
Submitted:
08 September 2023
Posted:
12 September 2023
You are already at the latest version
Abstract
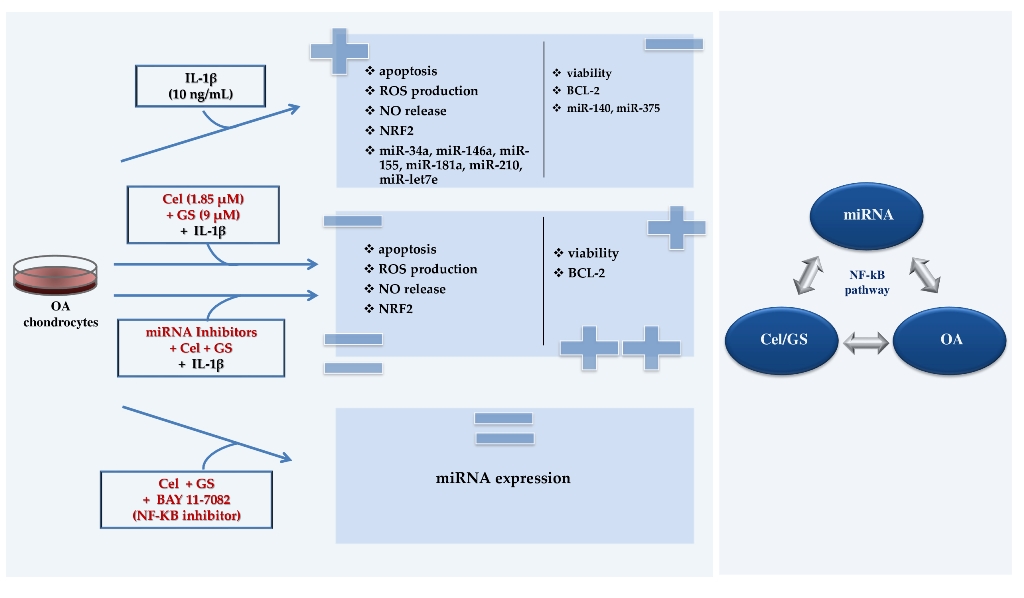
This study investigated the role of a pattern of microRNA (miRNA) as possible mediators of celecoxib and prescription-grade glucosamine sulfate (GS) effects in human osteoarthritis (OA) chondrocytes.
Chondrocytes were treated with celecoxib (1.85 µM) and GS (9 µM), alone or in combination, for 24 hrs, with or without interleukin (IL)-1β (10 ng/mL). Viability was detected by MTT assay, apoptosis and reactive oxygen species (ROS) by cytometry, nitric oxide (NO) by Griess method. Gene levels of miRNA, antioxidant enzymes, nuclear factor erythroid (NRF)2, and B-cell lym-phoma (BCL)2 expressions were analyzed by quantitative real time PCR. Protein expression of NRF2 and BCL2 was also detected at immunofluorescence and western blot.
Celecoxib and GS, alone or in combination, significantly increased viability, reduced apoptosis, ROS and NO production and the gene expression of miR-34a, -146a, -181a, -210, in comparison to baseline and to IL-1β. The transfection with miRNA specific inhibitors significantly counteracted the IL-1β activity and potentiated the properties of celecoxib and GS on viability, apoptosis and oxidant system, through NF-κB regulation. The observed effects were enhanced when the drugs were tested in combination.
Our data confirmed the synergistic anti-inflammatory and chondroprotective properties of celecoxib and GS, suggesting miRNA as possible mediators.
Keywords:
microRNA
; celecoxib
; glucosamine sulfate
; chondrocytes
; osteoarthritis
; NF-κB
; inflammation
; chondroprotection
1. Introduction
Non-coding RNAs constitute a large portion of molecules that act as epigenetic regulators of gene and protein expressions, and thus participate in fundamental biological processes [1]. Among them, in the last time, microRNA (miRNA) have received extensive attention for their role in the pathogenesis of various pathological disorders, including cancer, diabetes, cardiovascular, neurological and musculoskeletal diseases. Growing evidence underscore the implication of some miRNA in the development and progression of osteoarthritis (OA) [2,3,4,5,6]. Microarray analysis has identified different miRNA expression profile between normal and OA cartilage samples [7,8]; the evaluation of miRNA in synovial fluid, in serum, or in plasma has showed a significantly difference in patients with OA in comparison to healthy controls [9,10]. An aberrant expression of miRNA can influence chondrocytes phenotype and function, induce extracellular matrix degradation, apoptosis and oxidative stress, playing a pivotal role in OA pathogenesis [11,12,13,14,15]. Additionally, several miRNA affecting the expression of inflammatory mediators, such as the Interleukin-1β (IL-1β), which stimulates synovial inflammation, bone and cartilage destruction, characteristic features of OA disease [16,17].
To date, there isn’t an effective treatment for OA, the current strategies for its management have relied on a multimodal approach with a combination of pharmacological and/or non-pharmacological therapies focused on reducing pain and improving physical function [18]. Among the pharmacological options, high-quality prescription-grade glucosamine sulfate (GS), a symptomatic slow-acting drugs for osteoarthritis (SYSADOAs), has been approved from the ESCEO working group for the treatment of OA due to its symptomatic and disease-modifying properties [19].
GS has shown to be effective in the regulation of chondrocytes and synoviocytes metabolism and in the reduction of IL-1β-induced negative activities [20,21,22,23,24].
Oral non-steroidal anti-inflammatory drugs (NSAIDs), selective and non-selective, are recommended to treat severe pain and inflammation in OA [19]. Celecoxib, a selective cyclooxygenase (COX)-2 inhibitor (COXIB), is one of the most frequently used symptomatic drug in clinical practice for its low toxicity, especially at the gastrointestinal and cardiovascular levels [19].
Accumulating evidence reported that celecoxib, besides its analgesic and anti-inflammatory properties, had also disease-modifying activities [25,26]. Indeed, different studies showed its regulation of cell metabolism, apoptosis, and oxidative stress in human OA chondrocytes, fibroblast-like synoviocytes, and subchondral bone osteoblasts [27,28,29,30,31,32]. Recently, it has been found the ability of celecoxib to restore the altered plasma expression of miR-155 and miR-146a in patients with knee OA [33], as well as the transcriptional levels of miR-29 and miR-34a in human gastric cancer cells and osteosarcoma lines [34,35].
In a previous report, we demonstrated, for the first time, the synergistic chondroprotective role of celecoxib and prescription-grade GS on human OA chondrocytes; this combination treatment exerted a protective role against the detrimental activities induced by IL-1β, mainly reducing inflammation and apoptosis, and regulating oxidant/antioxidant balance and cartilage turnover, via nuclear factor (NF)-κB pathway [36].
As an extension of these preliminary data, the present study aimed at investigating the potential regulatory effect of the celecoxib and GS, alone or in combination, on the expression profile of a group of miRNA (miR-34a, miR-140, miR-146a, miR-155, miR-181a, miR-210, and miR-375), associated to OA pathogenesis, in human OA chondrocytes exposed to IL-1β. Moreover, we assessed the possible implication of miRNA in mediating celecoxib and GS-induced effects on viability, apoptosis and oxidant system.
2. Results
2.1. Celecoxib and GS modulate apoptosis and oxidant/antioxidant system
In Figure 1 are confirmed the effects induced by celecoxib (1.85 µM) and GS (9 µM), alone or in combination, for 24 hrs, on the regulation of apoptosis ratio, oxidant/antioxidant system, nitric oxide (NO) release and viability, in OA chondrocytes stimulated or not with IL-1β.
The results showed that the treatment of the cells with celecoxib or GS alone didn’t cause any significant modification on apoptosis, redox balance and survival in comparison to control cultures (CTRL). The incubation of chondrocytes with celecoxib and GS in combination significantly reduced the apoptotic rate, up-regulated the gene expression of the anti-apoptotic marker B-cell lymphoma 2 (BCL2), limited reactive oxygen species (ROS) production and the mRNA levels of the transcriptional factor nuclear factor erythroid 2 (NRF2), while increased viability (p < 0.05), with respect to CTRL (Figure 1A-G). Conversely, IL-1β significantly induced the activation of apoptosis and oxidative stress, while reduced BCL2 levels and the percentage of survival (p < 0.01, p < 0.001); the negative trend of IL-1β was significantly counteracted by the pre-treatment of chondrocytes with celecoxib or GS, especially when used in combination (p < 0.05, p < 0.01, p < 0.001) (Figure 1A-G).
To confirm the beneficial synergistic effect of the studied drugs on apoptosis and oxidant/antioxidant system, an immunofluorescence and a western blot analysis have been performed on the anti-apoptotic marker BCL2 and on the transcriptional factor NRF2 (Figure 2, Figure 3).
Figure 2A demonstrated that, at CTRL condition, the BCL2 signal was localized in the cytoplasm, and it appeared weak and diffuse in 48% of cells, while it appeared strong only in the 8% of chondrocytes; after the IL-1β stimulus, the strong localization was present in a percentage of 68 % of cells and sometimes it appeared dotted (Figure 2B). The treatment of chondrocytes with celecoxib and GS in combination reduced the presence of cells strongly positive to BCL2 protein in comparison to the only IL-1β stimulus (30%) (Figure 2D).
The image reported in Figure 2E-H showed the different localization of NRF2 protein in the studied experimental conditions, less evident compared BCL2 protein. A diffuse weak or absent cytoplasmic distribution was detected at CTRL in the majority of cells (79%) in which a clear signal was present in the 21% (Figure 2E). The incubation of chondrocytes with IL-1β determined an increase of number of cells with a strong localization (70%) (Figure 2F) that was reduced by the pre-treatment with celecoxib and GS in combination (35%) (Figure 2H).
No significant different localization of both antibodies were observed when the cells were incubated with the studied drugs, alone or in combination, compared to CTRL or to the stimulus of IL-1β (BCL2 Figure 2C: NFR2 Figure 2G).
A similar trend was obtained on the densitometric quantification of the bands assessed at western blot analysis (Figure 3A-D). The results demonstrated that the stimulus of chondrocytes with IL-1β induced a significant increase of BCL2 and NRF2 protein expression (p < 0.05) in comparison to CTRL. Celecoxib and GS tested in combination significantly reduced the expression of BCL2 (p <0.05) with respect to control condition, while no modifications on NRF2 were found. More interestingly, the combined effect of the studied drugs was able to counteract, in a significant manner, the negative activity of IL-1β on these proteins.
2.2. MiRNA profile modulation following celecoxib and GS treatment
Figure 4 showed the ability of celecoxib and GS to regulate the expression profile of a pattern of miRNA, involved in the pathogenesis of OA, modified by IL-1β.
The combination of the studied pharmacological compounds induced a significant reduction of miR-34a, miR-146a, miR-181a, and miR-210 gene expression (p < 0.05), while no changes in miR-140, miR-155, miR-375, and miR-let-7e levels were found, with respect to CTRL (Figure 4A-G). As expected, IL-1β stimulus significantly up-regulated the transcriptional levels of miR-34a, miR-146a, miR-155, miR-181a, miR-210, and miR-let-7e (p < 0.01), and decreased those of miR-140 and miR-375 (p < 0.01).
Interestingly, the pre-treatment of the cells with celecoxib or GS alone or in combination significantly limited the expression of miR-34a, miR-146a, mir-181a, and miR-210 induced by IL-1β, in comparison to the incubation with the cytokine alone, with a particular enhancement when the drugs were tested simultaneously (p < 0.05, p < 0.01, p < 0.001) (Figure 4A,C,E,F). Otherwise, miR-140, miR-155, miR-375, and miR-let-7e profiles appeared not to be influenced by the treatment with the drugs alone or in combination (Figure 4B,D,G,H).
To better understand the direct regulation of celecoxib and GS on miR-34a, miR-146a, miR-181a, and miR-210 expression profile, chondrocytes were transiently transfected with miRNA specific inhibitors for 24 hrs before the standard treatment with the drugs (Figure 5).
Figure 5A corroborated the ability of the inhibitors to significantly reduce the gene expression of miR-34a, miR-146a, miR-181a, and miR-210 (p < 0.01, p < 0.001) in comparison to CTRL cultures and negative control (NC). After miRNA silencing, the cells exposed to IL-1β showed a significant down-regulation of miR-34a, miR-146a, miR-181a, and miR-210 compared to the stimulus with IL-1β alone (p < 0.05) (Figure 5B–E). Interestingly, the effects induced by IL-1β resulted strongly neutralized by the concomitant exposure of the cells to miR-34a, miR-146a, and miR-210 inhibitors and the drugs, in comparison to their relative NC (p < 0.05, p < 0.01) (Figure 5B,C,E). On the other hand, celecoxib and GS activities seem to be not dependent by the presence of miR-181a inhibitor (Figure 5D).
2.3. MiRNA mediate the effect induced by celecoxib and GS
To verify the potential role of miRNA as mediators of celecoxib and GS alone or in combination effects on viability, apoptosis and oxidant system, our cultures were simultaneously incubated with miR-34a, miR-146a, miR-181a, and miR-210 inhibitors and treated with the pharmacological compounds.
Our results showed that the silencing of chondrocytes with miR-34a, miR-146a, miR-181a, and miR-210 inhibitors induced a significant reduction of apoptosis (p < 0.05, p < 0.01), an increase of viability (p < 0.05) while limited the production of ROS and NO (p < 0.05, p < 0.01), with respect to CTRL cultures and NC (Figure S1A-E, Figure 6A-H, Figure 7A-H, Figure 8A-D).
The stimulus with IL-1β caused opposite and detrimental effects increasing apoptosis and oxidative stress, which was significantly reduced by the pre-treatment of the cells with celecoxib and GS, or transiently transfected with miRNA specific inhibitors (p < 0.05, p < 0.01) (Figure 6A-H, Figure 7A-H, Figure 8A-D).
Interestingly, the co-incubation of the cells with miR-34a, miR-146a, miR-210 inhibitors and the pharmacological compounds significantly reinforced their inhibitory effect against IL-1β on the studied processes (p < 0.05, p < 0.01), than those induced by each single treatment (Figure 6A,B,D-F,H, Figure 7A,B,D-F,H, Figure 8A,B,D). Also in this case, celecoxib and GS didn’t alter miR-181a inhibitor activities on apoptosis and oxidative stress (Figure 6 C, G, Figure 7C,G, Figure 8C).
2.4. NF-κB involvement in drugs-induced effects on miRNA
On the basis of the obtained results and the direct interaction between celecoxib and GS with NF-κB found in our previous study, we hypothesized miRNA as possible mediators of drugs’ effects, via NF-κB. At this regard, our cultures were pre-incubated with a specific NF-κB inhibitor (BAY 11-7082, IKKα/β) and, then, treated with the studied drugs and IL-1β (Figure 9).
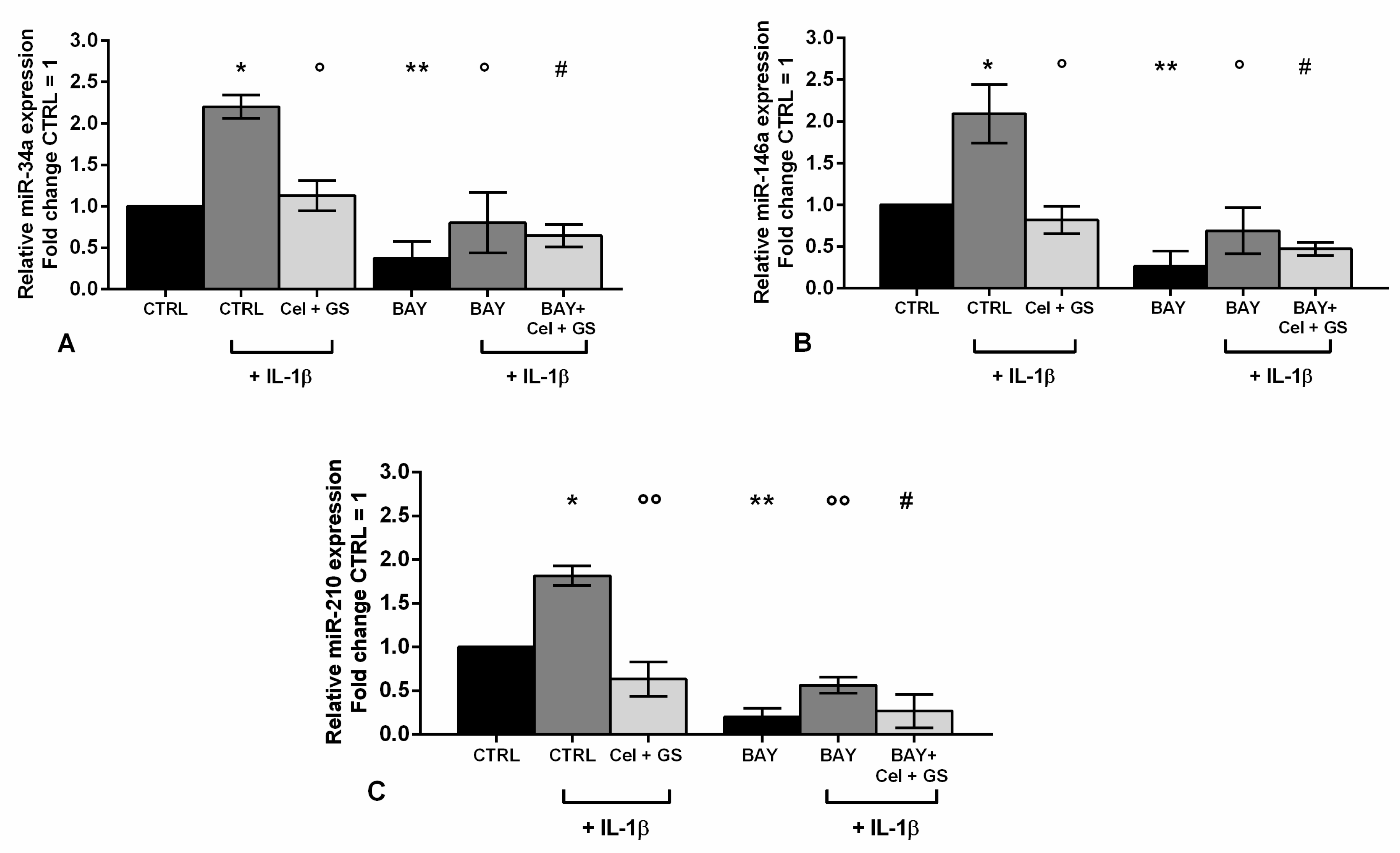
An analysis at real time PCR was performed on miR-34a, miR-146a and miR-210, which resulted already modulated by celecoxib and GS, to explore their modulation in presence of NF-κB inhibitor.
The data reported in Figure 9 showed a significant decrease in gene expression of miR-34a, miR-146a and miR-210 after incubation of the cells with BAY 11-7082 with respect to CTRL and, in particular, to IL-1β stimulus (p < 0.01) (Figure 9A-C). Very interestingly, miRNA up-regulation induced by IL-1β negative effect was significantly limited by the pre-treatment of the cells with BAY 11-7082 plus celecoxib and GS (p < 0.05), in a more effective way beyond that caused by BAY 11-7082 alone or by the treatment with the only drugs (Figure 9A-C).
3. Discussion
This study aimed at deeper investigating the molecular mechanism underlying the beneficial properties of celecoxib and prescription-grade GS on human OA chondrocytes, on the basis of the preliminary data obtained in our previous research [36]. In particular, we analyzed the regulatory effects of the two drugs, alone or in combination, on apoptosis, viability, and oxidant system, as well as on the expression profile of a pattern of miRNA involved in OA pathophysiology. In addition, we hypothesized miRNA as possible mediators of celecoxib and GS-induced effects.
The doses of celecoxib and GS used to carried out our experiments represent the most suitable concentrations to reproduce in vivo condition [37] and were selected in agreement with our prior data [36]; the treatment was applied for a period of 24 h on the basis of the best results in terms of viability.
This study confirmed the favorable effects of celecoxib and GS on apoptosis, viability and redox balance, according to our preliminary report [36]. These processes are well known as important hallmarks associated to OA unset and progression [38,39]. Indeed, during the pathogenesis of the disease, inflammatory mediators, such as cytokines, can alter mitochondrial structure and function causing the failure in oxidant/antioxidant system, leading to an excessive ROS generation. This condition contributes to activate cartilage degradation and synovial inflammation, as well as the apoptosis signaling [38,39,40]; the latter is controlled and regulated through a balance between the members of the BCL2 family characterized by proteins with pro-apoptotic roles and others, such as BCL2, which generally driving anti-apoptotic functions [41].
Our first data showed an increased production of mitochondrial, extracellular ROS species and of NO, and an up-regulation of the transcriptional factor NRF2, in OA chondrocytes exposed to IL-1β; also, this negative stimulus raised the ratio of apoptotic cells, reduced the percentage of viability, with a concomitant decreased gene expression of the anti-apoptotic marker BCL2. These data are consistent with other evidence from the literature conducted on chondrocyte cultures [32,36,42,43]. Surprisingly, our immunofluorescence and western blot assays showed opposite findings on BCL2 protein expression, after IL-1β stimulus, than those found at its mRNA levels. The apparent discrepancy among the results can be attributed to the complex interplay between the anti-apoptotic and pro-apoptotic members of BCL2 family during the apoptosis process. Indeed, it has been demonstrated that BCL2 exerts its role of anti-apoptotic protein by binding some pro-apoptotic molecules to inhibit the apoptosis signaling; once the binding among the proteins is dissociated, BCL2 can be phosphorylated in the cytoplasm and degraded by different kind of stimuli [41,44,45]. This can explain, in our opinion, the increased levels of BCL2 protein at the immunofluorescence analysis, since, for our experiments, we used an anti-BCL2 antibody not able to discriminate the active non-phosphorylated BCL2 protein from the inactivate phosphorylated form destined for degradation.
Furthermore, we corroborated the ability of celecoxib and GS, tested alone or in combination, to limit the negative effect of IL-1β, decreasing ROS and NO release, NRF2 expression, and cell death process, according to previous findings. In fact, different authors reported a reduction of ROS and NO production and antioxidant enzymes expression, induced by IL-1β, after treatment of human OA cartilage explants or chondrocytes with celecoxib and/or GS [32,36,46,47,48]. Similarly, both drugs limited the apoptosis rate and increased the anti-apoptotic marker expression, in IL-1β-stimulated OA cells [24,32,36,48].
These results confirm the anti-apoptotic and antioxidant activities of celecoxib and GS, in particular when used in combination. However, their exact molecular mechanism and, thus, the beneficial properties of these drugs still remains not completely defined.
In the second part of the study, we verified the hypothesis that miRNA could represent possible mediator factors by which celecoxib and GS exert their effects on apoptotic process and oxidant system.
Accumulating evidence proved that some of the most important miRNA associated to OA pathogenesis are strong regulators of apoptosis and oxidative stress signaling [12,13,49]. In particular, miR-34a and miR-181a were found to increase cell death and reduce cell viability, as well as to modulate the production of ROS and NO, in different cell types [12,13,49,50,51,52]. Also miR-146a has been recently recognized as an activator of these processes by its direct targeting on SMAD4 and NRF2 transcriptional factors, in human OA chondrocytes and synoviocytes [13,53,54]. In addition, miR-210 was found to be associated to OA due to its higher levels in synovial fluid of patients with knee OA compared to healthy individuals [55]. Furthermore, miR-210 contribute to chondrogenic differentiation, DNA damage response, and apoptosis in bone marrow mesenchymal stem cells and synovial fibroblasts [56,57].
In the present research, we showed the over-expression of miR-34a, miR-146a, miR-181a, and miR-210 after the stimulus of OA chondrocytes with IL-1β, consistently with the results derived from other in vitro studies [58,59,60]. Interestingly, we also observed that the pre-incubation of our cells with celecoxib and GS significantly reduced the expression of the studied miRNA, counteracting the negative effect induced by IL-1β. This is the first report revealing the ability of these drugs to regulate miR-34a, miR-146a, miR-181a, and miR-210 profiles in human OA chondrocytes, and, especially, highlighting their potentiated effect when used simultaneously. Previous studies confirmed the relevance of celecoxib in controlling the gene expression of some miRNA involved in the progression of human malignancies has been documented. In particular, Chen et al. [61], performing an analysis of miRNA expression profile in human colorectal cancer cells, reported that celecoxib limited the aberrant expression of 28 miRNA correlated to clinical stage of cancer, lymph node involvement and metastasis. More recently, other authors, through a miRNA microarray analysis on gastric cancer cells, proved the clinical relevance of celecoxib for the treatment of gastric cancer, fine-tuning the tumor suppressor miR-29 [34].
To attest the direct effect of celecoxib and GS on the studied miRNA, we carried out additional experiments of miRNA silencing. In fact, the transient transfection of our OA chondrocytes with miR-34a, miR-146a, miR-181a, and miR-210 inhibitors significantly reduced the apoptotic process, ROS generation and NO production, and increase the percentage of viability, in particular preventing their activation induced by IL-1β. In a similar manner, previous researches on OA chondrocyte and synoviocytes employing miRNA inhibitors attested a direct modulation of miR-34a, miR-146a and miR-181a on cell proliferation and apoptosis by targeting SIRT-1/p53 and on BCL2 signaling pathways [13,62]. The inhibition of these miRNA also limited the release of ROS species and the production of NO in different cell cultures, probably affecting NRF2 proteins and NF-kB pathway [13,49,63,64].
Intriguingly, we firstly demonstrated that the influence of miR-34a, miR-146a, and miR-210 silencing on apoptosis and redox balance was strongly reinforced in presence of celecoxib and GS, particularly when the drugs were used in combination. In support of our data, Chen and co-authors [63] demonstrated the synergistic, additive effect of celecoxib and miR-34a in the regulation of cell viability, cell migration and invasion in osteosarcoma cell lines.
As the last step of our research, we have tried to find out the regulatory network underlying the interaction between celecoxib and GS and miRNA, assuming that a key role could be played by NF-κB signaling pathway. Indeed, a number of studies, even our previous one, demonstrated that these pharmacological compounds exert a protective role on cartilage metabolism and inflammation through the regulation of NF-κB proteins [21,25,36]. In fact, evidence from cancer cell lines and human chondrocytes demonstrated the ability of celecoxib in controlling apoptosis and oxidative stress processes through a direct effect on p50 and p65 subunits of NF-kB pathway [36,65,66]. GS exerts its role on NF-κB via an epigenetic mechanism, regulating the demethylation of specific CpG sites of DNA [67,68], responsible for the expression of redox- and apoptosis-related factors, in human articular chondrocytes [36,67,69]. In addition, a number of in vitro evidence highlighted a direct effect between some miRNA, including miR-34a, miR-146a, and miR-181a, and NF-κB signaling, which reflects a consequent modulation of the downstream genes controlled by the pathway [13,70,71].
According to the current literature, in our previous report we confirmed the direct combined effect of celecoxib and GS on NF-κB pathway in OA chondrocytes, demonstrating the capacity of the drugs to reduce the expression of p50 and p65 subunits [36]. Very interestingly, in the present paper, for the first time, we showed that the inhibition of NF-κB signaling in our cultures, induced a reduction of the expression profile of miR-34a, miR-146a, and miR-210. Intriguingly, this trend resulted more favorable and strengthen in the presence of celecoxib and GS in combination.
On the other hand, celecoxib and GS properties seem to be not influenced by miR-181a, indeed, no detectable changes on apoptosis or redox balance were observed in our results following miR-181a silencing. To the best of our knowledge, there is no evidence from the literature explaining this aspect, so it’s difficult to draw any conclusion in this sense; therefore, it’s reasonable to assume that miR-181a and celecoxib and GS act through two independent mechanisms on cell death and ROS modulation.
4. Materials and Methods
4.1. Primary cultures of human OA chondrocytes
Human OA articular cartilage was obtained from femoral heads of five patients with hip OA according to ACR criteria [72], subjected to total arthroplasty. The donors were two men and three women with age ranging from 61 to 76. The samples were provided by the Orthopaedic Section from the University of Siena (Italy), and OA grades was defined by Mankin score [73]. Authorization at using human specimens was permitted by the Ethic Committee of Azienda Ospedaliera Universitaria Senese (decision no. 13931/18), after receiving the signed informed consent from the donor.
Chondrocytes were isolated immediately after surgery. Cartilage slices were aseptically dissected from the femoral heads and processed by a sequential enzymatic digestion using trypsin and type IV collagenase, as previously described [13]. Then cells were grown in Dulbecco’s Modified Eagle Medium (DMEM) (Euroclone, Italy) with phenol red and L-glutamine, supplemented with 10% fetal bovine serum (FBS) (Euroclone, Italy), and 200 U/mL penicillin and 200 µg/mL streptomycin (P/S) (Euroclone, Italy). After a test of viability with Tripan Blue (Sigma-Aldrich, Italy), primary cells at the first passage were employed for the experiments.
4.2. Treatment procedure
Human OA chondrocytes were plated in 6-well dishes at a starting density of 1 × 105 cells/well and cultured in DMEM containing 10% FBS and 2% P/S, until reaching about 85% of confluence.
Prescription-grade crystalline glucosamine sulfate (Dona®) and celecoxib (Celebrex®) were provided by Meda Pharma SpA (Viatris group). The powders were reconstituted in phosphate-buffered saline (PBS) (Euroclone, Italy) filtered and diluted in the culture medium (DMEM with 0.5% FBS and 2% P/S) to reach the final concentrations selected for the experiments. Celecoxib and GS were tested at the concentration of 1.85 µM and 9 µM, respectively, for 24 h, in agreement with previous studies [36,37,68]. The treatment was conducted in presence or not of IL-1β (10 ng/mL) (Sigma-Aldrich, Italy), added 2 h later than the incubation of the cells with celecoxib or GS, or their combination.
For the transient transfection procedure, chondrocytes were incubated with miR-34a, miR-146a, miR-181a, and miR-210 specific inhibitors (50 nM) (Qiagen, Germany), or with the respective negative controls siRNA (NC) (5 nM) (Qiagen, Germany), for 24 hrs, according to the kit instructions. Afterwards miRNA silencing, the standard treatment with celecoxib or/and GS in presence or not of IL-1β was applied.
Timing of treatments were chosen on the basis of the best results obtained in terms of viability (data not shown) and according to previous studies [36,68,74].
Finally, some cultures were processed for 2 h with a specific inhibitor of NF-κB kinase subunit alpha, named IKKα/β (BAY 11-7082, 1 μM) (Sigma–Aldrich, Italy), according to the manufacturer’s instructions, and, then, incubated for 24 h with celecoxib and GS in presence of IL-1β.
4.3. MTT assay
The viability of the cells was evaluated by MTT assay and expressed as the percentage of survival. The procedure has been performed according to previously established method [74]. The percentage of survival cells was measured as (absorbance of the sample) / (absorbance of control) × 100 by a microplate reader spectrophotometer at 570 nm. The obtained data were reported as optical density units per 104 adherent cells.
4.4. Apoptosis labeling and reactive oxygen species assessment
The detection of apoptosis was assessed through a commercial kit provided with annexin-V and propidium iodide (PI) probes (ThermoFisher Scientific, Italy). After treatment, the cells were harvested, collected into cytometry tubes, and centrifuged. The pellet was resuspended in a working solution of annexin-V and PI, according to the manufacturer’s instructions, and incubated at room temperature for 15 min in the dark.
The evaluation of mitochondrial superoxide anion and of intracellular ROS production was carried out by using a commercial kit of MitoSOX Red and 2′,7′-Dichlorodihydrofluorescein diacetate (DCFH-DA) probes (ThermoFisher Scientific, Italy). After treatment, the cells were incubated for 30 min at 37°C in the dark with a solution of MitoSOX Red, according to the instructions, and, then, collected into cytometry tubes and centrifuged. The pellet was resuspended in saline solution before the analysis at flow cytometry.
A total of 10,000 cells per assay was measured by the instrument both for apoptosis and ROS assessment. For the detection of apoptotic cells, the analysis was carried out measuring cells simultaneously stained and positive to each dye, and the results were reported as percentage of total apoptosis; for the production of superoxide anion the results were expressed as median fluorescence.
4.5. Nitric oxide detection
The release of NO in the culture medium of the cells was measured by using the Griess method. One hundred μl of supernatant from each experimental condition was transferred in a microplate with 100 μl of Griess reagent (1% of sulfanilamide, 0.1% of N-1-naphthylethylenediamide dihydrochloride in 5% of H3PO4) and were incubated at room temperature for 15 minutes. The absorbance was measured at 550 nm using a microplate reader. The obtained results were normalized with the number of the cells and the NO concentration was reported as ng/106 cells.
4.6. Quantitative real-time PCR
After treatment, the cells were harvested and total RNA was extracted using TriPure Isolation Reagent (Euroclone, Italy), according to the manufacturer’s instructions. The concentration, purity, and integrity of RNA were evaluated by Nanodrop-1000 (Celbio, Italy).
For the reverse-transcription of target genes and miRNA, 500 ng of RNA were processed into cDNA using specific commercial kits (Qiagen, Germany), and, then, employed for real-time PCR using specific commercial kits for SYBR Green assay (Qiagen, Germany). The primers used for PCR procedure were reported in Table S1.
All qPCR reactions were executed in glass capillaries by a LightCycler 1.0 instrument (Roche Molecular Biochemicals, Germany) using LightCycler Software Version 3.5.
For the data analysis, the Ct values of each sample were calculated and converted into relative quantities. The normalization was performed using housekeeping genes, Actin Beta (ACTB) for target genes and Small Nucleolar RNA, C/D Box 25 (SNORD-25) for miRNA.
4.7. Immunofluorescence determination
For this determination, cells were grown in specific and sterile coverslips mounted in multi-wells at a starting low density of 4 × 104 cells/chamber, to avoid any possible achievement of confluence or overlapping. Chondrocytes were treated for 24 h with celecoxib and GS in combination, in presence or not of IL-1β. After that, cells were fixed in 4% paraformaldehyde (ThermoFisher Scientific, Italy) for 15 minutes, and permeabilized with a blocking solution contained PBS, 1% bovine serum albumin (BSA) (Sigma–Aldrich, Italy) and 0.2% Triton X-100 (ThermoFisher Scientific, Italy) for 30 minutes. After these steps, the cells were incubated overnight, at 4 °C, with solutions of mouse monoclonal anti-BCL2 and anti-NRF2 primary antibodies (Santa Cruz Biotechnology, Italy), followed by 1 hr incubation with goat anti-mouse IgG-Texas Red conjugated antibody (Southern Biotechnology, Italy). After antibodies incubations, the coverslips were washed and a nuclear colorant was added before mounting in specific slides.
Fluorescence was examined with a Leitz Aristo plan fluorescence microscope and the epifluorescence was analyzed at 200× and 400× magnification. About 100 cells for each experimental conditions were considered. The fluorescent signal was evaluated as fair, medium, or strong label [75].
4.8. Western blot analysis
For protein extraction, cell pellets were suspended in ice-cold lysis buffer [50 mM Tris-HCl (pH 7.5), 50 mM EDTA (pH 8), 150 mM NaCl, 1% NP40, 2 mM Na3VO4, 10 mM NaF, 0.3 mM PMSF, and a protease inhibitor cocktail (Cat# 87785, Thermo Fisher Scientific, USA)]. Protein concentrations were determined using the Bradford Method (Cat# S-B6916, Sigma-Aldrich, Italy). 10 μg of proteins were loaded onto a 10% sodium dodecyl sulfate-polyacrylamide gel and then transferred onto a nitrocellulose membrane (Cat# 1620115, Bio-rad, CA, USA). Primary antibodies anti- BCL2 (sc-7382 Mouse Monoclonal antibody, Santa Cruz Biotechnology, Italy), anti-NRF2 (sc-365949 Mouse Monoclonal antibody, Santa Cruz Biotechnology, Italy) and β-actin (sc-47778 Mouse Monoclonal antibody, Santa Cruz Biotechnology, Italy) were incubated overnight at 4 °C. Membranes were washed with TBS with 0.1% Tween-20 and incubated with horseradish peroxidase-conjugated secondary antibodies for 1 hour at room temperature. Then, membranes were washed before chemiluminescence detection using Clarity ECL reagents (Cat #1705061, Bio-Rad, CA, USA) and the images were acquired with Chemidoc MP (Bio-Rad, CA, USA). Image Lab software (Bio-Rad, CA, USA) was used to quantify the images of the bands. Results were normalized with the relative loading control.
4.9. Statistical analysis
Three independent experiments were carried out and the results were expressed as the mean ± standard deviation of triplicate values for each experiment. Data normal distribution was evaluated by Shapiro–Wilk, D’Agostino and Pearson, and Kolmogorov–Smirnov tests. Statistical analysis was performed using analysis of variance followed by the Bonferroni multiple comparison test. All analyses were carried out through the SAS System (SAS Institute Inc., Cary, NC, USA) and GraphPad Prism 6.1. A p-value <0.05 will be defined as statistically significant.
5. Conclusions
In the present study, we confirmed, first of all, the regulatory synergistic effects of celecoxib and prescription-grade GS on apoptosis process and oxidant system, in human OA chondrocyte cultures.
In addition, we proved, for the first time, the ability of the studied compounds to modulate the expression profile of some miRNA, known to be implicated in apoptosis and oxidative stress processes, and, therefore, in OA pathophysiology.
Finally, we identified miR-34a, miR-146a, and miR-210 as possible mediators of celecoxib and GS-induced positive effects, via NF-κB pathway, providing additional information about their molecular mechanism. Furthermore, these data support the potential role of miRNA as therapeutic targets for the treatment of OA [76].
This study confirms the synergistic anti-inflammatory and chondroprotective effects of celecoxib and GS, as previously demonstrated in our in vitro experience, and support the therapeutic use of this combination in the multimodal approach for patient with OA [77].
Anyway, further studies are required to better comprehend this complex network, such as the identification of the direct targets of miRNA, controlling apoptosis and oxidative stress signaling, which may help to elucidate the downstream cascade triggered by celecoxib and GS.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1; Table S1: List of primers for quantitative real time PCR.
Author Contributions
Conceptualization, S.C., and A.F.; Laboratory Investigation, S.C, G.C, M. B., E.M.; R. C., M. B.; Methodology, S.C. and G.C., M.B.; Supervision, A.F.; Validation, N.V.; Writing—original draft, S.C., S.C. and A.F.; Writing—review and editing, S.C., N.V., and A.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was approved by the Ethic Committee of Azienda Ospedaliera Universitaria Senese (decision no. 13931/18).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
We would like to express our sincere gratitude to our supervisor, Professor Antonio Giordano, from Center for Biotechnology, Sbarro Institute for Cancer Research and Molecular Medicine, College of Science and Technology, Temple University, Philadelphia, PA 19122, USA, for his valuable guidance and support throughout the research process.
Conflicts of Interest
The Authors declare no conflicts of interest.
References
- Matsuyama, H.; Suzuki, H.I. Systems and Synthetic microRNA Biology: From Biogenesis to Disease Pathogenesis. Int. J. Mol. Sci. 2020, 21, 132–155. [Google Scholar] [CrossRef] [PubMed]
- De Palma, A.; Cheleschi, S.; Pascarelli, N.A.; Tenti, S.; Galeazzi, M.; Fioravanti, A. Do microRNAs have a key epigenetic role in osteoarthritis and in mechanotransduction? Clin. Exp. Rheumatol. 2017, 35, 518–526. [Google Scholar] [PubMed]
- Ratneswaran, A.; Kapoor, M. Osteoarthritis year in review: genetics, genomics, epigenetics. Osteoarthritis Cartilage. 2021, 29, 151–160. [Google Scholar] [CrossRef]
- Li, J.; Gao, X.; Zhu, W.; Li, X. Integrative Analysis of the Expression of microRNA, Long Noncoding RNA, and mRNA in Osteoarthritis and Construction of a Competing Endogenous Network. Biochem. Genet. 2022, 1141–1158. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Rao, W.; Huo, S.; Fan, T.; Qiu, M.; Zhu, H.; Chen, D.; Sheng, X. MicroRNAs and long non-coding RNAs in cartilage homeostasis and osteoarthritis. Front. Cell Dev. Biol. 2022, 10, 1092776. [Google Scholar] [CrossRef] [PubMed]
- Iulian Stanciugelu, S.; Homorogan, C.; Selaru, C.; Patrascu, J.M.; Patrascu, J.M. JR; Stoica, R.; Nitusca, D.; Marian, C. Osteoarthritis and microRNAs: Do They Provide Novel Insights into the Pathophysiology of This Degenerative Disorder? Life (Basel). 2022, 12, 1914. [Google Scholar] [CrossRef]
- Díaz-Prado, S.; Cicione, C.; Muiños-López, E.; Hermida-Gómez, T.; Oreiro, N.; Fernández-López, C.; Blanco, F.J. Characterization of microRNA expression profiles in normal and osteoarthritic human chondrocytes. BMC Musculoskelet. Disord. 2012, 13, 144. [Google Scholar] [CrossRef]
- Balaskas, P.; Goljanek-Whysall, K.; Clegg, P.D.; Fang, Y.; Cremers, A.; Smagul, A.; Welting, T.J.M.; Peffers, M.J. MicroRNA Signatures in Cartilage Ageing and Osteoarthritis. Biomedicines. 2023, 11, 1189. [Google Scholar] [CrossRef]
- Ali, S.A.; Gandhi, R.; Potla, P.; Keshavarzi, S.; Espin-Garcia, O.; Shestopaloff, K.; Pastrello, C.; Bethune-Waddell, D.; Lively, S.; Perruccio, A.V. , et al. Sequencing identifies a distinct signature of circulating microRNAs in early radiographic knee osteoarthritis. Osteoarthritis Cartilage. 2020, 28, 1471–1481. [Google Scholar] [CrossRef]
- Baloun, J.; Pekáčová, A.; Švec, X.; Kropáčková, T.; Horvathová, V.; Hulejová, H.; Prajzlerová, K.; Růžičková, O.; Šléglová, O.; Gatterová, J.; et al. Circulating miRNAs in hand osteoarthritis. Osteoarthritis Cartilage. 2023, 31, 228–237. [Google Scholar] [CrossRef]
- Prasadam, I.; Batra, J.; Perry, S.; Gu, W.; Crawford, R.; Xiao, Y. Systematic Identification, Characterization and Target Gene Analysis of microRNAs Involved in Osteoarthritis Subchondral Bone Pathogenesis. Calcif. Tissue Int. 2016, 99, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Cheleschi, S.; De Palma, A.; Pecorelli, A.; Pascarelli, N.A.; Valacchi, G.; Belmonte, G.; Carta, S.; Galeazzi, M.; Fioravanti, A. Hydrostatic Pressure Regulates MicroRNA Expression Levels in Osteoarthritic Chondrocyte Cultures via the Wnt/β-Catenin Pathway. Int. J. Mol. Sci. 2017, 18, 133. [Google Scholar] [CrossRef] [PubMed]
- Cheleschi, S.; Gallo, I.; Barbarino, M.; Giannotti, S.; Mondanelli, N.; Giordano, A.; Tenti, S.; Fioravanti, A. MicroRNA Mediate Visfatin and Resistin Induction of Oxidative Stress in Human Osteoarthritic Synovial Fibroblasts Via NF-κB Pathway. Int. J. Mol. Sci. 2019, 20, 5200. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Poulet, C.; Malaise, M.; Abak, A.; Mahmud Hussen, B.; Taheriazam, A.; Taheri, M.; Hallajnejad, M. The Emerging Role of Non-Coding RNAs in Osteoarthritis. Front. Immunol. 2021, 12, 773171. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.A.; Peffers, M.J.; Ormseth, M.J.; Jurisica, I.; Kapoor, M. The non-coding RNA interactome in joint health and disease. Nat. Rev. Rheumatol. 2021, 17, 692–705. [Google Scholar] [CrossRef] [PubMed]
- Chow, Y.Y.; Chin, K.Y. The Role of Inflammation in the Pathogenesis of Osteoarthritis. Mediators Inflamm. 2020, 8293921. [Google Scholar] [CrossRef]
- Yao, Q.; Wu, X.; Tao, C.; Gong, W.; Chen, M.; Qu, M.; Zhong, Y.; He, T.; Chen, S.; Xiao, G. Osteoarthritis: pathogenic signaling pathways and therapeutic targets. Signal Transduct. Target Ther. 2023, 8, 56. [Google Scholar] [CrossRef]
- Veronese, N.; Cooper, C.; Bruyère, O; Al-Daghri, N.M.; · Branco, J.; Cavalier, E.; Cheleschi, S.; da Silva Rosa, M.C.; Conaghan, P.G.; Dennison, E.M.; et al. Multimodal Multidisciplinary Management of Patients with Moderate to Severe Pain in Knee Osteoarthritis: A Need to Meet Patient Expectations. Drugs 2022, 82, 1347–1355. [Google Scholar] [CrossRef]
- Bruyère, O.; Honvo, G.; Veronese, N.; Arden, N.K.; Branco, J.; Curtis, E.M.; Al-Daghri, N.M.; Herrero-Beaumont, G.; Martel-Pelletier, J.; Pelletier, J.P.; et al. An updated algorithm recommendation for the management of knee osteoarthritis from the European Society for Clinical and Economic Aspects of Osteoporosis, Osteoarthritis and Musculoskeletal Diseases (ESCEO). Semin. Arthritis Rheum. 2019, 49, 337–350. [Google Scholar] [CrossRef]
- Bassleer, C.; Rovati, L.; Franchimont, P. Stimulation of proteoglycan production by glucosamine sulfate in chondrocytes isolated from human osteoarthritic articular cartilage in vitro. Osteoarthritis Cartilage. 1998, 6, 427–34. [Google Scholar] [CrossRef]
- Largo, R.; Alvarez-Soria, M.A.; Díez-Ortego, I.; Calvo, E.; Sánchez-Pernaute, O.; Egido, J.; Herrero-Beaumont, G. Glucosamine inhibits IL-1beta-induced NFkappaB activation in human osteoarthritic chondrocytes. Osteoarthritis Cartilage. 2003, 11, 290–298. [Google Scholar] [CrossRef]
- Chan, P.S.; Caron, J.P.; Rosa, G.J.; Orth, M.W. Glucosamine and chondroitin sulfate regulate gene expression and synthesis of nitric oxide and prostaglandin E(2) in articular cartilage explants. Osteoarthritis Cartilage. 2005, 13, 387–394. [Google Scholar] [CrossRef]
- Chiusaroli, R.; Piepoli, T.; Zanelli, T.; Ballanti, P.; Lanza, M.; Rovati, L.C.; Caselli, G. Experimental pharmacology of glucosamine sulfate. Int. J. Rheumatol. 2011, 2011, 939265. [Google Scholar] [CrossRef]
- Luo, M.; Xu, F.; Wang Q, Luo W. The inhibiting effect of glucosamine sulfate combined with loxoprofen sodium on chondrocyte apoptosis in rats with knee osteoarthritis. J. Musculoskelet. Neuronal. Interact. 2021, 21, 113–120. [Google Scholar]
- Zweers, M.C.; de Boer, T.N.; van Roon, J.; Bijlsma, J.W.; Lafeber, F.P.; Mastbergen, S.C. Celecoxib: Considerations regarding its potential disease-modifying properties in osteoarthritis. Arthritis Res. Ther 1186, 13, 239. [Google Scholar] [CrossRef]
- Nakata, K.; Hanai, T.; Take, Y.; Osada, T.; Tsuchiya, T.; Shima, D.; Fujimoto, Y. Disease-modifying effects of COX-2 selective inhibitors and non-selective NSAIDs in osteoarthritis: A systematic review. Osteoarthritis Cartilage 2018, 26, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- Cha, H.S.; Ahn, K.S.; Jeon, C.H.; Kim, J.; Koh, E.M. Inhibitory effect of cyclo-oxygenase-2 inhibitor on the production of matrix metalloproteinases in rheumatoid fibroblast-like synoviocytes. Rheumatol. Int. 2004, 24, 207–11. [Google Scholar] [CrossRef] [PubMed]
- Mastbergen, S.C.; Jansen, N.W.; Bijlsma, J.W.; Lafeber, F.P. Differential direct effects of cyclo-oxygenase-1/2 inhibition on proteoglycan turnover of human osteoarthritic cartilage: An in vitro study. Arthritis Res. Ther. 2006, 8, 1–9. [Google Scholar] [CrossRef]
- Tat, S.K.; Pelletier, J.P.; Lajeunesse, D.; Fahmi, H.; Duval, N.; Martel-Pelletier, J. Differential modulation of RANKL isoforms by human osteoarthritic subchondral bone osteoblasts: Influence of osteotropic factors. Bone. 2008, 43, 284–291. [Google Scholar] [CrossRef] [PubMed]
- de Boer, T.N.; Huisman, A.M.; Polak, A.A.; Niehoff, A.G.; van Rinsum, A.C.; Saris, D.; Bijlsma, J.W.; Lafeber, F.J.; Mastbergen, S.C. The chondroprotective effect of selective COX-2 inhibition in osteoarthritis: Ex vivo evaluation of human cartilage tissue afterin vivo treatment. Osteoarthritis Cartilage. 2009, 17, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Fioravanti, A.; Tinti, L.; Pascarelli, N.A.; Di Capua, A.; Lamboglia, A.; Cappelli, A.; Biava, M.; Giordani, A.; Niccolini, S.; Galeazzi, M.; Anzini, M. In Vitro effects of VA441, a new selective cyclooxygenase-2 inhibitor, on human osteoarthritic chondrocytes exposed to IL-1β. J. Pharmacol. Sci. 2012, 120, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Cheleschi, S.; Calamia, V.; Fernandez-Moreno, M.; Biava, M.; Giordani, A.; Fioravanti, A.; Anzini, M.; Blanco, F. In vitro comprehensive analysis of VA692 a new chemical entity for the treatment of osteoarthritis. Int. Immunopharmacol. 2018, 64, 86–100. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Jiang, H.; Jian, X.; Zhang, W. Change of miRNA expression profiles in patients with knee osteoarthritis before and after celecoxib treatment. J. Clin. Lab. Anal. 2019, 33, e22648. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Suzuki, H.; Imaeda, H.; Matsuzaki, J.; Hirata, K.; Tsugawa, H.; Hibino, S.; Kanai, Y.; Saito, H.; Hibi, T. The tumor suppressor microRNA-29c is downregulated and restored by celecoxib in human gastric cancer cells. Int. J. Cancer. 2013, 132, 1751–1760. [Google Scholar] [CrossRef]
- Chen, X.; Peng, D.; Shen, Y.; Liu, B.; Zhou, H.; Tao, H.; Huang, J. The potential combinational effect of miR-34a with celecoxib in osteosarcoma. Anticancer Drugs. 2017, 28, 888–897. [Google Scholar] [CrossRef]
- Cheleschi, S.; Tenti, S.; Giannotti, S.; Veronese, N.; Reginster, J.Y.; Fioravanti, A. A Combination of Celecoxib and Glucosamine Sulfate Has Anti-Inflammatory and Chondroprotective Effects: Results from an In Vitro Study on Human Osteoarthritic Chondrocytes. Int. J. Mol. Sci. 2021, 22, 8980. [Google Scholar] [CrossRef]
- Walter, M.F.; Jacob, R.F.; Day, C.A.; Dahlborg, R.; Weng, Y.; Mason, R.P. Sulfone COX-2 inhibitors increase susceptibility of human LDL and plasma to oxidative modification: comparison to sulfonamide COX-2 inhibitors and NSAIDs. Atherosclerosis. 2004, 177, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhang, Z.; Sheng, P.; Mobasheri, A. The role of metabolism in chondrocyte dysfunction and the progression of osteoarthritis. Ageing Res. Rev. 2021, 66, 101249. [Google Scholar] [CrossRef]
- Tudorachi, N.B.; Totu, E.E.; Fifere, A.; Ardeleanu, V.; Mocanu, V.; Mircea, C.; Isildak, I.; Smilkov, K.; Cărăuşu, E.M. The implication of reactive oxygen species and antioxidants in knee osteoarthritis. Antioxidants (Basel). 2021, 10, 985. [Google Scholar] [CrossRef]
- Coaccioli, S.; Sarzi-Puttini, P.; Zis, P.; Rinonapoli, G.; Varrassi, G. Osteoarthritis: New insight on its pathophysiology. J. Clin. Med. 2022, 11, 6013. [Google Scholar] [CrossRef]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: the balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Héraud, F.; Héraud, A.; Harmand, M.F. Apoptosis in normal and osteoarthritic human articular cartilage. Ann. Rheum. Dis. 2000, 59, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.J.; Lu, J.W.; Lee, C.H. , Lee, H.S.; Chu, Y.H.; Ho, Y.J.; Liu, F.C.; Huang, C.J.; Wu, C.C.; Wang, C.C. Cardamonin Attenuates Inflammation and Oxidative Stress in Interleukin-1β-Stimulated Osteoarthritis Chondrocyte through the Nrf2 Pathway. Antioxidants (Basel). 2021, 10, 862. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Yao, C.; Liu, Y.; Yuan, J.; Wu, L.; Hosoi, K.; Yu, S.; Huang, C.; Wei, H. , Chen, G. Arsenic trioxide induces expression of BCL-2 expression via NF-κB and p38 MAPK signaling pathways in BEAS-2B cells during apoptosis. Ecotoxicol. Environ. Saf. 2021, 222, 112531. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Matsuda, K.; Matsusaka, K.; Nakajima, M.; Takeno, Y.; Miyazaki, T.; Shintaku, T.; Yoda, N.; Saito, T.; Ikeda, E.; et al. Anti-proliferating and apoptosis-inducing activity of chemical compound FTI-6D in association with p53 in human cancer cell lines. Chem. Biol. Interact. 2023, 369, 110257. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Soria, M.A.; Herrero-Beaumont, G.; Moreno-Rubio, J.; Calvo, E.; Santillana, J.; Egido, J.; Largo, R. Long-term NSAID treatment directly decreases COX-2 and mPGES-1 production in the articular cartilage of patients with osteoarthritis. Osteoarthritis Cartilage. 2008, 16, 1484–1493. [Google Scholar] [CrossRef] [PubMed]
- Valvason, C.; Musacchio, E.; Pozzuoli, A.; Ramonda, R.; Aldegheri, R.; Punzi, L. Influence of glucosamine sulphate on oxidative stress in human osteoarthritic chondrocytes: effects on HO-1, p22(Phox) and iNOS expression. Rheumatology (Oxford). 2008, 47, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Tan, C.; An, H.; Jiang, D.; Quan, Z.; Tang, K.; Luo, X. Selective COX-2 inhibitor ameliorates osteoarthritis by repressing apoptosis of chondrocyte. Med. Sci. Monit. 2012, 18, BR247–52. [Google Scholar] [CrossRef]
- Zhong, X.; Li, P.; Li, J.; He, R.; Cheng, G.; Li, Y. Downregulation of microRNA-34a inhibits oxidized low-density lipoprotein-induced apoptosis and oxidative stress in human umbilical vein endothelial cells. Int. J. Mol. Med. 2018, 42, 1134–1144. [Google Scholar] [CrossRef]
- Feng, X.; Zhang, C.; Yang, Y.; Hou, D.; Zhu. Role of miR-181a in the process of apoptosis of multiple malignant tumors: A literature review. Adv. Clin. Exp. Med. 2018, 27, 263–270. [Google Scholar] [CrossRef]
- Cheng, D.L.; Fang, H.X.; Liang, Y.; Zhao, Y.; Shi, C.S. MicroRNA-34a promotes iNOS secretion from pulmonary macrophages in septic suckling rats through activating STAT3 pathway. Biomed. Pharmacother. 2018, 105, 1276–1282. [Google Scholar] [CrossRef] [PubMed]
- Endisha, H.; Datta, P.; Sharma, A.; Nakamura, S.; Rossomacha, E.; Younan, C. MicroRNA-34a-5p Promotes Joint Destruction During Osteoarthritis. Arthritis Rheumatol. 2021, 73, 426–439. [Google Scholar] [CrossRef] [PubMed]
- Cheleschi, S.; Barbarino, M.; Gallo, I.; Tenti, S.; Bottaro, M.; Frati, E.; Giannotti, S.; Fioravanti, A. Hydrostatic Pressure Regulates Oxidative Stress through microRNA in Human Osteoarthritic Chondrocytes. Int. J. Mol. Sci. 2020, 21, 3653. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.N.; Lu, S.; Fu, C.M. MiR-146a expression profiles in osteoarthritis in different tissue sources: a meta-analysis of observational studies. J. Orthop. Surg. Res. 2022, 17, 148. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Su, W.; Xia, H.; Wang, Z.; Su, C.; Su, B. Synovial Fluid MicroRNA-210 as a Potential Biomarker for Early Prediction of Osteoarthritis. Biomed. Res. Int. 2019, 2019, 7165406. [Google Scholar] [CrossRef]
- Liu, S.C.; Chuang, S.M.; Hsu, C.J.; Tsai, C.H.; Wang, S.W.; Tang, C.H. CTGF increases vascular endothelial growth factor-dependent angiogenesis in human synovial fibroblasts by increasing miR-210 expression. Cell Death Dis. 2014, 5, e1485. [Google Scholar] [CrossRef]
- Yang, M.; Yan, X.; Yuan, F.Z.; Ye, J.; Du, M.Z.; Mao, Z.M.; Xu, B.B.; Chen, Y.R.; Song, Y.F.; Fan, B.S.; Yu, J.K. MicroRNA-210-3p Promotes Chondrogenic Differentiation and Inhibits Adipogenic Differentiation Correlated with HIF-3α Signalling in Bone Marrow Mesenchymal Stem Cells Biomed. Res. Int. 2021, 2021, 6699910. [Google Scholar] [CrossRef]
- Kim, D.; Song, J.; Kim, S.; Chun, C.H.; Jin, E.J. MicroRNA-34a regulates migration of chondroblast and IL-1β-induced degeneration of chondrocytes by targeting EphA5. Biochem. Biophys Res. Commun. 2011, 415, 551–557. [Google Scholar] [CrossRef]
- Wang, W.; Wang, Z.; Chen, S.; Zang, X.; Miao, J. Interleukin-1β/nuclear factor-κB signaling promotes osteosarcoma cell growth through the microRNA-181b/phosphatase and tensin homolog axis. J. Cell Biochem. 2019, 20, 1763–1772. [Google Scholar] [CrossRef]
- Zhao, G.; Gu, W. Effects of miR-146a-5p on chondrocyte interleukin-1β-induced inflammation and apoptosis involving thioredoxin interacting protein regulation. J. Int. Med. Res. 2020, 48, 300060520969550. [Google Scholar] [CrossRef]
- Chen, W.C.; Lin, M.S.; Ye, Y.L.; Gao, H.J.; Song, Z.Y.; Shen, X.Y. microRNA expression pattern and its alteration following celecoxib intervention in human colorectal cancer. Exp. Ther. Med. 2012, 3, 1039–1048. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Wang, M.; Zhao, J.; Zhang, H.; Zhou, C.; Jin, L.; Zhang, Y.; Qiu, X.; Ma, B.; Fan, Q. MicroRNA-34a affects chondrocyte apoptosis and proliferation by targeting the SIRT1/p53 signaling pathway during the pathogenesis of osteoarthritis. Int. J. Mol. Med. 2016, 38, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.L.; Fu, Y.Y.; Shi, M.Y.; Li, H.X. Down-regulation of miR-181a can reduce heat stress damage in PBMCs of Holstein cows. In Vitro Cell. Dev. Biol. Anim. 2016, 52, 864–871. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Chu, A.; Feng, Y.; Chen, L.; Shao, Y.; Luo, Q.; Deng, X.; Wu, M.; Shi, X.; Chen, Y. A MicroRNA-146a: A Comprehensive Indicator of Inflammation and Oxidative Stress Status Induced in the Brain of Chronic T2DM Rats. Front. Pharmacol. 2018, 9, 478. [Google Scholar] [CrossRef]
- Cai, F,; Chen, M.; Zha, D.; Zhang, P.; Zhang, X.; Cao, N.; Wang, J.; He, Y.; Fan, X.; Zhang, W; et al. Curcumol potentiates celecoxib-induced growth inhibition and apoptosis in human non-small cell lung cancer. Oncotarget. 2017, 8, 115526-115545. [CrossRef]
- Tudor, D.V.; Bâldea, I.; Olteanu, D.E.; Fischer-Fodor, E.; Piroska, V.; Lupu, M.; Călinici, T.; Decea, R.M.; Filip, G.A. Celecoxib as a valuable adjuvant in cutaneous melanoma treated with trametinib. Int. J. Mol. Sci. 2021, 22, 4387. [Google Scholar] [CrossRef]
- Imagawa, K.; de Andrés, M.C.; Hashimoto, K.; Pitt, D.; Itoi, E.; Goldring, M.B.; Roach, H.I.; Oreffo, R.O. The epigenetic effect of glucosamine and a nuclear factor-kappa B (NF-kB) inhibitor on primary human chondrocytes–implications for osteoarthritis. Biochem. Biophys. Res. Commun. 2011, 405, 362–367. [Google Scholar] [CrossRef]
- Kucharz, E.J.; Kovalenko, V.; Szántó, S.; Bruyère, O.; Cooper, C.; Reginster, J.Y. A review of glucosamine for knee osteoarthritis: why patented crystalline glucosamine sulfate should be differentiated from other glucosamines to maximize clinical outcomes. Curr. Med. Res. Opin. 2016, 32, 997–1004. [Google Scholar] [CrossRef]
- Calamia, V.; Mateos, J.; Fernández-Puente, P.; Lourido, L.; Rocha, B.; Fernández-Costa, C.; Montell, E.; Vergés, J.; Ruiz-Romero, C.; Blanco, F. J Pharmacoproteomic study of the effects of chondroitin and glucosamine sulfate on human articular chondrocytes. Arthritis Res Ther. 2010, 12, R138. [Google Scholar] [CrossRef]
- Qi, J.; Qiao, Y.; Wang, P.; Li, S.; Zhao, W.; Gao, C. microRNA-210 negatively regulates LPS-induced production of proinflammatory cytokines by targeting NF-κB1 in murine macrophages. FEBS Lett. 2012, 586, 1201–1207. [Google Scholar] [CrossRef]
- Zhou, B.; Li, H.; Shi, J. miR-27 inhibits the NF-κB signaling pathway by targeting leptin in osteoarthritic chondrocytes. Int. J. Mol. Med. 2017, 40, 523–530. [Google Scholar] [CrossRef]
- Altman, R.; Alarcón, G.; Appelrouth, D.; Bloch, D.; Borenstein, D.; Brandt, K.; Brown, C.; Cooke, T.D.; Daniel, W.; Feldman, D.; et al. The American College of Rheumatology criteria for the classification and reporting of osteoarthritis of the hip. Arthritis Rheum. 1991, 34, 505–514. [Google Scholar] [CrossRef]
- Mankin, H.J.; Dorfman, H.; Lippiello, L.; Zarins, A. Biochemical and metabolic abnormalities in articular cartilage from osteoarthritic human hips. II. Correlation of morphology with biochemical and metabolic data. J. Bone Joint Surg. Am. 1971, 53, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Cheleschi, S.; Tenti, S.; Lorenzini, S.; Seccafico, I.; Barbagli, S.; Frati, E.; Fioravanti, A. Synovial Fluid Regulates the Gene Expression of a Pattern of microRNA via the NF-κB Pathway: An In Vitro Study on Human Osteoarthritic Chondrocytes. Int. J. Mol. Sci. 2022, 23, 8334. [Google Scholar] [CrossRef] [PubMed]
- Pascarelli, N.A.; Moretti, E.; Terzuoli, G.; Lamboglia, A.; Renieri, T.; Fioravanti, A.; Collodel, G. Effects of gold and silver nanoparticles in cultured human osteoarthritic chondrocytes. J. Appl. Toxicol. 2013, 33, 1506–1513. [Google Scholar] [CrossRef] [PubMed]
- Ho, P.T.B.; Clark, I.M.; and Le, L.T.T. MicroRNA-Based Diagnosis and Therapy. Int. J. Mol. Sci. 2022, 23, 7167. [Google Scholar] [CrossRef]
- Veronese, N.; Ecarnot, F.; Cheleschi, S.; Fioravanti, A.; Maggi, S. Possible synergic action of non-steroidal anti-inflammatory drugs and glucosamine sulfate for the treatment of knee osteoarthritis: a scoping review. BMC Musculoskelet. Disord. 2022, 23, 1084. [Google Scholar] [CrossRef]
Figure 1.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of treatment with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence or not of interleukin (IL)-1β (10 ng/ml). (A, C, E) Apoptosis detection and reactive oxygen species (ROS) production by flow cytometry. (B,D) Expression levels of B-cell lymphoma 2 (BCL2) and nuclear factor erythroid 2 (NRF2) by quantitative real time PCR. (F) Nitric oxide (NO) release by Griess method. (G) Viability by MTT assay. All the results, except for NO and viability, were expressed as fold change of the value of interest respect to CTRL, reported equal to 1. Data were represented as mean ± standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001 versus CTRL. °p < 0.05, °°p < 0.01, °°°p < 0.001 versus IL-1β.
Figure 1.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of treatment with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence or not of interleukin (IL)-1β (10 ng/ml). (A, C, E) Apoptosis detection and reactive oxygen species (ROS) production by flow cytometry. (B,D) Expression levels of B-cell lymphoma 2 (BCL2) and nuclear factor erythroid 2 (NRF2) by quantitative real time PCR. (F) Nitric oxide (NO) release by Griess method. (G) Viability by MTT assay. All the results, except for NO and viability, were expressed as fold change of the value of interest respect to CTRL, reported equal to 1. Data were represented as mean ± standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001 versus CTRL. °p < 0.05, °°p < 0.01, °°°p < 0.001 versus IL-1β.
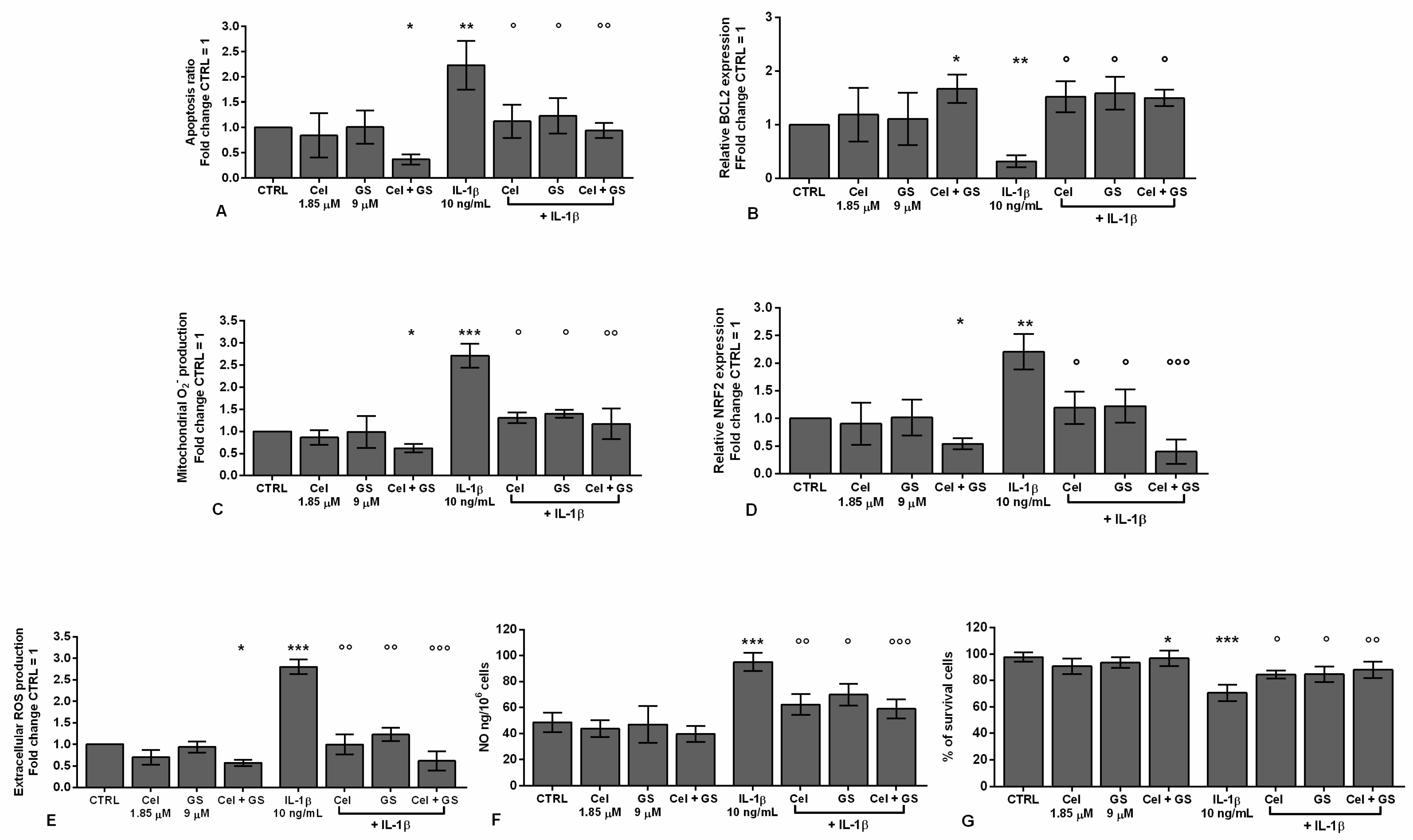
Figure 2.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of treatment with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence or not of interleukin (IL)-1β (10 ng/ml). (A-H) Indirect immunofluorescence microscopy of cells incubated with monoclonal anti- B-cell lymphoma 2 (BCL2) (A-D) and anti-nuclear factor erythroid 2 (NRF2) (E-H) primary antibodies. (A,E) CTRL: a fair fluorescence in the cytoplasm is shown; (B,F) IL-1β: an intense signal is evident in the cytoplasm; (C,G) Cel + GS; the label is almost absent. (D,H) Cel + GS + IL-1β: the signal is diffused in the cytoplasm but reduced with respect to IL-1β. Nuclei (blue) were stained with DAPI. Bars: A-H 50 µm.
Figure 2.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of treatment with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence or not of interleukin (IL)-1β (10 ng/ml). (A-H) Indirect immunofluorescence microscopy of cells incubated with monoclonal anti- B-cell lymphoma 2 (BCL2) (A-D) and anti-nuclear factor erythroid 2 (NRF2) (E-H) primary antibodies. (A,E) CTRL: a fair fluorescence in the cytoplasm is shown; (B,F) IL-1β: an intense signal is evident in the cytoplasm; (C,G) Cel + GS; the label is almost absent. (D,H) Cel + GS + IL-1β: the signal is diffused in the cytoplasm but reduced with respect to IL-1β. Nuclei (blue) were stained with DAPI. Bars: A-H 50 µm.
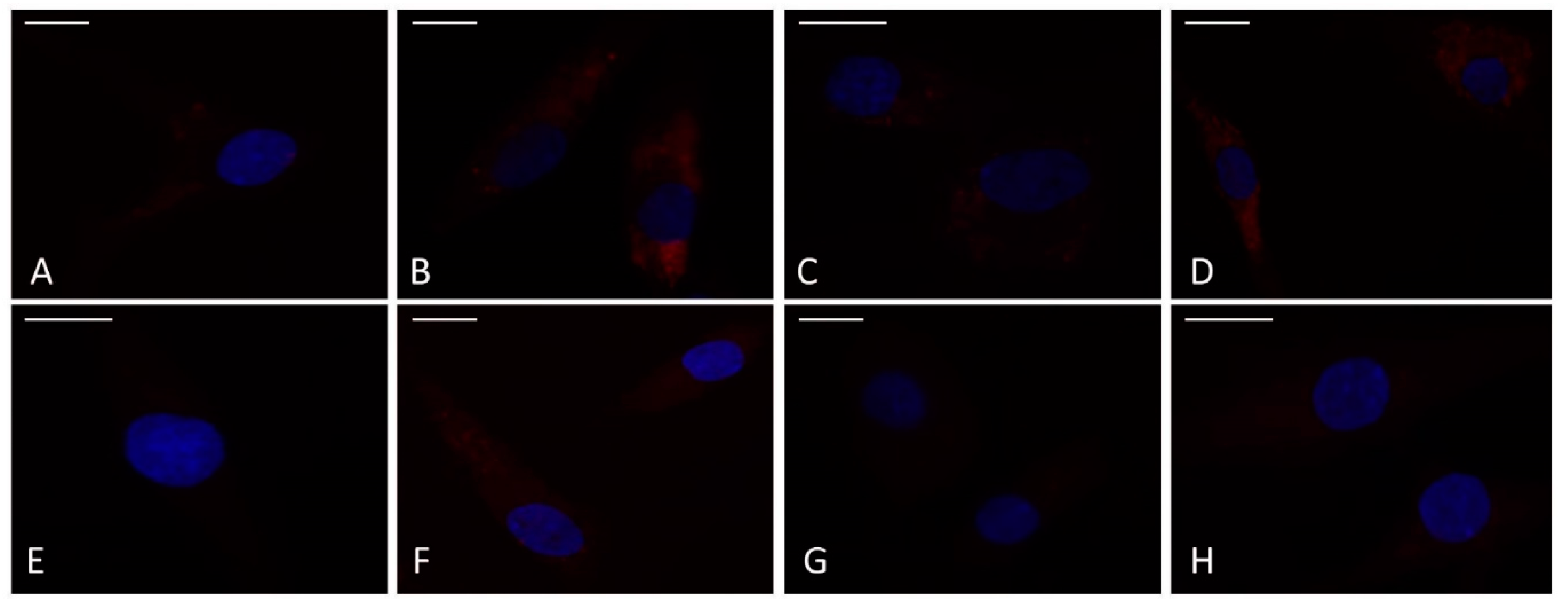
Figure 3.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of treatment with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence or not of interleukin (IL)-1β (10 ng/ml). (A-D) Representative immunoblotting image and densitometric analysis of B-cell lymphoma 2 (BCL2) and nuclear factor erythroid 2 (NRF2) protein levels by western blot. The results were expressed as fold change of the value of interest respect to CTRL, reported equal to 1. Data were represented as mean ± standard deviation. *p < 0.05 versus basal CTRL. °p < 0.05, °°p < 0.01 versus IL-1β.
Figure 3.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of treatment with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence or not of interleukin (IL)-1β (10 ng/ml). (A-D) Representative immunoblotting image and densitometric analysis of B-cell lymphoma 2 (BCL2) and nuclear factor erythroid 2 (NRF2) protein levels by western blot. The results were expressed as fold change of the value of interest respect to CTRL, reported equal to 1. Data were represented as mean ± standard deviation. *p < 0.05 versus basal CTRL. °p < 0.05, °°p < 0.01 versus IL-1β.
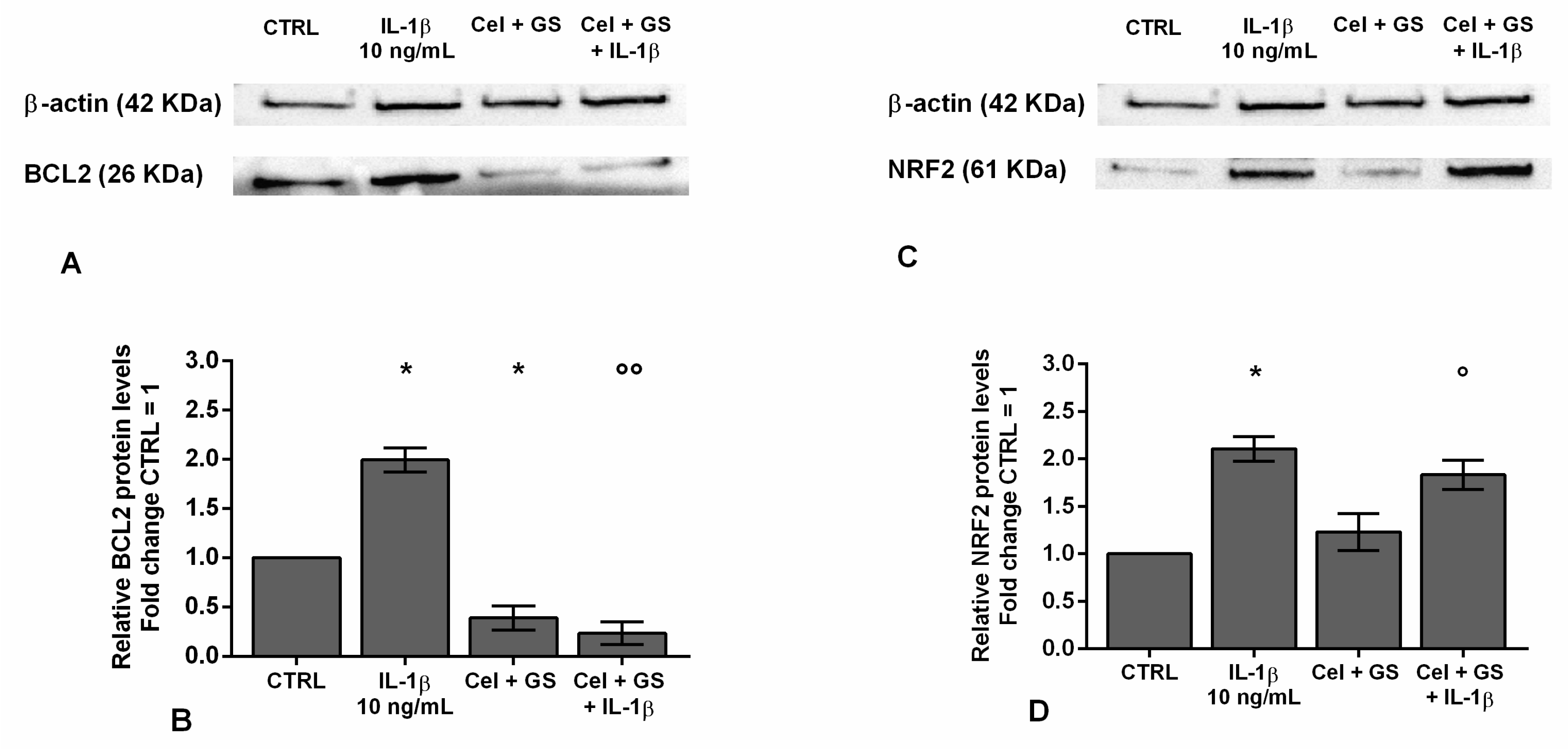
Figure 4.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of treatment with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence or not of interleukin (IL)-1β (10 ng/ml). (A-H) Expression levels of microRNA by quantitative real time PCR. The results were expressed as fold change of the value of interest respect to CTRL, reported equal to 1. Data were represented as mean ± standard deviation. *p < 0.05, **p < 0.01 versus basal CTRL. °p < 0.05, °°p < 0.01, °°°p < 0.001 versus IL-1β.
Figure 4.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of treatment with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence or not of interleukin (IL)-1β (10 ng/ml). (A-H) Expression levels of microRNA by quantitative real time PCR. The results were expressed as fold change of the value of interest respect to CTRL, reported equal to 1. Data were represented as mean ± standard deviation. *p < 0.05, **p < 0.01 versus basal CTRL. °p < 0.05, °°p < 0.01, °°°p < 0.001 versus IL-1β.
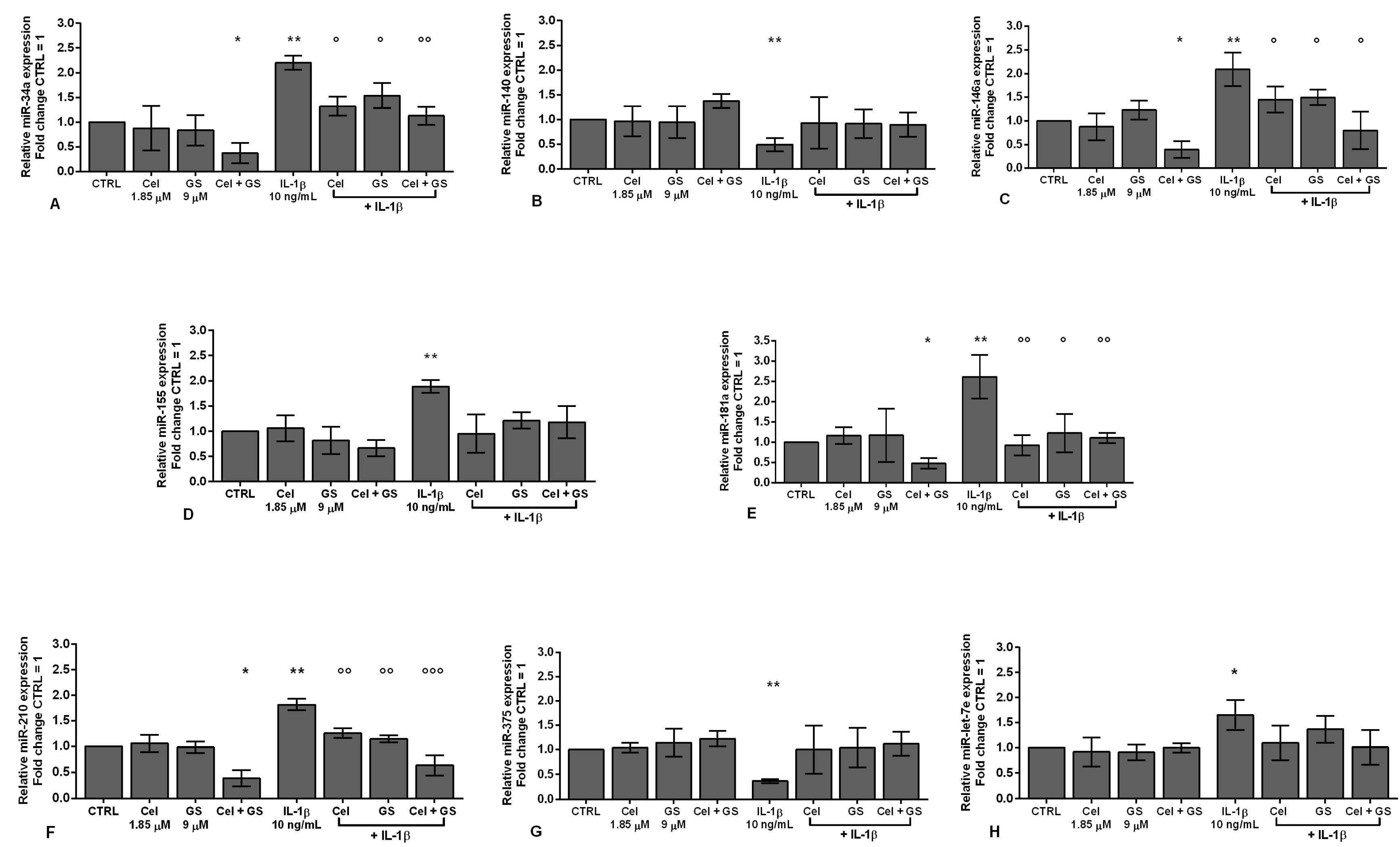
Figure 5.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of transient transfection with miR-34a, miR-146a, miR-181a, and miR-210 inhibitors (50 nM) or NC (5 nM), 24 h of incubation with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence of interleukin (IL)-1β (10 ng/ml). (A-E) Expression levels of microRNA by quantitative real time PCR. The results were expressed as fold change of the value of interest respect to CTRL, reported equal to 1. Data were represented as mean ± standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001 versus CTRL or NC. °p < 0.05, °°p < 0.01 versus IL-1β. # p < 0.05, ## p < 0.01 versus cel + IL, GS + IL and cel + GS + IL. INIB = Inhibitor, NC = negative control siRNA.
Figure 5.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of transient transfection with miR-34a, miR-146a, miR-181a, and miR-210 inhibitors (50 nM) or NC (5 nM), 24 h of incubation with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence of interleukin (IL)-1β (10 ng/ml). (A-E) Expression levels of microRNA by quantitative real time PCR. The results were expressed as fold change of the value of interest respect to CTRL, reported equal to 1. Data were represented as mean ± standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001 versus CTRL or NC. °p < 0.05, °°p < 0.01 versus IL-1β. # p < 0.05, ## p < 0.01 versus cel + IL, GS + IL and cel + GS + IL. INIB = Inhibitor, NC = negative control siRNA.
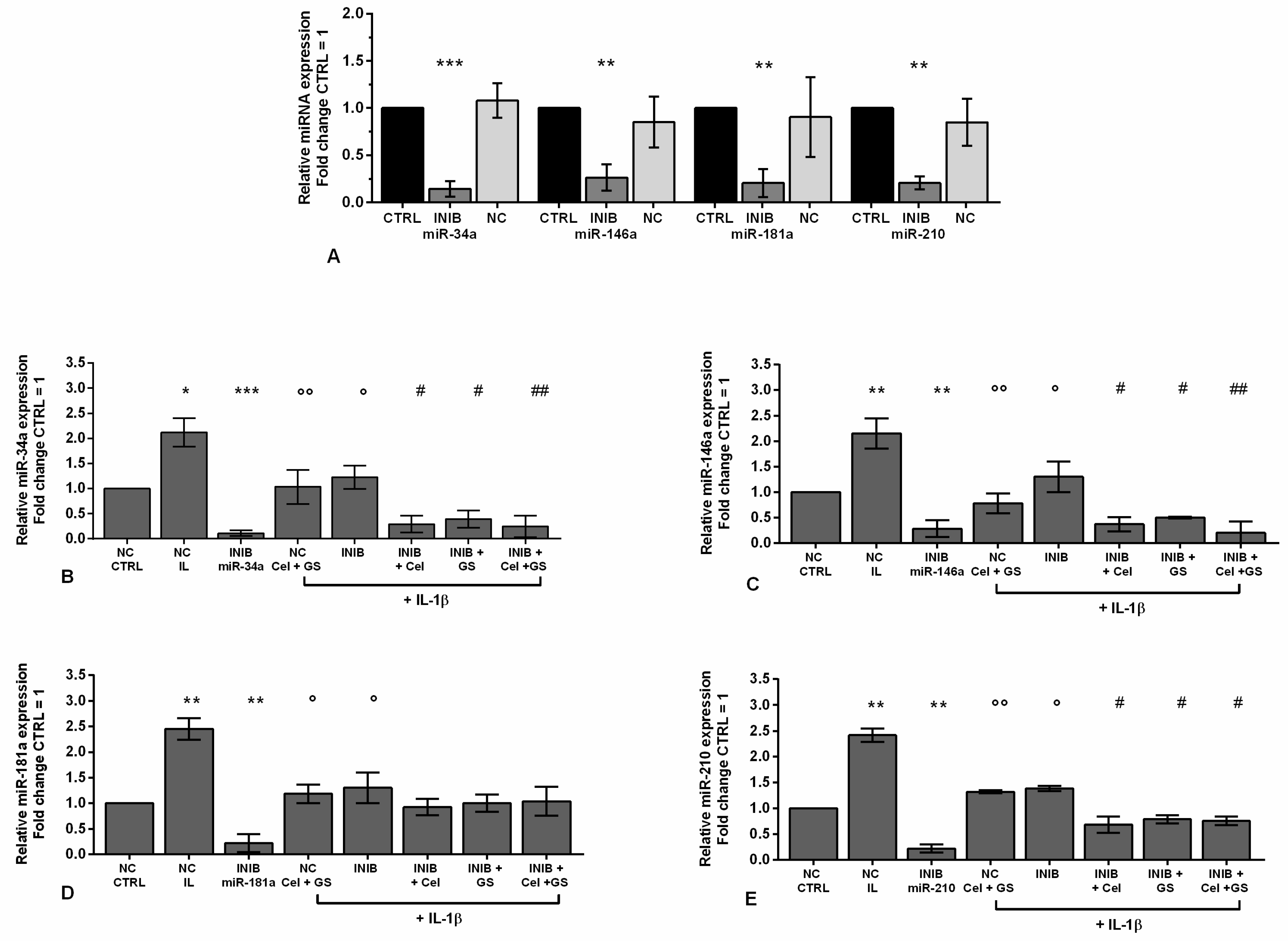
Figure 6.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of transient transfection with miR-34a, miR-146a, miR-181a, and miR-210 inhibitors (50 nM) or NC (5 nM), 24 h of incubation with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence of interleukin (IL)-1β (10 ng/ml). (A-D) Apoptosis detection by flow cytometry. The results were expressed as fold change of the value of interest respect to CTRL, reported equal to 1. (E-H) Viability by MTT assay. Data were represented as mean ± standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001 versus CTRL or NC. °p < 0.05, °°p < 0.01 versus IL-1β. # p < 0.05, ## p < 0.01 versus cel + IL, GS + IL and cel + GS + IL. INIB = Inhibitor, NC = negative control siRNA.
Figure 6.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of transient transfection with miR-34a, miR-146a, miR-181a, and miR-210 inhibitors (50 nM) or NC (5 nM), 24 h of incubation with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence of interleukin (IL)-1β (10 ng/ml). (A-D) Apoptosis detection by flow cytometry. The results were expressed as fold change of the value of interest respect to CTRL, reported equal to 1. (E-H) Viability by MTT assay. Data were represented as mean ± standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001 versus CTRL or NC. °p < 0.05, °°p < 0.01 versus IL-1β. # p < 0.05, ## p < 0.01 versus cel + IL, GS + IL and cel + GS + IL. INIB = Inhibitor, NC = negative control siRNA.
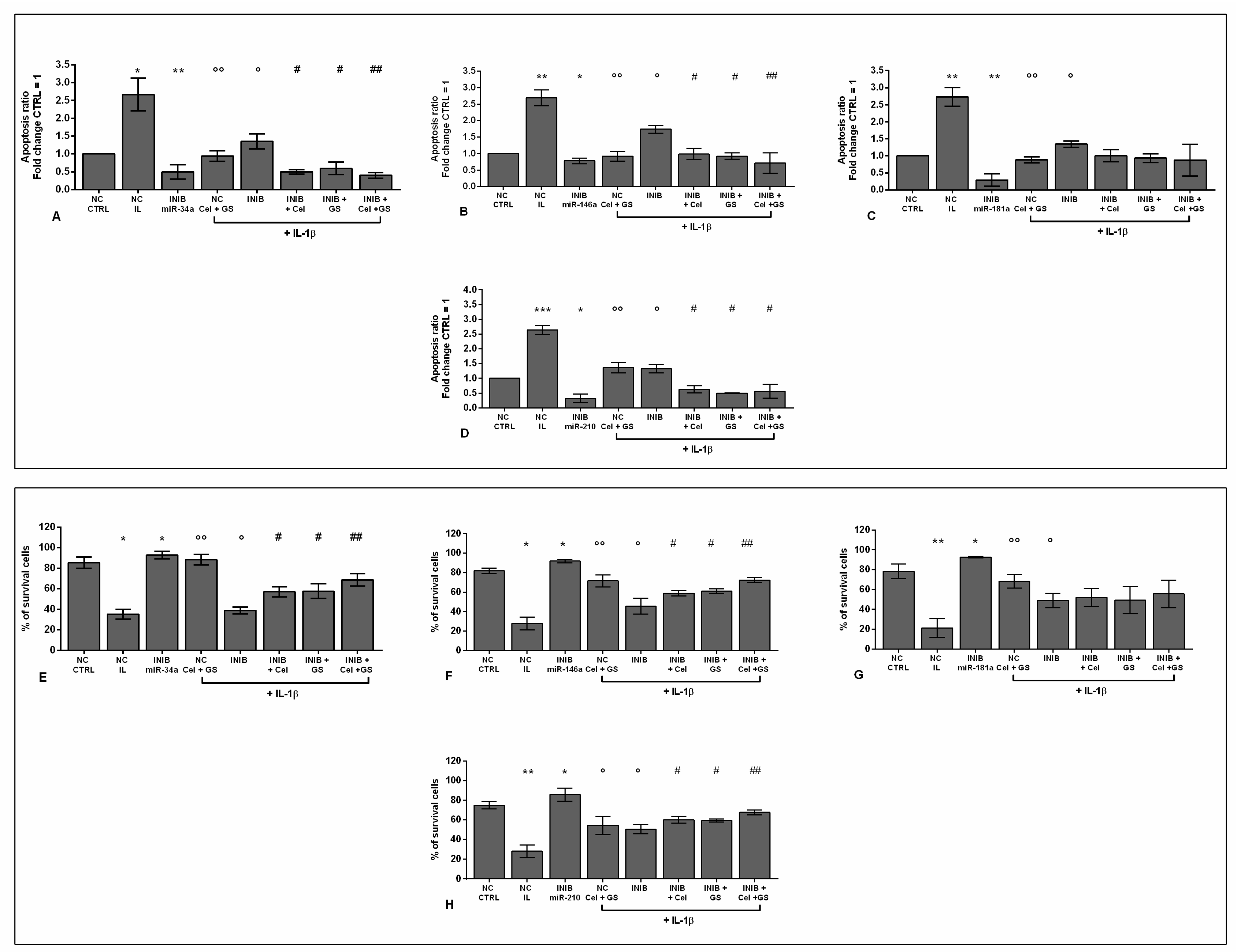
Figure 7.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of transient transfection with miR-34a, miR-146a, miR-181a, and miR-210 inhibitors (50 nM) or NC (5 nM), 24 h of incubation with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence of interleukin (IL)-1β (10 ng/ml). (A-D) Mitochondrial superoxide anion and (E-H) extracellular ROS production by flow cytometry. The results were expressed as fold change of the value of interest respect to CTRL, reported equal to 1. Data were represented as mean ± standard deviation. **p < 0.01, ***p < 0.001 versus CTRL or NC. °p < 0.05, °°p < 0.01 versus IL-1β. # p < 0.05, ## p < 0.01 versus cel + IL, GS + IL and cel + GS + IL. INIB = Inhibitor, NC = negative control siRNA.
Figure 7.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of transient transfection with miR-34a, miR-146a, miR-181a, and miR-210 inhibitors (50 nM) or NC (5 nM), 24 h of incubation with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence of interleukin (IL)-1β (10 ng/ml). (A-D) Mitochondrial superoxide anion and (E-H) extracellular ROS production by flow cytometry. The results were expressed as fold change of the value of interest respect to CTRL, reported equal to 1. Data were represented as mean ± standard deviation. **p < 0.01, ***p < 0.001 versus CTRL or NC. °p < 0.05, °°p < 0.01 versus IL-1β. # p < 0.05, ## p < 0.01 versus cel + IL, GS + IL and cel + GS + IL. INIB = Inhibitor, NC = negative control siRNA.
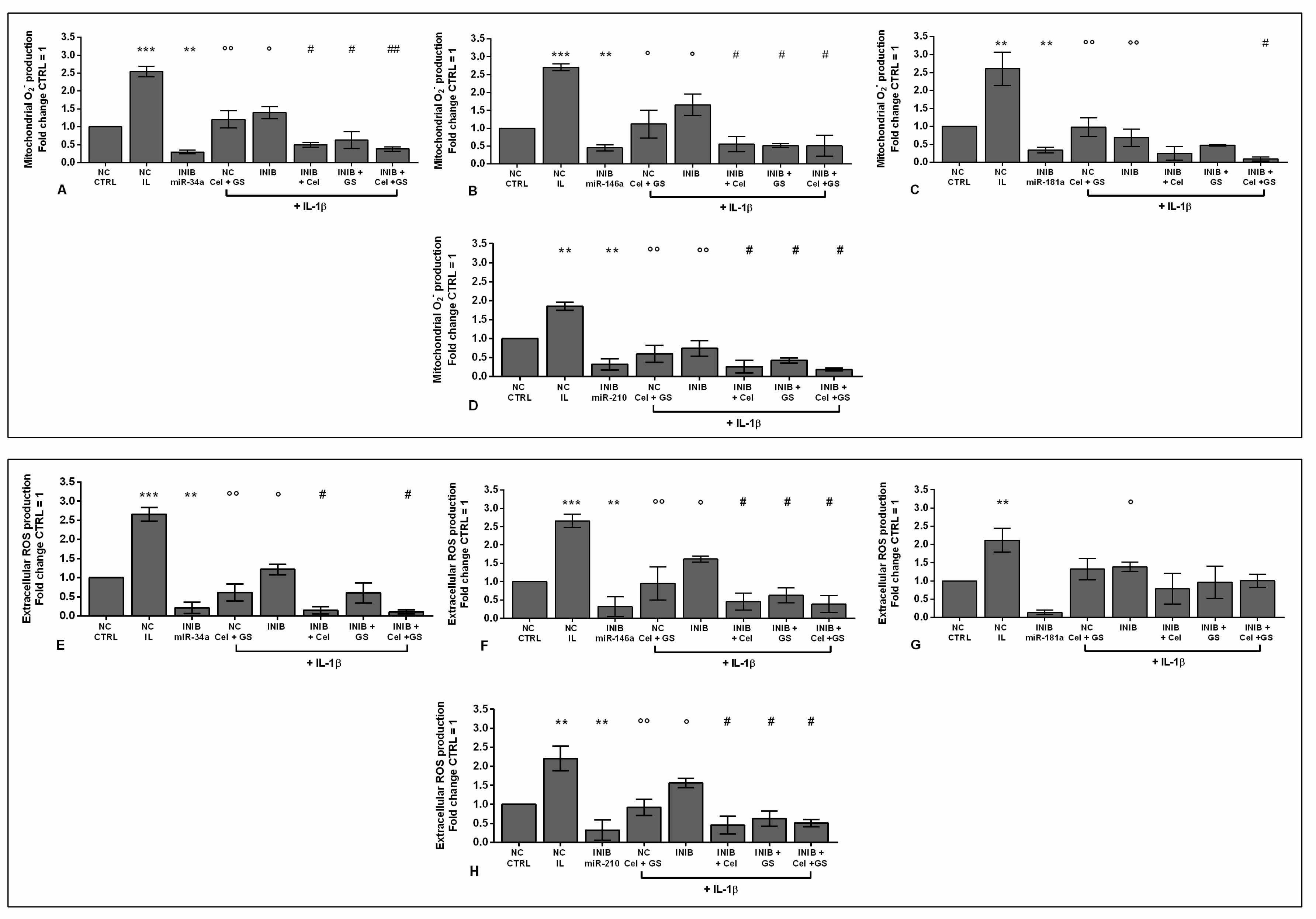
Figure 8.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of transient transfection with miR-34a, miR-146a, miR-181a, and miR-210 inhibitors (50 nM) or NC (5 nM), 24 h of incubation with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence of interleukin (IL)-1β (10 ng/ml). (A-D) Nitric oxide (NO) release by Griess method. Data were represented as mean ± standard deviation. *p < 0.05, **p < 0.01 versus CTRL or NC. °p < 0.05, °°p < 0.01 versus IL-1β. # p < 0.05 versus cel + IL, GS + IL and cel + GS + IL. INIB = Inhibitor, NC = negative control siRNA.
Figure 8.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of transient transfection with miR-34a, miR-146a, miR-181a, and miR-210 inhibitors (50 nM) or NC (5 nM), 24 h of incubation with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence of interleukin (IL)-1β (10 ng/ml). (A-D) Nitric oxide (NO) release by Griess method. Data were represented as mean ± standard deviation. *p < 0.05, **p < 0.01 versus CTRL or NC. °p < 0.05, °°p < 0.01 versus IL-1β. # p < 0.05 versus cel + IL, GS + IL and cel + GS + IL. INIB = Inhibitor, NC = negative control siRNA.
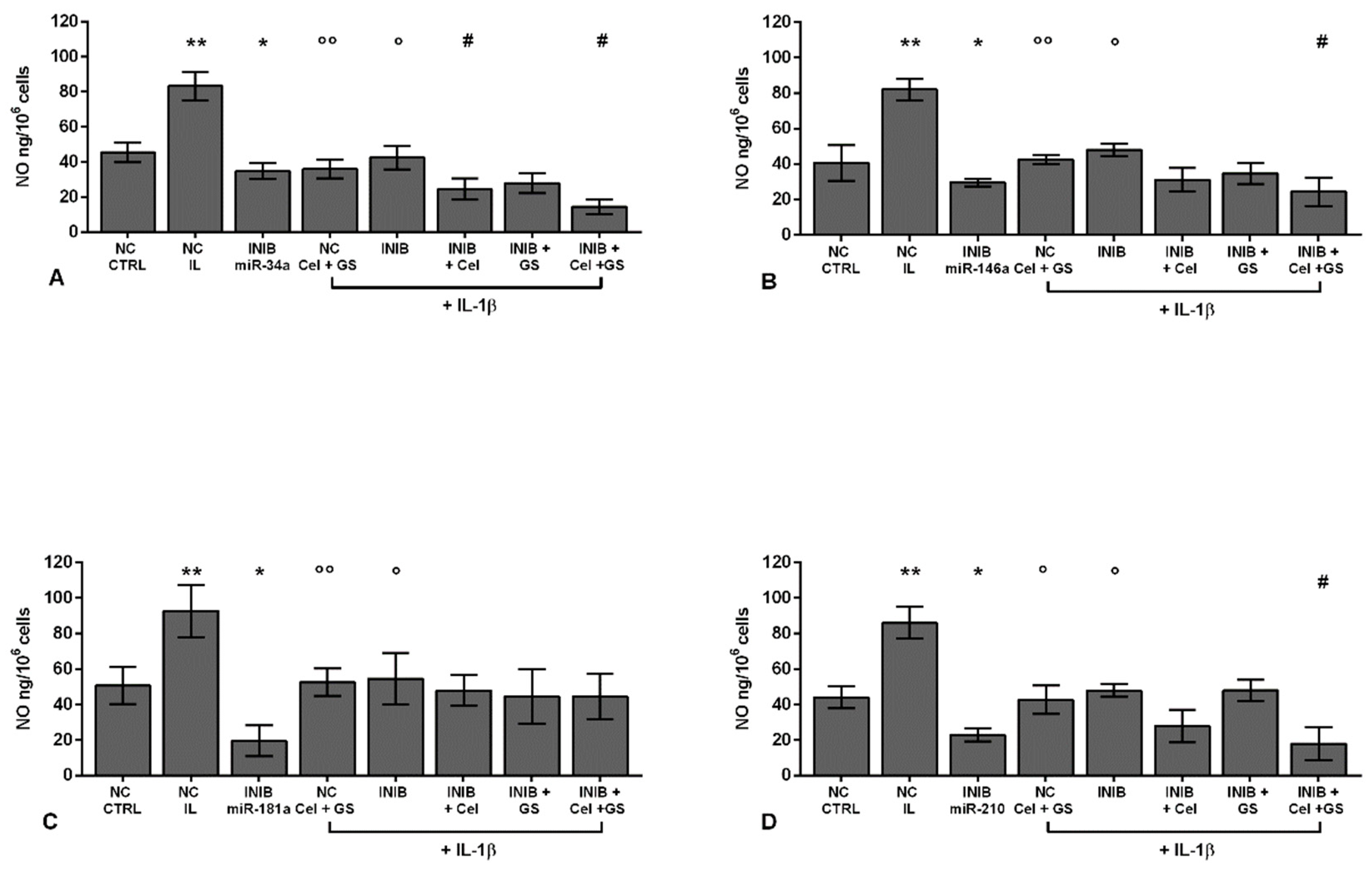
Figure 9.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of incubation with a specific nuclear factor (NF)-κB inhibitor (BAY 11-7082, IKKα/β, 1 μM), 24 h of treatment with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence of interleukin (IL)-1β (10 ng/ml). (A-C) Expression levels of microRNA by quantitative real time PCR. The results were expressed as fold change of the value of interest respect to CTRL, reported equal to 1. Data were represented as mean ± standard deviation. *p < 0.05, **p < 0.01 versus CTRL. °p < 0.05, °°p < 0.01 versus IL-1β. # p < 0.05 versus cel + GS + IL-1β.
Figure 9.
Chondrocytes were evaluated at control (CTRL) condition, after 24 h of incubation with a specific nuclear factor (NF)-κB inhibitor (BAY 11-7082, IKKα/β, 1 μM), 24 h of treatment with celecoxib (cel) (1.85 µM) and glucosamine sulfate (GS) (9 µM), in presence of interleukin (IL)-1β (10 ng/ml). (A-C) Expression levels of microRNA by quantitative real time PCR. The results were expressed as fold change of the value of interest respect to CTRL, reported equal to 1. Data were represented as mean ± standard deviation. *p < 0.05, **p < 0.01 versus CTRL. °p < 0.05, °°p < 0.01 versus IL-1β. # p < 0.05 versus cel + GS + IL-1β.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



