
Submitted:
18 August 2023
Posted:
12 September 2023
You are already at the latest version
Abstract
A comparative efficacy of apathogenic genotype I (V4) and lentogenic genotype II (LaSota) strains of live Newcastle disease virus (NDV) vaccines was done following vaccination with pathogen-associated molecular pattern (PAMP) H9N2 avian influenza vaccine and challenge with velogenic NDV genotype VII.1.1 (vNDV-VII.1.1). Eight groups (Gs) of day-old chicks were used (n=25). Groups 1-4 received a single dose of PAMP-H9N2 subcutaneously, while Gs (1, 5) and (2, 6) received eye drops of V4 and LaSota respectively as twice doses. All Gs except 4 and 8 were intramuscularly challenged with vNDV-VII.1.1 at 28th days of age. No signs were detected in Gs 1, 5, 4, and 8. The mortality rates were 0% in Gs 1, 4, 5, and 8; 40% in G2; 46.66% in G6; and 100% in Gs 3 and 7. Lesions were recorded as minimal in Gs 1 and 5, but mild to moderate in Gs 2 and 6. The lowest significant viral shedding was detected in Gs 1, 2, and 5. In conclusion, two successive vaccinations of broilers with a live V4 NDV vaccine provided a higher protection against vNDV-VII.1.1 challenge than LaSota. Besides, PAMP-H9N2 with live NDV vaccines induced more protection than live vaccine alone.
Keywords:
vNDV-VII.1.1 challenge
; broiler chickens
; live ND vaccines
; V4
; LaSota
; PAMP-H9N2
1. Introduction
Newcastle disease (ND) is an acute highly contagious viral disease of poultry. Over the last decade, ND has caused continuous devastating effects on the global poultry industry, thefore, the World Organization for Animal Health (WOAH) listed it as a notifiable terrestrial animal disease [1]. The incidence of ND has been increased because of improper vaccination programs with vaccination failure, presence of immunosuppressive diseases, besides the genetic mutations of the virus which led to changes in the biological characteristics and pathogenicity even in the vaccinated flocks [2].
Newcastle disease virus (NDV) or avian Orthoavulavirus 1 (AOAV-1) has been recently classified [3]. Based on the phylogenetic analysis of the fusion (F) protein gene sequence, NDV strains are divided into two major classes, namely class I and class II. Class I comprises non-virulent strains and consists of one genotype and 3 sub-genotypes [4]. However, class II includes virulent and non-virulent strains and consists of at least 20 genotypes (I–XXI) and multiple sub-genotypes following the exclusion of genotype XV which contains recombinant sequences [5,6]. Genotype VII.1.1 of NDV is now regarded as the predominant circulating genotype in various poultry species in Egypt as well as several African, Middle East, Asian, and European countries [7,8,9,10].
The outer surface proteins such as hemagglutinin-neuraminidase (HN) and F which are located on the NDV envelope are important for viral virulence, tropism, and immunoprotection through neutralizing antibodies [11,12,13]. The HN protein is important for the initial viral binding to sialic acid receptors of host cells, while F protein mediates the fusion of the viral envelope with the cell membrane [1].
Both vaccination and biosecurity measures are the major parallel strategies for the prevention and control of NDV infections in endemic areas. Avirulent and lentogenic strains of NDV have been extensively used as live vaccines that efficiently cross-protect against the velogenic and mesogenic pathotypes of the virus [14]. Although all NDV isolates are serologically grouped into a single serotype [1], the protection provoked by the vaccine is not always optimal. The inadequate efficacy of NDV vaccines may be owing to the poor vaccination practices, improper biosecurity measures, or the antigenic differences (neutralizing epitopes) between the circulating virulent viruses and the vaccine strains [13,14,15,16]. The antigenic similarity between the circulating velogenic (vNDV) and the vaccine strain could improve the induced vaccinal protection in terms of reduced infectivity and virus shedding [6,17,18,19]. Vaccination with a live genotype I-based vaccine (ND.TR.IR strain) protected broiler chickens against the local heterologous genotype VII vNDV and Herts 33 strain in Iran [20,21].
Chicken macrophages express several receptors for recognition of pathogens, including Toll-like receptors (TLRs). These receptors can bind to pathogen-associated molecular pattern (PAMP) or synthetic ligands leading to activation of macrophages which play critical role in the immunity against viruses [22].
To the best of our knowledge, scanty information is available about the comparative efficacy of genotypes I- and II-based live NDV vaccines in the protection of broiler chickens against vNDV-VII.1.1 challenge. Therefore, the objectives of this research were investigation of the protective efficacy of V4 (an apathogenic genotype I-) and LaSota (a lentogenic genotype II-) based live NDV vaccines against vNDV-VII.1.1 challenge in broiler chickens that were previously vaccinated with inactivated low pathogenic avian influenza virus subtype H9N2 prepared by PAMP technology (PAMP-H9N2).
2. Materials and Methods
2.1. Ethical Approval
All experimental procedures have complied with the general guidelines of the Local Ethics Commission of the Institutional Animal Care and Use Committee of Faculty of Veterinary Medicine, Cairo University (Vet IA CUC) and the ethical approval code is Vetcu09092023742. All efforts have been made to minimize chickens suffering.
2.2. Challenge virus
The challenge vNDV strain was isolated from a 26-days-old broiler chickens flock of 26,000 birds in El- Beheira governorate, Egypt. That flock was vaccinated against NDV using live vaccines only and it was exhibited mortality rate of 10% within 5 days before sampling. The virus was identified as AOAV-1 (NDV) genotype VII.1.1 (AOAV-1/Egy/Ch/MR78/2018), and the full F protein gene sequencing revealed the velogenic motif 112RRQKRF117 (GenBank accession No. MK984238). The intracerebral pathogenicity index of the isolated vNDV in chickens was 1.86 and the mean death time in the embryonated chicken eggs (ECEs) was 40 hours [23]. The challenge virus was propagated and titrated in 10-days-old specific pathogen free ECEs [24], and then the required challenge dose was prepared according to WOAH [25].
2.3. Birds' management, experimental design, and evaluation parameters
A total of 200, one-day-old, commercial broiler chicks (Ross 308) of mixed sex were divided into 8 equal groups (Gs), n=25 each (Table 1). Each group was floor reared in separate clean room. Groups 1-4 were subcutaneously vaccinated with an inactivated low pathogenic AIV vaccine using PAMP technology (H9N2®P, MSD, Intervet Int., Netherlands containing an inactivated avian influenza virus type A, subtype H9N2, strain A/CK/UAE/415/99; dose ≥ 8 Log2 HI) at the 4th day of age, while Gs 5-8 didn’t receive this vaccine. At the 8th and 15th days of age, chickens of Gs 1 and 5 were vaccinated with a live apathogenic V4 strain (Vaxsafe® ND, Bioproperties, Australia; dose ≥ 106 EID50), while those of Gs 2 and 6 were vaccinated with a live lentogenic LaSota strain (Avishield® ND, Decra, UK; dose 106-107 TCID50) using eye drop method for both vaccines. At 28-days-old (DO), each bird in Gs 1, 2, 3, 5, 6, and 7 was intramuscularly challenged with 0.1 ml containing 106 emberyo lethal dose (ELD50)/bird of vNDV-VII.1.1. However, Gs 4 and 8 were kept as controls without NDV vaccination or challenge. During the 38 days experimental period, the chickens were fed on a standard diet that met their nutritional requirements [26,27] and they had free access to freshwater ad-libitum.
2.3.1. Clinical observation and performance parameters
The clinical signs, gross lesions, and mortality were observed daily for 10 days post-challenge (dpc, 38 DO). Besides, the performance parameters including body weights (Bwts) and feed conversion ratio (FCR) were calculated weekly.
2.3.2. Histopathological assessment
Trachea, lung, thymus glands, spleen, and bursa of Fabricius of two birds/group, sacrificed humanely through cervical dislocation after intravenous injection of sodium pentobarbital with a dose of 50 mg/kg, were collected at 0, 3, 7, and 10 dpc. Organs were removed, flushed with sterile phosphate buffer saline (PBS, pH 7.4), and fixed in neutral buffered formaldehyde (10%) for 48 hours at 4°C. The fixed specimens were processed by the conventional paraffin embedding technique including dehydration through ascending grades of ethanol, clearing in three changes of xylene, and embedding in paraffin wax at 65°C. Four µm thick sections were stained by Hematoxylin and Eosin (H and E) as previously described by Bancroft and Layton [28]. The microscopical examination was done by using a digital camera (Leica EC3, Leica, Germany) connected with a microscope (Leica DM500) and a software (Leica LAS EZ). Semiquantitative lesion scoring of organs was calculated according to Gibson-Corley et al. [29]. Briefly, 10 fields were randomly chosen from each investigated group where the lesions were scored in a blind way by an expert (Score scale: 0= normal; 1≤ 25%; 2= 26–50%; 3= 51–75%; 4= 76–100%), and then averaged.
2.3.3. Evaluation of the virus shedding
The tracheal (n=3) and cloacal (n=3) individual swabs were collected at 0, 3, 7, and 10 dpc for the detection of NDV shedding using real-time polymerase chain reaction (rRT-PCR). Briefly, each swab was immersed in a tube containing 2 mL of PBS (supplemented with antibiotic and antifungal agents) and shaken to obtain a suspension which stored at -80oC until use. The viral RNA was extracted by viral RNA kit QIAamp Viral RNA Mini Kit (Qiagen, Germany) according to the manufacturer’s protocol. The used oligonucleotide primers and probes [30] were supplied from Metabion according to manufacturer instructions and the rRT-PCR reaction was conducted using Strategen 3005P, USA [8].
2.3.4. Serological evaluation
Eight blood samples were collected from the wing veins of chickens in each group on a weekly basis. The serum was separated to determine the antibody titers against NDV and AIV (H9N2) using hemagglutination inhibition (HI) test [25,31]. The standard antigens for NDV genotype II and low pathogenic (LPAIV) (H9N2) were used with haemagglutination titers of 8 log2 for both antigens, along with 4 haemagglutinating units were applied during the HI test.
2.3.5. Cytokines analysis
Five blood samples were collected from each chicken group at 17, 21, 28, and 31 DO for cytokines detection. The sera were examined for the innate and adaptive immune response parameters including interleukin-1β (IL-1β) [32], cluster of differentiation 4 (CD4) [33], lysozyme (LYZ) [34], and nitric oxide (NO) [35] using enzyme linked immuno-sorbent assay (ELISA) kits (Novatein Bio, Massachusetts, USA). Absorptions were measured at a wavelength of 450nm. In addition, 4 spleen samples were collected from each chicken group at 31 DO and their homogenates were examined for gene expression of IL-4, IL-10, and interferon-γ (IF-γ). Primers and cycling conditions were carried out [36,37,38].
3.4.6. Statistical analysis
All data calculations were statistically analyzed using the SPSS programming tool (IBM SPSS.20®) (SPSS Inc., Chicago, IL, USA) through one-way ANOVA followed by Duncan’s multiple range tests. All significant deviations were based on p≤0.05 [39].
3. Results
3.1. Clinical observation and performance parameters
The typical clinical picture of NDV such as sneezing, rales, nasal and ocular discharge, conjunctivitis, coughing, head swelling, greenish diarrhea, torticollis, and lateral recumbency, besides tracheitis, pneumonia, proventricular hemorrhages, enteritis, petechial hemorrhages on ileocecal tonsils, splenitis, hepatic congestion with distended gall bladder, and nephritis were observed from the 2nd dpc or the 5th dpc (nervous signs) in G3 (PAMP-challenged) and G7 (Non-vaccinated-challenged). However, less clinical picture has been observed in G2 (PAMP-LaSota-challenged) and G6 (LaSota-challenged) than control Gs 3 and 7. It is important to note that chickens of G1 (PAMP-V4-challenged) and G5 (V4-challenged) showed only a slight decrease in feed intake. The mortality rates were 0% in Gs 1, 4, 5, and 8; 40% in G2; 46.66% in G6, and 100% for both Gs 3 and 7 during the first 5 dpc (Figure 1). The highest significant (p≤0.05) average final Bwts were recorded in G4 (1985a g), G8 (1980a g), G5 (1968a g), and G1 (1944a g) compared to G6 (1496b g) and G2 (1465b g). Moreover, the FCR was recorded as 1.52, 1.55, 1.57, 1.58, 1.79, and 1.86 for Gs 1, 5, 4, 8, 2, and 6, respectively.
3.2. Histopathology
The tracheal epithelium and cilia were normal and well developed in chickens of control negative Gs 8 and 4 (Figure 2A and B). On the other hand, challenged chickens of G7 showed severe necrotic tracheitis, desquamation of hyperplastic hemorrhagic tracheal mucosa, and lymphocytic infiltration (Figure 2C). Likewise, chickens of challenged G3 revealed severe tracheal hyperplasia with a complete loss of the cilia and metaplasia of surface epithelia into squamous (Figure 2D). Despite LaSota vaccinated-challenged chickens of G6 displayed diffuse infiltration by lymphocytes, loss of cilia, and severe hemorrhage and degeneration of respiratory epithelia (Figure 2E), PAMP-LaSota vaccinated-challenged G2 showed less lesions in terms of slight hyperplasia and metaplasia of tracheal mucosa with slight degeneration of cilia (Figure 2F). In G5 (V4 vaccinated-challenged), chickens were exposed a relatively normal tracheal epithelia with slight lymphocytic infiltration and normal cilia (Figure 2G). Moreover, a complete protection of trachea in the form of normal epithelia was displayed in chickens of G1 (PAMP-V4-challenged) (Figure 2H).
The histopathological examination of the lung in chickens of Gs 4 and 8 revealed normal alveoli without any pathological lesions (Figure 3A and B). However, chickens in G7 had severe hemorrhage, interstitial fibrous pneumonia, and extensive lymphocytic infiltration (Figure 3C). Pre-vaccination with H9N2-PAMP in G3 slightly decreased the severity of lesions with no fibrous pneumonia, but severe hemorrhage and lymphocytic infiltration were present (Figure 3D). In contrast, pre-vaccination with LaSota either with (G2) or without (G6), H9N2-PAMP simply protected against the NDV challenge with a moderately congested blood vessels (Figure 3E and F). Furthermore, a relatively normal lungs were detected in V4 vaccinated G1 or G5 (Figure 3G and H).
The thymic architecture of the cortex and medulla in chickens of Gs 4 and 8 was normal without any detected lesions (Figure 4A and B). In contrast, NDV challenged chickens of G7 displayed destructed thymic architecture in the form of lymphoid depletion, necrosis in whole thymic lobule with macrophage accumulation, and severe congestion (Figure 4C). The thymic architecture was slightly protected in H9N2-PAMP vaccinated chickens of G3, but hemorrhage was still found and several Hassell’s corpuscles were extended to the thymic cortex (Figure 4D). In LaSota vaccinated Gs 2 and 6, the thymic architecture was more organized into the cortex and medulla with several congested blood vessels and many Hassell’s corpuscles (Figure 4E and F). The best protection for the thymic architecture was seen in both V4 vaccinated chickens of Gs 1 and 5 (Figure 4G and H).
Chickens of control negative Gs 4 and 8 did not have any lesions in the spleen (Figure 5A and B). On the other hand, severe multifocal lymphoid depletion and necrosis were detected in challenged chickens of both Gs 3 and 7 (Figure 5C and D). The vaccination with LaSota did not protect against NDV challenge in G6 (Figure 5E), but PAMP and LaSota vaccination induced mild necrosis and lymphoid depletion in G2 (Figure 5F). The splenic architecture was normal after the virus challenge and V4 vaccination of Gs 1 and 5 (Figure 5G and H).
The bursa of Fabricius was normal in chickens of both control negative Gs 4 and 8 (Figure 6A and B). However, NDV infection in G7 caused complete damage of the bursal architecture in terms of severe lymphoid depletion, necrosis, infiltration of inflammatory cells, and complete destruction of the covered epithelium (Figure 6C). In G3, H9N2-PAMP vaccination induced moderate lymphoid depletion and necrosis and the thickening of the interfollicular septum was filled by congested blood vessels (Figure 6D). LaSota-vaccination in G6 lowered the lesions caused by NDV and were represented by mild lymphoid depletion and necrosis (Figure 6E). However, H9N2-PAMP vaccinated chickens of G2 were more protected against the viral challenge (Figure 6F). The bursal architecture in V4-vaccinated Gs 1 and 5 was normal as the negative control groups (Figure 6G and H). More details for all examined organs were provided in a low power (supplementary Figures S1–S5). The histopathological lesion scores were significantly decreased (p≤0.05) in chickens of Gs 1 and 5, respectively compared to all other NDV challenged chickens. It was noticed that PAMP-H9N2 vaccinated chickens had lower lesion scores compared to non-vaccinated ones as indicated in Table 2.
3.3. Virus shedding
The lowest significant (p≤0.05) tracheal shedding was detected at the 3rd dpc in chickens of G1 (1.3 log10 viral copies) followed by G2 and G5 (2 log10) compared to G3, G6 (3.9 log10), and G7 (3.2 log10). The tracheal shedding continued till the 7th dpc in Gs 2, 5, and 6. Moreover, the cloacal shedding decreased significantly (p≤0.05) in Gs 5 (0.7 log10), 1 (1.6 log10), and 2 (2 log10) compared to Gs 3, 6 (3 log10), and 7 (2.7 log10) at the 3rd dpc. Chickens in both Gs 2 and 6 showed this cloacal shedding till the 7th and the 10th dpc (Figure 7).
3.4. Serology
At 28 DO, the highest significant (p≤0.05) HI titers for NDV were found in chickens of Gs 5, 1, and 2 as 5.25, 4.75, and 4.13 log2, respectively. However, at 35 DO (7th dpc), the titers increased significantly (p≤0.05) to 11, 10.63, 9.38, and 8 log2 in Gs 6, 2, 5, and 1, respectively compared to the other groups. In addition, at 35 DO, the highest significant (p≤0.05) HI titers for LPAIV (H9N2) were detected in Gs 1, 4, and 2 as 7.25, 6.88, and 6 log2, respectively compared to other groups (Table 3).
3.5. Cytokines
The results of IL-1β, CD4, LYZ, and NO parameters at 17, 21, and 28 DO indicated a higher significant difference (p≤0.05) in the expression of IL-1β in PAMP-vaccinated Gs 1, 4, and 3 compared to the non-PAMP-vaccinated Gs 5-8. However, at the 3rd dpc, the results of IL-1β didn’t differ significantly (p>0.05) in all groups but they were numerically higher in G2 (319.6 pg/ml) and G3 (318.4 pg/ml). Besides, at 31 DO, the highest significant difference (p≤0.05) in the CD4 level was in G8 (7.2 ng/ml), followed by Gs 5 and 7 (6 ng/ml), Gs 1 and 2 (5.6 ng/ml), as well as Gs 3 and 6 (5 ng/ml), respectively. For LYZ and NO expressions, G7 had the highest significant (p≤0.05) levels (132.6 ng/ml and 82.6 μmol/L, respectively) at the 3rd dpc (Table 4).
The gene expression of IL-4 and IL-10 in spleen showed a higher significant difference (p≤0.05) in Gs 1-6 in comparison to controls Gs 7 and 8. For IF-γ, the lowest gene expression was recorded in G8 (Ct value of 0), while the highest significant (p≤0.05) expression was recorded in G7 (the lowest Ct value of 5.6); followed by Gs 1, 2, and 6 (Ct values of 14, 15, and 15.75, respectively), and then Gs 5 and 3 (Ct values of 18 and 24.5, respectively). However, the lowest significant (p≤0.05) IF-γ gene expression [the highest significant (p≤0.05) Ct value of 27.5] was reported in G4 (Figure 8).
4. Discussion
Despite adoption of various vaccines and extensive vaccination programs for controlling ND, it still causes high and worldwide economic losses in the poultry industry. The low virulence strains of live NDV vaccines such as LaSota (pneumotropic) and B1 (V4 and VG/GA) (enterotropic) have been commonly used to control the virus infections. Generally, the pneumotropic NDV vaccines in chickens are more virulent and usually tend to be more immunogenic than enterotropic types [14,40,41]. Although all types of NDV vaccines are originating from different strains and genotypes, they are belonging to a single serotype and protect against morbidity and mortality. Matching of F and HN proteins of NDV vaccines and the homologous field virus is important in enhancing the vaccine efficiency and reducing the virus shedding rate [14,15,17,42,43,44]. Additionally, a high protection and reduction of the viral shedding have been reported in birds vaccinated with recombinant vaccines containing genotype I (D26) and challenged by heterologous genotypes VII.1.1, VII.2, and V of NDV [45,46,47,48]. Therefore, this study was planned to evaluate the protection induced by genotype I (V4) or genotype II (LaSota) strains of live NDV vaccines against vNDV genotype VII.1.1 challenge in broiler chickens. In addition, the efficacy of using a combined vaccination with PAMP-H9N2 and live NDV vaccines was demonstrated here.
During 10 dpc, genotype I (V4) vaccinated-challenged (Gs 1 and 5) were completely protected against NDV-induced clinical disease and mortalities, but genotype II (LaSota) vaccinated-challenged (Gs 2 and 6) showed some signs and lesions with mortality rates of 40% and 46.66%, respectively. However, non-vaccinated and challenged chickens (Gs 3 and 7) displayed severe signs with mortality rate of 100%. The study of Sultan et al. [49] showed mortality rate of 23% in 28-days-old broiler chickens following intranasal challenge by vND genotype VII.1.1 and vaccination with both live LaSota and inactivated genotype II NDV vaccines. The higher mortality rates in LaSota vaccinated chickens in our study might be attributed to the double vaccinations with LaSota vaccine without any inactivated vaccine and the intramuscular route of challenge. Absence or reducing of the clinical picture in vaccinated chickens Gs 1, 5, 2, and 6 was positively reflected on the Bwts and FCR. Histopathologically, chickens of V4 vaccinated (Gs 1 and 5) showed the best protection against the virus challenge with minimal pathologic lesions and nearly normal architectures of all organs during all days post challenge. However, some lesions including apoptosis, hemorrhages in the lung, spleen, and germinal center of the bursa of Fabricius, degenerated thymus glands, and detached tracheal epithelium have been detected in chickens of LaSota vaccinated (Gs 2 and 6) compared to challenged control (Gs 3 and 7) which displayed more excessive lesions at the 3rd and 7th dpc. Some pervious Egyptian reports confirmed decreasing in lesion scores with a high degree of protection in chickens vaccinated with different genotypes matching vaccines and challenged by vND-VII.1.1 [23,49,50,51]. Moreover, PAMP vaccination minimized the histopathological lesions in chickens in LaSota vaccinated (G2) compared to its parallel non-PAMP vaccinated (G6). This could be attributed to the enhancement of innate and adaptive immunity induced by PAMP vaccination. It is important to link the results of histopathology, virus shedding, serological immune response, and cytokines production in different groups. Complete reduction of the virus shedding was a main indication of the vaccine efficacy during the disease control in houses-rearing chickens [47]. Here, it was recorded that chickens V4-vaccinated G1 had the lowest and shortest tracheal and cloacal shedding. Moreover, chickens vaccinated with both PAMP-H9N2 and live NDV vaccines had shorter viral shedding compared to those received live NDV vaccine alone. In the same way, Bello et al. [52] reported that both the duration and load of the cloacal and oropharyngeal shedding of vND genotype VII virus challenge were significantly reduced when the genotype-matched vaccine was applied. Regarding the results of HI titers, the highest significant (p≤0.05) neutralizing antibodies against the challenge virus were found in V4-vaccinated G1 (HI titer of 8 Log2) and G5 (HI titer of 9.38 Log2), rather than LaSota-vaccinated G2 (HI titer of 10.63 Log2) and G6 (HI titer of 11 Log2). This result indicates the low efficacy of LaSota vaccine to induce a high matched antibodies to neutralize the challenge virus. It has been reported that the neutralization titers against NDV were always 3- to 6-fold higher against the homologous virus than the heterologous one, besides the cross-reactivity R-value between aSG10 (genotype VII NDV) and LaSota strain was 0.23, despite both aSG10 and LaSota strains are the same single serotype, they are loosely related with major antigenic differences [18]. Moreover, the vaccination-challenge study revealed that the aSG10 vaccine strain showed a significant higher protection rate than the LaSota vaccine strain when chickens were challenged with a genotype VII-NDV using intramuscular or eye drop/intranasal route [18]. Further, Shahar et al. [19] proved that the differences between NDV vaccine and the field viruses in the neutralizing epitopes (HN and F proteins) may enable escaping of the field variants strains from the vaccine antibodies. Based on the viral sequences, NDV isolates may be divided into “neutralization types” or “neutrotypes”, which can help in the development of mutation-adjusted vaccines [19]. From our point of view, the effective viral neutralization in both Gs 1 and 5 lead to low stimulation of IL-1β, mild pro-inflammation, and consequent mild stimulation of the antigen presenting cells. Such mild stimulation decreased the release of LYZ and NO in comparison to other groups, particularly G7 and LaSota vaccinated Gs 2 and 6. The critical innate and adaptive immunity of cytokines expression indicated a significant (p≤0.05) high enhancement of IL-4 and IL-10 in the spleen of PAMP-vaccinated chickens (Gs 1-4 ) and live NDV-vaccinated chickens (Gs 5 and 6). Besides, a significant (p≤0.05) high IF-γ gene expression was recorded in chickens of Gs 1, 2, 5, 3, and 4, respectively which indicates the anti-inflammatory effect. Previous research mentioned that IF-γ of chickens plays an important role in the immunogenicity and pathobiology of NDV [53,54]. The higher gene expression and better histopathology were reflected also on the viral shedding, as G1 had the lowest and shortest tracheal and cloacal shedding.
The main question of this study is how did V4 (genotype-I) vaccine of NDV provoke more protection of chcikens than LaSota did? To answer this question, it should consider the role of both F and HN proteins in the induction of neutralizing antibodies against vNDV. The HN protein prevents the viral attachment step (anti-HN antibodies), while the F protein inhibits the virus spread to the nearby cells (anti-F antibodies), even when the virus enters the cell and multiplies within it [55,56,57,58]. Further, Kim et al. [12] reported that the F protein plays the most important role in the protection of virus infection following the use of genotype-matched vaccines. Tabatabaeizadeh [59] demomstrated that the number of identical linear and conformational neutralizing epitopes of F and HN proteins (n=15) of the vaccine strains compared to the VII.2 NDV were the follows: (V4=14) > (I-2=11) > (Ulster, VG/GA, and R2B =10) > (F=9) > (LaSota=7) > (PHY.LMV.42=6) > (B1=5). In the same previous study, V4 and I-2 vaccine strains showed the highest number of identical epitopes compared to the other vaccine strains, while B1, PHY.LMV.42, and LaSota strains revealed the lowest epitope identity compared to VII.2 NDV “especially in F protein”. Under immunological pressure, the estimation of evolutionary distance confirmed that the number of amino acids substitutions per site between sequences was higher and changed more rapidly for HN compared to F protein [59,60,61,62,63,64,65,66]. Additionally, the decrease in the identity of common epitopes between the vaccine and the vNDV field strains may result in expanded protective antibodies [6,17,18,19]. In the recent study of Liu et al. [66], the results indicated that the amino acids similarity of LaSota strain with vNDV genotype VII strain was 88.4% for the F protein and 88.5% for the HN protein and that with vNDV genotype IX strain was 92.2% for the F protein and 91.1% for the HN protein. Moreover, genotype IX strain is genetically closer to LaSota strain than genotype VII strain and consequently this higher antigenic similarity can provide a better protection against the development of clinical disease and viral shedding [67].
Finally, it is essential to note that adoption of strict biosecurity measures before completion of the vaccine-derived immunity, controlling of the mixed viral/bacterial respiratory diseases and immunosuppressive agents, minimizing stressors, as well as application of efficient vaccines are crucial factors to prevent the infection with circulating NDV in poultry flocks [16].
5. Conclusions
Two successive vaccinations of commercial broiler chickens with apathogenic (genotype I-based, V4) live NDV vaccine fully protected chickens from the development of the clinical disease, mortalities, and histopathological lesions, lowered and shortened the viral shedding, and enhanced the antibodies and cytokines production compared to a lentogenic (genotype II-based, LaSota) live vaccine. Chickens received both PAMP-H9N2 and live NDV vaccines induced milder histopathological lesions and shorter viral shedding rate than those received a live NDV alone.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, A.R.E., H.F.E. and W.A.A.; methodology, A.R.E., W.A.A., H.F.E., A.R.G., A.E.N., M.A.A., M.E.S., A.O. and I.R.; writing—original draft preparation, A.R.E., and W.A.A.; writing—review and editing, A.R.E., .H.F.E., I.R., and W.A.A.; supervision, A.R.E., H.F.E., I.R., and W.A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement (Ethical approval)
All tests, procedures, and experimental work concerning the care of animals under study complied with the general guidelines and had approved by the Local Ethics Commission of the Institutional Animal Care and Use Committee of Cairo University, and all efforts were made to minimize suffering.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Miller, P.J.; Koch, G. Newcastle Disease. In Diseases of Poultry, 14th edition, Swayne, D. (Ed). John Wiley & Sons, Inc. Wiley Blackwell, Ames, IA, USA, 2020, pp. 112-129.
- Elfatah, K.S.A., Elabasy, M.A., El-Khyate, F., Elmahallawy, E.K., Mosad, S.M., El-Gohary, A.F., Abdo, W., Al-Brakati, A., Seadawy, M.G., Tahoon, A.E., El-Gohary, A.E. Molecular characterization of velogenic Newcastle disease virus (Sub-genotype VII.1.1) from wild birds, with assessment of its pathogenicity in susceptible chickens. Animals, 2021, 11(2), 505. [CrossRef]
- ICTV, International committee on taxonomy of viruses. Virus Taxonomy, 2019, Available at https://talk.ictvonline.org/taxonomy/.
- Dimitrov, H., Choi, K.S., Chvala, I., Diel, D.G., Durr, P.A., Ferreira, H.L., Fusaro, A., Gil, P., Goujgoulova, G.V., Grund, C., Hicks, J.T., Joannis, T.M., Torchetti, M.K., Kolosov, S., Lambrecht, B., Lewis, N.S., Liu, H., McCullough, S., Miller, P.J., Monne, I., Muller, C.P., Munir, M., Reischak, D., Sabra, M., Samal, S.K., de Almeida, R.S., Shittu, I., Snoeck, C.J., Suarez, D.L., van Borm, S., Wang, Z., Wong, F.Y.K. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infec. Gen. Evol., 2019, 74, 103917. [CrossRef]
- Afonso, C.L. Virulence during Newcastle disease viruses cross species adaptation. Viruses, 2021, 13(1), 252. [CrossRef]
- Miller, P.J., Afonso, C.L., El Attrache, J., Dorsey, K.M., Courtney, S.C., Guo, Z., Kapczynski, D.R. Effects of Newcastle disease virus vaccine antibodies on the shedding and transmission of challenge viruses. Dev. Comp. Immunol., 2013, 41(4), 505-513. [CrossRef]
- Miller, P.J., Haddas, R., Simanov, L., Lublin, A., Rehmani, S.F., Wajid, A., Bibi, T., Ahmad, S., Khan, T., Yaqub, T., Setiyaningsih, S., Afonso, C.L. Identification of new subgenotypes of virulent Newcastle disease virus with potential panzootic features. Infect. Gen. Evol., 2015, 29, 216-229. [CrossRef]
- Elbestawy, A.R., Ellakany, H.F., Abd El-Hamid, H.S., Zedan, R.E., Gado, A.R., Sedeik, M.E., Abd El-Hack, M.E., Saadeldin, I.M., Alowaimer, A.N., Ba-Awadh, H.A., Swelum, A.A. Muscovy ducks infected with velogenic Newcastle disease virus (genotype VIId) act as carriers to infect in-contact chickens. Poult. Sci., 2019, 98(10), 4441-4448. [CrossRef]
- Ellakany, H.F., Elbestawy, A.R., Abd El-Hamid, H.S., Zedan, R.E., Gado, A.R., Taha, A.E., Soliman, M.A., Abd El-Hack, M.E., Swelum, A.A., Saadeldin, I.M., Ba-Awadh, H., Hussein, E.O.S. Role of pigeons in the transmission of Avian Avulavirus (Newcastle disease-genotype VIId) to chickens. Animals, 2019a, 9(6), 338. [CrossRef]
- Abd-Ellatieff, H.A., Abd El Aziem, A.N., Elbestawy, A.R., Goda, A.R., Belih, S.M., Ellakany, H.F., Abd El-Hamid, H.S., Yanai, T., AbouRawash, A.A., El-Habashi, N. Efficacy of vaccination against infection with velogenic Newcastle disease virus genotypes VI and VII 1.1 strains in Japanese quails. J. Comp. Path. 2021, 186, 35-50. [CrossRef]
- Lee, Y.J., Sung, H.W., Choi, J.G., Lee, E.K., Yoon, H., Kim, J.H., Song, C.S. Protection of chickens from Newcastle disease with a recombinant baculovirus subunit vaccine expressing the fusion and hemagglutinin-neuraminidase proteins. J. Vet. Sci. 2008, 9(3), 301-308. [CrossRef]
- Kim, S.H., Wanasen, N., Paldurai, A., Xiao, S., Collins, P.L., Samal, S.K. Newcastle disease virus fusion protein is the major contributor to protective immunity of genotype-matched vaccine. PLoS One, 2013, 8(8), e74022. [CrossRef]
- Liu, J., Zhu, J., Xu, H., Li, J., Hu, Z., Hu, S., Wang, X., Liu, X. Effects of the HN antigenic difference between the vaccine strain and the challenge strain of Newcastle Disease virus on virus shedding and transmission. Viruses, 2017, 9(8), 225. [CrossRef]
- Dimitrov, K.M., Afonso, C.L., Yu, Q., Miller, P.J. Newcastle disease vaccines-A solved problem or a continuous challenge? Vet. Microbiol., 2017, 206, 126-136. [CrossRef]
- Miller, P.J., King, D.J., Afonso, C.L., Suarez, D.L. Antigenic differences among Newcastle disease virus strains of different genotypes used in vaccine formulation affect viral shedding after a virulent challenge. Vaccine, 2007, 25(41), 7238-7246. [CrossRef]
- Dortmans, J.C.F.M., Peeters, B.P.H., Koch, G. Newcastle disease virus outbreaks: Vaccine mismatch or inadequate application? Vet. Microbiol., 2012, 160(1-2), 17-22. [CrossRef]
- Miller, P.J., Estevez, C., Yu, Q., Suarez, D.L., King, D.J. Comparison of viral shedding following vaccination with inactivated and live Newcastle disease vaccines formulated with wild-type and recombinant viruses. Avian Dis., 2009, 53(1), 39-49. [CrossRef]
- Yang, H.M., Zhao, J., Xue, J., Yang, Y.L., Zhang, G.Z. Antigenic variation of LaSota and genotype VII Newcastle disease virus (NDV) and their efficacy against challenge with velogenic NDV. Vaccine, 2017, 35(1), 27-32. [CrossRef]
- Shahar, E., Haddas, R., Goldenberg, D., Lublin, A., Bloch, I., Hinenzon, N.B., Pitcovski, J. Newcastle disease virus: is an updated attenuated vaccine needed? Avian Pathol., 2018, 47(5), 467-478. [CrossRef]
- Abdoshah, M., Hassanzadeh, M., Masoudi, S., Ashtari, A., Yousefi, A.R., Nasr, M.P. Thermoresistant Newcastle disease vaccine effectively protects SPF, native, and commercial chickens in challenge with virulent virus. Vet. Med. Sci., 2022, 8(4), 1539-1546. [CrossRef]
- Hassanzadeh, M., Abdoshah, M., Yousefi, A.R., Moluki, I., Haghshenas, F. Evaluation of virulence and phylogenetic study of Newcastle disease virus isolated from 295 broiler poultry farms in Mazandaran province. Iran. Vet. J. 2022, In press. [CrossRef]
- Qureshi, M.A., Heggen, C.L., Hussain, I. Avian macrophage: effector functions in health and disease. Dev. Comp. Immunol., 2000, 24, 103–119.
- Ellakany, H.F., Abd El-Hamid, H.S., Nasef, S.A., Elbestawy, A.R., Nasr, S.M., Abd El Aziz, M.N., Gado, A.R., Zedan, R.E., Yonis, A.E. Evaluation of the protection of commercial live and inactivated NDV vaccines against Newcastle virus genotype VIId circulating in the field. Daman. J. Vet. Sci., 2019b, 1(1), 17-20. [CrossRef]
- Reed, L.J., Muench, L.H. A simple method of estimating fifty percent end points. Amer. J. Hyg., 1938, 27, 493-497.
- World Organization for Animal Health, WOAH, OIE. Newcastle disease (infection with Newcastle disease virus). Manual of diagnostic tests and vaccines for terrestrial animals, chapter 3.3.14., 2021a.
- Aviagen, R. Broiler Nutrition Specifications (All plant-protein based feeds). 2022. USA. Available online, https://aviagen.com/assets/Tech_Center/Ross_Broiler/Ross-PlantProteinBasedBroilerNutritionSpecifications2022-EN.pdf.
- NRC. Nutrient requirements of poultry. 10th Revised Edition. National Academy Press, Washington, DC, USA, 2016.
- Bancroft, J.D., Layton, C. The hematoxylin and eosin. In Theory and Practice of Histological Techniques. SK Suvarna, C Layton and JD Bancroft, eds. 7th ed. ElSevier, Churchill Livingstone, Pennsylvania, USA, 2013, pp. 179-220.
- Gibson-Corley, K.N., Olivier, A.K., Meyerholz, D.K. Principles for valid histopathologic scoring in research. Vet. Pathol. 2013, 50 (6), 1007-1015. [CrossRef]
- Wise, M.G., Suarez, D.L., Seal, B.S., Pedersen, J.C., Senne, D.A., King, D.J., Kapczynski, D.R., Spackman, E.E. Development of a real-time reverse-transcription PCR for detection of Newcastle disease virus RNA in clinical samples. J. Clin. Microbiol., 2004, 42 (1), 329-338. [CrossRef]
- World Organization for Animal Health, WOAH, OIE. Avian Influenza (including infection with high pathogenicity avian influenza viruses). Manual of diagnostic tests and vaccines for terrestrial animals, chapter 3.3.4. 2021b.
- Kaiser, P., Rothwell, L., Goodchild, M., Bumstead, N. The chicken proinflammatory cytokines interleukin-1beta and interleukin-6: differences in gene structure and genetic location compared with their mammalian orthologues. Anim. Genet., 2004, 35, 169-175. doi: 10.1111/j.1365-2052.2004.01121.x.
- Yu, X., Rui, L., Shao, Q., Liu, H., Lu, Y., Zhang, Y., Li, Z. Changes of CD4+CD25+ cells ratio in immune organs from chickens challenged with infectious bursal disease virus strains with varying virulences. Viruses, 2015, 7, 1357. doi: 10.3390/v7031357.
- Huang, X., Sang, S., Yuan, Z., Duan, Q., Guo, X., Zhang, H., Zhao, C. Magnetoelastic immunosensor via antibody immobilization for the specific detection of lysozymes. ACS Sensors, 2021, 6(11), 3933-3939. [CrossRef]
- Ariaans, M.P., Matthijs, M.G.R., van Haarlem, D., van de Haar, P., van Eck, J.H.H., Hensen, E.J., Vervelde, L. The role of phagocytic cells in enhanced susceptibility of broilers to colibacillosis after infectious bronchitis virus infection. Vet. Immunol. Immunopathol. 2008, 123, 240-250. doi: 10.1016/j.vetimm.2008.02.003.
- Hong, Y.H., Lillehoj, H.S., Lee, S.H., Dalloul, R.A., Lillehoj, E.P. Analysis of chicken cytokine and chemokine gene expression following Eimeria acervulina and Eimeria tenella infections. Vet. Immunol. Immunopathol. 2006a, 114, 209-223.
- Hong, Y.H., Lillehoj, H.S., Lillehoj, E.P., Lee, S.H. Changes in immune-related gene expression and intestinal lymphocyte subpopulations following Eimeria maxima infection of chickens. Vet. Immunol. Immunopathol., 2006b, 114, 259-272.
- Adams, S.C., Xing, Z., Li, J., Cardona, C.J. Immune-related gene expression in response to H11N9 low pathogenic avian influenza virus infection in chicken and Pekin duck peripheral blood mononuclear cells. Mol. Immunol., 2009, 46, 1744-1749.
- SAS. SAS User’s Guide Statistics. 2004, Cary, NC: SAS Institute, Inc.
- Westbury, H. Newcastle disease virus: An evolving pathogen? Avian Pathol., 2001, 30(1), 5-11. [CrossRef]
- Kim, J.Y., Kye, S.J., Lee, H.J., Gaikwad, S., Lee, H.S., Jung, S.C., Choi, K.S. Development of a highly immunogenic Newcastle disease virus chicken vaccine strain of duck origin. Poult. Sci. 2016, 95(4), 790-797. [CrossRef]
- Hu, Z., Hu, S., Meng, C., Wang, X., Zhu, J., Liu, X. Generation of a genotype VII Newcastle disease virus vaccine candidate with high yield in embryonated chicken eggs. Avian Dis., 2011, 55(3), 391-397. [CrossRef]
- Cheng, Y., Sheng, D., Li, X., Hong, S., Guo, L., Zhao, S., Yuan, Y., Xue, J., Tian, H., Ren, Y., Liu, W., Tian, K. Efficacy of a recombinant genotype VII vaccine against challenge with velogenic Newcastle disease virus. J. Vac. Immunol., 2016, 2(1), 019-022.
- Mahmoud, N.K., El-Deeb, A.H., Emara, M.M., Abd El-Khaleck, M.A., Hussein, H.A. Genotypes II and VIId-based inactivated Newcastle disease vaccine reduces virus shedding. Virus dis., 2019, 30(3), 453-461. [CrossRef]
- Palya, V., Kiss, I., Tatár-Kis, T., Mató, T., Felföldi, B., Gardin, Y. Advancement in vaccination against Newcastle disease: recombinant HVT NDV provides high clinical protection and reduces challenge virus shedding with the absence of vaccine reactions. Avian Dis., 2012, 56(2), 282-287. [CrossRef]
- Palya, V., Tatár-Kis, T., Mato, T., Felfoldi, B., Kovacs, E., Gardin, Y. Onset and long-term duration of immunity provided by a single vaccination with a turkey herpesvirus vector ND vaccine in commercial layers. Vet. Immunol. Immunopathol., 2014, 158(1-2), 105-115. [CrossRef]
- Palya, V., Tatár-Kis, T., Arafa, A.S.A., Felföldi, B., Mató, T., Setta, A. Efficacy of a turkey herpesvirus vectored Newcastle disease vaccine against genotype VII.1.1 virus: challenge route affects shedding pattern. Vaccines, 2021, 9(1), 37. [CrossRef]
- Tatar-Kis, T., Fischer, E.A.J., Cazaban, C., Walko-Kovacs, E., Homonnay, Z.G., Velkers, F.C., Palya, V., Stegeman, J.A. A herpesvirus of turkey-based vector vaccine reduces transmission of Newcastle disease virus in commercial broiler chickens with maternally derived antibodies. Vaccines, 2020, 8(4), 614. [CrossRef]
- Sultan, H.A., Elfeil, W.K., Nour, A.A., Tantawy, L., Kamel, E.G., Eed, E.M., El Askary, A., Talaat, S. Efficacy of the Newcastle disease virus genotype VII.1.1-matched vaccines in commercial broilers. Vaccines, 2021, 10(1), 29. https:// doi.org/10.3390/vaccines10010029.
- Sultan, H.A., Talaat, S., Elfeil, W.K., Selim, K., Kutkat, M.A., Amer, S.A., Choi, K.S.. Protective efficacy of the Newcastle disease virus genotype VII–matched vaccine in commercial layers. Poult. Sci., 2020, 99, 1275–1286.
- Dewidar, A.A.A., Kilany, W.H., El-Sawah, A.A., Shany, S.A.S., Dahshan, A.H.M., Hisham, I., Elkady, M.F., Ali, A. Genotype VII.1.1-based Newcastle disease virus vaccines afford better protection against field isolates in commercial broiler chickens. Animals, 2022, 12, 1696. [CrossRef]
- Bello, M.B., Mahamud, S.N.A., Yusoff, K., Ideris, A., Hair-Bejo, M., Peeters, B.P.H., Omar, A.R. Development of an effective and stable genotype-matched live attenuated Newcastle disease virus vaccine based on a novel naturally recombinant Malaysian isolate using reverse genetics. Vaccines, 2020, 8(2), 270. [CrossRef]
- Kapczynski, D.R., Afonso, C.L., Miller, P.J. Immune responses of poultry to Newcastle disease virus. Dev. Comp. Immunol., 2013, 41(3): 447-453. [CrossRef]
- Cardenas-Garcia, S., Dunwoody, R.P., Marcano, V., Diel, D.G., Williams, R.J., Jr. Gogal, R.M., Brown, C.C., Miller, P.J., Afonso, C.L. Effects of chicken interferon gamma on Newcastle disease virus vaccine immunogenicity. PLoS One, 2016, 11(7), e0159153. [CrossRef]
- Merz, D.C., Scheid, A., Choppin, P. Importance of antibodies to the fusion glycoprotein of paramyxoviruses in the prevention of spread of infection. J. Exp. Med., 1980, 151(2), 275-288. [CrossRef]
- Boursnell, M., Green, P., Samson, A., Campbell, J., Deuter, A., Peters, R., Millar, N., Emmerson, P., Binns, M. A recombinant fowlpox virus expressing the hemagglutinin-neuraminidase gene of Newcastle disease virus (NDV) protects chickens against challenge NDV. Virology, 1990, 178(1), 297-300. [CrossRef]
- Naohiro, K., Masahiro, N., Mitsuru, O., Chieko, K., Yoshiharu, M., Takeshi, M. Protective effect of individual glycoproteins of Newcastle disease virus expressed in insect cells: the fusion protein derived from an avirulent strain had lower protective efficacy. Virus Res., 1994, 32(3), 373-379. [CrossRef]
- Reynolds, D., Maraqa, A. Protective immunity against Newcastle disease: the role of antibodies specific to Newcastle disease virus polypeptides. Avian Dis. 2000, 44(1), 138-144. [CrossRef]
- Tabatabaeizadeh, S.E. Immunoinformatic analysis and antibody epitope comparison of Newcastle disease virus classical vaccines with a virus involved in the fifth NDV panzootic. Mapping Intimacies, (Res. Square), 2021, 1-16. DOI: 10.21203/rs.3.rs-456948/v1.
- Chambers, P., Nesbit, M., Yusoff, K., Millar, N., Samson, A., Emmerson, P. Location of a neutralizing epitope for the haemagglutinin-neuraminidase glycoprotein of Newcastle disease virus. J. Gen. Virol., 1988, 69(8), 2115-2122. [CrossRef]
- Toyoda, T., Gotoh, B., Sakaguchi, T., Kida, H., Nagai, Y. Identification of amino acids relevant to three antigenic determinants on the fusion protein of Newcastle disease virus that are involved in fusion inhibition and neutralization. J. Virol., 1988, 62(11), 4427-4430. [CrossRef]
- Neyt, C., Geliebter, J., Slaoui, M., Morales, D., Meulemans, G., Burny, A. Mutations located on both F1 and F2 subunits of the Newcastle disease virus fusion protein confer resistance to neutralization with monoclonal antibodies. J. Virol., 1989, 63(2), 952-954. [CrossRef]
- Iorio, R.M., Syddall, R.J., Sheehan, J.P., Bratt, M.A., Glickman, R.L., Riel, A.M. Neutralization map of the hemagglutinin-neuraminidase glycoprotein of Newcastle disease virus: domains recognized by monoclonal antibodies that prevent receptor recognition. J. Virol., 1991, 65(9), 4999-5006. [CrossRef]
- Iorio, R.M., Glickman, R.L., Sheehan, J.P. Inhibition of fusion by neutralizing monoclonal antibodies to the haemagglutinin-neuraminidase glycoprotein of Newcastle disease virus. J. Gen. Virol., 1992, 73(5), 1167-1176. [CrossRef]
- Gong, Y., Cui, Z. Epitope variation in the Newcastle disease virus HN gene under antibody immune selective pressure in cell culture. Sci. China Life Sci., 2011, 54(5), 474-479. [CrossRef]
- Gu, M., Liu, W., Xu, L., Cao, Y., Yao, C., Hu, S., Liu, X. Positive selection in the hemagglutinin neuraminidase gene of Newcastle disease virus and its effect on vaccine efficacy. Virol. J., 2011, 8, 150. [CrossRef]
- Liu, M., Shen, X., Li, J., Yu, Y., Fan, J., Jia, X., Dai, Y. Efficacy of Newcastle disease LaSota vaccine-induced hemagglutination inhibition antibodies against challenges with heterologous virulent strains of genotypes VII and IX. Vet. Immunol. Immunopathol., 2023, 259, 110591. [CrossRef]
Figure 1.
Survival rate in all chicken groups.
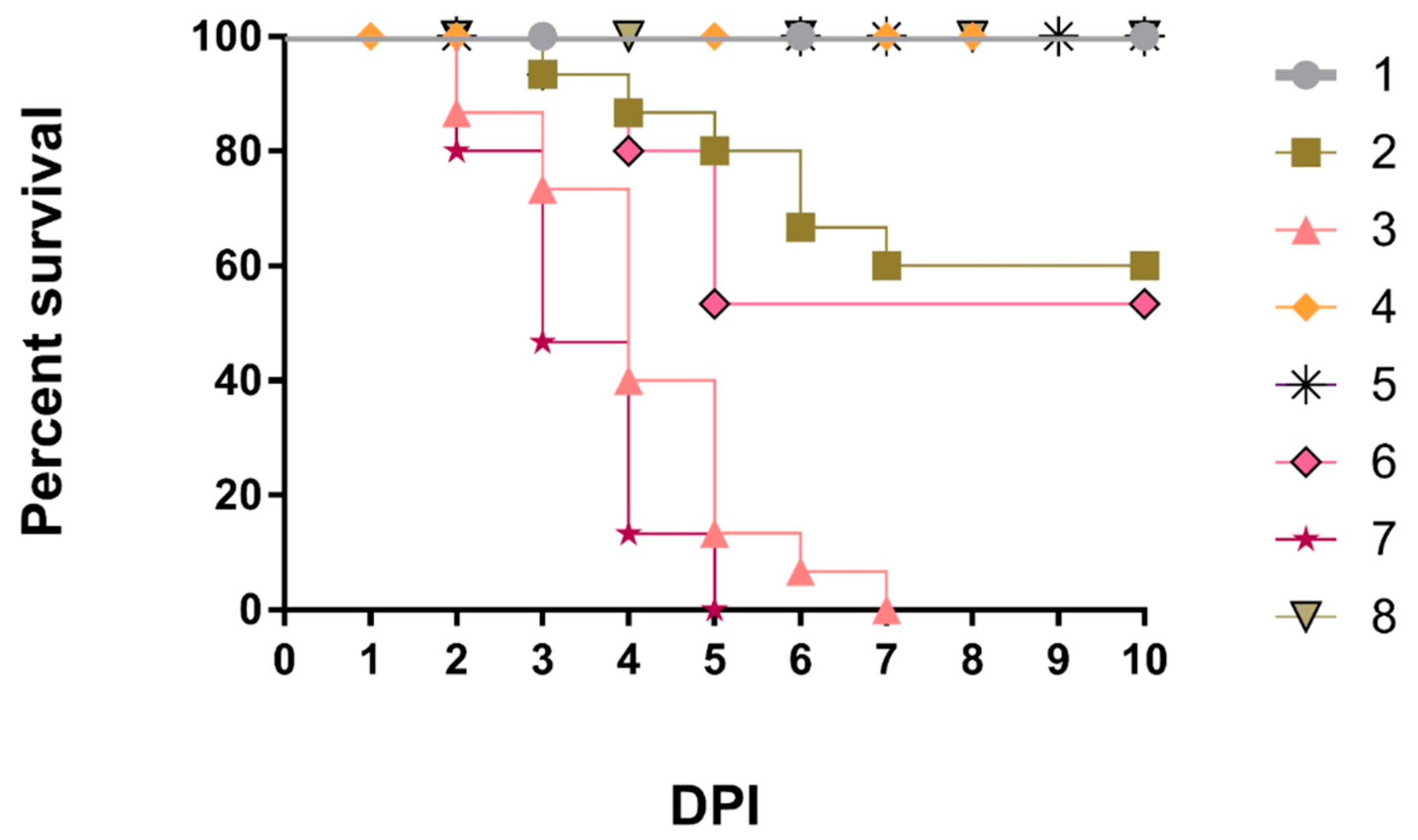
Figure 2.
Histopathological examination of chicken's trachea. (A) Negative control (G8). (B) H9N2-PAMP vaccination only (G4). (C) vND-VII.1.1 challenge only (G7). (D) H9N2-PAMP vaccination and vND-VII.1.1 challenge (G3). (E) LaSota vaccination and vND-VII.1.1 challenge (G6). (F) H9N2-PAMP, LaSota vaccination, and vND-VII.1.1 challenge (G2). (G) V4 vaccination and vND-VII.1.1 challenge (G5). (H) H9N2-PAMP, V4 vaccination, and vND-VII.1.1 challenge (G1). Normal cilia (thin arrows), hemorrhage (arrowheads), normal tracheal glands (stars), and mucosal hyperplasia and metaplasia (thick arrows). Scale bar =50 μm.
Figure 2.
Histopathological examination of chicken's trachea. (A) Negative control (G8). (B) H9N2-PAMP vaccination only (G4). (C) vND-VII.1.1 challenge only (G7). (D) H9N2-PAMP vaccination and vND-VII.1.1 challenge (G3). (E) LaSota vaccination and vND-VII.1.1 challenge (G6). (F) H9N2-PAMP, LaSota vaccination, and vND-VII.1.1 challenge (G2). (G) V4 vaccination and vND-VII.1.1 challenge (G5). (H) H9N2-PAMP, V4 vaccination, and vND-VII.1.1 challenge (G1). Normal cilia (thin arrows), hemorrhage (arrowheads), normal tracheal glands (stars), and mucosal hyperplasia and metaplasia (thick arrows). Scale bar =50 μm.

Figure 3.
Histopathological examination of chicken's lung. (A) G8. (B) G4. (C) G7. (D) G3. (E) G6. (F) G2. (G) G5. (H) G1. Normal alveoli (thin arrows), thick interstitial connective tissue filled with inflammatory cells (arrowheads), and hemorrhage (thick arrows). Scale bar =50 μm.
Figure 3.
Histopathological examination of chicken's lung. (A) G8. (B) G4. (C) G7. (D) G3. (E) G6. (F) G2. (G) G5. (H) G1. Normal alveoli (thin arrows), thick interstitial connective tissue filled with inflammatory cells (arrowheads), and hemorrhage (thick arrows). Scale bar =50 μm.
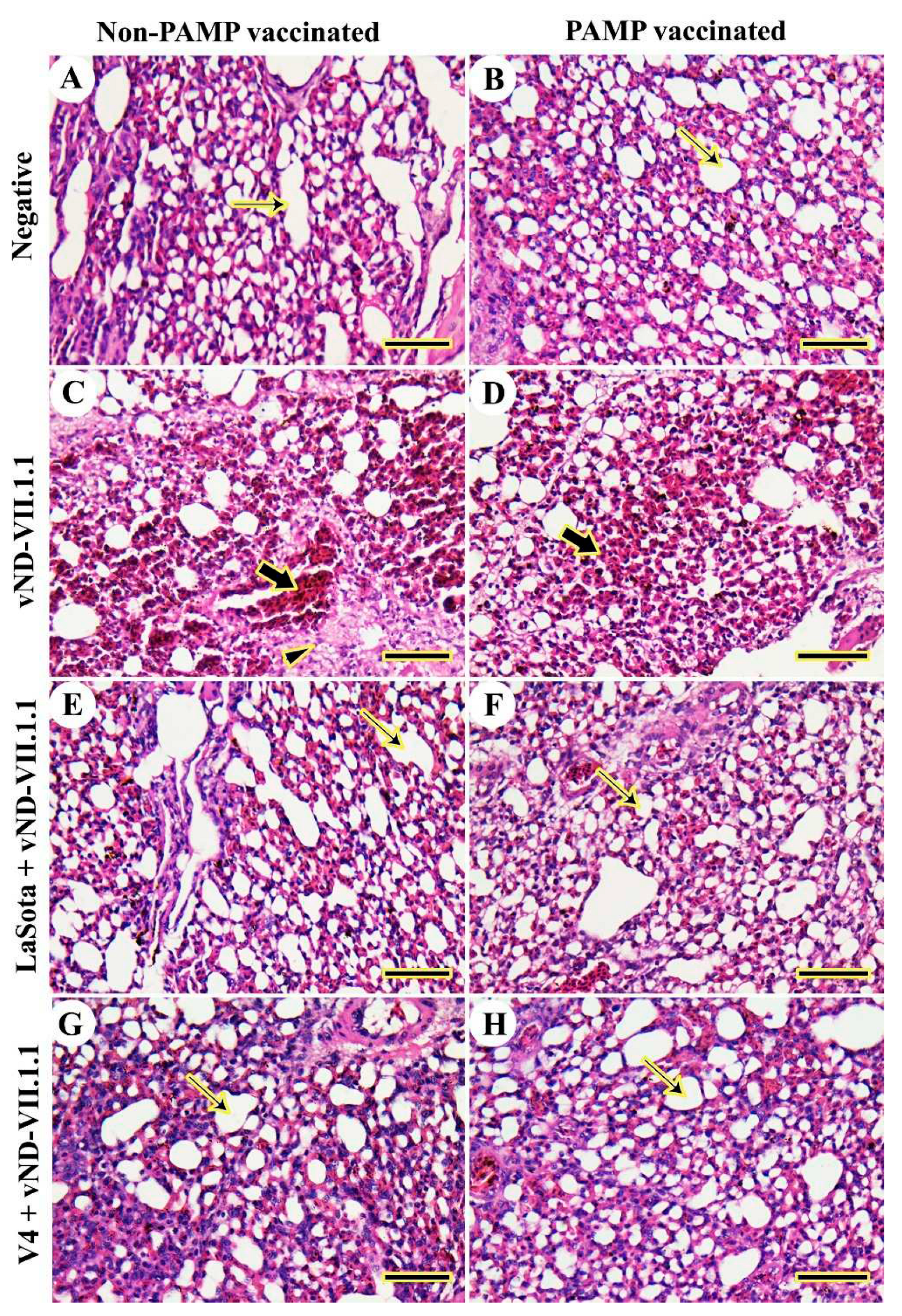
Figure 4.
Histopathological examination of chicken's thymus glands. (A) G8. (B) G4. (C) G7. (D) G3. (E) G6. (F) G2. (G) G5. (H) G1. Cortex (Cx), medulla (Md), Hassell’s corpuscles (thin arrows), severe congestion (arrowheads), and necrosis (thick arrows). Scale bar =50 μm.
Figure 4.
Histopathological examination of chicken's thymus glands. (A) G8. (B) G4. (C) G7. (D) G3. (E) G6. (F) G2. (G) G5. (H) G1. Cortex (Cx), medulla (Md), Hassell’s corpuscles (thin arrows), severe congestion (arrowheads), and necrosis (thick arrows). Scale bar =50 μm.
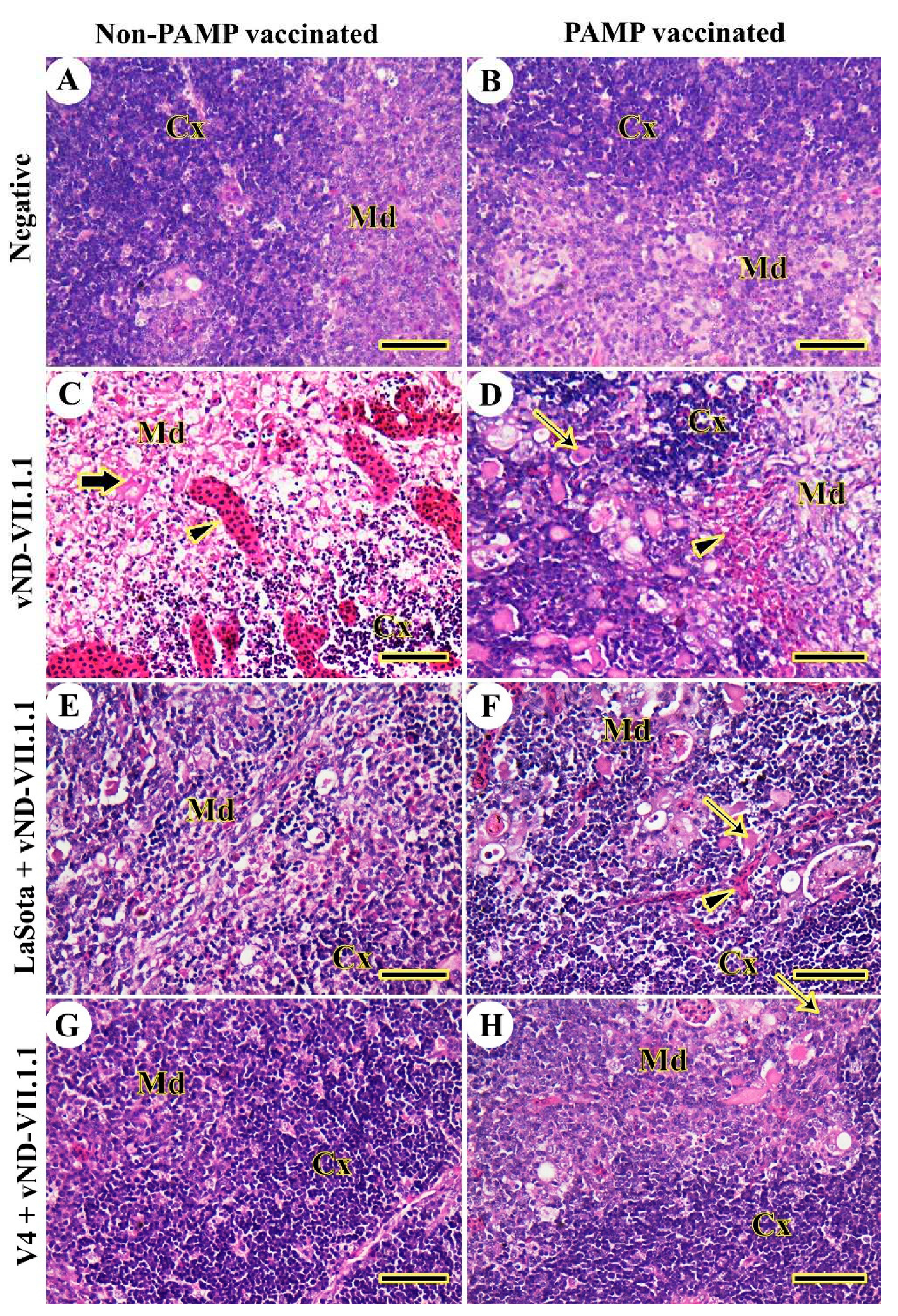
Figure 5.
Histopathological examination of chicken's spleen. (A) G8. (B) G4. (C) G7. (D) G3. (E) G6. (F) G2. (G) G5. (H) G1. Red pulp (Rp), white pulp (Wp), necrosis (thin arrows), and multifocal lymphoid depletion (arrowheads). Scale bar =50 μm.
Figure 5.
Histopathological examination of chicken's spleen. (A) G8. (B) G4. (C) G7. (D) G3. (E) G6. (F) G2. (G) G5. (H) G1. Red pulp (Rp), white pulp (Wp), necrosis (thin arrows), and multifocal lymphoid depletion (arrowheads). Scale bar =50 μm.
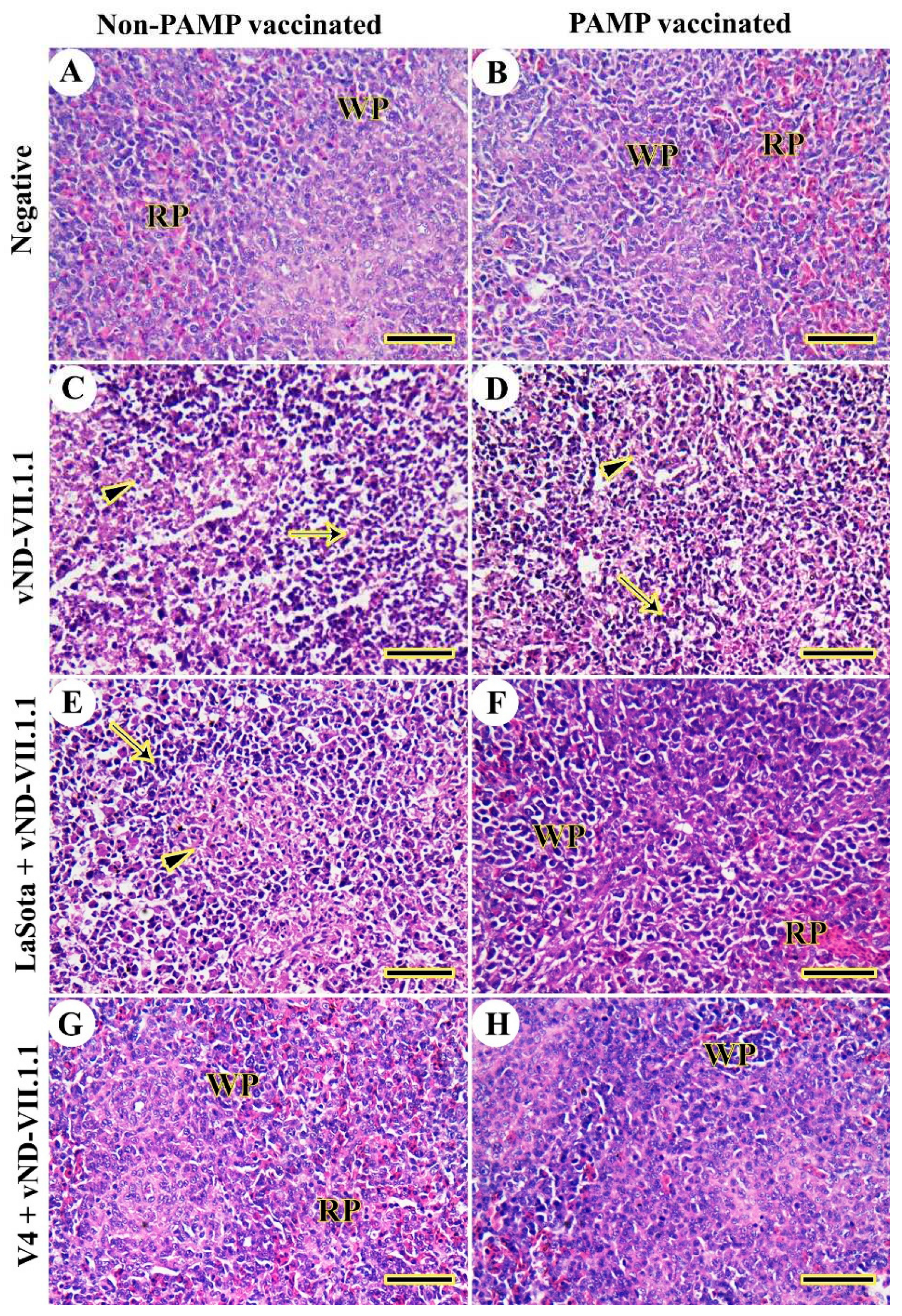
Figure 6.
Histopathological examination of chicken bursa of Fabricius. (A) G8. (B) G4. (C) G7. (D) G3. (E) G6. (F) G2. (G) G5. (H) G1. Lymphoid nodule (LN), normal epithelium (thin arrows), necrosis and lymphoid depletion (thick arrows), and congested blood vessels (arrowheads). Scale bar =50 μm.
Figure 6.
Histopathological examination of chicken bursa of Fabricius. (A) G8. (B) G4. (C) G7. (D) G3. (E) G6. (F) G2. (G) G5. (H) G1. Lymphoid nodule (LN), normal epithelium (thin arrows), necrosis and lymphoid depletion (thick arrows), and congested blood vessels (arrowheads). Scale bar =50 μm.

Figure 7.
Tracheal and cloacal viral shedding in all groups at 3, 6 and 10 dpc.
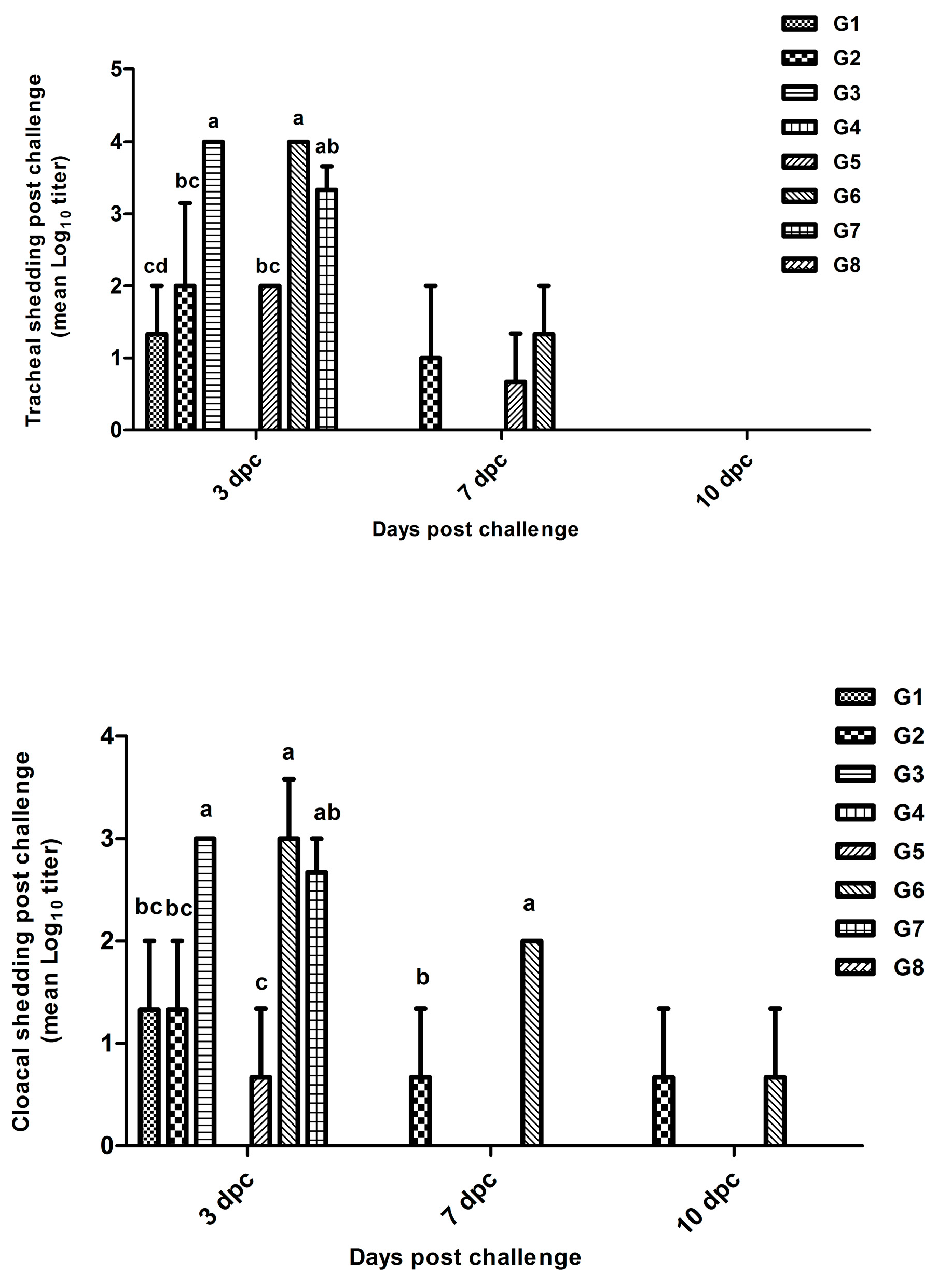
Figure 8.
The detection of IL-4 and IL-10 IF-γ genes expression in the spleen of chickens at 3rd dpc (31 DO) using rRT-PCR.
Figure 8.
The detection of IL-4 and IL-10 IF-γ genes expression in the spleen of chickens at 3rd dpc (31 DO) using rRT-PCR.
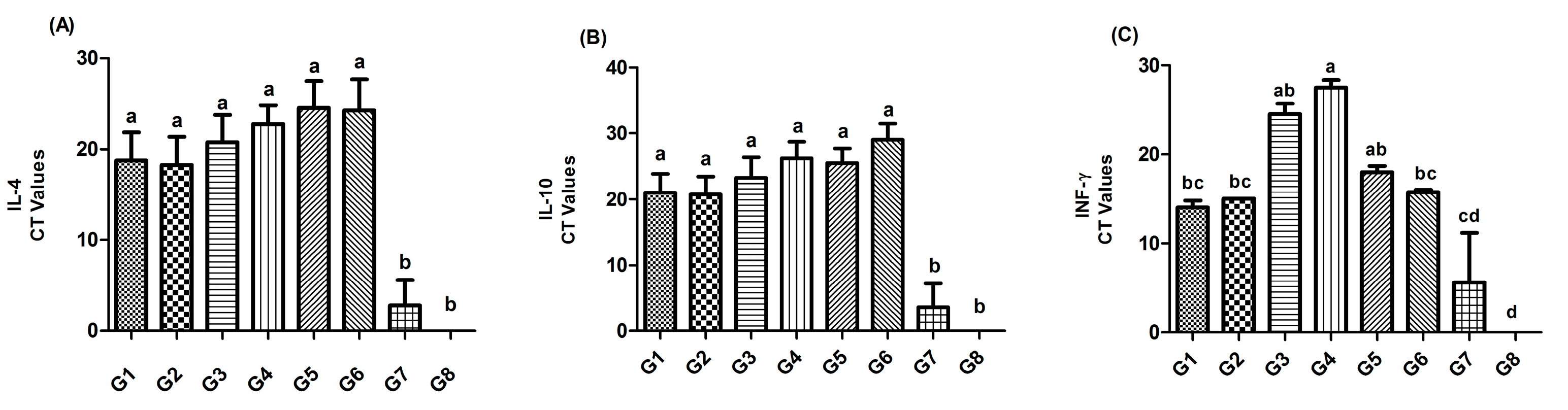

Table 1.
Experimental design.
| Chicken groups (Number) | Age (days) | ||||
| Vaccination | Challenge | ||||
| 4 | 8 | 15 | 28 | ||
| G1 | 25 | PAMP H9 | V4 | V4 | vND-VII.1.1 |
| G2 | 25 | PAMP H9 | LaSota | LaSota | vND-VII.1.1 |
| G3 | 25 | PAMP H9 | - | - | vND-VII.1.1 |
| G4 | 25 | PAMP H9 | - | - | - |
| G5 | 25 | - | V4 | V4 | vND-VII.1.1 |
| G6 | 25 | - | LaSota | LaSota | vND-VII.1.1 |
| G7 | 25 | - | - | - | vND-VII.1.1 |
| G8 | 25 | - | - | - | - |
-: Not applied.
Table 2.
Histopathological lesion scoring of tracheae, lung, thymus glands, spleen, and bursa of Fabricious in all chicken groups.
Table 2.
Histopathological lesion scoring of tracheae, lung, thymus glands, spleen, and bursa of Fabricious in all chicken groups.
| Groups | Organ histopathological lesion score | ||||||
|---|---|---|---|---|---|---|---|
| Trachea | Lung | Thymus glands | Spleen | Bursa of Fabricius | |||
| Hemorrhage | Mucosal hyperplasia and metaplasia | Hemorrhage | Congestion | Necrosis and lymphoid depletion | Necrosis and multifocal lymphoid depletion | Necrosis and lymphoid depletion | |
| 1 | 0.3±0.153de | 0.5±0.167d | 0.30±0.153de | 0.6±0.221c | 0.3±0.153de | 0.6±0.163c | 0.3±0.153de |
| 2 | 2.1±0.233c | 2.5±0.269c | 2.0±0.258c | 3.1±0.233ab | 1.8±0.249c | 2.2±0.2b | 1.7±0.213bc |
| 3 | 2.8±0.2bc | 3.5±0.167ab | 3.4±0.163ab | 2.6±0.163b | 2.5±0.167bc | 3.2±0.327a | 3.3±0.213a |
| 4 | 0.0±0.0e | 0.0±0.0d | 0.0±0.0e | 0.0±0.0c | 0.0±0.0e | 0.0±0.0c | 0.0±0.0e |
| 5 | 1.0±0.149de | 0.6±0.163d | 0.9±0.180d | 0.8±0.249c | 1.0±0.0d | 0.7±0.260c | 1.0±0.258cd |
| 6 | 3.0±0.333ab | 2.9±0.233bc | 2.8±0.291b | 2.9±0.233ab | 3.2±0.249ab | 3.0±0.211ab | 2.4±0.163b |
| 7 | 3.8±0.133ab | 3.9±0.1a | 3.7±0.153a | 3.5±0.167a | 3.7±0.213a | 3.5±0.167a | 3.8±0.2a |
| 8 | 0.0±0.0e | 0.0±0.0d | 0.0±0.0e | 0.0±0.0c | 0.0±0.0e | 0.0±0.0c | 0.0±0.0e |
Means within each column for each division with common superscript letters are significantly different (p ≤ 0.05).
Table 3.
Serological response through HI titers for NDV and LPAIV-H9N2 in all chicken groups.
| Groups | HI titers for ND | HI titers for LPAIV-H9N2 | ||||||||
| Age (days) | Age (days) | |||||||||
| 7 | 14 | 21 | 28 | 35 | 7 | 14 | 21 | 28 | 35 | |
| 1 | 7.38±1.017a | 3.13±0.350bcd | 4.63±0.420a | 4.75±0.366a | 8.00±0.327c | 7.00±0.000b | 4.00±0.267ab | 2.13±0.295ab | 3.25±0.881ab | 7.25±0.559a |
| 2 | 6.88±0.854ab | 3.00±0.000cd | 2.13±0.227bc | 4.13±0.515a | 10.63±0.263ab | 11.13±0.515a | 4.00±0.189ab | 1.63±0.324abc | 1.63±0.460bcd | 6.00±0.567a |
| 3 | 6.50±0.756ab | 3.75±0.313abc | 1.75±0.164c | 1.00±0.000b | 0.00±0.000d (NA) | 6.75±0.412b | 4.13±0.125a | 1.75±0.164abc | 5.00±0.500a | 0.00±0.000b(NA) |
| 4 | 4.50±0.189b | 3.13±0.125bcd | 2.63±0.653bc | 1.38±0.183b | 0.13±0.125d | 7.63±0.625b | 3.75±0.164ab | 1.00±0.000c | 2.88±0.350bc | 6.88±0.833a |
| 5 | 4.63±0.375ab | 3.00±0.000cd | 4.25±0.164a | 5.25±0.313a | 9.38±0.596b | 7.75±0.366b | 3.25±0.164b | 1.75±0.313abc | 1.25±0.164cd | 1.25±0.491b |
| 6 | 5.00±0.378ab | 4.25±0.164a | 4.63±0.460a | 2.13±0.125b | 11.00±0.378a | 7.00±0.000b | 4.00±0.189ab | 1.38±0.183bc | 1.00±0.000cd | 0.25±0.250b |
| 7 | 5.13±0.295ab | 2.63±0.263d | 2.63±0.324bc | 1.50±0.189b | 0.00±0.000d (NA) | 7.00±0.000b | 3.88±0.125ab | 2.13±0.227ab | 0.88±0.125d | 0.13±0.125b(NA) |
| 8 | 5.25±0.412ab | 4.13±0.295ab | 3.63±0.183ab | 1.00±0.000b | 0.38±0.183d | 8.13±0.441b | 3.88±0.227ab | 2.63±0.263a | 1.13±0.125b | 0.13±0.125b |
NA: not applied, means within each column for each division with common superscript letters are significantly different (p≤0.05).
Table 4.
Cytokines parameters through IL-1β, CD4, LYZ, and NO in all chicken groups.
| Age (days) | Groups | ||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| IL1β (pg/ml) |
17 | 280.2±7.838a | 220±4.899d | 274.2±1.960ab | 237.8±13.962cd | 256.6±0.980abc | 243.2±0.735bcd | 260.8±11.758abc | 263.4±0.245abc |
| 21 | 312.2±15.432bc | 376.8±1.715ab | 349.2±22.780abc | 410.8±0.490a | 256.2±46.50c | 377±3.674ab | 406.2±7.838ab | 374.4±22.290ab | |
| 28 | 364.8±13.962ab | 264.6±5.144c | 323.2±0.735a | 306±2.449ab | 256.4±1.470c | 268.6±9.553bc | 258.2±14.207c | 290.6±9.553abc | |
| 31 | 284.4±8.328 | 319.6±18.126 | 318.4±20.576 | 263.8±37.232 | 299.2±7.838 | 255.2±4.164 | 281.4±0.980 | 303.8±6.200 | |
| CD4 (ng/ml) |
17 | 4.4±0.245b | 5.0±0.0ab | 5.0±0.0ab | 4.6±0.245b | 4.4±0.245b | 5.0±0.0ab | 5.0±0.0ab | 5.6±0.245a |
| 21 | 5.0±0.0bc | 6.0±0.0a | 5.6±0.245ab | 6.0±0.0a | 4.6±0.245c | 4.6±0.245c | 5.0±0.0bc | 5.0±0.0bc | |
| 28 | 5.0±0.0c | 5.0±0.0c | 5.0±0.0c | 5.6±0.245bc | 5.2±0.2c | 6.0±0.0ab | 5.6±0.245bc | 6.4±0.245a | |
| 31 | 5.6±0.245b | 5.6±0.245b | 5.0±0.0b | 5.4±0.245b | 6.0±0.0ab | 5.0±0.0b | 6.0±0.0ab | 7.2±0.735a | |
| LYZ (ng/ml) |
17 | 112.2±1.715b | 113.8±0.735ab | 116.6±0.980ab | 115±1.225ab | 115±2.449ab | 111.4±0.980b | 115.6±0.245ab | 119.2±0.490a |
| 21 | 112.2±0.735cd | 115±0.0bc | 109.6±1.470d | 113.4±0.245c | 111.6±0.980cd | 120.6±0.245a | 112.2±0.735cd | 118.2±0.490ab | |
| 28 | 107.2±1.715c | 109±0.0bc | 111.4±2.694bc | 113.8±0.490ab | 106.4±0.245c | 109.2±0.490bc | 111.8±0.800bc | 119.8±1.715a | |
| 31 | 104.4±0.980c | 116.4±0.245bc | 113.4±2.694bc | 117.4±0.490bc | 113.2±0.490bc | 119.8±7.838ab | 132.6±3.429a | 123±0.0ab | |
| NO (μmol/L) |
17 | 62.2±1.715b | 63.8±0.735ab | 66.6±0.980ab | 65±1.225ab | 65±2.449ab | 61.4±0.980b | 65.6±0.245ab | 69.2±0.490a |
| 21 | 62.2±0.735cd | 65±0.0bc | 59.6±1.470d | 63.4±0.245c | 61.6±0.980cd | 70.6±0.245a | 62.2±0.735cd | 68.2±0.490ab | |
| 28 | 57.2±1.715c | 59±0.0bc | 61.4±2.694bc | 63.8±0.490ab | 56.8±0.374c | 59.2±0.490bc | 61.8±0.800bc | 69.8±1.715a | |
| 31 | 54.4±0.980c | 66.4±0.245bc | 63.4±2.694bc | 67.4±0.980bc | 63.2±0.490bc | 69.8±7.838ab | 82.6±3.429a | 73±0.0ab | |
Means within each row for each division with common superscript letters are significantly different (p≤0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



