome.
4. Materials and methods
4.1. Materials: Reagents, solvents and standards used
All chemicals and solvents were of analytical grade from commercial suppliers: Sigma-Aldrich (Saint-Quentin Fallavier, France) or Flandres Chimie (Villeneuve d'Ascq, France). Ultrapure water was prepared using a Milli-Q system in the laboratory. Purified bovine hemoglobin (H2625) powder, dark brown and purified human hemoglobin (H7379), and dark red were purchased from Sigma-Aldrich (Saint-Quentin Fallavier, France). The hemoglobins were stored at 4°C before use. Neokyotorfin (NKT) standard α137-141 and Alpha 1-32 were supplied by Genecust (Luxembourg) and stored at -20°C until use. Pepsin is a lyophilized powder derived from porcine gastric mucosa and was purchased from Sigma-Aldrich (P6887, (Saint-Quentin Fallavier, France)). Pepsin activity was measured at 3350 AU/mg protein according to a protocol established by the supplier Sigma-Aldrich. pepsin was stored at -20 °C. Watercress seed (Lepidium sativum) brand Truffaut (zone commercial Cora - Rue du 19 Mars 1962 - 02100 Saint-Quentin, France)
4.2. Preparation of bovine and human hemoglobin hydrolysates
Conventional enzymatic hydrolysis using pepsin has been used to compare two types of purified hemoglobin: bovine hemoglobin and purified human hemoglobin. Enzymatic hydrolysis of bovine and human hemoglobin has been shown in several studies to produce active peptides, including the peptide α137-141, which has a variety of biological activities [19,22,29,40,41].
4.3. Preparation of the stock solution
Stock solutions were prepared by adding 15 g of bovine (BH) or human (HH) hemoglobin to 100 ml of ultrapure water. After centrifugation at 4000 min -1 for 30 min (Eppendorf AG, Hamburg, Germany; Centrifuge 5804 R, Brinkmann Instruments, Westbury, NY, USA), the supernatant was collected. The Drabkin method, established by Crosby, Munn and Furth in 1954, was used to determine the actual concentrations of BH (CBH ) and HH (CHH) using a spectrophotometric approach to quantify hemoglobin. To do this, 20 µL of the sample was mixed with 10 mL of Drabkin D5941 reagent (Sigma-Aldrich), followed by a 15 min incubation at room temperature and protected from light. Absorbance was then measured at 540 nm using a UV spectrophotometer (ChemStation UV spectrophotometer - VIS 8453A, Agilent Technologies, USA). The results were adjusted according to the calibration curve. From the C concentrations of the bovine or human stock solution, various solutions of hemoglobin were obtained by dilution, reaching the following specific concentrations: 1%, 2%, 8% and 10% (w/v).
4.4. Hydrolysis process
To denature the hemoglobin solution, which was initially in its native, "globular" form, the pH was adjusted to 3.5 using 2 M hydrochloric acid, which was added gradually. The hydrolysis reaction was started by the addition of pepsin (EC 3.4.23.1, 3200-4500 units mg -1 of protein) which had previously been solubilized in ultrapure water, with an enzyme/substrate ratio equal to 1/11 (mole/mole). Samples were taken at T 0, T 2.5, T 30, T 60, T 120 and T 180 min of hydrolysis, corresponding to different degrees of hydrolysis. The peptic hydrolysis reaction was then stopped by adding sodium chloride NaCl 5M to a final pH of 9, which deactivated the enzyme. The temperature was maintained at a constant 30°C throughout the reaction. The samples were stored at -20°C, then lyophilized, and the powder recovered was ready for testing.
4.5. Fractionation of peptide hydrolysates by semi-preparative HPLC
In order to complete the research results, the total hydrolysates were fractionated. To do this, it was necessary to increase the concentration of the initial substrate used in order to recover more active peptides in a single step so that they could be used for cancer treatment. Consequently, a hydrolysate can be chosen, more specifically those with a DH of 10% and hydrolysis times of 3 hours. It has been shown that increasing the peptide concentration of the 10% (w/v) hydrolysate allows the recovery of up to a 10-fold higher concentration of active α137-141 peptide compared to the 10% (w/v) hydrolysate [22]. This results in a 10-fold enrichment compared with the initial hydrolysate and could suggest an interesting use for co-products. The fractions were collected every 5 minutes in a tube using a liquid chromatography system. This system consisted of a Waters 600E automated gradient controller pump module, a Waters Wisp 717 automated sampling device and a Waters 996 photodiode array detector. Waters software was used to plot, acquire and analyze the chromatographic data. All chromatographic procedures were carried out using a semi-preparative C4 column (250 mm x 4.6 mm, 3 mm internal diameter). The mobile phases were ultrapure water/trifluoroacetic acid (1000:1, v/v) as solvent A, and acetonitrile/trifluoroacetic acid (1000:1, v/v) as solvent B. Samples were filtered at 0.20 μm and then injected. On-line UV absorbance scans were performed between 200 and 390 nm at a rate of one spectrum per second with a resolution of 1.2 nm [18,42]. The injection volume was 60 µL. The flow rate was 0.6 mL min -1. The elution program was as follows: The mobile phases were LC-MS grade water with 0.1% trifluoroacetic acid (1000:1, v/v) as solvent A, and LC-MS grade acetonitrile, ACN with 0.1% trifluoroacetic acid (1000:1, v/v) as solvent B. A gradient was applied with solvent B increasing from 5% to 30% over 30 min, then to 60% for 10 min and maintained until 47min at 95%, then back to initial conditions. The tubes containing the different fractions were dried with speedVac and stored at -20°C.
4.6. RP-UPLC analysis and mass spectrometry
After centrifugation for 10 minutes at 8000 × g, 10 µL samples of bovine and human hemoglobin hydrolysate at a concentration of 30 mg-mL-1 were subjected to triplicate RP-HPLC-MS/MS analysis using an ACQUITY UPLC system (Waters Corporation, France). The peptides were separated on a C18 column (150 × 3.0 mm, 2.6 µm, Uptisphere CS Evolution, Interchim, France). The mobile phases consisted of solvent A (0.1% (v/v) formic acid / 99.9% (v/v) water) and solvent B (0.1% (v/v) formic acid / 99.9% (v/v) acetonitrile (ACN)). The ACN gradient (flow rate 0.5 mL-min-1) was as follows: from 5% to 30% solvent B in 40 minutes, from 30% to 100% solvent B in 10 minutes, followed by washes and equilibrations using 100% and 1% solvent B respectively for 5 minutes each. The eluate was introduced into the electrospray ionization source of the qTOFSynapt G2-Si™ (Waters Corporation, Manchester, UK), previously calibrated with sodium solution. Mass spectrometry (MS) measurements were performed in sensitivity, positive ion, and data-dependent analysis (DDA) mode using Mass Lynx 4.2 software (Waters). The source temperature was maintained at 150°C, and the capillary and cone voltages were set at 3000 and 60 V-MS. Data was collected for m/z values between 50 and 2000 Da, with a scan time of 0.2 s. Up to 10 precursor ions were selected for MS/MS analysis with an intensity threshold of 10,000. MS/MS data were collected in collision-induced dissociation (CID) fragmentation mode, with a scan time of 0.1 s and specified voltages of 8-9 V and 40-90 V for lower and higher molecular weight ions, respectively.
4.7. Determination of the anti-cancer activity of hydrolysates and peptide fractions
4.7.1. Lepidium sativum (LS) rootlet growth test
Description of the methods used.
The anti-cancer efficacy of peptic hydrolysates of human and bovine hemoglobin and their peptide fractions was determined using a patented method developed by our laboratory [23]. This method is used by pharmaceutical laboratories (TBC - France) for the rapid screening of candidate molecules for anticancer, antibiotic and antiparasitic drugs. The technique involves assessing the effect of the various products tested (human and bovine hemoglobin hydrolysates and fractions) on rootlet growth during germination by measuring length. The use of LS seeds for the screening of anti-cancer drug candidates appears to be a promising and ethically responsible method, offering an alternative to animal experimentation. Although this method cannot completely replace studies using animal models, it represents an important opportunity to reduce the number of animals used and cut preclinical research costs. Other alternatives to animal experimentation exist, such as sophisticated tests using cell cultures and human tissues (in vitro), advanced computer models (in silico) and studies involving human volunteers. In addition, the emergence of 'organs on a chip', which simulate the structure and function of human organs, offers new prospects for preclinical studies. However, all these experimental approaches remain complex to put into practice and are associated with high costs. Nevertheless, validating the efficacy of this model is not limited to compounds traditionally used in oncology, but also extends to antibiotics and antiparasitics. All molecules or xenobiotics that inhibit protein synthesis [23].
Preparation of vegetable seeds
LS seeds are pregerminated from seeds in Petri dishes on filter paper or blotting paper soaked in distilled water in total darkness for 24 hours. The aim of pre-germinating the seeds is to eliminate those that do not show signs of germination with a closed cuticle. Seeds that do not show these signs are not likely to germinate and are therefore removed before the tests begin. This ensures that only seeds suitable for germination are used in subsequent tests. The seeds are germinated in the presence of different concentrations of the products to be tested, then the results are compared with a negative control. This control consists of normal germination under the same conditions as the treated seeds but with distilled water and no added product.
Preparation of test concentrations
Different concentrations of the products to be tested are prepared, ranging from low to high concentrations, generally from picograms (pg/ml) to milligrams (mg/ml). Next, 5 ml of each product at different concentrations were added to Petri dishes containing watercress seeds. After incubation in the dark for 3 days (72 hours), the length of the rootlets was measured with a ruler and recorded in centimeters. To obtain more accurate results, it is recommended that more than 8 measurements are taken per concentration of test or control product, to allow a proper comparison of the effects of each product on rootlet growth.
4.7.2. Covalent labeling of proteins with tRNAox by Formylase
Preparation of tRNAox
In an Eppendorf tube, 4.8 µL of tRNA (419 µM) was taken and 4 µL of NaIO4 (0.5 M) and 2 µL of NH4OAc (5 M) at pH 5.0 were added. This mixture was made up to 50 µL with water (DNase/RNAase free) and incubated for 1 hour at 4°C. After removing the NaIO4 via a G-25 column, the sample was added to the center of the resin in the G-25 column. Centrifuging at 3000 rpm for 2 minutes recovered a sample of approximately 110 µL. Precipitation was achieved by adding 0.2 times the sample volume of NaCl (5 M) to the tRNAox, i.e. 22 µL of NaCl, and 2.2 times the sample volume of cold ethanol -20°C, i.e. 242 µL of ethanol. The mixture was then prepared by adding -20°C cold ethanol to 700 µL, homogenizing and leaving it to stand overnight at -20°C. Finally, by centrifugation at 13,000 rpm at 4°C for 45 minutes, the already oxidized tRNA was concentrated and the ethanol was allowed to evaporate. The precipitate obtained was re-suspended in 1 mL of water (without DNase/RNAase), giving tRNAox prepared at 2000 pmoles.
Figure 1.
tRNA oxidation reaction.
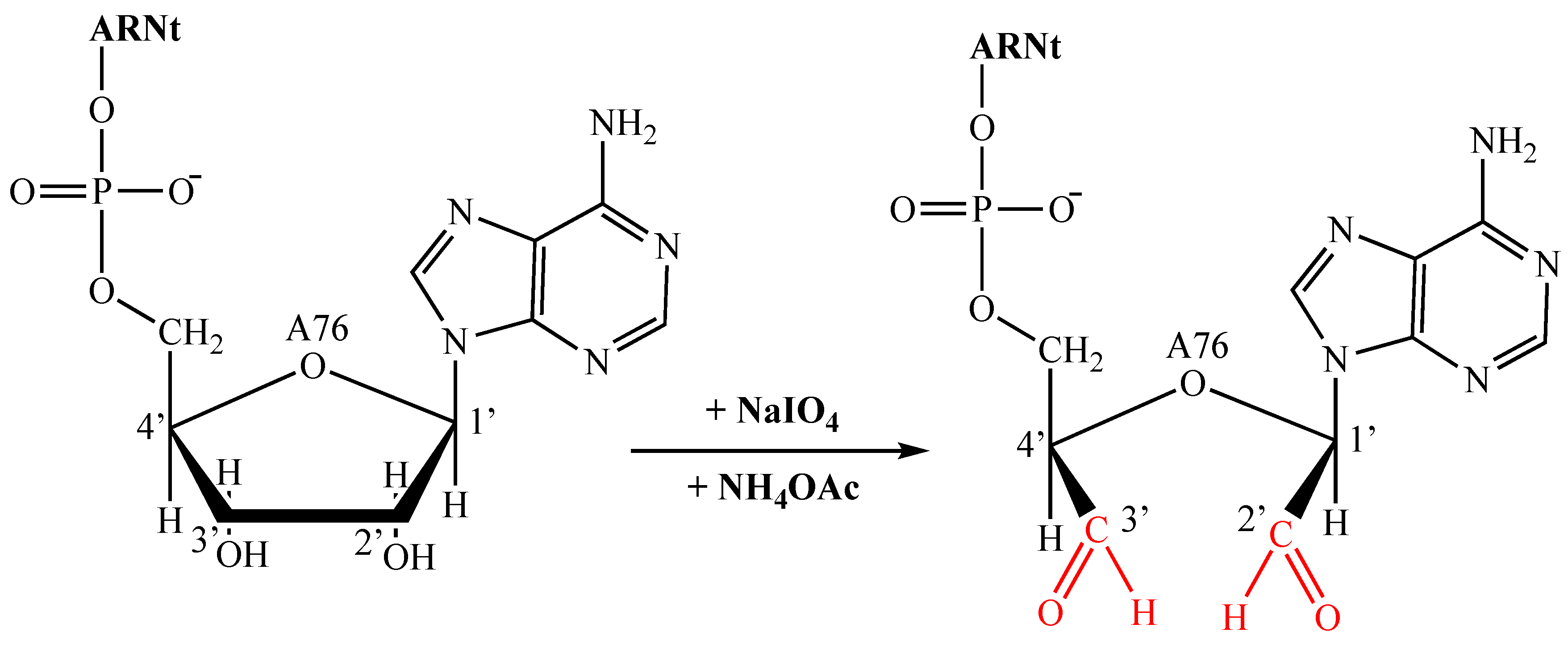
tRNAox oxidation and protein labeling reactions
The principle of covalent labeling involves the use of a reactive analog of tRNA as an affinity indicator targeted to the catalytic site of purified enzymes within the translation machinery, as well as to the A, P, and E sites of ribosomes. This analog is oxidized tRNA (tRNAox), which is tRNA with the 2', 3' cis-diol group of the 3'-terminal adenosine ribose converted to the 2', 3' dialdehyde group by sodium periodate [27]. The aldehyde function in the 2' or 3' position of the tRNAox can form a Schiff base with an amine function of a lysine (or arginine) residue of a protein. This Schiff base is then stabilized by a specific reducing agent, sodium cyanoborohydride (NaBH3CN), leading to the creation of a covalent complex between the tRNA and the lysine residue of the protein [27] See Figure 2. Labeling of formylase using tRNAox outside the ribosome was performed by adding non-radioactive tRNAox at a concentration of 100 µM to a 10 µL reaction mixture (two types were used: tRNA (Asp)4-ox and tRNA (Asp)15-ox). Formylase must be present at a concentration 10 times lower than that of tRNAox, i.e. at 10 µM in the 10 µL reaction mixture. This mixture also includes 1 µL of 50 mM sodium cyanoborohydride (NaBH3CN) and 1 µL of 10X buffer (a 10-fold more concentrated labeling buffer). The mixture was then incubated for 1 hour at 37°C. At the end of labelling, the mixtures were treated by adding 1.5 µL of freshly prepared 0.5 M sodium borohydride (NaBH4) in water (DNase/RNAase free) and 3.5 µL of 3X Laemmli. 15 µL portions of the NaBH4-treated samples were analyzed by electrophoresis on a 10% polyacrylamide gel under denaturing conditions (SDS-PAGE) revealed by Coomassie blue staining.
4.8. Statistical analysis
Rootlet lengths by concentration are presented as mean values ± SE (standard error) for separate experiments using n seeds. Graphs of concentration – responses curves were determined using nonlinear regression and were fitted to the Hill equation by an iterative least-squares method (GraphPad Prism 8.0 Software San Diego.CA. USA) to provide estimates of the maximum effective concentration IC50 (the negative logarithm of the agonist concentration producing 50% of maximum inhibition). For the comparison of the different effects against the control. One-way analysis of variance (ANOVA) was performed followed by multiple comparison t-tests. For a simple comparison of unpaired IC50, Statistical significance was determined using the Bonferroni-Dunn method, with alpha = 0.05. Each row was analyzed individually, without assuming a consistent SD.
5. Conclusion
In conclusion, this study explored the anticancer potential of bioactive peptides derived from the enzymatic hydrolysis of bovine and human hemoglobin. The investigation of key parameters such as the initial concentration of hemoglobin, the degree of hydrolysis and the structural characteristics of the antimicrobial peptides highlighted the influence of these factors on the antimitotic activity of the peptides. In particular, the NKT peptide emerged as a central player in the inhibition of rootlet sprouting, demonstrating exceptionally low IC50 values and suggesting a potentially novel pathway for curbing tumor proliferation. In vitro analyses reinforced the hypothesis that inhibition of protein synthesis plays a key role in the anti-cancer mechanism of these peptides. The interactions observed between the peptide fractions, total hydrolysates and formylase shed new light on the molecular mechanisms underlying these inhibitory effects. In short, this study broadens the horizon for future research into anti-cancer molecules. The specific properties of the NKT peptide, combined with its antimicrobial and antioxidant potential, make it an attractive candidate for the development of innovative treatments. However, to translate these discoveries into tangible clinical applications, further investigations and in vivo studies will be essential to confirm their efficacy and safety. These scientific advances are inspiring new hope in the fight against cancer, paving the way for effective targeted therapies.
Author Contributions
Conceptualization, NN, AO, CH and BE.; methodology, NN, AO, CH, BE.; software, MB MA, BE; validation, NN, AO, and BE.; formal analysis, AO, NN, BE; investigation, AO, NN, MB, BE.; resources, NN, BG, BE.; data curation, AO, BE, NN, MB, ASA, MA.; writing—original draft preparation, AO, OMN, CH, ASA, BE.; writing—review and editing, NN, AO, BE.; visualization, AO,NN, BE, MB, ASA, MA.; supervision, NN, BG., BE.; project administration, NN.; BG., BE. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
The authors express their gratitude to the Advanced High Throughput Technologies Platform for Biorefineries Catalysts Design, known as "REALCAT," for their support. The REALCAT platform is funded by a French governmental subsidy administered by the French National Research Agency (ANR) as part of the "Future Investments" program (ANR-11-EQPX-0037). The authors would also like to acknowledge the financial contributions from the Hauts-de-France region, the FEDER (European Regional Development Fund), the Ecole Centrale de Lille, and the Centrale Initiatives Foundation, which enabled the acquisition of equipment for the REALCAT platform. The authors extend their appreciation to the Deputyship for Research & Innovation, Ministry of Education in Saudi Arabia for funding this research through the project number IFP-IMSIU-2023097. The authors also appreciate the Deanship of Scientific Research at Imam Mohammad Ibn Saud Islamic University (IMSIU) for supporting this project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The ever-increasing importance of cancer as a leading cause of premature death worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef] [PubMed]
- Bakare, O.O.; Gokul, A.; Wu, R.; Niekerk, L.-A.; Klein, A.; Keyster, M. Biomedical Relevance of Novel Anticancer Peptides in the Sensitive Treatment of Cancer. Biomolecules 2021, 11, 1120. [Google Scholar] [CrossRef]
- Kolberg, H.C.; Villena-Heinsen, C.; Deml, M.M.; Kraemer, S.; Diedrich, K.; Friedrich, M. Relationship between chemotherapy with paclitaxel, cisplatin, vinorelbine and titanocene dichloride and expression of proliferation markers and tumour suppressor gene p53 in human ovarian cancer xenografts in nude mice. Eur. J. Gynaecol. Oncol. 2005, 26, 398–402. [Google Scholar] [PubMed]
- Pierard, G.E.; Focan, C.; Lapiere, C.M. Cell Proliferation in a Malignant Angioendothelioma during Sequential Chemotherapy. J. Cutan. Pathol. 1979, 6, 479–485. [Google Scholar] [CrossRef]
- Kato, Y.; Sato, J.; Kato, R.; Takata, R.; Obara, W. Side effect and supportive care to combination of gemcitabine and cisplatin chemotherapy for the advanced urothelial cancer. Nihon rinsho. Jpn. J. Clin. Med. 2015, 73, 609–613. [Google Scholar]
- Saxena, A. Cancer chemotherapy and its side effect management. Nurs. J. India 2006, 97, 109–110. [Google Scholar] [CrossRef]
- Chism, D.D.; De Silva, D.; E Whang, Y. Mechanisms of acquired resistance to androgen receptor targeting drugs in castration-resistant prostate cancer. Expert Rev. Anticancer. Ther. 2014, 14, 1369–1378. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The Different Mechanisms of Cancer Drug Resistance: A Brief Review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef]
- Deslouches, B.; Di, Y.P. Antimicrobial peptides with selective antitumor mechanisms: prospect for anticancer applications. Oncotarget 2017, 8, 46635–46651. [Google Scholar] [CrossRef]
- Tornesello, A.L.; Borrelli, A.; Buonaguro, L.; Buonaguro, F.M.; Tornesello, M.L. Antimicrobial Peptides as Anticancer Agents: Functional Properties and Biological Activities. Molecules 2020, 25, 2850. [Google Scholar] [CrossRef]
- Kordi, M.; Borzouyi, Z.; Chitsaz, S.; Asmaei, M.H.; Salami, R.; Tabarzad, M. Antimicrobial peptides with anticancer activity: Today status, trends and their computational design. Arch. Biochem. Biophys. 2023, 733, 109484. [Google Scholar] [CrossRef]
- Nedjar-Arroume, N.; Zouari, O.; Przybylski, R.; Hannioui, M.; Sion, L.; Dhulster, P. High Added-Value Co-Product: the Porcine Cruor is an Attractive Source of Active Peptides. J. Nutr. Heal. Food Sci. 2020, 8, 1–9. [Google Scholar] [CrossRef]
- Lignot, B.; Froidevaux, R.; Nedjar-Arroume, N.; Guillochon, D. Solvent effect on kinetics of appearance of neokyotorphin, VV-haemorphin-4 and a bradykinin-potentiating peptide in the course of peptic hydrolysis of bovine haemoglobin, Biotechnol. Appl. Biochem. 1999, 30, 201–207. [Google Scholar]
- Zhao, Q.; Piot, J. Investigation of inhibition angiotensin-converting enzyme (ACE) activity and opioid activity of two hemorphins, LVV-hemorphin-5 and VV-hemorphin-5, isolated from a defined peptic hydrolysate of bovine hemoglobin. Neuropeptides 1997, 31, 147–153. [Google Scholar] [CrossRef]
- Vercaigne-Marko, D.; Kosciarz, E.; Nedjar-Arroume, N.; Guillochon, D. Improvement of Staphylococcus aureus-V8-protease hydrolysis of bovine haemoglobin by its adsorption on to a solid phase in the presence of SDS: peptide mapping and obtention of two haemopoietic peptides. Biotechnol. Appl. Biochem. 2000, 31, 127–134. [Google Scholar] [CrossRef]
- Adje, E.Y.; Balti, R.; Kouach, M.; Guillochon, D.; Nedjar-Arroume, N. α 67-106 of bovine hemoglobin: a new family of antimicrobial and angiotensin I-converting enzyme inhibitory peptides. Eur. Food Res. Technol. 2011, 232, 637–646. [Google Scholar] [CrossRef]
- Daoud, R.; Dubois, V.; Bors-Dodita, L.; Nedjar-Arroume, N.; Krier, F.; Chihib, N.-E.; Mary, P.; Kouach, M.; Briand, G.; Guillochon, D. New antibacterial peptide derived from bovine hemoglobin. Peptides 2005, 26, 713–719. [Google Scholar] [CrossRef]
- Choisnard, L.; Froidevaux, R.; Nedjar-Arroume, N.; Lignot, B.; Vercaigne-Marko, D.; Krier, F.; Dhulster, P.; Guillochon, D. Kinetic study of the appearance of an anti-bacterial peptide in the course of bovine haemoglobin peptic hydrolysis. Biotechnol. Appl. Biochem. 2002, 36, 187–194. [Google Scholar] [CrossRef]
- Przybylski, R.; Firdaous, L.; Châtaigné, G.; Dhulster, P.; Nedjar, N. Production of an antimicrobial peptide derived from slaughterhouse by-product and its potential application on meat as preservative. Food Chem. 2016, 211, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Parish, C.A.; Jiang, H.; Tokiwa, Y.; Berova, N.; Nakanishi, K.; McCabe, D.; Zuckerman, W.; Xia, M.M.; Gabay, J.E. Broad-spectrum antimicrobial activity of hemoglobin. Bioorganic Med. Chem. 2001, 9, 377–382. [Google Scholar] [CrossRef]
- Liepke, C.; Baxmann, S.; Heine, C.; Breithaupt, N.; Ständker, L.; Forssmann, W.-G. Human hemoglobin-derived peptides exhibit antimicrobial activity: a class of host defense peptides. J. Chromatogr. B 2003, 791, 345–356. [Google Scholar] [CrossRef]
- Outman, A.; Deracinois, B.; Flahaut, C.; Diab, M.A.; Gressier, B.; Eto, B.; Nedjar, N. Potential of Human Hemoglobin as a Source of Bioactive Peptides: Comparative Study of Enzymatic Hydrolysis with Bovine Hemoglobin and the Production of Active Peptide α137–141. Int. J. Mol. Sci. 2023, 24, 11921. [Google Scholar] [CrossRef] [PubMed]
- Outman, A.; Elombo, F.K.; Abidi, C.; Bouhrim, M.; Hountondji, C.; Al-Zharani, M.; Nasr, F.A.; Aleissa, M.S.; Gressier, B.; Nedjar, N.; Eto, B. Protein synthesis by the plant rootlet as a target for the rapid screening of anticancer drugs. J. Biol. Regul. Homeost. Agents. In Press. 2023. [Google Scholar] [CrossRef]
- Outman, A.; Nedjar, N.; Gressier, B.; Eto, B. Method of rapid and simple screening of anticancer, antibiotic, antifungal and antiparasitic drugs and results obtained by this process., FR2213717, n.d.
- Kim, J.-H.; You, K.-R.; Kim, I.H.; Cho, B.-H.; Kim, C.-Y.; Kim, D.-G. Over-expression of the ribosomal protein L36a gene is associated with cellular proliferation in hepatocellular carcinoma. Hepatology 2004, 39, 129–138. [Google Scholar] [CrossRef]
- Hountondji, C.; Créchet, J.-B.; Tanaka, M.; Suzuki, M.; Nakayama, J.-I.; Aguida, B.; Bulygin, K.; Cognet, J.; Karpova, G.; Baouz, S. Ribosomal protein eL42 contributes to the catalytic activity of the yeast ribosome at the elongation step of translation. Biochimie 2018, 158, 20–33. [Google Scholar] [CrossRef]
- Hountondji, C.; Bulygin, K.; Woisard, A.; Tuffery, P.; Créchet, J.-B.; Pech, M.; Nierhaus, K.H.; Karpova, G.; Baouz, S. Lys53 of Ribosomal Protein L36AL and the CCA End of a tRNA at the P/E Hybrid Site Are in Close Proximity on the Human Ribosome. ChemBioChem 2012, 13, 1791–1797. [Google Scholar] [CrossRef]
- Hountondji, C.; Fayat, G.; Blanquet, S. Complete Inactivation and Labeling of Methionyl-tRNA Synthetase by Periodate-Treated Initiator tRNA in the Presence of Sodium Cyanohydridoborate. JBIC J. Biol. Inorg. Chem. 1979, 102, 247–250. [Google Scholar] [CrossRef]
- Nedjar-Arroume, N.; Dubois-Delval, V.; Adje, E.Y.; Traisnel, J.; Krier, F.; Mary, P.; Kouach, M.; Briand, G.; Guillochon, D. Bovine hemoglobin: An attractive source of antibacterial peptides. Peptides 2008, 29, 969–977. [Google Scholar] [CrossRef] [PubMed]
- V. Dubois, Préparation de peptides antimicrobiens à partir de l’hydrolyse enzymatique de deux protéines: l’hémoglobine bovine et l’α-lactalbumine bovine, Lille 1, 2006.
- Wang, R.E. Targeting heat shock proteins 70/90 and proteasome for cancer therapy, Curr. Med. Chem. 2011, 18, 4250–4264. [Google Scholar]
- Kovalski, J.R.; Kuzuoglu-Ozturk, D.; Ruggero, D. Protein synthesis control in cancer: selectivity and therapeutic targeting. EMBO J. 2022, 41, e109823. [Google Scholar] [CrossRef]
- Porse, B.T.; Kirillov, S.V.; Awayez, M.J.; Ottenheijm, H.C.; Garrett, R.A. Direct crosslinking of the antitumor antibiotic sparsomycin, and its derivatives, to A2602 in the peptidyl transferase center of 23S-like rRNA within ribosome-tRNA complexes, Proc. Natl. Acad. Sci. 1999, 96, 9003–9008. [Google Scholar] [CrossRef]
- Lockhead, S.; Moskaleva, A.; Kamenz, J.; Chen, Y.; Kang, M.; Reddy, A.R.; Santos, S.D.; Ferrell, J.E. The Apparent Requirement for Protein Synthesis during G2 Phase Is due to Checkpoint Activation. Cell Rep. 2020, 32, 107901–107901. [Google Scholar] [CrossRef]
- Awad, D.; Prattes, M.; Kofler, L.; Rössler, I.; Loibl, M.; Pertl, M.; Zisser, G.; Wolinski, H.; Pertschy, B.; Bergler, H. Inhibiting eukaryotic ribosome biogenesis. BMC Biol. 2019, 17, 46. [Google Scholar] [CrossRef]
- Dinos, G.P. The macrolide antibiotic renaissance. Br. J. Pharmacol. 2017, 174, 2967–2983. [Google Scholar] [CrossRef]
- Zhong, W.; Chen, S.; Zhang, Q.; Xiao, T.; Qin, Y.; Gu, J.; Sun, B.; Liu, Y.; Jing, X.; Hu, X.; et al. Doxycycline directly targets PAR1 to suppress tumor progression. Oncotarget 2017, 8, 16829–16842. [Google Scholar] [CrossRef]
- Wang, X.; Wu, X.; Zhang, Z.; Ma, C.; Wu, T.; Tang, S.; Zeng, Z.; Huang, S.; Gong, C.; Yuan, C.; et al. Monensin inhibits cell proliferation and tumor growth of chemo-resistant pancreatic cancer cells by targeting the EGFR signaling pathway. Sci. Rep. 2018, 8, 17914. [Google Scholar] [CrossRef] [PubMed]
- Kondo, J.; François, B.; Russell, R.J.; Murray, J.B.; Westhof, E. Crystal structure of the bacterial ribosomal decoding site complexed with amikacin containing the γ-amino-α-hydroxybutyryl (haba) group. Biochimie 2006, 88, 1027–1031. [Google Scholar] [CrossRef]
- Takagi, H.; Shiomi, H.; Fukui, K.; Hayashi, K.; Kiso, Y.; Kitagawa, K. Isolation of a novel analgesic pentapeptide, neo-kyotorphin, from bovine brain. Life Sci. 1982, 31, 1733–1736. [Google Scholar] [CrossRef]
- Catiau, L.; Traisnel, J.; Delval-Dubois, V.; Chihib, N.-E.; Guillochon, D.; Nedjar-Arroume, N. Minimal antimicrobial peptidic sequence from hemoglobin alpha-chain: KYR. Peptides 2010, 32, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Sannier, F.; Piot, J.M. Kinetics of appearance of four hemorphins from bovine hemoglobin peptic hydrolysates by HPLC coupled with photodiode array detection. Biochim. et Biophys. Acta (BBA) - Protein Struct. Mol. Enzym. 1996, 1295, 73–80. [Google Scholar] [CrossRef]
Figure 3.
Effect of initial bovine (A) and human (B) hemoglobin concentration on LS rootlet growth. (A) shows the effect of initial human hemoglobin concentration (1, 2, 8 and 10%) and (B) the effect of initial bovine hemoglobin concentration (1, 2, 8 and 10%).
Figure 3.
Effect of initial bovine (A) and human (B) hemoglobin concentration on LS rootlet growth. (A) shows the effect of initial human hemoglobin concentration (1, 2, 8 and 10%) and (B) the effect of initial bovine hemoglobin concentration (1, 2, 8 and 10%).
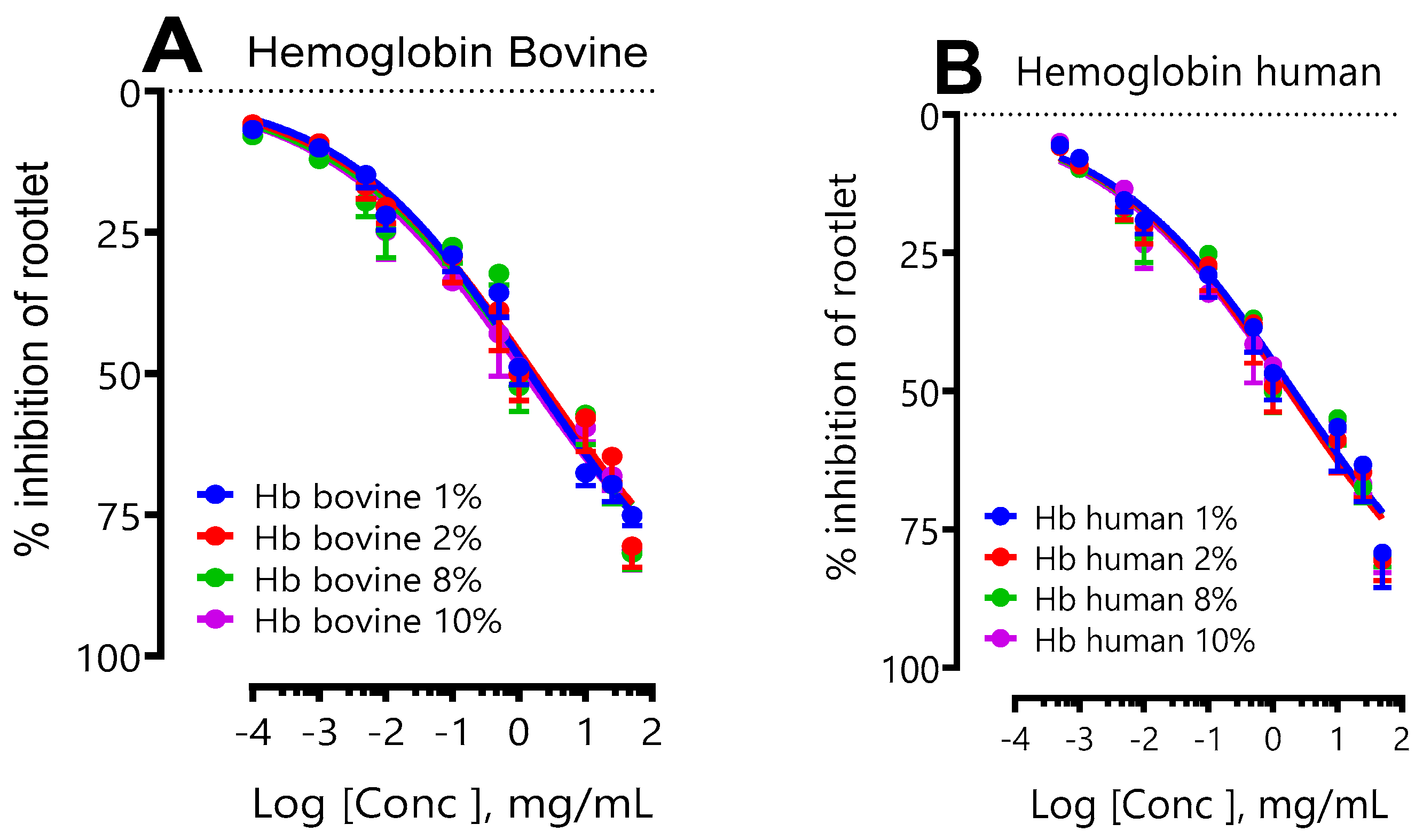
Figure 4.
Effect of the degree of hydrolysis (DH) of bovine (A-B) and human (C-D) hemoglobin, i.e. a DH ranging from 0 to 18%, on the inhibition of LS rootlet growth.
Figure 4.
Effect of the degree of hydrolysis (DH) of bovine (A-B) and human (C-D) hemoglobin, i.e. a DH ranging from 0 to 18%, on the inhibition of LS rootlet growth.
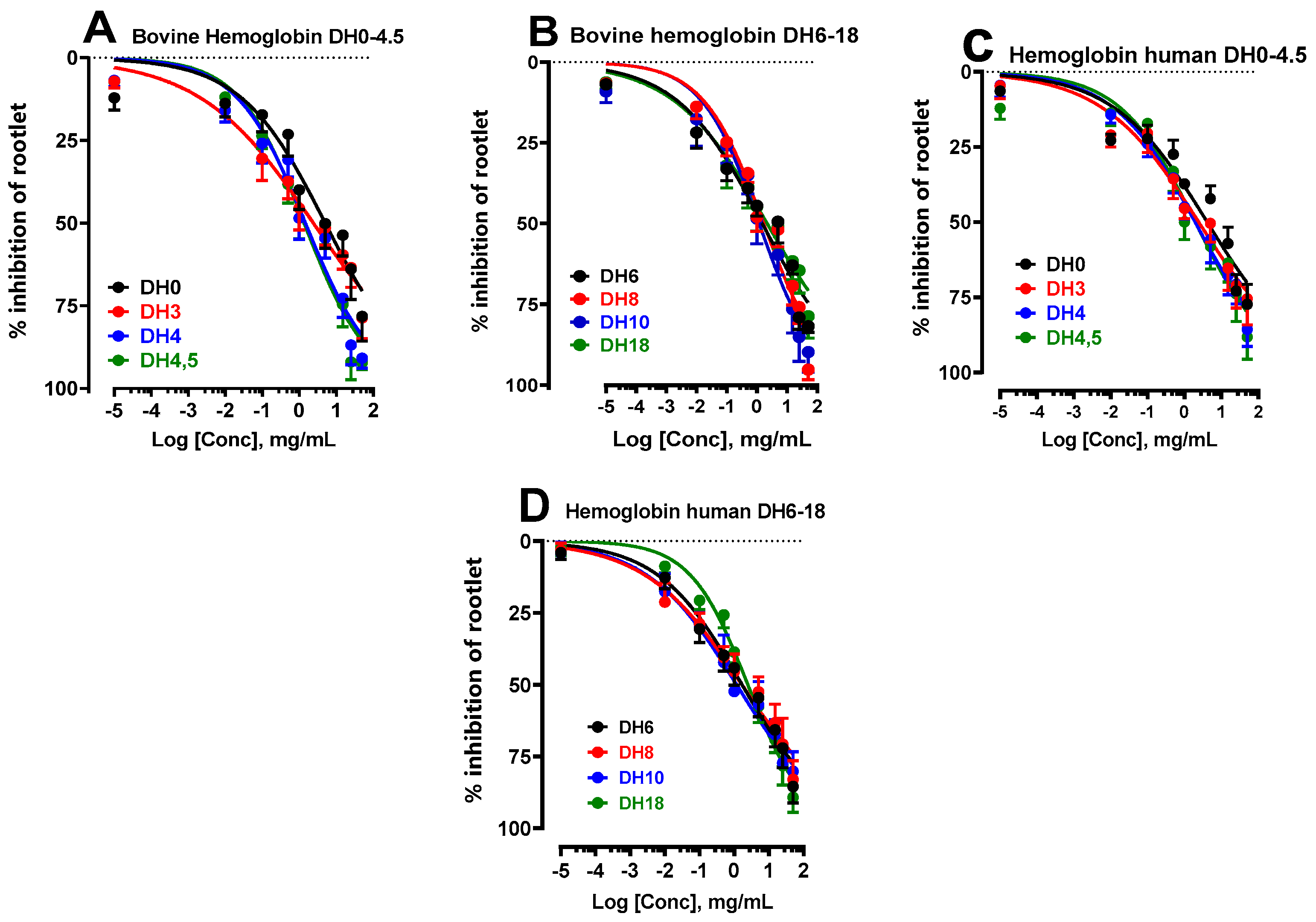
Figure 5.
Study of the antimitotic activity and peptidomic analysis of human and bovine hemoglobin hydrolysate fractions after a 3-hour hydrolysis period (pH 3.5, 23°C, E/S = 1/11, CBH = 10%, CHH= 10%, w/v). Fractions were collected every 5 minutes. (A) Chromatographic profiles of bovine and human hemoglobin hydrolysis acquired with Waters software at 215 nm by semi-preparative HPLC, analyzed by a semi-preparative C4 column. Effect of the influence of bovine (B-C) and human (D-E) hemoglobin hydrolysate fractions on the inhibition of LS rootlet growth. (F) Peptidomic analysis by UPLC-MS/MS and bioinformatics. Histogram showing the number of unique peptide sequences identified.
Figure 5.
Study of the antimitotic activity and peptidomic analysis of human and bovine hemoglobin hydrolysate fractions after a 3-hour hydrolysis period (pH 3.5, 23°C, E/S = 1/11, CBH = 10%, CHH= 10%, w/v). Fractions were collected every 5 minutes. (A) Chromatographic profiles of bovine and human hemoglobin hydrolysis acquired with Waters software at 215 nm by semi-preparative HPLC, analyzed by a semi-preparative C4 column. Effect of the influence of bovine (B-C) and human (D-E) hemoglobin hydrolysate fractions on the inhibition of LS rootlet growth. (F) Peptidomic analysis by UPLC-MS/MS and bioinformatics. Histogram showing the number of unique peptide sequences identified.
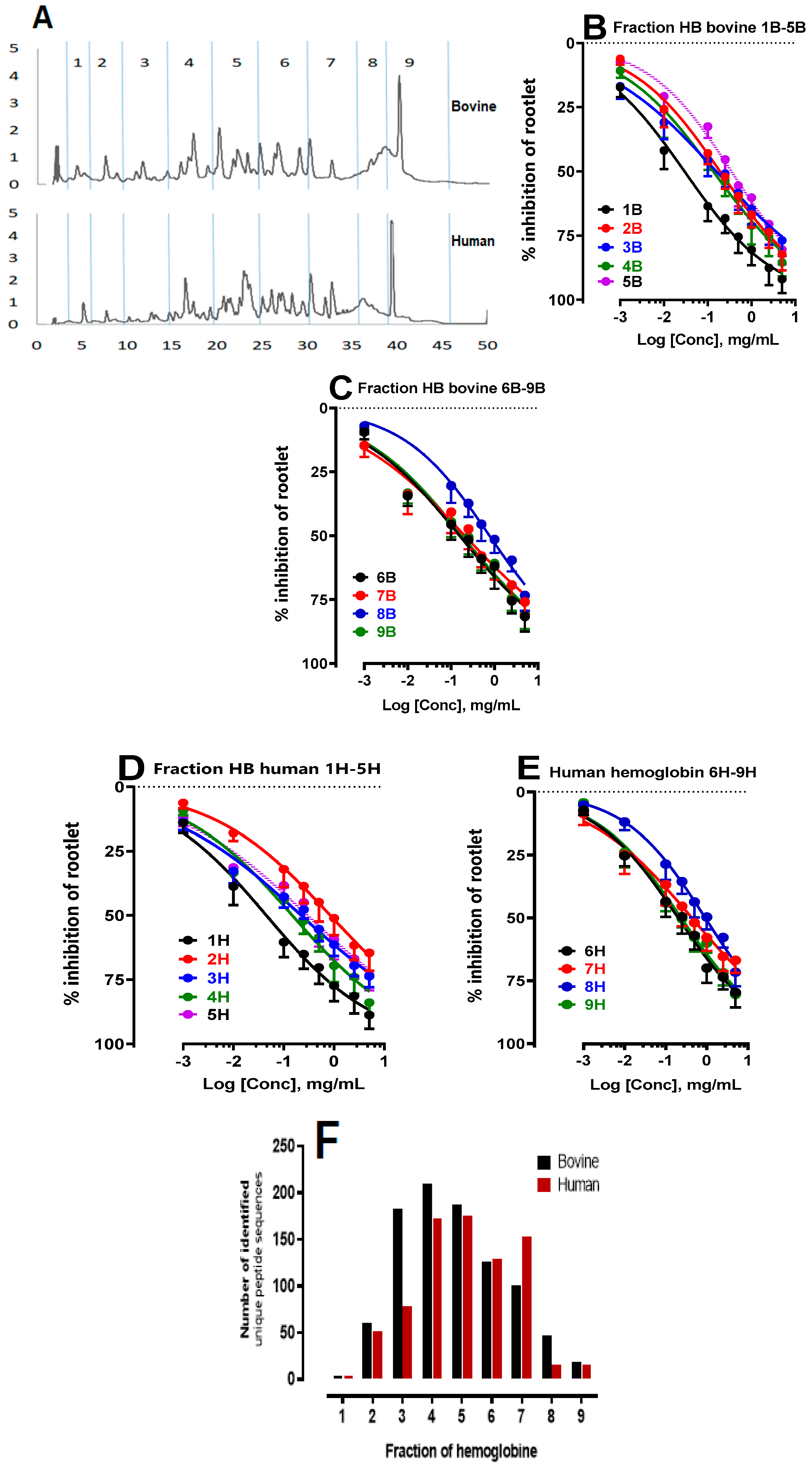
Figure 6.
Effect of the influence of standard peptides α 137-141 and α 1-32 on LS rootlet growth inhibition.
Figure 6.
Effect of the influence of standard peptides α 137-141 and α 1-32 on LS rootlet growth inhibition.
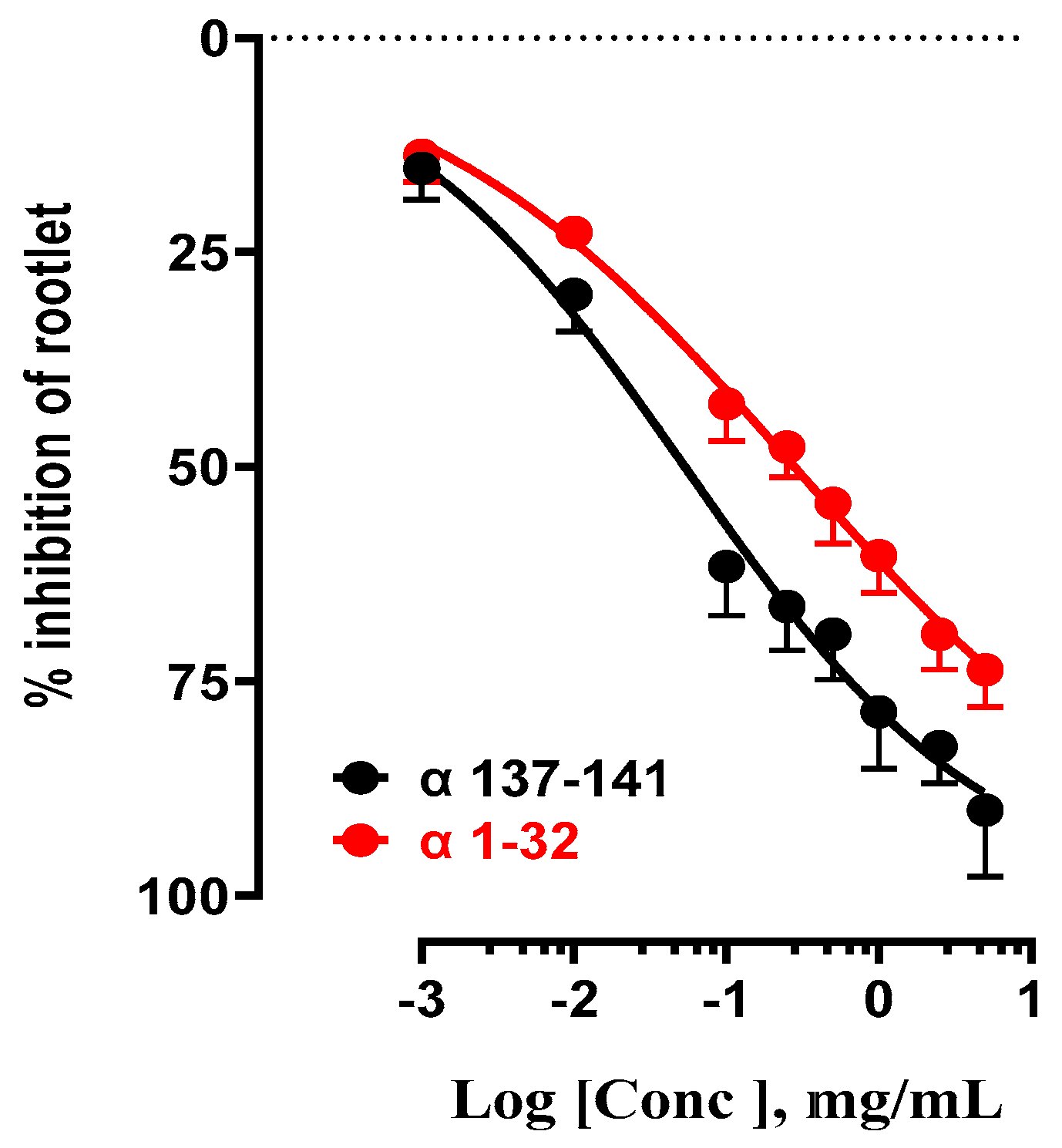
Figure 7.
Covalent labeling of Formylase by tRNAox in the absence and presence of UCK-36. Formylase (3µg/µL); tRNAox (36.5µg/µL) and 1 mM of the UCK-36 molecule.
Figure 7.
Covalent labeling of Formylase by tRNAox in the absence and presence of UCK-36. Formylase (3µg/µL); tRNAox (36.5µg/µL) and 1 mM of the UCK-36 molecule.
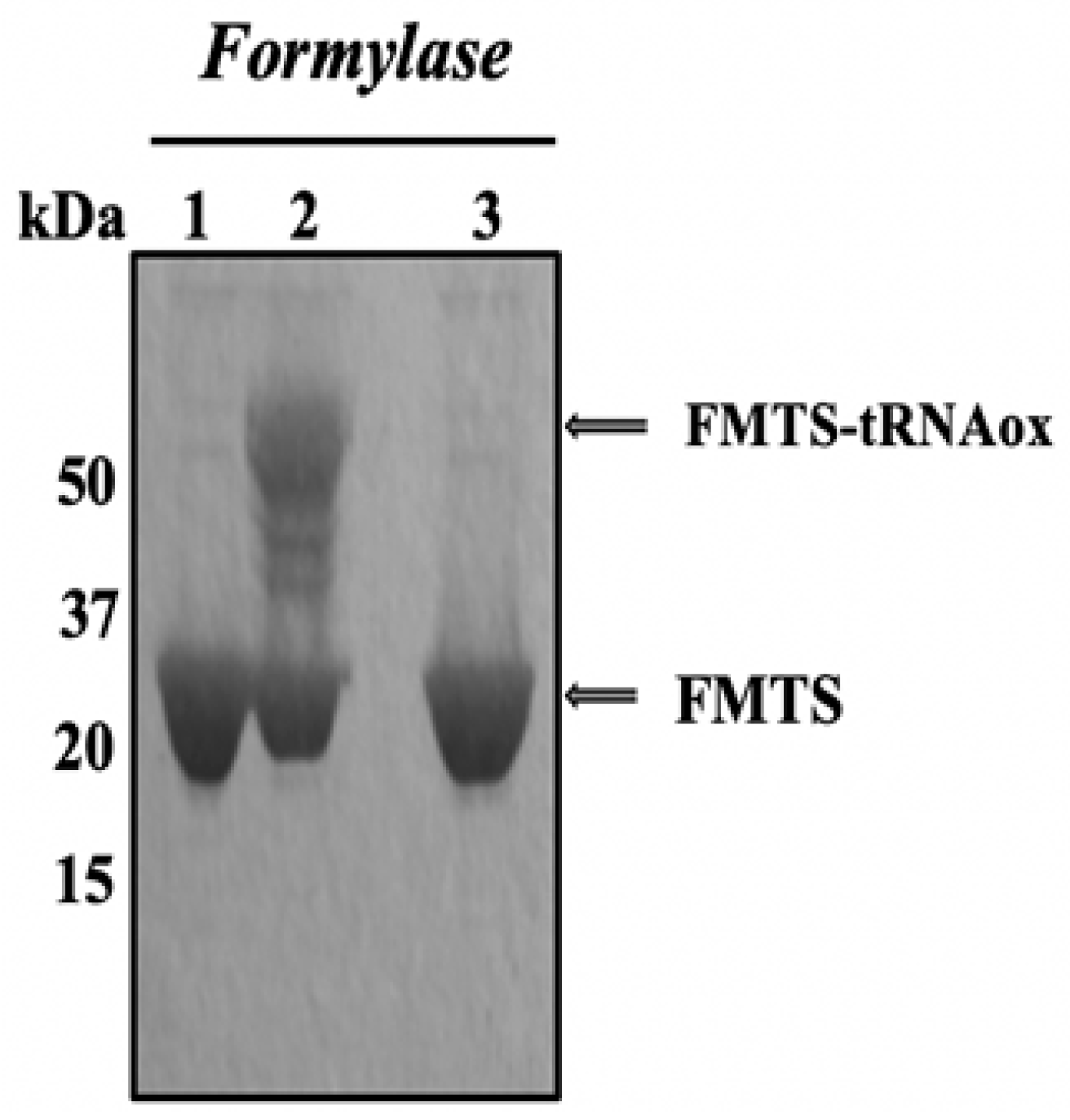
Figure 8.
Testing of bovine (A) and human (B) hemoglobin peptide hydrolysates and their peptide fractions. Analysis was carried out by electrophoresis on a 10% polyacrylamide gel under denaturing conditions (SDS-PAGE), revealed by Coomassie blue staining. The targeted protein is FMTs. 1: Marker (8 uL); 2: FMTs (0.3 uL); 3: FMTs (0.3 uL) + 6 uL of tRNAox; 4: FMTs (0.3 uL) + 6 uL of tRNAox + fraction 1 (NKT) (0.5 uL of 1/100); 5: FMTs (0.3 uL) + 6 uL of tRNAox + 2 (0.5 uL of 1/100); 6: FMTs (0.3 uL) + 6 uL of tRNAox + 3 (0.5 uL of 1/100); 7: FMTs (0.3 uL) + 6 uL of tRNAox + 4 (0.5 uL of 1/100) 8: FMTs (0.3 uL) + 6 uL of tRNAox + total hydrolysate (0.5 uL of 1/100)9: Marker (8 uL).
Figure 8.
Testing of bovine (A) and human (B) hemoglobin peptide hydrolysates and their peptide fractions. Analysis was carried out by electrophoresis on a 10% polyacrylamide gel under denaturing conditions (SDS-PAGE), revealed by Coomassie blue staining. The targeted protein is FMTs. 1: Marker (8 uL); 2: FMTs (0.3 uL); 3: FMTs (0.3 uL) + 6 uL of tRNAox; 4: FMTs (0.3 uL) + 6 uL of tRNAox + fraction 1 (NKT) (0.5 uL of 1/100); 5: FMTs (0.3 uL) + 6 uL of tRNAox + 2 (0.5 uL of 1/100); 6: FMTs (0.3 uL) + 6 uL of tRNAox + 3 (0.5 uL of 1/100); 7: FMTs (0.3 uL) + 6 uL of tRNAox + 4 (0.5 uL of 1/100) 8: FMTs (0.3 uL) + 6 uL of tRNAox + total hydrolysate (0.5 uL of 1/100)9: Marker (8 uL).
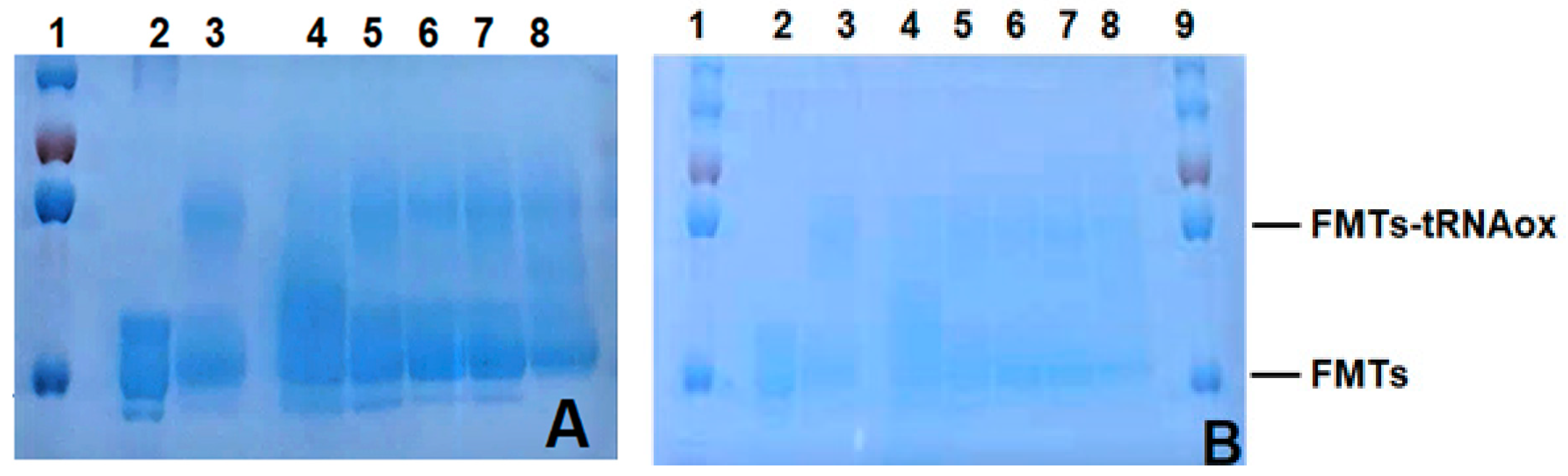
Figure 2.
Global tRNAox-Lys [Protein] labeling reaction.

Table 1.
represents the IC50 values of bovine and human hemoglobin hydrolysates according to the initial hemoglobin concentration (1, 2, 8 and 10%).
Table 1.
represents the IC50 values of bovine and human hemoglobin hydrolysates according to the initial hemoglobin concentration (1, 2, 8 and 10%).
| IC50 (mg/mL) | ||
| [HB] | Bovine (B) | Human (H) |
| 1 | 1.47 ± 0.65a | 2.12 ± 0.86a |
| 2 | 1.69 ± 0.70a | 1.80 ± 0.74a |
| 8 | 1.53 ± 0.61a | 2.24 ± 0.80a |
| 10 | 1.23 ± 0.52a | 1.77 ± 0.74a |
The IC50 values of the hydrolysates are compared with each other. The letters a and b are used to indicate significant differences between groups. Identical letters indicate no significant difference. Statistical significance was determined using the Bonferroni-Dunn method, with alpha = 0.05. Each row was analyzed individually.
Table 2.
Represent the IC50 values of bovine and human hemoglobin hydrolysates according to their DH.
Table 2.
Represent the IC50 values of bovine and human hemoglobin hydrolysates according to their DH.
| Hydrolysis | IC50 (mg/mL) | |
|---|---|---|
| DH | Bovine (B) | Human (H) |
| DH0 | 5.06 ± 2.00a | 3.97 ± 0.60a |
| DH3 | 2.42 ± 0.96a | 2.68 ± 0.67a |
| DH4 | 1.46 ± 0.59a | 1.94 ± 0.47a |
| DH4,5 | 1.35 ± 0.54a | 1.97 ± 0.84a |
| DH6 | 1.54 ± 0.60a | 1.68 ± 0.67a |
| DH8 | 1.68 ± 0.67a | 1.67 ± 0.65a |
| DH10 | 1.19 ± 0.47a | 1.17 ± 0.46a |
| DH18 | 2.25 ± 0.84a | 2.46 ± 1.06a |
The IC50 of the hydrolysates are compared with each other. The letters a and b are used to indicate significant differences between the groups. Identical letters indicate no significant difference. Statistical significance was determined using the Bonferroni-Dunn method, with alpha = 0.05. Each row was analyzed individually.
Table 3.
represents the IC50 values of bovine and human hemoglobin hydrolysates according to their DH.
Table 3.
represents the IC50 values of bovine and human hemoglobin hydrolysates according to their DH.
| Comparison of IC50 (mg/mL) | |||
|---|---|---|---|
| Bovine (B) | Human (H) | ||
| 1B | 0.029 ± 0.001a | 1H | 0.045 ± 0.002b |
| 2B | 0.19 ± 0.08a | 2H | 0.84 ± 0.35a |
| 3B | 0.15 ± 0.06a | 3H | 0.22 ± 0.09a |
| 4B | 0.13 ± 0.05a | 4H | 0.15 ± 0.06a |
| 5B | 0.34 ± 0.14a | 5H | 0.29 ± 0.11a |
| 6B | 0.20 ± 0.08a | 6H | 0.20 ± 0.08a |
| 7B | 0.43 ± 0.17a | 7H | 0.48 ± 0.19a |
| 8B | 0.92 ± 0.40a | 8H | 0.92 ± 0.40a |
| 9B | 0.23 ± 0.10a | 9H | 0.22 ± 0.09a |
The IC50 of the hydrolysates are compared with each other. The letters a and b are used to indicate significant differences between the groups. Identical letters indicate no significant difference. Statistical significance was determined using the Bonferroni-Dunn method, with alpha = 0.05. Each row was analyzed individually.
Table 4.
shows the IC50 the influence of standard peptides α 137-141 and α 1-32 on LS rootlet growth.
Table 4.
shows the IC50 the influence of standard peptides α 137-141 and α 1-32 on LS rootlet growth.
| Bioactifs peptides | IC50 (µg/mL) |
| α 137-141 | 53a |
| α 1-32 | 280 b |
The IC50s of the antimicrobial peptides are compared with each other. a and b are used to indicate significant differences between groups. Identical letters indicate no significant difference. Statistical significance was determined using the Bonferroni-Dunn method, with alpha = 0.05. Each row was analyzed individually.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



