
Submitted:
05 September 2023
Posted:
07 September 2023
You are already at the latest version
Abstract
Paleogenomics focuses on the recovery, manipulation, and analysis of ancient DNA (aDNA) from historical or long-dead organisms to reconstruct and analyze their genomes. The aDNA is commonly obtained from remains found in paleontological and archaeological sites, conserved in museums and in other archival collections. Herbarium collections represent a great source of phenotypic and genotypic information, and their exploitation allowed to infer and to clarify previously unsolved taxonomic and systematic relationships. Moreover, Herbarium specimens offered a new source to study phenological traits in plants and to disentangle biogeography and evolutionary scenarios of species. More recently, advances in the molecular technologies went in parallel with the decreasing costs of the next-generation sequencing (NGS) approaches, that paved the way to the utilization of aDNA for whole-genome studies. Although many studies have been carried out combining modern analytic techniques and Herbarium specimens, this research field is still relatively unexplored, due to the need of improving strategies for aDNA manipulation and exploitation. The higher susceptibility of aDNA to degradation and contamination during the Herbarium conservation and manipulation, and the occurrence of biochemical post-mortem damages, can result in the more challenging reconstruction of the original DNA sequence. Here, we review the methodological approaches that have been developed for the exploitation of historical Herbarium plant materials, such as the best practices for aDNA extraction, amplification and genotyping. We also focus on some strategies to overcome the main problems related to the utilization of Herbarium specimens for their exploitation in plant evolutionary studies.
Keywords:
plant genetic resources
; population genomics
; crop evolution
; food legumes
1. Introduction
1.1. History and state of the art of herbaria
Herbarium collections represent large and not entirely explored deposits of genetic and phenotypic information (Bebber et al., 2010) that are able to provide snapshots of the diversity that was present in the past. Initially, Herbaria consisted of figurative books describing the characteristics of medicinal plants (Zunic et al., 2017). During time, Herbaria evolved to contain preserved plants, seeds, fungi, and algae, collected at different phenological stages (Willis et al., 2017). Herbarium specimens are commonly preserved dried on paper sheets or stored in folded packets or small boxes, which can be treated with solvents or pesticides to protect the specimens. However, it is also possible to find plant material conserved in liquid solutions, that may contain acids, aldehydes, alcohols, or other organic solvents for specimen preservation (Taylor & Swan, 1994). More recently, Herbarium samples are usually provided with supplementary label notes, often including details about the collection site and observations made by the collector, as shown in Figure 1c (Heberling, 2022). The history of modern herbaria began in the 16th century, when Luca Ghini (1490–1556), a professor of medical botany at the University of Bologna and Pisa, developed a method to make plant specimens transportable and preservable over time (von Engelhardt, 2011, Bellorini C., 2016). However, his legacy mainly consists of books, manuscripts, and letters, since the dried plants collected by Ghini have been largely lost after his death. Ghini’s contribution to the field is highly recognized also thanks to the herbaria established by his disciples. Some notable examples include the oldest surviving herbarium, the “Erbario Cibo”, preserved at the Pontifical University Library in Rome, and the herbarium of Ulisse Aldrovandi (1522–1605) that is conserved in Bologna (Figure 1b) (Cristofolini & Mossetti, 1993; Nepi Chiara, 2007; Signorini, 1993; Stefanaki et al., 2018). In the 18th century, Carl Linnaeus developed his Herbarium “cabinet” collection, including approximately 14,000 sheets of plant specimens and several/some zoological specimens (Stearn, 1961). In addition to his herbaria, Linnaeus provided innovative instruction on how to establish an herbarium collection, including proper techniques for collecting, drying, pressing, and mounting plants onto paper. Linnaeus also emphasized the relevance of collecting closely related specimens and using standardized collecting procedures to facilitate the exchange of materials and information among botanists. His efforts laid the foundations for the establishment of modern herbarium collections, which are still essential resources for botanical research and education (Müller-Wille, 2006). In 1935, the International Association for Plant Taxonomy (IAPT) established the “Index Herbarium” (http://sweetgum.nybg.org/science/ih/), a comprehensive resource that serves as a global directory of herbaria from different independent contributors. Nowadays, it includes almost 400 million specimens provided by more than 3,500 active herbaria worldwide (Thiers M., 2021). Among them, the largest number of specimens are preserved at the “Muséum National d’Histoire Naturelle” (France), at the “New York Botanical Garden” (USA), at the “Komarov Botanical Institute” (Russia), and at the “Royal Botanic Gardens” (UK) (Thries, 2021). Until mid-20th Century, researchers had two options to examine herbarium collections: either by physically visiting the herbaria or by requesting specimens on loan for study. Towards the end of 1,900, significant advances took place with massive process of digitalization of herbarium collections (Thiers et al., 2016), which resulted in the development of virtual herbaria. Thus, Herbarium specimens were available as high-resolution images linked to electronically associated notes (Heberling, 2022; Nelson et al., 2012), that are accessible through online platforms. Such databases improved the exploitation and the exchange of plant specimens, facilitating collaborative research and conservation efforts, and providing a powerful source to investigate plant diversity and evolution dynamics.
1.2. Herbarium genomics
In recent decades, paleogenomics has greatly benefited from the significant advances in the field of molecular biology (Pont et al., 2019), including those in DNA extraction and amplification procedures, and the improvements in Next-Generation Sequencing (NGS) strategies. This has enabled researchers to obtain molecular markers, as well as the whole-genome sequences (WGS) from specimens dating back several centuries, including those preserved in herbaria. Paleogenomics studies were also applied to the analysis of pathogens preserved within these specimens, which allowed detailed explorations of temporal signals of divergence among modern and ancient strains (Saville et al., 2016; Yoshida et al., 2013). As stated above, the characterization and digitization processes of collections have greatly promoted the accessibility and utilization of ancient samples in a wide range of studies (Thiers et al., 2016). Traditionally, herbarium specimens have been employed for taxonomy and systematics studies. These collections still represent a significant resource for both the discovery and formal description of plant taxa. Moreover, molecular approaches have revolutionized the study of taxonomy, bringing significant advancements in resolving ambiguous phylogenetic relationships and classifications (Rodríguez 2019). With this regard, the possibility to extract and purify aDNA from dried historical specimens can provide essential support in such studies (Rogers & Bendich 1985). By exploiting aDNA from now-extinct species or ancient samples, researchers can gain valuable insights into the evolutionary history of different organism and species, and bridge gaps in our understanding of past biodiversity and present genetic diversity, also allowing for the reconstruction of phylogenetic trees (Rodríguez 2019). Moreover, taking into consideration that the dissemination of species in novel geographic areas is associated with molecular adaptative mechanisms, as well as significant phenotypic and phenological changes (Bellucci et al., 2022; Cortinovis et al., 2020), herbaria can be exploited to dissect plant response to the change of environmental conditions, across time (Damerval et al., 2018; Davis et al., 2015). Indeed, recruiting phenotypic and genotypic information from historical materials, collected in different geographical areas is crucial to reconstruct the evolutionary dynamics and adaptation processes of a species. For example, Myers et al. (2022) analyzed phenotypic data acquired from illustrations of common bean, from the XVI century, to provide historical support to the dynamics of the introduction of the two main gene pools of this species (i.e., Mesoamerican and Andean) from the Americas to Europe, following the Columbus travels. The application of molecular tools to ancient biological samples provides the opportunity to investigate population dynamics across different time points, as a result of migration processes, gene flow and occurrence of novel mutations, with changes at the genome level driven also by genetic drift and by natural selection behind adaptation. That is the possibility to investigate the complex interplay between genetics, environment and evolution of a population across time (Bakker, 2018; Gutaker & Burbano, 2017). In genomic studies, the comparison of data from herbaria and modern samples can provide valuable information to optimize the management of plant genetic resources (Rosche et al., 2022). Cozzolino et al. (2007) evaluated the changes in the genetic diversity of Anacamptis palustris population because of habitat transformation in Italy over time. To do so, authors sequenced and compared a plastid region between Herbarium specimens, collected prior to the Second World War, and modern samples. Thus, this approach could be very useful as a complement to large genotyping and phenotyping programs of modern accessions aimed at understanding the environmental adaptation and the evolutionary dynamics of genetic diversity as the INCRESAE project (Bellucci et al., 2021), which is focused on food legumes as chickpeas (Rocchetti et al., 2022) common bean (Cortinovis et al., 2022), lupin (Kroc et al.,2021) and lentil (Guerra-García et al.,2021). However, from the work carried out by Cozzolino et al. (2007), some possible limitations of herbarium collections have arisen, such as limited sample size and potential methodological biases in collecting procedures. For instance, only few individuals could have been collected from each collection site, which may not capture the full extent of genetic diversity within a population. Despite these limitations, herbaria can be considered a valuable source for detecting variations in the distribution and frequency of haplotypes and alleles.
1.3. Challeneges in the use of herbaria and ancient samples for genomics studies
The advent of NGS platforms have opened unprecedented possibilities for studying genome level diversity from Herbaria and ancient samples. However, for this purpose, the successful implementation of NGS may be challenging due to the unique nature and the issues associated to the manipulation of ancient samples and aDNA. The development of robust protocols for aDNA purification, amplification, and sequence data analysis is crucial for obtaining reliable results (Pont et al., 2019). The aDNA from Herbaria and ancient samples is often degraded and fragmented (Paabo et al., 2004), thus, the optimization of extraction protocols can represent a crucial bottleneck to obtain high-quality aDNA suitable for sequencing. Additionally, the risk of contamination from modern DNA, while handling and processing is also a main concern (Cooper et al., 2000). Moreover, aDNA in Herbarium-preserved samples can be susceptible to biochemical post-mortem damage (Willerslev & Cooper, 2005). Indeed, over time, the aDNA molecules can undergo various forms of degradation, including fragmentation and chemical modifications; these damages pose additional challenges when the original DNA sequence must be assembled and in the analysis of sequences. The biochemical post-mortem damages in Herbarium samples can result from different factors, such as light exposure, not controlled moisture, temperature fluctuations, and the presence of reactive chemicals. These factors can lead to DNA strand breaks, base modifications, and cross-linking of DNA molecules, which can affect the quality and integrity of the assembled sequence (Weiß et al., 2016). Efficient protocols have been developed to overcome these issues, especially regarding aDNA purification. Here, we report the most relevant protocols for aDNA purification from Herbaria specimens (Table 1). Furthermore, the choice of the bioinformatics pipeline and of the strategy for the analysis of NGS data from Herbaria and ancient samples is particularly relevant, especially in the case of low-quality and fragmented aDNA. In the present review, we want to give an insight about the best strategies for the extraction, purification, amplification and sequencing of Herbarium aDNA, which can be exploited in genetic diversity and evolutionary studies of plant species as well as in the reconstruction of phylogenetic and taxonomical relationships.
2. Extraction and purification of aDNA from Herbarium specimens
The utilization of herbarium specimens in molecular studies poses significant challenges due to the difficulties in obtaining a substantial amount of high-quality purified aDNA (Wang, 2018). Working with herbarium specimens involves the handling of a limited amount of tissues, that are often stored under suboptimal conditions for long periods, leading to aDNA damage and an increased risk of contamination (Drabkova et al., 2002). Doyle and Dickson (1987) raised the importance of testing different methods to preserve aDNA integrity within herbarium specimens and several subsequent studies have consistently found that air-drying of plant material is a reliable preservation method, preventing degradation and better preserving aDNA, when compared to alternative practices, such as microwaving, boiling, or immersion in chemical solutions (Staats et al., 2011). Moreover, it has been observed that well-preserved older materials tend to yield higher quantities of purified aDNA of better quality when compared to younger materials that have been subjected to unfavorable storage conditions (Doyle & Dickson, 1987; Drabkova et al., 2002; Särkinen et al., 2012). As a general consideration, we also have to take into account that the initial conditions of the tissues at the sampling stage, before storage, inevitably impacts the quality and the quantity of the purified aDNA (Jankowiak et al., 2005). Overall, the purification from herbarium specimens results in a lower yield of DNA, when compared to the utilization of fresh plant tissues (Savolainen et al., 1995; Taylor & Swan, 1994). In their pioneering work, Rogers and Bendich (1985) employed a modified CTAB (cetyltrimethylammonium bromide) nucleic acid extraction protocol to purify DNA from a small number of mummified seeds, herbarium specimens, and fresh plant tissues. Although this extraction method confirmed a higher yield when applied to fresh tissue, it has also proven to be effective for the extraction and purification of aDNA from ancient plant materials. A relevant step during aDNA purification is the grinding of the plant material obtained from the herbarium specimen. One of the most adopted practices rely on bead-based homogenization equipment, such as a mixer mill or beads mill. This approach minimizes aDNA loss and reduces the risk of contamination that may arise when using alternative methods, like griding in mortars with liquid nitrogen (Drábková, 2014). Additionally, the use of a bead-based homogenization approach enables the simultaneous processing of multiple samples, ensuring consistent and homogenous results (Drabkova et al., 2002). Over the past two decades, numerous aDNA extraction protocols have been developed, tested, and compared to optimize the quality and quantity of purified aDNA from herbaria specimens, for which a report is provided in Table 1. Drabkova et al. (2002) tested seven DNA extraction protocols in herbarium specimens from Juncus and Luzula genera (Juncaceae) species collected during the twentieth century, including fresh tissue materials as a control. They evaluated quality parameters, such as the aDNA A260/280 and A230/280 absorbance ratios, and estimated the quantity of aDNA through spectrometry. From their results, they concluded that the use of a DNeasy Plant Kit (Qiagen), with some modifications, was the best approach for the aDNA purification. In detail, the authors found that both the optimal homogenization of the plant tissue and the extension of the precipitation time were crucial steps. The modifications to the manufacturer’s protocol regarded in particular: a longer cell lysis time (i.e., 30 minutes), an increased volume of AP1 buffer (i.e., 450 μl), a reduced volume of elution buffer (i.e., 50 μl) that increases aDNA concentration, and an extended elution time (i.e., 10 minutes). They also pointed out the protocols that gave the worst results, particularly, the phenol extraction, due to the potential contamination of aDNA with phenol, which can negatively affect downstream reactions, including amplification and sequencing, and, similarly, the CsCl gradients protocol, as aDNA could be lost within the gradient, due to the limited amount of tissue (Drabkova et al., 2002).
One of the major challenges when dealing with the DNA extraction from ancient material is the complete loss of the original sample, contradicting one of the fundamental principles of museum collections, i.e., preserving specimens in their original state (Freedman et al., 2018). For this reason, efficient DNA extraction protocols, that require a very limited amount of tissue, have been set up for historical specimens, with the aim of preserving the integrity of the Herbaria. Shepherd (2017) developed a non-disruptive approach to sample material directly from herbaria, by using a Staedtler “Mars Plastic” eraser, that minimize the damages to the specimens while obtaining the necessary amount of tissue for DNA extraction. Sugita et al. (2020), taking as example a previously published protocol for DNA extraction in arthropods, established a plant-suitable non-disruptive protocol, particularly useful in species with small (i.e., <25 mm2) and fragile leaves (Gilbert et al., 2007, Santos et al., 2018). Nevertheless, despite the importance of non-disruptive methods, CTAB and modified DNeasy Plant Mini Kit (QIAGEN) protocols are the most frequently adopted methods for aDNA extraction (Drábková, 2014). Among the reasons for such choice, aDNA-specific protocols are generally more expensive, more time-consuming, and require specific protocols and facilities to avoid contamination. As stated above, age, storage conditions of herbarium specimens (e.g., fluctuating temperatures or high humidity), and the species from which the tissue is obtained, can influence the efficiency of the aDNA extraction (Zeng et al., 2018), thus, such factors must be taken into account when choosing the aDNA extraction protocol. With this regard, Marinček et al. (2022) compared the standard Qiagen DNeasy Plant Mini Kit and a specific dithiothreitol (PTB)-dithiothreitol (DTT) protocol, previously developed by Gutaker et al. (2017), with the aim to evaluate the efficiency of these protocols on specimens from the genera Xanthium L. and Salix L.. They showed a lower efficiency of the Qiagen DNeasy Plant Mini Kit when working with older specimens (i.e., collected before 1900). Thus, a researcher might want to consider the PTB-DTT protocol as an alternative for aDNA extraction in older and more challenging specimens, for example due to the higher concentration of secondary metabolites in the tissue. Indeed, several protocols have been developed specifically taking into account the issues related to the presence of secondary metabolites that can inhibit PCR reactions. Ribeiro & Lovato (2007) tested five DNA extraction protocols on fresh and herbarium leaves of various species belonging to the genus Dalbergia, known to contain significant amounts of secondary metabolites, potentially interfering with DNA amplification. Based on their results, the most efficient protocol was developed by Jobes et al. (1995). This protocol uses three key reagents: polyvinylpyrrolidone (PVP), that binds phenolic compounds, sodium chloride with a high molar concentration, that inhibit the co-precipitation of polysaccharides and DNA and, in turn enhances the solubility of polysaccharides in ethanol, and, finally, lithium chloride, which is useful to selectively precipitate and subsequently remove RNA. Krinitsina et al. (2015) proposed a cost-effective DNA extraction method specifically designed for herbarium specimens, that involves the utilization of AMPure XP magnetic beads, diluted in a buffer containing PEG 8000. The authors suggest that the use of magnetic beads can reduce the concentration of PCR inhibitors in the aDNA. Shapiro et al. (2016) adapted a protocol that has been originally designed for isolating aDNA from human hair samples (Thomas et al., 2004) to ancient plant specimens. In particular, the protocol includes two key steps: a phenol-chloroform extraction followed by a silica spin column purification. The phenol-chloroform extraction has proven to be effective for those samples containing low amounts of DNA and that may contain polymerase chain reaction (PCR)-inhibitors, affecting the enzymatic activity of the Taq polymerase (Kumar Shasany & Darokar, 1999; Särkinen et al., 2012). Overall, selecting the most appropriate aDNA purification protocols involves careful consideration of factors such as specimen age, preservation conditions, and the specific goals of the research. By taking these factors into account, researchers can optimize their DNA purification methods and enhance the chances of obtaining high-quality aDNA for subsequent analysis (Figure 2).
3. DNA amplification by PCR from Herbarium specimens
Since its development, PCR has become a routinely and indispensable technique in several protocols for molecular biology studies, that can be extended to the analysis of ancient samples. Several studies reported about the successful amplification of aDNA from plant herbarium material collected over a wide range of ages, resulting in variable lengths of amplified fragments (Cota-Sánchez et al., 2006; Lister et al., 2008; Särkinen et al., 2012). Staats et al. (2011) also reported that there are no significant differences in the PCR amplification efficiency among plastid, mitochondrial, and nuclear aDNA from herbarium material, similarly to what can be generally observed in the amplification from plastid, mitochondrial and nuclear DNA obtained from fresh tissue. As mentioned, DNA extraction from herbarium material can often yield low quantities of fragmented genetic material. However, the quality and purity, rather than the quantity, of aDNA template, mostly affect PCR amplification efficiency (Särkinen et al., 2012). Quality control procedures, including rigorous purification steps, and measures for minimizing and identifying contaminations, are crucial to ensure the accuracy and reliability of the PCR amplification. As stated above, the presence of secondary metabolites in herbarium samples can have a negative impact on the amplification efficiency. To address this issue, various extraction protocols have been developed with the aim of reducing the concentration of these metabolites (Krinitsina et al., 2015; Riahi et al., 2010; Ribeiro & Lovato, 2007). In order to mitigate the effects of PCR inhibitors, as a post aDNA purification step, a potential solution is diluting DNA extracts. However, this approach is not always feasible for herbarium samples due to the typically low yield and concentration of the eluted aDNA, obtained from these specimens (Samarakoon et al., 2013). An alternative approach to improve the quality and quantity of the aDNA template available for PCR amplification, in presence of potential inhibitors, has been suggested by Samarakoon et al. (2013); it involves the use of a reagent called TBT-PAR, which contains trehalose, bovine serum albumin (BSA), and polysorbate-20 (Tween-20). Multiple studies investigated the effects of using high concentrations of BSA on PCR amplification efficiency, when working with poor-quality template aDNA. Overall, these studies agreed that a higher concentration of BSA can have a strong positive impact on PCR efficiency (Drábková, 2014; Ribeiro & Lovato, 2007). In particular, Särkinen et al. (2012) suggested the utilization of high concentration of BSA when amplifying aDNA from herbarium specimens, routinely. Several factors can indeed affect PCR amplification efficiency, and among these factors are the purity and the specific type of polymerase, the purity of buffers and reagents used. It is crucial to consider these factors to ensure reliable and consistent PCR results, especially to amplify aDNA (tell et al., 2008). A suitable and cost-effective alternative to mitigate the effects of potential PCR inhibitors is the adoption of a polymerase enzyme that is less susceptible to their presence (Monroe et al., 2013). Särkinen et al. (2012) suggests that the utilization of a polymerase without proofreading activity may perform better than one with proofreading activity. Moreover, it is essential to investigate and optimize the performance of different polymerases to improve the amplification efficiency for aDNA. Furthermore, Särkinen et al. (2012) reported a significant negative correlation between the length of the aDNA template fragments and PCR efficiency, when amplifying aDNA from herbarium specimens. Thus, this suggests that shorter fragments can be easily amplified, that is an interesting aspect considering the fragmented nature of the aDNA template usually obtained from these specimens. However, in the case of extreme aDNA fragmentation, amplification of certain loci may be challenging. It is worth noting that mitochondrial aDNA (mtDNA) can be successfully retrieved also from severely degraded samples, since it is present in high copy numbers within a cell, there is a higher chance that some mtDNA fragments are long enough for amplification, even when nuclear DNA is highly fragmented (Burrell et al., 2015). Another relevant aspect is the instability of nucleic acids during long-term preservation, that can lead to the formation of various post-mortem damages, affecting the quantity and quality of template aDNA for the amplification; such damages can also introduce potential errors that make challenging the reconstruction of the aDNA sequence. An example of post-mortem damage is depurination, which can result in the formation of single or double-strand breaks in the DNA molecule. Additionally, crosslinking reactions can occur, limiting or even preventing DNA amplification altogether (Binladen & Willerslev, 2010). Thus, it is crucial to identify the best methodologies and strategies to take into account the occurrence of post-mortem damages when working with herbarium samples, in order to mitigate their effects on the quality and amplification efficiency. Indeed, oxidative and hydrolytic modifications of bases can lead to the formation of miscoding lesions, such as the deamination of cytosine to uracil. Thus, these lesions can lead to the incorporation of incorrect bases during the amplification process (Staats et al., 2011). Certain oxidative damage can also create lesions that block the polymerase enzyme and promote the generation of chimeric sequences through ‘jumping PCR’ (Paabo et al., 1990). To address the PCR artifact caused by misincorporation, Uracil-N-glycosylase (UNG) can be employed prior to amplification to remove deaminated cytosines (Willerslev & Cooper, 2005). UNG creates an abasic site that is subsequently hydrolyzed through β-elimination, resulting in a strand break (Hofreiter et al., 2001). However, it is important to carefully consider the use of UNG treatments since it inevitably leads to a reduction in the aDNA sequence length, that is often highly fragmented (Pruvost et al., 2005). In addition to the sensitivity of PCR amplification, the low template aDNA concentration, and the fragmented nature of potential target loci in the aDNA, the presence of exogenous DNA contamination is also a common challenge in aDNA manipulation. Indeed, exogenous DNA might have been introduced during the collection and conservation of the plant material or by the presence of microorganisms or pathogens (Bieker et al., 2020), or by handling samples during molecular processes. Cooper and Poinar (2000) emphasized the importance of manipulating herbarium specimens in a physically isolated laboratory during all the steps. Nevertheless, DNA contaminants can still be introduced into the experimental workflow through various means, as summarized in Figure 3, including the use of contaminated reagents or samples, as well as by the presence of residual DNA and PCR amplification products from previous experiments (Champlot et al., 2010).
To minimize the risk of contamination, Knapp et al. (2012) developed a set of guidelines for establishing a molecular laboratory exclusively dedicated to studies on ancient specimens. The guidelines recommend to conduct the different steps of aDNA extraction and amplification in dedicated rooms and/or hoods, expanding the concept of spatial isolation within the same laboratory. Furthermore, the authors highlight the importance of implementing a limited access policy, granting facility access only to qualified personnel who are well aware of the risks of contamination.
4. Next-generation sequencing (NGS) and genotyping on Herbarium specimens to disentangle relevant aspects of the evolutionary history of a species
Several studies investigated the genetic diversity preserved in herbarium collections, providing insights into relevant aspects of plant species evolutionary histories, including adaptation processes and the effects of environmental changes over time. Several approaches have been employed to genotype herbarium specimens (Beck and Semple 2015, Hart et al. 2016, Rowe et al. 2011). Malenica et al. (2011) successfully genotyped a 90-year-old Tribidrag grapevine herbarium specimen by utilizing a set of nine microsatellite markers (SSRs) and a whole genome amplification (WGA) procedure. Other amplification-based genotyping approaches have proven to be suitable for molecular analysis of herbarium samples, despite of the limitations in the aDNA amplification from such materials, as described above (Burrell et al., 2015; Cota-Sánchez et al., 2006). As a way of example, while AFLP fingerprinting can be utilized to analyze herbarium material, the fragmented nature of the purified aDNA can introduce a bias when utilizing such approaches, since AFLP is based on the presence and distribution of restriction sites. In such cases, Lambertini et al. (2008) suggested to use only those markers that can be detected also in fresh tissue, and, in parallel, to increase the number of primer combinations to ensure the amplification of an adequate number of polymorphic fragments. By studying chloroplast and nuclear microsatellites diversity on 57 historical herbarium specimens, Roullier et al. (2013) were able to trace relevant events in the evolutionary history of sweet potato over time, such as effect of the domestication process, migration patterns, and genetic interactions. By comparing the genetic data from historical herbarium collections with that of modern samples, researchers can assess changes in the level of genetic diversity and identify potential genetic bottlenecks or shifts that occurred during domestication and cultivation. The emergence of Next-Generation Sequencing (NGS) led to a revolutionary change in our ability to obtain multi-locus genetic data from natural historical collections (Burrell et al., 2015). An approach that has significantly impacted this field of research is the Sequencing by Synthesis (SBS) strategy, which has been developed to enable cost-effective shotgun sequencing of whole genomes (Liu et al., 2012). High-throughput SBS technologies involves the preparation of sequencing libraries, which entails attaching artificial DNA segments, known as adapters, to both ends of those template fragments characterized by a specific range of fragment (Hart et al. 2016). This strategy allows for efficient and accurate sequencing of the aDNA fragments. When working with aDNA, differently from the SBS procedure for library preparation in modern samples, the fragmentation step that comes before adapter ligation might be skipped (Hart et al., 2016). The adapters used in sequencing library preparation can serve multiple purposes. Indeed, they enable the priming of both whole-genome shotgun sequencing or the enrichment of specific genomic regions of interest through hybridization capture techniques (Hart et al. 2016). These approaches allow for the characterization of various types of DNA, such as organellar DNA or nuclear loci, as well as the detection of a vast number of single nucleotide polymorphisms (SNPs) distributed throughout the nuclear genome. Generally, library preparation protocols for herbarium DNA templates do not require significant modifications compared to standard ones. Most of these protocols were originally developed for Illumina sequencing-based platforms and can be broadly categorized into two main types: the single-stranded library (ss-library), and the double-stranded library (ds-library) construction methods. Briefly, ss-library preparation starts with heat denaturation of DNA, followed by the attachment of a biotinylated adapter oligonucleotide to the 3′ ends. A second adapter is then blunt-end ligated to complete the library preparation, which is further amplified through PCR (Gansauge & Meyer, 2013). Whereas, ds-library preparation can be further categorized into Blunt-End and Y-adapter methods, both involving the end-repairing of the DNA fragments and ligation of double-strand adapters, but differing in the type of adapter used. Bennett et al. (2014) tested all the different methodologies for library preparation, starting from aDNA faunal and Human remains, founding that the Y-adapter method led to the formation of adapter-dimer artifacts, while the ss-library approach allowed for an increased proportion of shorter endogenous molecules incorporated into the libraries. Their outcome has been further confirmed in recent researches, suggesting that ss-library preparation approach better suits to the aDNA features compared to double-stranded library protocols (Orlando et al., 2021), exhibiting greater sensitivity to degraded and poorly preserved ancient samples, such as herbarium specimens (Psonis et al., 2021). In protocols that require the amplification of libraries by PCR, the presence of artifacts caused by misincorporation can impact sequencing accuracy and efficiency. To mitigate the effects of post-mortem damage, UDG enzyme-based protocols (see previous section) can be employed during library preparation (Briggs & Heyn, 2012). One of the key advantages in using UDG is that it helps eliminating misincorporations (C to T and G to A) in the recovered sequences, improving the mapping efficiency. This is particularly useful when the reference sequence belongs to a species that is distantly related to the query sequence, by reducing the number of possible gaps and mismatches in the alignment (Psonis et al., 2021). However, the presence of uracil and other types of post-mortem damage can also serve as an indicator to assess the presence of modern contamination in aDNA samples (Prüfer et al., 2010). Damage profiles can be inferred using analytical tools such as mapDamage 2.0, which allows to distinguish ancient molecules from modern contaminants (Ginolhac et al., 2011; Jónsson et al., 2013). Another strategy to address errors in sequence reconstruction, resulting from misincorporations in aDNA, is to focus downstream analysis on transversions rather than transitions, since the latter are more susceptible to post-mortem damage. Trucchi et al. (2021) applied this strategy in their paleontological study on ancient bean seeds from different archaeological sites in Argentina. They analyzed the temporal dynamics of genetic variation and selection during the domestication process of the common bean (Phaseolus vulgaris L.) in the southern Andes, by comparing WGS data from a panel of 15 ancient beans dating between 2,500 and 600 years ago and modern wild and domesticated common bean accessions (from both Mesoamerican and Andean gene pools). The work of Trucchi et al. (2021) clearly demonstrated how by considering transversions, researchers can significantly reduce the impact of post-mortem damage on sequence analysis, and improve the accuracy of genetic variation inference in aDNA studies.
5. Conclusion
The employment of herbaria in paleogenomics can offer the opportunity of investigating genetic and phenotypic diversities of ancient samples compared to modern ones, being available plant genetic resources that are largely unexplored. This requires optimized strategies for herbaria handling and for the analysis of the aDNA, including the employment of advanced next-generation sequencing approaches, to access an unexplored source of genetic information. However, the quality of attainable genetic data can be strongly affected by the protocol used for aDNA extraction, amplification, and sequencing, due to several factors, such as preservation state of the specimens and of the aDNA. Consequently, an in-depth understanding of the strengths and limitations of available protocols for aDNA exploitation become crucial. Here, we reviewed the most recommended strategies and approaches for handling herbaria specimens and aDNA. Moreover, we discussed the relevance of integrating available high-quality ancient sequence data from Herbarium specimens with modern sequences, to perform population genetics and genomics studies and inferences on the evolution of a species.
Author Contributions
S.P., V.D.V., A.P. and R.P. conceived the structure, wrote and contributed to the drafting of the manuscript. All the authors contributed to the editing and approved the final version of the manuscript.
Funding
This work was supported by the INCREASE project (European Union’s Horizon 2020 research and innovation programme under grant agreement No. 862862) and by the Italian Government (MIUR) Grant n. 20177RL4KL, Project Progetti di Ricerca di Interesse Nazionale (PRIN PARDOM) 2017.
References
- Bakker: F. T. (2018). Herbarium Genomics: Plant Archival DNA Explored (pp. 205–224). https://doi.org/10.1007/13836_2018_40. [CrossRef]
- Barrett, C. F., Huebner, C. D., Bender, Z. A., Budinsky, T. A., Corbett, C. W., Latvis, M., McKain, M. R., Motley, M., Skibicki, S. v., Thixton, H. L., Santee, M. v., & Cumberledge, A. N. (2022). Digitized collections elucidate invasion history and patterns of awn polymorphism in Microstegium vimineum. American Journal of Botany, 109(5), 689–705. https://doi.org/10.1002/ajb2.1852. [CrossRef]
- Bebber, D. P., Carine, M. A., Wood, J. R. I., Wortley, A. H., Harris, D. J., Prance, G. T., Davidse, G., Paige, J., Pennington, T. D., Robson, N. K. B., & Scotland, R. W. (2010). Herbaria are a major frontier for species discovery. Proceedings of the National Academy of Sciences of the United States of America, 107(51), 22169–22171. https://doi.org/10.1073/pnas.1011841108. [CrossRef]
- Beck, J. B., & Semple, J. C. (2015). Next-Generation Sampling: Pairing Genomics with Herbarium Specimens Provides Species-Level Signal in Solidago (Asteraceae). Applications in Plant Sciences, 3(6), 1500014. https://doi.org/10.3732/apps.1500014. [CrossRef]
- Bellorini C. (2016). The World of PlanTs in renaissance Tuscany.
- Bellucci, E., Benazzo, A., Xu, C., Bitocchi, E., Rodriguez, M., Alseekh, S., di Vittori, V., Gioia, T., Neumann, K., Cortinovis, G., Frascarelli, G., Murube, E., Trucchi, E., Nanni, L., Ariani, A., Logozzo, G., Shin, J. H., Liu, C., Jiang, L., … Papa, R. (2022). Selection and adaptive introgression guided the complex evolutionary history of the European common bean. BioRxiv 2022.09.28.509856. https://doi.org/10.1101/2022.09.28.509856. [CrossRef]
- Bennett, E. A., Massilani, D., Lizzo, G., Daligault, J., Geigl, E. M., & Grange, T. (2014). Library construction for ancient genomics: Single strand or double strand? BioTechniques, 56(6), 289–300. https://doi.org/10.2144/000114176. [CrossRef]
- Bi, K., Linderoth, T., Vanderpool, D., Good, J. M., Nielsen, R., & Moritz, C. (2013). Unlocking the vault: Next-generation museum population genomics. Molecular Ecology, 22(24), 6018–6032. https://doi.org/10.1111/mec.12516. [CrossRef]
- Binladen, J., & Willerslev, E. (2010). Why study ancient DNA damage? In Journal of Nordic Archaeological Science (Vol. 17).
- Briggs, A. W., & Heyn, P. (2012). Preparation of next-generation sequencing libraries from damaged DNA. Methods in Molecular Biology, 840, 143–154. https://doi.org/10.1007/978-1-61779-516-9_18. [CrossRef]
- Burrell, A. S., Disotell, T. R., & Bergey, C. M. (2015). The use of museum specimens with high-throughput DNA sequencers. Journal of Human Evolution, 79, 35–44. https://doi.org/10.1016/j.jhevol.2014.10.015. [CrossRef]
- Butler, S. L., & Falke, J. J. (1996). Effects of protein stabilizing agents on thermal backbone motions: A disulfide trapping study. Biochemistry, 35(33), 10595–10600. https://doi.org/10.1021/bi961107v. [CrossRef]
- Champlot, S., Berthelot, C., Pruvost, M., Andrew Bennett, E., Grange, T., & Geigl, E. M. (2010). An efficient multistrategy DNA decontamination procedure of PCR reagents for hypersensitive PCR applications. PLoS ONE, 5(9). https://doi.org/10.1371/journal.pone.0013042. [CrossRef]
- Cooper A., & Poinar N. H. (2000). Science’s compass letters. SCIENCE, 289(1139).
- Cortinovis, G., di Vittori, V., Bellucci, E., Bitocchi, E., & Papa, R. (2020). Adaptation to novel environments during crop diversification. In Current Opinion in Plant Biology (Vol. 56, pp. 203–217). Elsevier Ltd. https://doi.org/10.1016/j.pbi.2019.12.011. [CrossRef]
- Cota-Sánchez, J. H., Remarchuk, K., & Ubayasena, K. (2006). Ready-to-Use DNA Extracted with a CTAB Method Adapted for Herbarium Specimens and Mucilaginous Plant Tissue.
- Cozzolino, S., Cafasso, D., Pellegrino, G., Musacchio, A., & Widmer, A. (2007). Genetic variation in time and space: The use of herbarium specimens to reconstruct patterns of genetic variation in the endangered orchid Anacamptis palustris. Conservation Genetics, 8(3), 629–639. https://doi.org/10.1007/s10592-006-9209-7. [CrossRef]
- Cristofolini, G., & Mossetti, U. (1993). Pre-linnean herbaria in Bologna: some newly discovered collections from the time of Ulisse Aldrovandi. https://www.researchgate.net/publication/260179625.
- Cronn, R., Knaus, B. J., Liston, A., Maughan, P. J., Parks, M., Syring, J. v., & Udall, J. (2012). Targeted enrichment strategies for next-generation plant biology. American Journal of Botany, 99(2), 291–311. https://doi.org/10.3732/ajb.1100356. [CrossRef]
- Damerval, C., ben Othman, W., Manicacci, D., & Jabbour, F. (2018). Distribution area of the two floral morphs of Nigella damascena L. (Ranunculaceae): a diachronic study using herbarium specimens collected in France. Botany Letters, 165(3–4), 396–403. https://doi.org/10.1080/23818107.2017.1422437. [CrossRef]
- Davis, C. C., Willis, C. G., Connolly, B., Kelly, C., & Ellison, A. M. (2015). Herbarium records are reliable sources of phenological change driven by climate and provide novel insights into species’ phenological cueing mechanisms. American Journal of Botany, 102(10), 1599–1609. https://doi.org/10.3732/ajb.1500237. [CrossRef]
- Doyle, J. J., & Dickson, E. E. (1987). Preservation of Plant Samples for DNA Restriction Endonuclease Analysis. In Source (Vol. 36, Issue 4).
- Drabkova, L., Kirschner, J., & Vl~k, C. (2002). Comparison of Seven DNA Extraction and Amplification Protocols in Historical Herbarium Specimens of Juncaceae. In International Society for Plant Molecular Biology (Vol. 20).
- Drábková, L. Z. (2014). DNA extraction from herbarium specimens. Methods in Molecular Biology, 1115, 69–84. https://doi.org/10.1007/978-1-62703-767-9_4. [CrossRef]
- Exposito-Alonso, M., Becker, C., Schuenemann, V. J., Reiter, E., Setzer, C., Slovak, R., Brachi, B., Hagmann, J., Grimm, D. G., Chen, J., Busch, W., Bergelson, J., Ness, R. W., Krause, J., Burbano, H. A., & Weigel, D. (2018). The rate and potential relevance of new mutations in a colonizing plant lineage. PLoS Genetics, 14(2). https://doi.org/10.1371/journal.pgen.1007155. [CrossRef]
- Freedman, J., Dorp, V., & Brace, L. &. (2018). Title: Destructive sampling natural science collections: An overview for museum professionals and researchers. http://www.natsca.orgURL:http://www.natsca.org/article/2440.
- Gansauge, M. T., & Meyer, M. (2013). Single-stranded DNA library preparation for the sequencing of ancient or damaged DNA. Nature Protocols, 8(4), 737–748. https://doi.org/10.1038/nprot.2013.038. [CrossRef]
- Gibbs, R. A. (1990). PERSPECTIVE: ANALYTICAL BIOTECHNOLOGY DNA Amplification by the Polymerase Chain Reaction. In Anal. Chem (Vol. 62). https://pubs.acs.org/sharingguidelines.
- Gilbert, M. T. P., Moore, W., Melchior, L., & Worebey, M. (2007). DNA extraction from dry museum beetles without conferring external morphological damage. PLoS ONE, 2(3). https://doi.org/10.1371/journal.pone.0000272. [CrossRef]
- Ginolhac, A., Rasmussen, M., Gilbert, M. T. P., Willerslev, E., & Orlando, L. (2011). mapDamage: Testing for damage patterns in ancient DNA sequences. Bioinformatics, 27(15), 2153–2155. https://doi.org/10.1093/bioinformatics/btr347. [CrossRef]
- Gutaker, R. M., & Burbano, H. A. (2017). Reinforcing plant evolutionary genomics using ancient DNA. In Current Opinion in Plant Biology (Vol. 36, pp. 38–45). Elsevier Ltd. https://doi.org/10.1016/j.pbi.2017.01.002. [CrossRef]
- Gutaker, R. M., Reiter, E., Furtwängler, A., Schuenemann, V. J., & Burbano, H. A. (2017b). Extraction of ultrashort DNA molecules from herbarium specimens. BioTechniques, 62(2), 76–79. https://doi.org/10.2144/000114517. [CrossRef]
- Hart, M. L., Forrest, L. L., Nicholls, J. A., & Kidner, C. A. (2016a). Retrieval of hundreds of nuclear loci from herbarium specimens. Taxon, 65(5), 1081–1092. https://doi.org/10.12705/655.9. [CrossRef]
- Hart, M. L., Forrest, L. L., Nicholls, J. A., & Kidner, C. A. (2016b). Retrieval of hundreds of nuclear loci from herbarium specimens. Taxon, 65(5), 1081–1092. https://doi.org/10.12705/655.9. [CrossRef]
- Heberling, J. M. (2022). HERBARIA AS BIG DATA SOURCES OF PLANT TRAITS. International Journal of Plant Sciences, 183(2), 87–118. https://doi.org/10.1086/717623. [CrossRef]
- Heberling, J. M., Prather, L. A., & Tonsor, S. J. (2019). The Changing Uses of Herbarium Data in an Era of Global Change: An Overview Using Automated Content Analysis. In BioScience (Vol. 69, Issue 10, pp. 812–822). Oxford University Press. https://doi.org/10.1093/biosci/biz094. [CrossRef]
- Jankowiak, K., Buczkowska, K., & Szweykowska-Kulinska, Z. (2005). Successful extraction of DNA from 100-year-old herbarium specimens of the liverwort Bazzania trilobata. Taxon, 54(2), 335–336. https://doi.org/10.2307/25065361. [CrossRef]
- Jobes, D. v, Hurley, D. L., & Thien, L. B. (1995). Plant DNA Isolation: A Method to Efficiently Remove Polyphenolics, Polysaccharides, and RNA. In Source (Vol. 44, Issue 3).
- Jónsson, H., Ginolhac, A., Schubert, M., Johnson, P. L. F., & Orlando, L. (2013). MapDamage2.0: Fast approximate Bayesian estimates of ancient DNA damage parameters. Bioinformatics, 29(13), 1682–1684. https://doi.org/10.1093/bioinformatics/btt193. [CrossRef]
- Kistler, L., Bieker, V. C., Martin, M. D., Pedersen, M. W., Ramos Madrigal, J., & Wales, N. (2020). Ancient Plant Genomics in Archaeology, Herbaria, and the Environment. https://doi.org/10.1146/annurev-arplant-081519. [CrossRef]
- Knapp, M., Clarke, A. C., Horsburgh, K. A., & Matisoo-Smith, E. A. (2012). Setting the stage—Building and working in an ancient DNA laboratory. Annals of Anatomy, 194(1), 3–6. https://doi.org/10.1016/j.aanat.2011.03.008. [CrossRef]
- Kreader, C. A. (1996). Relief of Amplification Inhibition in PCR with Bovine Serum Albumin or T4 Gene 32 Protein. In APPLIED AND ENVIRONMENTAL MICROBIOLOGY (Vol. 62, Issue 3).
- Krinitsina, A. A., Sizova, T. v., Zaika, M. A., Speranskaya, A. S., & Sukhorukov, A. P. (2015). A rapid and cost-effective method for DNA extraction from archival herbarium specimens. Biochemistry (Moscow), 80(11), 1478–1484. https://doi.org/10.1134/S0006297915110097. [CrossRef]
- Kumar Shasany, A., & Darokar, M. (1999). Rapid Isolation of DNA from Dry and Fresh Samples of Plants Producing Large Amounts of Secondary Metabolites and Essential Oils. https://doi.org/10.1023/A:1007528101452. [CrossRef]
- Lambertini, C., Frydenberg, J., Gustafsson, M. H. G., & Brix, H. (2008). Herbarium specimens as a source of DNA for AFLP fingerprinting of Phragmites (Poaceae): Possibilities and limitations. Plant Systematics and Evolution, 272(1–4), 223–231. https://doi.org/10.1007/s00606-007-0633-z. [CrossRef]
- Lister, D. L., Bower, M. A., Howe, C. J., & Jones, M. K. (2008). Extraction and amplification of nuclear DNA from herbarium specimens of emmer wheat: a method for assessing DNA preservation by maximum amplicon length recovery. In TAXON (Vol. 57, Issue 1). https://doi.org/10.2307/25065966. [CrossRef]
- Liu, L., Li, Y., Li, S., Hu, N., He, Y., Pong, R., Lin, D., Lu, L., & Law, M. (2012). Comparison of next-generation sequencing systems. In Journal of Biomedicine and Biotechnology (Vol. 2012). https://doi.org/10.1155/2012/251364. [CrossRef]
- Malenica, N., Šimon, S., Besendorfer, V., Maletic, E., Kontić, J. K., & Pejić, I. (2011). Whole genome amplification and microsatellite genotyping of herbarium DNA revealed the identity of an ancient grapevine cultivar. Naturwissenschaften, 98(9), 763–772. https://doi.org/10.1007/s00114-011-0826-8. [CrossRef]
- Marinček, P., Wagner, N. D., & Tomasello, S. (2022). Ancient DNA extraction methods for herbarium specimens: When is it worth the effort? Applications in Plant Sciences, 10(3). https://doi.org/10.1002/aps3.11477. [CrossRef]
- Martin, M. D., Quiroz-Claros, E., Brush, G. S., & Zimmer, E. A. (2018). Herbarium collection-based phylogenetics of the ragweeds (Ambrosia, Asteraceae). Molecular Phylogenetics and Evolution, 120, 335–341. https://doi.org/10.1016/j.ympev.2017.12.023. [CrossRef]
- Monroe, C., Grier, C., & Kemp, B. M. (2013). Evaluating the efficacy of various thermo-stable polymerases against co-extracted PCR inhibitors in ancient DNA samples. Forensic Science International, 228(1–3), 142–153. https://doi.org/10.1016/j.forsciint.2013.02.029. [CrossRef]
- Müller-Wille, S. (2006). Linnaeus’ herbarium cabinet: a piece of furniture and its function. In Endeavour (Vol. 30, Issue 2, pp. 60–64). https://doi.org/10.1016/j.endeavour.2006.03.001. [CrossRef]
- Myers, J. R., Formiga, A. K., & Janick, J. (2022). Iconography of Beans and Related Legumes Following the Columbian Exchange. Frontiers in Plant Science, 13. https://doi.org/10.3389/fpls.2022.851029. [CrossRef]
- Nelson, G., Paul, D., Riccardi, G., & Mast, A. R. (2012). Five task clusters that enable efficient and effective digitization of biological collections. ZooKeys, 209, 19–45. https://doi.org/10.3897/zookeys.209.3135. [CrossRef]
- Nepi Chiara. (2007). La “slegatura” dell’erbario di Andrea Cesalpino (1525-1603). MUSEOLOGIA SCIENTIFICA Nuova Serie, 1, 50–54.
- Orlando, L., Allaby, R., Skoglund, P., der Sarkissian, C., Stockhammer, P. W., Ávila-Arcos, M. C., Fu, Q., Krause, J., Willerslev, E., Stone, A. C., & Warinner, C. (2021). Ancient DNA analysis. In Nature Reviews Methods Primers (Vol. 1, Issue 1). Springer Nature. https://doi.org/10.1038/s43586-020-00011-0. [CrossRef]
- Pääbo S., Irwin D. M., Wilson A. C., (1990) DNA damage promotes jumping between templates during enzymatic amplification. Journal of Biological Chemistry. Volume 265, Issue 8,. https://doi.org/10.1016/S0021-9258(19)39621-8. [CrossRef]
- Pääbo S, Poinar H, Serre D, Jaenicke-Despres V, Hebler J, Rohland N, Kuch M, Krause J, Vigilant L, Hofreiter M. Genetic analyses from ancient DNA. Annu Rev Genet. 2004;38:645-79. doi: 10.1146/annurev.genet.37.110801.143214. PMID: 15568989. [CrossRef] [PubMed]
- Pont, C., Wagner, S., Kremer, A., Orlando, L., Plomion, C., & Salse, J. (2019). Paleogenomics: Reconstruction of plant evolutionary trajectories from modern and ancient DNA. In Genome Biology (Vol. 20, Issue 1). BioMed Central Ltd. https://doi.org/10.1186/s13059-019-1627-1. [CrossRef]
- Porebski, S., Bailey, L. G., & Baum, B. R. (1997). Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. In Plant Molecular Biology Reporter (Vol. 15, Issue 1).
- Pruvost M, Grange T, Geigl EM. Minimizing DNA contamination by using UNG-coupled quantitative real-time PCR on degraded DNA samples: application to ancient DNA studies. Biotechniques. 2005 Apr;38(4):569-75. doi: 10.2144/05384ST03. PMID: 15884675. [CrossRef] [PubMed]
- Prüfer, K., Stenzel, U., Hofreiter, M., Pääbo, S., Kelso, J., & Green, R. E. (2010). Open Access METHOD Computational challenges in the analysis of ancient DNA. In Genome Biology (Vol. 11). http://genomebiology.com/2010/11/5/R47.
- Psonis, N., Vassou, D., & Kafetzopoulos, D. (2021). Testing a series of modifications on genomic library preparation methods for ancient or degraded DNA. Analytical Biochemistry, 623. https://doi.org/10.1016/j.ab.2021.114193. [CrossRef]
- Riahi, M., Zarre, S., Maassoumi, A. A., Attar, F., & Kazempour Osaloo, S. (2010). An inexpensive and rapid method for extracting papilionoid genomic DNA from herbarium specimens. Genetics and Molecular Research : GMR, 9(3), 1334–1342. https://doi.org/10.4238/vol9-3gmr839. [CrossRef]
- Ribeiro, R., & Lovato, M. (2007a). DNA extraction in fresh and herbarium specimens of Dalbergia Comparative analysis of different DNA extraction protocols in fresh and herbarium specimens of the genus Dalbergia. In Genetics and Molecular Research (Vol. 6, Issue 1). www.funpecrp.com.brwww.funpecrp.com.br.
- Ribeiro, R., & Lovato, M. (2007b). DNA extraction in fresh and herbarium specimens of Dalbergia Comparative analysis of different DNA extraction protocols in fresh and herbarium specimens of the genus Dalbergia. In Genetics and Molecular Research (Vol. 6, Issue 1). www.funpecrp.com.brwww.funpecrp.com.br.
- Rogers, S. O., & Bendich, A. J. (1985). Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. In Plant Molecular Biology (Vol. 5).
- Rosche, C., Baasch, A., Runge, K., Brade, P., Träger, S., Parisod, C., & Hensen, I. (2022). Tracking population genetic signatures of local extinction with herbarium specimens. Annals of Botany, 129(7), 857–868. https://doi.org/10.1093/aob/mcac061. [CrossRef]
- Roullier, C., Benoit, L., Mckey, D. B., & Lebot, V. (2013). Historical collections reveal patterns of diffusion of sweet potato in Oceania obscured by modern plant movements and recombination. PNAS. https://doi.org/10.5061/dryad. [CrossRef]
- Rowe, K. C., Singhal, S., Macmanes, M. D., Ayroles, J. F., Morelli, T. L., Rubidge, E. M., Bi, K., & Moritz, C. C. (2011). Museum genomics: Low-cost and high-accuracy genetic data from historical specimens. Molecular Ecology Resources, 11(6), 1082–1092. https://doi.org/10.1111/j.1755-0998.2011.03052.x. [CrossRef]
- Samarakoon, T., Wang, S. Y., & Alford, M. H. (2013). Enhancing PCR Amplification of DNA from Recalcitrant Plant Specimens Using a Trehalose-Based Additive. Applications in Plant Sciences, 1(1), 1200236. https://doi.org/10.3732/apps.1200236. [CrossRef]
- Santos, D., Ribeiro, G. C., Cabral, A. D., & Sperança, M. A. (2018). A non-destructive enzymatic method to extract DNA from arthropod specimens: Implications for morphological and molecular studies. PLoS ONE, 13(2). https://doi.org/10.1371/journal.pone.0192200. [CrossRef]
- Särkinen, T., Staats, M., Richardson, J. E., Cowan, R. S., & Bakker, F. T. (2012). How to Open the Treasure Chest? Optimising DNA Extraction from Herbarium Specimens. PLoS ONE, 7(8). https://doi.org/10.1371/journal.pone.0043808. [CrossRef]
- Saville, A. C., Martin, M. D., & Ristaino, J. B. (2016). Historic late blight outbreaks caused by a widespread dominant lineage of Phytophthora infestans (Mont.) de Bary. In PLoS ONE (Vol. 11, Issue 12). Public Library of Science. https://doi.org/10.1371/journal.pone.0168381. [CrossRef]
- Savolainen, V., Cui~noud, P., Sp1chiger, R., Martinez, M. D. P., Cri~vecoeur, M., & Manen, J.-F. (1995). Systematics and Evolution The use of herbarium specimens in DNA phylogenetics: evaluation and improvement. In P1. Syst. Evol (Vol. 197).
- Sawyer, S., Krause, J., Guschanski, K., Savolainen, V., & Pääbo, S. (2012). Temporal patterns of nucleotide misincorporations and DNA fragmentation in ancient DNA. PLoS ONE, 7(3). https://doi.org/10.1371/journal.pone.0034131. [CrossRef]
- Shapiro, B., Barlow, A., Heintzman, P. D., Hofreiter, M., Paijmans, J. L. A., & Soares Editors, A. E. R. (n.d.). Ancient DNA Methods and Protocols Second Edition Methods in Molecular Biology 1963. http://www.springer.com/series/7651.
- Shepherd, L. D. (2017). A non-destructive DNA sampling technique for herbarium specimens. PLoS ONE, 12(8). https://doi.org/10.1371/journal.pone.0183555. [CrossRef]
- Signorini, M. A. (1993). SULLE PIANTE DIPINTE DAL BACHIACCA NELLO SCRITTOIO DI COSIMO I A PALAZZO VECCHI. https://www.jstor.org/stable/27654359.
- Smith, J. P. (2017). The Herbarium. https://digitalcommons.humboldt.edu/botany_jps/79.
- Staats, M., Cuenca, A., Richardson, J. E., Ginkel, R. V. van, Petersen, G., Seberg, O., & Bakker, F. T. (2011). DNA damage in plant herbarium tissue. PLoS ONE, 6(12). https://doi.org/10.1371/journal.pone.0028448. [CrossRef]
- Stearn, W. T. (1961). A New Photographic Record of the Linnaean Herbarium. In Source (Vol. 10, Issue 1).
- Stefanaki, A., Porck, H., Grimaldi, I. M., Thurn, N., Pugliano, V., Kardinaal, A., Salemink, J., Thijsse, G., Chavannes-Mazel, C., Kwakkel, E., & van Andel, T. (2018). Breaking the silence of the 500-year-old smiling garden of everlasting flowers: The En Tibi book herbarium. PLoS ONE, 14(6). https://doi.org/10.1371/journal.pone.0217779. [CrossRef]
- Sugita, N., Ebihara, A., Hosoya, T., Jinbo, U., Kaneko, S., Kurosawa, T., Nakae, M., & Yukawa, T. (2020). Non-destructive DNA extraction from herbarium specimens: a method particularly suitable for plants with small and fragile leaves. Journal of Plant Research, 133(1), 133–141. https://doi.org/10.1007/s10265-019-01152-4. [CrossRef]
- Taylor, W., & Swan, E. C. (1994). DNA from Herbarium Specimens. In: Herrmann, B., Hummel, S. (eds) Ancient DNA. https://doi.org/10.1007/978-1-4612-4318-2_11. [CrossRef]
- Thiers, B. M., Tulig, M. C., & Watson, K. A. (2016). Digitization of The New York Botanical Garden Herbarium. Brittonia, 68(3), 324–333. https://doi.org/10.1007/s12228-016-9423-7. [CrossRef]
- Thiers M. (2021). The World’s Herbaria 2020: A Summary Report Based on Data from Index Herbariorum. http://sweetgum.nybg.org/science/ih/.
- Thomas, M., Gilbert, P., Wilson, A. S., Bunce, M., Hansen, A. J., Willerslev, E., Shapiro, B., Higham, T. F. G., Richards, M. P., O’connell, T. C., Tobin, D. J., Janaway, R. C., & Cooper, A. (2004). Ancient mito-chondrial DNA from hair. http://www.current-.
- Thornhill, A. H., Baldwin, B. G., Freyman, W. A., Nosratinia, S., Kling, M. M., Morueta-Holme, N., Madsen, T. P., Ackerly, D. D., & Mishler, B. D. (2017). Spatial phylogenetics of the native California flora. BMC Biology, 15(1). https://doi.org/10.1186/s12915-017-0435-x. [CrossRef]
- Trucchi, E., Benazzo, A., Lari, M., Iob, A., Vai, S., Nanni, L., Bellucci, E., Bitocchi, E., Raffini, F., Xu, C., Jackson, S. A., Lema, V., Babot, P., Oliszewski, N., Gil, A., Neme, G., Michieli, C. T., de Lorenzi, M., Calcagnile, L., … Bertorelle, G. (2021). Ancient genomes reveal early Andean farmers selected common beans while preserving diversity. Nature Plants, 7(2), 123–128. https://doi.org/10.1038/s41477-021-00848-7. [CrossRef]
- Von Engelhardt, D. (2011). LUCA GHINI (1490-1556) IL PADRE FONDATORE DELLA BOTANICA MODERNA NEL CONTESTO DEI RAPPORTI SCIENTIFICI EUROPEI DEL SEDICESIMO SECOLO (1) (Vol. 27).
- Wales, N., Andersen, K., Cappellini, E., Ávila-Arcos, M. C., & Gilbert, M. T. P. (2014). Optimization of DNA recovery and amplification from non-carbonized archaeobotanical remains. PLoS ONE, 9(1). https://doi.org/10.1371/journal.pone.0086827. [CrossRef]
- Wang, W. (2018). A primer to the use of herbarium specimens in plant phylogenetics. Botany Letters, 165(3–4), 404–408. https://doi.org/10.1080/23818107.2018.1438311. [CrossRef]
- Willerslev, E., & Cooper, A. (2005). Ancient DNA. In Proceedings of the Royal Society B: Biological Sciences (Vol. 272, Issue 1558, pp. 3–16). Royal Society. https://doi.org/10.1098/rspb.2004.2813. [CrossRef]
- Willis, C. G., Ellwood, E. R., Primack, R. B., Davis, C. C., Pearson, K. D., Gallinat, A. S., Yost, J. M., Nelson, G., Mazer, S. J., Rossington, N. L., Sparks, T. H., & Soltis, P. S. (2017). Old Plants, New Tricks: Phenological Research Using Herbarium Specimens. In Trends in Ecology and Evolution (Vol. 32, Issue 7, pp. 531–546). Elsevier Ltd. https://doi.org/10.1016/j.tree.2017.03.015. [CrossRef]
- Yoshida, K., Schuenemann, V. J., Cano, L. M., Pais, M., Mishra, B., Sharma, R., Lanz, C., Martin, F. N., Kamoun, S., Krause, J., Thines, M., Weigel, D., & Burbano, H. A. (2013). The rise and fall of the Phytophthora infestans lineage that triggered the Irish potato famine. ELife, 2013(2). https://doi.org/10.7554/eLife.00731. [CrossRef]
- Zedane, L., Hong-wa, C., Murienne, O., Eline Jeziorski, C., Baldwin, B. G., & Besnard, G. (2015). Museomics illuminate the history of an extinct, paleoendemic plant lineage (Hesperelaea, Oleaceae) known from an 1875 collection from Guadalupe Island, Mexico. https://academic.oup.com/biolinnean/article/117/1/44/2440216.
- Zeng, C. X., Hollingsworth, P. M., Yang, J., He, Z. S., Zhang, Z. R., Li, D. Z., & Yang, J. B. (2018). Genome skimming herbarium specimens for DNA barcoding and phylogenomics. Plant Methods, 14(1). https://doi.org/10.1186/s13007-018-0300-0. [CrossRef]
- Zunic, L., Skrbo, A., & Dobraca, A. (2017). Historical Contribution of Pharmaceutics to Botany and Pharmacognosy Development. Materia Socio Medica, 29(4), 291. https://doi.org/10.5455/msm.2017.29.291-300. [CrossRef]
Figure 1.
Example of Herbaria evolution over time. (a) The oldest Illustration of an European Common Bean plant (Phaseolus vulgaris) from “Di Historias Stirpium” (Fuchs, 1542); “Courtesy of Hunt Institute for Botanical Documentation, Carnegie Mellon University, Pittsburgh, PA”. (b) Phaseolus vulgaris specimen from the herbarium of Ulisse Aldrovandi; “Courtesy of Alma Mater Studiorum University of Bologna—University Museum System—Herbarium and Botanical Garden”. (c) Phaseolus vulgaris specimen from the Muséum National d’Histoire Naturelle (MNHN) in Paris dated 1833. The label notes include information about the collector and the collection site and the date.
Figure 1.
Example of Herbaria evolution over time. (a) The oldest Illustration of an European Common Bean plant (Phaseolus vulgaris) from “Di Historias Stirpium” (Fuchs, 1542); “Courtesy of Hunt Institute for Botanical Documentation, Carnegie Mellon University, Pittsburgh, PA”. (b) Phaseolus vulgaris specimen from the herbarium of Ulisse Aldrovandi; “Courtesy of Alma Mater Studiorum University of Bologna—University Museum System—Herbarium and Botanical Garden”. (c) Phaseolus vulgaris specimen from the Muséum National d’Histoire Naturelle (MNHN) in Paris dated 1833. The label notes include information about the collector and the collection site and the date.

Figure 2.
Workflow for the exploitation of Herbarium specimens for genomic studies. (a) A preliminary investigation allows to gain a comprehensive understanding of the available information on Herbarium specimens. The use of web tools, such as web Herbaria, academic databases, and online libraries, allows to find reliable and up-to-date sources and conduct proper research before making any material requests; (b) the identification and selection of the best protocol—aDNA extraction, purification, genotyping—to obtain molecular information from a specimen should take into account the characteristics of the material, available resources, and research objectives; (c) Bioinformatics approaches are needed for data processing; (d) Maximizing the quality of sequencing data is of great importance, as it significantly enhances the chances of reconstructing the phylogeny, understanding the demographic history of a species, and identifying selection signatures in response to natural or human-driven selection.
Figure 2.
Workflow for the exploitation of Herbarium specimens for genomic studies. (a) A preliminary investigation allows to gain a comprehensive understanding of the available information on Herbarium specimens. The use of web tools, such as web Herbaria, academic databases, and online libraries, allows to find reliable and up-to-date sources and conduct proper research before making any material requests; (b) the identification and selection of the best protocol—aDNA extraction, purification, genotyping—to obtain molecular information from a specimen should take into account the characteristics of the material, available resources, and research objectives; (c) Bioinformatics approaches are needed for data processing; (d) Maximizing the quality of sequencing data is of great importance, as it significantly enhances the chances of reconstructing the phylogeny, understanding the demographic history of a species, and identifying selection signatures in response to natural or human-driven selection.
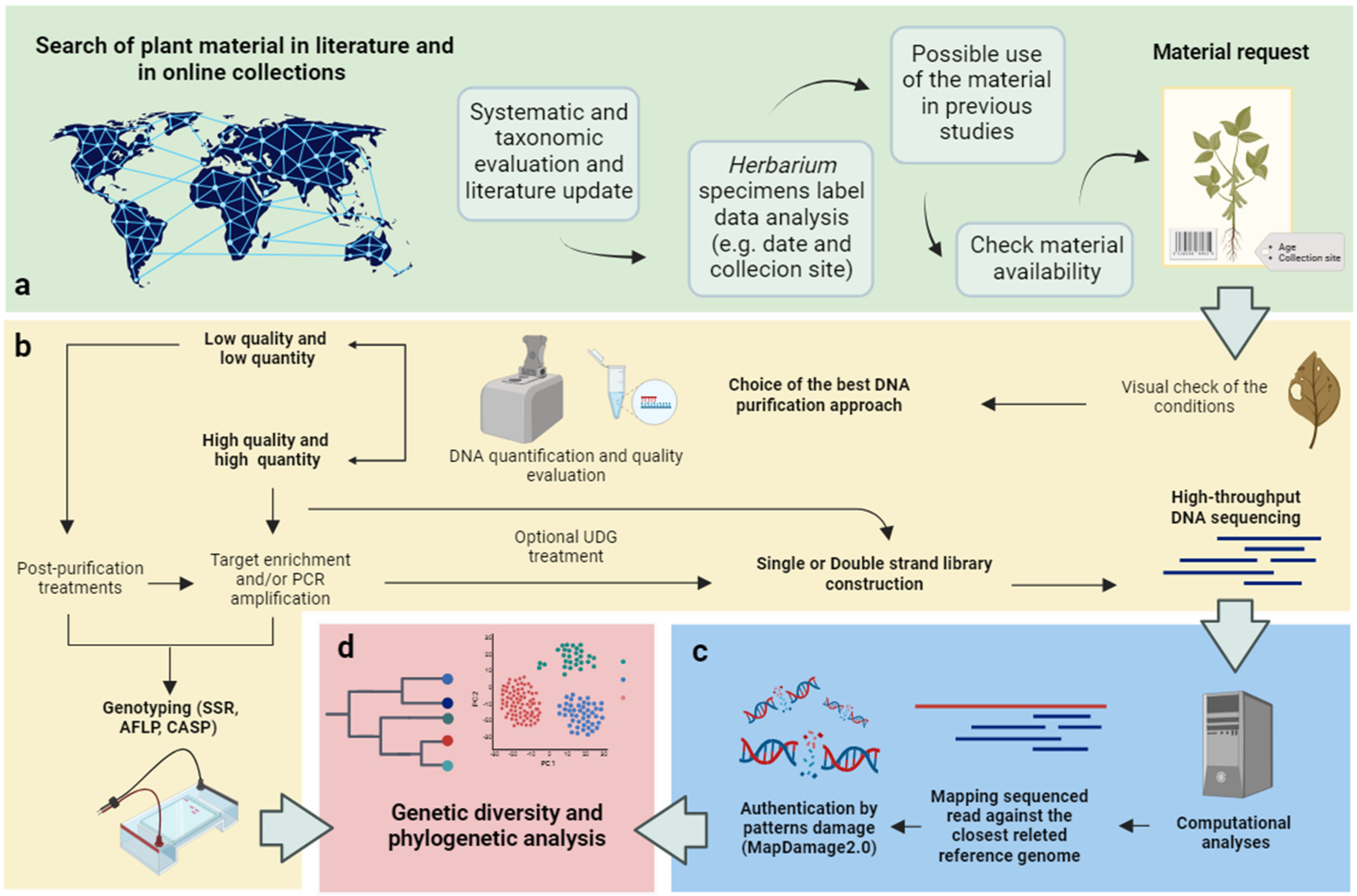
Figure 3.
Contamination sources and guidelines for minimizing negative impact during molecular manipulation of herbarium samples.
Figure 3.
Contamination sources and guidelines for minimizing negative impact during molecular manipulation of herbarium samples.


Table 1.
Summary of relevant aDNA purification protocols from herbaria specimens. The source of plant material, timing of the sampling before storage, success with the tested samples, procedures for the evaluation of aDNA quality, and the reference are provided for each of the tested protocols.
Table 1.
Summary of relevant aDNA purification protocols from herbaria specimens. The source of plant material, timing of the sampling before storage, success with the tested samples, procedures for the evaluation of aDNA quality, and the reference are provided for each of the tested protocols.
| DNA extraction/purification protocol | Source of plant material | Timing of sampling | Suitable for extracting Herbarium DNA | Quality evaluation approach | Reference |
|---|---|---|---|---|---|
| CTAB according to the protocol of Doyle and Doyle (1990) | Juncus and Luzula genera (Juncaceae) | * | Yes | PCR amplification | Drabkova et al., 2002 |
| CTAB + pre-wash with a sorbitol-containing buffer | Lafoensia spp. | N.A. | Yes | PCR amplification | Inglis et al., 2018 |
| Modified CTAB | Agropyronjunceum (Gramineae), Poa juncifolia (Gramineae), Poa palustris, Triticum aestivum (Gramineae), Vicia faba (Fabaceae), Zea mays ssp. mays | N.A. | Yes | Restriction enzymes | Rogers & Bendich 1985 |
| Juncus and Luzula genera (Juncaceae) | N.A. | Yes, but may present CTAB contamination | PCR amplification | Drabkova et al., 2002 | |
| N.A. | ≥60 years | Yes | PCR amplification | Cota-Sánchez et al., 2006 | |
| Species from nine genera of the Papilionoideae | N.A. | Yes | PCR amplification | Riahi et al. (2010) | |
| DNeasy Plant Mini Kit (QIAgen) | Juncus and Luzula genera (Juncaceae) | N.A. | Yes | PCR amplification | Drabkova et al., 2002 |
| DNA extraction with phenol purification and liquid nitrogen | Juncus and Luzula genera (Juncaceae) | N.A. | No | PCR amplification | Drabkova et al., 2002 |
| Long term precipitation in isopropanol and CsCl gradient | Juncus and Luzula genera (Juncaceae) | N.A. | No | PCR amplification | Drabkova et al., 2002 |
| Proteinase K and sodium dodecyl sulfate (SDS) | Scripus hattorianus | 1934 | Yes | PCR amplification | Sugita et al. 2020 |
| N-phenacylthiazolium bromide (PTB)—dithiothreitol (DTT) | Arabidopsis thaliana | Between 1839 and 1898 | Yes | NGS | Gutaker et al. 2017 |
| Phenol-chloroform and silica spin column purification | Herbarium grape leaf tissue (unpublished data) | N.A. | Yes | PCR amplification | Wales et al. 2019 |
| Polyvinylpyrrolidone PVP | genus Dalbergia | N.A. | Yes | PCR amplification | Ribeiro & Lovato 2007 |
| AMPure XP magnetic beads/PEG 8000-containing buffer | genus Scorzonera | Between 1920 and 1960 | Yes | PCR amplification | Krinitsina et al. 2015 |
* Modern plant samples that have been dried using sheets of paper to simulate herbarium specimens preparation. N.A.; not available.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



