
Submitted:
21 March 2024
Posted:
22 March 2024
You are already at the latest version
Abstract
The effects of the climate crisis are affecting ecosystems at different scales and magnitudes. This paper focuses on a massive Mediterranean mussel die-off observed along the middle Italian Adriatic coast in the summer of 2022. We considered the possible environmental causes of this phenomenon and carried out a climatic analysis of the last decade. We performed field surveys in different locations along a 16 Km coastal stretch (from Martinsicuro (TE) on the South, to Grottammare (AP) on the North). The study area includes two marine Sites of Community Importance under the European Natura 2000 network. The mussels die-off has interested practically all the natural mussel-beds colonizing the study area. Mussels are sessile filter-feeding organisms inhabiting the intertidal zone, therefore, are highly exposed to variations in environmental conditions such as temperature and nutrient load. We discuss the possible causes of this die-off, proposing that high temperature and the scarce availability of food acted simultaneously as stress factors, generating local unsustainable living conditions for this species.
Keywords:
Mussel die-off
; food availability
; marine heatwaves
; Adriatic Sea
; climatic crisis
; synergistic effect
1. Introduction
The climate crisis is a global threat to species, biodiversity, and ecosystems; it affects individual organisms and their interactions with other species and habitats, thus altering ecosystem functions and structure [1]. In fact, the Intergovernmental Panel on Climate Change stated that the recent rapid global warming has affected weather, climate, the economy and human society [2]. The impacts of the climate crisis are widespread but not uniform over time and space, as the responses of species and habitats vary according to their relative vulnerability, degree of exposure, sensitivity, and capacity to adapt [3,4,5,6]. The climate crisis affects not only the average temperature but also the duration, magnitude, and frequency of extreme events such as droughts, forest fires, and heat waves [7]. This synergy places great pressure on ecosystems, reducing their resilience [8].
The marine heatwaves (MHWs; anomalously warm water events) that have been striking marine ecosystems throughout the world in recent decades have affected local environmental dynamics [9]. Significant events were observed in the northern Mediterranean Sea in 2003 [10,11], on the west coast of Australia in 2011 [12], in the northwestern Atlantic Ocean in 2012 [13], in the north-eastern Pacific Ocean between 2013 and 2015 [14,15], off Southeast Australia in 2015/16 [16] and across Northern Australia in 2016 [17]. The impacts of MHWs on ecological communities include the loss of kelp forests [18], coral bleaching [19], reduction of surface chlorophyll levels [14], mass mortality of marine invertebrates [20,21] and changes both in species distribution and community structure [18,20,22].
The climate crisis is particularly intense in the semi-enclosed Mediterranean Sea basin where the average warming rate is more intense than the global one; indeed, significant increases in average temperatures, extreme temperatures, number of heat waves and more frequent and intense drought periods are being recorded here [2,23]. The combination of the climate crisis and local climate variations affects the Adriatic ecosystems included in the Mediterranean basin, [24]. The Adriatic Sea is a basin into which most of the rivers of northern and central Italy flow and which communicates with the Mediterranean Sea only through the Otranto channel, which is just 70 km wide. The effects of climatic anomalies are seen more quickly in this marine system because of its small extension and shallow depth, especially in the North and Central Adriatic Sea. For example, studies have detailed how discharge from rivers has influenced phytoplankton bloom dynamics, explored the relationship between atmospheric variability and marine ecosystems productivity, and examined the interplay between air circulation patterns and the occurrence of hypoxia conditions [25,26,27,28,29,30]. Several MHW events have been reported in the Adriatic Sea [31], but no reports have addressed the latest MHW events or their potential association with mass mortality events (MMEs) of marine organisms.
Various climate-induced mass mortalities of marine organisms have been recorded in the Mediterranean Sea over the past 15 years and studies on mass mortality events have explored this phenomenon in the Mediterranean and on both sides of the Adriatic [32]. Mass mortality events of benthic organisms have been well described for the Italian Adriatic coast along the Conero Promontory (about 70 km north of our study area) [33], but the cause of these events was not established. The authors reported that the last episode (2011) occurred during a period with the highest average surface sea-water temperature recorded in Italy in the 30 years from 1961 to 1990 [33]. On the other side of the Adriatic, [32] associated the mortality outbreak event that occurred on coral Balanophyllia europaea in the Mljet National Park (Croatian southern Adriatic Sea) with positive thermal anomalies of the sea temperature.
M.galloprovincialis serves important ecological functions in the Mediterranean Sea, performing fundamental functions in the marine ecosystems, for example bioremediation, substrate structure engineering and feeding, and representing an important food resource for the human population thanks to farms [34,35,36,37].
The aim of our study was to propose potential causes and foresee ecological consequences of an MME of the Mediterranean mussel (Mytilus galloprovincialis) observed along the Piceno Adriatic coast (Marche Region, central Italy) at the end of the summer of 2022. Our main hypothesis is that this MME was caused by a MHW event preceded by a long period of drought, based on the integration of climatological analysis with collected biological data. To our knowledge, this report is the first observation of a mussel MME within this coastal area, which hosts two marine Sites of Community Importance (SCI) and one mussel farm. Two other farms are located one slightly south and one slightly north of the area.
2. Materials and Methods
2.1. Study Area
The study area extends for about 16 km along the coast of the middle Adriatic Sea, locally known as the Piceno coast (Figure 1). The climate is marked by warm and moderately stormy summers, cool winters with rare snowfall, and mild and humid springs and autumns. The absolute maximum precipitation usually occurs in autumn and a slight secondary maximum generally is recorded in the month of May [38]. The rainfall regime is bimodal sublittoral Adriatic and from 1997 to 2022 the annual average rainfall was 593 mm and the temperature averaged 15.7°C [39]. Storms from the NNE to ESE characterize the wave regime of the area. The short tides have an average amplitude of 0.4-0.5 m with a maximum of around 0.75 m.
The study area is characterized by shallow sandy bottoms that slowly deepen towards the open sea: at 300 mt from the coast the average depth is about -4.5 m. There are artificial reefs (linear barriers placed to defend the coast against marine erosion) and a natural reef of about 7 hectares, which is included in the boundaries of the SCI IT534022 "Costa del Piceno - San Nicola a Mare" [40]. Mytilus galloprovincialis strongly colonizes these hard substrates in a depth band ranging from the sea surface to around -2.5 m. In the upper band, from sea surface to -1.5 m, they form almost monospecific populations. These bivalves perform several ecosystem functions such as: a) the increase of the substrate three-dimensional complexity (ecosystem engineering function), b) the control of seston abundance in the water column by filtration (control particle abundance function), c) the transfer of organic matter and nutrients from the water column to the seabed through the production of biodeposits such as feces and pseudofeces (benthic-pelagic coupling and nutrient cycling function), d) the removal of waste from the water (bioremediation function) and, e) being a food resource for other marine organisms (feeding function) [36,41,42,43]. Observation and the biological data collection took place in the following reef environments: Site 1, Grottammare, in the Marine Site of Community Importance (4761502N, 408104E) (natural reef); Site 2, San Benedetto del Tronto harbor (4756933N, 409848E) (artificial reef); Sites 3, San Benedetto del Tronto beach (4754238N, 409930E) (artificial reef); Site 4, Martinsicuro beach (4746627N, 412413E) (artificial reef) (Figure 1). All sites are characterized by a maximum depth of -4.5 meters, and thus temperatures are almost constant throughout the short water column.
Data was also obtained for a mussel farm about three nautical miles from the coast, facing the town of San Benedetto del Tronto, and in two others located just south and north of the study area.
2.2. Observations
For Site 1 the pre- and post- summer 2022 status was assessed through underwater surveys carried out in May/June and September 2022, respectively. Percentage covers of mussels in site 1 were estimated by the visual methods detailed below, which are considered valid procedures for obtaining an accurate representation of relative coverage of sessile organisms [44]. Briefly, ten 50 x 50 cm quadrants with a 10 cm sub-grating were randomly placed in the rocky zones colonized by mussels, in the band depth from sea surface to about -2.5 m. Each square "filled" by mussels counted as 4 % cover, a square 3/4 filled was considered 3 % cover and so on. Data from each quadrant were then used for estimating the percentage of the total area covered by mussels in Site 1. We did not use software programs for digital image analysis because we were interested in evaluating exclusively the layers of mussels on the rock that can be masked by a dense overlying canopy of algae (i.e. Ulva lactuca). This approach was applied only in Site 1 as a representative site of the entire study area and was supported by underwater images and video. Given the consistency of the observed phenomena within the study area, all other sites were monitored by using observational methods which include only underwater visual surveys for qualitative inspection. To evaluate the reduction in mussel farm productivity, data was drawn from the production register of the company that manages the three mussel farms, one in the study area and two close by.
2.3. Climatic Data
In order to analyze the possible causes of the massive death of M. galloprovincialis observed in the study area, we carried out an analysis of the available climatic data. Given the ecological characteristics of this species, we chose the parameters of air and sea temperature, precipitations and river flows into the Sea.
[45] define a MHW as a period of at least five days with temperatures in excess of the threshold of the 90th percentile relative to long-term local climatology, with no more than five days below the threshold. In order to identify MHWs in the last ten summers in our study area, we used three sources for marine weather data, first, the San Benedetto del Tronto measuring station (4756389N, 409228E) and second, the Italian Institute for Environmental Protection and Research, a government agency. The latter provided marine weather data with a variable time step between 10 and 60 minutes, in line with recommendations from the 2008 World Meteorological Organization guide [46,47]. We concentrated on data from 1st January 2011 to 31st December 2022. This signal is homogeneous and almost continuous, except for very short recording gaps. The third source of information was the Copernicus Marine Environment Monitoring Service, whose dataset we used to define the 90th percentile for each day of the summer months (June, July, August) with excellent approximation, drawing upon the Sea Surface Temperature (SST) data for the 30-year from 1993 to 2022 (CMEMS - Reprocessed (REP) Mediterranean (MED) dataset - see SST_MED_SST_L4_REP_OBSERVATIONS_010_021 product). After carrying out a nesting operation, we cross-referenced these data with the hourly data noted above [48].
The biological impacts of MHWs can vary greatly and appear to be context and target species dependent [9]. Therefore, based on both the results of [49], who observed that mussel mortality was 80% and 30% following an exposure for 15 days at 30°C and for 30 days at 28°C respectively, and of [50], who found that the distribution of M. galloprovincialis was defined by an extreme SST of around 29-30°C, we paid particular attention to the MHWs in which the sea surface temperatures exceeded 30°C. To understand how climatologically stressful a summer was, we evaluated the chronic heat stress of organisms (CHSO), quantified as the number of MHW days a site was exposed [51].
Moreover, based on the results of [52,53], who focused on how food availability affects mussel mortality, and considering that in the Italian Adriatic coastal area the abundance of organic matter consumed by these bivalves is mainly supplied by rivers [54], we analyzed the flows of the main rivers along the western coast of the upper Adriatic Sea and the available data about the sea organic load.
3. Results
3.1. Climatic Data Analysis
Detailed analysis was conducted on the historical series relating to thermal parameters for water and air, for a total of more than 1.2 mln of numerical data (period 2011-2022). Of note is the rather significant thermal increase of the SST, quantifiable at about 0.37°C and therefore in line with the indications from the 17th Report on the climate in Italy, published by the Higher Institute for Environmental Protection and Research [55]. The air temperature recorded in San Benedetto del Tronto during the same period increased about 1.08°C. Figure 2 shows the number and amplitude of MHWs in the study area from 2011 to 2022, during the summer months of June, July, and August.
The year 2022 had the greatest number of MHWs (4) and the highest CHSO (34 days of MHW) in the period 2011- 2022. Of the four MHWs detected, one exceeded the 30°C surface sea temperature threshold for 10 days out of 14, and during the remaining 4 days the temperature remained high, between 29.5 and 29.9°C. The maximum temperature detected in 2022 was 30.6°C on August 6th.
The summer of 2015 had the second highest number of MHWs (3) and the greatest CHSO (25 days). Another very long and intense MHW was observed in 2019, when the 90th percentile was exceeded for 14 consecutive days, and the temperature exceeded 30°C for 3 of these days. The highest absolute temperature value (31.2°C) for the 2011 - 2022 period was recorded on August 6th, 2017. Only one 10-day MHW occurred this summer, with temperatures greater than or equal to 30°C. Finally, also in the summer of 2019 there was a 14-day MHW in which the temperature exceeded 30°C for 3 days and ranged between 27.8 and 29.7°C for the remaining 11 days.
3.2. Mussel MME
In the present study, we observed a dramatic disappearance of Mediterranean mussel beds that previous to the summer 2022 had covered the rocky substrates in a depth band between the sea surface and about -2.5 m. In Site 1, the mean percentage of substrate covered by mussels dropped from 81.6% ± 16.3 SD to 0% in little more than two months (Figure 3 and video).
This phenomenon, which affected both intertidal and subtidal zones, was observed after June-August 2022, when four MHWs hit the western central Adriatic coast. Water temperatures reached 30.6°C, and during the last MHW (July 25-August 7), remained above 29.5°C for 14 days and above 30°C for 10 days. Only sporadic live individuals were detected in September in areas previously occupied by mussel beds. Some of these were in the intertidal zone on the north side of a partially submerged rock; a few others ones in the subtidal zone were incorporated into the structure of the bryozoan Schizoporella sp. (Figure 4 and video). Although not quantified, the same scenario was observed at sites 2, 3 and 4, where the mussel populations disappeared at the same time.
There was a 30% drop in mussel production recorded after the summer of 2022 in the mussel farms located in and near the study area.
4. Discussion
4.1. Climatic Data
The air temperature increase of about 1.08°C recorded in San Benedetto del Tronto harbor station from 2011 to 2022 is confirmed by the data recorded in the nearby harbors of Ancona and Ortona, respectively 1.27°C and 1.22 °C. At the meso-scale level we can observe a strong acceleration of climate forcing. The increase of 0.37°C of the SST in the same period is comparable to the data acquired from satellite observations within the Copernicus Marine Environment Monitoring Service (CMEMS) project - Mediterranean SST [56] and with those acquired directly in the Croatian surface waters of the Central Adriatic Sea [23]. From this study two fundamental points emerged. First, in contrast to trends in the previous 20 years, the period after 2005 would be characterized by constant increases in temperature (about 0.04ºC/year) without cooling phases or breaks. Second, there has been a change in the width of the seasonal cycle, characterized by a sharp increase in the uptrend of SST in summer (about 0.056°C/year), and a slowdown in winter (~0.029°C/ year). The increase in SSTs on the eastern coast of the Adriatic Sea, with a total increase exceeding 1°C between 1979 to 2015, was also shown by [57,58].
The 2022 MHWs were caused by the high air temperatures recorded at the end of June along the middle and upper Adriatic Italian coast. Indeed, July 2022 was the hottest July in the last 50 years [59]. These sea heat waves confirm a trend of increasing temperatures in the Mediterranean basin in the last twenty years [31].
The status of the study area sea waters depends not only on the conditions at the mesoscale (the basin of the Tronto and Aso Rivers) but especially on phenomena in the upper Adriatic basin, with freshwater from the main Italian rivers, the most important of which in terms of water flow are the Po, Adige, and Brenta Rivers. In fact, a dominant sea current runs from north to south along the Italian Adriatic coast, transporting the runoff southward.
The reduced river runoff was a crucial aspect of the Central Adriatic waters during 2022. The largest Italian river, the Po, is the main contributor to the nutrient load in the central-northern Adriatic waters [26], and its typical summer flow falls below the range of 550 and 700 mc/sec. Interestingly, the slowest flow of 104 mc/sec was recorded exactly in July 2022 [60]. In 2022 the Po River (Figure 5) and, simultaneously, the Adige, Piave and Brenta Rivers, which are normally of the nivo-glacial type, showed a typical Mediterranean fluvial regime, with a slight maximum in winter and a slight recovery in the flow rate in May.
Analyzing the data of the above-cited ISPRA Report [55], it is possible to observe that the flow rates of these rivers and especially the rainfall that occurred in the catchment areas of the Po and the Adige were extremely low even in the first half of the year, due to a long phase of climatic drought from December 2021 to July 2022. Moreover, this phase was also characterized by significant nivometric deficit on the Alpine and Apennine mountains. The negative anomalies related to the first 6 months of the 2021-22 meteorological years were about 55% in the regions of Piemonte and Valle d'Aosta and about 50% in Lombardy and Tridentine Venice (in northern Italy).
During 2022, the Po River reached the absolute minimum of the last 200 years, with a flow of 104 mc/sec on July 24th at the Ferrara-Pontelagoscuro hydrometric station (Figure 5). Other exceptionally dry periods for the Po River were in June 2022 with a range of 303 mc/sec, in June 2006 with 320 mc/sec, in June 2005 with 444 mc/sec and in June 2003 with 521 mc/sec [60].
As a consequence of all this, the chlorophyll-a was always much lower than the average of the period from January to August 2022 in the mid-Adriatic Sea (south of outlet of the Po River) [61].
In addition, examining the hottest summers, 2017 was marked by a slightly lower negative anomaly in winter and spring precipitation than 2022, but the hydrological data for the Po River flows in spring and summer showed values much higher than those of 2022. Regarding the summer 2015, we observed that the first half of the year was characterized by the average rainfall typical of the last thirty years, with moderate snowfall in the central and eastern Alps and alpine river flows significantly higher than in 2022. In 2019, because of winter snowfalls in line with seasonal averages and abundant rainfall in the months of May and April, the Po River flows had no significant minimums, always remaining above 900 m3/sec until the end of the spring and reaching an absolute minimum of 620 m3/sec during the last ten days of July [60].
4.2. Mussels MME
The death of mussel beds observed in the study area during the summer 2022 is not attributable to phenomena of illegal fishing. First, entire populations died off in little more than two months, which would not be typical results of illegal fishing. Second, illegal mussel harvesting is rare in this area, and in any case, would be easily identified by recognizable holes in the dense mussel beds (Figure 6), which were observed in this study.
Here the main marine predators of the mussels are Sparus aurata, Rapana venosa and Murix sp. Although ecosystem alterations are attributed to the alien Asian gastropod R. venosa [62], predation by this species must be excluded as a possible cause, considering the rapidity with which the mussels totally disappeared (little more than two months), the normal rate at which R. venosa consumes bivalves in the Adriatic Sea [57] and the fact that there was no observed increase of the R. venosa population in the study area before the summer 2022.
Neither polluting events nor harmful algal blooms (HABs) were recorded before and during the summer of 2022 [63]. Similarly, in this period no pathologies were detected in mussels by farmers and authorities during government mandated analyses.
Our findings align with previous studies demonstrating that HMWs correlate with MMEs in the Mediterranean Sea [21,64,65]. However, there is still limited knowledge about this association in the Adriatic Sea area. In the northwestern Mediterranean Sea, mortality incidence between 20% and 100% was registered after MHWs that lasted from 25 to 50 days [65]. This finding suggests that the level of mortality associated with MHWs is highly variable, and demonstrates the need to consider further factors in analyzing the relationship between heat stress and MMEs, for example, the ecological memory of the previous exposures to MHWs over the same geographic areas [66], and potential species or population-related differences [67,68]. For instance, benthic organisms colonizing intertidal zones, such as M. galloprovincialis, are exposed to a variety of harsh marine and terrestrial conditions that could affect their survival [69,70]. In fact, [48] demonstrated that the Mediterranean mussel exposed to high temperatures and a low food load had a 33% survival rate, while those exposed to the same thermal conditions but with a high food load had the significantly better survival rate of 57%.
Previous experimental studies [45] to measure the death levels of M. galloprovincialis in response to increasing temperature showed that individuals exposed to 30ºC reached an 80% mortality rate after 10 days of exposure, peaking at 100% on the 12th day. Indeed, [71] suggests that natural populations of Mediterranean mussels already live in conditions close to their thermal acclimation limits, considering 24–25°C as the upper limit for optimal physiological processes.
Physiological alterations that may result from increased stress include changes in heart rate; [72] observed that rising temperatures increase the heartbeat frequency until a certain critical temperature, after which the heart rate drops. Therefore, the extreme thermal variations of the climate crisis are challenging the physiological limits of these bivalves [72].
Another aspect to consider for both natural and farmed mussel populations is the status of the byssus. Byssal threads are composed of extracellular proteinaceous (collagenous) fibers and are synthesized along the mussel's foot starting from a mussel foot protein [73]. The byssus serves to prevent dislodgement of the bivalve in harsh hydrodynamic conditions, resist mobile competitors, and compete for space [74]. Environmental conditions like the warming of waters and ocean acidification may alter byssal production processes, causing a reduction in strength, stiffness, and extension [75,76]. [77] further demonstrated that in Mytilus coruscus, the expression of mussel foot protein genes is affected by elevated temperature, making the byssus susceptible to ocean warming and leading to a risk of dislodgement at high sea temperature (up to 27°C). Similarly, [78] found that the attachment strength of M. galloprovincialis was negatively correlated with sea surface temperature. The fact that the few live specimens found in the subtidal zone were partially incorporated into the bryozoan Schizoporella sp. (Figure 4), and therefore not anchored to the substrate by byssus, can be seen as a confirmation that the other mussels had been dislodged due to the weakening of the byssus during the heat waves of the summer of 2022. In the same way, the few living specimens in the intertidal zone were observed on the north-facing side of a semi-submerged rock (Figure 4), where the limited daytime solar radiation and the nocturnal exposure to air cooler than sea water probably favored their survival.
One further consequence of increased ocean warming is decreased marine productivity [79]. [52] have suggested that depending on the timing of phytoplankton blooms, marine bivalves may face periods of limited food availability and elevated temperatures, at the same time. These authors reported that with a controlled increase of water temperature, there was a decrease in enzyme kinetics in the Mediterranean mussels in anoxia or normoxia conditions. Interestingly, the authors found that the effects of severe environmental conditions can be limited or countered by sufficient food availability.
River flows and their transportation of nutrients to the sea may have been an important factor in the mussel die off. Our observations show that during the summer of 2022, the study area experienced a hot and dry period which resulted in a low concentration of nutrients released into the sea by rivers. In fact, that year was marked by strong nivometric deficit in winter, low rainfall in spring, and an absolute minimum of river flow in spring and summer. Also 2019, 2017 and especially 2015 were hot years, but only in the first half of 2022 was a much lower organic load (chl-a) observed [61]. Local mussel farmers recorded a reduction of 30% after the summer of 2022, unlike previous years when no decrease in productivity was observed, in line with [80]. Although the environmental factors of temperature, depth and currents differ between the coastal zone study area and the three-mile zone where the farms are located, the decrease in productivity in the same period as the observed mussel disappearance is nevertheless a significant fact, especially considering that farmers have recently implemented procedures to reduce damage from heat waves.
However, the consequences of the stressor combination of MHW, CHSO and low nutrient load on mussel farms are extremely different from those observed in natural settlements because of the good farming practices adopted by farmers. They have reduced damage in certain periods and allowed production to resume by choosing to harvest the mussels before the hottest periods and using biodegradable braided knit or cotton nets called socks to counter dislodgement. On the contrary, alterations of natural habitats driven by the climate crisis can be dramatic and require a longer time to restore. There is the negative possibility that natural habitat reassembly may occur with components and structures same from the those of the initial condition.
Ultimately, we hypothesize that the MME of mussels observed in our study area could be due to a vicious cycle of MHW and CHSO thermal stressors, combined with low nutrient availability, which resulted in the increased metabolic demand not being met.
5. Conclusions
Based on our field survey, climatic data analysis, and also considering previous findings on M. galloprovincialis tolerance under certain stress conditions, we suggest that the MME observed in the summer of 2022 in the study area was likely due to prolonged anomalously warm water combined with reduced nutrient supply.
It is hoped that these results should offer a useful contribution to the monitoring of ecosystems and the management of M. galloprovincialis mussel farms.
Considering that these extreme climatological events will probably intensify [81], our preliminary analysis stresses the need for adopting specific mitigation and restoration measures in order to improve the management of marine natural ecosystems and fisheries.
Author Contributions
Conceptualization, L.B. and F.A.P.; formal analysis, L.B., M.F.; investigation, L.B., M.C. and P.C.; methodology, L.B., M.C., M.F. and P.C.; software, L.B. and M.F.; Data Curation, L.B., M.C.; supervision, L.B. and F.A.P.; visualization, L.B. and P.C.; Writing – Original Draft Preparation, L.B., M.C., P.C. and F.A.P.; writing-review and editing, L.B., M.C. and F.A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available as they are partly owned by local governments.
Acknowledgments
We would like to thank the “Autorità di bacino distrettuale fiume Po” for the historical flows of the Po River data provided, and the Mediterranean Aquaculture Association (AMA) for both the mussel productivity data and for sharing their experience on mussel farming and techniques. We would also like to thank Sheila Beatty for editing the English usage in the manuscript.
Conflicts of Interest
This research received no external funding.
References
- Díaz, S.; Settele, J.; Brondizio, E.; Ngo, H.T.; Guèze, M.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.A.; Butchart, S.H.M. Summary for Policymakers of the Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-policy Platform on Biodiversity and Ecosystem Services. Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services 2019, Bonn, Germany, 39.
- Shukla, P.R.; Skea, J.; Calvo Buendia, E.; Masson-Delmotte, V.; Pörtner, H.O. Special report - Climate change and land. Intergovernmental Panel on Climate Change 2019. Available online: https://www.ipcc.ch/report/srccl/ (accessed on 20 October 2022).
- Beever, E.A.; O’Leary, J.; Mengelt, C.; West, J.M.; Julius, S.; Green, N.; Managness, D.; Petes, L.E.; Stein, B.A.; Nicotra, A.B.; et al. Improving conservation outcomes with a new paradigm for understanding species’ fundamental and realized adaptive capacity. Conserv. Lett. 2016, 9, 131–137. [Google Scholar] [CrossRef]
- Foden, W.B.; Young, B.E. IUCN SSC Guidelines for Assessing Species’ Vulnerability to Climate Change. Version 1.0. Occasional Paper of the IUCN Species Survival Commission No. 59. Cambridge, UK and Gland, Switzerland, 2016; 114. [CrossRef]
- Glick, P.; Stein, B.A.; Edelson, N.A. Scanning the Conservation Horizon: A Guide to Climate Change Vulnerability Assessment. National Wildlife Federation, Washington, DC, 2011; 168.
- Kovach, R.P.; Dunham, J.B.; Al-Chokhachy, R.; Snyder, C.D.; Letcher, B.H.; Young, J.A.; Beever, E.A.; Pederson, G.T.; Lynch, A.J.; Hitt, N.P.; et al. An integrated framework for ecological drought across riverscapes of North America. Bioscience 2019, 69, 418–431. [Google Scholar] [CrossRef]
- Jay, A.; Reidmiller, D.R.; Avery, C.W.; Barrie, D.; DeAngelo, B.J. 2018. Impacts, Risks, and Adaptation in the United States: Fourth National Climate Assessment, Volume II. U.S. Global Change Research Program, Washington, D.C., USA, pp. 33–71.
- Malhi, Y.; Franklin, J.; Seddon, N.; Solan, M.; Turner, M. G.; Field, C. B.; Knowlton, N. Climate change and ecosystems: Threats, opportunities and solutions. Philos. Trans. Royal Soc. A 2020, 375(1794), 20190104. [Google Scholar] [CrossRef] [PubMed]
- Hobday, A.J.; Oliver, E.C.; Gupta, A.S.; Benthuysen, J.A.; Burrows, M.T.; Donat, M.G.; Holbrook, N.J; Moore, P.J.; Thomsen, M.S.; Wernberg, T.; et al. Categorizing and naming marine heatwaves. Oceanogr. 2018, 31(2), 162–173. [Google Scholar] [CrossRef]
- Sparnocchia, S.; Schiano, M. E.; Picco, P.; Bozzano, R.; Cappelletti, A. The anomalous warming of summer 2003 in the surface layer of the Central Ligurian Sea (Western Mediterranean). Ann. Geophys. 2006, 24, 443–452. [Google Scholar] [CrossRef]
- Olita, A.; Sorgente, R.; Ribotti, A.; Natale, S.; Gaberšek, S. Effects of the 2003 European heatwave on the Central Mediterranean Sea surface layer: a numerical simulation. Ocean Sci. 2006, 3, 85–125. [Google Scholar]
- Pearce, A.F.; Feng, M. The rise and fall of the ‘marine heat wave’ off Western Australia during the summer of 2010/11. J. Mar. Syst. 2013, 112, 139–156. [Google Scholar] [CrossRef]
- Chen, K.; Gawarkiewicz, G.G.; Lentz, S.J.; Bane, J.M. Diagnosing the warming of the Northeastern US Coastal Ocean in 2012: a linkage between the atmospheric jet stream variability and ocean response. J. Geophys. Res. Oceans 2014, 119, 218–227. [Google Scholar] [CrossRef]
- Bond, N.A.; Cronin, M.F.; Freeland, H.; Mantua, N. Causes and impacts of the 2014 warm anomaly in the NE Pacific. Geophys. Res. Lett. 2015, 42, 3414–3420. [Google Scholar] [CrossRef]
- Di Lorenzo, E.; Mantua, N. Multi-year persistence of the 2014/15 North Pacific marine heatwave. Nat. Clim. Change 2016, 6, 1042–1047. [Google Scholar] [CrossRef]
- Oliver, E.C.J.; Donat, M.G.; Burrows, M.T.; Moore, P.J.; Smale, D.A.; Alexander, L.V.; Benthuysen, J.A.; Feng, M.; Gupta, A.S.; Hobday, A.J.; et al. Longer and more frequent marine heatwaves over the past century. Nat. Commun. 2018, 9, 1324. [Google Scholar] [CrossRef]
- Benthuysen, J.A.; Oliver, E.C.J.; Feng, M.; Marshall, A.G. Extreme marine warming across tropical Australia during austral summer 2015-16. J. Geophys. Res. Oceans 2018, 123, 1301–1326. [Google Scholar] [CrossRef]
- Wernberg, T.; Bennett, S.; Babcock, R.C.; De Bettignies, T.; Cure, K.; Depczynski, M.; Dufois, F.; Fromont, J.; Fulton, C.J.; Hovey, R.K.; et al. Climate-driven regime shift of a temperate marine ecosystem. Science 2016, 353, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.; Álvarez-Noriega, M.; Álvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Badcock, R.C.; Beger, M.; Bellwood, D.R.; et al. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Oliver, E.; Benthuysen, J.; Bindoff, N.; Hobday, A.J.; Holbrook, N.J.; Mundy, C.N.; Perkins-Kirkpatrick, S.E. The unprecedented 2015/16 Tasman Sea marine heatwave. Nat. Commun. 2017, 8, 16101. [Google Scholar] [CrossRef] [PubMed]
- Garrabou, J.; Coma, R.; Bensossan, N.; Bally, M.; Chevaldonné, P.; Cigliano, M.; Diaz, D.; Harmelin, J.G.; Gambi, M.C.; Kersting, D.K.; et al. Mass mortality in Northwestern Mediterranean rocky benthic communities: effects of the 2003 heat wave. Glob. Chang. Biol. 2009, 15, 1090–1103. [Google Scholar] [CrossRef]
- Cavole, L.M.; Demko, A.M.; Diner, R.E.; Giddings, A.; Koester, I.; Pagniello, C.M.L.S.; Paulsen, M.L.; Ramirez-Valdez, A.; Schwenck, S.M.; Yen, N.K.; et al. Biological impacts of the 2013--2015 warm-water anomaly in the Northeast Pacific: winners, losers, and the future. Oceanography 2016, 29, 273–285. [Google Scholar] [CrossRef]
- Bonacci, O.; Bonacci, D.; Patekar, M.; Pola, M. Increasing Trends in Air and Sea Surface Temperature in the Central Adriatic Sea (Croatia). J. Mar. Sci. Eng. 2021, 9(4), 358. [Google Scholar] [CrossRef]
- Grbec, B.; Morović, M.; Matić, F.; Ninčević Gladan, Ž.; Marasović, I.; Vidjak, O.; Bojanić, N.; Čikes Keć, V.; Zorica, B.; Kusplić, G; et al. Climate regime shifts and multi-decadal variability of the Adriatic Sea pelagic ecosystem. Acta Adriat. 2015, 56, 47––66. [Google Scholar]
- Grilli, F.; Accoroni, S.; Acri, F.; Bernardi Aubry, F.; Bergami, C.; Cabrini, M.; Campanelli, A.; Giani, M.; Guicciardi, S.; Marini, M.; et al. Seasonal and Interannual Trends of Oceanographic Parameters over 40 Years in the Northern Adriatic Sea in Relation to Nutrient Loadings Using the EMODnet Chemistry Data Portal. Water 2020, 12, 2280. [Google Scholar] [CrossRef]
- Penna, N.; Cappellacci, S.; Ricci, F. The influence of the Po River discharge on phytoplankton bloom dynamics along the coastline of Pesaro (Italy) in the Adriatic Sea. Mar. Poll. Bull. 2004, 48, 321–326. [Google Scholar] [CrossRef]
- Schiano, M.E.; Sparnocchia, S.; Cappa, C.; Bozzano, R. An analysis of the climate variability over the Mediterranean Sea by means of the surface water vapour density. Int. J. Climatol. 2005, 25, 1731–1748. [Google Scholar] [CrossRef]
- Grbec, B.; Morović, M.; Beg Paklar, G.; Kušpilić, G.; Matijević, S.; Matić, F.; Gladan, Ž. The relationship between the atmospheric variability and productivity in the Adriatic Sea area. J. Mar. Biol. Assoc. U.K. 2009, 89, 1549–1558. [Google Scholar] [CrossRef]
- Giani, M.; Djakovac, T.; Degobbis, D.; Cozzi, S.; Solidoro, C.; Umani, S.F. Recent changes in the marine ecosystems of the northern Adriatic Sea. Estuar. Coast. Shelf Sci. 2012, 115, 1–13. [Google Scholar] [CrossRef]
- Djakovac, T.; Supić, N.; Bernardi Aubry, F.; Degobbis, D.; Giani, M. Mechanisms of hypoxia frequency changes in the northern Adriatic Sea during the period 1972–2012. J. Mar. Syst. 2015, 141, 179–189. [Google Scholar] [CrossRef]
- Juza, M.; Fernández-Mora, À.; Tintoré, J. Sub-Regional Marine Heat Waves in the Mediterranean Sea from Observations: Long-Term Surface Changes, Sub-Surface and Coastal Responses. Front. Mar. Sci. 2022, 9, 785771. [Google Scholar] [CrossRef]
- Kružić, P.; Popijač, A. Mass mortality events of the coral Balanophyllia europaea (Scleractinia, Dendrophylliidae) in the Mljet National Park (eastern Adriatic Sea) caused by sea temperature anomalies. Coral reefs 2015, 34, 109–118. [Google Scholar] [CrossRef]
- Di Camillo, C.G.; Cerrano, C. Mass mortality events in the NW Adriatic Sea: phase shift from slow-to fast-growing organisms. PloS one 2015, 10(5), e0126689. [Google Scholar] [CrossRef]
- Cerrano, C.; Pica, D.; Di Camillo, C.; Bastari, A.; Torsani, F. Caratterizzazione biocenotica e restituzione cartografica per l’individuazione di habitat e specie di interesse comunitario lungo la costa marchigiana. Regione Marche, Ancona, Italy, 2014; pp. 55.
- Gazeau, F.; Alliouane, S.; Bock, C.; Bramanti, L.; López Correa, M.; Gentile, M.; Hirse, T.L.; Portner, H.O.; Ziveri, P. Impact of ocean acidification and warming on the Mediterranean mussel (Mytilus galloprovincialis). Front. Mar. Sci. 2014, 1, 62. [Google Scholar] [CrossRef]
- Bracchetti, L.; Capriotti, M. Le Formazioni a Reff della costa Picena (The reef formations of the Piceno coast, central Italy). Studi Costieri 2021, 30, 83–92. [Google Scholar]
- EUMOFA, E.U. The EU fish market. Eur. Mark. Obs. Fish. Aquac. Prod, 2023. https://eumofa.eu.
- Pesaresi, S.; Biondi, E.; Casavecchia, S. Bioclimates of Italy. J. Maps 2017, 13(2), 955–960. [Google Scholar] [CrossRef]
- Fazzini, M.; Beltrando, G.; Billi, P. Intense rainfalls and flooding problems in the beach resort of San Benedetto del Tronto, Adriatic Sea, Central Italy. Proc. 3°GEOMED 2013, Antalya Turkey, 10-14 june; Ibrahim Athalay and Recept EYE, 128-131.
- Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:31992L0043 (accessed on 15 September 2022).
- Dame, R.F., Kenneth, M.J. Ecology of marine bivalves: an ecosystem approach. CRC Press, Taylor & Francis, Boca Raton, FL, US, 2011; pp. 284.
- Ward, J.E.; Shumway, S.E. Separating the grain from the chaff: particle selection in suspension-and deposit-feeding bivalves. J. Exp. Mar. Biol. Ecol. 2004, 300(1-2), 83–130. [Google Scholar] [CrossRef]
- Broszeit, S.; Hattam, C.; Beaumont, N. Bioremediation of waste under ocean acidification: Reviewing the role of Mytilus edulis. Mar. Poll. Bull. 2016, 103(1-2), 5–14. [Google Scholar] [CrossRef]
- Murray, S.N.; Ambrose, R.F.; Dethier, M.N. Methods for Performing Monitoring, Impact, and Ecological Studies on Rocky Shores; MMS U.S. Department of the Interior Minerals Management Service: Pacific OCS Region, 2002; pp. 116-147.
- Hobday, A. J.; Alexander, L. V.; Perkins, S. E.; Smale, D. A.; Straub, S. C.; Oliver, E. C. J.; Benthuysen, J. A.; Burrows, M. T.; Donat, M. G.; Feng, M.; Holbrook, N. J.; Moore, P. J.; Scannell, H. A.; Sen Gupta, A; Wernberg, T. A hierarchical approach to defining marine heatwaves. Progress in Oceanography 2016, 141, 227–238. [Google Scholar] [CrossRef]
- WMO (World Meteorological Organization), 2008. Guide to Meteorological Instruments and Methods of Observation - No.8 (Seventh edition). https://www.weather.gov/media/epz/mesonet/CWOP-WMO8.pdf.
- National tide gauge network of ISPRA (Istituto Superiore per la Protezione e la Ricerca Ambientale). Available online: https://mareografico.it/?session=0S71809491267K766873MJG&syslng=ita&sysmen=-1&sysind=-1&syssub=-1&sysfnt=0&code=STAZ&idst=1W (accessed on 20 June 2023).
- Pastor, F.; Valiente, J.A.; Khodayar, S. A Warming Mediterranean: 38 Years of Increasing Sea Surface Temperature. Remote Sens. 2020, 12, 2687. [Google Scholar] [CrossRef]
- Anestis, A.; Lazou, A.; Portner, H O.; Michaelidis, B. Behavioral, metabolic, and molecular stress responses of marine bivalve Mytilus galloprovincialis during long-term acclimation at increasing ambient temperature. Am. J. Physiol. Regul. Integr. Comp. Physiol 2007, 293(2), R911-21. [Google Scholar] [CrossRef]
- Fly, E.K.; Hilbish, T.J.; Wethey, D.S.; Rognstad, R.L. Physiology and Biogeography: The Response of European Mussels (Mytilus spp.) to Climate Change. Am. Malacol. Bull. 2015, 33(1), 136–149. [Google Scholar] [CrossRef]
- Garrabou, J.; Gómez-Gras, D.; Medrano, A.; Cerrano, C.; Ponti, M.; Schlegel, R.; Bensoussan, N.; Turicchia, E.; Sini, M.; Gerovasileiou, M; et al. Marine heatwaves drive recurrent mass mortalities in the Mediterranean Sea. Global Change Biology 2022, 28(19), 5708–5725. [Google Scholar] [CrossRef]
- Parisi, M.G.; Mauro, M.; Sarà, G.; Cammarata, M. Temperature increases, hypoxia, and changes in food availability affect immunological biomarkers in the marine mussel Mytilus galloprovincialis. J. Comp. Physiol. B 2017, 187, 1117–1126. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K. R.; Van Thiel, L. E.; Helmuth, B. Interactive effects of food availability and aerial body temperature on the survival of two intertidal Mytilus species. J. Therm. Biol. 2010, 35(4), 161–166. [Google Scholar] [CrossRef]
- Rampazzo, F.; Berto, D.; Giani, M.; Brigolin, D.; Covelli, S.; Cacciatore, F.; Boscolo Brusà, R.; Bellucci, L.C.; Pastres, R. Impact of mussel farming on sedimentary geochemical properties of a Northern Adriatic area influenced by freshwater inflows. Estuarine, Coastal and Shelf Science, 2013, 129, 49–58. [Google Scholar] [CrossRef]
- ISPRA (Istituto Superiore per la Protezione e la Ricerca Ambientale), 2021. Gli indicatori del Clima in Italia nel 2021 Anno XVII - Stato dell’ambiente 98/2022. In italian, available only in electronic format at: https://www.isprambiente.gov.it/files2022/pubblicazioni/stato-ambiente/rapporto_clima_2021.pdf.
- Pisano, A.; Marullo, S.; Artale, V.; Falcini, F.; Yang, C.; Leonelli, F.E.; Santoleri, R.; Buongiorno Nardelli, B. New Evidence of Mediterranean Climate Change and Variability from Sea Surface Temperature Observations. Remote Sens. 2020, 12, 132. [Google Scholar] [CrossRef]
- Šolić, M.; Grbec, B.; Matić, F.; Šantić, D.; Šestanović, S.; Gladan, Ž.N.; Bojanić, N.; Ordulj, M.; Jozić, S.; Vrdoljak, A. Spatio-temporal reproducibility of the microbial food web structure associated with the change in temperature: Long-term observations in the Adriatic Sea. Oceanography 2018, 161, 87–101. [Google Scholar] [CrossRef]
- Grbec, B.; Matić, F.; Beg Paklar, G.; Morović, M.; Popović, R.; Vilibić, I. Long-Term Trends, Variability and Extremes of In Situ Sea Surface Temperature Measured Along the Eastern Adriatic Coast and its Relationship to Hemispheric Processes. Pure Aappl. Geophys. 2018, 175, 4031–4046. [Google Scholar] [CrossRef]
- Fazzini, M.; Fiore, A.; Sammartino, G. Preliminary analysis of climate data and geo-hydrological instability that affected the island of Ischia on November 26, 2022. Geologia dell’Ambiente 2023, 1/2023 ISSN 1591-5352; 12-25. 26 November.
- ARPAE, 2023. Agenzia Regionale Protezione Ambientale Emilia-Romagna. Data reserved.
- www.uniurb.it. Available online (bulletins only in Italian): https://www.uniurb.it/ricerca/organizzazione-della-ricerca/strutture-della-ricerca/qualita-delle-acque-della-costa.
- Savini, D.; Occhipinti-Ambrogi, A. Consumption rates and prey preference of the invasive gastropod Rapana venosa in the Northern Adriatic Sea. Helgol Mar Res 2006, 60, 153–159. [Google Scholar] [CrossRef]
- Regional Agency for Environmental Protection of Marche Region. Relazione annuale sulla qualità delle acque di balneazione stagione 2022 (Annual report on the quality of bathing water season 2022). ARPAM, Ancona, Italy, 2022, 62, 75.
- Cerrano, C.; Bavestrello, G.; Bianchi, C.N.; Cattaneo-Vietti, R.; Bava, S.; Morganti, C.; Morri, C.; Picco, P.; Sara, G.; Schiaparelli, S.; et al. Catastrophic mass-mortality episode of gorgonians and other organisms in the Ligurian Sea (North-Western Mediterranean), summer 1999. Ecol. Lett. 2000, 3(4), 284–293. [Google Scholar] [CrossRef]
- Garrabou, J.; Gómez-Gras, D.; Medrano, A.; Cerrano, C.; Ponti, M.; Schlegel, R.; Bensoussan, N.; Turicchia, E; Sini, M.; Gerovasileiou, V.; et al. Marine heatwaves drive recurrent mass mortalities in the Mediterranean Sea. Glob. Chang. Biol. 2022, 28(19), 5708–5725. [Google Scholar] [CrossRef]
- Turner, M. G.; Calder, W. J.; Cumming, G. S.; Hughes, T. P.; Jentsch, A.; LaDeau, S.L.; Lenton, T.M.; Shuman, B.N.; Turetsky, M.R.; Ratajczak, M.; et al. Climate change, ecosystems and abrupt change: Science priorities. Philos. Trans. Royal Soc. B 2020, 375, 20190105. [Google Scholar] [CrossRef] [PubMed]
- Pagès-Escolà, M.; Hereu, B.; Garrabou, J.; Montero-Serra, I.; Gori, A.; Gómez-Gras, D.; Figuerola, B.; Linares, C. Divergent responses to warming of two common co-occurring Mediterranean bryozoan. Sci. Rep. 2018, 8(1), 17455. [Google Scholar] [CrossRef]
- Gómez-Gras, D.; Linares, C.; de Caralt, S.; Cebrian, E.; Frleta-Valić, M.; Montero-Serra, I.; Pagès-Escolà, M.; López-Sendino, P.; Garrabou, J. Response diversity in Mediterranean coralligenous assemblages facing climate change: Insights from a multi-specific thermotolerance experiment. Ecol. Evol. 2019, 9(7), 4168–4180. [Google Scholar] [CrossRef] [PubMed]
- Somero, G.N. Thermal Physiology and Vertical Zonation of Intertidal Animals: Optima, Limits, and Costs of Living. Integrative and Comp. Biol. 2002, 42(4), 780–789. [Google Scholar] [CrossRef]
- Harley, C.D. Tidal dynamics, topographic orientation, and temperature-mediated mass mortalities on rocky shores. Mar. Ecol. Prog. Ser. 2008, 371, 37–46. [Google Scholar] [CrossRef]
- Anestis, A.; Pörtner, H.O.; Karagiannis, D.; Angelidis, P.; Staikou, A.; Michelidis, B. Response of Mytilus galloprovincialis (L.) to increasing seawater temperature and to marteliosis: metabolic and physiological parameters. Comp. Biochem.Physiol. Part A Mol. Integr. Physiol. 2010, 156, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Brady, C.E.; Somero, G.N. Following the heart: temperature and salinity effects on heart rate in native and invasive species of blue mussels (genus Mytilus). J. Exp. Biol. 2006, 209, 2554–2566. [Google Scholar] [CrossRef]
- Priemel, T.; Degtyar, E.; Dean, M.; Harrington, M.J. Rapid self-assembly of complex biomolecular architectures during mussel byssus biofabrication. Nat. Commun. 2017, 8, 14539. [Google Scholar] [CrossRef] [PubMed]
- Carrington, E.; Waite, J.H.; Sarà, G.; Sebens, K.P. Mussels as a Model System for Integrative Ecomechanics. Annu. Rev. Mar. Sci. 2015, 7, 443–469. [Google Scholar] [CrossRef]
- Lachance, A.A.; Myrand, B.; Tremblay, R.; Koutitonsky, V.; Carrington, E. Biotic and abiotic factors influencing attachment strength of blue mussels Mytilus edulis in suspended culture. Aquat. Biol. 2008, 2, 119–129. [Google Scholar] [CrossRef]
- O’Donnell, M.J.; George, M.N.; Carrington, E. Mussel byssus attachment weakened by ocean acidification. Nat. Clim. Change 2013, 3(6), 587–590. [Google Scholar] [CrossRef]
- Li, Y.F.; Yang, X.Y.; Cheng, Z.Y.; Wang, L.Y.; Wang, W.X.; Liang, X.; Yang, J.L. Near-future levels of ocean temperature weaken the byssus production and performance of the mussel Mytilus coruscus. Sci. Total Environ. 2020, 1(733), 139347. [Google Scholar] [CrossRef] [PubMed]
- Zardi, G.I.; McQuaid, C.D.; Nicastro, K.R. Balancing survival and reproduction: seasonality of wave action, attachment strength and reproductive output in indigenous Perna perna and invasive Mytilus galloprovincialis mussels. Mar. Ecol. Prog. Ser. 2007, 334, 155–163. [Google Scholar] [CrossRef]
- Watson, W.G.; Margarita, E.C.; Ginoux, P.; O'Reilly, J.E.; Casey, N.W. Ocean primary production and climate: Global decadal changes. Geophys. Res. Lett. 2003, 30(15), 1809. [Google Scholar] [CrossRef]
- Martinez, M.; Mangano, M.C.; Maricchiolo, G.; Genovese, L.; Mazzola, A.; Sarà, G. Measuring the effects of temperature rise on Mediterranean shellfish aquaculture. Ecol. Indic. 2018, 88, 71–78. [Google Scholar] [CrossRef]
- Stocker, T.F.; Qin, D.; Plattner, G.K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. Climate Change 2013: The Physical Science Basis. Contribution of Working Group 1 to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK and New York, USA, 2013, pp. 1523.
Figure 1.
Sites where the massive mussel die-off was observed during summer 2022. 1: Grottammare natural reef; 2: San Benedetto del Tronto harbor; 3: San Benedetto del Tronto beach artificial barriers; 4: Martinsicuro beach artificial barriers. SCI: Marine Sites of Community Importance. Photos by Google Earth.
Figure 1.
Sites where the massive mussel die-off was observed during summer 2022. 1: Grottammare natural reef; 2: San Benedetto del Tronto harbor; 3: San Benedetto del Tronto beach artificial barriers; 4: Martinsicuro beach artificial barriers. SCI: Marine Sites of Community Importance. Photos by Google Earth.
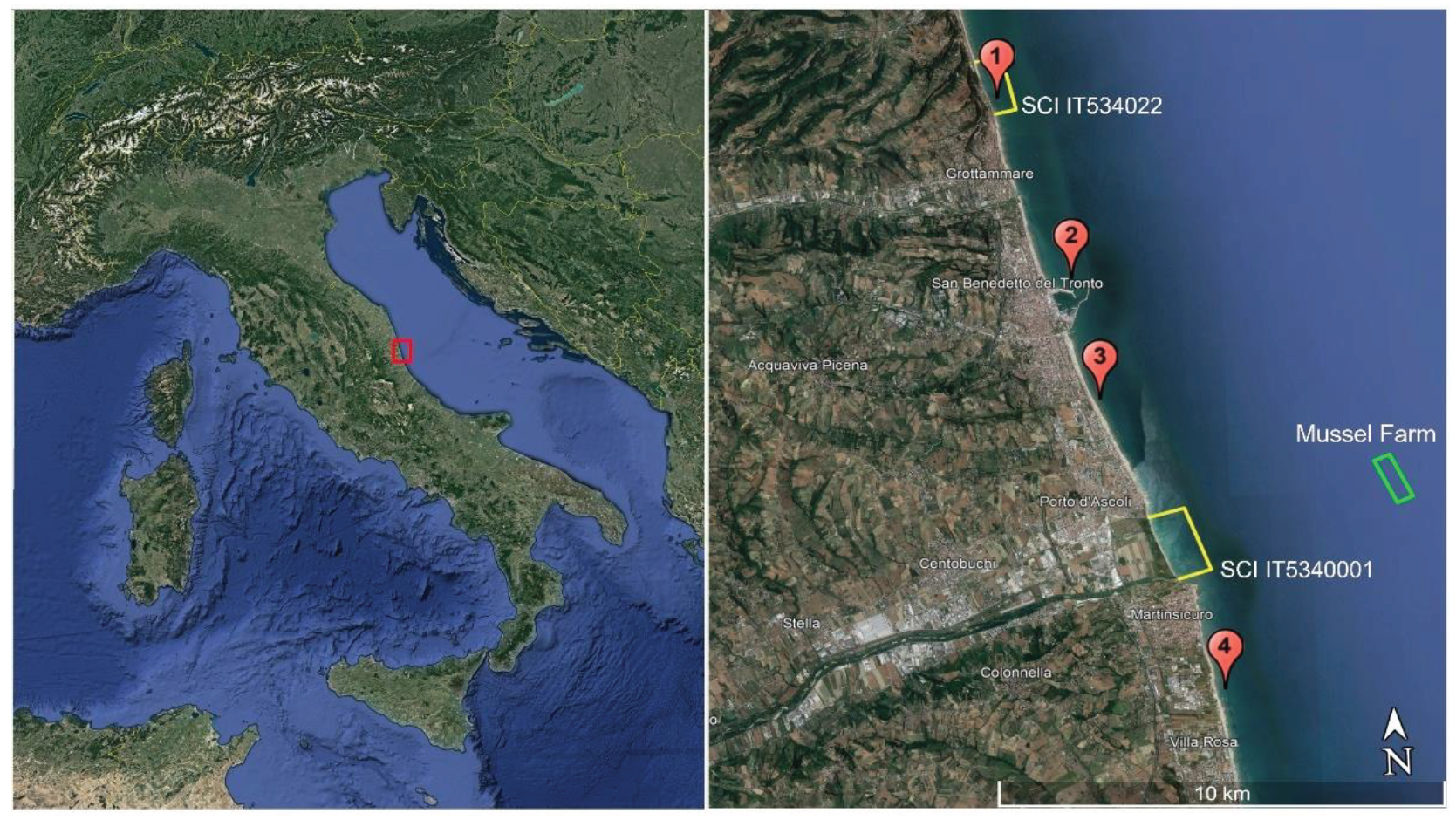
Figure 2.
MHWs occurred in the study area in the period 2011 – 2022. The bars represent the MHWs recorded for each year in the months of June, July and August; in red the MHWs in which the temperature exceeded 30°C. No MHWs were detected in 2013, 2016, 2020 and 2021.
Figure 2.
MHWs occurred in the study area in the period 2011 – 2022. The bars represent the MHWs recorded for each year in the months of June, July and August; in red the MHWs in which the temperature exceeded 30°C. No MHWs were detected in 2013, 2016, 2020 and 2021.
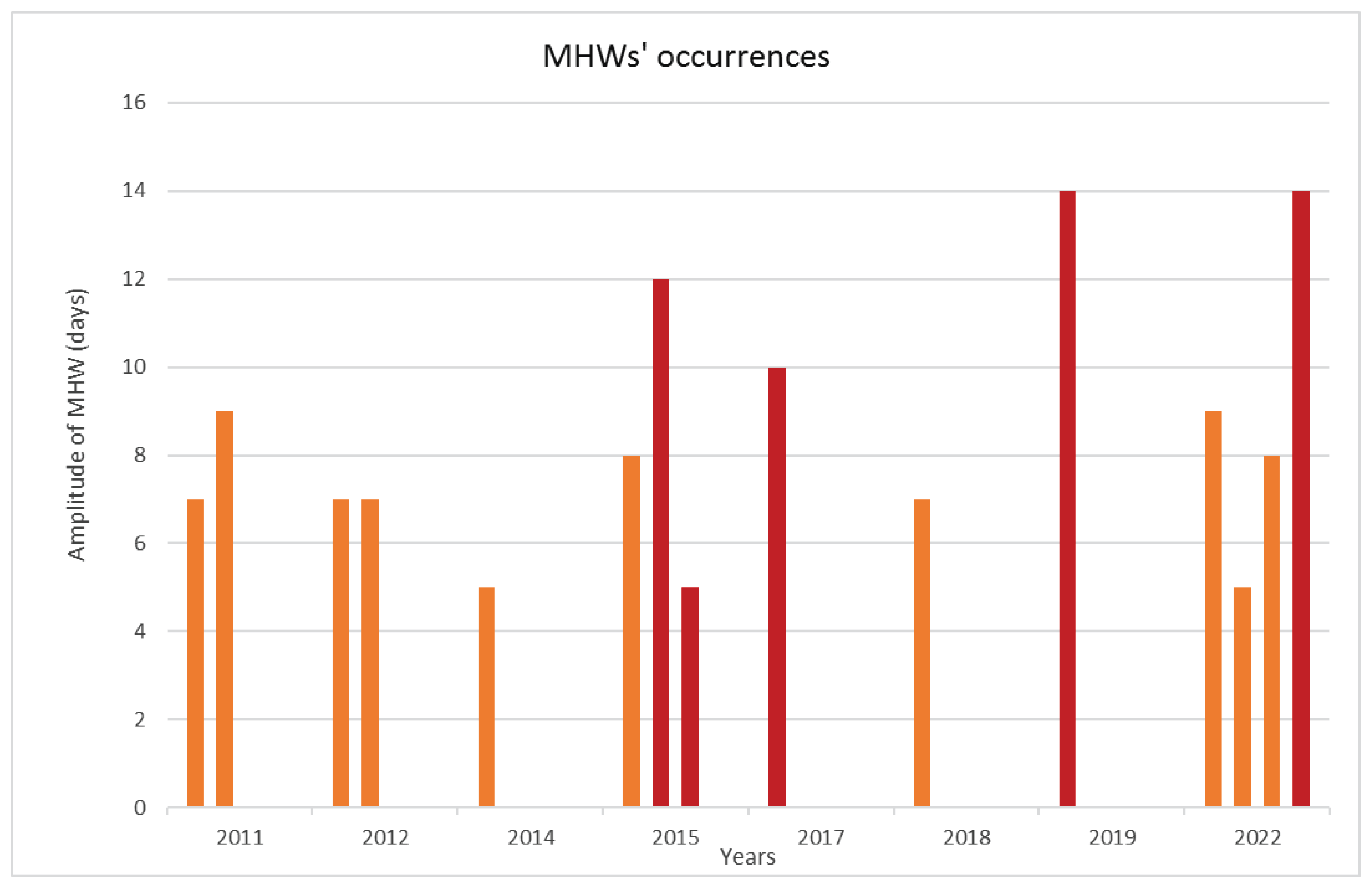
Figure 3.
Pre (A) and post (B) summer 2022 rocky substrate covered by mussel beds in Site 1.

Figure 4.
Surviving specimens of M. galloprovincialis detected in September 2022 in the subtidal (A) and intertidal (B) zone of Site 1.
Figure 4.
Surviving specimens of M. galloprovincialis detected in September 2022 in the subtidal (A) and intertidal (B) zone of Site 1.

Figure 5.
Comparison of river flow between the year 2022 and the two previous periods, at the Pontelagoscuro (FE) station (data source [60]).
Figure 5.
Comparison of river flow between the year 2022 and the two previous periods, at the Pontelagoscuro (FE) station (data source [60]).
Figure 6.
Effects of illegal mussel removal resulting in localized holes in the dense cover of mussels (photo of Grottammare Site1, June 2020); note the difference compared to Figure 3, photo B, in which the absence of mussels is total.
Figure 6.
Effects of illegal mussel removal resulting in localized holes in the dense cover of mussels (photo of Grottammare Site1, June 2020); note the difference compared to Figure 3, photo B, in which the absence of mussels is total.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



