
Submitted:
02 September 2023
Posted:
06 September 2023
You are already at the latest version
Abstract
Background: It is known that Preeclampsia affects the lactogenesis. However, data on effects of this pathology on the human milk neurobiomarkers composition are not available. The aim of this study is to investigate the effects of this gestational pathology on Activin A levels, a neurobi-omarker known to play an important role in the development and protection of the central nerv-ous system. Methods: The women recruited were divided in two different study groups: preeclamptic or normotensive women. All the human milk samples were collected using the same procedure. Activin A was quantified using an ELISA test. To investigate the effect of Preeclampsia on the Activin A concentration in the 3 lactation phases, mixed linear model with unistructural covariance structure, mother as random effect, and fixed effects were performed. Results: Activin A was detected in all samples. There were no significant differences between Preeclamptic and Normotensive women. The only significant effect is related to the lactation phase: it is significant the difference between colostrum and mature milk (p<0.01). In conclusion these results allow us to affirm that breast milk beneficial properties are maintained even if preeclampsia occurs.
Keywords:
human milk
; breastfeeding
; activin A
; preeclampsia
; newborn nutrition
; neuro-biomarker
1. Introduction
Preeclampsia (PE) is a gestational hypertensive syndrome characterized by a complex disease with variable clinical manifestation [1]. In most cases, the symptoms occur after the 20th week of Gestational Age (GA); the earlier its onset, the more serious it is [2,3]. PE is a common pregnancy complication that occurs in about 2-8% of pregnancies globally. At the basis of PE there is an early functional alteration of unknown origin to the development of placental-vascularization[3]. Among the etiological hypotheses advanced, it can be found an unbalance in the pro- and antiangiogenic factors, a reduced tolerance towards the child or the father, or insulin resistance[4]. PE is one of the primary causes of fetal-maternal morbidity and mortality. In addition, infants born to preeclamptic mothers are at high risk for several disorders: not only endocrine, nutritional, and metabolic but also cognitive due to neurodevelopmental impairments [5,6].
An aspect of interest is the possible effect that PE may have on the mammary gland, and therefore on breast milk composition. In fact, it is well known that Human milk (HM) is a peculiar food owing unique properties and resulting the ideal nourishment during neonatal period for the growing infant. HM is an individual species-specific biological “dynamic” system characterized by an extreme variability in its composition, with regard to both nutritional and bioactive components, changing according to lactation phase and adapting composition to different conditions such as GA, gestational pathologies and/or maternal diet [7]. From an evolutionary perspective, its composition has evolved over time to provide the infant a well-balanced nutrition and protection against potential infectious pathogens while the neonatal immune system completes its development. Mother’s own milk is always considered the first choice for nutrition of all neonates, including preterm newborns, thanks to the HM specific biological active factors (i.e. hormones, immunoglobulins, lysozyme, lactoferrin, saccharides, nucleotides and neuro-biomarkers) that improved several neonatal outcomes both in the short and the long-term[8,9,10]. Neuro-biomarkers are important HM components and between these, Activin A can play a relevant role as a growth factor[11,12]. Activin A is a dimeric protein belonging to the transforming growth factor beta (TGF-beta) superfamily and its receptors are widely distributed in the central nervous system (CNS) [13]. Studies in humans and animal model showed that Activin A can play a trophic and neuroprotective role on the CNS [13,14].
Although it is known that PE affects the lactogenesis, literature data on effects of this syndrome on the neuro-biomarkers composition and Activin A of HM of the lactating mother are not available.
Thus, the aim of this study is to integrate and to expand the available literature data by investigating the association between the composition of human milk and PE, considering the variations of this key biochemical marker during different lactation phases in mothers having delivered term and preterm infants.
2. Metods
2.1. Setting and Population
Mothers admitted into the study gave signed and informed consent. Newborns’ mothers were recruited after delivery at Neonatal Unit of Department of Public Health and Pediatrics, University of Turin, Italy. Written informed consent was obtained from all participants and approval from the local ethics committee was obtained.
The women recruited in the study were divided in the two different study groups: preeclamptic, according to the PE definition (Artery blood pressure >140/90 after 20 weeks of gestational age and proteinuria >290 mg/l, possibly associated with headache, edema, scotomas and epigastric pain [1]) or normotensive women.
A minimum of 30 women for each group will be recruited.
Exclusion Criteria: presence of diabetes mellitus, chorioamnionitis, all CNS pathologies or psychiatric syndromes; use of illicit drugs/alcohol during pregnancy; mastitis or continuous use of medication; newborn with congenital anomalies or infection.
The control group was formed at the same time and made up of normotensive mothers who met the same exclusion criteria.
2.2. Collection human milk samples
According to standard criteria, we classify “colostrum” the milk collected in the first three days after the delivery; “transition milk” the milk collected from the 8th day to 14th day after the delivery and “mature milk” the milk collected after the 30th day [15]. All the breast milk samples were collected using the same procedure outlined below. Fresh milk samples were collected in the morning (between 9 a.m. and 12 a.m.) into sterile disposable. Milk was collected with standard extraction methods by emptying one breast completely by means of an electric breast pump (Medela Symphony, Baar, Switzerland). A minimum of 10 ml of milk were collected and immediately frozen at -80°C before the analysis. Milk expression by the other breast was performed only in cases in which it is not possible to obtain 10 ml from a single breast.
2.3. Activin A measurements
Activin A levels were determined using a specific ELISA test (ELH-ActivinA-1 Human Activin A ELISA) according to the manufacturer's instructions (RayBiotech, Inc.; USA). Investigators who performed the laboratory tests were blind to storage modalities. The assay detection limit is 15.00 pg/ml, the intra-assay CV is ≤ 5.0%, and the inter-assay CV is ≤ 10%. The assay is specific for Activin A, having been tested by the manufacturer for lack of cross-reactivity with other proteins of the Activin family.
2.4. Statistical methods
In the description of the sample, the categorical variables were presented as frequencies (percent), while the continuous variables were presented as mean (standard deviation) or median (interquartile range) (IQR) according to their distribution.
Birth weight was transformed in z-score using as reference INeS charts [16]. Newborns with a birth weight lower than the 10th or higher than the 90th centile were classified as Small for GA (SGA) or Large for GA (LGA) respectively. The continuous variables were summarized as mean (Standard Deviation) or median [Inter Quartile Range] according to their distribution, the categorical variables were summarized as absolute frequency (percent). To investigate the distribution by HM phase and pathology of Activin A, specific box plot was done. Then Activin A concentration was normalized with the more appropriate Box-Cox transformation. To investigate the effect of pathology in the Activin A concentration in the 3 phases, mixed linear model with unistructural covariance structure, mother as random effect, and fixed effects were performed. The fixed effects were: hm phase, pathology, smoke, type of delivery, GA and mother age (continuous), and the interaction between phase hm×pathology.
3.Results
A total of 82 mothers were recruited for our study, divided as follow: 36 in the PE group and 49 in the normotensive group. Table 1 reports the basal characteristic of mothers and newborns included in this study. Regarding the maternal characteristics, in both group the median age is quite similar as well as the fraction of primigravida. Moreover, the percentage of Caesarean section is high into two groups.
Concerning the neonatal characteristics, as expected, the women with PE have a higher fraction of IUGR and SGA newborns. In both groups similar percentage of twin pregnancies is observed.
3.1. Characteristics of the Human Milk samples
A total of 158 HM samples were collected. In particular, concerning the Normotensive group, a total of 79 samples were collected, of which 30 were colostrum, 24 of transitional milk and 25 of mature milk. In the PE group, a total of 79 samples were collected, of which 30 were colostrum, 27 of transitional milk and 22 of mature milk.
3.2. Activin A Concentrations
Activin A has been detected in all samples and in all types of HM, regardless of the lactation phase, gestational pathologies and the GA at childbirth.
In the Normotensive group, the Activin A median concentration was respectively: 232.47 pg/ml [ IQR 96.13-771.46] in colostrum samples, 122.47 pg/ml [ IQR 74.80-254.80] in transitional milk samples, 147.46 pg/ml [ IQR 82.80-260.80] in mature milk samples, 142.13 pg/ml [ IQR 71,46-280.63] in HM samples of women who delivered at term of GA and 232,46 pg/ml [ IQR 132,96-744,46] in HM samples of women who delivered preterm of GA. In the PE group, the Activin A median concentration was respectively 553.80 pg/ml [ IQR 340.13-751.46] in colostrum samples, 238.80 pg/ml [ IQR 152.80-428.13] in transitional milk samples 108.13 pg/ml [ IQR37.46-274.80] in mature milk samples, 703,46 pg/ml [ IQR 452,13-1141.58] in HM samples of women who delivered at term of GA and 475,46 pg/ml [ IQR 206,13-577,46] in HM samples of women who delivered preterm of GA.
Figure 1 shows the box-plot Activin A distribution by HM phase and groups. The variability in HM phase 1 (colostrum) is higher than in phase 2 (Transitional milk) and 3 (Mature milk). λ=0 resulted the more appropriate Box-Cox transformation to normalized Activin A distribution. The mixed linear model resulted in no significant effect of pathology and phase hm×pathology. The only significant effect is related to the HM phase, in particular is significant the difference between Colostrum and Mature milk (p<0.01).
4. Discussion
Among the maternal organs affected by the PE there are the mammary glands. As well as in the rest of the body, even at this level there could be changes in the endothelium and blood vessels: these would lead a reduction in the development of the gland and changes in the mechanisms of production of milk [17]. It is also known that the children of preeclamptic mothers, exposed to intrauterine stress, may have special nutritional needs in addition a greater risk of complications [2,18]. In view of these considerations, it is interesting to evaluate the potentially differences in composition between HM of PE women and normotensive women, in the different lactation phases.
Our study is the first that provide data on the association between PE and HM Activin A levels. Our results shown the absence of significant differences between the two women groups.
Considering the importance of the HM in newborn nutrition, previous studies have focused their attention on PE lactating mother. Data shown alteration in the levels of several components: macronutrients ( i.e. proteins, carbohydrates, lipids and energy metabolites) and pro- and anti-inflammatory cytokines, oxidative stress markers and antioxidant molecules [19,20,21,22,23,24,25]. Specifically, two previous studies evaluated the effects of PE on two similar panel of cytokines of the HM, and both considered only the variations between colostrum to mature milk [21,22]. Data of both studies demonstrated that PE modifies the inflammatory cytokine levels in HM, as well as the cytokine profile and these modifications depended on the lactation stage. First study showed that pro-inflammatory cytokines (IL-1β, sIL-2R, IL-6, IL-8 and TNF-α) in HM display biological differences in different periods of lactation: that is, higher cytokine levels in the colostrum versus mature milk following normal pregnancy. In PE, high cytokine levels perdure in mature milk but all cytokines’ concentrations were not significant different in the PE versus the control group in colostrum. However, IL-8 and TNF- α levels were higher in the PE group versus control in mature milk [21]. The data of other study showed that in the colostrum of PE group, IL-1b and IL-6 levels increased, and IL-12 levels decreased, whereas in the mature milk IL-6 and IL-8 levels decreased more than those of the control. Regarding the differences during the lactation period, in the control group the levels of IL-8, IL-10, and IL-12 are lower in mature milk than in colostrum, whereas the IL-6 concentration was higher in mature milk. In opposition, many cytokine levels in PE are stable and show no differences between colostrum and mature milk; only IL-1β and IL-8 decrease during postpartum period [22]. The levels of most cytokines did not decrease as lactation progressed, which may reflect the persistence of the systemic inflammatory response or a change in the immune system of the mammary gland in women with PE [21,22]. PE is a systemic inflammatory disease, so it is interesting to speculate whether the inflammatory response also occurs in the mammary gland, leading to increased levels of inflammatory cytokines in the human milk. Moreover, cytokine production in the mammary gland is an active process, so the reduction of IL-12 in the milk of mothers with PE may represent a defense mechanism for a neonate exposed to a chronic inflammatory condition during fetal life [26,27]. Another interesting finding in the PE group was the lower concentrations of IL-6 and IL-8 in mature milk. These cytokines are produced in the mammary gland, so this decrease during the progression of lactation may reflect persistent adaptations in the mother’s body to protect the newborn. However, IL-8 plays a trophic role in the intestinal mucosa of the infant, so decreased values can mean less protection for the gastrointestinal tract [26]. IL-1β increased levels in colostrum can be beneficial to the newborn because it appears to be involved in human milk defense mechanisms, including the production of IgA and other cytokines [28].
Regarding neurotrophic factors, Dangat et al. examined the levels of brain-derived neurotrophic factor (BDNF) and nerve growth factor (NGF) [19,29]. At first, they evaluated the neurotrophins levels only in colostrum and observed that NGF levels were similar, whereas BDNF levels were higher in the PE group as compared to controls[19]. In the second time, they extended the evaluation of these agents through the other phases of lactation and they found that the NGF concentrations at 1.5 and 3.5 months and BDNF levels at 1.5 months were lower in the PE group as compared to control group [29]. BDNF and NGF are known to play a critical role in the development and maintenance of the nervous system. The significant quantitative differences in this neurotrophin at several time points during lactation probably indicate that the milk programming by mother’s breast is altered by preeclampsia [29]. It is likely that these changes are most likely adaptive changes of the mother; some of these changes are not normalized even up to 6 months [29].
Anyway, these findings in Activin A levels herein reported warrants further consideration. In particular, Activin A probably acts in HM as a growth factor. In the brain, the expression of Activin A is uneven: its levels vary according to the specific regions considered. In previous studies demonstrated that Activin A have a neurotrophic function in differentiations of many CNS target cell-types[13,30]. In the last years, the hypothesis of a possible role of Activin A as a neuroprotective factor is emerging, in the light of the multiple actions it performs at the level of this tissue [31]. Several in vitro and in vivo researches have investigated the effect of Activin A on the nervous system, showing that: i) acts as a nerve tissue survival factor [32], a neural differentiation inhibitor [33] and a mitogen factor [34]; ii) is a powerful survival factor for neurogenic clonal cell lines, retinal neurons and dopaminergic neurons of the midbrain [35]; iii) promotes in vitro survival of hippocampal neurons from rat embryos [35]. In addition, Activin A promotes the survival of specific populations of damaged neurons and its expression is crucial for neuronal protection in case of brain injury [36,37]. In fact, it has been shown to protect the dopaminergic neurons of the midbrain from neurotoxic damage [38] and, in experiments on rats, to recover striatal neurons undergoing neurotoxicity [39]. It seems exert a role of CNS protection from antidepressant treatment side-effects[40]. It also performs, on the other hand, a biomarker function of damage, especially at the brain level, so it has also been suggested to use it as an early neonatal indicator of neurological insults, caused for example by asphyxia and intraventricular hemorrhage after birth[41]. There is also evidence that the protective role of Activin A is also extended to the heart tissue. In particular, it has been reported that the protein can participate in a cascade of events promoting tissue protection and regeneration in patients who have undergone ischemia/reperfusion injury [42,43]
In addition, a fairly large number of researchers have evaluated, in other types of samples (i.e blood, plasma, urine), the variation in the Activin A concentration. During pregnancy, the predominant source of Activin A is the placenta, which expresses both βA-mRNA and molecule receptors. High levels of Activin A are also present in the amniotic fluid, in which the concentration increases as pregnancy progresses, and in the celomatic fluid, which performs the function of reservoir of Activin A for the development of the fetus, although the molecule is also produced in fetal tissues [31]. Its levels rise in the maternal plasma during gestation, reaching maximum values at the end of the same: in particular, the serum level of Activin is higher in women who give birth with vaginal delivery than those who undergo a caesarean section, suggesting therefore an effect of the molecule on the mechanisms of childbirth (through the liberation of prostaglandins and oxytocin). In case of PE many studies agree that there is a significant correlation between blood and Activin A levels [44]. In PE, in particular, Activin A free in the maternal circulation could suggest a role in the processes of adaptation of the maternal organism in response to pathology; Moreover, high levels of Activin A in mid-trimester may be helpful in predicting which patients will develop preeclampsia during pregnancy. HM Activin A is expressed by the mammary gland (βa-subunit and βa-mrna have been localized in ductal and lobular epithelial cells). It therefore appears to have at least two other important functions [12]: i) is necessary for the proper development of the mammary gland itself, acting as an autocrine and paracrine factor: if the gene encoding Activin A is inactivated, the development of the breast is incomplete, there is no milk production, the elongation of the ducts is not complete, and the morphogenesis of the secreting alveolar ducts is altered; ii) is likely to have a growth factor function on various tissues of the newborn (including the brain and heart tissues mentioned above) and an immune function. Activin A, in fact, increases the production of cytokines by mononucleate cells, regulates the development of T cells and performs both pro-and anti-inflammatory actions.
The presence of Activin A was evaluated for the first time by Luisi et al. [12] in HM of women that delivered at term of GA. The results of this study show no differences in Activin A levels for type of delivery, maternal age, gestational age. There are also no differences between colostrum and transitional milk samples. The only significant difference lies in the molecule concentrations in mature milk, which are significantly lower than those of colostrum [12]. Our data show, like this previous study, differences in Activin A concentration in the different lactation phases, with a significant decrease in levels from colostrum to mature milk, in mother having delivered also preterm and term of GA. In addition this current study confirm the presence of Activin A also in HM of woman that delivered preterm, as demonstrated only in a single previous study [11], and our data shown no differences between term and preterm levels.
Bearing in mind these considerations, these data may suggest a trophic role of Activin A also in breast milk. Moreover, the absence of differences in HM Activin A composition is an important finding: in fact, thanks to these data, it can be said that the beneficial properties of milk are maintained even in the event of the onset of PE. This is a very encouraging fact, especially considering the high vulnerability, already mentioned, of the children of hypertensive mothers.
Future studies are needed to confirm the present findings and to obtain a more comprehensive evaluation of the effects of this important pathology on the HM. It will be also important to analyze the effect of different drugs of PE on HM and their potential interaction on the different biological components. These future findings are relevant to individualize and modify the maternal therapies and supplement properly the nutrition especially of the preterm newborns.
5. Conclusions
Breast milk is the ideal food for newborns and infants. Breastfeeding has positive implications for health of the newborn and the mother, and its benefits are greater the more the feeding with breast milk is prolonged and exclusive. Moreover, the effects of such nutrition are not only limited to the period in which the infant is actually fed with human milk but are also evident after many years: the composition of the milk is in fact such as to affect the child’s own development, both at an organic and psycho-cognitive level, thus causing beneficial effects in the short and long-term [8,9,10]. Given the importance of nutrition of newborns with breast milk, many studies have investigated the characteristics of HM of women with gestational pathologies. Some of them have focused on women with gestational hypertension and/or preeclampsia, comparing the composition of their milk with that of Normotensive mothers. The main observation on the data reported in literature is that PE altered the composition of HM milk. The effect of PE on HM is present not only in the first day post-partum but continue throughout all the lactating phases. In some cases, the modification in concentration of biological factors of HM may appear as a benefit for the newborns but the variation of other components is a disadvantage for the development of these babies [25]. Currently, however, the influence of this pathology on the composition of milk is still little known. The objective of our study was to fill part of this gap in literature. Our study is the first that extensively evaluates the concentration of these molecules in the milk of women with PE, regardless of the gestational age at delivery and taking into account all stages of lactation.
Our data show that there is not significant difference in HM Activin A composition from PE and normotensive women.
Further studies are needed to confirm our findings and to identify the potential correlations between the composition of the HM and the different gestational hypertension pathologies. In conclusion, our study confirms that the biological value of human milk associated with the Activin A content is maintained in preeclamptic mothers.
Author Contributions
“Conceptualization, C.P., A.C. and D.G.; methodology, C.P., E.S, I.B., G.L. and F.G.; formal analysis, I.B., G.L. and F.G.; data curation, E.S. and S.S.; writing—original draft preparation, C.P., S.S., L.R., E.S. E.B; writing—review and editing, C.P., L.R., A.C., E.B. and D.G. All authors have read and agreed to the published version of the manuscript.”.
Funding
“This research received no external funding”
Institutional Review Board Statement
“The study was conducted in accordance with the Declaration of Helsinki, and approved by the local Ethics Committee: Comitato di Bioetica di Ateneo, University of Turin, protocol number 169849, Approval date: 24/02/2021”
Informed Consent Statement
“Informed consent was obtained from all subjects involved in the study.”.
Conflicts of Interest
“The authors declare no conflict of interest.”.
References
- Gestational Hypertension and Preeclampsia: ACOG Practice Bulletin, Number 222. Obstetrics & Gynecology 135(6):p e237-e260, June 2020. | . [CrossRef]
- Lisonkova S, Joseph KS. Incidence of preeclampsia: risk factors and outcomes associated with early- versus late-onset disease. Am J Obstet Gynecol. 2013 Dec;209(6):544.e1-544.e12. Epub 2013 Aug 22. [CrossRef] [PubMed]
- Uzan J, Carbonnel M, Piconne O, Asmar R, Ayoubi JM. Pre-eclampsia: pathophysiology, diagnosis, and management. Vasc Health Risk Manag. 2011;7:467-74. Epub 2011 Jul 19. [CrossRef] [PubMed] [PubMed Central]
- Hod T, Cerdeira AS, Karumanchi SA. Molecular Mechanisms of Preeclampsia. Cold Spring Harb Perspect Med. 2015 Aug 20;5(10):a023473. [CrossRef] [PubMed] [PubMed Central]
- Backes CH, Markham K, Moorehead P, Cordero L, Nankervis CA, Giannone PJ. Maternal preeclampsia and neonatal outcomes. J Pregnancy. 2011;2011:214365. Epub 2011 Apr 4. [CrossRef] [PubMed] [PubMed Central]
- Leitner Y, Harel S, Geva R, Eshel R, Yaffo A, Many A. The neurocognitive outcome of IUGR children born to mothers with and without preeclampsia. J Matern Fetal Neonatal Med. 2012 Nov;25(11):2206-8. Epub 2012 May 4. [CrossRef] [PubMed]
- Andreas NJ, Kampmann B, Mehring Le-Doare K. Human breast milk: A review on its composition and bioactivity. Early Hum Dev. 2015 Nov;91(11):629-35. Epub 2015 Sep 12. [CrossRef] [PubMed]
- Gartner LM, Morton J, Lawrence RA, Naylor AJ, O'Hare D, Schanler RJ, Eidelman AI; American Academy of Pediatrics Section on Breastfeeding. Breastfeeding and the use of human milk. Pediatrics. 2005 Feb;115(2):496-506. [CrossRef] [PubMed]
- Horta BL, and Victora CG. Long-term health effects of breastfeeding: a systematic review. World Health Organization 2013.
- Bar S, Milanaik R, Adesman A. Long-term neurodevelopmental benefits of breastfeeding. Curr Opin Pediatr. 2016 Aug;28(4):559-66. [CrossRef] [PubMed]
- Peila C, Coscia A, Bertino E, Li Volti G, Galvano F, Barbagallo I, Visser GH, Gazzolo D. The Effect of Holder Pasteurization on Activin A Levels in Human Milk. Breastfeed Med. 2016 Nov;11:469-473. Epub 2016 Aug 16. [CrossRef] [PubMed]
- Luisi S, Calonaci G, Florio P, Lombardi I, De Felice C, Bagnoli F, Petraglia F. Identification of activin A and follistatin in human milk. Growth Factors. 2002 Sep;20(3):147-50. [CrossRef] [PubMed]
- Gazzolo D, Abella R, Frigiola A, Giamberti A, Tina G, Nigro F, Florio P, Colivicchi M, Temporini F, Ricotti A, Li Volti G, Galvano F. Neuromarkers and unconventional biological fluids. J Matern Fetal Neonatal Med. 2010 Oct;23 Suppl 3:66-9. [CrossRef] [PubMed]
- Tretter YP, Hertel M, Munz B, ten Bruggencate G, Werner S, Alzheimer C. Induction of activin A is essential for the neuroprotective action of basic fibroblast growth factor in vivo. Nat Med. 2000 Jul;6(7):812-5. [CrossRef] [PubMed]
- Playford RJ, Macdonald CE, Johnson WS. Colostrum and milk-derived peptide growth factors for the treatment of gastrointestinal disorders. Am J Clin Nutr. 2000 Jul;72(1):5-14. [CrossRef] [PubMed]
- Bertino E, Di Nicola P, Varalda A, Occhi L, Giuliani F, Coscia A. Neonatal growth charts. J Matern Fetal Neonatal Med. 2012 Apr;25 Suppl 1:67-9. Epub 2012 Mar 12. [CrossRef] [PubMed]
- Demir D., Demirel Sezer E, Turan V., Ozturk S., Canbay E. and E. Yıldırım Sozmen E. How Preeclampsia Affects Oxidant Status and Antiiflammatory Potential of Breast Milk? Free Radic. Biol. Med. 2016; 100:S59 . [CrossRef]
- Duley L. The global impact of pre-eclampsia and eclampsia. Semin Perinatol. 2009 Jun;33(3):130-7. [CrossRef] [PubMed]
- Dangat K, Kilari A, Mehendale S, Lalwani S, Joshi S. Higher levels of brain derived neurotrophic factor but similar nerve growth factor in human milk in women with preeclampsia. Int J Dev Neurosci. 2013 May;31(3):209-13. Epub 2013 Jan 18. [CrossRef] [PubMed]
- Cekmen MB, Balat A, Balat O, Aksoy F, Yurekli M, Erbagci AB, Sahinoz S. Decreased adrenomedullin and total nitrite levels in breast milk of preeclamptic women. Clin Biochem. 2004 Feb;37(2):146-8. [CrossRef] [PubMed]
- Erbağci AB, Cekmen MB, Balat O, Balat A, Aksoy F, Tarakçioğlu M. Persistency of high proinflammatory cytokine levels from colostrum to mature milk in preeclampsia. Clin Biochem. 2005 Aug;38(8):712-6. [CrossRef] [PubMed]
- Freitas NA, Santiago LTC, Kurokawa CS, Meira Junior JD, Corrente JE, Rugolo LMSS. Effect of preeclampsia on human milk cytokine levels. J Matern Fetal Neonatal Med. 2019 Jul;32(13):2209-2213. Epub 2018 Jan 25. [CrossRef] [PubMed]
- Dangat K, Upadhyay D, Kilari A, Sharma U, Kemse N, Mehendale S, Lalwani S, Wagh G, Joshi S, Jagannathan NR. Altered breast milk components in preeclampsia; An in-vitro proton NMR spectroscopy study. Clin Chim Acta. 2016 Dec 1;463:75-83. Epub 2016 Oct 12. [CrossRef] [PubMed]
- Dangat KD, Mehendale SS, Yadav HR, Kilari AS, Kulkarni AV, Taralekar VS, Joshi SR. Long-chain polyunsaturated fatty acid composition of breast milk in pre-eclamptic mothers. Neonatology. 2010;97(3):190-4. Epub 2009 Oct 28. [CrossRef] [PubMed]
- Peila C, Bertino E, Cresi F, Coscia A. Interactions between preeclampsia and composition of the human milk: what do we know? J Matern Fetal Neonatal Med. 2021 Jun 13:1-7. [CrossRef]
- Zhou Y, Sun X, Jin L, Stringfield T, Lin L, Chen Y. Expression profiles of adiponectin receptors in mouse embryos. Gene Expr Patterns. 2005 Jun;5(5):711-5. [CrossRef]
- Saito S, Sakai M, Sasaki Y, Tanebe K, Tsuda H, Michimata T. Quantitative analysis of peripheral blood Th0, Th1, Th2 and the Th1:Th2 cell ratio during normal human pregnancy and preeclampsia. Clin Exp Immunol. 1999 Sep;117(3):550-5. [CrossRef]
- Jonsson Y, Rubèr M, Matthiesen L, Berg G, Nieminen K, Sharma S, Ernerudh J, Ekerfelt C. Cytokine mapping of sera from women with preeclampsia and normal pregnancies. J Reprod Immunol. 2006 Jun;70(1-2):83-91. [CrossRef]
- Dangat K, Kilari A, Mehendale S, Lalwani S, Joshi S. Preeclampsia alters milk neurotrophins and long chain polyunsaturated fatty acids. Int J Dev Neurosci. 2014 Apr;33:115-21. Epub 2013 Dec 25. [CrossRef] [PubMed]
- Serpero LD, Frigiola A, Gazzolo D. Human milk and formulae: neurotrophic and new biological factors. Early Hum Dev. 2012 Mar;88 Suppl 1:S9-12. Epub 2012 Jan 17. [CrossRef] [PubMed]
- Luisi S, Florio P, Reis FM, Petraglia F. Expression and secretion of activin A: possible physiological and clinical implications. Eur J Endocrinol. 2001 Sep;145(3):225-36. [CrossRef]
- Schubert D, Kimura H, LaCorbiere M, Vaughan J, Karr D, Fischer WH. Activin is a nerve cell survival molecule. Nature. 1990 Apr 26;344(6269):868-70. [CrossRef] [PubMed]
- Hashimoto M, Kondo S, Sakurai T, Etoh Y, Shibai H, Muramatsu M. Activin/EDF as an inhibitor of neural differentiation. Biochem Biophys Res Commun. 1990 Nov 30;173(1):193-200. [CrossRef]
- Schubert D, Kimura H. Substratum-growth factor collaborations are required for the mitogenic activities of activin and FGF on embryonal carcinoma cells. J Cell Biol. 1991 Aug;114(4):841-6. [CrossRef]
- Iwahori Y, Saito H, Torii K, Nishiyama N. Activin exerts a neurotrophic effect on cultured hippocampal neurons. Brain Res. 1997 Jun 20;760(1-2):52-8. [CrossRef] [PubMed]
- Wu DD, Lai M, Hughes PE, Sirimanne E, Gluckman PD, Williams CE. Expression of the activin axis and neuronal rescue effects of recombinant activin A following hypoxic-ischemic brain injury in the infant rat. Brain Res. 1999 Jul 24;835(2):369-78. [CrossRef]
- Tretter YP, Hertel M, Munz B, ten Bruggencate G, Werner S, Alzheimer C. Induction of activin A is essential for the neuroprotective action of basic fibroblast growth factor in vivo. Nat Med. 2000 Jul;6(7):812-5. [CrossRef]
- Krieglstein K, Suter-Crazzolara C, Fischer WH, Unsicker K. TGF-beta superfamily members promote survival of midbrain dopaminergic neurons and protect them against MPP+ toxicity. EMBO J. 1995 Feb 15;14(4):736-42. [CrossRef]
- Hughes PE, Alexi T, Williams CE, Clark RG, Gluckman PD. Administration of recombinant human Activin-A has powerful neurotrophic effects on select striatal phenotypes in the quinolinic acid lesion model of Huntington's disease. Neuroscience. 1999;92(1):197-209. [CrossRef]
- Duman RS, Monteggia LM. A neurotrophic model for stress-related mood disorders. Biol Psychiatry. 2006 Jun 15;59(12):1116-27. Epub 2006 Apr 21. [CrossRef] [PubMed]
- Serpero LD, Bellissima V, Colivicchi M, Sabatini M, Frigiola A, Ricotti A, Ghiglione V, Strozzi MC, Li Volti G, Galvano F, Gazzolo D. Next generation biomarkers for brain injury. J Matern Fetal Neonatal Med. 2013 Oct;26 Suppl 2:44-9. [CrossRef] [PubMed]
- Hedger MP, Phillips DJ, de Kretser DM. Divergent cell-specific effects of activin-A on thymocyte proliferation stimulated by phytohemagglutinin, and interleukin 1beta or interleukin 6 in vitro. Cytokine. 2000 Jun;12(6):595-602. [CrossRef]
- Schultz Jel J, Witt SA, Glascock BJ, Nieman ML, Reiser PJ, Nix SL, Kimball TR, Doetschman T. TGF-beta1 mediates the hypertrophic cardiomyocyte growth induced by angiotensin II. J Clin Invest. 2002 Mar;109(6):787-96. [CrossRef]
- Giguère Y, Charland M, Bujold E, Bernard N, Grenier S, Rousseau F, Lafond J, Légaré F, Forest JC. Combining biochemical and ultrasonographic markers in predicting preeclampsia: a systematic review. Clin Chem. 2010 Mar;56(3):361-75. Epub 2009 Dec 31. [CrossRef] [PubMed]
Figure 1.
Box-Plot Activin A by Phase HM and pathology. Median and [IQR] were reported for each HM Phases: 1 Colostrum; 2 Transitional milk; 3 Mature Milk.
Figure 1.
Box-Plot Activin A by Phase HM and pathology. Median and [IQR] were reported for each HM Phases: 1 Colostrum; 2 Transitional milk; 3 Mature Milk.
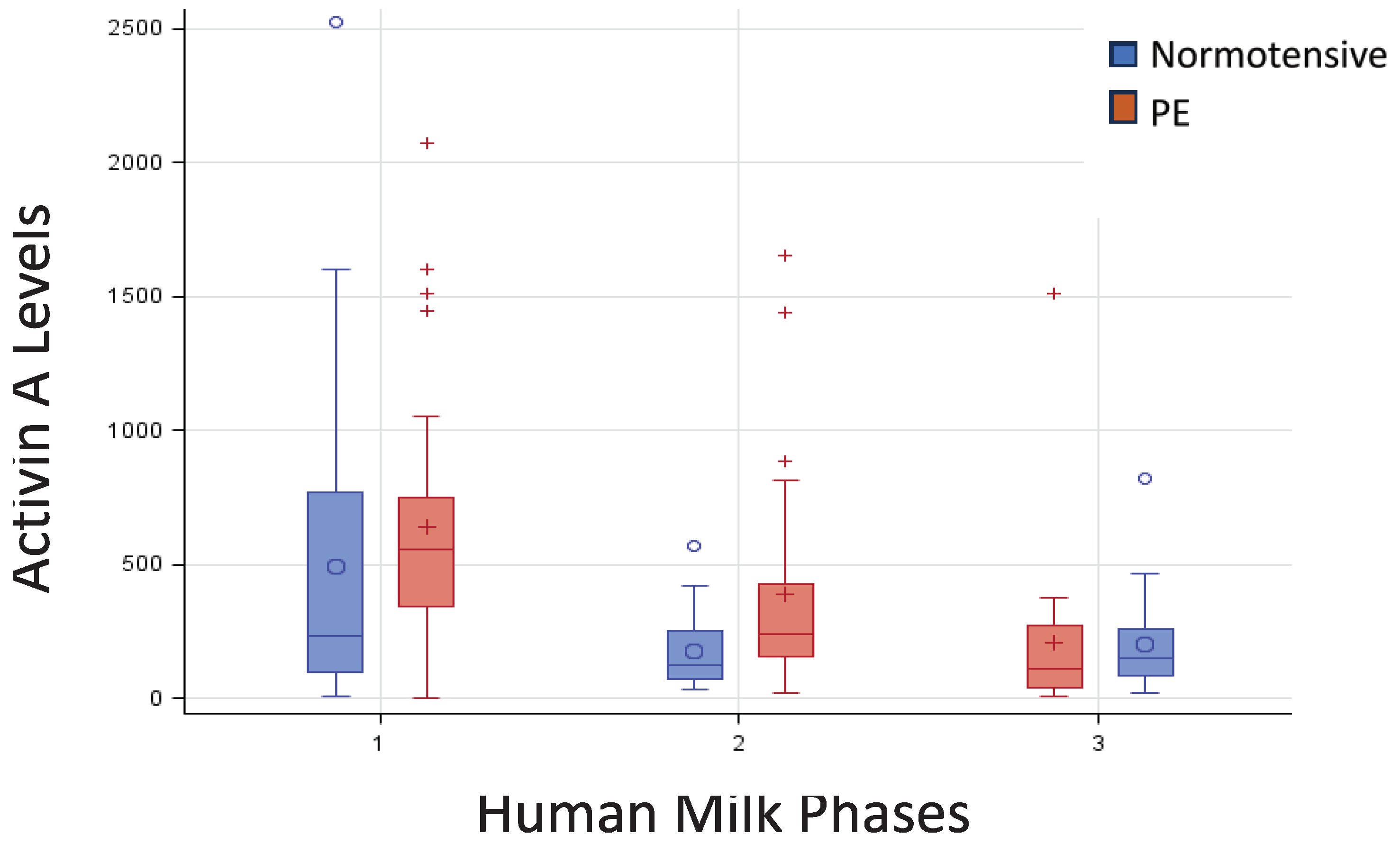

Table 1.
Basal characteristics of mothers and newborns.
| Normotensive N=46 |
Preeclamptic N=39 |
||
|---|---|---|---|
| Maternal characteristics | |||
| Age (years) | median [IQR] | 33.5 [31,32,33,34,35,36,37] | 35 [31,32,33,34,35,36,37,38] |
| Italian | n (%) | 35 (76.1) | 31 (79.5) |
| Caesarian Section | n (%) | 25 (54.4) | 28 (71.8) |
| Weight gain (kg) | mean (SD) | 10.9 (4.75) | 10.4 (5.65) |
| Primigravida | n (%) | 29 (63.0) | 25 (64.0) |
| Smoker | n (%) | 6 (13.0) | 2 (5.1) |
| Newborn characteristics | |||
| Singleton | n (%) | 38 (82.6) | 36 (92.3) |
| IUGR | n (%) | 2 (4.4) | 16 (41.0) |
| GA (weeks) | median [IQR] | 37 [31;39] | 32 [29,30,31,32,33,34,35] |
| Girls | n (%) | 19 (41.3) | 19 (48.7) |
| Birth weight (g) | mean (SD) | 2345 (1028) | 1542 (720) |
| Birth weight (z-score) | mean (SD | -0.21 (0.934) | -1.16 (0.810) |
| SGA | n (%) | 6 (13.0) | 19 (50.0) |
| LGA | n (%) | 2 (4.4) | 0 (0.0) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



