Submitted:
01 September 2023
Posted:
05 September 2023
You are already at the latest version
Abstract
The globose scale (GS) (Sphaerolecanium prunastri Boyer de Fonscolombe) is a sucking insect that feeds on saps of wild apricot tree (Prunus armeniaca L.) in the Xinjiang Uygur Autonomous Region of northwestern China. It feeds on branches of wild apricot leading to poor growth, leaf yellowing and leaf drop, and sometimes mortality. Since the initial infestation in 2018, all the main valleys of wild apricot trees have been affected, but there is little research on the host’s physiological response to GS infestation. We measured the differences in growth between infested and non-infested wild apricots. The results showed that the diameter of shoot branches, the fresh weight, dry weight, length, width, area, and chlorophyll content of leaves, and the longitudinal diameter of fruit, were all significantly less for infested wild apricot trees than for un-infested wild apricot. The branch length of shoots, cross diameter, and weight of fruit also decreased, although the differences were not significant. Overall, GS infestation significantly reduced the growth of wild apricot trees, and may accelerate the damage by GS to wild apricot over time.
Keywords:
Sphaerolecanium prunastri (Boyer de Fonscolombe)
; Prunus armeniaca L.
; branch and leaf growth
; chlorophyll SPAD values
; fruit characteristics
1. Introduction
All kinds of biological and abiotic factors in the forest restrict and coordinate simultaneously, and determine a dynamic in the forest ecosystem. Among them, insects and plants are important components of forest ecosystem, and their interaction has a great influence on species diversity, which is related to forest productivity [1]. The change of insect community has a great influence on composition of forest ecosystem, and can directly or indirectly affect the species and quantity distribution of other species [2]. Increasing of insect populations, has a significant negative impact on forest ecosystems, reducing the growth of trees [3,4]. Studies have shown that insects feeding on plants can affect plant physiology and biochemistry such as chlorophyll content and photosynthesis, thus affecting growth of plants, and even lead to death of plants [5,6,7]. The relationship between insect feeding and plant growth has always been the focus of research on forest ecosystem health, as well as on insect tolerance of plants [8,9].
The wild apricot (Prunus armeniaca L.) is one of ancestors of the cultivated apricot in the world [10]. It is mainly distributed in the Tianshan and Himalayan regions of Central Asia, including the mountainous areas of northern and northeastern China, the northern and western Himalayas of India, and the Dzunggar and Zagara mountains of Central Asia [11,12]. It is also found in small quantities at high altitudes in Afghanistan and Pakistan [13]. It is an important species in the wild fruit forest of Yili, Tianshan, China, and also a relict species in Tertiary deciduous broad-leaved forest [14]. It is an important germplasm resource for protecting natural ecosystem and cultivating and improving cultivated apricot [13,15], and has been listed as a national key protected species at level Ⅱ and a key protected wild plant at the sub-level of Xinjiang priority protected species list in China [16]. However, with the influence of environmental change, human factors and pests, the area of wild fruit forest in Yili, Xinjiang has shrunk, the community has degenerated due to the difficulty in plant regeneration [17,18]. Among them, the survival of wild apricot is under serious threats from the piercing-sucking pest globose scale (GS) (Sphaerolecanium prunastri Boyer de Fonscolombe) (Hemiptera: Coccidae), a wax bug that sucks sap from the younger branches of a host plant.
GS is native to the subtropical areas of the Palearctic and now widely distributed in the Mediterranean, southern Europe, the Middle East, North America, China, and some areas of Central Asia and the Far East [19,20]. In China, GS has caused damages to drupes such as apricots and peaches in Liaoning, Hebei, Shandong and Shaanxi provinces [19, 21]. Since the 2018 , GS has been widespread in the wild apricot forests in the Yili River Valley. Its infestation has resulted in a large amount of dead branches and lead to some trees dead in the wild apricot forests, which has reduced the health and ecological services of the wild fruit forest ecosystem. It can be predicted that feeding of GS will slow down the growth of wild apricot trees, and the infestation of this scale also pose a great threat to the survival of wild apricot forests. At present, researches on GS mainly were focused on the biological characteristics and management including occurrence, damage, prevention and control techniques [22,23,24], but the relationship between the damage of GS and the growth of its host plants has not been reported. Here, effects of the damage of GS on the growth of its host wild apricot were studied. This study is helpful to reveal the relationship between GS and wild apricot, and to provide some theoretical basis for the study of the effects of the piercing-sucking insects on growth and health of plants.
2. Materials and Methods
2.1. Study site and treatment
Experiments were conducted at in kuerdening town in Gongliu county, Yili, China(43°14'53.41"N-43°15'25.40"N and 82°49'39.79"E-82°49'40.15"E). We investigate the impact of infestation by GS on status of branch and leaf growth and chlorophyll and fruit characters in wild apricot (Prunus armeniaca). The region is a typical temperate continental climate with alpine climate. The climate is mild and humid, large temperature difference between day and night, and the average annual temperature is 5-7 ℃. The rainfall concentrate in spring and summer, and the average annual precipitation ranges from 200 mm to 800 mm. The frost free period is about 150 days [17, 25]. Height of trees investigated are 3.5 m - 4.5 m, and base diameters are 6.5 cm - 8.5 cm. At the stage of female adult enlargement of S. prunastri on the 25 May, 2020, infested mature leafs was collected from wild apricot trees, and applied as above to control trees. 120 leafs collected from six the GS-infested trees and 60 leafs collected from three control wild apricot trees (There were very few healthy trees in the study area). The diameter of the fruit was measured at the wild apricot fruit ripening stage. Each treatment collected 30 fruits from six the GS-infested trees and three control wild apricot trees, respectively.
2.2. Shoot growth
For all treatments, the length and basal diameter of the current year branches were measured. There were 3 branches per tree. Measure the length from the base of the new shoot to the shoot with a steel tape. Measure the diameter 1 cm away from the base with digital electronic travel scale, cross measure twice, and take the average value.
2.3. Leaf traits
The leaf area of wild apricot was measured by portable YMJ-B living leaf area meter (Hangzhou Tuopu), and the leaf length and width were recorded. Each leaf was measured three times and the average value was taken.
2.4. Chlorophyll SPAD values
The chlorophyll SPAD value was measured by using the portable daily chlorophyll tester spad-502 plus (KONICA MINOLTA, Japan). Each leaf was tested 3 times in 6 different parts, including upper, middle and lower parts, and the average value was taken.
2.5. Leaf biomass
Use electronic balance (HUAZHI PTX-FA210, China, 0.0001g) to measure the fresh weight of the leafs. After the measurement, put it in the oven and dry it to constant weight at 80℃. Take it out and cool it. Weigh the dry weight immediately and calculate the average value.
2.6. Fruit traits
The vertical diameter, transverse diameter and lateral diameter of fruit were measured by digital electronic vernier caliper (0-300mm), and the fresh weight of single fruit was measured by electronic balance.
2.7. Data analysis
Microsoft Office Excel 2010 software was used to complete the data processing, SPSS 19.0 software was used to conduct Independent sample t-test, and origin 7.0 software was used to draw graphs. Prior to all analyses, in order to satisfy the hypothesis of normal distribution of data in statistical test, all the original data are processed after normal transformation. Independent sample t-test was used to analyze the difference between the treatment and the control. All analyses were done at a 5% significance level (p<0.05).
3. Results
3.1. Shoot growth
The branch length and branch diameter of GS-infested trees decreased by 17.92% and 16.61% compared with that control wild apricot, respectively. Although there was a trend for higher rates of branch length in GS trees than in control trees, there was no significant difference between treatments (P=0.202; Figure 1A). The branch diameter differed between treatments (P<0.01; Figure 1B), with lower rates in GS-infested than in control trees.
3.2. Leaf traits
GS-infested trees had extremely significantly lower relative leaf length and leaf width than control trees (P<0.01; Fig.2A, Figure 2B). GS-infested trees had extremely significantly lower relative leaf area than control trees, which we interpret as evidence of GS-induced wild apricot leaf loss (P<0.01; Figure 2C).
3.3. Chlorophyll SPAD values
The chlorophyll SPAD values of GS-infested trees decreased by 8.49% compared with that control wild apricot. GS-infested trees had extremely significantly lower chlorophyll SPAD values than control trees (P<0.01; Figure 3).
3.4. Leaf biomass
The leaf fresh weight and leaf dry weight of GS-infested trees decreased by 13.14% and 11.89% compared with that control wild apricot, respectively. GS-infested trees had extremely significantly lower leaf fresh weight and leaf dry weight than control trees (P<0.01; Figure 4A,B).
3.5. Fruit traits
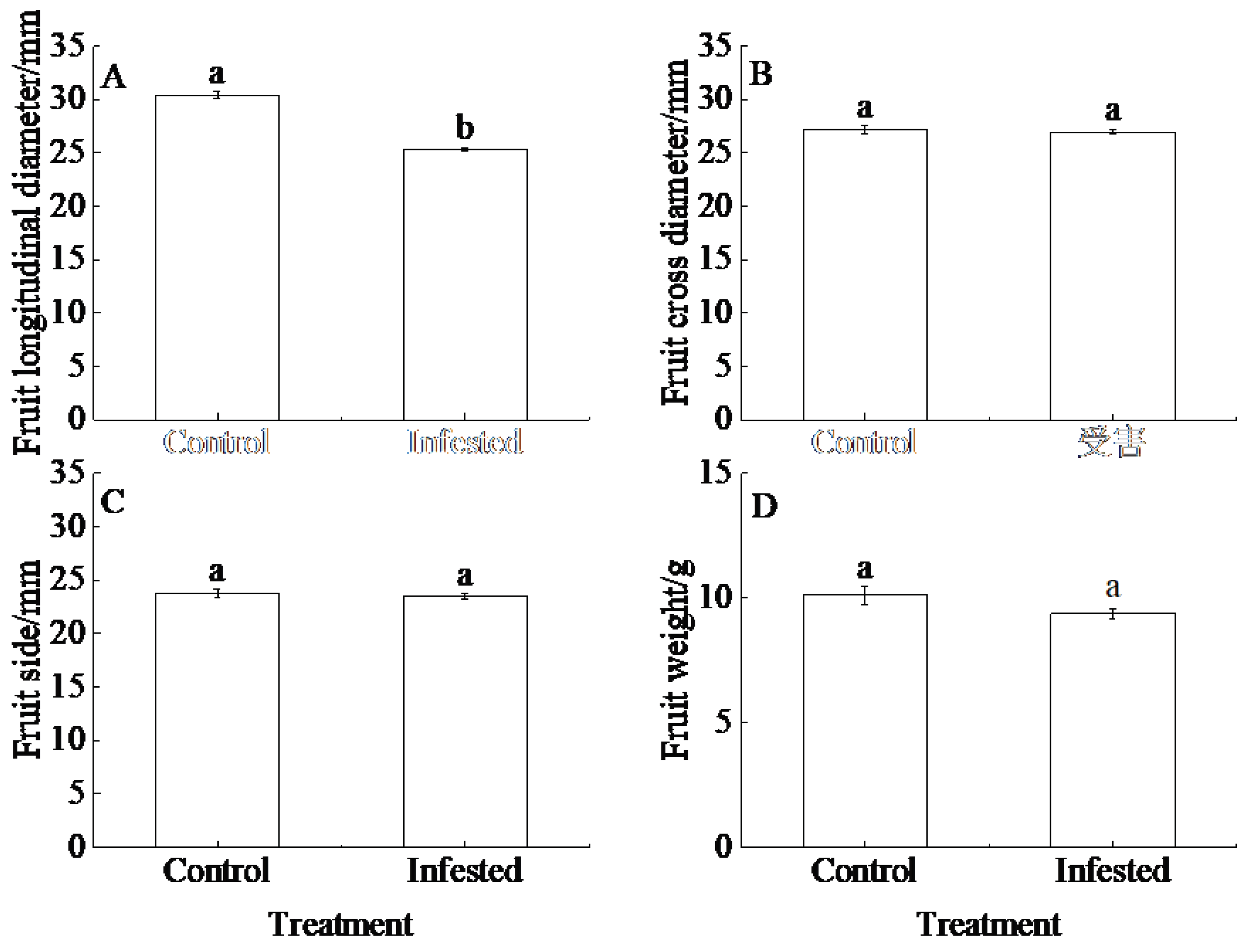
GS-infested trees had extremely significantly lower relative fruit longitudinal diameter than control trees (P<0.01; Figure 5A). Although there was a trend for higher rates of fruit cross diameter, fruit side and fruit weight in GS trees than in control trees, there was no significant difference between treatments (P=0.668, Figure 5B; P=0.530, Figure 5C; P=0.151, Figure 5D).
4. Discussion
4.1. Effects of the GS infection on growth of wild apricot
Insects can affect the overall physiological health, appearance and photosynthetic performance of plants, and thus change the species diversity and social and economic value of forest ecosystems. For example, bud moth larvae (Lasiognatha cellifera) infection reduced the leaf area, increased the amount of deciduous leafs, and decreased the amount of flowering in the mangrove ecosystem [3]. In the forests of Southern California, the invasion of goldspotted oak borer (Agrilus coxalis) resulted in the death of oak trees and reduced the landscape value of the forest [26]. Our study showed that GS had a significant adverse effect on the growth of wild apricot shoots and leafs. The growth of new shoot length and branch diameter of wild apricot decreased and the leaf area lost. The results indicated that GS had an inhibitory effect on the growth of wild apricot. Similar results have also been reported in date palm trees infected by Parlatoria blanchardii [27], Vitis vinifera infected by Parthenolecanium pruinosum [28] and Hibiscus rosa-sinensis infected by Phenacoccus solenopsis [29]. These studies suggest that GS infection may seriously damage the growth and leafs of wild apricot. GS infection leads to poor growth of wild apricot, decreased nutrients in the tree, weakened resistance to stress, and more vulnerable to GS or other diseases and insect pests [8]. This is a "vicious cycle" until the current host dies, and GS is transferred to the next host. It is not conducive to the survival of wild apricot, and reduces its economic or landscape value of forests.
4.2. Effect of GS infestation leaf traits of wild apricot
Leafs are the largest organ for plants to sense the environment, and their morphology is susceptible to the different ecological stress. Therefore, under pest stress, leaf morphology will also change, which will lead to leaf yellowing or even falling off. The leafs of Euonymus fortunei infected by euonymus scale (Unaspis euonymi) were susceptible to wither and fall off, and the transpiration rate of infected leafs decreased [30,31]. Leaf yellowing of E. fortunei was caused by the destruction of chloroplasts caused by euonymus scale, and the chlorophyll content of leafs decreased by 49% on average due to the feeding of two generations of scale [32]. After being fed by Toumeyella sp., the leaf area, leaf dry weight and chlorophyll content of Guaiacum sanctum were all lower than those of uninfected trees, leading to the decrease of tree growth [33]. Similar results were found for palm trees fed on Parlatoria blanchardii [27]. The same results have also been reported in other piercing-sucking pests, such as the feeding of adults of Bagrada hilaris significantly reduced the leaf area, relative chlorophyll content and dry weight of host plants [34]. Our study also found that the fresh and dry weight, leaf length and width, leaf area and relative chlorophyll content (chlorophyll SPAD value) of GS-infected wild apricot trees were significantly lower than those of uninfected trees, which was basically consistent with those results. The above studies could explain that the infection of GS was the cause of the leaf diminution, yellowing and even shedding of wild apricot. Evidence showed that deciduous leafs would inhibit the growth of branches in the tree body, and the survival rate of branches with fewer leafs was lower, which was easy to dry up and die [35], which could explain the large area of withered branches in the wild apricot at present.
4.3. Effect of GS infection on Photosynthetic products of wild apricot
Source-sink relationships of plants are relative and will change at different stages of plant development. In the stage of plant reproductive growth, the leaf is the “source” and the fruit is the main “sink” [36]. Pests can change the proportion of source-sink tissues in plants and affect photosynthesis [33]. Leafs are the main organs of photosynthesis and transpiration in green plants, and also have certain functions of absorption, reproduction and storage. In the study of the relationship between insect pest and plant growth, the change of chlorophyll content can reflect the damage level of the host plant [37]. Under pest stress, leaf size decreased, chlorophyll content decreased, and then photosynthesis decreased, and dry matter accumulation decreased [27,38,39]. After feeding on Solanum lycopersicum by Phenacoccus solenopsis [40], Mango shield scale (Kilifia acuminata) infected Mango [41], and Coccus hesperidum infected Citrus Limon and Nephrolepis Biserrata [42] later reported similar results. Scale insects disgreen plant leafs and ultimately reduce photosynthesis [43]. It has been shown that scale insects reduce photosynthesis by reducing the maximum rate of electron transport in plants [44]. Our research results also showed that the leaf area and relative chlorophyll content of wild apricot trees decreased significantly after GS infection, so it can be inferred that the photosynthesis of wild apricot trees also decreased. However, Retuerto et al. (2004) found that the photosynthetic rate of European Holly (Ilex aquifolium) was enhanced after being infected with Coccus sp., and its photosynthetic performance was improved, which was considered to be the compensatory effect of plants [45]. Plant compensation effect is common, but under the high density population or long harm, plant compensation effect is not enough to make up for the loss caused by plant hoppers, plants show serious victims [46,47,48]. This phenomenon can be clearly observed in the wild from wild apricot trees seriously infected with GS. And more and more studies have proved that herbivorous insects inhibit the photosynthesis of plants and reduce photosynthesis, while the compensatory enhancement of plant photosynthesis is rarely found [49]. The study of Velikova et al. (2010) also showed that both feeding and oviposition behaviors of herbivorous insects would inhibit the photosynthesis of host plants [50]. In addition, the wild apricot trees seriously infected by GS in the wild fruit forest have large areas of dead branches (fallen leafs), which will reduce the photosynthesis of the plant [51].
When the "source" of the plant changes, the corresponding "sink" will also be affected and changed. The response of plants to scale insect infestation can be explained by thinking of scale insects as additional pools of plants that compete with the plant's own absorption pools and consume nutrients while changing the distribution of plant nutrients [46]. There is some evidence that the production of honeydew by scale insects not only inhibits photosynthesis, but also consumes plant nutrients [52,53,54]. The fruit yield and quality of host plants decreased after being affected by scale insects [55], such as sugarcane plants (Saccharum officinarum) infected with Red-Striped soft scale insect (Pulvinaria tenuivalvata) [56], Mango plants infected by Icerya Seychellarum and Aonidiella aurantii [57,58], coconut trees infected by Aspidiotus rigidus [7]. Similar results have been reported for other stinger-sucking pests, such as a decline in the yield of Brassica carinata infested with Brevicoryne brassicae and Lipahis eyrsimi [59]. The decrease in leaf dry weight can also be used as an argument for the decrease in photosynthate accumulation after reduced photosynthesis in wild apricot trees, which is also a result of the decrease in leaf area [60,61]. Lu and Ge (1988) observed the mouth needles of Matsucoccus matsumurae and the slices of its harmful branches and found that the mouth organs of scale insects punctures the phloem cambium of plant branches and at the same time destroy xylem and tracheids [62]. They suggest that scale insects are not only declining nutrients of plants , they are also impeding the transport of water and nutrients in the tree itself, weakening growth and, in severe cases, killing plants. Our study also found that the fruit appearance and yield of wild apricot decreased after GS infection, which was basically consistent with the above research results. Similarly, the competition or distribution of nutrients by GS to wild apricot trees can also explain the decline in the growth of wild apricot branches and even the large number of dead branches of wild apricot trees [63].
4.4. Effect of GS infection on wild fruit forest
The growth and survival of wild apricot are threatened by GS, and the ecological health of wild fruit forest is also affected. The foliar damage, crown loss, mortality of host trees, and the composition, ecological function of forest were serious damaged due to the aggravation of insect occurring [64, 65]. Similar studies have been reported on wild apples (Malus sieversii) [66] and mangroves in South China [3], and eastern hemlock (Tsuga canadensis) in Pelham, Massachusetts, USA [67]. Compared with mechanical damage, herbivorous insects caused a greater reduction in physiological processing of plants [68]. As opposed to the leaf-eaters, pests with sap-sucking or stem feeding have more adverse effects on host plants [67,69,70]. But no matter what kind of herbivorous insect, its damage will have an impact on growth and reproduction of the host plant, and then on the ecosystem, and for the most cases, this effect is negative to ecosystem [71,72]. Delucia et al. (2012) also believed that effects of herbivorous insects on host plants can unbalance ecosystem [73]. Liu et al. (2010) pointed out that the mobility of herbivorous insects enables their damage ability to spread to a wider ranges [74]. Ancheta and Heard (2011) reviewed 37 articles on the relationship between herbivorous insects and rare plants, and they pointed out that herbivorous insects would greatly reduce the survival ability and reproduction, and even seriously cause the extinction of some species [75]. Myers and Sarfeaz (2016) also reported similar review results [76]. From our results, we can also draw the same conclusion that GS infected wild apricot has a negative impact on the growth and reproduction of wild apricot, which is not conducive to the survival of wild apricot trees, destroys the ecological balance of wild fruit forest, and finally has a negative impact on the ecosystem of wild fruit forest. Domec et al. (2013) found that the death mechanism of eastern hemlock was the decrease of water use and carbon assimilation caused by hemlock woolly adelgid (Adelges tsugae) infection [77]. Haavik et al. (2015) suggested that carbon imbalance is a potential mechanism for the death of declining oaks infected by pests and diseases [78]. However, the mechanism of GS infection leading to the death of wild apricot trees needs further study. However, it is predictable that a large number of wild apricot branches or tree deaths caused by GS will lead to the decline of apricot population [46], which also aggravates the imbalance of wild fruit forest ecosystem.
5. Conclusions
In general, our results showed that after GS infection, the growth status of branches and leafs, relative chlorophyll content and fruit quality of wild apricot trees decreased significantly. These physiological deterioration are helpful to elucidate the mechanism of GS-induced leaf shrinkage, leaf wither and body death of wild apricot trees. It is suggested to carry out long-term monitoring of GS in wild fruit forests, from GS infestation to the mortality of wild apricot trees, from wild apricot individual plant to wild apricot population and the ecosystem, so as to better clarify the mechanism of death of trees and ecosystem collapsing induced by GS.
Author Contributions
Conceptualization, G.G. and L.Z. designed this study; L.W. and W. Y. collected the data; L.W. and G.G. wrote the first draft of the manuscript. All authors have read and agree to the published version of the manuscript.
Funding
This research was funded by the Central finance forest and grass science and technology popularization demonstration project, grant number Xin 2020TG09 and Tianshan Talents Program of Xinjiang Uygur Autonomous Region.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Liang, J.J. ; Crowther, T.W.; Picard, N.; Wiser, S.K.; Zhou, M.; Alberti, G.; Schulze, E.D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; de-Miguel, S.; Paquette, A.; Hérault, B.; et al. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354(6309), aaf8957. [CrossRef]
- Zhang, H.Y.; Ou, X.H. Using insect for indicator to monitor and assess forest ecosystm health. World Forestry Res. 2006, 19(4), 22-25.
- Lu, W. Z.; Xiao, J. F.; Cui, X.W.; Xu, F.H.; Lin, G.X.; Lin, G.H. Insect occurs have transient effects on carbon fluxes and vegetative growth but longer-term impacts on reproductive growth in a mangrove forest. Agr. Forest Meteorol. 2019, 279(4), 107747. [CrossRef]
- Michael, A. Are changes in plants due to enhanced CO2 contributing to insect population declines? Environ. Entomol. 2019, 48(2), 274-275. [CrossRef]
- Smith, J.P.; Schowalter, T.D. Aphid-induced reduction of shoot and root growth in Douglas-fir seedlings. Ecol. Entomol. 2010, 26(4), 411-416. [CrossRef]
- Rubino, L.; Charles, S.; Sirulnik, A.G.; Tuininga, A.R.; Lewis, J.D. Invasive insect effects on nitrogen cycling and host physiology are not tightly linked. Tree Physiol. 2015, 35(2), 124–133. [CrossRef]
- Watson, G.W.; Adalla, C.B.; Shepard, B.M.; Carner, G.R. Aspidiotus rigidus Reyne (Hemiptera: Diaspididae): A devastating pest of coconut in the Philippines. Agr. Forest Ento. 2015, 17(1), 1-8. [CrossRef]
- Han, P.; Becker, C.; Bot, J.L.; Larbat, R.; Lavoir, A.V.; Desneux, N. Plant nutrient supply alters the magnitude of indirect interactions between insect herbivores: From foliar chemistry to community dynamics. J. Ecol. 2019, 108(4), 1497-1510. [CrossRef]
- You, Y.R.; An, C.P.; Li, C.Y. Insect feeding assays with Spodoptera exigua on Arabidopsis thaliana. Bio-protocol 2020, 10(5), e3538. [CrossRef]
- Li, W.W.; Liu, L.Q.; Wang, Y.N.; Zhang, Q.P.; Fan, G.Q.; Zhang, S.K.; Wang, Y.T.; Liao, K. Genetic diversity, population structure, and relationships of apricot (Prunus) based on restriction site-associated DNA sequencing. Hortic. Res. 2020, 7(1), 69-82. [CrossRef]
- Zeven, A.C.; Zhukovsky, P.M. Dictionary of cultivated plants and their centres of diversity: Excluding ornamentals, forest trees and lower plants. Wageningen: Centre for Agricultural Publishing and Documentation, 1975, 36.
- Zhebentyayeva, T.; Reighard, G.; Gorina, V.; Abbott, A. Simple sequence repeat (SSR) analysis for assessment of genetic variability in apricot germplasm. Theor. Appl. Genet. 2003, 106(3), 435-444. [CrossRef]
- Malik, S.K.; Chaudhury, R.; Dhariwal, O.P.; Mir, S. Genetic diversity and traditional uses of wild apricot (Prunus armeniaca L.) in high-altitude north-western Himalayas of India. Plant Genet. Resour. 2010, 8(3), 249-257. [CrossRef]
- Zhang, X.S. On the eco-geographical characters and the problems of classification of the wild fruit-tree forest in the Ili Valley of Sinkiang. Acta Bot. Sin. 1973, 15(2), 239-253.
- Khadivi-Khub, A.; Khalili, Z. A breeding project: The selection of promising apricot (Prunus armeniaca L.) genotypes with late blooming time and high fruit quality. Sci. Hortic. 2017, 216, 93-102. [CrossRef]
- Liu, W.; Chen, X.S.; Liu, G.J.; Liang, Q.; He, T.M.; Feng, J.R. Interspecific hybridization of Prunus persica with P. armeniaca and P. salicina using embryo rescue. Plant Cell Tis. Org. 2007, 88(3), 289-299. [CrossRef]
- Hou, B.; Xu, Z. Relation ship of the occurences and evolutions of wild-fruit forests with climatic factors in the Tinshan Mountain. Acta Bot. Boreali-Occidentalia Sin. 2005, 25(11), 2266-2271.
- Fang, Z.Y.; Li, L.Y.; Maola, A.; Zhou, L.; Lu, B. Effects of human disturbance on plant diversity of wild fruit forests in western Tianshan Mountain. Bull. Soil Water Conserv. 2019, 39(2), 267-274.
- Wang, Z.Q. Zootaxa of China. Insecta XXII. Beijing: Science Press, 2001, 237-243.
- Malumphy, C.; Anderson, H. Plant pest factsheet: Globose scale, Sphaerolecanium prunastri[EB/OL]. https://planthealthportal.defra.gov.uk/assets/factsheets/Plant-Pest-Factsheet-S.-prunastri-June-2016v3.pdf, 2016-09-28/2021.04.03.
- Wang, C. Investigation on the species of Coccaidea from Shaanxi garden (Hemiptera: Coccoidea)[M]. Yang Ling: Northwest A & F University, 2010, 21.
- Karaca, I.; Japoshvili, G.; Demirozer, O. The chalcid parasitoid complex (Hymenoptera: Chalcidoidea) associated with the globose scale (Sphaerolecanium prunastri Fonscolombe) (Hemiptera: Coccoidea) in Isparta Province, Turkey and some east European countries. J. Plant Dis. Prot. 2003, 110(5), 505-511. [CrossRef]
- Aksit, T.; Apak, F.K. Effects of some insecticides with infestation rate and some biological characteristics of Sphaerolecanium prunastri (Fonscolombe, 1834) (Hemiptera: Coccidae) on plum. Turk. J. Entom. 2013, 37(1), 133-144.
- Yiğit, T.; Tunaz, H. Determination of Sphaerolecanium prunastri Fonscolombe, 1834 ( Hemiptera: Coccidae ), and its parasitoids and predators in apricot areas of Malatya province, Turkey. Acta Hortic. 2018, 1214, 67-71. [CrossRef]
- Yang, Y.H.; Chen, Y.N.; Li, W.H.; Chen, Y.P. Distribution of soil organic carbon under different vegetation zones in the Ili River Valley, Xinjiang. J. Geogr. Sci. 2010, 20(5), 729-740. [CrossRef]
- Coleman, T.W.; Seybold, S. J. Previously unrecorded damage to oak, Quercus spp., in southern California by the goldspotted oak borer, Agrilus coxalis Waterhouse (Coleoptera: Buprestidae). The Pan-Pac. Entomol. 2008, 84(4), 288-300. [CrossRef]
- Moussa S, Salman A, Bakry M. The negative effects of Parlatoria blanchardii (Targ.) infestation on the morphological and chemical characters of certain varieties leaflets of date palm trees at Luxor governorate. Egypt. Acad. J. Biolog. Sci., 2012, 5(1), 169-181.
- Simbiken, N.A.; Cooper, P.D.; Powell, K.S. Development and feeding effect of frosted scale Parthenolecanium pruinosum Cocquillet (Hemiptera: Coccidae) on selected Vitis vinifera L. cultivars. Aust. J. Grape Wine R. 2015, 21(3), 451-457. [CrossRef]
- Sun, F.; Ye, W.F.; Pan, Z.P.; Lu, Y.Y. Effect on population density of Phenacoccus solenopsis Tinsley on the growth of Hibiscus rosa-sinensis. J. Environ. Entomol. 2021, 43(1), 60-65.
- Cockfield, S.D.; Potter, D.A. Interaction of Euonymus scale (Homoptera: Diaspididae) feeding damage and severe water stress on leaf abscission and growth of Euonymus fortunei. Oecologia, 1986, 71(1), 41-46. [CrossRef]
- Cockfield, S.D.; Potter, D.A. Euonymus scale (Homoptera: Diaspididae) effects on plant growth and leaf abscission and implications for differential site selection by male and female scales. J. Econ. Entomol. 1990, 83(3), 995-1001. [CrossRef]
- Cockfield, S.D.; Potter, D.A.; Houtz, R.L. Chlorosis and reduced photosynthetic CO2 assimilation of Euonymus fortunei infested with Euonynlus scale (Homoptera: Diaspididae). Environ. Entomol. 1987, 16(3), 1314-1318. [CrossRef]
- Schaffer, B.; Mason, L.J. Effects of scale insect herbivory and shading on net gas exchange and growth of a subtropical tree species (Guaiacum sanctum L.). Oecologia, 1990, 84(4), 468-473. [CrossRef]
- Huang, T.I.; Reed, D.A.; Perring, T.M.; Palumbo, J.C. Feeding damage by Bagrada hilaris (Hemiptera: Pentatomidae) and impact on growth and chlorophyll content of Brassicaceous plant species. Arthropod-Plant Inte. 2014, 8(2), 89-100. [CrossRef]
- Canelo, T.; Gaytán, Á.; González-Bornay, G.; Bonal, R. Seed loss before seed predation: Experimental evidence of the negative effects of leaf feeding insects on acorn production. Integr. Zool. 2018, 13(3), 238-250. [CrossRef]
- Proietti, P. Effect of fruiting on leaf gas Exchange in Olive (Olea Europaea L.). Photosynthetica, 2000, 38(3), 397-402. [CrossRef]
- Hura, T.; Hura, K.; Grzesiak, M.; Rzepka, A. Effect of long-term drought stress on leaf gas exchange and fluorescence parameters in C3 and C4 plants. Acta Physiol. Plant. 2007, 29(2), 103-113. [CrossRef]
- Sharma, S.; Kumar, H.; Prasad, F.M.; Singh, K. Chlorophyll and carotenoid loss in Psidium guajva caused by Abgrallaspis cyanophylli. Ann. Plant Protect. Sci. 2009, 17(1), 245-246.
- Goławska, S.; Krzyżanowski, R.; Łukasik, I. Relationship between aphid infestation and chlorophyll content in fabaceae species. Acta Biol. Cracov. Bot. 2010, 52(2), 76-80. [CrossRef]
- Huang, J.; Zhang, P.J.; Zhang, J.; Lu, Y.B.; Huang, F.; Li, M.J. Chlorophyll content and chlorophyll fluorescence in tomato leafs tnfested with an invasive mealybug, Phenacoccus solenopsis (Hemiptera: Pseudococcidae). Environ. Entomol. 2013, 42(5): 973-979. [CrossRef]
- Nabil, H.A. Relationship Between Kilifia acuminata (Signoret) and Chlorophyll Percentage Loss on Mango leafs. J. Entomol. 2013, 10(2): 110-114. [CrossRef]
- Golan, K.; Rubinowska, K.; Kmiec, K.; Kot, I.; Gorska-Drabik, E.; Lagowska, B.; Michalek, W. Impact of scale insect infestation on the content of photosynthetic pigments and chlorophyll fluorescence in two host plant species. Arthropod-Plant Inte. 2015, 9(1), 55-65. [CrossRef]
- Walstad, J.D.; Nielsen, D.G.; Johnson, N.E. Effect of the pine needle scale on photosynthesis of Scots pine. Forest Sci. 1973, 19(2), 109-111.
- Moore, G.W.; Watts, D.A.; Goolsby, J.A. Ecophysiological responses of giant reed (Arundo donax) to herbivory. Invas. Plant Sci. Mana. 2010, 3(4), 521-529. [CrossRef]
- Retuerto, R.; Fernandez-Lema, B.; Rodriguez-Roiloa, Obeso, J.R. Increased photosynthetic performance in holly trees infested by scale insects. Funct. Ecol. 2004, 18(5), 664-669. [CrossRef]
- Vranjic, J.O.; Ash, J.E. Scale insects consistently affect roots more than shoots: The impact of infestation size on growth of eucalypt seedlings. J. Ecol. 1997, 85(2), 143-149. [CrossRef]
- Garcia, L.C.; Eubanks, M.D. Overcompensation for insect herbivory: A review and meta-analysis of the evidence. Ecology, 2019, 100(3), 2585-2625. [CrossRef]
- Nordkvist, M.; Klapwijk, M.J.; Edenius, L.R.; Björkman, C. Interacting effects of insect and ungulate herbivory on Scots pine growth. Sci. Rep. 2020, 10(1): 1-11. [CrossRef]
- Kerchev, P.I.; Fenton, B.; Foyer, C.H.; Hancock, R.D. Plant responses to insect herbivory: Interactions between photosynthesis, reactive oxygen species and hormonal signalling pathways. Plant Cell Environ. 2012, 35(2), 441-453. [CrossRef]
- Velikova, V.; Salerno, G.; Frati, F.; Peri, E.; Conti, E.; Colazza, S.; Loreto, F. Influence of feeding and oviposition by phytophagous pentatomids on photosynthesis of herbaceous plants. J. Chem. Ecol. 2010, 36(6), 629-641. [CrossRef]
- Delaney, K.J.; Haile, F.J.; Peterson, R.K.D.; Higley, L.G. Seasonal patterns of leaf photosynthesis after insect herbivory on Common Milkweed, Asclepias syriaca: Reflection of a physiological cost of reproduction, not defense? Am. Midl. Nat. 2009, 162(2), 224-238. [CrossRef]
- Dungan, R.J.; Kelly, D.; Turnbull, M. Separating host-tree and environmental determinants of honeydew production by Ultracoelostoma scale insects in a Nothofagus forest. Ecol. Entomol. 2007a, 32(4), 338-348. [CrossRef]
- Dungan, R.J.; Turnbull, M.H.; Kelly, D. The carbon costs for host trees of a phloem-feeding herbivore. J. Ecol. 2007b, 95(4), 603-613. [CrossRef]
- Miezite, O.; Okmanis, M.; Indriksons, A.; Indriksons, A.; Ruba, J.; Polmanis, L.; Freimane, L. Assessment of sanitary conditions in stands of Norway spruce (Picea abies Karst.) damaged by spruce bud scale (Physokermes piceae Schrnk.). iForest. 2013, 6(2), 73-78. [CrossRef]
- Martins, D.S.; Fornazier, M.J.; Culik, M.P.; Ventura, J.A.; Ferreira, P.S.F.; Zanuncio, J.C. Scale insect (Hemiptera: Coccoidea) pests of Papaya (Carica papaya) in Brazil. Ann. Entomol. Soc. Am. 2014, 107(8), 1-8. [CrossRef]
- Dimetry, N.; Abdel-Moniem, A. Physical and chemical variability in sugarcane fields infested with the red-striped soft scale insect, Pulvinaria Tenuivalvata(Newstead). Arch.Phytopathol. Plant Protect. 2004, 37(4), 327-337. [CrossRef]
- Salman, A.M.A.; Bakry, M.M.S. Relationship between the rate of infestation with the Mealybug, Icerya Seychellarum (Westwood) (Margarodidae: Homoptera) and the yield loss of seedy Balady Mango trees at Luxor governorate. World Rural Observ. 2012, 4(4), 50-56.
- Bakry, M.M.S.; Mohamed, G.H. Relationship between the rates of infestation with the California red scale insect, Aonidiella aurantii (Mask.) (Hemiptera: Diaspidae) and the yield loss of mango trees at Luxor governorate, Egypt. Egypt. J. Agric. Res., 2015, 93(3), 41-60.
- Hussain, A.; Razaq, M.; Zaka, S.M.; Shahzad, W.; Mahmood, K. Effect of aphid infestation on photosynthesis, growth and yield of Brassica carinata A. Braun. Pak. J. Zool. 2015, 47(5), 1335-1340.
- May, B.M.; Carlyle, J.C. Effect of defoliation associated with Essigella californica on growth of mid-rotation Pinus radiata. Forest Ecol. Manag. 2003, 183(1), 297-312. [CrossRef]
- Mustafa, F. Switchgrass (Panicum virgatum L.) yield and photosynthetic response to simulated insect defoliation and Grasshopper (Orthoptera: Acrididae) feeding. Nebraska: The University of Nebraska - Lincoln, 2013, 85-98.
- Lu, Q.H.; Ge, Z.H. Observation on the stylet and the parasitical site segment of Japanese pine bast scale. Forest Res. 1988, 1(2), 191-194.
- Miller-Pierce M.R.; Orwig, D.A.; Preisser, E. Effects of hemlock woolly adelgid and elongate hemlock scale on eastern hemlock growth and foliar chemistry. Environ. Entomol. 2010, 39(2), 513-519. [CrossRef]
- Orwig, D.A.; Thompson, J.R.; Povak, N.A.; Manner, M.; Niebyl, D, Forter, D.R. A foundation tree at the precipice: Tsuga canadensis health after the arrival of Adelges tsugae in central New England. Ecosphere, 2012, 3(1), 1-16. [CrossRef]
- Maclean, D.A. Impacts of insect occurs on tree mortality, productivity, and stand development. Can. Entomol. 2015, 148(1). 1-22.
- Zhang, P,; Cui, Z.J.; Xu, H.; Ali, A; Zhang, X.; Liu, X.X.; Zhang, Y.M.; Zhou, X.B.; Lu, Z.Z. Thirst or malnutrition: The impacts of invasive insect Agrilus mali on the physiological status of wild apple trees. Forests, 2020, 11(440), 1-10. [CrossRef]
- Radville, L.; Chaves, A.; Preisser, E.L. Variation in plant defense against invasive herbivores: Evidence for a hypersensitive response in eastern hemlocks (Tsuga canadensis). J. Chem. Ecol. 2011, 37(6), 592-597. [CrossRef]
- Visakorpi, K.; Gripenberg, S.; Malhi, Y.; Bolas, C.; Oliveras, I.; Harris, N.; Rifai, S.; Riutta, T. Small-scale indirect plant responses to insect herbivory could have major impacts on canopy photosynthesis and isoprene emission. New Phytol. 2018, 220(3), 799-810. [CrossRef]
- Zvereva, E.L.; Lanta, V.; Kozlov, M.V. Effects of sap-feeding insect herbivores on growth and reproduction of woody plants: A meta-analysis of experimental studies. Oecologia, 2010, 163(4), 949-960. [CrossRef]
- Stephens, A.E.A.; Westoby, M. Effects of insect attack to stems on plant survival, growth, reproduction and photosynthesis. OIKOS, 2014, 124(3), 266-273. [CrossRef]
- Takahashi, M.; Huntly, N. Herbivorous insects reduce growth and reproduction of big sagebrush (Artemisia tridentata). Arthropod-Plant Inte. 2010, 4(4), 257-266. [CrossRef]
- Halitschke, R.; Hamilton, J.G.; Kessler, A. Herbivore-specific elicitation of photosynthesis by mirid bug salivary secretions in the wild tobacco Nicotiana attenuata. New Phytologist, 2011, 191(2), 528-535. [CrossRef]
- DeLucia, E.H.; Nabity, P.D.; Zavala, J.A.; Berenbaum, M.R. Climate change: Resetting plant-insect interactions. Plant Physiol. 2012, 160(4), 1677-1685. [CrossRef]
- Liu, Z.G.; Cai, Y.L.; Fang, Y.; Jing, J.; Li, K. Induced response in Schima superba: Effects of early-season herbivory on leaf traits and subsequent insect attack. Afr. J. Biotechnol. 2010, 9(51), 8731-8738.
- Ancheta, J.; Heard, S.B. Impacts of insect herbivores on rare plant populations. Biol. Conserv. 2011, 144(10), 2395-2402. [CrossRef]
- Myers, J.H.; Sarfraz, R.M. Impacts of insect herbivores on plant populations. Annu. Rev. Entomol. 2016, 62(1), 207-230. [CrossRef]
- Domec, J.C.; Rivera, L.N.; King, J.S.; Peszlen, I.; Hain, F.; Smith, B.; Frampton, J. Hemlock woolly adelgid (Adelges tsugae) infestation affects water and carbon relations of eastern hemlock (Tsuga canadensis) and Carolina hemlock (Tsuga caroliniana). New Phytol. 2013, 199(2), 452-463.
- Haavik, L.J.; Billings, S.A.; Guldin, J.M.; Stephen, F.M. Emergent insects, pathogens and drought shape changing patterns in oak decline in North America and Europe. Forest Ecol. Manag. 2015, 354, 190-205. [CrossRef]
Figure 1.
Treatment differences in (A) branch length and (B) branch diameter of Prunus armeniaca infested with Sphaerolecanium prunastri (infested) and uninfested controls. Different lowercase letters are significantly different according to an independent two-sample t-test (P<0.05).
Figure 1.
Treatment differences in (A) branch length and (B) branch diameter of Prunus armeniaca infested with Sphaerolecanium prunastri (infested) and uninfested controls. Different lowercase letters are significantly different according to an independent two-sample t-test (P<0.05).
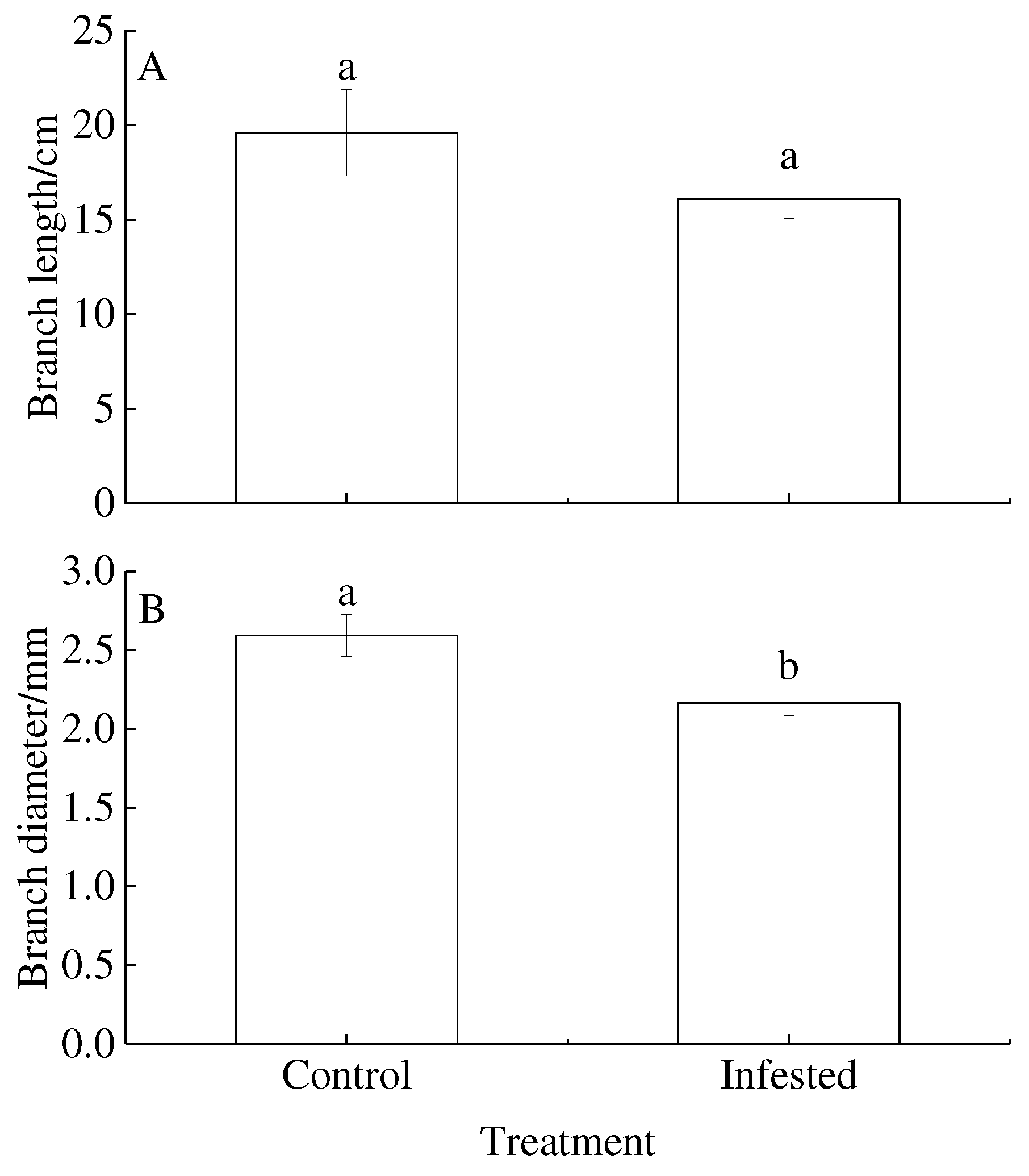
Figure 2.
Treatment differences in (A) leaf length, (B) leaf width and (C) leaf area of Prunus armeniaca infested with Sphaerolecanium prunastri (infested) and non-infested controls. Different lowercase letters are significantly different according to an independent two-sample t-test (P<0.05).
Figure 2.
Treatment differences in (A) leaf length, (B) leaf width and (C) leaf area of Prunus armeniaca infested with Sphaerolecanium prunastri (infested) and non-infested controls. Different lowercase letters are significantly different according to an independent two-sample t-test (P<0.05).
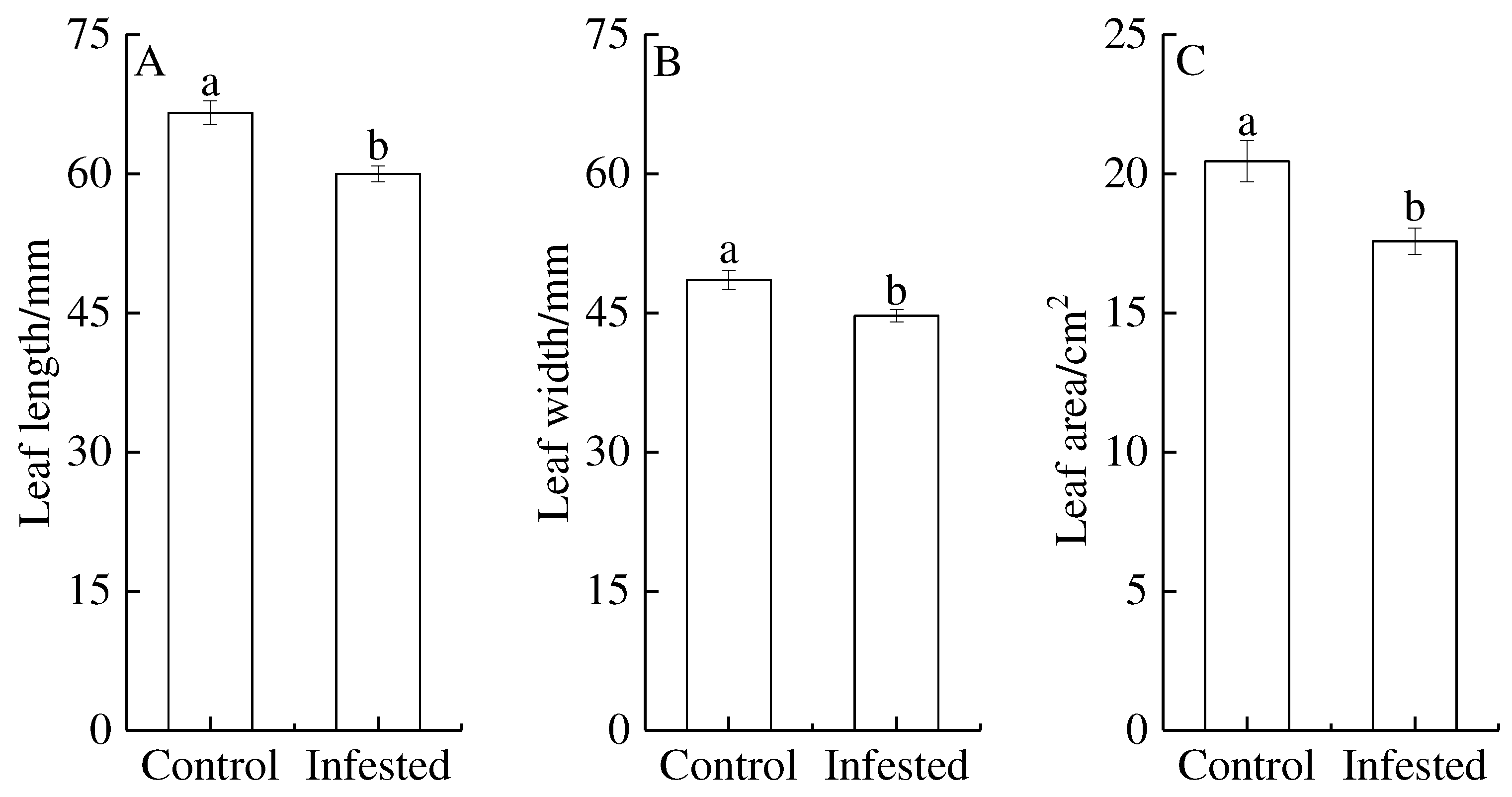
Figure 3.
Comparison of chlorophyll SPAD values in Prunus armeniaca across treatments with Sphaerolecanium prunastri (infested) and the control. Different lowercase letters are significantly different according to an independent two-sample t-test (P<0.05).
Figure 3.
Comparison of chlorophyll SPAD values in Prunus armeniaca across treatments with Sphaerolecanium prunastri (infested) and the control. Different lowercase letters are significantly different according to an independent two-sample t-test (P<0.05).
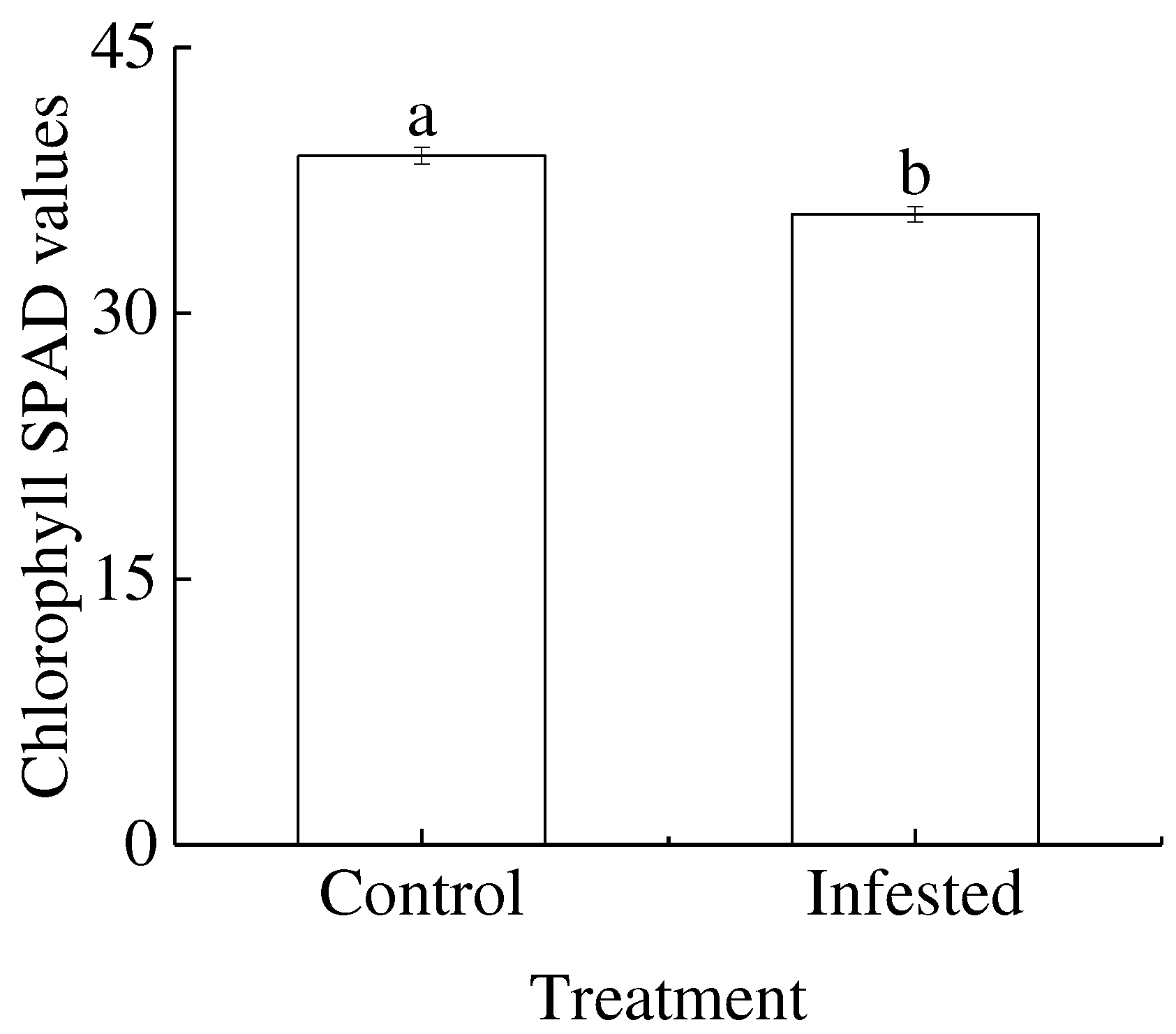
Figure 4.
Treatment differences in (A) leaf fresh weight and (B) leaf dry weight of Prunus armeniaca infested with Sphaerolecanium prunastri (infested) and uninfested controls. Different lowercase letters are significantly different according to an independent two-sample t-test (P<0.05).
Figure 4.
Treatment differences in (A) leaf fresh weight and (B) leaf dry weight of Prunus armeniaca infested with Sphaerolecanium prunastri (infested) and uninfested controls. Different lowercase letters are significantly different according to an independent two-sample t-test (P<0.05).
Figure 5.
Treatment differences in (A) fruit longitudinal diameter, (B) fruit cross diameter, (C) fruit side and (D) fruit weight of Prunus armeniaca infested with Sphaerolecanium prunastri (infested) and non-infested controls. Different lowercase letters are significantly different according to an independent two-sample t-test (P<0.05).
Figure 5.
Treatment differences in (A) fruit longitudinal diameter, (B) fruit cross diameter, (C) fruit side and (D) fruit weight of Prunus armeniaca infested with Sphaerolecanium prunastri (infested) and non-infested controls. Different lowercase letters are significantly different according to an independent two-sample t-test (P<0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



