
Submitted:
05 September 2023
Posted:
07 September 2023
You are already at the latest version
Abstract
Agriculture (crop production, land use, and livestock) is the second most important greenhouse gas (GHG) emitting sector after the energy sector. Agriculture is also recognized as the source and sink of GHGs. Evidence demonstrates that the application of high amounts of nitrogen-rich fertilizers enhances methane (CH4) and nitrous oxide (N2O) emissions, which are potent GHGs with a high global warming potential (GWP). Considering its global contribution to the climate crisis, reducing GHG emissions in agriculture would considerably lower its share of the global GHG emission records, which may lead to enormous benefits for the environment and food production systems. Several diverging and controversial views questioning the actual role of plants in the current global GHG budget continue to nourish the debate globally. We must acknowledge that considering the beneficial roles of major GHGs to plants at a certain level of accumulation, implementing GHG mitigation measures from agriculture is indeed a complex task. This review seeks to provide key approaches for GHG mitigation in the literature (environmentally friendly crop cultivation and residue management practices, improvement of plants nutrients/fertilizer use efficiency, exploring the genetic diversity for low GHG emission, soil methane-producing bacteria, integrated soil fertility management, improved livestock feeding efficiency, and production, etc.). This work gathers key approaches from 275 peer-reviewed publications, including experimental research papers, review articles, and books, discussing greenhouse gas emissions mechanisms and mitigation to unravel effective strategies for GHG mitigation, proven to be effective or carry the potential to mitigate GHG generation from agriculture. This review also discusses in depth the significance and the dynamics of strategies regarded as game changers with a high potential to enhance, in a sustainable manner, the resilience of agricultural systems. Agricultural GHG mitigation approaches discussed in this work can serve as game changers in global efforts to reducing GHG emissions and alleviating the impact of climate change through sustainable agriculture and informed-decision making.
Keywords:
nitrogen
; livestock feeding
; biochar
; greenhouse gas
; sustainability
; agriculture
; game changer
; environment
1. Introduction
Climate change is one of the most life-threatening challenges humanity has ever faced, which puts at risk our common future [1]. The impact of climate change has been recorded on five dimensions known as the 5Ps: the people, planet, partnership, prosperity, and peace [2,3]. The major effects of climate change are persistent global warming and episodes of abiotic and biotic stresses that exacerbate the economic crisis [4], aggravate inequalities and social vulnerability [5,6], and increase food insecurity [7,8]. Empirical data reveals that the recorded gradual and persistent emission of greenhouse gases (GHGs: carbon dioxide (CO2), methane (CH4), nitrous oxide (N2O), etc.) into the atmosphere are the major cause of the observed global warming events and climate crisis. GHGs are produced biologically or naturally (through the action of specific microorganisms in soil or via chemical reactions) and by the action of humans (anthropogenic source: energy sector (73.2%), industry (5.2%), waste (3.2%)), and the remaining is attributed to agriculture (crop production, agricultural soils, livestock), land use and forests.
Agriculture, land use, and forests (herein referred to as ALF) are sinks and sources of GHGs. Sinks of GHGs are reservoirs of carbon removed from the atmosphere through biological carbon sequestration [9,10]. As sources, ALF account for about 18.4% of global GHG emissions. In the same way, agricultural soils and cultivation practices, especially due to the excessive applications of nitrogen (N)-rich fertilizers and livestock, are identified as leading sources of atmospheric GHG emissions that possess a high global warming potential (GWP) and the potential to exacerbate climate change effects [11]. Nevertheless, agriculture remains the economic sector that suffers the most from climate change. Of the well-identified GHGs emitted from agriculture (crop production and management, agricultural soils, agricultural practices and livestock), CH4 ranks number one and accounts for nearly 67%, followed by N2O and CO2 with 32% and 1%, respectively.
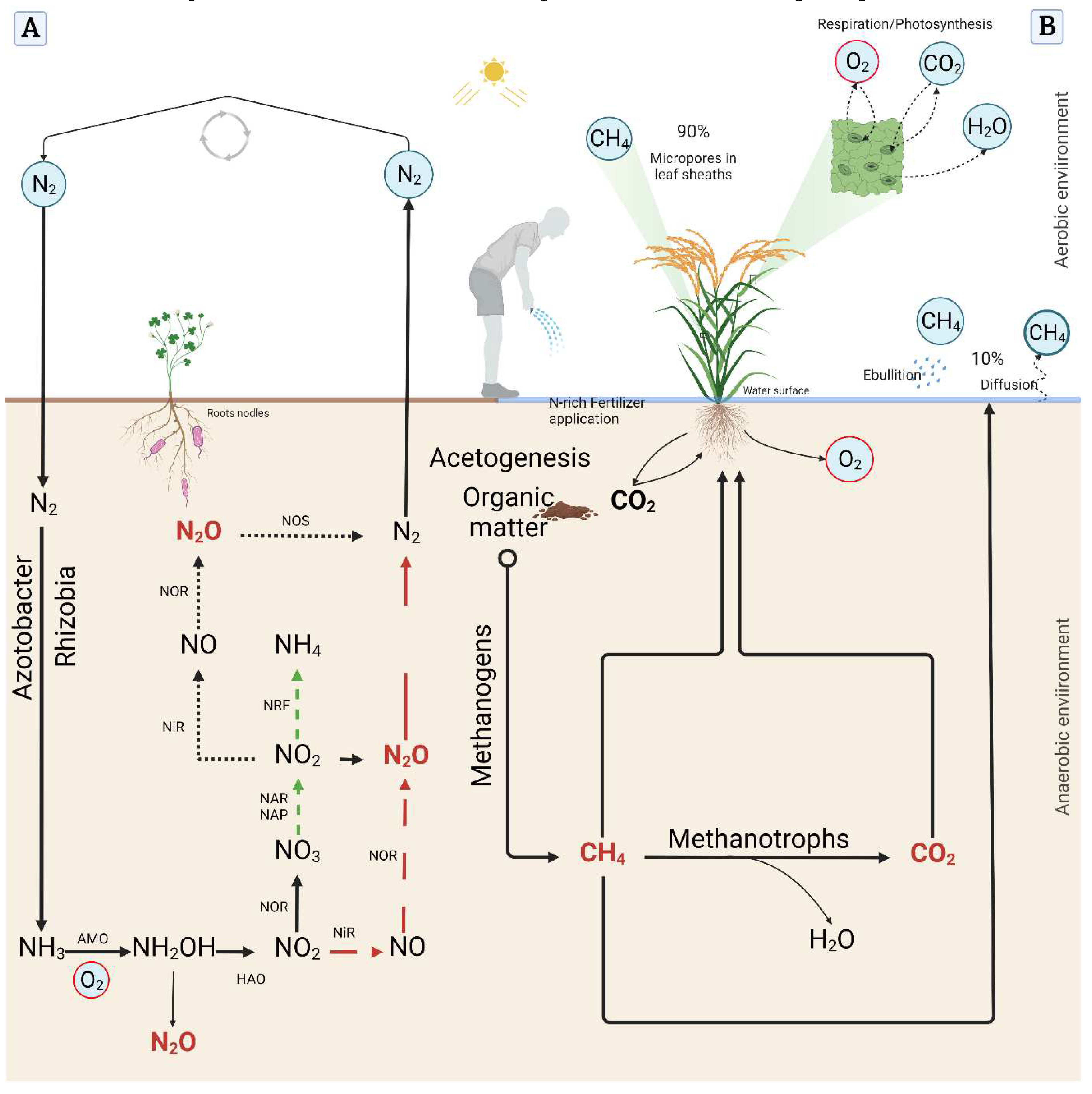
Despite the observed historical preference for CH4 to coal and oil, CH4 has emerged as a leading GHG right behind CO2 and before N2O, with a GWP 28–34 times higher than that of CO 2 in a 30 year-period [12]. An estimated indicates that approximately 90% of CH4 emitted during crop cultivation, especially in flooded-prone cultivation areas such as paddy fields (including rice production), are conveyed by crops through plant’s gas exchange, majorly through micropores of leaf sheaths, not via stomata. Whereas, nearly 10% would be released to the atmosphere through ebullition or diffusion from the water surface or soil [13,14]. However, several diverging debates question on the actual role of plants in the current global GHG budget, especially CH4 [15].
Some research groups argue that plants do not make CH4, while supporting that plants do not contain a known biochemical pathway for CH4 synthesis [16]. In contrast, plants only play a role of conveying gases produced by the action of specific soil microorganisms in the rhizosphere of roots and its immediate environment that move throughout the plant vessels and later released to the atmosphere [17-19]. Others think that this question remains open, while pointing to a strong relationship between CH4 production and the ultraviolet radiation, and supporting that CH4 would come from parts of plants that do not give off water [20-22]. A third voice nourishes the idea that, based on some lines of evidence, the production of CH4 by plants would be part of a survival strategy during stress conditions [23,24]. According to the UNEP, reducing global GHG emissions from agriculture caused by linear food production systems, as part of the net zero carbon emissions targets, may help slow down the current global warming rate in the short term, reduce peak warming during this century, and bridge the GHGs emissions gap between current emission pathways or trajectories and those consistent with the 1.5 °C temperature goals [25].
Furthermore, agricultural practices such as stubble burning or crop residues burning is widely considered as another important source of atmospheric GHG emissions from agriculture. This farming practice was widely used by many farmers across the world to quickly get rid of crop residues after harvest is complete without employing an important work force. However, several reports have warned against serious damages of crop residues burning on the air quality and human health, in addition to GHGs carried (CO2, CH4, N2O, etc.) in the smoke. Currently, there is strong consensus that farmers must abandon the burning of residues to adopt environmental friendly residues management practices.
Although studies question the net impact of N-rich fertilization regimes on the emission of CH4 and N2O from agriculture, a strong consensus exists on the fact that different rates of N applications differentially influence the activity of soil microorganisms, including methanogens and methanotrophs [26,27]. Like nitrogen, even if they do not belong to the class of mineral macronutrients, carbon, hydrogen, and oxygen are considered essential to the life of plants as they are required in large quantities to build the large organic molecules of the cell [28,29]. The O2 level in the soil is essential for both plants and soil microorganisms [30]. In the flooded-prone or wetlands cultivation areas, the diffusion of O2 is much lower compared to that of the upland, which conditions favor, in most cases, the formation of GHGs, including CH4 [31]. During gas exchange, plants release O2 through aerenchyma to the rhizosphere of the root system and the immediate environment. This process called radial oxygen loss (ROL) helps reduce the accumulation of phytotoxins (sulfides and ferrous iron) but also helps in the reduction of CH 4 generation in the soil. ROL is regarded as a promising trait, which helps supply O2 to hypoxic soil environments for efficient plant root growth and development [14]. In the same way, O2 level in the rhizosphere is proposed to influence the activity of methanogens and methanotrophs [31], while O2 deficiency, especially in waterlogging conditions, has long been suggested to cause injury to plants [32]. A recent study conducted by Duyen, et al. [33], aiming at exploring the genetic basis for ROL suggested a set of genes with the potential to regulate ROL in plants.
Like certain crop management practices, livestock production and management are an important source of atmospheric GHGs. Livestock (ruminants: cattle and sheep, etc.) contributes significantly to GHG emissions, especially CH4, throughout their life (feeding process: digestion, burps, flatulence). At the latest COP26 and COP27 in Glasgow in 2021 and in Sharm el-Shekh in November 2022, global leaders expressed their concern over the impact of livestock-mediated CH4 emissions to the atmosphere and their contribution to the global GHG emissions record and climate crisis (https://www.zurich.com/en/media/magazine/2022/is-there-more-to-methane-than-cow-farts, accessed on February 1, 2023). Because of its high GWP 30 times than CO2 in a relatively short period, CH4 is considered one of the most potent GHGs. Given the GWP of CH4 and its contribution to global warming events, countries have signed a Global Methane Pledge (https://ec.europa.eu/commission/presscorner/detail/en/statement_21_5766, accessed on February 1, 2023) to Keep 1.5 °C within reach.
This review seeks to increase global awareness of the necessity to rethink agriculture and food production systems while highlighting key approaches that can effectively play the role of game-changers or carry the potential needed to support global efforts to reduce GHG emissions from agriculture, which offer multiple beneficial outcomes for plants, the environment, and the people.
2. Major sources of greenhouse gases in agriculture
2.1. Agricultural Land Use and Management of Crop Residues
Crop production involves several land preparatory activities prior to cultivation. Economists have identified factors of production resources or inputs commonly used in the production process to produce goods and services. The production function is largely determined by the utilized amounts of the various inputs. Reports indicate that there are four basic resources or factors of production, which are land, labor, capital and enterprise. Labor, capital, and land are considered primary factors of production. Land includes not only the site of production but natural resources as well.
In agriculture (crop production, fisheries and livestock), these factors of production are essential resources and components of the production system to achieve a desired productivity and profitability. Practicing agriculture involves opening new cultivation lands or utilizing already existing and exploited cultivation fields. In many parts of the world, farmers set fire to agricultural lands to clear stubble, weeds and wastes, after harvest is complete and before sowing a new crop or when opening newly acquired agricultural land that is to be cultivated for the first time. On the one hand, voices claim that this practice appears to be the easiest and most economical way to open a new cultivation land or get rid of wastes and crop residues [34]. On the other hand, another trend of thought highlights that the environmental and human cost of agricultural open burning of crop residues far overweigh the near-term economic benefits for farmers. Burning of crop residues after harvest is used by many farmers as an alibi to quickly prepare the land for other crops in a crop rotation scheme for instance or is considered as a low-cost straw-disposal practice to reduce the turnaround time between harvesting and sowing for the next season. However, this practice is more and more castigated or event prohibited because of its negative impact on the environment (by degrading air quality in producing black carbon and reducing the fertility of soil) [35] and one of the largest causes of air pollution-associated illnesses and deaths after cookstoves (https://www.ccacoalition.org/en/activity/open-agricultural-burning, accessed on January 13, 2023). In turn, black carbon would be responsible for about a third of all black carbon emissions globally, and constitute a short-lived climate pollutant that causes air pollution, climate change, and increase melting in the cryosphere (regions of snow and ice). In the same perspective, Singh, et al. [36] highlights that burning of crop residues is regarded as a serious threat to the environment and human health and wellbeing, induces loss of nutrients (nitrogen, phosphorus, potassium, sulfur, organic carbon, etc.) essential for soil fertility, plant growth and productivity. Crop burning is also counted among the major causes of air pollution in several parts of the world, therefore contributing to enhance mortality rates and slumping agricultural productivity.
Reports suggest that burning of crop residues produces heat that elevates soil temperature, which causes death of beneficial soil microorganisms. Successive burning of crop residues on an agricultural field may lead to a complete loss of the microbial population and reduces the level of nutrients such as nitrogen, destroy the organic matter and carbon in the top soil layer (0–15 cm) profile that makes soil fertile, causing yields to decrease over time and increasing the need for costly fertilizers. This soil layer is important for crop root development. Consequently, there could be a slow and steady reduction in soil health, which will eventually result in decreased productivity. Figure 1 displays a map of crop stubble burning globally.
On the one hand, Andini, et al. [37] assessed the impact of open burning of crop residues on air pollution and climate change. The authors observed that on average, 90% CO2 and 8% CO were the most abundantly emitted GHGs over a period of one year, while nearly 2% included CH4, SO2, NOx, NH3, N2O, and others. At the global scale, the assessment of climate charging emissions suggests that crop residue burning to contribute to 12–14% toward global warming potential. On the other hand, Romasanta, et al. [38] employed various rice straw management practices (SRt-straw retained including stubbles and incorporated, PSRm-partial straw removal only stubbles incorporated, CSRm-complete straw removal including removal of stubbles, and SB-straw burned followed by incorporation) to study their effects on GHG emissions and GWP contributions. The authors argued that despite the fact that open burning of straw residues emits, through smoke, high amounts of CO2, the latter is not considered as net GHG emissions. Rather, CO2 from smoke concludes the annual carbon cycle that started with photosynthesis. This could justify the observed focus in favoring investigations of burning residues-mediated CH4 and N2O emissions over other GHGs. In addition, the above study found that SRt recorded the highest GWP value per unit area. In contrast, CRSm yielded the lowest GWP, which eventually depends on the ensuing utilization of straw and the off-field emissions involved.
2.2. Farming Practices and Fertilizers
Agriculture (farming practices/crop production and livestock production) is the second biggest GHG emitting sector after the Energy sector. An increasing trend in developing innovative agricultural practices and tools for more food production and animal feeding has long been at the core center of several agricultural-related research globally. Until before the last two decades, less attention was given to the environmental aspects of these farming practices, until the last few decades. The damages caused to the environment, health, and people's lives drew the attention of many when the situation became alarming and when consequences became evident and life-threatening (https://agriculture.vic.gov.au/climate-and-weather/understanding-carbon-and-emissions/greenhouse-gas-cycles-in-agriculture).
Figure 2A,B (adapted from Baggs and Philippot [39]) and S1 illustrate the share of agriculture in the global GHG budget in agriculture, including crops production and livestock and manure, land use, and forestry are responsible for about 18.4% of global GHGs emissions. Based on their release amounts, CO2, CH4, and N2O are considered as the major GHGs contributing the most to the global warming. However, in agriculture, CH4 is the most abundant, followed by N2O, and CO2 (Figure 2B). With regard to their respective GWP, N2O and CH4 have higher GWP compared to CO2. Concerning crop production systems, synthetic or mineral N-rich application regimes are reported as major factors enhancing GHGs emissions from agriculture [40]. Figure 3A,B and Figure S2 provides insights into the generation and emissions of GHGs from agriculture (crop production), such as N2O, CH4, and CO2.
A review on factors controlling N2O and CH4 emissions from soil indicated that nitrogen fertilizers, biological N fixation by associative free-living and mutualistic bacteria, organic N and the excreta of grazing animals are sources of N, which can lead to CH4 and N2O emissions from soil [41,42]. In essence, factors having a great potential to influence agriculture N2O and CH4 emissions are fertilizer type, N application rate, crop type, soil organic carbon content, soil pH and texture. On the one hand, production of GHG in agriculture is expected to increase with the increase in food demands (including animal products) following the relatively high population growth, the intensification of agricultural activities and the expansion of agricultural land coverage globally. On the other hand, monitoring and quantification of GHGs from agriculture under open field conditions has been a challenging task. Nevertheless, reducing the emission of GHGs is gaining momentum and is at the center of global initiatives aiming at establishing effective strategies to lower the contribution of agriculture to the global warming.
2.3. Livestock production
The share of livestock alone in the agricultural GHG emission record surpasses that of crop production, agricultural soils, crop burning taken individually [43]. An estimate from various sources (foodandagricultureorganization.shinyapps.io/GLEAMV3_Public/, accessed on June 13, 2023) suggest that livestock would be responsible for nearly 11.1‒19.6% of global GHG emissions. Other sources indicate that agriculture produces and emits nearly 18.4% of total GHG emissions recorded globally, including 6.2% is attributed to livestock and manure (Figure S1, https://www.climatewatchdata.org, accessed on February 24, 2022). A lot of critics point out animals, especially ruminants, for the part they play in climate change, and most often cattle and sheep are mostly indexed. Nevertheless, reducing GHG emissions from livestock remains the most important target.
3. Mechanisms of greenhouse gases production and emission from crop cultivation
3.1. Methanogenesis and Methanotrophy
Methane (CH4) was first identified by the Italian physicist Alessandro Volta in the late 18th century [11]. Methanogenesis is an anaerobic respiration that generates CH4 as the final product of this metabolism, through the exclusive action of methanogens, from substrates such as H2/CO2 , acetate, formate, methanol, and methylamines [44]. These bacteria are strictly anaerobic and are common in wetland environments. Several reports indicate that plant-mediated transport is the primary mechanism for CH4 emission from agriculture, especially in flooded-prone crop cultivation areas such as rice paddy fields, which accounts for about 90% of CH4 emitted to the atmosphere [14]. As the diffusivity of O2 of gases is much slower in water than in air, direct exchange of gases between submerged tissues and the environment is impeded in flooded fields. Under these conditions, the aerenchyma play a dual role; one is the supply of O2 to the roots and rhizosphere, while the other is the transport of gases such as CO2, ethylene, and CH4 from the soil to the shoots and release to the atmosphere [14]. Under anaerobic conditions, organic matter such as glucose is oxidized to CO2, while the molecular oxygen (O2) is reduced to water.
Three major pathways for CH4 generation have been reported, including acetoclastic, methylotrophic, and hydrogenotrophic methanogenesis [45]. Acetoclastic methanogenesis is assumed to be the major pathway through which CH4 is generated (acetoclastic methanogenesis). Acetoclastic Methanosaeta are dominant methanogens in organic-rich Antarctic marine sediments. In addition, two genes required for acetoclastic methanogenesis, ackA and pta, were identified and functionally characterized [46]. During hydrogenotrophic methanogenesis, hydrogen (H2) is oxidized to hydrogen proton or ion (H+), formic acid (CH2O2) or other simple alcohols are also oxidized in the process, and CO2 is reduced to CH4.
Studies have shown that the most effective, if not the only biological option of degrading CH4 is by microbial oxidation [47,48]. Kirschke, et al. [49] indicated that due to their characteristics (tight-aquatic terrestrial coupling and large organic matter accumulation), wetland or flood-prone cultivation environments are hotspots of biogeochemical processing. The authors supported that the high methanogenic activity in their anoxic, carbon-rich soils make wetlands or flood-prone cultivation areas source of global atmospheric CH4, emitting about 142–284 Tg CH4 per year.
Methane is oxidized by methane oxidizing bacteria (MOB) or methanotrophs, in the presence of O2 through methanotrophy. This process is restricted to prokaryotes (Methylosinus, Methylocystis, Methanomonas, Methylomonas, Methanobacter, and Methylococcus). Eukaryotic microorganisms, such as algae and fungi do not oxidize CH4. Methanotrophs require CH4 as their sole source of carbon majorly through a process called “nitrifier denitrification” or from abiotic and biotic transformations of their metabolic intermediates, with the nitrifier denitrification being predominant and more favored in soils containing a high NH4 supply [50,51]. In the process, they unlock the energy of oxygen, nitrate, sulfate, or other oxidized species [52,53]. These bacteria occur mostly in soils, rice paddles, mud, landfills, among other places where CH4 is available. In aerobic environments, MOB use O2 and CH4 to form formaldehyde. Reim, et al. [54] supported that anaerobic MOB found in paddy soils act as a bio-filter in mitigating emissions of CH4 to the atmosphere, as O2 is available in soils the atmospheric CH4 is corroded. Aerobic methanotrophs has the ability to oxidize CH4 via the MMO enzyme [55]. As per some evidence, there is strong linkage between the consumption of CH4 and the composition of MOB communities. According to Walkiewicz, et al. [56], methanotrophy of arable soils may be affected by N fertilization. The authors observed that CH4 oxidation was complete concomitant to the reduction of O2 level in soil without NH4 application; therefore supporting that methanotrophs would be favored under hypoxia in ammonium-fertilized soils.
Stein, Roy and Dunfield [55] supported that methanotrophs and ammonia (NH3) oxidizers or nitrifiers share many similarities, such as having in common key enzymes including ammonia monooxygenase or particulate methane monooxygenase; they occupy similar ecological niches, and compete for nitrogen. Both CH4 and NH3 are highly reduced molecules and are suitable growth substrates for microbes, and they can be oxidized (aerobically or anaerobically) to yield energy. The authors further indicated that ammonia oxidizers are enzymatically capable of oxidizing CH4. In the same way, methanotrophs are capable of nitrification.
3.2. Nitrous Oxide generation and emission
Nitrous oxide (N2O) is the third most important GHG behind CO2 and CH4. Although it is less abundant and present at a concentration of about 320 parts per billion (ppb) in the atmosphere, N2O is said to have a GWP nearly 300 folds greater than that of CO2 and is regarded as the dominant stratospheric ozone-depleting substance [57]. Thomson, et al. [58] argued that N2O emissions are difficult to estimate due to their predominant biogenic origin, and the N2O-production and –consumption pathways occurring simultaneously in different microenvironments in the same soil (Figure 3A). N2O is formed predominantly in soils and oceans [59], and is mediated by microbial processes [39]. The production of N2O is the result of multiple biological pathways, including nitrification using the reactive N compound NH4 [60,61], and denitrification, where NO3 is converted back to N2 in the biological N fixation [40,62-65] in the presence of O2 [66], dissimilatory NO3 reduction to NH4, nitrifier denitrification, and non-biological chemodenitrification. However, this event is particularly dominated by nitrification and denitrification [67,68]. Agricultural practices, climatic conditions, and soil properties have been recognized as the major source of N2O emissions, driven by the soil moisture and temperature [69], aeration NH4, and NO3 concentrations [70], and pH [71]. Other sources of agricultural N2O and other nitrogen oxides (NOx) release into the atmosphere are biomass burning and NH3 from livestock manure accounting for about 65% of global N2O emissions [72], land use and management [73], and leaching of agricultural N fields [74].
Nitrification, a major component of the global nitrogen cycle, is initiated with the oxidation of ammonia (NH3), governed by two specialized groups of ammonia oxidizers named ammonia-oxidizing bacteria (AOB) and ammonia-oxidizing archaea (AOA) [75]. Stein [76] highlighted that ammonia-oxidizers contribute to the generation of N2O, with AOB prevailing over AOA that produces lower yields of N2O than bacteria during aerobic NH4 oxidation in soil [77,78]. Hink, Nicol and Prosser [50] supported that nitrogen fertilization during crops cultivation increases significantly N2O emissions in the presence of O2. For several decades, until recent reports showed evidence of AOA being active in the nitrification process [79-83], it was believed that AOB were the only microbes responsible for the oxidation of NH3 to NO2. Then, the conversion of NO2 to NO3 is regulated by nitrite oxidoreductase [84]. As per to some evidence, a complete oxidation of NH3 to NO3 (complete NH3 oxidation, camammox) is mediated by a single organism [85,86].
Microbial production in soils is the prevalent source of N2O, which is enhanced with the application of N-rich fertilizers [87,88]. Nevertheless, fertilizers alone cannot account for the historical global trends of atmospheric N2O concentrations [89]. Soils account for 56%–70% of total global annual N2O budget, while anthropogenic N2O sources, especially due to N-rich fertilization, is responsible for nearly 40% of global N2O emissions [89-91]. Cost-effective inhibitors, which have the potential to regulate nitrogen processes in soils, are highly recommended. Sufficient knowledge of sources of GHGs production via various microbial processes in the soil is a prerequisite.
Figure 3.
Simplified illustration of the nitrogen biological cycle and greenhouse gases emissions in plants. (A) Nitrogen gas N2 fixation and nitrous oxide (N2O) generation during nitrification and denitrification. Black continuous lines with an arrow represent the nitrification process. Black dotted lines with an arrow indicate the denitrification process. Green medium dash lines show the nitrate ammonification process. Red long dash lines with an arrow denote the nitrifier denitrification. (B) Methane (CH4) generation and emission from nitrogen-rich fertilizers and organic matter decomposition under the action of methanogens, and CH4 oxidation mediated by methanotrophs. Greenhouse gases have a red font. This model was created using biorender design platform (https://app.biorender.com/, accessed on 2023.03.25).
Figure 3.
Simplified illustration of the nitrogen biological cycle and greenhouse gases emissions in plants. (A) Nitrogen gas N2 fixation and nitrous oxide (N2O) generation during nitrification and denitrification. Black continuous lines with an arrow represent the nitrification process. Black dotted lines with an arrow indicate the denitrification process. Green medium dash lines show the nitrate ammonification process. Red long dash lines with an arrow denote the nitrifier denitrification. (B) Methane (CH4) generation and emission from nitrogen-rich fertilizers and organic matter decomposition under the action of methanogens, and CH4 oxidation mediated by methanotrophs. Greenhouse gases have a red font. This model was created using biorender design platform (https://app.biorender.com/, accessed on 2023.03.25).
3.3. Carbon dioxide emission and sequestration
Carbon dioxide (CO2) is the best-know anthropogenic GHG. High throughput scientific data suggest that the increased emissions of CO2 resulting from various sources (majorly from fossil fuel burning and much less from agriculture) have the potential to affect global temperatures, considering the radiative effects of CO2 and water vapor on the surface temperature of the earth. Likewise, based on theoretical scientific predictions, an increase of CO2 by 250–300% over time would cause an increase in temperatures in the Arctic by 8–9% [92]. Agricultural soils and cultivation activities are identified as leading sources of atmospheric GHG emissions, which have a high GWP and the potential to exacerbate climate change effects. Agriculture is both as sink and as source for GHG. Agriculture sinks of GHG are reservoirs of carbon removed from the atmosphere through biological carbon sequestration. Carbon sequestration here refers to the capacity of agricultural lands and forests to capture and long-term storage of atmospheric CO2 in the soil. Data indicate that the curve of CO2 increase over time has a zigzag pattern rather than an increasing as steady rate, which suggests a signal of a sink (the massive uptake of CO2 by terrestrial and oceanic plants in specific seasons). These data provide evidence of the importance of sinks in the control of atmospheric GHGs concentrations, while showing at the same time that this control might be limited and underlying upward trend that the sinks are no longer balancing the sources [92]. In this regard, CO2 sequestration currently presents the best solution to counterbalance the increase of GHGs. This includes enhancing biomass production, application of low-cost plant growth regulators and bio-fertilizers, agricultural conservation practices (no-till, use of biochar, no crop residue burning, etc.). CO2 is removed from the atmosphere by plants and converted to organic carbon through the photosynthesis process. During the decomposition of organic carbon, it is converted back to CO2 through the respiration process.
4. Mechanisms of GHG production and emission from livestock
Livestock-mediated GHG emissions occur at different levels, including (i) enteric fermentation, (ii) manure, (iii) feed production, (iv) land use change, (v) energy, and (vi) processing. During the enteric fermentation (a regular digestive process of ruminants), ruminants such as cattle and sheep produce CH4 within the rumen (fore-stomach) during digestion, through a reaction between carbon and hydrogen. Reports suggest that nearly 90‒95% of CH4 produced during enteric fermentation (a digestive process that convert sugars into simple molecules for absorption into the bloodstream, which in turn produces CH4 as a by-product) and released to the atmosphere, occurs through burps (burping). In essence, cow belching due to enteric fermentation is the largest CH4 source in livestock as illustrated in Figure 4. Another portion of CH4 is produced in the intestine and later expelled. The latter accounts for about 5‒10% of CH4 emissions, which occur during farting. In addition, setting ponds and lagoons for processing manure produce a significant amount of CH4 as well (https://climate.nasa.gov/faq/33/which-is-a-bigger-methane-source-cow-belching-or-cow-flatulence/, accessed on February 1, 2023). In this case, solid waste produces both CH4 and N2O, when manure is tore in liquid systems such manure lagoons. Likewise, feed production is another source of GHG gas emissions. A small amount is associated to manufacturing fertilizers and other farm inputs (CO2), and fertilizing crops (N2O) [93]. GHG are also emitted during feed transportation and processing. Furthermore, expansion of pasture for grazing animals and cropland for growing feed crops (land use change) results in conversion of forest, grassland, and other land, causing emissions of CO2 stored in biomass and soils [94]. Although several studies assumed that GHG emissions balances out by carbon sequestration, Xu, Sharma, Shu, Lin, Ciais, Tubiello, Smith, Campbell and Jain [43] found that, in addition to CH4 and N2O, CO2 is release from livestock production than it is sequestered in vegetation and soil via pasture, rangeland, and feed crops. The authors indicated that soil and livestock respiration, tillage, and manure among others are important sources of CO2. In the same way, energy used to produce farm inputs and feed serves in animal production for ventilation, cooling, and other activities, and constitutes a source of GHGs. Although GHG emissions related to processing (slaughtering of livestock, processing and packing the meat for consumers, which extends beyond the farm gate) is not a major component of raising livestock per se, it is included in most global estimates [95,96].
Unlike the forest, environmental sciences, and the energy sector, measuring and quantifying GHG emissions from crop cultivation in the field remains a challenge, due to the complexity of sampling procedures and data collection, design of effective and convenient chamber size and plant growth, the variation in weather conditions during the day, among other factors. Although some progress has been recorded in recent years, GHG emissions measurement tools and technology is not yet accessible to all. In crop production, scientists and environmentalists commonly use customized PVC chambers various shapes and dimensions (square, cubic, cylindrical, etc.) for field applications used for grass plant species, cereal crop species, or livestock (cow, sheep, etc.) to condition the gas and syringe for gas samples collection, thermometer temperature data acquisition, etc. For the laboratory analysis, GC MS is commonly used.
5. Approaches to Reduce GHG Emissions in Agriculture
Farming practices are not always the same around the world, although there are some common practices and similarities shared among certain regions of the world. The possible reasons explaining in part this situation may include the diversity of soils properties and characteristics, rainfall patterns and climates varying from one region to another as well as cultural and social dimensions. In addition, different farming practices or methods may work better in a given environment but perform differently in other places. Furthermore, different areas of the world are better for growing certain types of crops, and some farms are huge, while others are small. Besides, there are also cases where farms are operated by large corporates or companies, middle-scale or small-scale farmers, with modern technologies or secular practices with limited resources [97]. In essence, agriculture is the process of producing food, including grains, fiber, fruits, and vegetables, raising livestock producing feed for animals, among others. Since the invention of agriculture (about 10,000 BCE), humans took control of their environment to produce their own food. As of today, modern agriculture, characterized by a linear production system, is a subject of controversies because of its contribution to global GHG emissions. Given the importance of the subject, and considering the necessity to address the agricultural-associated GHG emissions (about 18.4% of global GHG emissions), we could think of several potential solutions that may be regarded as game changers. Scientists and agricultural practitioners have come up with suggestions that may have a greater impact on lowering GHG emission records over time. In the below paragraphs, we attempted to collect and propose diverse approaches, not to be considered in the order of importance, which may exert transversal effects on GHG emissions and economical aspects of food production, while targeting agricultural practices, crop management habits and fertilizer application regimes or plant nutrition schemes, plant breeding methods and technology, livestock management and feeding, etc. Below are some of the areas identified with the potential to contribute to the reduction of GHG emissions from agriculture. A summary of most promising approaches is provided in Table 1.
5.1. Enhancing Nitrogen Use Efficiency in Plants
Nitrogen use efficiency (NUE), also referred to as N uptake, transport, translocation, assimilation, and remobilization, is regarded as a way of understanding the relationships between the total nitrogen inputs compared to the nitrogen output (Figure 5). Breeding for enhanced NUE in plants is essential but a challenging task regarding the complexity surrounding N acquisition and assimilation by plants. Improving NUE would imply targeting genetic loci controlling various aspects of the NUE using a forward genetic approach, targeting specific genes or transcription factors encoding genes associated with N acquisition, transport, and assimilation events. These could be identified through quantitative trait locus (QTL) analysis and fine mapping of detected QTLs or genome-wide association studies. In addition, the application of reverse genetics that employ molecular techniques to elucidate the function of genes through genetic engineering, coupled with sequencing technologies have gained momentum in the scientific community [98-100]. These techniques offer a wide range of opportunities and open new paths to investigating genetic factors controlling important traits in plants under various environmental conditions. Nevertheless, developing crop varieties with a high NUE is a promising approach to reducing application rates of synthetic fertilizers especially in wetlands cultivation areas.
Although the mechanism of N acquisition, uptake, and assimilation by plants is well described [101,102], the molecular basis of NUE in plants have not been fully elucidated, and continue to be investigated. Studies aiming at investigating mechanisms underlying NUE identified key protein families with a high potential to controlling NUE in plants under various cultivation conditions [103,104], while others suggested methods for assessing and estimating NUE in plant crops [105-107]. NO3 and NH4 are the major forms of N taken up by plants, with NO3 being the most abundant. N is acquired from soil through a combined action of low- and high-affinity NO3 and NH4 transporters. The latter are found within five protein families, including NO3 transporter 1 (NRT1) and 2 (NRT2), chloride channel (CLC), and slow anion channel-associated/slow anion channel-associated homologs (SLAC/SLAH) , while assimilation primarily involves glutamine and glutamate synthase encoding genes but not limited to [103,108,109] . The enzyme glutamate dehydrogenase (GDH), which protects the mitochondrial functions during episodes of high N metabolism takes part in N remobilization [110].
Application of synthetic N-rich fertilizers during crops cultivation dramatically increased the last decades. This common agricultural practice has been shown to contribute to GHG emissions. In this regard, several strategies to reducing the emissions of GHGs from agriculture have been proposed. Number of methods employed to assess the NUE in different crop species are reported [111-113]. Of this number, various strategies aiming at improving NUE have been implemented, and their efficiency varies with crop species [114-120]. With the recent advances in plant breeding techniques and the advent of sequencing technologies, a wide range of opportunities are explored to identify high NUE in crop plants in various breeding populations. Screening for chlorates (ClO3) sensitivity may also help identify rice varieties with an enhanced NUE [106].
Furthermore, in higher plants, phytohormones were originally known as a group of naturally occurring organic substances, which positively or negatively regulate plant growth and development. In addition to their basic roles, plant hormones are recognized as key players in coordinating multiple (both local and long-distance) signaling pathways at the whole-plant level [121]. As per some evidence, plant hormones interact with nitrogen (N) as well as other nutrients such as iron, sulphur, and phosphorus [122-126]. Among the well-studied phytohormones, abscisic acid (ABA), auxin, and cytokinin (CK) are closely associated with the N signaling. NO3 availability differentially affects phytohormones accumulation. For instance, NO3 signaling was proposed to interact with AtIPT3 in Arabidopsis and regulate N acquisition events, while inhibiting auxin (AUX signaling and basipetal transport (translocation from shoot to root). Meanwhile, Vidal, et al. [127] suggested that NO3 induces the activity of the auxin receptor gene AFB3, which in turn promotes lateral root, N acquisition and uptake. In contrast, NO3 was observed to repress the transcript accumulation of the auxin response factor ARF8.
Moreover, several studies target key N transporters and assimilation related genes to attempt improving the NUE in plants. Nitrate reductase (NR), nitrite reductase (NiR), plastidic glutamate synthase (GS2), and Fd-GOGAT are involved in the primary NO3 assimilation events. In contrast, the cytosolic glutamate synthase (GS1) and nicotinamide dinucleotide hydrogen (NADH)-GOGAT are involved in the secondary ammonia (NH3) assimilation and remobilization. In this regard, Chen, et al. [128] suggested that genetic manipulation of NO3 remobilization in plants, a key component of the N metabolism, would help improve NUE, while critically reducing N fertilizer demand and alleviating environmental pollution. To date, genetic engineering techniques are used with the purpose of improving NUE in plants crops [129-131]. Figure 6 highlights some of the tools and methods employed to investigate the mechanisms and key players in the N metabolism with the purpose of improving NUE in plants, as well as its beneficial outcomes.
Heuermann, et al. [132] showed that NO3 stimulates Cytokinin (CK) synthesis. However, elevated CK levels may delay plant senescence, while favoring a prolonged N uptake. Likewise, Ruffel, et al. [133] reported a NO3-CK relay and distinct systemic signaling for N supply and demand. Gu, et al. [134] supported that nitrogen and CK signaling play a role in root and shoot communication, which maximizes plant productivity. CK biosynthetic genes include IPTs, play a key role in root development, bud outgrowth and shoot branching, and plant development. CK activity occurs in two stages. During the initial stage, CK is produced in the outer layer of the roots and translocate inward. In the second stage, the inner part of the root pushes outward, and forms the nodule. This stage has been proposed to be controlled by ITP3. A study revealed that a knockout mutant plant lacking the IPT3 gene failed to form nodules in the roots [135], which suggests that IPT3 would play a key role in the formation of nodule and nitrogen fixation. Lin, et al. [136] recently observed that NO3 restricted nodule organogenesis through CK biosynthesis inhibition. Similarly, Sasaki, et al. [137] supported that CK regulates root nodulation in plants. Moreover, growth-promoting microorganisms are widely used in agriculture for their roles in the promotion of plant growth and productivity. In their report, Singh, et al. [138] revealed that Trichoderma spp., known as plant growth promoter and biocontrol fungal agents, can enhance NO3 acquisition events, and were shown to encompass the ability to regulate transcripts level of high-affinity NO3 transporters, in a crosstalk with phytohormones.
5.2. Improving Abiotic Stress Tolerance in Plants
Nitrogen acquisition and uptake can be restricted under abiotic stress conditions, such as drought and salinity. External fluctuations of N supply to plants caused by abiotic stress occurrence have been shown to hinder NO3 acquisition as well as other subsequent events due to water scarcity [139]. As per to some evidence, high- and low-affinity NO3 transporters and glutamate synthase-encoding genes [140], and phytohormones biosynthetic and signaling pathway genes [141,142] (in addition to well-characterized abscisic acid (ABA), jasmonic acid (JA)), would play important roles in the adaptive response mechanism towards abiotic stress tolerance in plants. In addition, Zhong, et al. [143] revealed that over expression of a bZIP (basic leucine zipper) transcription factor encoding gene in Arabidopsis, AtTGA4 conferred drought tolerance through the increase in NO3 transport and assimilation mediated by high- and low-affinity NO3 transporters and NO3 encoding genes. Owing to the above, capitalizing on the recorded progress in terms of understanding plant nutrition and abiotic stress tolerance in plants, exploring the interplay between NUE and abiotic stress tolerance could serve as novel and exciting research direction. The outputs would give more insights that allow breeding for abiotic stress tolerance, such as drought, while addressing NUE using an integrated or system thinking approach.
5.3. Exploring Radial Oxygen Loss and Intermittent Drainage
The importance of oxygen (O2) in the life of plants has been established. O2 plays a fundamental role in plants metabolism. For instance, O2 serves as a terminal electron acceptor of the electron transport, and its concentration plays an important role in regulating cellular respiration [144]. The internal transport of gases is said to be crucial for vascular plants inhibiting aquatic, wetland, or flood-prone environments [145]. O2 is the rate-limiting substrate for the efficient production of energy in aerobic organisms. Therefore, they need to adjust their metabolism to the availability of O2.
Plants have the ability to produce oxygen in the presence of light. However, when the O2 diffusion from the environment cannot satisfy the demand set by metabolic rates, plants can experience low O2 availability [146]. Flooding or waterlogging induces hypoxic conditions in plants, which may lead to a reduced energy production. Under these conditions, the direct exchange of O2 between the submerged tissues and the environment is strongly impeded and other programmed cell death (PCD) [147]. The diffusivity of O2 in water is about 10,000 times slower than in the air. In addition, the transport of O2 and other gases across the plant increases by tissues high porosity [148], which results from the intercellular gas-filled spaces formed as a constitute part of development [149-152], and may be enhanced further by the formation of aerenchyma [153]. The aerenchyma facilitate the flow of O2 in and outside the plant, which provide roots with O 2 under flood-mediated hypoxia [14]. Colmer et al. [14] also indicated that aerenchyma provides a low-resistance internal pathway for gas transport between shoot and root extremities, and by this pathway O2 is supplied to the roots and rhizosphere; whereas, CO2 and CH4 move from the soil to the shoot and atmosphere by the same means. The O2 that is released to the rhizosphere of the root system and the immediate environment through the aerenchyma is known as radial oxygen loss (ROL) [154]. In the same perspective, Mohammed, et al. [155] revealed that rice overexpressing the EPIDERMAL PATTERNING FACTOR 1 (OsEPF1)-mediated reduction of stomatal conductance resulted in an increased formation of root cortical aerenchyma, which would be in part explained by reduced O2 diffusion from shoot to the root where EPF signaling may be involved.
Furthermore, flood-prone and wetland cultivation areas, where anaerobic conditions are prevailing and a relatively high amounts of N-rich fertilizers are often applied, have proven to be major sources of GHG gases emissions during crops cultivation [156,157]. The flood status produces anoxic environments that are conductive to the production and emissions of CH4. According to Bodelier, et al. [158], the only biological way of degrading CH4, the second most important GHG globally but the first in agriculture, is by microbial oxidation. In the same way, Reim, Lüke, Krause, Pratscher and Frenzel [54] studied methane-oxidizing bacteria (MOB) under oxic-anoxic conditions in a flooded paddy soil, and suggested that MOB act as a bio-filter in mitigating CH4 emissions to the atmosphere. Biological emissions of CH 4 from wetlands are major uncertainty in CH4 budgets. The MOB use CH4 as their sole source of carbon and energy, as long as oxygen is available [159], contrasting with the methanogenesis by Archaea that is known as an anaerobic process accounting for most biological CH4 production in nature. .
According to Dalal, Allen, Livesley, Richards and Soil [42], aerobic well-drained soils are generally a sink for CH4, due to the high CH4 diffusion rate into such soils and subsequent oxidation by methanotrophs. The capacity of soils to uptake CH4 varies with land use, management practices [160], and soil conditions [161]. In contrast, large CH4 emissions are usually observed in anaerobic conditions, such as wetlands, rice paddy fields, and landfills. Warm temperatures and the presence of soluble carbon provide optimal conditions for CO2 production and incompletely oxidized substrates, thus enhancing the activity of methanogens. Likewise, a close relationship between the increase in atmospheric CO2 levels and the subsequent increase in CH4 emissions has been proposed. In this regard, studies suggested intermittent drainage to reduce the activity of anaerobic methanogens in the soil, especially in flooded crop cultivation systems, which may have a direct impact on the amount of CH4 produced and released by up to 80%. Although in-season or intermittent drainage can result in significant reduction in CH4 production and emissions, this crop management technique aiming to mitigate CH4 emissions can cause an increased N2O emissions, even if the overall warming potential remains lowered [88,162].
As for Walkiewicz, Brzezińska, Bieganowski and Soils [56], the activity of methanotrophs is favored under hypoxia in ammonium (NH4) fertilized soils. In Figure 7, we illustrate the action of ROL on methanogens and methanotrophs activity, which influences CH4 production through oxidation process to yield water and CO2. Studies revealed that there are factors that may cause the reduction of ROL with the formation of a ROL barrier. Colmer, et al. [163] reporter that low concentrations of organic acids may help trigger a barrier to ROL in roots. Ejiri and Shiono [164] supported that the prevention of ROL would be associated with exodermal suberin along adventitious roots. Abiko, et al. [165] observed the formation of a ROL barrier on lateral roots, in addition to adventitious roots, and reported a major locus controlling the formation of ROL barrier in maize. The authors argued that the enhanced formation of aerenchyma and induction of a ROL barrier would confer waterlogging tolerance, which argument was supported by Ejiri, et al. [166] suggesting that a barrier to ROL helps the root system cope with waterlogging-induced hypoxia. In their study, Peralta Ogorek, et al. [167] reported a novel function of the root barrier to ROL in conferring diffusion resistance to H2 and water vapor. In rice, the first genetic locus associated with ROL was recently identified, with a set of genes suggested to be involved in aerenchyma-mediated ROL in plants [33]. Therefore, with the growing concern about mitigating GHG emissions from agriculture, exacerbated by the application of excessive amounts of N-rich fertilizers, coupled with the hypoxic conditions and low diffusion of O2 in waterlogged or flooded cultivation areas, breeding for high ROL in plants could serve as an alternative to conventional techniques such as intermittent drainage that are rarely employed in wetlands. This could be essential for areas such as paddy fields that require efficient water management and where drainage could not be applicable due to evident circumstances such as limited access to a water source. Moreover, it has been evidenced that respiration and nitrogen assimilation in plants are tightly linked. In this regard, studies exploring the interplay between the above factors supported that mitochondrial-associated metabolism can be used as a mean to enhance NUE in plants [168-170].
Carbon dioxide (CO2) is the most abundantly emitted of all GHGs. However, CO2 has a global warming potential 25 times less than that of CH4 and 300 times less than that of N2O. Global leaders and scientists, among other, stressed at the COP26 that CH4 is a great threat to accelerate global warming over a 30-year period, which makes CH4 much more potent than CO2 and the greater climate change hazard. As indicated earlier, irrigated or flood-prone cultivation, systems are favorable environments for CH4 production, which is by far the most abundantly emitted in agriculture. Rice (staple food for nearly half of the world’s population) production occurs through irrigation/flooded or wet environments or upland/rainfed system. For instance, the use of system of rice intensification (SRI) [171], which focuses on changing the management of plants, soil water, and nutrients to create more productive and sustainable rice cultivation, while tending to reduce environmental impacts, could serve as a relevant alternative to reducing GHG emissions. Some of the fundamental concepts of SRI include the use of a smaller amount of seeds and greater planting distances, less use of inputs and intermittent irrigation instead of flood irrigation (savings in irrigation water and inputs) and reduced environmental footprint of rice farming. Regardless of the benefits of SRI, it is overly labor-intensive, requires a higher level of technical knowledge and skill than conventional methods or rice cultivation [172].
Pereira-Mora, et al. [173] investigated the response of plants to organic acids, found that organic acids the abundance of methanogenic arechea and the mcrA gene in plants was reduced in treatment with organic acid under the SRI-rotational cultivation system.
5.4. Biochar reduces mineral fertilizers use, improves soil properties and mitigate GHG emissions
Biochar is widely used as a soil amendment in different agricultural ecosystems. The application of biochar in agriculture increased over the years for various purposes [174], and their recognition as an effective tool for reducing soil GHG emissions has been reinforced in recent years [175-179]. Joseph, et al. [180] define biochar as the carbon-rich product obtained when biomass, such as wood manure or leaves, is heated in a closed container with little or no available air. In other words, biochar is produced by thermal decomposition of organic material under limited O2 supply, and at relatively low temperatures. Unlike charcoal, biochar is mainly produced to improve soil properties, carbon storage or filtration of percolating soil water. Reports indicate that biochar is not only more stable than any other amendment to soil [181], but it helps increase the availability of nutrients beyond a fertilizer effect [174]. Biochar also contributes to (the): (i) improvement of water-holding capacity and other physical properties [182,183], (ii) increase in the stable pool of carbon [184], absorption/complexation of soil organic matter and toxic compounds [185], (iii) absorption and reaction with gases within the soil [186], affect carbon and nitrogen transformation and retention processes in soil [174,187], and (iv) promotion of the growth of beneficial soil microorganisms.
A number of studies proposed that incorporating biochar within soil reduces N2O emissions and impacts on CH4 uptake from soil [188-190]. However, the mechanisms through which biochar influences CH4 and N2O fluxes are not yet well elucidated. Studies suggest that the properties of biochar and its effects within agricultural ecosystems largely depend on feedstock and pyrolysis conditions. As biochar ages, it is incorporated into soil aggregates, and promotes the stabilization of rhizodeposits and microbial products [190]. In addition, Joseph, et al. [191] indicated that the properties of biochar can vary with their element compositions, ash content, and composition, density, water absorbance, pore size, toxicity, ion absorption and release, recalcitrance to microbial or abiotic decay, surface chemical properties (i.e. pH), or surface area. Biochar can catalyze abiotic and biotic reactions in the rhizosphere, which may increase nutrient availability and uptake by plants, reduced phytotoxins, stimulate plant development, and increase resilience to disease and environmental stimuli [190]. Recent evidence suggest that biochar generally increase soil CO2 emission, reduce N2O emissions and NO3 leaching [192,193], and have varying effects on CH4 emissions [194,195]. Kalu, et al. [196] reported an increased CO2 efflux after applying biochar 2‒8 years before planting but did not observe any significant effect on the fluxes of N2O or CH4 in soil with a high soil organic carbon (SOC). A tendency of biochar to reduce N2O fluxes was observed in soils with high silt content and lower soil carbon. The authors recorded as well an increased NUE in the long term, while soils with a high SOC underwent continuous freeze-thaw cycles, which may lead to differential effects of biochar. Thus, biochar is emerging as a sustainable source of plant nutrients for crops and soil quality, with interesting environmental benefits.
5.5. Enhancing Sink Strength
A growing interest in investigating the starch metabolism in plants to explore the possibility to reducing GHG emissions from agriculture, especially CH4 has been observed [197,198]. A study by Su, et al. [199] suggested that increasing sink strength would help enhance the sugar metabolism, while reducing the substrate required for methanogenesis, therefore lowering the activity of methanogens, and consequently affecting CH4 generation in the soil. However, a pending question on how the methanotrophs population would be affected in their role of contributing to the nitrification and denitrification processes [162,200], while relying on CH4 as their sole carbon source for their metabolism remains unanswered.
Root exudation is an important process determining plant interactions with the soil environment [201]. On the one hand, the exudates (low molecular weight compounds: amino acids, organic acids, sugars, phenolics, and other secondary metabolites [202]; high molecular weight compounds: mucilage (polysaccharides) and proteins [203,204]) continuously secreted to the rhizosphere by the roots of plants, are involved in several processes [205]. Plants can modify soil properties to adapt and ensure their survival under adverse conditions, by modulating the composition of the root exudates [206]. Plant root exudates are important factors that structure the bacterial community and their interactions in the rhizosphere [204], or promoting the interactions between plants and soil microorganisms [207], and enhancing resources use efficiency in the rhizosphere [208]. In addition, root exudates are involved in the inhibition of harmful microorganisms [209] or stimulating beneficial micro-organisms [201], keeping the soil moist and wet, mobilizing nutrients, stabilizing soil aggregates around the roots, changing the chemical properties of the soil, inhibiting the growth of competitor of plants [203,210], etc. It is well established that root exudates provide nutrients that favor enhanced growth and a higher prevalence of degrading strains of bacteria [211].
On the other hand, Lu, et al. [212] suggested that stronger roots could secrete more carbon-containing root exudates into the rhizosphere for methanogenesis. The authors found that soils amended with acetate or glucose, root exudates, and straw caused an increased CH4 production. Likewise, Moscôso, et al. [213] recorded an increased CH4 emission induced by short-chain organic acids in lowland soil. In the same way, Aulakh, et al. [214] assessed the impact of root exudates on CH4 production revealed that CH4 production commenced soon after treatment, and the emission increased over time.
For grain crops, yield is the cumulative result of both source and sink strength for photoassimilates and nutrients during seed development. Source strength is determined by the net photosynthetic rate and the rate of photoassimilates remobilization from sources tissues [215]. The long distance transport (sugar export from leaves) and the corresponding demand by sinks has been examined as a possible target for improving plant productivity. The transfer of materials from source to sink is governed by a highly regulated signaling network elicited by resource availability. Sink strength is regarded as the function of size and sink activity, which is tightly related to the source availability. It is accepted that carbon allocation to various sinks is controlled by both sink demand (activity and size) and source control of photosynthate production [216].
Furthermore, Studies indicated that carbohydrates signaling gives insights into the understanding of changes in resources such as N. Increased N uptake and inorganic N availability in leaf tissue favors the synthesis amino acids over gluconeogenesis. As a result, carbohydrates are retained in source tissue at the expense of allocation to heterotrophic tissues such as roots [216,217]. Similarly, a decreased leaf inorganic N leads to decreased amino acids synthesis but increases carbohydrate availability for transport to heterotrophic tissues, including roots. With the increase in carbon availability, genes involved in storage and use are induced [218], leading to root growth and increased N acquisition, more exudates secretion and GHG production.
5.6. Use of Nitrification Inhibitors or Low GHG-Emitting Crop Cultivars
It is widely accepted that excessive application of N-rich fertilizers (mineral N source or organic matter) [219] significantly exacerbate CH4 and N2O production and emissions, especially during nitrification and denitrification processes (the microbial reduction of NO3 to intermediate gases nitric oxide (NO) and N2O and finally to N2). Although N is an indispensable macronutrient for plant growth and development, productivity, quality of products, as well as plant defense, and knowing that doing agriculture without N is nearly utopic; however reducing N application, while optimizing its use, remains one of the major target and one the best options with multiple benefits for the environment and production costs. Organic matter are commonly applied to satisfy soil fertility and improve water retention capacity. The application of green manure, crop residues, manure and composted products contribute to reducing CH4 emissions as discussed earlier.
The application of straw often reduces N2O emissions [88]. Generally, straw with a high carbon/nitrogen (C/N) ratio likely immobilizes available N, thus reducing its availability for both nitrification and denitrification [220,221]. However, the reducing effect of straw on N2O emissions varies from one crop species to another [221], and long-term application of high C/N straw may result in increased N availability which, in turn, may increase N2O emission [222]. Additionally, farmers can take advantage of the nitrification inhibitors, which have been widely shown to reduce N2O emissions in a wide range of crop species [223-225]. Evidence showed that GHG emissions from crops production is also crop variety-dependent.
5.7. Improving Management of Crop Residues
Burning of crop residues is a global issue rooted in many farming systems, although in many countries, several initiatives are being implemented and measures are being taken to curb the use of this linear type of agricultural practice. Burning of crop residues remains harmful to the environment, human being and negatively affect agriculture and food production. To tackle this issue, a global attention is required. To reduce significantly the practice of stubble burning, governing authorities and scientists in several countries are encouraging or introducing effective crop residue management practices as alternative solutions to crop residue burning. Scientists and governments have suggested a number of techniques of crop residue management to efficiently transition to a more friendly agriculture. These include the use of technological interventions to: (i) incorporate crop residues into soils through adoption of conservation agriculture practices (although straw incorporation and organic matter amendments can increase CH4 and N2O production [88,162]) and to prevent soil erosion from wind and water, and augment the soil moisture; (ii) promote the use of crop residue for preparation of bio-enriched compost or vermi-compost and its utilization as farm yard manure; (iii) use of agri-machineries such as Happy seeder (used for sowing of crop in standing stubble), rotavator (used for land preparation and incorporation of crop stubble in the soil), zero till seed drill (used for land preparation directly sowing of seeds in the previous crop stubble), baler (used for collection of straw and making of bales for cereal crops stubble), paddy straw chopper (cutting of crop stubble for easily mixing with the soil), or reaper binder (used for harvesting paddy stubble and making into bundles), zero-seed-cum fertilizer drill, to facilitate in-situ management of crop residue and retaining the straw as surface mulching; (iv) use crop residue for mushroom cultivation.
There is a strong consensus that practicing conservation agriculture (minimize soil disturbance by not tilling, maintaining soil cover, and diversifying crop species) can be an effective, sustainable and productive method of agriculture that can play an important role in containing and curbing the practice of crop residues burning, which is regarded as this environmentally unjustifiable practice. Other alternative solutions considered include the (i) diversification of crop residue as fuel (for power plants, production of cellulosic ethanol, etc.); (ii) use of crop residue in paper, board, panel and packing material industry; (iii) collection of crop residue for feed, brick making, etc. (https://www.downtoearth.org.in/blog/agriculture/stubble-burning-a-problem-for-the-environment-agriculture-and-humans-64912, accessed on January 12, 2023). Agricultural residues equally offer a valuable resource worth saving, since crop stubble can be used as an energy source when converted into pellets, and straw is useful in livestock feed or bedding (https://www.ccacoalition.org/en/activity/open-agricultural-burning, accessed on January 13, 2023).
5.8. Improving Livestock Production and Feeding Efficiency
The global demand for meat and dairy products is growing, and over the past 50 years, meat production has significantly increased in the recent years and is projected to increase by two to three folds by 2050 [226], reaching about 340 million tons each year. The contribution of livestock to the recorded global CH4 emissions is high (https://ourworldindata.org/meat-production, accessed on April 26, 2023). Meat and dairy products are important sources of proteins and vitamins and essential minerals useful to human health in many countries [227-229] but also present potential risks for health [230-232]. Likewise, the production of meat and dairy products has environmental impacts, as it contributes to GHG emissions such as CH4, among others. Today, one of the most pressing global challenges is the sustainable production and consumption of meat, dairy, and other protein products.
The major source of GHG emissions from agricultural production is enteric fermentation of ruminant livestock, and the interest to reducing CH4 production in ruminants continues to grow globally [233]. According to the UNEP Emissions gap report 2022 [234], beyond the necessity to change diets, the reduction of CH4 emissions from ruminants can be achieved via changes in feed level and feed composition, which can also increase animal productivity. Frank, et al. [235] found that the adoption of technical and structural mitigation options could help agriculture achieve at a carbon price of 25$/tCO2 non-CO2 reductions of around 1GtCO2eq by 2030. In the same way, Arndt, et al. [236] indicated that to meet the 1.5 °C target, CH4 from ruminants must be reduced by 11‒30% by 2030 or 24‒47% by 2050 as compared to the record in 2010. The authors identified strategies to decrease product-based (PB, CH4 per unit meat or milk) and absolute (ABS) enteric CH4 emissions, while maintaining or increasing animal productivity (AP, weight gain or milk yield). Other independent studies [237] claimed that enhancing the activity of the major ruminal sulfate-reduction bacteria (SRB: Desulfovibrio, Desulfohalobium, Sulfobus) through dietary sulfate addition, can be used as an effective approach to mitigate CH4 emissions in ruminants, which may lead to a decreased ruminal CH4 production. The major target would be helium (H2), which is the primary substrate for CH4 production during ruminal methanogenesis. In the rumen, SRB have the ability to compete with methanogens for H2, thus resulting in the inhibition of methanogenesis.
From another perspective, research indicates that CH4 emission is also associated with dietary energy loss that reduces feed efficiency [238]. Another way of mitigating ruminal CH4 identified in the literature is the use of saponins. According to Newbold, et al. [239], low concentrations of saponins act as antiprotozoal. In contrast, at higher concentrations, saponins are able to suppress methanogens [240] and inhibit ruminal bacterial and fungal species [241], limiting the H2 availability for methanogenesis in the rumen, thereby lowering CH4 production by up to 50% [240,242]. Other methods for ruminal CH4 mitigation include forage quality [243-245], type of silage [246-249], proportion of concentrates [250-252] and composition [253-256], the use of organic acids [257-259], essential oils (secondary metabolites) [260-262], or probiotics [263-266]. Additionally, exogenous enzymes, such as cellulase and hemicellulose, are used in ruminant diets. These enzymes can improve the digestibility of fiber as well as animal productivity. They are also capable of lowering the acetate: propionate ratio in the rumen, ultimately resulting in the reduction of CH4 production [267,268].
An indirect approach to reduce CH4 production could be the use of antibiotics such as the antimicrobial monensin. The latter enhances the acetate: propionate ratio in the rumen [246] when added to the diet as a premix and has methanogenic effect. According to Hook, et al. [269], ionophores do not alter the diversity of methanogens but change the bacterial population from Gram-positive to Gram-negative, therefore resulting in the change in the fermentation from acetate to propionate, and reducing CH4 [270-272]. Researcher are thinking of employing breeding to explore the possibility for developing low CH4/GHG-emitting cows/ruminants. Numerous studies have shown a substantial variation in CH4 production from cows and sheep [238,273,274], which is associated with phenotypic traits and heritability. Thus, this variation suggests a possibility of breeding animals with low CH4 emission. However, a different view from Eckard, et al. [275] suggested that breeding for reduced CH4 production is unlikely to be compatible with other breeding objectives.
5. Conclusions and Future Prospects
Developing high-yielding and quality crop varieties have been for decades the target of many agricultural-related research programs globally. Although significant progress has been recorded in this regard, the current global warming pattern exposed the linearity and imbalances of current food production systems that are responsible for exacerbating the impact of climate change by contributing to enhancing GHGs emissions. There is a global hunt for effective strategies that allow to lower GHG emissions while maintaining a balanced food production level to meet the increasing food demands. This review presents key approaches previously employed to improve plant nutrient use, fitness, and soil fertility, and here proposed as strategic because they carry a great potential to mitigate GHG emissions from agriculture. Among them, improving nitrogen use efficiency using forward or reverse genetics, and genetic engineering approaches offer the best perspectives for sustainability due to multidimensional impact on fertilizers application, and may confer abiotic stress tolerance, among others. In addition, enhancing radial oxygen loss in plants influences the activity of soil microorganisms, especially methane producing bacteria. Likewise, despite being a major component of agriculture, livestock production produces nearly 30% of GHG emitted from agriculture globally. This challenge offers a great opportunity to improve the livestock production chain and feed use efficiency (dairy farming, beef cattle, etc.), while attempting to lower GHG production and emissions, building on advances in breeding and genetic engineering and feed use efficiency and best nutrients use practices.
These areas of great interest carry the potential to serve as leading approaches with greater impact on the current global efforts to mitigate the effects of climate change and climate crisis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Authors’ contributions: Conceptualization, methodology, and data curation: N.R.K. and J.-H.L.; Writing-original draft: N.R.K.; Visualization and Writing-review & editing. D.S. and J.W.K.; Investigation/Text mining: N.R.K., Y.K., S.M.L., J.K.C., H.P., and G.D.D.; Software: N.R.K; Validation, Funding acquisition, Project Administration and Supervision: J.H.L. and K.W.O. mobilized resources and supervised the project. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Project No. PJ016030032023 of the Rural Development Administration, Korea.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not Applicable.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Holden, E.; Linnerud, K.; Banister, D.J.G.e.c. Sustainable development: Our common future revisited. 2014, 26, 130-139.
- Sam, S.J.E. The people, planet, prosperity, peace and partnership: why the sustainable development goals should matter to everyone. 2016, 30, 7-12.
- Mpabanga, D.; Sesa, L.J.A.J.o.P.A. ; Management. Imperatives: The Five P’s: People, Planet, Prosperity, Peace and Partnerships and Sustainable Development Goals-The Need to Transform Public Administration and Management. 2020, 17, 44–58. [Google Scholar]
- Papoulis, D.; Kaika, D.; Bampatsou, C.; Zervas, E.J.C. Public perception of climate change in a period of economic crisis. 2015, 3, 715-726.
- Adger, W.N.; Kelly, P.M.J.M.; change, a.s.f.g. Social vulnerability to climate change and the architecture of entitlements. 1999, 4, 253-266.
- Otto, I.M.; Reckien, D.; Reyer, C.P.; Marcus, R.; Le Masson, V.; Jones, L.; Norton, A.; Serdeczny, O.J.R.e.c. Social vulnerability to climate change: A review of concepts and evidence. 2017, 17, 1651-1662.
- Hasegawa, T.; Fujimori, S.; Havlík, P.; Valin, H.; Bodirsky, B.L.; Doelman, J.C.; Fellmann, T.; Kyle, P.; Koopman, J.F.; Lotze-Campen, H.J.N.C.C. Risk of increased food insecurity under stringent global climate change mitigation policy. 2018, 8, 699-703.
- Ericksen, P.J.; Thornton, P.K.; Notenbaert, A.M.O.; Cramer, L.; Jones, P.G.; Herrero, M.T.J.C.r. Mapping hotspots of climate change and food insecurity in the global tropics. 2011.
- Crutzen, P.J.J.N. Methane's sinks and sources. 1991, 350, 380-381.
- Reay, D. Greenhouse gas sinks; CABI: 2007.
- Reay, D.; Smith, P.; Van Amstel, A. Methane sources and the global methane budget. In Methane and climate change; Earthscan London and Washington, DC: 2010; pp. 1-13.
- Ming, T.; Davies, P.; Liu, W.; Caillol, S.J.P.i.E.; Science, C. Removal of non-CO2 greenhouse gases by large-scale atmospheric solar photocatalysis. 2017, 60, 68-96.
- Schütz, H.; Holzapfel-Pschorn, A.; Conrad, R.; Rennenberg, H.; Seiler, W.J.J.o.G.R.A. A 3-year continuous record on the influence of daytime, season, and fertilizer treatment on methane emission rates from an Italian rice paddy. 1989, 94, 16405-16416.
- Colmer, T.J.P. , Cell; Environment. Long-distance transport of gases in plants: a perspective on internal aeration and radial oxygen loss from roots. 2003, 26, 17–36. [Google Scholar]
- Schiermeier, Q. Study fails to catch plants making methane. 2009.
- Nisbet, R.; Fisher, R.; Nimmo, R.; Bendall, D.; PM, C.; Gallego-Sala, A.; Hornibrook, E. L {ó} pez-Juez. E., Lowry, D., Nisbet, P., Shuckburgh, E., Sriskantharajah, S., Howe, CJ, and Nisbet, EG: Emission of methane from plants, Proc. R. Soc 2009, 276, 1347–1354. [Google Scholar]
- Nouchi, I.; Mariko, S.; Aoki, K. Mechanism of methane transport from the rhizosphere to the atmosphere through rice plants. Plant Physiology 1990, 94, 59–66. [Google Scholar] [CrossRef]
- Wassmann, R.; Aulakh, M.S. The role of rice plants in regulating mechanisms of methane missions. Biology and Fertility of Soils 2000, 31, 20–29. [Google Scholar] [CrossRef]
- Garnet, K.N.; Megonigal, J.P.; Litchfield, C.; Taylor Jr, G.E. Physiological control of leaf methane emission from wetland plants. Aquatic Botany 2005, 81, 141–155. [Google Scholar] [CrossRef]
- McLeod, A.R.; Fry, S.C.; Loake, G.J.; Messenger, D.J.; Reay, D.S.; Smith, K.A.; Yun, B.W. Ultraviolet radiation drives methane emissions from terrestrial plant pectins. New Phytologist 2008, 180, 124–132. [Google Scholar] [CrossRef]
- Putkinen, A.; Siljanen, H.M.; Laihonen, A.; Paasisalo, I.; Porkka, K.; Tiirola, M.; Haikarainen, I.; Tenhovirta, S.; Pihlatie, M. New insight to the role of microbes in the methane exchange in trees: evidence from metagenomic sequencing. New Phytologist 2021, 231, 524–536. [Google Scholar] [CrossRef]
- Nisbet, R.; Fisher, R.; Nimmo, R.; Bendall, D.; Crill, P.; Gallego-Sala, A.V.; Hornibrook, E.; López-Juez, E.; Lowry, D.; Nisbet, P.J.P.o.t.R.S.B.B.S. Emission of methane from plants. 2009, 276, 1347-1354.
- Li, L.; Wei, S.; Shen, W. The role of methane in plant physiology: a review. Plant cell reports 2020, 39, 171–179. [Google Scholar] [CrossRef]
- Wang, N.; Huang, D.; Li, C.; Deng, Y.; Li, W.; Yao, Y.; Liao, W. Regulatory roles of methane in plants. Scientia Horticulturae 2020, 272, 109492. [Google Scholar] [CrossRef]
- UNEP, U.J.U.E.P. Emissions gap report 2020. 2020.
- SUN, B.-f.; Hong, Z.; Lü, Y.-z.; Fei, L.; WANG, X.-k.J.J.o.i.a. The effects of nitrogen fertilizer application on methane and nitrous oxide emission/uptake in Chinese croplands. 2016, 15, 440-450.
- Xu, X.; Wu, Z.; Dong, Y.; Zhou, Z.; Xiong, Z.J.S.r. Effects of nitrogen and biochar amendment on soil methane concentration profiles and diffusion in a rice-wheat annual rotation system. 2016, 6, 1-13.
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M.J.M.p. Metabolic and signaling aspects underpinning the regulation of plant carbon nitrogen interactions. 2010, 3, 973-996.
- Stokstad, E. The nitrogen fix. 2016.
- Tokarz, E.; Urban, D.J.J.o.E.E. Soil redox potential and its impact on microorganisms and plants of wetlands. 2015, 16, 20--30.
- Wilmoth, J.L.; Schaefer, J.K.; Schlesinger, D.R.; Roth, S.W.; Hatcher, P.G.; Shoemaker, J.K.; Zhang, X.J.G.C.B. The role of oxygen in stimulating methane production in wetlands. 2021, 27, 5831-5847.
- Morard, P.; Silvestre, J.J.P. ; soil. Plant injury due to oxygen deficiency in the root environment of soilless culture: a review. 1996, 184, 243–254. [Google Scholar]
- Duyen, D.V.; Kwon, Y.; Kabange, N.R.; Lee, J.-Y.; Lee, S.-M.; Kang, J.-W.; Park, H.; Cha, J.-K.; Cho, J.-H.; Shin, D.J.P. Novel QTL Associated with Aerenchyma-Mediated Radial Oxygen Loss (ROL) in Rice (Oryza sativa L. ) under Iron (II) Sulfide. 2022, 11, 788. [Google Scholar]
- Shan, Y.; Johnson-Beebout, S.; Buresh, R.J.A.i.a. Crop residue management for lowland rice-based cropping systems in Asia. 2008, 98, 117-199.
- Dobermann, A.; Fairhurst, T.J.B.C.I. Rice straw management. 2002, 16, 7-11.
- Singh, D.; Dhiman, S.K.; Kumar, V.; Babu, R.; Shree, K.; Priyadarshani, A.; Singh, A.; Shakya, L.; Nautiyal, A.; Saluja, S.J.A. Crop Residue Burning and Its Relationship between Health, Agriculture Value Addition, and Regional Finance. 2022, 13, 1405.
- Andini, A.; Bonnet, S.; Rousset, P.; Hasanudin, U.J.C.S. Impact of open burning of crop residues on air pollution and climate change in Indonesia. 2018, 115, 2259-2266.
- Romasanta, R.R.; Sander, B.O.; Gaihre, Y.K.; Alberto, M.C.; Gummert, M.; Quilty, J.; Castalone, A.G.; Balingbing, C.; Sandro, J.; Correa Jr, T.J.A., ecosystems; et al. How does burning of rice straw affect CH4 and N2O emissions? A comparative experiment of different on-field straw management practices. 2017, 239, 143–153. [Google Scholar]
- Baggs, E.M.; Philippot, L. Microbial terrestrial pathways to nitrous oxide. Nitrous oxide and climate change 2010, 256. [Google Scholar]
- Khalil, K.; Mary, B.; Renault, P.J.S.B. ; Biochemistry. Nitrous oxide production by nitrification and denitrification in soil aggregates as affected by O2 concentration. 2004, 36, 687–699. [Google Scholar]
- Dalal, R.C.; Wang, W.; Robertson, G.P.; Parton, W.J.J.S.R. Nitrous oxide emission from Australian agricultural lands and mitigation options: a review. 2003, 41, 165-195.
- Dalal, R.; Allen, D.; Livesley, S.; Richards, G.J.P. ; Soil. Magnitude and biophysical regulators of methane emission and consumption in the Australian agricultural, forest, and submerged landscapes: a review. 2008, 309, 43–76. [Google Scholar]
- Xu, X.; Sharma, P.; Shu, S.; Lin, T.; Ciais, P.; Tubiello, F.; Smith, P.; Campbell, N.; Jain, A. Global greenhouse gas emissions from animal-based foods are twice those of plant-based foods. Nature Food 2 (9): 724–732. 2021.
- Bridgham, S.D.; Cadillo-Quiroz, H.; Keller, J.K.; Zhuang, Q. Methane emissions from wetlands: biogeochemical, microbial, and modeling perspectives from local to global scales. Global change biology 2013, 19, 1325–1346. [Google Scholar] [CrossRef]
- Lyu, Z.; Shao, N.; Akinyemi, T.; Whitman, W.B. Methanogenesis. Current Biology 2018, 28, R727–R732. [Google Scholar] [CrossRef]
- Fournier, G.P.; Gogarten, J.P. Evolution of acetoclastic methanogenesis in Methanosarcina via horizontal gene transfer from cellulolytic Clostridia. Journal of bacteriology 2008, 190, 1124–1127. [Google Scholar] [CrossRef]
- Tveit, A.T.; Hestnes, A.G.; Robinson, S.L.; Schintlmeister, A.; Dedysh, S.N.; Jehmlich, N.; von Bergen, M.; Herbold, C.; Wagner, M.; Richter, A.J.P.o.t.N.A.o.S. Widespread soil bacterium that oxidizes atmospheric methane. 2019, 116, 8515-8524.
- Hanson, R.S.; Hanson, T.E.J.M.r. Methanotrophic bacteria. 1996, 60, 439-471.
- Kirschke, S.; Bousquet, P.; Ciais, P.; Saunois, M.; Canadell, J.G.; Dlugokencky, E.J.; Bergamaschi, P.; Bergmann, D.; Blake, D.R.; Bruhwiler, L.J.N.g. Three decades of global methane sources and sinks. 2013, 6, 813-823.
- Hink, L.; Nicol, G.W.; Prosser, J.I.J.E.m. Archaea produce lower yields of N2O than bacteria during aerobic ammonia oxidation in soil. 2017, 19, 4829-4837.
- Hink, L.; Lycus, P.; Gubry-Rangin, C.; Frostegård, Å.; Nicol, G.W.; Prosser, J.I.; Bakken, L.R.J.E.M. Kinetics of NH3-oxidation, NO-turnover, N2O-production and electron flow during oxygen depletion in model bacterial and archaeal ammonia oxidisers. 2017, 19, 4882-4896.
- Schmidt-Rohr, K.J.A.o. Oxygen is the high-energy molecule powering complex multicellular life: Fundamental corrections to traditional bioenergetics. 2020, 5, 2221-2233.
- Murrell, J. The aerobic methane oxidizing bacteria (methanotrophs). In Handbook of hydrocarbon and lipid microbiology; 2010.
- Reim, A.; Lüke, C.; Krause, S.; Pratscher, J.; Frenzel, P.J.T.I.j. One millimetre makes the difference: high-resolution analysis of methane-oxidizing bacteria and their specific activity at the oxic–anoxic interface in a flooded paddy soil. 2012, 6, 2128-2139.
- Stein, L.Y.; Roy, R.; Dunfield, P.F.J.e. Aerobic methanotrophy and nitrification: processes and connections. 2012.
- Walkiewicz, A.; Brzezińska, M.; Bieganowski, A.J.B.; Soils, F.o. Methanotrophs are favored under hypoxia in ammonium-fertilized soils. 2018, 54, 861-870.
- Ravishankara, A.; Daniel, J.S.; Portmann, R.W.J.s. Nitrous oxide (N2O): the dominant ozone-depleting substance emitted in the 21st century. 2009, 326, 123-125.
- Thomson, A.J.; Giannopoulos, G.; Pretty, J.; Baggs, E.M.; Richardson, D.J. Biological sources and sinks of nitrous oxide and strategies to mitigate emissions. Philosophical Transactions of the Royal Society B: Biological Sciences 2012, 367, 1157–1168. [Google Scholar] [CrossRef]
- Bange, H.W.; Freing, A.; Kock, A.; Löscher, C.R. Marine pathways to nitrous oxide. Nitrous oxide and climate change 2010, 2, 36–62. [Google Scholar]
- Inatomi, M.; Hajima, T.; Ito, A.J.P.o. Fraction of nitrous oxide production in nitrification and its effect on total soil emission: A meta-analysis and global-scale sensitivity analysis using a process-based model. 2019, 14, e0219159.
- Liu, R.; Hu, H.; Suter, H.; Hayden, H.L.; He, J.; Mele, P.; Chen, D.J.F.i.m. Nitrification is a primary driver of nitrous oxide production in laboratory microcosms from different land-use soils. 2016, 7, 1373.
- Codispoti, L.; Christensen, J. Nitrification, denitrification and nitrous oxide cycling in the eastern tropical South Pacific Ocean. Marine chemistry 1985, 16, 277–300. [Google Scholar] [CrossRef]
- Ji, Q.; Buitenhuis, E.; Suntharalingam, P.; Sarmiento, J.L.; Ward, B.B. Global nitrous oxide production determined by oxygen sensitivity of nitrification and denitrification. Global Biogeochemical Cycles 2018, 32, 1790–1802. [Google Scholar] [CrossRef]
- Scala, D.J.; Kerkhof, L.J. Nitrous oxide reductase (nosZ) gene-specific PCR primers for detection of denitrifiers and three nosZ genes from marine sediments. FEMS Microbiology Letters 1998, 162, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Bouwman, A.; Stehfest, E.; van Kessel, C. Nitrous oxide emissions from the nitrogen cycle in arable agriculture: estimation and mitigation. Nitrous oxide and climate change 2010, 85–106. [Google Scholar]
- Ji, Q.; Babbin, A.R.; Jayakumar, A.; Oleynik, S.; Ward, B.B.J.G.R.L. Nitrous oxide production by nitrification and denitrification in the Eastern Tropical South Pacific oxygen minimum zone. 2015, 42, 10,755-710,764.
- Wrage, N.; Velthof, G.L.; Van Beusichem, M.L.; Oenema, O.J.S.b. ; Biochemistry. Role of nitrifier denitrification in the production of nitrous oxide. 2001, 33, 1723–1732. [Google Scholar]
- Zhang, J.; Mueller, C.; Cai, Z.J.S.B. ; Biochemistry. Heterotrophic nitrification of organic N and its contribution to nitrous oxide emissions in soils. 2015, 84, 199–209. [Google Scholar]
- Livesley, S.; Kiese, R.; Graham, J.; Weston, C.; Butterbach-Bahl, K.; Arndt, S.J.P. ; Soil. Trace gas flux and the influence of short-term soil water and temperature dynamics in Australian sheep grazed pastures of differing productivity. 2008, 309, 89–103. [Google Scholar]
- Jørgensen, C.J.; Elberling, B.J.S.B. ; Biochemistry. Effects of flooding-induced N2O production, consumption and emission dynamics on the annual N2O emission budget in wetland soil. 2012, 53, 9–17. [Google Scholar]
- Mørkved, P.T.; Dörsch, P.; Bakken, L.R.J.S.B. ; Biochemistry. The N2O product ratio of nitrification and its dependence on long-term changes in soil pH. 2007, 39, 2048–2057. [Google Scholar]
- de Klein, C.A.; Eckard, R.J.; van der Weerden, T.J. Nitrous oxide emissions from the nitrogen cycle in livestock agriculture: estimation and mitigation. Nitrous oxide and climate change 2010, 107–144. [Google Scholar]
- Conen, F.; Neftel, A. Nitrous oxide emissions from land-use and land-management change. Nitrous Oxide and Climate Change 2010, 143–159. [Google Scholar]
- Well, R.; Butterbach-Bahl, K. Indirect emissions of nitrous oxide from nitrogen deposition and leaching of agricultural nitrogen. Nitrous Oxide and Climate Change (ed Smith K), 2010; 162, 75. [Google Scholar]
- Gubry-Rangin, C.; Nicol, G.W.; Prosser, J.I.J.F.m.e. Archaea rather than bacteria control nitrification in two agricultural acidic soils. 2010, 74, 566-574.
- Stein, L.Y.J.C.o.i.c.b. Insights into the physiology of ammonia-oxidizing microorganisms. 2019, 49, 9-15.
- Di, H.J.; Cameron, K.C.; Shen, J.-P.; Winefield, C.S.; O’Callaghan, M.; Bowatte, S.; He, J.-Z.J.N.G. Nitrification driven by bacteria and not archaea in nitrogen-rich grassland soils. 2009, 2, 621-624.
- Di, H.J.; Cameron, K.C.; Sherlock, R.R.; Shen, J.-P.; He, J.-Z.; Winefield, C.S.J.J.o.S. ; Sediments. Nitrous oxide emissions from grazed grassland as affected by a nitrification inhibitor, dicyandiamide, and relationships with ammonia-oxidizing bacteria and archaea. 2010, 10, 943–954. [Google Scholar]
- Treusch, A.H.; Leininger, S.; Kletzin, A.; Schuster, S.C.; Klenk, H.P.; Schleper, C.J.E.m. Novel genes for nitrite reductase and Amo-related proteins indicate a role of uncultivated mesophilic crenarchaeota in nitrogen cycling. 2005, 7, 1985-1995.
- Leininger, S.; Urich, T.; Schloter, M.; Schwark, L.; Qi, J.; Nicol, G.W.; Prosser, J.I.; Schuster, S.; Schleper, C.J.N. Archaea predominate among ammonia-oxidizing prokaryotes in soils. 2006, 442, 806-809.
- Offre, P.; Prosser, J.I.; Nicol, G.W.J.F.M.E. Growth of ammonia-oxidizing archaea in soil microcosms is inhibited by acetylene. 2009, 70, 99-108.
- He, J.z.; Shen, J.p.; Zhang, L.m.; Zhu, Y.g.; Zheng, Y.m.; Xu, M.g.; Di, H.J.E.m. Quantitative analyses of the abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea of a Chinese upland red soil under long-term fertilization practices. 2007, 9, 2364-2374.
- Shen, J.p.; Zhang, L.m.; Zhu, Y.g.; Zhang, J.b.; He, J.z.J.E.M. Abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea communities of an alkaline sandy loam. 2008, 10, 1601-1611.
- Freitag, A.; Rudert, M.; Bock, E.J.F.M.L. Growth of Nitrobacter by dissimilatoric nitrate reduction. 1987, 48, 105-109.
- Daims, H.; Lebedeva, E.V.; Pjevac, P.; Han, P.; Herbold, C.; Albertsen, M.; Jehmlich, N.; Palatinszky, M.; Vierheilig, J.; Bulaev, A.J.N. Complete nitrification by Nitrospira bacteria. 2015, 528, 504-509.
- Van Kessel, M.A.; Speth, D.R.; Albertsen, M.; Nielsen, P.H.; Op den Camp, H.J.; Kartal, B.; Jetten, M.S.; Lücker, S.J.N. Complete nitrification by a single microorganism. 2015, 528, 555-559.
- Zou, J.; Huang, Y.; Qin, Y.; Liu, S.; Shen, Q.; Pan, G.; Lu, Y.; Liu, Q.J.G.C.B. Changes in fertilizer-induced direct N2O emissions from paddy fields during rice-growing season in China between 1950s and 1990s. 2009, 15, 229-242.
- Ma, J.; Li, X.; Xu, H.; Han, Y.; Cai, Z.; Yagi, K.J.S.R. Effects of nitrogen fertiliser and wheat straw application on CH4 and N2O emissions from a paddy rice field. 2007, 45, 359-367.
- Davidson, E.A.J.N.G. The contribution of manure and fertilizer nitrogen to atmospheric nitrous oxide since 1860. 2009, 2, 659-662.
- Syakila, A.; Kroeze, C.J.G.g.m. ; management. The global nitrous oxide budget revisited. 2011, 1, 17–26. [Google Scholar]
- Prather, M.J.; Holmes, C.D.; Hsu, J.J.G.R.L. Reactive greenhouse gas scenarios: Systematic exploration of uncertainties and the role of atmospheric chemistry. 2012, 39. 39.
- Reay, D.S.; Grace, J. Carbon dioxide: importance, sources and sinks. In Greenhouse gas sinks; CABI Wallingford UK: 2007; pp. 1-10.
- Xu, X.; Sharma, P.; Shu, S.; Lin, T.-S.; Ciais, P.; Tubiello, F.N.; Smith, P.; Campbell, N.; Jain, A.K.J.N.F. Global greenhouse gas emissions from animal-based foods are twice those of plant-based foods. 2021, 2, 724-732.
- Chang, J.; Ciais, P.; Gasser, T.; Smith, P.; Herrero, M.; Havlík, P.; Obersteiner, M.; Guenet, B.; Goll, D.S.; Li, W.J.N.C. Climate warming from managed grasslands cancels the cooling effect of carbon sinks in sparsely grazed and natural grasslands. 2021, 12, 118.
- Adopted, I.J.I.G. , Szwitzerland. Climate change 2014 synthesis report. 2014. [Google Scholar]
- Poore, J.; Nemecek, T.J.S. Reducing food’s environmental impacts through producers and consumers. 2018, 360, 987-992.
- Casper, J.K. Agriculture: the food we grow and animals we raise; Infobase Publishing: 2007.
- Rakotoson, T.; Dusserre, J.; Letourmy, P.; Frouin, J.; Ratsimiala, I.R.; Rakotoarisoa, N.V.; Vom Brocke, K.; Ramanantsoanirina, A.; Ahmadi, N.; Raboin, L.-M.J.R.S. Genome-wide association study of nitrogen use efficiency and agronomic traits in upland rice. 2021, 28, 379-390.
- Bandyopadhyay, T.; Swarbreck, S.M.; Jaiswal, V.; Maurya, J.; Gupta, R.; Bentley, A.R.; Griffiths, H.; Prasad, M.J.J.o.A.R. GWAS identifies genetic loci underlying nitrogen responsiveness in the climate resilient C4 model Setaria italica (L. ). 2022. [Google Scholar] [CrossRef]
- Akhatar, J.; Goyal, A.; Kaur, N.; Atri, C.; Mittal, M.; Singh, M.P.; Kaur, R.; Rialch, I.; Banga, S.S.J.S.r. Genome wide association analyses to understand genetic basis of flowering and plant height under three levels of nitrogen application in Brassica juncea (L. ) Czern & Coss. 2021, 11, 1–14. [Google Scholar]
- Nazish, T.; Arshad, M.; Jan, S.U.; Javaid, A.; Khan, M.H.; Naeem, M.A.; Baber, M.; Ali, M.J.T.R. Transporters and transcription factors gene families involved in improving nitrogen use efficiency (NUE) and assimilation in rice (Oryza sativa L. ). 2021, 1–20. [Google Scholar] [CrossRef]
- Wang, Q.; Su, Q.; Nian, J.; Zhang, J.; Guo, M.; Dong, G.; Hu, J.; Wang, R.; Wei, C.; Li, G.J.M.P. The Ghd7 transcription factor represses ARE1 expression to enhance nitrogen utilization and grain yield in rice. 2021, 14, 1012-1023.
- Han, M.-L.; Lv, Q.-Y.; Zhang, J.; Wang, T.; Zhang, C.-X.; Tan, R.-J.; Wang, Y.-L.; Zhong, L.-Y.; Gao, Y.-Q.; Chao, Z.-F.J.M.P. Decreasing nitrogen assimilation under drought stress by suppressing DST-mediated activation of Nitrate Reductase 1. 2 in rice. 2022, 15, 167–178. [Google Scholar] [CrossRef]
- Zhang, Z.; Chu, C.J.M.P. Nitrogen-use divergence between indica and japonica rice: variation at nitrate assimilation. 2020, 13, 6-7.
- Teng, W.; He, X.; Tong, Y.J.P. Genetic Control of Efficient Nitrogen Use for High Yield and Grain Protein Concentration in Wheat: A Review. 2022, 11, 492.
- Kabange, N.R.; Park, S.-Y.; Shin, D.; Lee, S.-M.; Jo, S.-M.; Kwon, Y.; Cha, J.-K.; Song, Y.-C.; Ko, J.-M.; Lee, J.-H.J.A. Identification of a novel QTL for chlorate resistance in rice (Oryza sativa L. ). 2020, 10, 360. [Google Scholar] [CrossRef]
- Anas, M.; Liao, F.; Verma, K.K.; Sarwar, M.A.; Mahmood, A.; Chen, Z.-L.; Li, Q.; Zeng, X.-P.; Liu, Y.; Li, Y.-R. Fate of nitrogen in agriculture and environment: agronomic, eco-physiological and molecular approaches to improve nitrogen use efficiency. Biological Research 2020, 53, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lu, X.; Wang, C.; Shen, L.; Dai, L.; He, J.; Yang, L.; Li, P.; Hong, Y.; Zhang, Q.J.T.C.J. Genome-wide association study and transcriptome analysis reveal new QTL and candidate genes for nitrogen-deficiency tolerance in rice. 2022.
- Fan, X.; Naz, M.; Fan, X.; Xuan, W.; Miller, A.J.; Xu, G.J.J.o.E.B. Plant nitrate transporters: from gene function to application. 2017, 68, 2463-2475.
- Lea, P.J.; Miflin, B.J.J.P.P. ; Biochemistry. Glutamate synthase and the synthesis of glutamate in plants. 2003, 41, 555–564. [Google Scholar]
- Prasad, L.R.V.; Mailapalli, D.R.J.C.i.S.S.; Analysis, P. Evaluation of nitrogen fertilization patterns using DSSAT for enhancing grain yield and nitrogen use efficiency in rice. 2018, 49, 1401-1417.
- Nakidakida, T.; Hayashi, H. Nitrogen recovery and nitrogen use efficiency of potatoes in an integrated compost fertilization system in an Andosol soil. In Proceedings of the III International Symposium on Organic Matter Management and Compost Use in Horticulture 1146; 2015; pp. 41–48. [Google Scholar]
- Li, H.; Hu, B.; Chu, C.J.J.o.E.B. Nitrogen use efficiency in crops: lessons from Arabidopsis and rice. 2017, 68, 2477-2488.
- Duan, Y.; Zhang, Y.; Ye, L.; Fan, X.; Xu, G.; Shen, Q.J.A.o.B. Responses of rice cultivars with different nitrogen use efficiency to partial nitrate nutrition. 2007, 99, 1153-1160.
- Djaman, K.; Mel, V.; Ametonou, F.; El-Namaky, R.; Diallo, M.; Koudahe, K.J.J.o.A.S.; Research, F. Effect of nitrogen fertilizer dose and application timing on yield and nitrogen use efficiency of irrigated hybrid rice under semi-arid conditions. 2018.
- Gallais, A.; Hirel, B.J.J.o.e.b. An approach to the genetics of nitrogen use efficiency in maize. 2004, 55, 295-306.
- Liu, X.; Hu, B.; Chu, C.J.J.o.G. ; Genomics. Nitrogen assimilation in plants: current status and future prospects. 2021.
- Hirel, B.; Tétu, T.; Lea, P.J.; Dubois, F.J.S. Improving nitrogen use efficiency in crops for sustainable agriculture. 2011, 3, 1452-1485.
- Yamaya, T.; Obara, M.; Nakajima, H.; Sasaki, S.; Hayakawa, T.; Sato, T.J.J.o.e.b. Genetic manipulation and quantitative-trait loci mapping for nitrogen recycling in rice. 2002, 53, 917-925.
- Zhang, S.; Zhu, L.; Shen, C.; Ji, Z.; Zhang, H.; Zhang, T.; Li, Y.; Yu, J.; Yang, N.; He, Y.J.T.P.C. Natural allelic variation in a modulator of auxin homeostasis improves grain yield and nitrogen use efficiency in rice. 2021, 33, 566-580.
- Kiba, T.; Kudo, T.; Kojima, M.; Sakakibara, H.J.J.o.e.b. Hormonal control of nitrogen acquisition: roles of auxin, abscisic acid, and cytokinin. 2011, 62, 1399-1409.
- Séguéla, M.; Briat, J.F.; Vert, G.; Curie, C.J.T.P.J. Cytokinins negatively regulate the root iron uptake machinery in Arabidopsis through a growth-dependent pathway. 2008, 55, 289-300.
- Maruyama-Nakashita, A.; Nakamura, Y.; Yamaya, T.; Takahashi, H.J.T.P.J. A novel regulatory pathway of sulfate uptake in Arabidopsis roots: implication of CRE1/WOL/AHK4-mediated cytokinin-dependent regulation. 2004, 38, 779-789.
- Wagner, B.M.; Beck, E.J.P. Cytokinins in the perennial herb Urtica dioica L. as influenced by its nitrogen status. 1993, 190, 511–518. [Google Scholar]
- Franco-Zorrilla, J.M.; Martin, A.C.; Solano, R.; Rubio, V.; Leyva, A.; Paz-Ares, J.J.T.P.J. Mutations at CRE1 impair cytokinin-induced repression of phosphate starvation responses in Arabidopsis. 2002, 32, 353-360.
- Nacry, P.; Canivenc, G.; Muller, B.; Azmi, A.; Van Onckelen, H.; Rossignol, M.; Doumas, P.J.P.P. A role for auxin redistribution in the responses of the root system architecture to phosphate starvation in Arabidopsis. 2005, 138, 2061-2074.
- Vidal, E.A.; Araus, V.; Lu, C.; Parry, G.; Green, P.J.; Coruzzi, G.M.; Gutiérrez, R.A.J.P.o.t.N.A.o.S. Nitrate-responsive miR393/AFB3 regulatory module controls root system architecture in Arabidopsis thaliana. 2010, 107, 4477-4482.
- Chen, K.-E.; Chen, H.-Y.; Tseng, C.-S.; Tsay, Y.-F. Improving nitrogen use efficiency by manipulating nitrate remobilization in plants. Nature Plants 2020, 6, 1126–1135. [Google Scholar] [CrossRef]
- Ueda, Y.; Yanagisawa, S. Transcription factor-based genetic engineering to increase nitrogen use efficiency. In Engineering Nitrogen Utilization in Crop Plants; Springer: 2018; pp. 37-55.
- Wu, J.; Zhang, Z.S.; Xia, J.Q.; Alfatih, A.; Song, Y.; Huang, Y.J.; Wan, G.Y.; Sun, L.Q.; Tang, H.; Liu, Y. Rice NIN-LIKE PROTEIN 4 plays a pivotal role in nitrogen use efficiency. Plant Biotechnology Journal 2021, 19, 448–461. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ding, G.; Yang, N.; White, P.J.; Ye, X.; Cai, H.; Lu, J.; Shi, L.; Xu, F. Comparative genome and transcriptome analysis unravels key factors of nitrogen use efficiency in Brassica napus L. Plant, cell & environment 2020, 43, 712–731. [Google Scholar]
- Heuermann, D.; Hahn, H.; Von Wirén, N.J.F.i.p.s. Seed yield and nitrogen efficiency in oilseed rape after ammonium nitrate or urea fertilization. 2021, 11, 2197.
- Ruffel, S.; Krouk, G.; Ristova, D.; Shasha, D.; Birnbaum, K.D.; Coruzzi, G.M.J.P.o.t.N.A.o.S. Nitrogen economics of root foraging: transitive closure of the nitrate–cytokinin relay and distinct systemic signaling for N supply vs. demand. 2011, 108, 18524–18529. [Google Scholar] [CrossRef]
- Gu, J.; Li, Z.; Mao, Y.; Struik, P.C.; Zhang, H.; Liu, L.; Wang, Z.; Yang, J.J.P.S. Roles of nitrogen and cytokinin signals in root and shoot communications in maximizing of plant productivity and their agronomic applications. 2018, 274, 320-331.
- Azarakhsh, M.; Lebedeva, M.A.; Lutova, L.A.J.F.i.P.S. Identification and expression analysis of Medicago truncatula isopentenyl transferase genes (IPTs) involved in local and systemic control of nodulation. 2018, 9, 304. 9.
- Lin, J.; Roswanjaya, Y.P.; Kohlen, W.; Stougaard, J.; Reid, D.J.N.c. Nitrate restricts nodule organogenesis through inhibition of cytokinin biosynthesis in Lotus japonicus. 2021, 12, 1-12.
- Sasaki, T.; Suzaki, T.; Soyano, T.; Kojima, M.; Sakakibara, H.; Kawaguchi, M.J.N.c. Shoot-derived cytokinins systemically regulate root nodulation. 2014, 5, 1-9. 5.
- Singh, B.N.; Dwivedi, P.; Sarma, B.K.; Singh, G.S.; Singh, H.B.J.B. A novel function of N-signaling in plants with special reference to Trichoderma interaction influencing plant growth, nitrogen use efficiency, and cross talk with plant hormones. 2019, 9, 1-13.
- Mundim, F.M.; Pringle, E.G.J.F.i.p.s. Whole-plant metabolic allocation under water stress. 2018, 9, 852. 9.
- Rolly, N.K.; Yun, B.-W.J.P. Regulation of Nitrate (NO3) Transporters and Glutamate Synthase-Encoding Genes under Drought Stress in Arabidopsis: The Regulatory Role of AtbZIP62 Transcription Factor. 2021, 10, 2149.
- Rolly, N.K.; Mun, B.-G.; Yun, B.-W.J.G. Insights into the Transcriptional Regulation of Branching Hormonal Signaling Pathways Genes under Drought Stress in Arabidopsis. 2021, 12, 298.
- Sun, L.; Di, D.-W.; Li, G.; Li, Y.; Kronzucker, H.J.; Shi, W.J.J.o.P.P. Transcriptome analysis of rice (Oryza sativa L. ) in response to ammonium resupply reveals the involvement of phytohormone signaling and the transcription factor OsJAZ9 in reprogramming of nitrogen uptake and metabolism. 2020, 246, 153137. [Google Scholar]
- Zhong, L.; Chen, D.; Min, D.; Li, W.; Xu, Z.; Zhou, Y.; Li, L.; Chen, M.; Ma, Y. AtTGA4, a bZIP transcription factor, confers drought resistance by enhancing nitrate transport and assimilation in Arabidopsis thaliana. Biochemical and Biophysical Research Communications 2015, 457, 433–439. [Google Scholar] [CrossRef]
- Gupta, K.J. Plant Respiration and Internal Oxygen.
- Armstrong, W.; Armstrong, J. Plant internal oxygen transport (diffusion and convection) and measuring and modelling oxygen gradients. In Low-oxygen stress in plants; Springer: 2014; pp. 267-297.
- Kosmacz, M.; Weits, D.A. Oxygen perception in plants. In Low-Oxygen Stress in Plants; Springer: 2014; pp. 3-17.
- Guan, B.; Lin, Z.; Liu, D.; Li, C.; Zhou, Z.; Mei, F.; Li, J.; Deng, X.J.F.i.p.s. Effect of waterlogging-induced autophagy on programmed cell death in Arabidopsis roots. 2019, 10, 468.
- Armstrong, W. Advances in Botanical Research, Volume 7. Aeration in higher plants. Woolhouse HW. 1979. [Google Scholar]
- Sifton, H.J.T.B.R. Air-space tissue in plants. 1945, 11, 108-143.
- Roland, J.J.J.o.C.S. Cell wall differentiation and stages involved with intercellular gas space opening. 1978, 32, 325-336.
- Jeffree, C.; Dale, J.; Fry, S.J.P. The genesis of intercellular spaces in developing leaves ofPhaseolus vulgaris L. 1986, 132, 90-98.
- RAVEN, J.A.J.A.o.B. Into the voids: the distribution, function, development and maintenance of gas spaces in plants. 1996, 78, 137-142.
- Wegner, L.H. Oxygen transport in waterlogged plants. In Waterlogging signalling and tolerance in plants; Springer: 2010; pp. 3-22.
- Lai, W.-L.; Zhang, Y.; Chen, Z.-H.J.E.E. Radial oxygen loss, photosynthesis, and nutrient removal of 35 wetland plants. 2012, 39, 24-30.
- Mohammed, U.; Caine, R.; Atkinson, J.; Harrison, E.; Wells, D.; Chater, C.; Gray, J.; Swarup, R.; Murchie, E.J.S.r. Rice plants overexpressing OsEPF1 show reduced stomatal density and increased root cortical aerenchyma formation. 2019, 9, 1-13.
- Scheehle, E.A.; Kruger, D.J.T.E.J. Global anthropogenic methane and nitrous oxide emissions. 2006.
- Denmead, O.; Freney, J.; Simpson, J.J.S.S.S.o.A.J. Nitrous oxide emission during denitrification in a flooded field. 1979, 43, 716-718.
- Bodelier, P.L.; Pérez, G.; Veraart, A.J.; Krause, S. Methanotroph ecology, environmental distribution and functioning. In Methanotrophs; Springer: 2019; pp. 1-38.
- Trotsenko, Y.A.; Murrell, J.C.J.A.i.a.m. Metabolic aspects of aerobic obligate methanotrophy⋆. 2008, 63, 183-229.
- Saggar, S.; Tate, K.; Giltrap, D.; Singh, J.J.P. ; Soil. Soil-atmosphere exchange of nitrous oxide and methane in New Zealand terrestrial ecosystems and their mitigation options: a review. 2008, 309, 25–42. [Google Scholar]
- Schütz, H.; Seiler, W.; Rennenberg, H.J.S. ; effect., t.g. Soil and land use related sources and sinks of methane (CH4) in the context of the global methane budget. 1990, 269-285.
- Horwath, W. Greenhouse gas emissions from rice cropping systems. In Understanding greenhouse gas emissions from agricultural management; ACS Publications: 2011; pp. 67-89.
- Colmer, T.D.; Kotula, L.; Malik, A.I.; Takahashi, H.; Konnerup, D.; Nakazono, M.; Pedersen, O.J.P. , Cell; Environment. Rice acclimation to soil flooding: low concentrations of organic acids can trigger a barrier to radial oxygen loss in roots. 2019, 42, 2183–2197. [Google Scholar]
- Ejiri, M.; Shiono, K.J.F.i.p.s. Prevention of radial oxygen loss is associated with exodermal suberin along adventitious roots of annual wild species of Echinochloa. 2019, 10, 254.
- Abiko, T.; Kotula, L.; Shiono, K.; Malik, A.I.; Colmer, T.D.; Nakazono, M.J.P. , Cell; Environment. Enhanced formation of aerenchyma and induction of a barrier to radial oxygen loss in adventitious roots of Zea nicaraguensis contribute to its waterlogging tolerance as compared with maize (Zea mays ssp. mays). 2012, 35, 1618–1630. [Google Scholar]
- Ejiri, M.; Fukao, T.; Miyashita, T.; Shiono, K.J.B.s. A barrier to radial oxygen loss helps the root system cope with waterlogging-induced hypoxia. 2021, 71, 40-50.
- Peralta Ogorek, L.L.; Pellegrini, E.; Pedersen, O.J.N.P. Novel functions of the root barrier to radial oxygen loss–radial diffusion resistance to H2 and water vapour. 2021, 231, 1365-1376.
- Foyer, C.H.; Noctor, G.; Hodges, M.J.J.o.E.B. Respiration and nitrogen assimilation: targeting mitochondria-associated metabolism as a means to enhance nitrogen use efficiency. 2011, 62, 1467-1482.
- Maier, R.J. Nitrogen fixation and respiration: Two processes linked by the energetic demands of Nitrogenase. In Respiration in archaea and bacteria; Springer: 2004; pp. 101-120.
- Weger, H.G.; Turpin, D.H.J.P.p. Mitochondrial respiration Can support NO3− and NO2− reduction during photosynthesis: interactions between photosynthesis, respiration, and N assimilation in the N-limited green alga Selenastrum minutum. 1989, 89, 409-415.
- De Laulanié, H.J.T. Intensive rice farming in Madagascar. 2011, 29, 183-187.
- Sinclair, T.R.J.R.T. Agronomic UFOs waste valuable scientific resources. 2004, 3. 3.
- Pereira-Mora, L.; Terra, J.A.; Fernández-Scavino, A.J.A.S.E. Methanogenic community linked to organic acids fermentation from root exudates are affected by rice intensification in rotational soil systems. 2022, 176, 104498.
- Chan, K.Y.; Xu, Z.J.B.f.e.m.s. ; technology. Biochar: nutrient properties and their enhancement. 2009, 1, 67–84. [Google Scholar]
- Lehmann, J.; Cowie, A.; Masiello, C.A.; Kammann, C.; Woolf, D.; Amonette, J.E.; Cayuela, M.L.; Camps-Arbestain, M.; Whitman, T.J.N.G. Biochar in climate change mitigation. 2021, 14, 883-892.
- Roe, S.; Streck, C.; Beach, R.; Busch, J.; Chapman, M.; Daioglou, V.; Deppermann, A.; Doelman, J.; Emmet-Booth, J.; Engelmann, J.J.G.C.B. Land-based measures to mitigate climate change: Potential and feasibility by country. 2021, 27, 6025-6058.
- Yang, W.; Feng, G.; Miles, D.; Gao, L.; Jia, Y.; Li, C.; Qu, Z.J.S.o.t.T.E. Impact of biochar on greenhouse gas emissions and soil carbon sequestration in corn grown under drip irrigation with mulching. 2020, 729, 138752.
- Lyu, H.; Zhang, H.; Chu, M.; Zhang, C.; Tang, J.; Chang, S.X.; Mašek, O.; Ok, Y.S.J.L.D. ; Development. Biochar affects greenhouse gas emissions in various environments: A critical review. 2022, 33, 3327–3342. [Google Scholar]
- Lin, X.; Xie, Z.; Zheng, J.; Liu, Q.; Bei, Q.; Zhu, J.J.E.J.o.S.S. Effects of biochar application on greenhouse gas emissions, carbon sequestration and crop growth in coastal saline soil. 2015, 66, 329-338.
- Joseph, S.; Lehmann, J.J.B.f.e.m.S. ; technology. Biochar for environmental management: an introduction. 2015, 1.
- Lehmann, J.; Czimczik, C.; Laird, D.; Sohi, S.J.B.f.e.m.s. ; technology. Stability of biochar in soil. 2009, 183–206. [Google Scholar]
- Major, J.; Steiner, C.; Downie, A.; Lehmann, J.; Joseph, S.J.B.f.e.m.S. ; technology. Biochar effects on nutrient leaching. 2009, 271. [Google Scholar]
- Turunen, M.; Hyväluoma, J.; Heikkinen, J.; Keskinen, R.; Kaseva, J.; Hannula, M.; Rasa, K.J.B.E. Quantifying the pore structure of different biochars and their impacts on the water retention properties of Sphagnum moss growing media. 2020, 191, 96-106.
- Gaunt, J.; Cowie, A.J.B.f.e.m.S. ; technology. Biochar, greenhouse gas accounting and emissions trading. 2009, 317-340.
- Smernik, R.J.J.B.f.e.m.s. ; technology. Biochar and sorption of organic compounds. 2009, 289–300. [Google Scholar]
- Thies, J.E.; Rillig, M.C.J.B.f.e.m.S. ; technology. Characteristics of biochar: biological properties. 2009, 1, 85–105. [Google Scholar]
- DeLuca, T.H.; Gundale, M.J.; MacKenzie, M.D.; Jones, D.L.J.B.f.e.m.s. , technology; implementation. Biochar effects on soil nutrient transformations. 2015, 2, 421–454. [Google Scholar]
- Rondon, M.A.; Molina, D.; Hurtado, M.; Ramirez, J.; Lehmann, J.; Major, J.; Amezquita, E. Enhancing the productivity of crops and grasses while reducing greenhouse gas emissions through bio-char amendments to unfertile tropical soils. In Proceedings of the 18th world congress of soil science; 2006; pp. 9–15. [Google Scholar]
- Yanai, Y.; Toyota, K.; Okazaki, M.J.S.s.; nutrition, p. Effects of charcoal addition on N2O emissions from soil resulting from rewetting air-dried soil in short-term laboratory experiments. 2007, 53, 181-188.
- Joseph, S.; Cowie, A.L.; Van Zwieten, L.; Bolan, N.; Budai, A.; Buss, W.; Cayuela, M.L.; Graber, E.R.; Ippolito, J.A.; Kuzyakov, Y.J.G.B. How biochar works, and when it doesn't: A review of mechanisms controlling soil and plant responses to biochar. 2021, 13, 1731-1764.
- Joseph, S.; Peacocke, C.; Lehmann, J.; Munroe, P.J.B.f.e.m.s. ; technology. Developing a biochar classification and test methods. 2009, 1, 107–126. [Google Scholar]
- Borchard, N.; Schirrmann, M.; Cayuela, M.L.; Kammann, C.; Wrage-Mönnig, N.; Estavillo, J.M.; Fuertes-Mendizábal, T.; Sigua, G.; Spokas, K.; Ippolito, J.A.J.S.o.t.T.E. Biochar, soil and land-use interactions that reduce nitrate leaching and N2O emissions: a meta-analysis. 2019, 651, 2354-2364.
- Riley, W.; Ortiz-Monasterio, I.; Matson, P.J.N.C.i.A. Nitrogen leaching and soil nitrate, nitrite, and ammonium levels under irrigated wheat in Northern Mexico. 2001, 61, 223-236.
- Song, X.; Pan, G.; Zhang, C.; Zhang, L.; Wang, H.J.E.H. ; Sustainability. Effects of biochar application on fluxes of three biogenic greenhouse gases: a meta-analysis. 2016, 2, e01202. [Google Scholar]
- He, Y.; Zhou, X.; Jiang, L.; Li, M.; Du, Z.; Zhou, G.; Shao, J.; Wang, X.; Xu, Z.; Hosseini Bai, S.J.G.B. Effects of biochar application on soil greenhouse gas fluxes: A meta-analysis. 2017, 9, 743-755.
- Kalu, S.; Kulmala, L.; Zrim, J.; Peltokangas, K.; Tammeorg, P.; Rasa, K.; Kitzler, B.; Pihlatie, M.; Karhu, K.J.F.i.E.S. Potential of biochar to reduce greenhouse gas emissions and increase nitrogen use efficiency in boreal arable soils in the long-term. 2022, 630.
- Sass, R.L.; Cicerone, R.J.J.P.o.t.N.A.o.S. Photosynthate allocations in rice plants: Food production or atmospheric methane? 2002, 99, 11993-11995.
- van Der Gon, H.D.; Kropff, M.; Van Breemen, N.; Wassmann, R.; Lantin, R.; Aduna, E.; Corton, T.; Van Laar, H.J.P.o.t.N.A.o.S. Optimizing grain yields reduces CH4 emissions from rice paddy fields. 2002, 99, 12021-12024.
- Su, J.; Hu, C.; Yan, X.; Jin, Y.; Chen, Z.; Guan, Q.; Wang, Y.; Zhong, D.; Jansson, C.; Wang, F.J.N. Expression of barley SUSIBA2 transcription factor yields high-starch low-methane rice. 2015, 523, 602-606.
- Stein, L.Y.; Klotz, M.G.J.B.S.T. Nitrifying and denitrifying pathways of methanotrophic bacteria. 2011, 39, 1826-1831.
- Canarini, A.; Kaiser, C.; Merchant, A.; Richter, A.; Wanek, W.J.F.i.P.S. Root exudation of primary metabolites: mechanisms and their roles in plant responses to environmental stimuli. 2019, 10, 157.
- Rougier, M.J.P.C.I.E.C. Secretory activity of the root cap. 1981, 542-574.
- Abbott, L.K.; Murphy, D.V. Soil biological fertility: a key to sustainable land use in agriculture; Springer Science & Business Media: 2003.
- Walker, T.S.; Bais, H.P.; Grotewold, E.; Vivanco, J.M.J.P.p. Root exudation and rhizosphere biology. 2003, 132, 44-51.
- Nardi, S.; Concheri, G.; Pizzeghello, D.; Sturaro, A.; Rella, R.; Parvoli, G.J.C. Soil organic matter mobilization by root exudates. 2000, 41, 653-658.
- Vives-Peris, V.; De Ollas, C.; Gómez-Cadenas, A.; Pérez-Clemente, R.M.J.P.c.r. Root exudates: from plant to rhizosphere and beyond. 2020, 39, 3-17.
- De-la-Pena, C.; Badri, D.V.; Lei, Z.; Watson, B.S.; Brandao, M.M.; Silva-Filho, M.C.; Sumner, L.W.; Vivanco, J.M.J.J.o.B.C. Root secretion of defense-related proteins is development-dependent and correlated with flowering time. 2010, 285, 30654-30665.
- Chai, Y.N.; Schachtman, D.P.J.T.i.P.S. Root exudates impact plant performance under abiotic stress. 2022, 27, 80-91.
- Salem, M.A.; Wang, J.Y.; Al-Babili, S.J.F.i.P.S. Metabolomics of plant root exudates: From sample preparation to data analysis. 2022, 13, 5035.
- Narula, N.; Kothe, E.; Behl, R.K.J.J.o.A.B.; Quality, F. Role of root exudates in plant-microbe interactions. 2012, 82, 122-130.
- Kumar, R.; Bhatia, R.; Kukreja, K.; Behl, R.K.; Dudeja, S.S.; Narula, N.J.J.o.b.m. Establishment of Azotobacter on plant roots: chemotactic response, development and analysis of root exudates of cotton (Gossypium hirsutum L. ) and wheat (Triticum aestivum L.). 2007, 47, 436–439. [Google Scholar]
- Lu, Y.; Wassmann, R.; Neue, H.; Huang, C.; Bueno, C.J.S.B. ; Biochemistry. Methanogenic responses to exogenous substrates in anaerobic rice soils. 2000, 32, 1683–1690. [Google Scholar]
- Moscôso, J.S.C.; Silva, L.S.d.; Pujol, S.B.; Giacomini, S.J.; Severo, F.F.; Marzari, L.B.; Molin, G.D.J.R.B.d.C.d.S. Methane emission induced by short-chain organic acids in lowland soil. 2019, 43. 43.
- Aulakh, M.S.; Wassmann, R.; Bueno, C.; Rennenberg, H.J.P. ; Soil. Impact of root exudates of different cultivars and plant development stages of rice (Oryza sativa L.) on methane production in a paddy soil. 2001, 230, 77–86. [Google Scholar]
- Smith, M.R.; Rao, I.M.; Merchant, A.J.F.i.P.S. Source-sink relationships in crop plants and their influence on yield development and nutritional quality. 2018, 9, 1889.
- Andersen, C.P.J.N.p. Source–sink balance and carbon allocation below ground in plants exposed to ozone. 2003, 157, 213-228.
- Wingler, A.; Einig, W.; Schaeffer, C.; Wallenda, T.; Hampp, R.; Wallander, H.; NYLUND, J.E.J.N.p. Influence of different nutrient regimes on the regulation of carbon metabolism in Norway spruce [Picea abies (L. ) Karst.] seedlings. 1994, 128, 323–330. [Google Scholar] [CrossRef]
- Koch, K.J.A.r.o.p.b. Carbohydrate-modulated gene expression in plants. 1996, 47, 509-540.
- Mathur, J.; Hülskamp, M.J.C.B. Microtubules and microfilaments in cell morphogenesis in higher plants. 2002, 12, R669-R676.
- Yao, Z.; Zhou, Z.; Zheng, X.; Xie, B.; Mei, B.; Wang, R.; Butterbach-Bahl, K.; Zhu, J.J.P. ; Soil. Effects of organic matter incorporation on nitrous oxide emissions from rice-wheat rotation ecosystems in China. 2010, 327, 315–330. [Google Scholar]
- Zou, J.; Huang, Y.; Jiang, J.; Zheng, X.; Sass, R.L.J.G.b.c. A 3-year field measurement of methane and nitrous oxide emissions from rice paddies in China: Effects of water regime, crop residue, and fertilizer application. 2005, 19. 19.
- Eagle, A.J.; Bird, J.A.; Horwath, W.R.; Linquist, B.A.; Brouder, S.M.; Hill, J.E.; van Kessel, C.J.A.J. Rice yield and nitrogen utilization efficiency under alternative straw management practices. 2000, 92, 1096-1103.
- Li, X.; Zhang, G.; Xu, H.; Cai, Z.; Yagi, K.J.C. Effect of timing of joint application of hydroquinone and dicyandiamide on nitrous oxide emission from irrigated lowland rice paddy field. 2009, 75, 1417-1422.
- Akiyama, H.; Yan, X.; Yagi, K.J.G.C.B. Evaluation of effectiveness of enhanced-efficiency fertilizers as mitigation options for N2O and NO emissions from agricultural soils: meta-analysis. 2010, 16, 1837-1846.
- Majumdar, D.J.C.S. Methane and nitrous oxide emission from irrigated rice fields: proposed mitigation strategies. 2003, 1317-1326.
- Mahesh, M.; Mohini, M.J.A.i.D.R. Crop residues for sustainable livestock production. 2014, 1-2.
- Ritchie, H.; Rosado, P.; Roser, M.J.O.W.i.D. Meat and dairy production. 2017.
- Williams, P.J.N. ; Dietetics. Nutritional composition of red meat. 2007, 64, S113–S119. [Google Scholar]
- Porter, J.J.P.o.t.N.S. Milk as a source of lactose, vitamins and minerals. 1978, 37, 225-230.
- Shaheen, N.; Ahmed, M.K.; Islam, M.S.; Habibullah-Al-Mamun, M.; Tukun, A.B.; Islam, S.; MA Rahim, A.T.J.E.s.; research, p. Health risk assessment of trace elements via dietary intake of ‘non-piscine protein source’foodstuffs (meat, milk and egg) in Bangladesh. 2016, 23, 7794-7806.
- McAfee, A.J.; McSorley, E.M.; Cuskelly, G.J.; Moss, B.W.; Wallace, J.M.; Bonham, M.P.; Fearon, A.M.J.M.s. Red meat consumption: An overview of the risks and benefits. 2010, 84, 1-13.
- Willett, W.C.; Ludwig, D.S.J.N.E.J.o.M. Milk and health. 2020, 382, 644-654.
- Tricarico, J.; de Haas, Y.; Hristov, A.; Kebreab, E.; Kurt, T.; Mitloehner, F.; Pitta, D.J.J.o.d.s. Symposium review: Development of a funding program to support research on enteric methane mitigation from ruminants. 2022.
- Nairobi., U.N.E.P.J. Nairobi., U.N.E.P.J. Emissions gap report 2022: The closing window—Climate crisis calls for rapid transformation of societies. 2022.
- Frank, S.; Beach, R.; Havlík, P.; Valin, H.; Herrero, M.; Mosnier, A.; Hasegawa, T.; Creason, J.; Ragnauth, S.; Obersteiner, M.J.N.c. Structural change as a key component for agricultural non-CO2 mitigation efforts. 2018, 9, 1060.
- Arndt, C.; Hristov, A.N.; Price, W.J.; McClelland, S.C.; Pelaez, A.M.; Cueva, S.F.; Oh, J.; Dijkstra, J.; Bannink, A.; Bayat, A.R.J.P.o.t.N.A.o.S. Full adoption of the most effective strategies to mitigate methane emissions by ruminants can help meet the 1. 5 C target by 2030 but not 2050. 2022, 119, e2111294119. [Google Scholar]
- Zhao, Y.; Zhao, G.J.A.N. Decreasing ruminal methane production through enhancing the sulfate reduction pathway. 2022.
- Haque, M.N.J.J.o.a.s. ; technology. Dietary manipulation: a sustainable way to mitigate methane emissions from ruminants. 2018, 60, 1–10. [Google Scholar]
- Newbold, C.; El Hassan, S.; Wang, J.; Ortega, M.; Wallace, R.J.B.J.o.N. Influence of foliage from African multipurpose trees on activity of rumen protozoa and bacteria. 1997, 78, 237-249.
- Bodas, R.; Prieto, N.; García-González, R.; Andrés, S.; Giráldez, F.J.; López, S.J.A.F.S. ; Technology. Manipulation of rumen fermentation and methane production with plant secondary metabolites. 2012, 176, 78–93. [Google Scholar]
- Patra, A.K.; Saxena, J.J.A.v.L. Dietary phytochemicals as rumen modifiers: a review of the effects on microbial populations. 2009, 96, 363-375.
- Patra, A.; Saxena, J.J.N.r.r. The effect and mode of action of saponins on the microbial populations and fermentation in the rumen and ruminant production. 2009, 22, 204-219.
- Milich, L.J.G.E.C. The role of methane in global warming: where might mitigation strategies be focused? 1999, 9, 179-201.
- Benchaar, C.; Pomar, C.; Chiquette, J.J.C.J.o.A.S. Evaluation of dietary strategies to reduce methane production in ruminants: a modelling approach. 2001, 81, 563-574.
- Boadi, D.; Wittenberg, K.J.C.J.o.A.S. Methane production from dairy and beef heifers fed forages differing in nutrient density using the sulphur hexafluoride (SF6) tracer gas technique. 2002, 82, 201-206.
- Beauchemin, K.; Kreuzer, M.; O’mara, F.; McAllister, T.J.A.J.o.E.A. Nutritional management for enteric methane abatement: a review. 2008, 48, 21-27.
- Tamminga, S.; Bannink, A.; Dijkstra, J.; Zom, R. Feeding strategies to reduce methane loss in cattle; 1570-8616; Animal Sciences Group: 2007.
- O'mara, F.; Fitzgerald, J.; Murphy, J.; Rath, M.J.L.P.S. The effect on milk production of replacing grass silage with maize silage in the diet of dairy cows. 1998, 55, 79-87.
- Hassanat, F.; Gervais, R.; Julien, C.; Massé, D.; Lettat, A.; Chouinard, P.; Petit, H.; Benchaar, C.J.J.o.D.S. Replacing alfalfa silage with corn silage in dairy cow diets: Effects on enteric methane production, ruminal fermentation, digestion, N balance, and milk production. 2013, 96, 4553-4567.
- Martin, C.; Morgavi, D.P.; Doreau, M.J.A. Methane mitigation in ruminants: from microbe to the farm scale. 2010, 4, 351-365.
- Ferris, C.; Gordon, F.; Patterson, D.; Porter, M.; Yan, T.J.T.J.o.A.S. The effect of genetic merit and concentrate proportion in the diet on nutrient utilization by lactating dairy cows. 1999, 132, 483-490.
- Lovett, D.; Lovell, S.; Stack, L.; Callan, J.; Finlay, M.; Conolly, J.; O'Mara, F.J.L.P.S. Effect of forage/concentrate ratio and dietary coconut oil level on methane output and performance of finishing beef heifers. 2003, 84, 135-146.
- Johnson, K.A.; Johnson, D.E.J.J.o.a.s. Methane emissions from cattle. 1995, 73, 2483-2492.
- Murphy, M.; Baldwin, R.; Koong, L.J.J.o.A.S. Estimation of stoichiometric parameters for rumen fermentation of roughage and concentrate diets. 1982, 55, 411-421.
- Hindrichsen, I.; Wettstein, H.; Machmüller, A.; Jörg, B.; Kreuzer, M.J.E.m. ; assessment. Effect of the carbohydrate composition of feed concentratates on methane emission from dairy cows and their slurry. 2005, 107, 329–350. [Google Scholar]
- Hindrichsen, I.; Kreuzer, M.J.J.o.a.p.; nutrition, a. High methanogenic potential of sucrose compared with starch at high ruminal pH. 2009, 93, 61-65.
- Castillo, C.; Benedito, J.; Méndez, J.; Pereira, V.; Lopez-Alonso, M.; Miranda, M.; Hernández, J.J.A.F.S. ; Technology. Organic acids as a substitute for monensin in diets for beef cattle. 2004, 115, 101–116. [Google Scholar]
- McAllister, T.; Newbold, C.J.A.J.o.E.A. Redirecting rumen fermentation to reduce methanogenesis. 2008, 48, 7-13.
- Beauchemin, K.; McGinn, S.J.J.o.a.s. Methane emissions from beef cattle: Effects of fumaric acid, essential oil, and canola oil. 2006, 84, 1489-1496.
- Burt, S.J.I.j.o.f.m. Essential oils: their antibacterial properties and potential applications in foods—a review. 2004, 94, 223-253.
- Benchaar, C.; Calsamiglia, S.; Chaves, A.V.; Fraser, G.; Colombatto, D.; McAllister, T.A.; Beauchemin, K.A.J.A.F.S. ; Technology. A review of plant-derived essential oils in ruminant nutrition and production. 2008, 145, 209–228. [Google Scholar]
- Benchaar, C.; Greathead, H.J.A.F.S. ; Technology. Essential oils and opportunities to mitigate enteric methane emissions from ruminants. 2011, 166, 338–355. [Google Scholar]
- Moss, A.R.; Jouany, J.-P.; Newbold, J. Methane production by ruminants: its contribution to global warming. In Proceedings of the Annales de zootechnie; 2000; pp. 231–253. [Google Scholar]
- Lopez, S.; McIntosh, F.; Wallace, R.; Newbold, C.J.A.F.S. ; Technology. Effect of adding acetogenic bacteria on methane production by mixed rumen microorganisms. 1999, 78, 1–9. [Google Scholar]
- McAllister, T.; Beauchemin, K.; Alazzeh, A.; Baah, J.; Teather, R.; Stanford, K.J.C.J.o.A.S. The use of direct fed microbials to mitigate pathogens and enhance production in cattle. 2011, 91, 193-211.
- Newbold, C.J.; Rode, L. Dietary additives to control methanogenesis in the rumen. In Proceedings of the International congress series; 2006; pp. 138–147. [Google Scholar]
- Beauchemin, K.; Colombatto, D.; Morgavi, D.; Yang, W.J.J.o.A.S. Use of exogenous fibrolytic enzymes to improve feed utilization by ruminants. 2003, 81, E37-E47.
- Eun, J.-S.; Beauchemin, K.J.A.F.S. ; Technology. Assessment of the efficacy of varying experimental exogenous fibrolytic enzymes using in vitro fermentation characteristics. 2007, 132, 298–315. [Google Scholar]
- Hook, S.E.; Northwood, K.S.; Wright, A.-D.; McBride, B.W.J.A.; Microbiology, E. Long-term monensin supplementation does not significantly affect the quantity or diversity of methanogens in the rumen of the lactating dairy cow. 2009, 75, 374-380.
- Patra, A.K.J.E.M. ; Assessment. Enteric methane mitigation technologies for ruminant livestock: a synthesis of current research and future directions. 2012, 184, 1929–1952. [Google Scholar]
- McGinn, S.; Beauchemin, K.; Coates, T.; Colombatto, D.J.J.o.a.s. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast, and fumaric acid. 2004, 82, 3346-3356.
- Guan, H.; Wittenberg, K.; Ominski, K.; Krause, D.J.J.o.a.s. Efficacy of ionophores in cattle diets for mitigation of enteric methane. 2006, 84, 1896-1906.
- Clark, H.; Pinares-Patiño, C.; De Klein, C.J.G.A.G.R. Methane and nitrous oxide emissions from grazed grasslands. 2005, 279, 293.
- Madsen, J.; Lassen, J.; Hvelplund, T.; Weisbjerg, M.J.E.p. A fast, easy, reliable and cheap method to measure the methane production from ruminants. 2010, 121-122.
- Eckard, R.; Grainger, C.; De Klein, C.J.L.s. Options for the abatement of methane and nitrous oxide from ruminant production: A review. 2010, 130, 47-56.
Figure 1.
Illustration of the fire map in the world. Each dot (not to scale) represents a crop-stubble fire detected in a single day by a NASA (National Aeronautics and Space Administration) infrared imaging satellite (Fire Information for Resource Management System, FIRMS: https://firms.modaps.eosdis.nasa.gov/map/#d:24hrs;@0.0,0.0,3z, accessed on January 12, 2023).
Figure 1.
Illustration of the fire map in the world. Each dot (not to scale) represents a crop-stubble fire detected in a single day by a NASA (National Aeronautics and Space Administration) infrared imaging satellite (Fire Information for Resource Management System, FIRMS: https://firms.modaps.eosdis.nasa.gov/map/#d:24hrs;@0.0,0.0,3z, accessed on January 12, 2023).

Figure 2.
Sources of greenhouse gases from agriculture and the global share. (A) Cumulative greenhouse gases (GHGs) deriving from agriculture, land use and forestry globally (all GHGs considered), and (B) major GHGs emissions from agriculture by their importance. This illustration was created with BioRender.com.
Figure 2.
Sources of greenhouse gases from agriculture and the global share. (A) Cumulative greenhouse gases (GHGs) deriving from agriculture, land use and forestry globally (all GHGs considered), and (B) major GHGs emissions from agriculture by their importance. This illustration was created with BioRender.com.
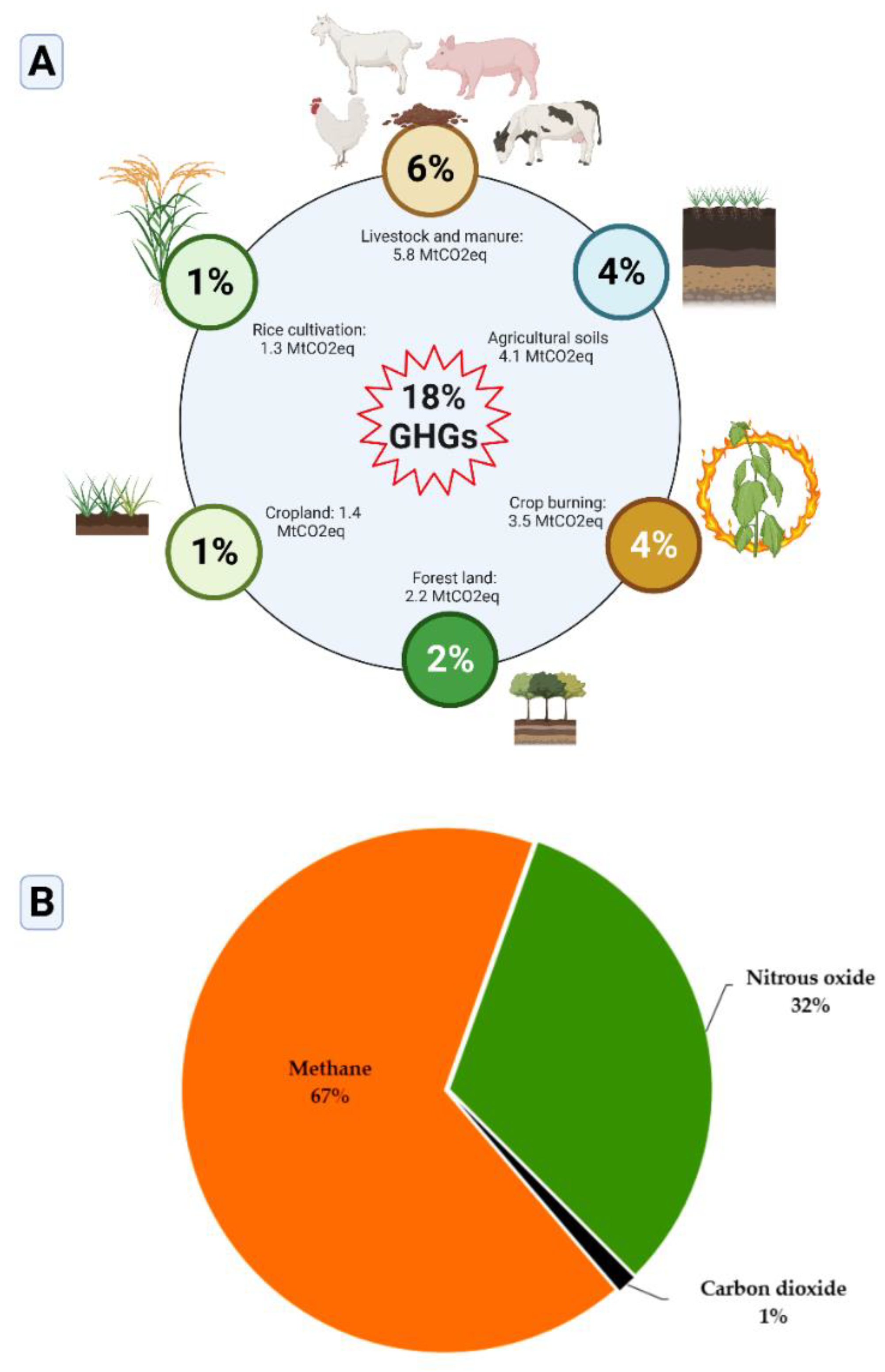
Figure 4.
Illustration of greenhouse gas production and emissions from livestock (ruminant animals).
Figure 4.
Illustration of greenhouse gas production and emissions from livestock (ruminant animals).
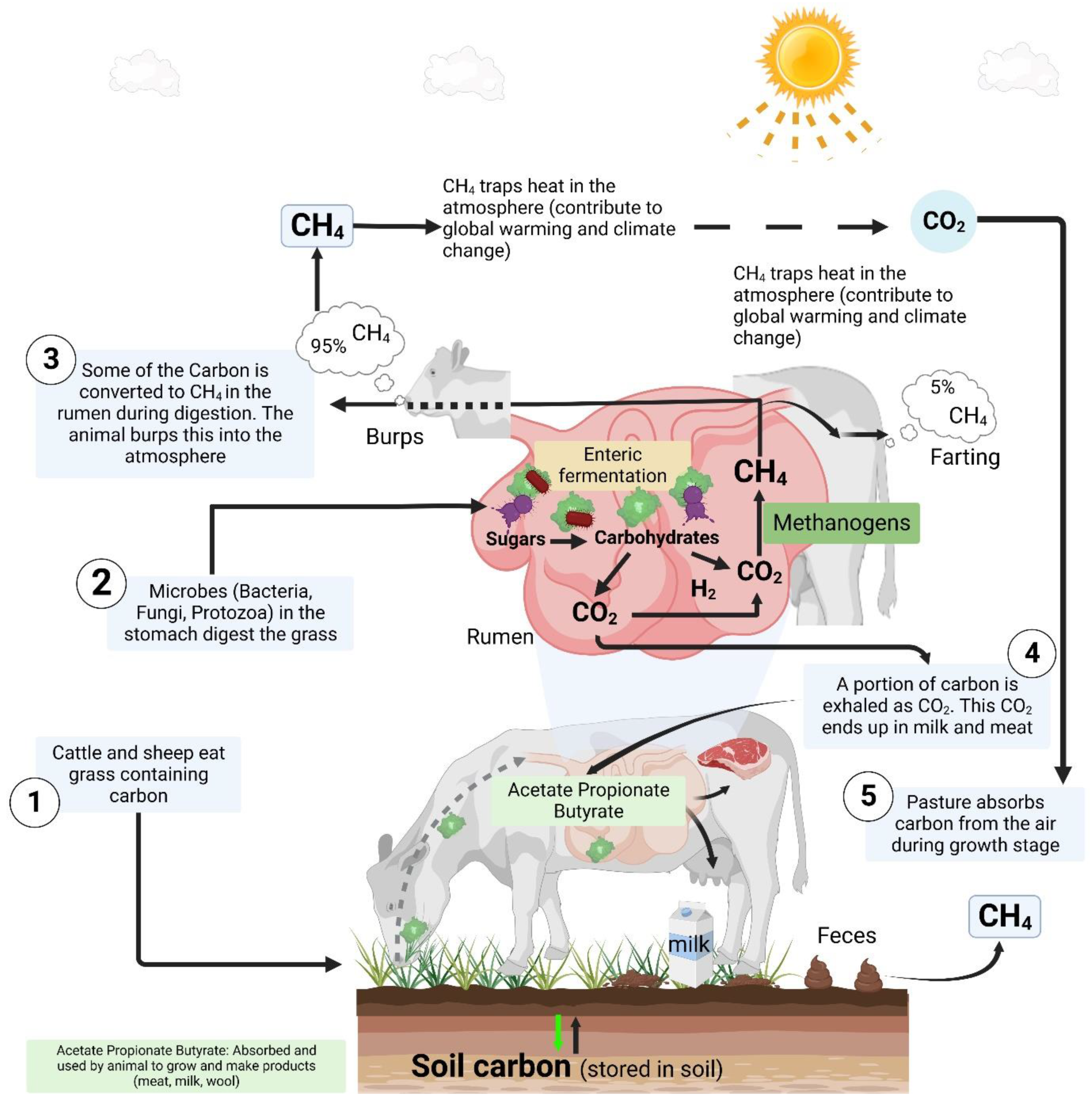
Figure 5.
Schematic representation of nitrogen use efficiency in plants. This model was created using biorender design platform (https://app.biorender.com/, accessed on 2023.05.25).
Figure 5.
Schematic representation of nitrogen use efficiency in plants. This model was created using biorender design platform (https://app.biorender.com/, accessed on 2023.05.25).
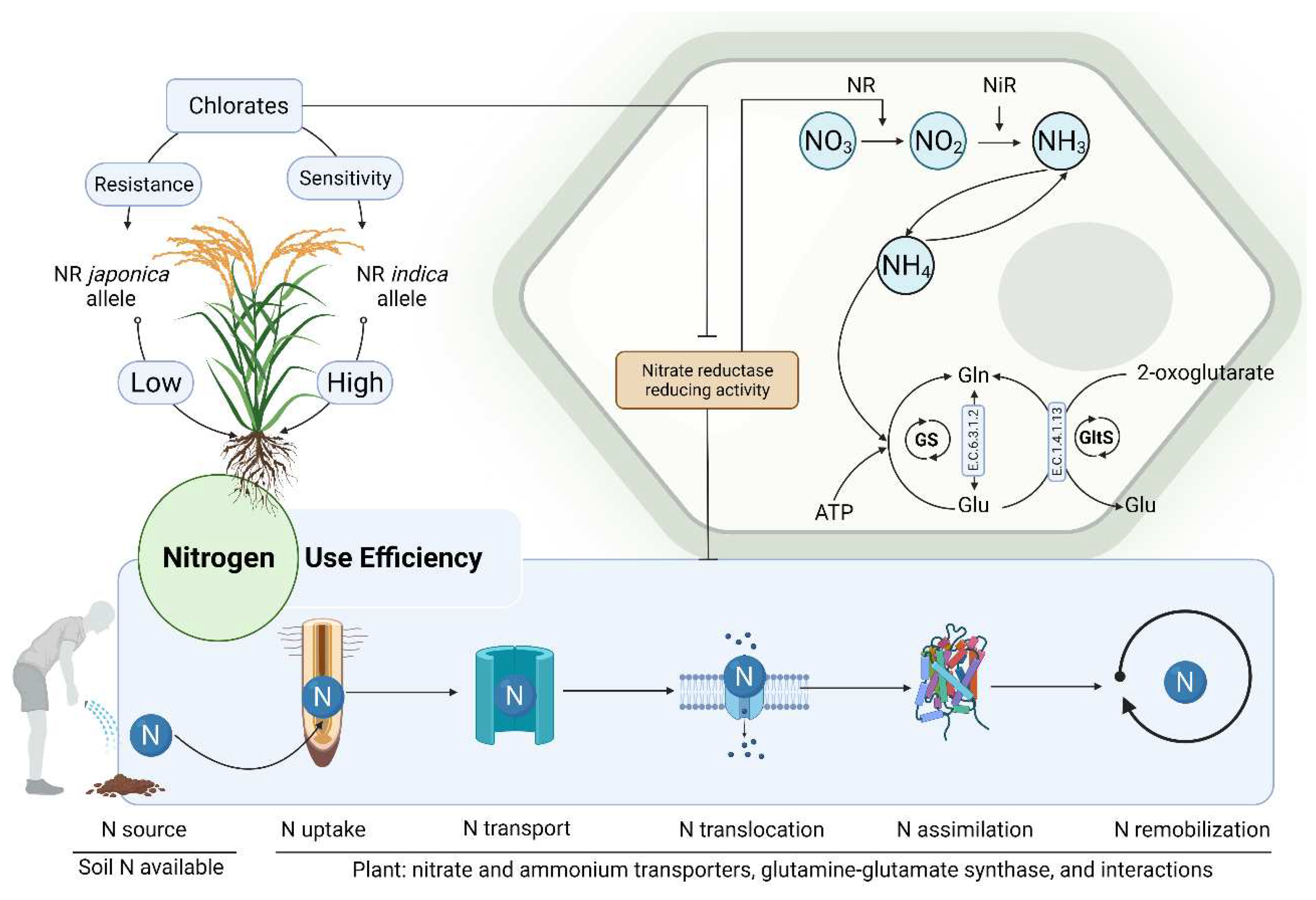
Figure 6.
Illustration of different methods and tools employed to understand N metabolism and improve NUE in plants. Highlighted circular zone in different segments shows various methods, technologies and tools employed to enhance the understanding of N metabolism in order to improve NUE. Continuous lines with an arrow connected to the origin denotes the possible outcomes, but limited to, of an improved NUE for plants and the environment. This model was created using biorender design platform (https://app.biorender.com/, accessed on 2023.06.11).
Figure 6.
Illustration of different methods and tools employed to understand N metabolism and improve NUE in plants. Highlighted circular zone in different segments shows various methods, technologies and tools employed to enhance the understanding of N metabolism in order to improve NUE. Continuous lines with an arrow connected to the origin denotes the possible outcomes, but limited to, of an improved NUE for plants and the environment. This model was created using biorender design platform (https://app.biorender.com/, accessed on 2023.06.11).
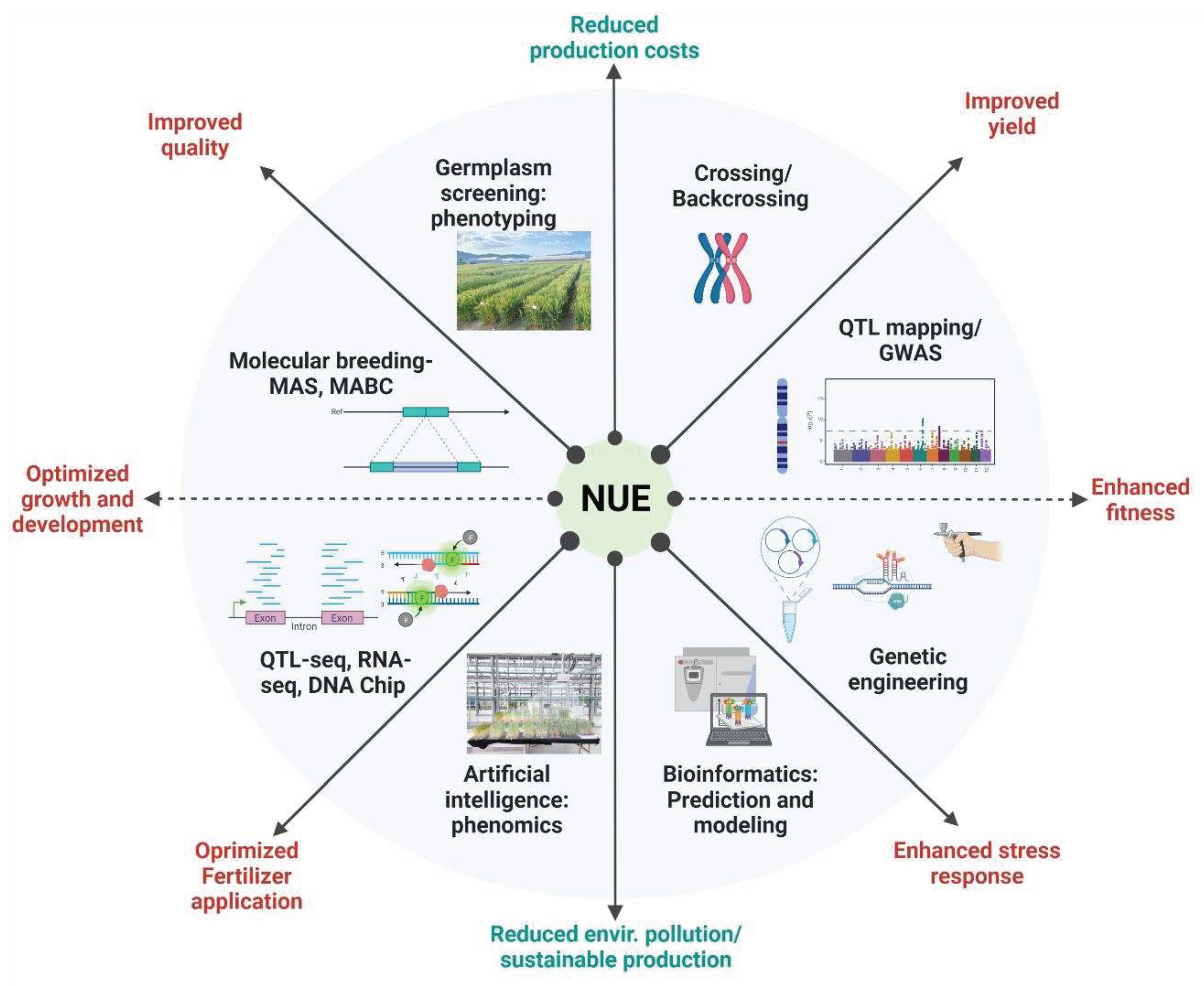
Figure 7.
Illustration of ROL in plants and mitigation of GHG emissions. Internal transport of gases is crucial for vascular plants in wetland or flooded prone environments. The direct exchange of gases between submerged tissues and the environment is impeded due to the slow diffusivity of gases in water. Soil aeration can fluctuate and zones of low O2 are widely spread in soil. In many wetland plants, aerenchyma is well developed even in drained conditions, and further enhanced in waterlogged conditions. Aerenchyma formation increases porosity above level due to the usual intercellular spaces. The O2 released to the rhizosphere of roots and the immediate environment (ROL) exerts differential effects on soil microbial activity. ROL inhibits the metabolism of anaerobic microorganisms such as methanogens (Archaea). Consequently, ROL abundance reduces CH4 production. In contrast, ROL promotes the metabolism of obligate aerobic organisms (methanotrophs or methane-oxidizing bacteria), allowing them to oxidize CH4 as their sole source of energy. Aerenchyma provides a low-resistance internal pathway for gas transport between shoot and roots extremities, and this pathway supplies O2 to the roots and rhizosphere. About 90% of CH4 emitted by crop cultivation is conveyed by plants during gas exchange events and long-distance transport. Whereas, nearly 10% is released through ebullition or diffusion from water or soil surface. This model was created using the biorender design platform (https://app.biorender.com/, accessed on 2023.05.25).
Figure 7.
Illustration of ROL in plants and mitigation of GHG emissions. Internal transport of gases is crucial for vascular plants in wetland or flooded prone environments. The direct exchange of gases between submerged tissues and the environment is impeded due to the slow diffusivity of gases in water. Soil aeration can fluctuate and zones of low O2 are widely spread in soil. In many wetland plants, aerenchyma is well developed even in drained conditions, and further enhanced in waterlogged conditions. Aerenchyma formation increases porosity above level due to the usual intercellular spaces. The O2 released to the rhizosphere of roots and the immediate environment (ROL) exerts differential effects on soil microbial activity. ROL inhibits the metabolism of anaerobic microorganisms such as methanogens (Archaea). Consequently, ROL abundance reduces CH4 production. In contrast, ROL promotes the metabolism of obligate aerobic organisms (methanotrophs or methane-oxidizing bacteria), allowing them to oxidize CH4 as their sole source of energy. Aerenchyma provides a low-resistance internal pathway for gas transport between shoot and roots extremities, and this pathway supplies O2 to the roots and rhizosphere. About 90% of CH4 emitted by crop cultivation is conveyed by plants during gas exchange events and long-distance transport. Whereas, nearly 10% is released through ebullition or diffusion from water or soil surface. This model was created using the biorender design platform (https://app.biorender.com/, accessed on 2023.05.25).
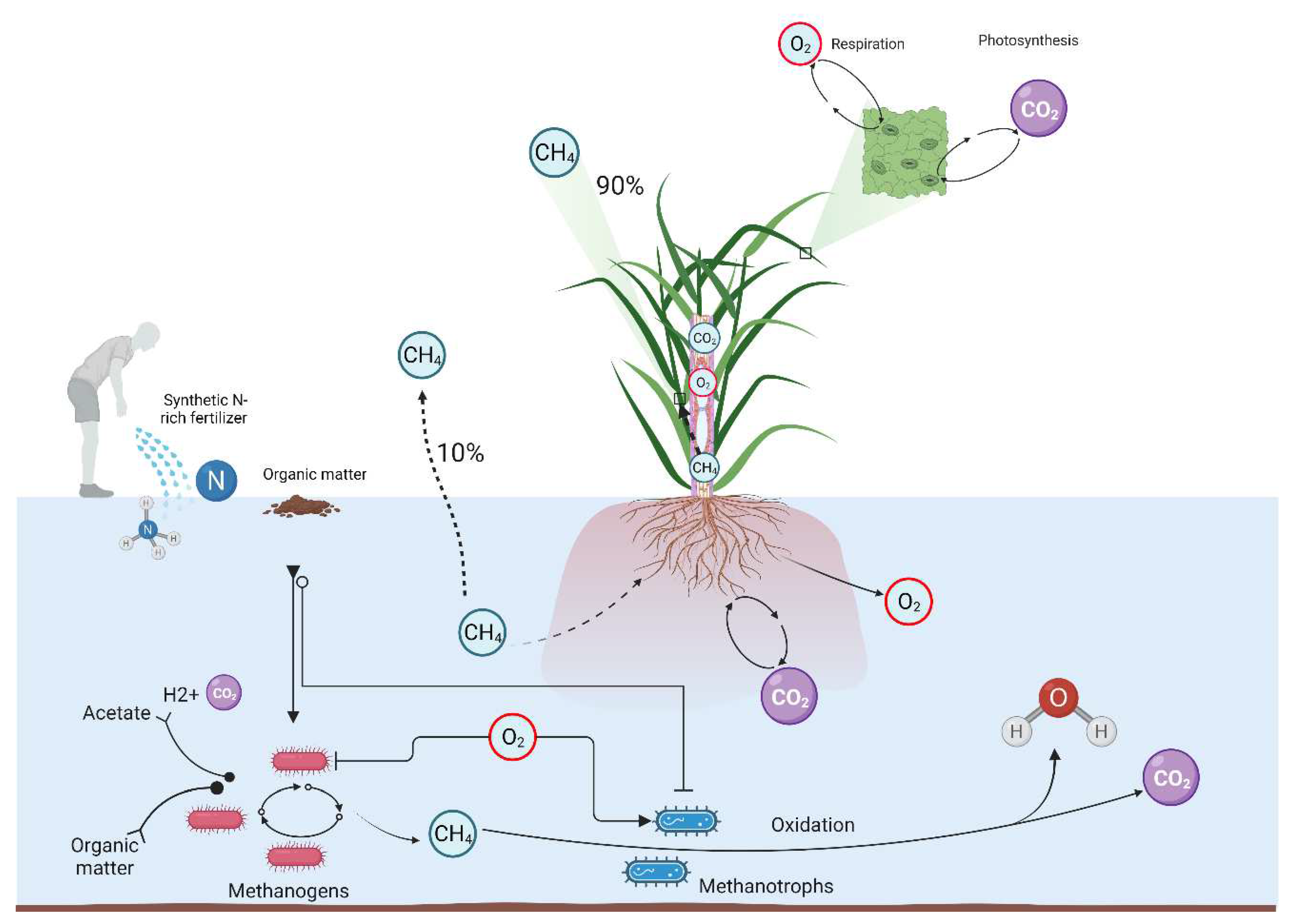

Table 1.
Summary key approaches for GHG reduction in agriculture.
| Advantages | Prerequisite | Application | Outcomes |
|---|---|---|---|
| Nitrogen use efficiency | |||
|
|
|
|
| Radial oxygen loss (ROL) | |||
|
|
|
|
| Application of biochar | |||
|
|
|
|
| Best Agricultural Practices | |||
|
|
|
|
| Improving ruminant feeding efficiency and enhancing target-specific ruminal bacterial activity | |||
|
|
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



