
Submitted:
31 August 2023
Posted:
06 September 2023
You are already at the latest version
Abstract
Babesia species infect a very wide range of mammal hosts across the globe and zoonotic infections are of growing concern. Several species of the Babesia genus infect dogs and some of these cause significant morbidity and mortality. The Apicomplexan parasite resides within the red cell and infections result in direct damage to the host through intra- and extravascular hemolysis. An exuberant inflammatory response by the host to some species of Babesia parasites also result in significant collateral damage to the host. Canine infections have been the subject of much study as the wellbeing of these companion animals is increasingly threatened by the spread of tick vectors and an increasingly mobile dog population. There are no widely available and effective vaccines and effective treatment can be challenging. Understanding disease pathogenesis underlies the development of new treatments. The varying pathogenicity of the various Babesia parasite species that infect dogs offers an opportunity to explore the molecular basis for the wide range of disease caused by infection with this parasite genus. In this review we focus on what has been reported about the clinical presentation of Babesia infected dogs in an attempt to compare the severity of disease caused by different Babesia species.
Keywords:
Babesiosis
; canine
; Babesia rossi
; Babesia canis
; Babesia gibsoni
; Babesia vogeli
; Babesia conradae
; Babesia vulpes
; disease comparison
Introduction
Canine babesiosis is the most common vector-borne infection in many parts of the developing world where infections are probably very under reported and where veterinary care for dogs is often non-existent. Around 10% of the small animal case load across surveyed practices in South Africa was attributed to canine babesiosis—and that in a country where veterinary care and ectoparasite control is wide spread [1]. It is a less common, albeit still important, cause of disease in the developed world. Several species of the Babesia genus infect dogs and result in variable clinical manifestations. Literature comparing clinical features of the disease caused by infections with the various Babesia species is sparse. Many factors determine the severity of the disease resulting from an infection and the interplay of these factors is obviously complex. The species of parasite infecting a dog is an important factor and does play a crucial (and probably dominant) role in the nature of the disease seen and this is the main focus of this review.
It is common for different parasite species of the same genus to induce clinically distinct diseases. The bovine Babesia, B. bigemina, causes a relatively mild acute disease followed by persistent infection. B. bovis on the other hand causes a severe acute disease characterized by neurological signs and a high mortality [2]. The neurovirulence of B. bovis is associated with adhesion of infected red cells to the endothelium of the microvasculature of the brain which initiates inflammation and disturbs normal cerebral blood flow. This is not seen in B. bigemina infections. Another relevant example is the varied disease caused by the important malaria parasite species (genus Plasmodium) that infect humans. Malaria parasites are also intraerythrocytic Apicomplexa and Babesia and malaria have been likened to one another for decades [3]. The disease caused by the malaria parasites that commonly infect humans (Plasmodium falciparum, P. vivax, P ovale and P. malariae) varies in many ways and the Plasmodium species responsible for the infection is strongly associated with clinical severity. Plasmodium falciparum is the most common cause of severe disease and also carries the highest mortality [4]. Plasmodium vivax typically causes a severe disease but with a significantly lower mortality [5]. Plasmodium malariae generally causes a mild disease. Severe disease with P. ovale is extremely rare, with infections often ending in spontaneous cure [6].
Classifying a disease severity has typically involved the development a composite clinical score. Various scoring systems have been developed in both human and veterinary care [7,8,9]. A system has been developed to assess the severity of immune mediated hemolytic anemia in dogs, a disease with similarities to babesiosis [10]. Scoring systems usually make use of a few crucial and easy to measure variables that, through regression analysis, result in the development of a set of numerically weighted criteria that can predict outcome or assist in assessing disease severity. There is no such scoring system for human malaria or canine babesiosis. Years of research however have created awareness amongst clinicians that certain findings are associated with less complicated disease (with a better outcome) or more complicated disease (with a poorer outcome, requiring more intensive treatment). The disease caused by some of the Babesia species that infect dogs has been classified as uncomplicated or complicated in a very similar vein to Falciparum malaria in humans. Criteria for this classification of canine babesiosis have been published for the disease caused by Babesia rossi [11] and Babesia canis [12].
Recognizing that the various canine Babesia parasites do in fact cause fairly distinct diseases (despite overlap) is important to try to explore the molecular basis for parasite species pathogenicity. It may be possible by comparing the genomes of various Babesia parasites with widely varying disease severities, that pathogenicity associated genes or gene families could be identified. This could be helpful in understanding disease pathogenesis, leading to the identification of mechanisms or gene products amenable to treatment, or identifying vaccine targets. In the bovine Babesia example cited above, parasite proteins that are expressed on the surface of infected red cells of the so-called Variant Erythrocyte Surface Antigen (VESA) family (products of the VESA genes) have been identified as possible candidates for mediating red cell cytoadhesion. The ability of a particular parasite to cause cytoadhesion plays an important part in the the varied pathogencities of different bovine babesia parasites [13]. Preliminary work with B. rossi has demonstrated a similar gene family (BrEMA1) and suggests that polymorphisms in these genes may result in different disease phenotypes [14]. Early innate immune responses are crucial to infectious disease outcomes. Hemoprotozoa express Pathogen Associated Molecular Pattern molecules (PAMP) that are early signals to Pathogen Associated Recognition Receptors (PARR) such as Toll-like receptors [15]. Genetic diversity in the Toll-like receptor gene family has been associated with malarial disease variability in humans [16]. Similar parasite genetic variability may also play a role in the varied disease phenotype seen in dogs infected with different Babesia species.
The clinical description of babesiosis provided here was gleaned from published case reports and case series of naturally and experimentally infected dogs. Reviews of the literature were seldom used. It is important to recognize that over the last few decades nomenclature has evolved. Small Babesia organisms were morphologically identified as B. gibsoni (such as in Conrad et al. 1991 [17]). Molecular techniques now recognize that this report was in fact describing B. conradae infections [18]. A similar situation arose with the large Babesia organisms which were all typically identified as B. canis. An example of this is seen in the work of Malherbe et al. in South Africa where the very severe babesiosis described is attributed to B. canis [19,20,21] which we now know described disease caused by B. rossi. More recent molecular techniques recognize 3 large parasites infecting dogs, namely B. canis, B. vogeli and B. rossi [22,23]. The early and seminal work by Maegraith et al. published in 1957 [24] attributes the disease they studied to B. canis when in all likelihood it actually describes an infection caused by a South African isolate now known to be B. rossi. B. vulpes (previously called B. annae, B. microti-like, and Babesia Spanish isolate [25]) infections in dogs have been reported in the USA and Europe [26,27,28,29,30]. There is some clinical description of the disease (many cases are associated with co-infections) but, where it exists, it is provided. A large Babesia parasite that infectes dogs (Babesia ‘coco’) has been described but too little is known about this infection to draw conclusions about the clinical manifestation of this infection to compare it to the disease caused by other Babesia parasites that infect dogs [31].
Things all Babesia infections in dogs have in common
Irrespective of which Babesia parasite is responsible for an infection, there are features that are broadly similar across the genus.
Not all cases of all Babesia infections make dogs clinically ill. Although probably rare, even in infections with what has traditionally been described as the most pathogenic parasite, B. rossi, subclinical parasitemia has recently been demonstrated in dogs [32]. The same has been described for B. canis [33]. Subclinical infection is common with B. vogeli infections seen in 5/12 cases in one study [34], all 4 cases described in a Chile study were assymptomatic [35], however an Italian study reported 11 dogs with B. vogeli infection, and all were ill. A large number of dogs (20/29, 69%) infected with B. canis were found to be asymptomatic in a Croatian study [36]. A significant proportion of B. gibsoni infected dogs are also subclinical. In a Korean study, 10% of 60 infected dogs showed no clinical signs [37]. In an experimental infection study, 2/10 dogs remained subclinical after needle infection [38]. In 9 cases described from North Carolina (USA), 1 was parasitemic but healthy [39]. In another American study, 9/10 parasitemic dogs were healthy [40]. A description of 58 cases of B. vulpes infections from Spain describes a severe disease with no mention of asymptomatic infection [27]. Evidence suggests that for B. rossi and B. canis, if a parasitemia is demonstrated in a sick dog, the default would be to consider the illness to be caused by Babesia infection. This is not always true for B. conradae [41] or B. gibsoni or B. vogeli.
Lethargy and anorexia are described for infections caused by all species of canine Babesia. Vomiting and diarrhea is also occasionally seen. In the chronic relapsing infections (such as what is described for B. gibsoni), weight loss may be a feature [38,42]. Lethargy and anorexia form part of the owners chief complaint in almost all B. rossi infected dogs [11]. It is also described in the majority of B. canis infections, seen in 63/63 cases from Hungary, in 49/49 cases from Germany and 50/50 cases from Croatia [12,33,43]. Anorexia was described in 76% of dogs (23/30) and depression in 93% (28/30) in an Italian study [44] and in 43/50 (86%) cases from Croatia [43]. Anorexia was described in 76% of dogs (23/30) and depression in 93% (28/30) in an Italian study [44]. These signs are significantly less common in B. vogeli infected dogs. None of 4 infected dogs from Chile showed any clinical illness [35]. Only splenectomised dogs (n = 3) in an experimental B. vogeli infection became depressed and anorexic and one of these dogs self-cured [45]. Five of 11 naturally infected dogs from Italy were described as lethargic and anorexic [44]. Lethargy was observed in 48/60 (80%) and anorexia in 51/60 (85%) naturally B. gibsoni infected dogs from Taiwan [37]. A smaller proportion of these dogs were also noted to have vomiting (18/60) and diarrhea (6/60) as part of their clinical histories [37]. In a description of 11 naturally infected B. conradae dogs, vomiting and lethargy are described as common [17]. B. vulpes infection would appear to be associated with obvious illness in a case series of 58 infections and several other case reports [26,27,46,47]. Babesia rossi and B. canis infections would appear to be equally likely to cause lethargy and anorexia. B. gibsoni may be a subclinical infection and B. vogeli is frequently identified as a subclinical infection. In the limited description B. vulpes infections, it would appear to cause a relatively severe disease with a pathogenicity somewhere between B. canis and B. gibsoni.
Pyrexia is a common finding with the more pathogenic parasites and is typically the result of a host response to an endogenous pyrogen. The association between a fever, TNF production and the cyclic growth of malaria parasites is a clear example of this [48]. It is described in all cases of B. rossi infection besides those with who are close to death where a low rectal temperature was a poor prognostic indicator (probably a terminal shock phenomenon) [11]. In B. canis it is also common but not uniformly. One study documented it in 84% (27/32) [49] and another in 43% (13/30) of cases [44]. Although B. vogeli infection is commonly subclinical, pyrexia was common in 11 dogs with evidence of disease [44]. All 8 dogs experimentally infected showed a mild pyrexia which self-resolved in a matter of days despite the infection persisting [45]. Five of 11 naturally infected Italian dogs developed pyrexia [44]. It is also not a consistent finding in B. gibsoni infections and when it is found it is described as being poorly correlated with parasitemia [17,38] and seldom rising above 40 °C [42]. Temperature is frequently described as normal despite parasitemia [38]. In an experimental infection pyrexia developed on days 13 and 14 post infection before parasites were seen on blood smear. Fever also resolved within days and never recurred despite a climbing parasitemia [50]. In another study, 8/10 dogs developed a transient fever which mostly resolved [38]. A study of 79 naturally infected dogs in India did not regard it as a cardinal clinical finding as only 31% (5/16) of dogs with positive blood smears were febrile [51]. In another study pyrexia was only detected in 2/8 naturally infected dogs that were anemic and PCR positive for B. gibsoni DNA [52]. Although pyrexia is noted to occur in B. conradae infections, it appears not to be a consistent finding [17,41]. Babesia rossi and B. canis appear equally likely to induce a fever whilst in B. gibsoni, B. vogeli and B. conradae this is an inconsistent finding.
All Babesia species that infect dogs can cause anemia. The severity, rate at which anemia develops following infection and strength of association between infection and anemia does seem to vary however. Babesia rossi almost always presents with anemia. Over a third of 320 dogs presented for care were severely anemic (hematocrit < 15%, requiring blood transfusions), a quarter were moderately so, and just under a quarter were mildly anemic. A small proportion of cases had normal hematocrits at presentation. Mortality was only marginally higher in the severely anemic dogs compared to the other groups [11]. All studies that evaluated the hematocrit of B. canis infected dogs described a significantly greater proportion of dogs with mild to moderate anemia and a very small proportion of these were treated with blood transfusions [33,44,49,53,54]. Case series reporting on the clinical disease caused by B. vogeli infections are less common. Anemia is reported but appears to be a feature of the infection in puppies (where it described as hemolytic and severe) or immunocompromised rather than immunocompetent adult dogs (where the infections is usually subclinical or reported as a co-infection) [44,45,55,56]. B. gibsoni caused anemia in over 80% of 60 infected dogs. A quarter of these dogs had mild to moderate anemia whilst just over 10% of dogs had hematocrits <20%. Similar findings in other studies describe a severe life-threatening anemia rarely with mild to moderate anemia being more characteristic of the infection [50,51,52,57,58]. There is more limited description of the clinical disease caused by B. conradae. Severe anemia is described in 11 cases (before B. gibsoni and B. conradae were understood to be separate species) [17]. The anemia is described as more pronounced than what is observed in B. gibsoni infections [58]. One study from California demonstrated mild anemia in 13/29 infected dogs [41] while another demonstrated severe anemia in 3/12 infected dogs [59]. Moderate regenerative anemia was present in 95% (20/21) cases of B. vulpes infections [27]. It would seem from these descriptions that B. rossi probably causes a more consistently severe anemia that likely evolves more quickly than what is seen in infections from other parasites. This is followed in severity by B. canis, then B. gibsoni and B. conradae and B. vulpes with B. vogeli being the infection least likely to cause life-threatening anemia.
Together with anemia, hemolysis is described in B. rossi (Figure 1). This is at times so rapid and overwhelming as to cause black urine (akin to the ‘Black Water Fever’ of Falciparum malaria in humans [60]) with port wine colored plasma [11,61,62,63]. Cell free hemoglobin plays a role in disease pathogenesis and is potentially an important measure of disease severity [64]. Eighty-four percent (269/320) of cases had hemoglobinuria at presentation [11]. A rare form of B. rossi is associated with hemoconcentration in the face of obvious hemolysis and carries a very poor prognosis [11,63]. Hemolysis is reported to be common in B. canis infections with macroscopically visible hemoglobin in urine and/or blood in >2/3rds of cases [12], in 24/49 dogs [33] and in 63% of 63 cases [49]. Hemolytic anemia is reported in 11 cases of B. vogeli infection although this is less commonly reported than for either B. rossi of B. canis [44]. Clinical evidence of rapid intravascular hemolysis is reported for B. gibsoni but in a small percentage of cases [57] and evidence for this in B. vulpes infections is lacking. It is likely that the more slowly developing infections (such as B. gibsoni and B. vulpes) result in slower extravascular hemolysis. It appears that B. rossi may be responsible for more severe and more sudden hemolysis compared to B. canis although hemolysis in B. canis is nevertheless clinically obvious in the majority of cases. The other parasites, although reported to be cause hemolysis, typically show variable and less marked clinical signs of this process.
It is no surprise that dogs infected with Babesia develop a reactive splenomegaly given the spleen’s role as the primary immune organ to detect and remove foreign antigens from the blood. This has been described in B. rossi [65], B. canis [49], B. gibsoni [37], B. vulpes [46] and B. conradae [41]. Although it has not specifically been described in B. vogeli infections, splenectomy has been described to worsen the infection [45]. It is possible that the significantly milder disease caused by this parasite does not evoke the same degree of splenic pathology. A detailed description of splenic pathology has only been reported for B. rossi infections [65]. Some cursory comments on the splenic pathology caused by B. conradae have also been made and from these it appears that the pathology caused by B. rossi in the spleen are significantly worse than what is described for B. conradae [66].
The Babesia rossi and Babesia canis examples
Large case series have been published for both B. canis [12,33,54,67,68] and B. rossi [11]. These descriptions have allowed for a good understanding of these two infections. Sadly, there is a distinct gap in the description of the gross, histological and immunohistological pathology of both diseases although in the case of B. rossi, this is slowly being rectified. There are aspects of these two infections that appear to be fairly unique to their clinical presentation and that distinguish them from the diseases caused by the other parasites that need to be highlighted. B. rossi is usually referred to as being responsible for the most severe disease and so we will make use of the markers of disease severity for this infection to compare and contrast with the diseases caused by the other parasites.
Mortality
The greatest majority of B. rossi infected dogs presented for care that die, succumb to the infection within the first 24 hours of hospitalization, despite intensive treatment [11,63]. Time to death following admission for care has not been reported for other Babesia species besides for B. vulpes where deaths occurred within a week of presentation [27]. From this it would appear that the rate at which complicated disease develops in B. rossi infections is generally much faster than with infections with the other parasites. Mortality in B. rossi infections has been reported to range between 5 and 35% with a rate of over 80% of cerebral or hemoconcentrating cases [1]. Others have reported mortality of 45% for complicated cases with death in 10-12% of all admitted cases [69]. Lower mortality rates have also been reported with 1–3% of cases euthanized because of a grave prognosis and about 5% of all cases dying [63]. In a series of 320 cases the overall mortality rate was 11%. The odds ratios for death were significantly increased for certain complications (see below). In a large study of B. canis infections, 10% of dogs presented for care were diagnosed with multiple organ dysfunction syndrome and 67% of these died. Five percent of dogs that did not develop MODS died [67]. The mortality across all 332 dogs included in the study was around 6%—not dissimilar to the B. rossi study [11]. Most B. vogeli infected dogs are reported as being subclinical or only very mildly affected [35]. One of 11 dogs in a case series died as a result of B. vogeli infection. Severe disease (and the single death) were only seen in puppies [44]. In a Korean study of B. gibsoni, 9 (31%) of 39 dogs, were regarded as subclinical and none were reported to have had complicated disease or to have died [57]. In a study of 60 B. gibsoni infected dogs from Taiwan, the majority of dogs has mild to moderate disease, 10 dogs were severely anemic and relieved a blood transfusion and no deaths were reported [37]. In an American study of 150 cases, most were reported as mild or moderate disease and there were no reported deaths [52]. One of 9 naturally infected dogs from North Carolina (USA) died with a B. gibsoni infection [39]. In an Indian study, 10% of B. gibsoni infected dogs died whilst 34% of dual B. gibsoni/B. vogeli infected dogs died. None of the dogs with B. vogeli infection alone died [70]. In an experimental infection study, 2 of 9 B. gibsoni infected dogs died [38]. Babesia conradae can cause significant mortality with between 25-40% of naturally infected dogs dying or being euthanized because of severe illness in two separate studies [58,59]. Other studies indicate a significantly lower mortality than this [17,41] but data from large study populations involving this infection is lacking. From the available mortality data it is possible that B. rossi may more frequently be peracute/acute than B. canis although the mortality rates do not appear dissimilar enough to say with certainty that B. rossi is unequivocally more often fatal than B. canis. In a case series of 58 B. vulpes infected dogs the mortality rate was very high with 13/58 (22%) of cases dying within the first week of presenting for care. All of these appear to have succumbed to proteinuric renal failure which is likely a slowly developing glomerular pathology.
Systemic Inflammatory Response Syndrome (SIRS) and multiple Organ Dysfunction Syndrome (MODS)
The concept of the Systemic Inflammatory Response Syndrome (SIRS) was developed for use in human medicine in 1991 and was intended to provide physicians with a set of easily measurable clinical parameters that could help identify patients at risk and track host response to a wide variety of insults such assepsis [71]. These criteria were adapted for veterinary patients [72]. The usefulness of this syndrome has however been questioned [73,74]. Despite this, the veterinary criteria for the identification of SIRS in dogs have been studied in dogs infected with B. canis [67,75,76] and B. rossi [11,69]. Although the identification of this syndrome identifies dogs that are ill with Babesia, there are many cases with the syndrome that do not have complicated disease or that succumb to the infection but never qualify to be characterized as having SIRS. A more useful syndrome that identifies multiple organ dysfunction (Multiple Organ Dysfunction Syndrome—MODS) is recognized in human and veterinary medicine [77,78]. Because of the inflammatory and multiorgan nature of babesiosis in dogs, MODS has been studied in B. canis [67] and B. rossi [11,69]. It should be recognized that just because there is evidence of organ damage (such as a elevated liver or muscle enzyme activity), this does not indicate an organ dysfunction. Biochemical evidence of damage does not equate to whole organ dysfunction or failure. It is clear that the more evidence there is of the more organs that are dysfunctional or failing, the more likely a dog is to die of its infection [11,67]. There is also clear evidence for B. rossi that certain single organ failures are more predictive of death than a cluster of organs that show biochemical evidence of some level of dysfunction [11,69]. The odds ratio for death (all statistically significant) was 62.39 for cerebral disease, 32.7 for hemoconcentrating disease, 8.36 for a collapsed state at presentation, 4.9 for cases with an increased band cell count, 3.47 for cases that were hypoglycemic at presentation, 7.55 for cases with elevated creatinine and 2.89 for cases with elevated urea [11]. Neither SIRS of MODS have been convincingly identified and published for B. gibsoni, B. vogeli, B. vulpes or B. conradae infections. Cerebral disease (which carries a very poor prognosis) appears to be more common with B. rossi infections. Hemoconcentration, which also carries a very poor prognosis, has only been described in B. rossi infections. The fact that single organ failures with such high mortality rates in B. rossi infections is likely a reflection of the peracute and severe nature of this infection. The single organ failure (proteinuric renal failure) causing high mortality seen with B. vulpes infection is a slowly developing pathology [27].
Organ Pathology
In both B. rossi and B. canis infections, cerebral disease is rare and almost uniformly fatal. The cerebral pathology caused by B. rossi has been carefully described (Figure 1) [79]. There is a description of the pathology of 56 cases of cerebral babesiosis collected in South Africa over a period of 3 years that were presumably caused by B. rossi, indicating that this form of B. rossi infection may be significantly more common than what is described for B. canis [80]. The clinical effect of both infections on the brain appears to be broadly similar although the incidence of cerebral complications may be more common in B. rossi infections. There is also very little reported pathology for B. canis cerebral disease for comparison. Renal dysfunction appears to be common in both B. rossi and B. canis infections. Indeed, 91% of B. canis infected dogs identified with MODS demonstrated biochemical evidence of azotemia (elevations in creatinine). It is possible that many of these dogs had a prerenal azotemia which results in acute kidney injury, albeit potentially more easily remedied that intrinsic injuries [67]. Recoverable acute kidney injury is common in B. rossi [62] infections but elevations in creatinine are not common (just under 10% of 320 cases had elevated creatinine) [11]. This is aligns with the IRIS AKI score of I (http://www.iris-kidney.com/guidelines/grading.html). Elevations in creatinine are typically associated with a non-recoverable state of acute renal injury in B. rossi infections [11,63] and are typical of an IRIS AKI score of II, III or IV (http://www.iris-kidney.com/guidelines/grading.html).In the early work of Maegraith anuric renal failure, uremia and ‘Black Water Fever’ is described in what is now believed to be B. rossi and not B. canis infection [24]. An elevation in creatinine is a negative prognostic indicator in B rossi infections [11]. Sixteen percent (4/25) of B. rossi infected cases that died had post mortem evidence of nephrosis. Elevations in creatinine are however common in severe B. canis infection but what impact this has on outcome is not known as there is no indication of what proportion of azotemic B. canis infected dogs develop acute renal failure. A reversable protein losing nephropathy has been identified in a small number of B. gibsoni infected American pit bull terriers [81]. An IgM positive membranoproliferative glomerulonephritis (consistent with a type III hypersensitivity) has been described in B. conradae [66]. Increased serum urea concentration without a concomitant increase in creatinine is common in severe B. rossi [82]. This suggests that its origins are not reduced glomerular filtration. Increased urea has been correlated with poor outcome [11]. This has not been described for B. canis infection. Because the incidence of true acute renal failure is unknown for B. canis, it is difficult to say whether, in respect to this organ system, the infections are similar. It is likely that the renal lesion that may result from B. canis infection is common but usually reversable whilst that with results from B. rossi is less common but more likely to develop progressive azotemia rather than a reversable kidney injury. Azotemia with proteinuria appears common in B. vulpes infections with 21/58 (36%) affected in this way. This was associated 13 of these 21 affected dogs dying within the first week of presenting for care. Azotemia was significantly associated with death [27]. Acute B. rossi infections that rarely result in life-threatening AKI have quite a different pathophysiology to the azotemic and proteinuric renal failure described for the more slowly developing disease seen with B. gibsoni and B. vulpes infections.
Although liver dysfunction has been described, this is usually a reflection of modest elevations in various liver enzymes which are not necessarily a reflection of liver function. Hyperbilirubinemia and icterus are likely prehepatic in many cases due to the profound hemolytic anemia caused by infection. That said, the liver was the single most common organ showing biochemical evidence of injury in B. rossi [69] and the second most common organ affected in B. canis [67] as well as in B. gibsoni infections [37,70]. Only 1/11 dogs presenting ill with B. vogeli was icteric [44]. Icterus and hyperbilirubinemia has been reported for B. conradae however elevated liver enzyme activity appears to be rare [17,41]. The liver pathology in cases of B. rossi infection that died was fairly stereotypical and characterized by edema, bile stasis, extramedullary hematopoiesis, vacuolar hepatopathy with some centrilobular necrosis and mild to moderate macrophage and lymphocytic inflammation (Horrell H, et al., manuscript in preparation). Broadly similar pathology has been described in B. conradae [66] infections and may well also be present in B. canis infections [12]. Icterus was observed in almost two thirds of B. rossi infected dogs and elevated serum bilirubin concentrations were predictive of a poor outcome. Icterus was present on post mortem in 68% (17/25)of cases at post mortem and 16% (52/320) of a large cohort study [11]. Icterus is also reported in 80% (39/49) of B. canis infected dogs although there does not appear to correlate with outcome [33]. Icterus is also common in B. gibsoni infections with the incidence ranging from 14-25% of cases [37,51,57]. It is quite likely that the icterus and hyperbilirubinemia be mostly a pre-hepatic but, considering some of the changes seen in B. rossi infected dogs’ liver histology, there is likely also a hepatic component in the more severe and rapidly developing diseases. It is likely that the liver pathology is similar in incidence and severity for B. rossi, B. canis and B. gibsoni infections and to a lesser degree in B. conradae infections. There is no mention of icterus in B. vulpes infections.
Lung pathology that manifests clinically as hypoxemia and is diagnosed clinically as Acute Respiratory Distress Syndrome (ARDS) is rare in babesiosis but has been reported for both B. rossi [83] and B. canis [84]. There are no references to lung pathology for B. gibsoni, B. vogeli or B. conradae. In a large study of B. canis infection, 16/331 (just <5%) of dogs demonstrated diagnostic criteria consistent with ARDS. The proportion of ARDS in B. rossi infections may be higher with 18% (18/98) of cases having an arterial pO2 <60mmHg in one study [11] and 9% (3/34) in another [85]. Lung pathology in B. rossi cases that died is surprisingly common with acute interstitial pneumonia described in 64% (16/25) of cases on which post mortems were performed (Figure 1). In early work done by Maegraith et al. (now believed to be studies of B. rossi and not B. canis) 74% (25/34) of cases showed lung pathology [24]. The pathology of these lung lesions has also been described with the lesions being typified by acute interstitial pneumonia characterized by alveolar edema and hemorrhage with the inflammatory response dominated by mononuclear leukocytes in the alveolar walls and lumens [83]. There are no descriptions of the pathology caused by any other Babesia species infection. It seems likely that the lung lesions caused by B. rossi are more common and more severe than what has been described for B. canis. There is no mention of lung pathology in B vulpes infections.
Pancreatitis is described in a study of B. rossi infected dogs in which 28% of admitted dogs had a canine pancreatic lipase immunoreactivity level (cPLI) in the range diagnostic for pancreatitis. In another study, pancreatitis was histologically confirmed [86]. Pancreatitis was suspected in an old study (based on amylase and lipase concentrations) in 33% of 31 B. canis infected dogs although it was never the only organ with signs of damage [49]. In 13/46 (28%) an increased lipase DDGR was found and 8 of these were clinical for pancreatitis (including ultrasound findings) [33]. A much lower incidence (2% of 20 dogs) was described for B. gibsoni by means of canine specific pancreatic lipase activity [87]. There are no reports of pancreatitis in B. vogeli, B. vulpes, or B. conradae. The incidence of pancreatitis appears to be similar for B. rossi and B.canis but very low or not occurring in infections caused by the other parasites.
Coagulation and inflammation are inextricably linked and hence Babesia infections can be expected to result in a coagulopathy. Thrombocytopenia (sometimes severe) has been described for all Babesia species that infect dogs. Despite this, clinical hemorrhage is very rare although obvious hemorrhage was common internally during post mortem examination of dogs that died of B. rossi infections (seen in 22/25 post mortems [11]). Coagulation has been quite extensively studied in B. rossi in which disseminated intravascular coagulation (DIC) and its association with mortality has been described [88,89]. Uncomplicated cases of B. rossi showed no alteration in coagulation [90] DIC has also been described in B. canis infections [91,92]. There are no descriptions of a coagulopathy caused by other species of Babesia infecting dogs. It would appear that B. rossi and B. canis have the ability to cause an equally severe coagulopathy when clotting is evaluated clinicopathologically but at post mortem it seems possible that a far more severe coagulopathy is present with B. rossi infections.
Inflammation
The very presence of a circulating protozoal infection that results in red cell lysis would dictate that an inflammatory host response would typify a Babesia infection. Indeed, much of the pathology seen in babesiosis is arguably caused by an excessive and poorly regulated inflammatory host response: the more severe this host response, the poorer the outcome. Measures of inflammation that are clinically useful include the differential white cell count and the concentration of acute phase proteins, most notably C-reactive protein. Measuring cytokines is not clinically useful but numerous studies have evaluated these.
The differential white cells count (WCC) has been evaluated in B. rossi. Total counts are typically higher in the more severely affected dogs but often still within the normal range. Severe leukocytosis is occasionally seen [93,94]. The band cell count however was significantly higher in complicated cases and case that died [11,95]. Interestingly, dogs with complicated disease had significantly depressed CD3+, CD3+/CD4+ and CD3+/CD8+ lymphocytes in circulation [96]. This may be consistent with an immune dysregulation consistent with a state of hyperinflammation [97,98]. In B. canis the WCC are usually described as normal or low with only an occasional finding of an increased band cell count. A lymphopenia has been described but its association with disease severity or immunophenotyping have not been reported [33,49]. In B. vogeli an increase in the band cell count was described in 4/11 cases presented with illness. An increased WCC was common in the 11 sick dogs [44]. No comment is made about the WCC in other published case series of B. vogeli infection. The WCC in B. gibsoni infections are generally unremarkable [50,66] or mildly elevated due to neutrophilia with the dogs more severely affected having moderately elevated counts without remarkable changes in the band cell numbers [37]. There is a report of an obviously inflammatory WCC with an increased band cell count in a B. gibsoni dog that was euthanized with ARDS [39]. The WCC in B. conradae infected dogs is described and appears to be varied though 41% of infected dogs in one study were leukopenic [17,41]. B. vulpes does also not seem to be associated with an obvious white cell response [27]. Unfortunately there is too little data to draw clear clear conclusions about the association between inflammation reflected in the WCC and disease severity or outcome for these two infections. From these data it would seem that B. rossi causes the most severe inflammation, followed by B. canis, B. conradae and B. gibsoni which appear comparable, and then finally B. vogeli resulting in the least hematologic evidence of inflammation.
C-reactive protein (CRP) is a well-established veterinary marker of inflammation in the dog [99]. In B. rossi CRP elevates significantly during infection in concert with disease progression and decreases with resolution, but does not predict outcome [95,100]. The levels CRP reaches in B. canis infections are similar to what is described for B. rossi [33,43,44]. B. gibsoni also induces a sudden rise in CRP which coincides with the appearance of the peripheral parasitemia in experimental infections (which is very delayed compared to similar experimental infections with other parasite species) [50]. In B. vogeli CRP was elevated in 4/5 cases that presented ill [44]. CRP has not been assessed in B. conradae infections. CRP is a sensitive indicator of inflammation and elevates quickly in babesia infections and is unlikely to be a good measure to distinguish the disease severity caused by infection with the various Babesia parasites.
The role of cytokines has been quite widely investigated in the various canid Babesia infections. B. rossi is an excellent example of a cytokine storm, hyperinflammation and cytokine mediated immune dysregulation with proinflammatory cytokine levels correlating with disease severity [95,101,102]. Very similar cytokine responses have been described for B. canis infections in which complicated disease and poor outcome are associated with higher concentrations of proinflammatory cytokines [103]. Similar cytokine profiles were described in 2 experimentally B. gibsoni infected dogs but, as with CRP, the onset of increases coincided with the very delayed onset of parasitemia [50]. There are no published studies that evaluate the role of cytokines in either B. vogeli or B. conradae. It would appear that there are only small differences between the cytokine profiles in B. rossi and B. canis infected dogs making it difficult to decide if cytokine concentrations differentiate the two infections. Babesia gibsoni does however appear to drive a less virulent cytokine profile than either B. rossi or B. canis.
Evaluation of the gross, histological and immunohistochemical pathology caused by infection is another way to establish the severity of inflammation and disease. Sadly, despite the incidence of Babesia species infecting dogs in many parts of the world, the only infection that has detailed organ pathology described is B. rossi. Here organ pathology is dominated by macrophage/monocyte inflammation with some tissues showing lymphocyte accumulation. Organ damage has been described in all the organs studied thus far (spleen, bone marrow, brain, liver and lung [65,79,83,104]). There is a single report describing some pathology caused by B. conradae infection [66]. The findings for this infection were similar to B. rossi in the liver but changes that appear unique were multifocal segmental necrotizing arteritis see in the small and medium sized arteries of the gastrointestinal tract and muscle [66]. It is probable that proteinuria and azotemia described in 34% of 35 dogs with B. gibsoni infection was caused by an immune complex mediated glomerulonephritis [81]. It seems that the pathology of at least two of these infections have some things in common but some marked differences that reflect a very different pathogenesis. The paucity of basic descriptive pathology of canine Babesia infection is an important knowledge gap.
Serum Biochemistry Markers of Disease Severity
Other biochemical markers of disease severity include hypoglycemia and hyperlactatemia. Hypoglycemia in B. rossi infections is associated with a poor outcome and hyperglycemia (which is common) is less strongly correlated with a poor outcome [11,105,106]. Hypoglycemia is present in similar proportions of B. canis infected dogs but its association with outcome is unknown [107]. Hyperglycemia is not described. Hyperlactatemia that is treatment refractory is a good indicator of a poor outcome in B. rossi infections [108]. Hyperlactatemic metabolic acidosis is described in B. canis but there is no report on its association with outcome [109]. There is one report of a small number of B. gibsoni infected dogs in which hyperlactatemia was associated with a poor outcome [70]. Both B. rossi and B. canis infections can present with mixed acid base disturbances but B. rossi is associated with more severe imbalances than B. canis [85,109].The use of urea, creatinine and bilirubin have been discussed above.
Endocrine markers of disease severity
Serum cortisol and thyroxine (T4) are freely available and easily measurable in many small animal clinical settings. Hypercortisolemia and a low T4 in B. rossi infections are well correlated with disease severity and have been reported in several studies [11,110,111,112]. Very similar findings for cortisol have been reported for B. canis infections [113]. It is very likely that the endocrine reponses described for these 2 diseases are reflective of the degree of metabolic stress the dogs are enduring. It would be interesting to explore the possibility of T4 and cortisol concentration in B. gibsoni, B. vogeli and B. conradae. It is possible that these markers of stress reflect a graded disease severity.
Conclusions
B. rossi appears to cause a more severe disease than the other Babesia parasites that infect dogs. This is reflected in the speed of disease progression, mortality, nature of organ failure, the rarity of asymptomatic or waxing and waning infections, the severity of the anemia (and likely the rate of fall in hematocrit) and the degree of inflammation (host repose) evoked by an infection (Table 1 and Figure 1).
References
- Collett: M.G. Survey of canine babesiosis in South Africa. Journal of the South African Veterinary Association 2000, 71, 180-186.
- Nevils, M.A.; Figueroa, J.V.; Turk, J.R.; Canto, G.J.; Le, V.; Ellersieck, M.R.; Carson, C.A. Cloned lines of Babesia bovis differ in their ability to induce cerebral babesiosis in cattle. Parasitology research 2000, 86, 437-443. [CrossRef]
- Krause, P.J.; Daily, J.; Telford, S.R.; Vannier, E.; Lantos, P.; Spielman, A. Shared features in the pathobiology of babesiosis and malaria. Trends in parasitology 2007, 23, 605-610. [CrossRef]
- Severe falciparum malaria. World Health Organization, Communicable Diseases Cluster. Transactions of the Royal Society of Tropical Medicine and Hygiene 2000, 94 Suppl 1, S1-90.
- Anstey, N.M.; Douglas, N.M.; Poespoprodjo, J.R.; Price, R.N. Plasmodium vivax: clinical spectrum, risk factors and pathogenesis. Advances in parasitology 2012, 80, 151-201.
- S., G.L. Malaria. Clinics in Laboratory Medicine 2010, 30, 93-129.
- Rapsang, A.G.; Shyam, D.C. Scoring systems in the intensive care unit: a compendium. Indian journal of critical care medicine: peer-reviewed, official publication of Indian Society of Critical Care Medicine 2014, 18, 220.
- Ruys, L.J.; Gunning, M.; Teske, E.; Robben, J.H.; Sigrist, N.E. Evaluation of a veterinary triage list modified from a human five-point triage system in 485 dogs and cats. Journal of Veterinary Emergency and Critical Care 2012, 22, 303-312. [CrossRef]
- Hayes, G.; Mathews, K.; Kruth, S.; Doig, G.; Dewey, C. Illness severity scores in veterinary medicine: what can we learn? Journal of Veterinary Internal Medicine 2010, 24, 457-466.
- Goggs, R.; Dennis, S.; Di Bella, A.; Humm, K.R.; McLauchlan, G.; Mooney, C.; Ridyard, A.; Tappin, S.; Walker, D.; Warman, S. Predicting outcome in dogs with primary immune-mediated hemolytic anemia: results of a multicenter case registry. Journal of Veterinary Internal Medicine 2015, 29, 1603-1610. [CrossRef]
- Leisewitz, A.L.; Goddard, A.; Clift, S.; Thompson, P.N.; de Gier, J.; Van Engelshoven, J.; Schoeman, J.P. A clinical and pathological description of 320 cases of naturally acquired Babesia rossi infection in dogs. Veterinary parasitology 2019, 271, 22-30. [CrossRef]
- Mathe, A.; Voros, K.; Papp, L.; Reiczigel, J. Clinical manifestations of canine babesiosis in Hungary (63 cases). Acta Vet Hung 2006, 54, 367-385. [CrossRef]
- Jackson, A.P.; Otto, T.D.; Darby, A.; Ramaprasad, A.; Xia, D.; Echaide, I.E.; Farber, M.; Gahlot, S.; Gamble, J.; Gupta, D. The evolutionary dynamics of variant antigen genes in Babesia reveal a history of genomic innovation underlying host–parasite interaction. Nucleic acids research 2014, 42, 7113-7131. [CrossRef]
- Matjila, P.T.; Carcy, B.; Leisewitz, A.L.; Schetters, T.; Jongejan, F.; Gorenflot, A.; Penzhorn, B.L. Preliminary evaluation of the BrEMA1 gene as a tool for associating babesia rossi genotypes and clinical manifestation of canine Babesiosis. Journal of clinical microbiology 2009, 47, 3586-3592. [CrossRef]
- Gazzinelli, R.T.; Denkers, E.Y. Protozoan encounters with Toll-like receptor signalling pathways: implications for host parasitism. Nature Reviews Immunology 2006, 6, 895-906. [CrossRef]
- Leoratti, F.M.; Farias, L.; Alves, F.P.; Suarez-Mútis, M.C.; Coura, J.R.; Kalil, J.; Camargo, E.P.; Moraes, S.L.; Ramasawmy, R. Variants in the toll-like receptor signaling pathway and clinical outcomes of malaria. The Journal of infectious diseases 2008, 198, 772-780. [CrossRef]
- Conrad, P.; Thomford, J.; Yamane, I.; Whiting, J.; Bosma, L.; Uno, T.; Holshuh, H.J.; Shelly, S. Hemolytic anemia caused by Babesia gibsoni infection in dogs. Journal of the American Veterinary Medical Association 1991, 199, 601-605.
- Kjemtrup, A.M.; Conrad, P.A. A review of the small canine piroplasms from California: Babesia conradae in the literature. Veterinary parasitology 2006, 138, 112-117. [CrossRef]
- Malherbe, W.D.; Immelman, A.; Haupt, W.H.; Walzl, H.J. The diagnosis and treatment of acid-base deranged dogs infected with Babesia canis. Journal of the South African Veterinary Association 1976, 47, 29-33.
- Malherbe, W.D. A clinico-pathological study of Babesia canis infection in dogs. University of Pretoria, Pretoria, 1968.
- Malherbe, W.D.; Parkin, B.S. Atypical symptomatology in Babesia canis infection. Journal of the South African Veterinary Association 1951, 22, 25-36.
- Uilenberg, G.; Franssen, F.F.; Perie, N.M.; Spanjer, A.A. Three groups of Babesia canis distinguished and a proposal for nomenclature. The Veterinary quarterly 1989, 11, 33-40. [CrossRef]
- Carret, C.; Walas, F.; Carcy, B.; Grande, N.; Precigout, E.; Moubri, K.; Schetters, T.P.; Gorenflot, A. Babesia canis canis, Babesia canis vogeli, Babesia canis rossi: differentiation of the three subspecies by a restriction fragment length polymorphism analysis on amplified small subunit ribosomal RNA genes. J Eukaryot Microbiol 1999, 46, 298-303. [CrossRef]
- Maegraith, B.; Gilles, H.M.; Devakul, K. Pathological processes in Babesia canis infections. Zeitschrift fur Tropenmedizin und Parasitologie 1957, 8, 485-514.
- Baneth, G.; Cardoso, L.; Brilhante-Simões, P.; Schnittger, L. Establishment of Babesia vulpes n. sp.(Apicomplexa: Babesiidae), a piroplasmid species pathogenic for domestic dogs. Parasites & vectors 2019, 12, 1-8. [CrossRef]
- Barash, N.R.; Thomas, B.; Birkenheuer, A.J.; Breitschwerdt, E.B.; Lemler, E.; Qurollo, B.A. Prevalence of Babesia spp. and clinical characteristics of Babesia vulpes infections in North American dogs. Journal of veterinary internal medicine 2019, 33, 2075-2081.
- Camacho, A.T.; Guitian, F.J.; Pallas, E.; Gestal, J.J.; Olmeda, A.S.; Goethert, H.K.; III, S.R.T.; Spielman, A. Azotemia and mortality among Babesia microti-like infected dogs. Journal of veterinary internal medicine 2004, 18, 141-146. [CrossRef]
- Miró, G.; Checa, R.; Paparini, A.; Ortega, N.; González-Fraga, J.L.; Gofton, A.; Bartolomé, A.; Montoya, A.; Gálvez, R.; Mayo, P.P. Theileria annae (syn. Babesia microti-like) infection in dogs in NW Spain detected using direct and indirect diagnostic techniques: clinical report of 75 cases. Parasites & vectors 2015, 8, 1-11. [CrossRef]
- Simões, P.B.; Cardoso, L.; Araújo, M.; Yisaschar-Mekuzas, Y.; Baneth, G. Babesiosis due to the canine Babesia microti-like small piroplasm in dogs-first report from Portugal and possible vertical transmission. Parasites & vectors 2011, 4, 1-6. [CrossRef]
- Zahler, M.; Rinder, H.; Schein, E.; Gothe, R. Detection of a new pathogenic Babesia microti-like species in dogs. Veterinary parasitology 2000, 89, 241-248. [CrossRef]
- Birkenheuer, A.J.; Neel, J.; Ruslander, D.; Levy, M.; Breitschwerdt, E. Detection and molecular characterization of a novel large Babesia species in a dog. Veterinary parasitology 2004, 124, 151-160. [CrossRef]
- Morters, M.K.; Archer, J.; Ma, D.; Matthee, O.; Goddard, A.; Leisewitz, A.L.; Matjila, P.T.; Wood, J.L.N.; Schoeman, J.P. Long-term follow-up of owned, free-roaming dogs in South Africa naturally exposed to Babesia rossi. International journal for parasitology 2020, 50, 103-110. [CrossRef]
- Weingart, C.; Helm, C.S.; Müller, E.; Schäfer, I.; Skrodzki, M.; von Samson-Himmelstjerna, G.; Krücken, J.; Kohn, B. Autochthonous Babesia canis infections in 49 dogs in Germany. Journal of Veterinary Internal Medicine 2023. [CrossRef]
- Rojas, A.; Rojas, D.; Montenegro, V.; Gutiérrez, R.; Yasur-Landau, D.; Baneth, G. Vector-borne pathogens in dogs from Costa Rica: first molecular description of Babesia vogeli and Hepatozoon canis infections with a high prevalence of monocytic ehrlichiosis and the manifestations of co-infection. Veterinary parasitology 2014, 199, 121-128. [CrossRef]
- Di Cataldo, S.; Ulloa-Contreras, C.; Cevidanes, A.; Hernandez, C.; Millan, J. Babesia vogeli in dogs in Chile. Transboundary and emerging diseases 2020, 67, 2296-2299. [CrossRef]
- Beck, R.; Vojta, L.; Mrljak, V.; Marinculić, A.; Beck, A.; Živičnjak, T.; Cacciò, S.M. Diversity of Babesia and Theileria species in symptomatic and asymptomatic dogs in Croatia. International journal for parasitology 2009, 39, 843-848. [CrossRef]
- Liu, P.-C.; Lin, C.-N.; Su, B.-L. Clinical characteristics of naturally Babesia gibsoni infected dogs: A study of 60 dogs. Veterinary Parasitology: Regional Studies and Reports 2022, 28, 100675. [CrossRef]
- Botros, B.A.; Moch, R.W.; Barsoum, I.S. Some observations on experimentally induced infection of dogs with Babesia gibsoni. American journal of veterinary research 1975, 36, 293-296.
- Birkenheuer, A.J.; Levy, M.G.; Savary, K.C.; Gager, R.B.; Breitschwerdt, E.B. Babesia gibsoni infections in dogs from North Carolina. Journal of the American Animal Hospital Association 1999, 35, 125-128. [CrossRef]
- Macintire, D.K.; Boudreaux, M.K.; West, G.D.; Bourne, C.; Wright, J.C.; Conrad, P.A. Babesia gibsoni infection among dogs in the southeastern United States. Journal of the American Veterinary Medical Association 2002, 220, 325-329. [CrossRef]
- Dear, J.D.; Owens, S.D.; Lindsay, L.L.; Biondo, A.W.; Chomel, B.B.; Marcondes, M.; Sykes, J.E. Babesia conradae infection in coyote hunting dogs infected with multiple blood-borne pathogens. Journal of veterinary internal medicine 2018, 32, 1609-1617. [CrossRef]
- Groves, M.G.; Dennis, G.L. Babesia gibsoni: field and laboratory studies of canine infections. Experimental parasitology 1972, 31, 153-159. [CrossRef]
- Matijatko, V.; Mrljak, V.; Kis, I.; Kucer, N.; Forsek, J.; Zivicnjak, T.; Romic, Z.; Simec, Z.; Ceron, J.J. Evidence of an acute phase response in dogs naturally infected with Babesia canis. Veterinary parasitology 2007, 144, 242-250. [CrossRef]
- Solano-Gallego, L.; Trotta, M.; Carli, E.; Carcy, B.; Caldin, M.; Furlanello, T. Babesia canis canis and Babesia canis vogeli clinicopathological findings and DNA detection by means of PCR-RFLP in blood from Italian dogs suspected of tick-borne disease. Veterinary parasitology 2008, 157, 211-221. [CrossRef]
- Wang, J.; Zhang, J.; Kelly, P.; Zheng, X.; Li, M.; You, J.; Huang, K.; Qiu, H.; Wang, Y.; Zhang, R. First description of the pathogenicity of Babesia vogeli in experimentally infected dogs. Veterinary parasitology 2018, 253, 1-7. [CrossRef]
- Arsenault, A.C.; Foley, P.M.; Clancey, N.P. Babesia vulpes in a dog from Prince Edward Island, Canada. The Canadian Veterinary Journal 2022, 63, 589.
- Radyuk, E.; Karan, L. A case of Babesia vulpes infection in a dog in Russia. Veterinary Parasitology: Regional Studies and Reports 2020, 22, 100467.
- Oakley, M.S.; Gerald, N.; McCutchan, T.F.; Aravind, L.; Kumar, S. Clinical and molecular aspects of malaria fever. Trends in parasitology 2011, 27, 442-449. [CrossRef]
- Máthé, A.; Vörös, K.; Papp, L.; Reiczigel, J. Clinical manifestations of canine babesiosis in Hungary (63 cases). Acta Veterinaria Hungarica 2006, 54, 367-385. [CrossRef]
- Brown, A.L.; Shiel, R.E.; Irwin, P.J. Clinical, haematological, cytokine and acute phase protein changes during experimental Babesia gibsoni infection of beagle puppies. Experimental parasitology 2015, 157, 185-196. [CrossRef]
- Bilwal, A.K.; Mandali, G.C.; Tandel, F.B. Clinicopathological alterations in naturally occurring Babesia gibsoni infection in dogs of Middle-South Gujarat, India. Vet World 2017, 10, 1227-1232. [CrossRef]
- Birkenheuer, A.J.; Correa, M.T.; Levy, M.G.; Breitschwerdt, E.B. Geographic distribution of babesiosis among dogs in the United States and association with dog bites: 150 cases (2000-2003). Journal of the American Veterinary Medical Association 2005, 227, 942-947. [CrossRef]
- Crnogaj, M.; Cerón, J.J.; Šmit, I.; Kiš, I.; Gotić, J.; Brkljačić, M.; Matijatko, V.; Rubio, C.P.; Kučer, N.; Mrljak, V. Relation of antioxidant status at admission and disease severity and outcome in dogs naturally infected with Babesia canis canis. BMC Veterinary Research 2017, 13, 1-9. [CrossRef]
- Zygner, W.; Gójska, O.; Rapacka, G.; Jaros, D.; Wędrychowicz, H. Hematological changes during the course of canine babesiosis caused by large Babesia in domestic dogs in Warsaw (Poland). Veterinary parasitology 2007, 145, 146-151. [CrossRef]
- Zaki, A.A.; Attia, M.M.; Ismael, E.; Mahdy, O.A. Prevalence, genetic, and biochemical evaluation of immune response of police dogs infected with Babesia vogeli. Veterinary World 2021, 14, 903. [CrossRef]
- Gülanber, A.; Gorenflot, A.; Schetters, T.P.; Carcy, B. First molecular diagnosis of Babesia vogeli in domestic dogs from Turkey. Veterinary parasitology 2006, 139, 224-230.
- Lee, M.J.; Yu, D.H.; Yoon, J.S.; Li, Y.H.; Lee, J.H.; Chae, J.S.; Park, J. Epidemiologic and clinical surveys in dogs infected with Babesia gibsoni in South Korea. Vector Borne Zoonotic Dis 2009, 9, 681-686. [CrossRef]
- Meinkoth, J.H.; Kocan, A.A.; Loud, S.D.; Lorenz, M.D. Clinical and hematologic effects of experimental infection of dogs with recently identified Babesia gibsoni-like isolates from Oklahoma. Journal of the American Veterinary Medical Association 2002, 220, 185-189. [CrossRef]
- Di Cicco, M.F.; Downey, M.E.; Beeler, E.; Marr, H.; Cyrog, P.; Kidd, L.; Diniz, P.P.V.; Cohn, L.A.; Birkenheuer, A.J. Re-emergence of Babesia conradae and effective treatment of infected dogs with atovaquone and azithromycin. Veterinary parasitology 2012, 187, 23-27. [CrossRef]
- Bodi, J.; Nsibu, C.; Hirayama, K. Immunogenetic mechanisms of black water fever: article review. Gene Technol 2021, 10, 1-8.
- Smith, R.L.; Goddard, A.; Boddapati, A.; Brooks, S.; Schoeman, J.P.; Lack, J.; Leisewitz, A.; Ackerman, H. Experimental Babesia rossi infection induces hemolytic, metabolic, and viral response pathways in the canine host. BMC genomics 2021, 22, 1-16.
- Defauw, P.; Schoeman, J.P.; Leisewitz, A.L.; Goddard, A.; Duchateau, L.; Aresu, L.; Meyer, E.; Daminet, S. Evaluation of acute kidney injury in dogs with complicated or uncomplicated Babesia rossi infection. Ticks Tick Borne Dis 2020, 11, 101406. [CrossRef]
- Jacobson, L.S. The South African form of severe and complicated canine babesiosis: clinical advances 1994-2004. Veterinary parasitology 2006, 138, 126-139. [CrossRef]
- Wang, J.; Applefeld, W.N.; Sun, J.; Solomon, S.B.; Feng, J.; Couse, Z.G.; Risoleo, T.F.; Danner, R.L.; Tejero, J.; Lertora, J. Mechanistic Insights into Cell-free Hemoglobin-induced Injury During Septic Shock. American Journal of Physiology-Heart and Circulatory Physiology 2021. [CrossRef]
- Henning, A.; Clift, S.J.; Leisewitz, A.L. The pathology of the spleen in lethal canine babesiosis caused by Babesia rossi. Parasite immunology 2020, 42, e12706. [CrossRef]
- Wozniak, E.J.; Barr, B.C.; Thomford, J.W.; Yamane, I.; McDonough, S.P.; Moore, P.F.; Naydan, D.; Robinson, T.W.; Conrad, P.A. Clinical, anatomic, and immunopathologic characterization of Babesia gibsoni infection in the domestic dog (Canis familiaris). The Journal of parasitology 1997, 83, 692-699. [CrossRef]
- Matijatko, V.; Kiš, I.; Torti, M.; Brkljačić, M.; Barić Rafaj, R.; Žvorc, Z.; Mrljak, V. Systemic inflammatory response syndrome and multiple organ dysfunction syndrome in canine babesiosis. Veterinarski Arhiv 2010, 80, 611-626.
- Adaszek, Ł.; Winiarczyk, S.; Skrzypczak, M. The clinical course of babesiosis in 76 dogs infected with protozoan parasites Babesia canis canis. Polish Journal of Veterinary Sciences 2009, 12, 81-87.
- Welzl, C.; Leisewitz, A.L.; Jacobson, L.S.; Vaughan-Scott, T.; Myburgh, E. Systemic inflammatory response syndrome and multiple-organ damage/dysfunction in complicated canine babesiosis. Journal of the South African Veterinary Association 2001, 72, 158-162. [CrossRef]
- Kandasamy, R.; Venkatasubramanian, L.; Loganathasamy, K.; Latha, B.R.; Mani, B. Prognostic markers and their discriminant score in predicting the outcome of Babesia gibsoni infection. Veterinary Record 2021, 188, e29. [CrossRef]
- Bone, R.C.; Sibbald, W.J.; Sprung, C.L. The ACCP-SCCM consensus conference on sepsis and organ failure. Chest 1992, 101, 1481-1483. [CrossRef]
- Okano, S.; Yoshida, M.; Fukushima, U.; Higuchi, S.; Takase, K.; Hagio, M. Usefulness of systemic inflammatory response syndrome criteria as an index for prognosis judgement. The Veterinary record 2002, 150, 245. [CrossRef]
- Vincent, J.-L. Dear SIRS, I’m sorry to say that I don’t like you. Critical care medicine 1997, 25, 372-374.
- Balk, R.A. Systemic inflammatory response syndrome (SIRS): where did it come from and is it still relevant today? Virulence 2014, 5, 20-26. [CrossRef]
- Beletić, A.; Janjić, F.; Radaković, M.; Spariosu, K.; Andrić, J.F.; Chandrashekar, R.; Tyrrell, P.; Radonjić, V.; Balint, B.; Ajtić, J. Systemic inflammatory response syndrome in dogs naturally infected with Babesia canis: Association with the parasite load and host factors. Veterinary parasitology 2021, 291, 109366.
- Schetters, T.P.M.; Kleuskens, J.A.G.M.; Van De Crommert, J.; De Leeuw, P.W.J.; Finizio, A.L.; Gorenflot, A. Systemic inflammatory responses in dogs experimentally infected with Babesia canis; a haematological study. Veterinary parasitology 2009, 162, 7-15. [CrossRef]
- Gourd, N.M.; Nikitas, N. Multiple organ dysfunction syndrome. Journal of intensive care medicine 2020, 35, 1564-1575.
- Osterbur, K.; Mann, F.; Kuroki, K.; DeClue, A. Multiple organ dysfunction syndrome in humans and animals. Journal of veterinary internal medicine 2014, 28, 1141-1151. [CrossRef]
- Lewis, M. Histopathology of cerebral babesiosis in dogs with naturally acquired Babesia rossi infection. University of Pretoria, Pretoria, South Africa, 2022.
- Pardini, D.A. The pathology and pathogenesis of canine cerebral babesiosis. University of Pretoria, Pretoria, South Africa, 2000.
- Ullal, T.; Birkenheuer, A.; Vaden, S. Azotemia and Proteinuria in Dogs Infected with Babesia gibsoni. Journal of the American Animal Hospital Association 2018, 54, 156-160. [CrossRef]
- de Scally, M.P.; Leisewitz, A.L.; Lobetti, R.G.; Thompson, P.N. The elevated serum urea:creatinine ratio in canine babesiosis in South Africa is not of renal origin. Journal of the South African Veterinary Association 2006, 77, 175-178. [CrossRef]
- Martin, C.; Clift, S.; Leisewitz, A. Lung pathology of natural Babesia rossi infection in dogs. Journal of the South African Veterinary Association 2023, 94, 59-69. [CrossRef]
- DeStefano, I.; Wayne, A.; Cudney, S.; Rozanski, E. Successful treatment of suspect Babesia-induced ARDS in a dog using lung-protective positive-pressure ventilation and neuromuscular blockade. Authorea Preprints 2022.
- Leisewitz, A.L.; Jacobson, L.S.; de Morais, H.S.; Reyers, F. The mixed acid-base disturbances of severe canine babesiosis. Journal of veterinary internal medicine/American College of Veterinary Internal Medicine 2001, 15, 445-452.
- Mohr, A.J.; Lobetti, R.G.; van der Lugt, J.J. Acute pancreatitis: a newly recognised potential complication of canine babesiosis. Journal of the South African Veterinary Association 2000, 71, 232-239. [CrossRef]
- Masuda, M.; Otsuka-Yamasaki, Y.; Shiranaga, N.; Iguchi, A.; Uchida, N.; Sato, R.; Yamasaki, M. Retrospective study on intercurrent pancreatitis with Babesia gibsoni infection in dogs. Journal of Veterinary Medical Science 2019, 81, 1558-1563. [CrossRef]
- Goddard, A.; Wiinberg, B.; Schoeman, J.P.; Kristensen, A.T.; Kjelgaard-Hansen, M. Mortality in virulent canine babesiosis is associated with a consumptive coagulopathy. Vet J 2012. [CrossRef]
- Goddard, A.; Wiinberg, B.; Schoeman, J.P.; Kristensen, A.T.; Kjelgaard-Hansen, M. Mortality in virulent canine babesiosis is associated with a consumptive coagulopathy. Vet J 2013, 196, 213-217. [CrossRef]
- Liebenberg, C.; Goddard, A.; Wiinberg, B.; Kjelgaard-Hansen, M.; van der Merwe, L.L.; Thompson, P.N.; Matjila, P.T.; Schoeman, J.P. Hemostatic abnormalities in uncomplicated babesiosis (Babesia rossi) in dogs. Journal of veterinary internal medicine/American College of Veterinary Internal Medicine 2013, 27, 150-156. [CrossRef]
- Dubova, O.; Feshchenko, D.; Bakhur, T.; Zghozinska, O.; Antipov, A.; Rublenko, S.; Goncharenko, V.; Shahanenko, R.; Shahanenko, V. Disseminated intravascular coagulation syndrome as a complication in acute spontaneous canine babesiosis. 2020. [CrossRef]
- de Gopegui, R.R.; Peñalba, B.; Goicoa, A.; Espada, Y.; Fidalgo, L.E.; Espino, L. Clinico-pathological findings and coagulation disorders in 45 cases of canine babesiosis in Spain. The veterinary journal 2007, 174, 129-132. [CrossRef]
- Weltan, S.M.; Leisewitz, A.L.; Goddard, A. A case-controlled retrospective study of the causes and implications of moderate to severe leukocytosis in dogs in South Africa. Veterinary clinical pathology/American Society for Veterinary Clinical Pathology 2008, 37, 164-172. [CrossRef]
- Lobetti, R. Leukaemoid response in two dogs with Babesia canis infection. Journal of the South African Veterinary Association 1995, 66, 182-184.
- Atkinson, B.K.; Thompson, P.; Van Zyl, E.; Goddard, A.; Rautenbach, Y.; Schoeman, J.P.; Mukorera, V.; Leisewitz, A. Kinetics of the inflammatory response during experimental Babesia rossi infection of beagle dogs. Veterinary parasitology 2022, 109717. [CrossRef]
- Rautenbach, Y.A.G.P.N.T.R.J.M.; Leisewitz, A.L. A flow cytometric assessment of the lymphocyte immunophenotypes in dogs naturally infected with Babesia rossi. Veterinary parasitology 2017, 241, 26-34. [CrossRef]
- Yadav, H.; Cartin-Ceba, R. Balance between Hyperinflammation and Immunosuppression in Sepsis. Semin Respir Crit Care Med 2016, 37, 42-50. [CrossRef]
- Inoue, M.; Shinohara, M.L. Hyperinflammation, T cells, and endotoxemia. Oncotarget 2015, 6, 23040-23041. [CrossRef]
- Christensen, M.B.; Langhorn, R.; Goddard, A.; Andreasen, E.B.; Moldal, E.; Tvarijonaviciute, A.; Kirpensteijn, J.; Jakobsen, S.; Persson, F.; Kjelgaard-Hansen, M. Comparison of serum amyloid A and C-reactive protein as diagnostic markers of systemic inflammation in dogs. The Canadian Veterinary Journal 2014, 55, 161.
- Koster, L.S.; Van Schoor, M.; Goddard, A.; Thompson, P.N.; Matjila, P.T.; Kjelgaard-Hansen, M. C-reactive protein in canine babesiosis caused by Babesia rossi and its association with outcome. Journal of the South African Veterinary Association 2009, 80, 87-91. [CrossRef]
- Leisewitz, A.; Goddard, A.; De Gier, J.; Van Engelshoven, J.; Clift, S.; Thompson, P.; Schoeman, J.P. Disease severity and blood cytokine concentrations in dogs with natural Babesia rossi infection. Parasite immunology 2019, 41, e12630. [CrossRef]
- Goddard, A.; Leisewitz, A.L.; Kjelgaard-Hansen, M.; Kristensen, A.T.; Schoeman, J.P. Excessive Pro-Inflammatory Serum Cytokine Concentrations in Virulent Canine Babesiosis. PloS one 2016, 11, e0150113. [CrossRef]
- Galan, A.; Mayer, I.; Rafaj, R.B.; Bendelja, K.; Susic, V.; Ceron, J.J.; Mrljak, V. MCP-1, KC-like and IL-8 as critical mediators of pathogenesis caused by Babesia canis. PloS one 2018, 13, e0190474. [CrossRef]
- Bumby, M.M. Cytological and histopathological bone marrow findings in dogs with natural Babesia rossi infection. University of Pretoria, Pretoria, 2021.
- Rees, P.; Schoeman, J.P. Plasma insulin concentrations in hypoglycaemic dogs with Babesia canis rossi infection. Veterinary parasitology 2008, 152, 60-66. [CrossRef]
- Keller, N.; Jacobson, L.S.; Nel, M.; de Clerq, M.; Thompson, P.N.; Schoeman, J.P. Prevalence and risk factors of hypoglycemia in virulent canine babesiosis. Journal of veterinary internal medicine/American College of Veterinary Internal Medicine 2004, 18, 265-270.
- Zygner, W.; Rapacka, G.; Gojska-Zygner, O.; Dlugosz, E.; Wedrychowicz, H. Biochemical abnormalities observed in serum of dogs infected with large Babesia in Warsaw (Poland). Pol J Vet Sci 2007, 10, 245-253.
- Nel, M.; Lobetti, R.G.; Keller, N.; Thompson, P.N. Prognostic value of blood lactate, blood glucose, and hematocrit in canine babesiosis. Journal of veterinary internal medicine/American College of Veterinary Internal Medicine 2004, 18, 471-476.
- Torti, M.; Kuleš, J.; Matijatko, V.; Brkljačić, M.; Kiš, I.; Gotić, J.; Mrljak, V.; Šmit, I. Acid-base status in canine babesiosis caused by Babesia canis. Veterinarski arhiv 2020, 90, 603-610. [CrossRef]
- Schoeman, J.P.; Herrtage, M.E. Adrenal response to the low dose ACTH stimulation test and the cortisol-to-adrenocorticotrophic hormone ratio in canine babesiosis. Veterinary parasitology 2008, 154, 205-213. [CrossRef]
- Schoeman, J.P.; Herrtage, M.E. Serum thyrotropin, thyroxine and free thyroxine concentrations as predictors of mortality in critically ill puppies with parvovirus infection: a model for human paediatric critical illness? Microbes and infection/Institut Pasteur 2008, 10, 203-207. [CrossRef]
- Schoeman, J.P.; Rees, P.; Herrtage, M.E. Endocrine predictors of mortality in canine babesiosis caused by Babesia canis rossi. Veterinary parasitology 2007, 148, 75-82. [CrossRef]
- Matijatko, V.; Torti, M.; Kiš, I.; Šmit, I.; Štoković, I.; Vranješ-Đurić, S.; Milanović, S.; Mrljak, V.; Brkljačić, M. Serum cor tisol and insulin concentrations in dogs naturally infected Serum cor tisol and insulin concentrations in dogs naturally infected with Babesia canis. Veterinarski arhiv 2014, 84, 551-562.
Figure 1.
Infection with Babesia rossi causes severe disease. (A): Collapse at presentation is an indication of severe disease and increases the odds of a poor outcome. (B): Blood transfusion is a common form of supportive treatment required in the majority of dogs admitted to a hospital for care. (C): Overt signs of intravascular hemolysis are common resulting in hemoglobinemia and (D) hemoglobinuria. (E): Acute Respiratory Distress Syndrome is associated with a significant risk of death as evidenced here by a blood tinged froth pouring from the nose of an agonal case of B. rossi infection. (F): On post mortem examination lungs are characterized grossly as edematous with extensive hemorrhage. (G): Cerebral babesiosis is uncommon but results in hemorrhagic encephalomalacia. (H): Histologically classic ring hemorrhages around small vessels are frequently observed.
Figure 1.
Infection with Babesia rossi causes severe disease. (A): Collapse at presentation is an indication of severe disease and increases the odds of a poor outcome. (B): Blood transfusion is a common form of supportive treatment required in the majority of dogs admitted to a hospital for care. (C): Overt signs of intravascular hemolysis are common resulting in hemoglobinemia and (D) hemoglobinuria. (E): Acute Respiratory Distress Syndrome is associated with a significant risk of death as evidenced here by a blood tinged froth pouring from the nose of an agonal case of B. rossi infection. (F): On post mortem examination lungs are characterized grossly as edematous with extensive hemorrhage. (G): Cerebral babesiosis is uncommon but results in hemorrhagic encephalomalacia. (H): Histologically classic ring hemorrhages around small vessels are frequently observed.
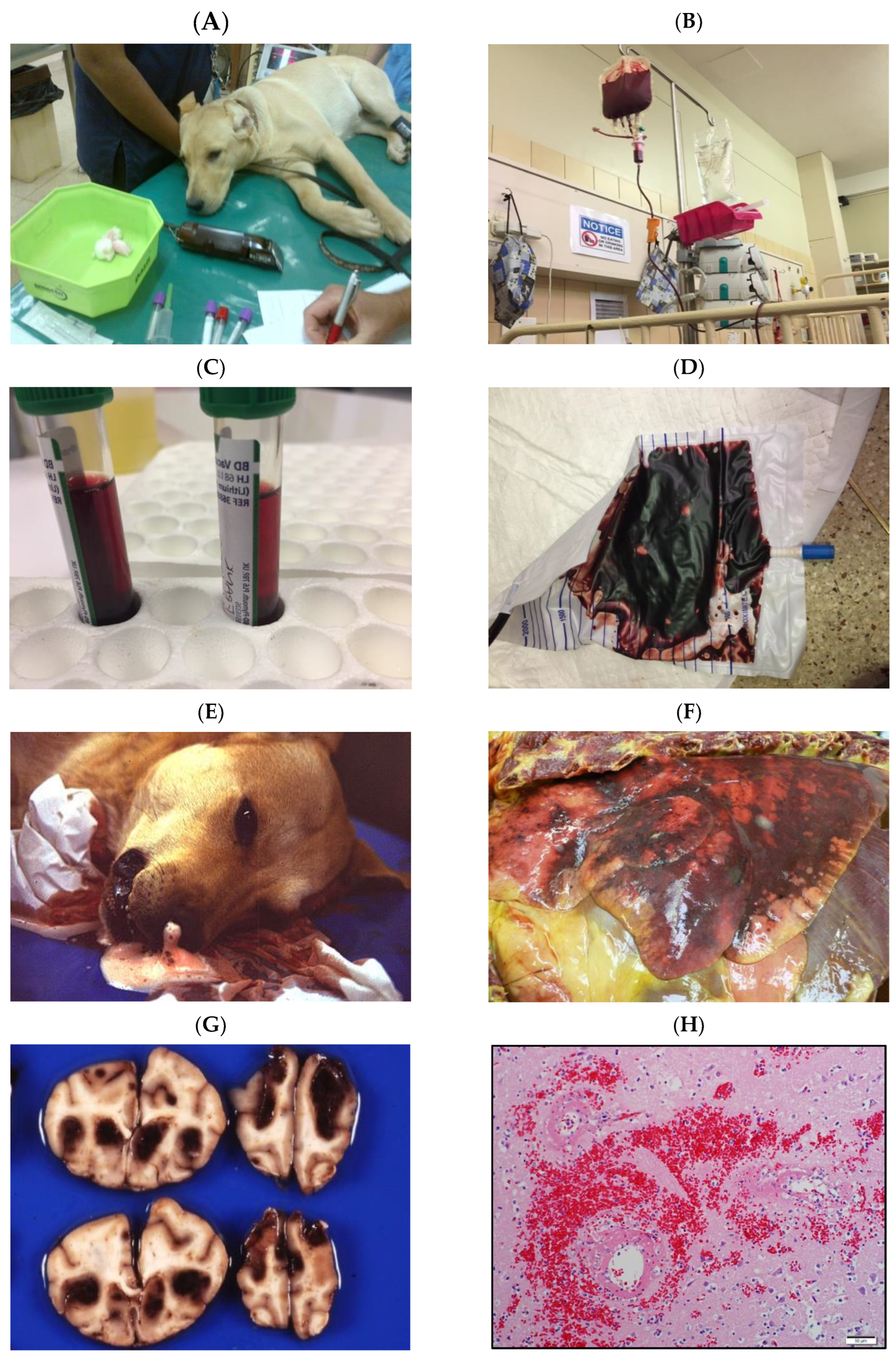

Table 1.
Comparing the clinical measures of disease severity between Babesia rossi, Babesia canis, Babesia gibsoni, Babesia vogeli, Babesia vulpes and Babesia conradae. SIRS: systemic inflammatory response syndrome; MODS: multiple organ dysfunction syndrome.
Table 1.
Comparing the clinical measures of disease severity between Babesia rossi, Babesia canis, Babesia gibsoni, Babesia vogeli, Babesia vulpes and Babesia conradae. SIRS: systemic inflammatory response syndrome; MODS: multiple organ dysfunction syndrome.
| Measure of severity | B. rossi | B. canis | B. gibsoni | B. conradea | B. vulpes | B. vogeli |
|---|---|---|---|---|---|---|
| Mortality | 5-35% (multiple studies, hundreds of cases) | 1.5-20% (multiple studies, hundreds of cases) | Low (multiple studies, hundreds of cases) | 25-40% (two small studies) | 22% (one study of 58 cases) | Very low |
| Complicated disease seen | Yes, around 20% of 320 cases | Yes, around 10% of 226 cases. | Occasionally | Occasionally | Unlikely | Very rarely |
| SIRS | Well reported but poorly correlated with outcome | Well reported but poorly correlated with outcome | Not reported but seems possible | Not reported but seems possible | Unlikely | Very rare |
| Single organ failure or MODS | Catastrophic single organ failure of the brain, lungs or kidney are uncommon but carry a very high mortality. MODS well described with much poorer outcomes |
Catastrophic single organ failure of the brain, lungs or kidney are less common than with B. rossi and carry a high mortality. | Evidence of organ dysfunction but catastrophic single organ failure or MODS appears rare. | Not reported but seems possible | Evidence of proteinuric renal failure being the most common cause of death | Very rare. |
| Disease course | Rapidly evolving disease common with most deaths occurring within 24 hours of hospital admission | Disease evolved less acutely than B. rossi | Chronic and waxing and waning disease well described | Appears to be sub-acute | Probably chronic | Infection rarely causes disease |
| Asymptomatic or Self resolving infection | Very rare | Rare | Common | Unknown | Very likely | Asymptomatic infection very common |
| Anemia | Severe anemia is common with blood transfusions frequently required as part of management | Severe anemia is less common and blood transfusions are not usual as part of the management | Severe anemia possible but rare. Blood transfusions rarely required as part of the treatment | Severe anemia possible | Usually mild to moderate | Severe anemia very rare. Blood transfusions not described as part of treatment |
| Extravascular hemolysis | Clinically overt and common | Clinically overt and common | Occurs rarely | Is described | Not described | Occurs rarely |
| Inflammation | Very obviously a profoundly inflammatory disease | Infection results in inflammation but less severe than B. rossi | Evidence of inflammation but less severe and of a more chronic nature | Likely to be moderately inflammatory | Not desxcribed | Host damaging inflammation very rare |
| Hallmarks of poor outcome | Collapse Single organ failure (brain, lung, kidney) Collapse Low body temperature Increased band cell count Elevations in urea (independent of creatinine), creatinine, bilirubin, lactate. Hypoglycemia Increased cortisol, depressed thyroid hormone |
The presence of MODS | Low hemoglobin and elevations in urea, creatine, ALT, ALP and lactate. | Not known | Proteinuria and azotemia | Not known |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



