
Submitted:
10 September 2023
Posted:
12 September 2023
You are already at the latest version
Abstract
The root-colonizing endophytic fungus Piriformospora indica promotes root and shoot growth of its host plants. We show that growth promotion of Arabidopsis leaves is abolished when the seedlings are grown on media with nitrogen (N) limitation. The fungus neither stimulated the total N content nor did it promote 15NO3- uptake from agar plates to the leaves of the host under N-sufficient or N-limiting conditions. However, when the roots were co-cultivated with 15N-labelled P. indica, more label can be detected in the leaves of N-starved host plants, but not of plants supplied with sufficient N. Amino acid and primary metabolite profiles, as well as expression analyses of N metabolite transporter genes suggest that the fungus alleviates the adaptation of its host to the N limitation condition. P. indica alters the expression of transporter genes which participate in relocation of NO3-, NH4+ and N metabolites from the roots to the leaves under N limitation. We propose that P. indica participates in the plant´s metabolomic adaptation to N limitation by delivering reduced N metabolites to the host, alleviating metabolic N starvation responses, and reprogramming the expression of N-metabolism related genes.
Keywords:
Piriformospora indica
; nitrogen starvation
; nitrogen metabolism
; nitrate transporter
; ammonium transporter
; amino acid transporter
; endophyte
1. Introduction
Nitrogen is a key mineral nutrient playing a crucial role in plant growth and development. The soil microbiome contributes to nitrogen acquisition, and among the best studied endosymbiotic interactions are those with N-fixing rhizobia and arbuscular mycorrhizal (AM) fungi. Legumes gain access to N through symbiotic association with rhizobia which convert N2 gas into ammonia in nodules. Although several efforts have been made to incorporate biological N fixation capacity into non-legume plants [1], agricultural crop production without N fertilization is currently not conceivable. AM fungi help plants in nutrient acquisition and much progress has been made in understanding the molecular basis of P and N transfer from the fungal partner to the host plant [cf. 2]. Less is known about endophytes, although they show relatively little host specificity and have therefore a great potential for agricultural applications [3].
A well-studied endophytic fungus is Piriformospora (Serendipita) indica, which interacts with numerous host plants and promotes their growth and resistance against biotic and abiotic stresses [4, 5]. Stimulation of growth of its hosts suggests that the fungus promotes nutrient acquisition, including nitrogen. An effect of P. indica on nitrate uptake and the nitrogen metabolism in the hosts has been reported repeatedly. On full medium, the fungus promotes nitrogen accumulation and the expression of nitrate reductase in Arabidopsis [6]. In sunflower, P. indica increases the absorption of nitrogen by the root [7]. Strehmel et al. [8] showed that the concentration of nitrogen-rich amino acids decreased in inoculated Arabidopsis plants. Ghaffari et al. [9] proposed that the nitrogen metabolism plays an important role in systemic salt-tolerance in leaves of P. indica-colonized barley. Furthermore, Lahrmann et al. [10] showed that the P. indica ammonium transporter Amt1 functions as a nitrogen sensor mediating the signal that triggers the in planta activation of the saprotrophic program. In Chinese cabbage, especially the amino acid γ-amino butyrate is de novo synthesized in colonized roots [11]. Bandyopadhyay et al. [12] demonstrated that P. indica together with Azotobacter chroococcum facilitates higher acquisition of N and P in rice. P. indica also improves chickpea productivity and N metabolism in a tripartite combination with Mesorhizobium [13]. Finally, Serendipita williamsii does not affect P status but C and N dynamics in AM tomato plants [14]. These examples highlight the importance of the N metabolism on numerous beneficial effects of P. indica for different plant species, however, how the fungus influences the host N metabolism is not clear. In this study, we use the model plant Arabidopsis thaliana to investigate how P. indica interferes with N uptake and metabolism under N limiting conditions.
2. Results
2.1. Shoot growth promotion by P. indica requires external N supply
P. indica colonizes Arabidopsis roots and induces visible growth promotion of Arabidopsis seedlings after 4-7 days in full medium [15]. After 5 days, the fresh weight of the shoots was significantly increased (+ 41.9 %) by the fungus, while barely any growth promotion was detectable on N-limited medium (Figure 1 shoot). Root growth was neither affected by the fungus nor by N availability (Figure 1 root). We conclude that under these experimental conditions, shoot, but not root growth of Arabidopsis seedlings is promoted by P. indica, and this requires N in the medium.
2.2. P. indica colonisation did not change the total N content in the shoots and transfer of 15N from the medium to the shoots
To test whether P. indica interferes with N accumulation or uptake into the plant under N-limiting conditions, the total N content in the shoots and the amount of 15N from 15NO3--labelled growth medium in the shoots were compared for uncolonized and colonized seedlings, grown on either full or N-limiting media. As expected, the total N content in shoots of seedlings which were exposed to N limitation, was lower than in the shoots of seedlings grown on full medium (Figure 2 left). Furthermore, accumulation of 15N in the shoots was much higher on full medium than on medium with low N (Figure 2 right). However, we did not observe significant differences for uncolonized and colonized seedlings. This suggests that the fungus does not stimulate nitrate uptake from the medium under N-sufficient and N-limiting growth conditions.
N limitation might influence the colonisation of the roots. We observed that roots on N-limiting conditions were ~ 2-times more colonized than roots on full medium (Supplemental Figure S1), although the difference was not significant. This indicates that in spite of a higher colonisation rate, transport of 15N label from the 15NO3- containing medium to the leaves was not stimulated by the fungus under N-limiting conditions.
2.3. 15N label is transferred from P. indica to the host under N-limiting conditions
Since P. indica did not promote NO3- uptake, we tested whether labelled 15N metabolites are translocated from the fungus to the plant. As shown in Figure 3A, P. indica was cultured on 15N-containing medium for 14 days before co-culture with Arabidopsis seedlings on full or N-limited media.
The 15N-labelled fungal mycelium was positioned about 1 cm away from the roots. Establishing contact between the two partners and initiation of root colonisation started approximately 24 h later, after the growing hyphae have reached the roots (Figure 3A). Since label could be detected in the aerial parts of all analysed seedlings which were in contact with P. indica, the fungus transfers N-containing metabolites to the roots of its host, and the label is further translocated to the aerial parts of the seedlings (Figure 3B). Interestingly, approximately 4-times more 15N accumulated in the aerial parts of the plants under N-limiting conditions (Figure 3B). This indicates that the fungus helps the host with reduced N metabolites to compensate N limitation during growth on NO3--limiting medium.
2.4. Reprogramming of the metabolite profiles to N limitation conditions is alleviated by P. indica
Next, we tested whether the fungus affects the host´s N metabolism under sufficient N and N-limitation conditions. We measured the levels of primary metabolites by GC-MS in the rosettes after 2 days of transfer to N-limitation condition compared to N-sufficient condition in the absence and presence of P. indica (Supplementary Table S1). We then calculated the metabolite ratios for plants grown under limiting versus sufficient N and compared these ratios for plants grown in the absence and the presence of the fungus. Although the amino acid profiles were comparable for colonized and uncolonized shoots, we observed slight differences for several amino acids (Table 1A). The content of aspartate and alanine decreased under N-limiting condition in both the absence and the presence of the fungus, and these decreases were less pronounced in colonized shoots (Figure 4). Similar tendencies were observed for amino acid contents that increased under N-limitation condition; these increases were less distinct in the presence of the fungus in the case of isoleucine, lysine, tryptophan, phenylalanine, leucine, and arginine (Figure 4). The alterations in serin contents that were triggered by N limitation varied strongly in colonized plants in comparision to uncolonized plants. We then analysed the effect of N-limitation on soluble sugars (Table 1B). In the presence of P. indica N-limitation triggered stronger increases of monosaccharides in particular of glucose and fructose. The stress related sugars trehalose and raffinose showed strong variation between the 3 independent replicates, however raffinose tends to accumulate to higher levels under N limitation when the roots were colonized (cf. Discussion). Overall, the slight alteration of the metabolite profiles in response to N-limitation by the colonisation with P. indica suggest a lessening of the effects of N limitation on several steps of central metabolism.
2.5. P. indica stimulates expression of specific host´s transporter genes under N limitation
Incorporation of 15N into the aerial parts of colonized seedlings is ~4-fold higher under N starvation when compared to seedlings grown on full medium (Figure 2), and a comparable stimulation is observed for the translocation of labelled 15N from P. indica to the leaves under N limitation (Figure 3). To test whether genes for N metabolite transporters are regulated by P. indica, we performed expression profiles with RNA from roots and shoots of seedlings which were either grown on full or N-limitation medium in the presence or absence of P. indica (Table 2).
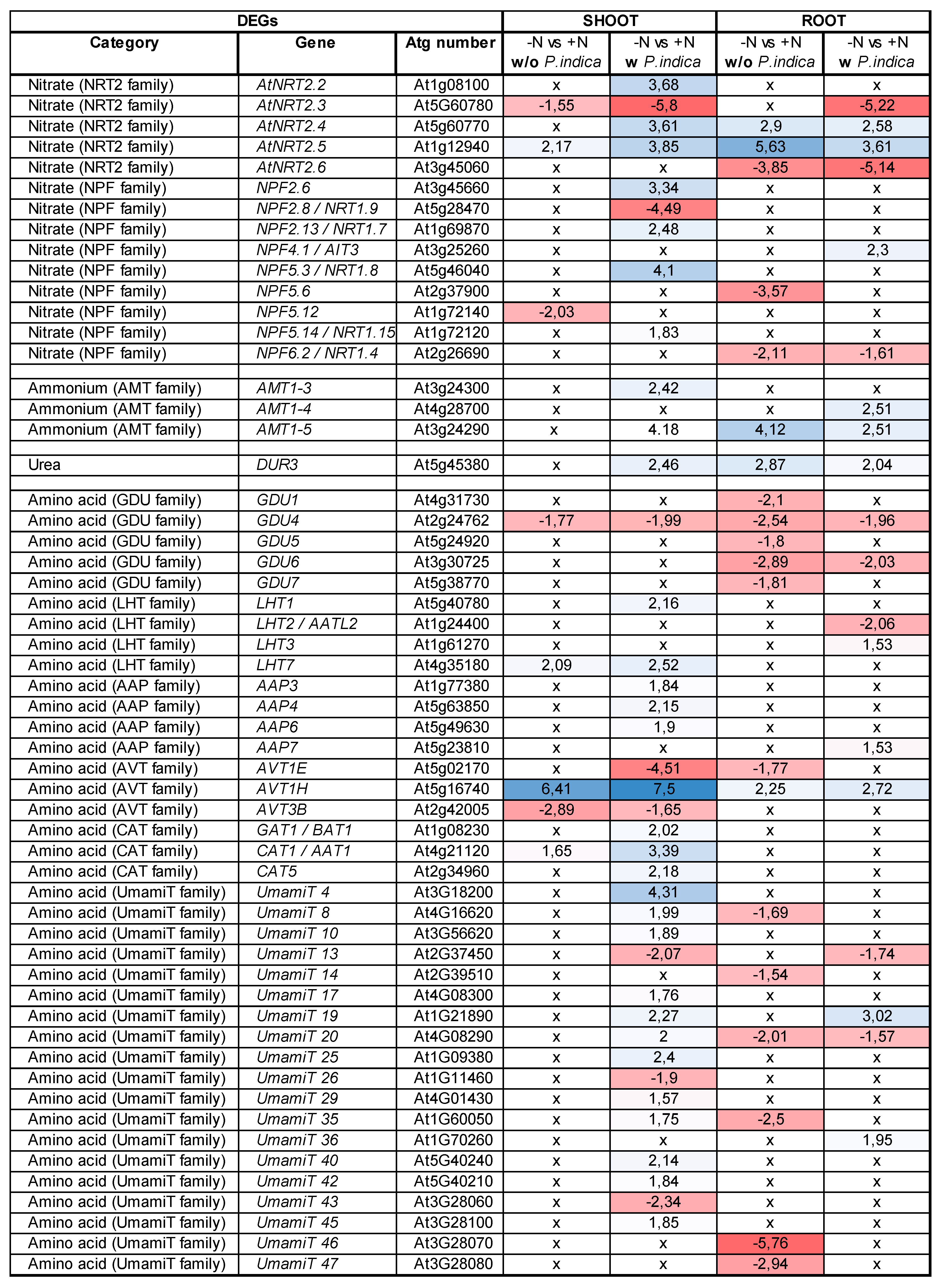
Among the 56 investigated genes, which code for NO3-, NH4+, amino acid or peptide transporters, 33 genes were differentially expressed in either roots and shoots or both of colonized and uncolonized seedlings grown on full or N-limited media (Table 2). In the shoots, this included genes for two NH4 transporters (AMT1-3 and AMT1-5), three NO3- transporters (NRT2.2, NRT2.4 and NRT2.5), 5 members of the NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTERs gene family (NPF2.6, NPF2.13, NPF5.3, NPF5.12 and NPF5.14) as well as the urea transporter DUR3. Furthermore, 21 amino acid transporters, including members of the LHT and AAP families, as well as 12 UmamiT putative amino acid transporters responded to the fungus. In contrast, 7 transporter genes were downregulated by P. indica in the shoots of N-starved seedlings (Table 2). In the roots, six of these genes showed the same regulation (Table 2). This clearly demonstrates that the expression of genes for NO3-, NH4+, amino acid and peptide transporters are major targets of the fungus under N limitation conditions (cf. Discussion).
3. Discussion
N limitation has severe consequences for plant performance [16], and endophytes may help plants to better adapt to the shortage. We used the well-investigated symbiotic interaction between the model plant Arabidopsis and P. indica to address this question. We demonstrate that under severe N limitation, the fungus does not stimulate the uptake of nitrate into the host plant, but rather N-label from fungal metabolites appears in the leaves of the host. Since our N limiting medium contains barely any nitrate, the absence of a detectable stimulatory effect of the fungus on nitrate uptake into the host is not surprising. The N metabolites which are translocated from the hyphae to the plants under N limitation did not results in fungus-induced growth promotion (Figure 1), suggesting that the N supply to the host by the fungus might only compensate deficits. Furthermore, N-translocation from the fungus to the host occurs only under N limitation conditions suggesting the involvement of an N sensing system [cf. 10]. Successful transfer of 15N by arbuscular mycorrhizal fungi to host plants has been shown earlier [17-19]. More recently, Hoysted et al. [20] investigated clover (Trifolium repens) colonized by Mucoromycotina fungi and showed that the host gained both 15N and 33P tracers directly from the fungus in exchange for plant-fixed C. Whether the N supply to the host in our study system has comparable symbiotic features with profit for both partners or it is just a stress-related withdrawal of N from the fungus by the plant without any profit for the microbe remains to be investigated. However, since the fungus can grow and propagate on the host under our –N conditions, the N translocation to the host does not restrict hyphal growth. It appears that the conditions are not strong enough to induce changes in the symbiotic interaction [10]. It is also not clear which metabolites and how they are transported from the microbe to the plant. In Medicago truncatula, three AMT2 family ammonium transporters (AMT2;3, AMT2;4, and AMT2;5) are involved in the uptake of N in form of ammonium from the periarbuscular space between the fungal plasma membrane and the plant-derived periarbuscular membrane [21]. In exchange, host plants transfer reduced carbon to the fungi [22-24]. Also Cope et al. [25] showed that colonization of M. truncatula with R. irregularis led to an elevated expression of the mycorrhiza-induced AMT2;3 and the nitrate transporter NPF4.12 as well as the putative ammonium transporter NIP1;5 in the roots. A dipeptide transporter from the arbuscular mycorrhizal fungus Rhizophagus irregularis is upregulated in the intraradical phase [26]. To investigate how the fungus manipulates the host N metabolism we performed a comprehensive metabolome and transcriptome analysis for N-related metabolites and genes (Table 2 and Table 3).
No major impact of the colonization by P. indica on the changes of shoot metabolite levels in response to N limitation has been observed in this study. Liu et al. [27] demonstrated that raffinose positively regulates maize drought tolerance by reducing leaf transpiration. The raffinose family oligosaccharides are associated with various abiotic and biotic stress responses in different plant species [e.g., 28-32]. It is conceivable that the stimulatory effect of P. indica on the raffinose level in N-limited leaves reduces stress.
Nitrate transporter genes are often upregulated under N starvation, however, the role of endophytic microorganisms in nitrate acquisition is not fully understood. In rice, the arbuscular mycorrhizal fungus R. irregularis remarkably promoted growth and N acquisition, and about 42% of the overall N could be delivered via the symbiotic route under nitrate limiting condition [33]. Nitrate uptake occurs via NITRATE TRANSPORTER1/PEPTIDE TRANSPORTER FAMILY (NPF)4.5, a member of the low affinity nitrate transporter family which is exclusively expressed in arbuscles of Gramineous species [33]. A comparable mechanism does not exist in our endophyte/Arabidopsis model, and the putative Arabidopsis NPF4.5 homolog is not upregulated in colonized roots under N limitation. However, we observed a highly specific response of several NPF/NRT1 and NRT2 family members to P. indica colonisation which allow conclusions how the fungus interferes with the plant N metabolism. The nitrate transporters NRT2.2 and NRT2.4 [34] are only upregulated in the rosettes when the roots are colonized by P. indica, while their expression in the roots is not responding to the fungus. This suggests that the fungus promotes nitrate scavenging that is released from the vacuole in response to N starvation. In fact, NRT2.4 has been shown to be expressed close to the phloem in rosettes and to contribute to nitrate homeostasis in the phloem under limiting nitrate supply, since in nitrate-starved nrt2.4 mutants, nitrate content in shoot phloem exudates was decreased [34]. Likewise, NRT1.7 (NPF2.13) and NRT1.8 (NPF5.3) are upregulated by P. indica in leaves, but not in roots. NRT1.7 loads excess nitrate stored in source leaves into phloem and facilitates nitrate allocation to sink leaves. Under N starvation, the nrt1.7 mutant exhibits growth retardation, indicating that NRT1.7-mediated source-to-sink remobilization of stored nitrate is important for sustaining growth in plants [35]. NRT1.8 is expressed predominantly in xylem parenchyma cells within the vasculature and functional disruption of NRT1.8 significantly increased the nitrate concentration in xylem sap [36]. In contrast, NRT2.3 and -2.6 are down-regulated under N limiting conditions and this is further promoted by the fungus. NRT2.6 has been linked to biotic and abiotic stress responses [37], and it appears that downregulation of NRT2.6 expression by P. indica alleviates the stress responses in the roots. Finally, NRT1.9 (NPF2.9) is strongly downregulated by P. indica in the leaves. NRT1.9 is expressed in the companion cells of phloem. In nrt1.9 mutants, downward nitrate transport was reduced, suggesting that NRT1.9 facilitates loading of nitrate into the phloem and enhances downward nitrate transport to the roots [38], apparently a process that is restricted by the fungus. Taken together, the analysis of the regulation of the Arabidopsis nitrate transporter genes by P. indica suggests that the root-colonizing fungus supports nitrate transport to and availability in the aerial parts of the host under our nitrate limiting conditions. This is further supported by the up-regulation of NRT1.15 (NPF5.14) by P. indica in the leaves. NRT1.15 is a tonoplast-localized low-affinity nitrate transport [39] and overexpression of the gene significantly decreased vacuolar nitrate contents and nitrate accumulation in Arabidopsis shoots. NRT1.15 regulates vacuolar nitrate efflux, and the reallocation might also contribute to osmotic stress responses other than mineral nutrition [39].
Since the medium does not contain NH4+, the plant can only receive NH4+ from the fungus via ammonium transporters (AMTs) [35,40,41]. AMT1-4 expression is upregulated by P. indica in roots under N limitation. Since AMT1-4 is root-specific [42], this suggests that the plant tries to compensate its N limitation by stimulating NH4+ uptake. NH4+ might also originate from the fungus and it is conceivable that withdrawal of this ion from the fungus might ultimately result in a change of the symbiotic interaction towards saprophytism [cf. 10]. Furthermore, expression of AMT1-3 and AMT1-5 as well as DUR3 coding for an urea transporter [43,44] is stimulated by P. indica in the shoots. Root colonization might create a metabolite environment in the host that requires these transporters for proper distribution of the N metabolites in the aerial parts.
Seven amino acid transporters are regulated >log2-fold by P. indica colonisation in nitrate-deprived Arabidopsis seedlings. In roots, the fungus prevents down-regulation of the gene for glutamine secreting GLUTAMINE DUMPER (GDU)1 [45] suggesting that the microbe wants to become access to the plant glutamine. Furthermore, the broad-specificity high affinity amino acid transporter LYSINE HISTIDINE TRANSPORTER (LHT)1 [46], AMINO ACID PERMEASE (AAP)4, γ-AMINOBUTYRIC ACID TRANSPORTER (GAT)1 and CATIONIC AMINO ACID TRANSPORTER (CAT)5 are upregulated in the leaves of P. indica-colonized seedlings. These transporters have been proposed to be involved in nitrogen recycling in plants [47]. Apparently, a better or different N metabolism management is required for the plant when the roots are associated with the endophyte. LHT1 and -2 are also involved in the transport of 1-aminocyclopropane carboxylic acid, a biosynthetic precursor of ethylene [48] which might indicate an increased stress by the interaction with the fungus under N limiting conditions. An involvement in nitrogen recycling has also been proposed for 5 of the 10 USUALLY MULTIPLE ACIDS MOVE IN AND OUT TRANSPORTERS (UMANIT13, -20, -40, -45 and -47) [47]. which are regulated >log2-fold in either roots or shoots of P. indica-colonized seedlings under N limitation.
4. Materials and Methods
4.1. Plant and fungus material, corresponding growth conditions
Arabidopsis thaliana seeds (Col-0) were surface-sterilized and sown on N-free MGRL medium supplemented with 2.5 mM NH4NO3 and 3 g/L gelrite [49]. The KNO3 and Ca(NO3)2 in the MGRL medium were replaced by KCl and CaCl2 to ensure ion equilibrium. After 48 h of stratification at 4 °C in the dark, the seeds were transferred to long-day conditions with 22 °C, 16 h light/8 h dark, 80 μmol m−2 s−1 for 10 days.
Piriformospora indica was cultured on Kaefers medium as described before [50,51]. As described previously, plugs of a 4-week-old fungal culture were used for co-cultures with the seedlings. The fungus was pre-grown for 7 days on PNM medium (PNM+N) with a nylon membrane in the dark at 22 °C. For N limiting conditions (0 mM total N, PNM-N), KNO3 and Ca(NO3)2 were replaced by KCl and CaCl2. For control plates without fungus, only empty KM plugs were placed on top of the membrane.
4.2. Plant-fungus co-cultures and determination of growth promotion
For plant-fungus co-cultures for 5 days, 4 plants (per petri dish) were placed on top of the pre-grown fungal lawn as described previously [51], with some adaptations. Plates were sealed with 3MTM Micropore tape to reduce the condensation and 10 d-old plants were used for co-cultivation, to reduce the amount of N which accumulates in the plants on MGRL medium before the co-culture. In pilot experiments, we showed that the reduced age did not affect the establishment of the symbiosis with the fungus. The co-cultures were incubated at 22 °C, 16 h light/8 h dark, 80 μmol m−2 s−1 with light from the top.
After 5 days, roots and shoots of the plants were harvested separately. For that, 5 plates (= 20 plants) were harvested as 1 sample. Both roots and shoots were washed in sterile distilled water and carefully dried before weighing and direct freezing in liquid nitrogen. Samples were stored in - 80 °C until further use. These experiments were repeated 3-4 times independently.
To determine growth promotion by the fungus, the weight of the sample with fungus was normalized (divided) to the weight without fungus. This was done for total weights sampled from full medium (PNM+N) as well as from N-limited medium (PNM-N). Final growth promotion values presented in the figures are averages of 3 replicates from independent cultures.
4.3. 15N Labeling experiments in the medium
To analyze the uptake of nitrogen by the plant, 2.5 % of the total KNO3 (which equals 0.125 mM KNO3) of the PNM medium was replaced by K15NO3 (Eurisotop, Saint-Aubin, France) dissolved in distilled water. For proper comparison, the 2.5 % of K15NO3 was also added to the N-free medium (PNM-N) resulting in a final concentration of 0.125 mM nitrate. Finally, PNM-N control plates without 15N were used and contained 0.125 mM unlabeled KNO3. Plants grown on these plates were compared to those grown on PNM+N plates to analyze the natural abundance of 15N in the plant tissue. As described before, the fungus or control plug was pre-incubated on the PNM with nylon membrane for 1 week before plants were placed on the plates. The co-cultures were incubated for 5 days to ensure that enough 15N was taken up by the plant.
4.4. 15N fungus labeling experiments
To analyze if the fungus can directly transfer N or N-containing metabolites to the plant, it was labeled with 15N before the co-culture. A modified KM medium without the N-containing components (20 g/L dextrose, 50 mL/L macronutrients, 10 mL/L micronutrients and 1 mL/L Fe-EDTA, 1 mL/L vitamin mix, pH 6.5) was prepared and supplemented with 10g/L ISOGRO®-15N (CortecNet, Les Ulis, France) according to the manufacturers protocol. P. indica plugs of 2 mm diameter were incubated (23 °C, 50 rpm, dark) in 2 mL of KMISOGRO in Greiner CELLSTAR® 12-well plates (Greiner Bio-One, Frickenhausen, Germany) sealed with 3MTM Micropore tape. After 14 days of growth, the fungal tissue was separated from the remaining medium and carefully washed 3 times with N-free liquid PNM to remove 15N bound to the hyphal surface. A 76.66 % enrichment in 15N was achieved using this protocol. The fungus was carefully cut in 5x5 mm pieces and placed on PNM-N and PNM+N plates to start the co-cultures. To minimize 15N uptake by the plant from dead fungal material due to the washing and handling procedure, the fungal plugs were placed in minimum 1 cm distance from the roots. Under these conditions, contact between the two symbionts requires active growth of the hyphae towards the roots. Co-cultivation was performed with 3 plants per plate for 14 days to ensure that enough 15N was taken up by the plant.
4.5. Isolation and clean-up of RNA
Samples of root or shoot material were stored in -80 °C. For homogenization, the samples were ground with mortar and pistil in liquid nitrogen. A maximum of 100 mg material was used for RNA extraction. RNA was extracted with TrizolTM (ThermoFisher Scientific, Waltham, USA) and chloroform according to the manufacturers protocol. Briefly, the plant material was mixed with 1 mL of TrizolTM and incubated on a shaker at room temperature for 15 min. After addition of 250 μl chloroform and a second incubation phase, the sample was centrifuged (30 min, 4 °C). The supernatant was mixed with isopropanol and incubated on ice, followed by centrifugation. The pellet was washed twice with 80 % ethanol, dried and resuspended in RNAse-free water. The RNA isolation was followed by an additional cleaning step to remove access salts originating from the fungus tissue. For that, the sample was mixed with 3 M sodium acetate [1/10 (v/v) in RNAse-free water, pH= 5.2] and 600 µL of ice-cold 100 % ethanol and incubated at -20 °C for at least 1 hr. After centrifugation and 2 cleaning steps with 80 % ethanol the sample was resuspended in RNAse-free water. The quality and concentration of the extracted RNA was tested by absorbance analysis using a NanoVue (GE Healthcare, Uppsala, Sweden).
4.6. RNAseq and data analysis
After transfer of samples to Novogene Genomics Service (Cambridge, UK), the RNA sample integrity was checked with a Bioanalyzer 2100 (Agilent). After samples passed the quality check, the service laboratory proceeded with the library construction and RNA sequencing (PE150) on Illumina NovaSeq™ 6000 platforms as described in a previous study [52].
The RNAseq libraries were filtered and quality-trimmed with fastp (v0.23.2) [53], i.e., read ends were truncated to achieve a Phred quality score of 30 or more. Reads below 15 nt length or those comprising at least 2 ambiguous N bases were removed from the dataset. Read qualities were monitored by FastQC (v0.11.3; https://www.bioinformatics.babraham.ac.uk/projects/fastqc/). Hisat2 (v2.2.1) [54] was used with default parameters to map the quality-trimmed RNAseq libraries to the A. thaliana reference genome (TAIR10, Ensembl release 51). The mapping allowed spliced reads and single-read mapping to multiple best-fitting locations. FeatureCounts (v1.5.3) [55] was applied to perform read counting based on the A. thaliana reference annotation (TAIR10, Ensembl release 51). The Bioconductor DESeq2 (v1.10.0) package [56] was utilized to identify DEGs in the different pairwise mutant and wild type comparisons. Benjamini and Hochberg’s False Discovery Rate (FDR) approach [57] was employed to adjust the calculated p-values for multiple testing.
To identify DEGs of transporters predicted to transport major N compounds, the obtained results were in a first step filtered according to their p-value (p<0.05). Next, DEGs were sorted according to their log2-fold change, here only changes with numbers >= +1.5 and <= -1.5 were further analyzed. This list was cross-checked with targets identified from a search in the UniProt database (https://www.uniprot.org) using keywords like “NH4 transport”.
4.7. Analysis of fungal colonization by qPCR
1 mg of RNA was used for the synthesis of cDNA. The Omniscript RT Kit (Qiagen, Hilden, Germany) was used according to the manufacturers protocol with the oligo(dT)18 primer (ThermoFisher Scientific, Waltham, USA). qPCR was performed with fifty nanograms of the synthesized cDNA as template in a Bio-Rad CFX96 Real-Time PCR Detection System (Feldkirchen, Germany) by use of DreamTaq Polymerase (ThermoFisher Scientific, Waltham, USA) and Evagreen (Biotium, Fremont, USA). The data were normalized with respect to the Arabidopsis RPS18B (At1g34030) gene by the 2-∆∆CT method [58]. To quantify the P. indica colonization level of Arabidopsis roots, the expression of PiTEF1 [59] was analysed in comparison to the plant´s housekeeping gene RPS18B (At1g 34030). Following primers were used: PiTEF1: CGCAGAATACAAGGAGGCC and CGTATCGTAGCTCGCCTGC; RPS18B: GTCTCCAATGCCCTTGACAT and TCTTTCCTCTGCGACCAGTT [60]. The colonization was compared between plants grown on PNM-N and PNM+N media (set as 1.0), by the 2-∆∆CT method.
4.8. Determination of total nitrogen and 15N enrichment
Total N and 15N contents were quantified on 1–2 mg aliquots of dry tissue, after drying a ground tissue aliquot at 65-70 °C for at least 48 h. N elements were detected by gas chromatography on a FLASH 2000 Organic Elemental Analyzer (Thermo Fisher Scientific, Villebon, France). The 15N/14N isotopic ratio was subsequently quantified by a coupled mass spectroscope (Delta V advantage IRMS; Thermo Fisher Scientific, Villebon, France). The total N content was only determined in plant shoots, because a discrimination of N from plant or fungus was not possible in root material.
4.9. Metabolomic analysis
For GC-MS-based quantifications, 25 mg of finely ground plant material was resuspended in 1 mL of frozen (-20 °C) water: acetonitrile: isopropanol (2:3:3, v/v/v) containing Ribitol at 4 ug/mL and analysed as described in [61].
5. Conclusion
We observed an unexpected complexity in the plant N metabolism when N-deprived Arabidopsis seedlings are colonized by P. indica. Our initial observations highlight a few aspects which need to be investigated in greater details. (1) Which N metabolites are transported from the fungus to a plant suffering under N limitation? (2) The plant appears to adapt its N metabolism under N limitation by transporting N metabolites shootwards, a process that is supported by the fungus. Is the fungal support for the plant specific for the symbiotic phase of the interaction? (3) Are our observations P. indica-specific or do they occur also in other endophyte/plant interactions?
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
S.S., E.B., G.C., A.M. and A.K. performed the experiments and analysed the data, J. L-M., H. S., T. K., J. V.-C, S. P., A. K. and R.O. developed the concept and wrote the manuscript.
Funding
This project was financially supported by the collaborative ICPS research project executed in the framework of the EIG CONCERT-Japan joint call on Food Crops and Biomass Production Technologies and the related national funding agencies: grants 01DR17007A and 01DR17007B from the Federal Ministry of Education and Research (BMBF), Germany, to R. O., respectively; grant EIG_JC1JAPAN-045 from the Centre National de la Recherche Scientifique (CNRS), France, to A. K.; grant PCIN-2016–037 from the Ministry of Economy and Competitiveness (MINECO), Spain, to J. V. C. and S. P.; and grant JPMJSC16C3 from the Japan Science and Technology Agency (JST) to H. S. This work was further supported by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) CRC1127 ChemBioSys (project ID: 239748522) to R. O. This work has benefited from the support of IJPB's Plant Observatory technological platforms. The IJPB benefits from additional support of Saclay Plant Sciences-SPS (ANR-17-EUR-0007).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data set is available in the NCBI GEO repository, under accession number GSE239281.
Acknowledgments
We thank Sarah Mußbach, Claudia Röppischer and Christin Weilandt for excellent technical support in the lab.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Guo, K.; Yang, J.; Yu, N.; Luo, L.; Wang, E. Biological nitrogen fixation in cereal crops: Progress, strategies, and perspectives. Plant Commun. 2023, 4(2),100499. Epub 2022 Nov 28. PMID: 36447432; PMCID: PMC10030364. [CrossRef]
- Rui, W.; Mao, Z.; Li, Z. The Roles of Phosphorus and Nitrogen Nutrient Transporters in the Arbuscular Mycorrhizal Symbiosis. Int. J. Mol. Sci. 2022, 23(19),11027. PMID: 36232323; PMCID: PMC9570102. [CrossRef]
- Chaudhary, P.; Agri, U.; Chaudhary, A.; Kumar, A.; Kumar, G. Endophytes and their potential in biotic stress management and crop production. Front. Microbiol. 2022, 13, 933017. PMID: 36325026; PMCID: PMC9618965. [CrossRef]
- Saleem, S.; Sekara, A.; Pokluda, R. Serendipita indica-A Review from Agricultural Point of View. Plants (Basel). 2022, 11(24), 3417. PMID: 36559533; PMCID: PMC9787873. [CrossRef]
- Del Barrio-Duque, A.; Ley, J.; Samad, A.; Antonielli, L.; Sessitsch, A.; Compant, S. Beneficial Endophytic Bacteria-Serendipita indica Interaction for Crop Enhancement and Resistance to Phytopathogens. Front. Microbiol. 2019, 10, 2888. PMID: 31921065; PMCID: PMC6930893. [CrossRef]
- Sherameti, I.; Shahollari, B.; Venus, Y.; Altschmied, L.; Varma, A.; Oelmüller, R. The endophytic fungus Piriformospora indica stimulates the expression of nitrate reductase and the starch-degrading enzyme glucan-water dikinase in tobacco and Arabidopsis roots through a homeodomain transcription factor that binds to a conserved motif in their promoters. J. Biol. Chem. 2005, 280(28), 26241-7. Epub 2005 Feb 14. PMID: 15710607. [CrossRef]
- Eliaspour, S.; Seyed Sharifi, R.; Shirkhani, A. Evaluation of interaction between Piriformospora indica, animal manure and NPK fertilizer on quantitative and qualitative yield and absorption of elements in sunflower. Food Sci. Nutr. 2020, 8(6), 2789-2797. PMID: 32566196; PMCID: PMC7300063. [CrossRef]
- Strehmel, N.; Mönchgesang, S.; Herklotz, S.; Krüger, S.; Ziegler, J.; Scheel, D. Piriformospora indica Stimulates Root Metabolism of Arabidopsis thaliana. Int. J. Mol. Sci. 2016, 17(7),1091. PMID: 27399695; PMCID: PMC4964467. [CrossRef]
- Ghaffari, M.R.; Ghabooli, M.; Khatabi, B.; Hajirezaei, M.R.; Schweizer, P.; Salekdeh, G.H. Metabolic and transcriptional response of central metabolism affected by root endophytic fungus Piriformospora indica under salinity in barley. Plant Mol. Biol. 2016, 90(6), 699-717. Epub 2016 Mar 7. PMID: 26951140. [CrossRef]
- Lahrmann, U.; Ding, Y.; Banhara, A.; Rath, M.; Hajirezaei, M.R.; Döhlemann, S.; von Wirén, N.; Parniske, M.; Zuccaro, A. Host-related metabolic cues affect colonization strategies of a root endophyte. Proc. Natl. Acad. Sci. U S A. 2013, 110(34), 13965-70. Epub 2013 Aug 5. PMID: 23918389; PMCID: PMC3752250. [CrossRef]
- Hua, M.D.; Senthil Kumar, R.; Shyur, L.F.; Cheng, Y.B.; Tian, Z.; Oelmüller, R.; Yeh, K.W. Metabolomic compounds identified in Piriformospora indica-colonized Chinese cabbage roots delineate symbiotic functions of the interaction. Sci. Rep. 2017, 7(1), 9291. PMID: 28839213; PMCID: PMC5571224. [CrossRef]
- Bandyopadhyay, P.; Yadav, B.G.; Kumar, S.G., Kumar, R.; Kogel, K.H.; Kumar, S. Piriformospora indica and Azotobacter chroococcum Consortium Facilitates Higher Acquisition of N, P with Improved Carbon Allocation and Enhanced Plant Growth in Oryza sativa. J. Fungi (Basel). 2022, 8(5), 453. PMID: 35628709; PMCID: PMC9146537. [CrossRef]
- Mansotra, P.; Sharma, P.; Sharma, S. Bioaugmentation of Mesorhizobium cicer, Pseudomonas spp. and Piriformospora indica for Sustainable Chickpea Production. Physiol. Mol. Biol. Plants 2015, 21(3), 385-93. Epub 2015 Apr 16. PMID: 26261403; PMCID: PMC4524863. [CrossRef]
- Hallasgo, A.M.; Spangl, B.; Steinkellner, S.; Hage-Ahmed, K. The Fungal Endophyte Serendipita williamsii Does Not Affect Phosphorus Status but Carbon and Nitrogen Dynamics in Arbuscular Mycorrhizal Tomato Plants. J. Fungi (Basel). 2020, 6(4), 233. PMID: 33086650; PMCID: PMC7711999. [CrossRef]
- Pérez-Alonso, M.M.; Guerrero-Galán, C.; Scholz, S.S.; Kiba, T.; Sakakibara, H.; Ludwig-Müller, J.; Krapp, A.; Oelmüller, R.; Vicente-Carbajosa, J.; Pollmann, S. Harnessing symbiotic plant-fungus interactions to unleash hidden forces from extreme plant ecosystems. J. Exp. Bot. 2020, 71(13), 3865-3877. PMID: 31976537; PMCID: PMC7316966. [CrossRef]
- Krapp, A.; Berthomé, R.; Orsel, M.; Mercey-Boutet, S.; Yu, A.; Castaings, L.; Elftieh, S.; Major, H.; Renou, J.P.; Daniel-Vedele, F. Arabidopsis roots and shoots show distinct temporal adaptation patterns toward nitrogen starvation. Plant Physiol. 2011, 157(3),1255-1282. Epub 2011 Sep 7. PMID: 21900481; PMCID: PMC3252138. [CrossRef]
- Hodge, A.; Stewart, J.; Robinson, D.; Griffiths, B.S.; Fitter, A.H. Competition between roots and soil microorganisms for nutrientsfrom nitrogen-rich patches of varying complexity. J. Ecology 2000, 88: 150–164.
- Hodge, A.; Campbell, C.D.; Fitter AH. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature 2001, 413, 297–299.
- Thirkell, T.J.; Cameron, D.D.; Hodge, A. Resolving the ‘nitrogen paradox’ of arbuscular mycorrhizas: fertilisation with organic matter brings considerable benefits for plant nutrition and growth. Plant, Cell Environ. 2016, 39, 1683–1690.
- Hoysted, G.A.; Field, K.J.; Sinanaj, B.; Bell, C.A.; Bidartondo, M.I.; Pressel, S. Direct nitrogen, phosphorus and carbon exchanges between Mucoromycotina 'fine root endophyte' fungi and a flowering plant in novel monoxenic cultures. New Phytol. 2023, 238(1), 70-79. Epub 2023 Feb 5. PMID: 36739554. [CrossRef]
- Breuillin-Sessoms, F.; Floss, D.S.; Karen Gomez, S.; et al. Suppression of arbuscule degeneration in Medicago truncatula phosphate transporter4 mutants is dependent on the ammonium transporter 2 family protein AMT2;3. Plant Cell 2015, 27, 1352–1366.
- Paul, E.A.; Kucey, R.M.N. Carbon flow in plant microbial associations incriminated in perinatal morbidity and mortality. Science 1981, 213, 473–474.
- Wright, D.P.; Read, D.J.; Scholes, J.D. Mycorrhizal sink strength influences whole plant carbon balance of Trifolium repens L. Plant, Cell Environ. 1998, 21, 881–891.
- Lendenmann, M.; Thonar, C.; Barnard, R.L.; et al. Symbiont identity matters: carbon and phosphorus fluxes between Medicago truncatula and different arbuscular mycorrhizal fungi. Mycorrhiza 2011, 21, 689–702.
- Cope, K.R.; Kafle, A.; Yakha, J.K.; et al. Physiological and transcriptomic response of Medicago truncatula to colonization by high- or low-benefit arbuscular mycorrhizal fungi. Mycorrhiza 2022, 32, 281–303. https://doi.org/10.1007/s00572-022-01077-2.
- Belmondo, S.; Fiorilli, V.; Pérez-Tienda, J.; Ferrol, N. Marmeisse, R., Lanfranco, L. A dipeptide transporter from the arbuscular mycorrhizal fungus Rhizophagus irregularis is upregulated in the intraradical phase. Front. Plant Science 2014, 5, 436.
- Liu, Y.; Li, T.; Zhang, C.; Zhang, W.; Deng, N.; Dirk, L.M.A.; Downie, A.B.; Zhao, T. Raffinose positively regulates maize drought tolerance by reducing leaf transpiration. Plant J. 2023, 114(1), 55-67. Epub 2023 Feb 7. PMID: 36703577. [CrossRef]
- ElSayed, A.I.; Rafudeen, M.S.; Golldack, D. Physiological aspects of raffinose family oligosaccharides in plants: protection against abiotic stress. Plant Biol. (Stuttg). 2014,16(1), 1-8. Epub 2013 Aug 12. PMID: 23937337. [CrossRef]
- Gu, L.; Jiang, T.; Zhang, C.; Li, X.; Wang, C.; Zhang, Y.; Li, T.; Dirk, L.M.A.; Downie, A.B.; Zhao, T. Maize HSFA2 and HSBP2 antagonistically modulate raffinose biosynthesis and heat tolerance in Arabidopsis. Plant J. 2019, 100(1), 128-142. Epub 2019 Jul 12. PMID: 31180156. [CrossRef]
- Han, Q.; Qi, J.; Hao, G.; Zhang, C.; Wang, C.; Dirk, L.M.A.; Downie, A.B.; Zhao, T. ZmDREB1A Regulates RAFFINOSE SYNTHASE Controlling Raffinose Accumulation and Plant Chilling Stress Tolerance in Maize. Plant Cell Physiol. 2020, 61(2), 331-341. PMID: 31638155. [CrossRef]
- Li, C.H.; Tien, H.J.; Wen, M.F.; Yen, H.E. Myo-inositol transport and metabolism participate in salt tolerance of halophyte ice plant seedlings. Physiol. Plant. 2021, 172(3), 1619-1629. Epub 2021 Mar 5. PMID: 33511710. [CrossRef]
- Yang, J.; Ling, C.; Liu, Y.; Zhang, H.; Hussain, Q.; Lyu, S.; Wang, S.; Liu, Y. Genome-Wide Expression Profiling Analysis of Kiwifruit GolS and RFS Genes and Identification of AcRFS4 Function in Raffinose Accumulation. Int. J. Mol. Sci. 2022, 23(16), 8836. PMID: 36012101; PMCID: PMC9408211. [CrossRef]
- Wang, S.; Chen, A.; Xie, K.; Yang, X.; Luo, Z.; Chen, J.; Zeng, D.; Ren, Y.; Yang, C.; Wang, L.; Feng, H.; López-Arredondo, D.L.; Herrera-Estrella, L.R.; Xu, G. Functional analysis of the OsNPF4.5 nitrate transporter reveals a conserved mycorrhizal pathway of nitrogen acquisition in plants. Proc Natl Acad Sci U S A. 2020, 117(28), 16649-16659. Epub 2020 Jun 25. PMID: 32586957; PMCID: PMC7368293. [CrossRef]
- Kiba, T., Feria-Bourrellier, A.B.; Lafouge, F.; Lezhneva, L.; Boutet-Mercey, S.; Orsel, M.; Bréhaut, V.; Miller, A.; Daniel-Vedele, F.; Sakakibara, H.; Krapp, A. The Arabidopsis nitrate transporter NRT2.4 plays a double role in roots and shoots of nitrogen-starved plants. Plant Cell 2012, 24(1), 245-58. Epub 2012 Jan 6. PMID: 22227893; PMCID: PMC3289576. [CrossRef]
- Chen, H.Y.; Chen, Y.N.; Wang, H.Y.; Liu, Z.T.; Frommer, W.B.; Ho, C.H. Feedback inhibition of AMT1 NH4+-transporters mediated by CIPK15 kinase. BMC Biol. 2020 18(1), 196. PMID: 33317525; PMCID: PMC7737296. [CrossRef]
- Li, J.Y.; Fu, Y.L.; Pike, S.M.; Bao, J.; Tian, W.; Zhang, Y.; Chen, C.Z.; Zhang, Y.; Li, H.M., Huang, J.; Li, L.G.; Schroeder, J.I.; Gassmann, W.; Gong, J.M. The Arabidopsis nitrate transporter NRT1.8 functions in nitrate removal from the xylem sap and mediates cadmium tolerance. Plant Cell 2010, 22(5),1633-46. Epub 2010 May 25. PMID: 20501909; PMCID: PMC2899866. [CrossRef]
- Dechorgnat, J.; Patrit, O.; Krapp, A.; Fagard, M.; Daniel-Vedele, F. Characterization of the Nrt2.6 gene in Arabidopsis thaliana: a link with plant response to biotic and abiotic stress. PLoS One 2012;7(8):e42491. Epub 2012 Aug 7. PMID: 22880003; PMCID: PMC3413667. [CrossRef]
- Wang, Y.Y.; Tsay, Y.F. Arabidopsis nitrate transporter NRT1.9 is important in phloem nitrate transport. Plant Cell 2011, 23(5), 1945-57. Epub 2011 May 13. PMID: 21571952; PMCID: PMC3123939. [CrossRef]
- Lu, Y.T.; Liu, D.F.; Wen, T.T.; Fang, Z.J.; Chen, S.Y.; Li, H.; Gong, J.M. Vacuolar nitrate efflux requires multiple functional redundant nitrate transporter in Arabidopsis thaliana. Front Plant Sci. 2022, 13, 926809. PMID: 35937356; PMCID: PMC9355642. [CrossRef]
- Wu, X.; Liu, T.; Zhang, Y.; Duan, F.; Neuhäuser, B.; Ludewig, U.; Schulze, W.X.: Yuan, L. Ammonium and nitrate regulate NH4+ uptake activity of Arabidopsis ammonium transporter AtAMT1;3 via phosphorylation at multiple C-terminal sites. J. Exp. Bot. 2019, 70(18), 4919-4930. PMID: 31087098; PMCID: PMC6760267. [CrossRef]
- Yuan, L.; Loqué, D.; Kojima, S.; Rauch, S.; Ishiyama, K.; Inoue, E.; Takahashi, H.; von Wirén, N. The organization of high-affinity ammonium uptake in Arabidopsis roots depends on the spatial arrangement and biochemical properties of AMT1-type transporters. Plant Cell 2007, 19(8), 2636-52. Epub 2007 Aug 10. PMID: 17693533; PMCID: PMC2002620. [CrossRef]
- Yuan, L.; Graff, L.; Loqué, D.; Kojima, S.; Tsuchiya, Y.N.; Takahashi, H.; von Wirén, N. AtAMT1;4, a pollen-specific high-affinity ammonium transporter of the plasma membrane in Arabidopsis. Plant Cell Physiol. 2009, 50(1),13-25. Epub 2008 Dec 10. PMID: 19073648; PMCID: PMC2638712. [CrossRef]
- Zanin, L.; Tomasi, N.; Wirdnam, C.; Meier, S.; Komarova, N.Y.; Mimmo, T.; Cesco, S.; Rentsch, D.; Pinton. R. Isolation and functional characterization of a high affinity urea transporter from roots of Zea mays. BMC Plant Biol. 2014, 14, 222. PMID: 25168432; PMCID: PMC416055. [CrossRef]
- Wang, W.H.; Köhler, B.; Cao, F.Q.; Liu, G.W.; Gong, Y.Y.; Sheng, S.; Song, Q.C.; Cheng, X.Y.; Garnett, T.; Okamoto, M.; Qin, R.; Mueller-Roeber, B.; Tester, M.; Liu, L.H. Rice DUR3 mediates high-affinity urea transport and plays an effective role in improvement of urea acquisition and utilization when expressed in Arabidopsis. New Phytol. 2012, 193(2), 432-44. Epub 2011 Oct 19. PMID: 22010949. [CrossRef]
- Yu, S.; Pratelli, R.; Denbow, C.; Pilot, G. Suppressor mutations in the Glutamine Dumper1 protein dissociate disturbance in amino acid transport from other characteristics of the Gdu1D phenotype. Front Plant Sci. 2015, 6, 593. PMID: 26300894; PMCID: PMC4523740. [CrossRef]
- Hirner, A.; Ladwig, F.; Stransky, H.; Okumoto, S.; Keinath, M.; Harms, A.; Frommer, W.B.; Koch, W. Arabidopsis LHT1 is a high-affinity transporter for cellular amino acid uptake in both root epidermis and leaf mesophyll. Plant Cell 2006, 18(8),1931-46. Epub 2006 Jun 30. PMID: 16816136; PMCID: PMC1533986. [CrossRef]
- Havé, M.; Marmagne, A.; Chardon, F.; Masclaux-Daubresse, C. Nitrogen remobilization during leaf senescence: lessons from Arabidopsis to crops. J. Exp. Bot. 2017, 68(10), 2513-2529. PMID: 27707774. [CrossRef]
- Choi, J.; Eom, S.; Shin, K.; Lee, R.A.; Choi, S.; Lee, J.H.; Lee, S.; Soh, M.S. Identification of Lysine Histidine Transporter 2 as an 1-Aminocyclopropane Carboxylic Acid Transporter in Arabidopsis thaliana by Transgenic Complementation Approach. Front. Plant Sci. 2019, 10:1092. PMID: 31572413; PMCID: PMC6749071. [CrossRef]
- Fujiwara, T.; Hirai, M.Y.; Chino, M.; Komeda, Y.; Naito, S. Effects of Sulfur Nutrition on Expression of the Soybean Seed Storage Protein Genes in Transgenic Petunia. Plant Physiol. 1992, 99 (1), 263-268.
- Johnson, J.M.; Sherameti, I.; Ludwig, A.; Nongbri, P.L.; Sun, C.; Varma, A.; Oelmüller, R. Protocols for Arabidopsis thaliana and Piriformospora indica co-cultivation: a model system to study plant beneficial traits. Endocyt. Cell Res. 2011, 21, 101–113.
- Johnson, J.M.; Sherameti, I.; Nongbri, P.L.; Oelmüller, R. Standardized conditions to study beneficial and nonbeneficial traits in the Piriformospora indica/Arabidopsis thaliana interaction. In A Varma, G Kost, R Oelmüller, eds, Piriformospora indica: Sebacinales and Their Biotechnological Applications. Soil Biology, 2013, 33, 325–343.
- Pérez-Alonso, M.-M.; Guerrero-Galán, C.; González Ortega-Villaizán, A.; Ortiz-García, P.; Scholz, S.S.; Ramos, P.; et al. The calcium sensor CBL7 is required for Serendipita indica-induced growth stimulation in Arabidopsis thaliana, controlling defense against the endophyte and K+ homoeostasis in the symbiosis. Plant, Cell Environ. 2022, 45, 3367–3382. [CrossRef]
- Chen, A.; Gu, M.; Wang, S.; Chen, J.; Xu, G. Transport properties and regulatory roles of nitrogen in arbuscular mycorrhizal symbiosis. Seminars in Cell and Developmental Biology 2018, 74, 80–88.
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nature Biotechnol. 2019, 37(8), 907–915. [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30(7), 923-30.
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq. 2. Genome Biology 2014, 15(12), 550. [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. Royal Statistical Soc.:Series B. 1995, 57(1), 289–300. [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45.
- Bütehorn, B.; Rhody, D.; Franken, P. Isolation and characterisation of Pitef1 encoding the translation elongation factor EF-1α of the root endophyte Piriformospora indica. Plant Biol. 2000, 2(6), 687–692. [CrossRef]
- Scholz, S. S.; Vadassery, J.; Heyer, M.; Reichelt, M.; Bender, K. W.; Snedden, W. A.; Boland, W.; Mithöfer, A., Mutation of the Arabidopsis Calmodulin-Like Protein CML37 Deregulates the Jasmonate Pathway and Enhances Susceptibility to Herbivory. Mol Plant, 2014, 7(12), 1712-1726. [CrossRef]
- Forzani, C.; Duarte, G.T.; Van Leene, J.; Clément, G.; Huguet, S.; Paysant-Le-Roux, C.; Mercier, R.; De Jaeger, G.; Leprince, A.-S.; Meyer, C.. Mutations of the AtYAK1 Kinase Suppress TOR Deficiency in Arabidopsis. Cell Rep. 2019, 27, 3696-3708.e5.
Figure 1.
Shoot (A) and root (B) weights of Arabidopsis seedlings which were either grown on full medium (+N, white) or N-limited medium (-N, grey), in the absence or presence of P. indica for 5 days. % Growth promotion by the fungus was determined for 20 shoots and roots, whereas the plant material from full medium without the fungus was set as 100%. Based on 3 independent experiments, error bars are SEs. Statistic significant differences were analyzed by One-way ANOVA (Holm-Sidak test). Different small letters indicate statistic significant differences.
Figure 1.
Shoot (A) and root (B) weights of Arabidopsis seedlings which were either grown on full medium (+N, white) or N-limited medium (-N, grey), in the absence or presence of P. indica for 5 days. % Growth promotion by the fungus was determined for 20 shoots and roots, whereas the plant material from full medium without the fungus was set as 100%. Based on 3 independent experiments, error bars are SEs. Statistic significant differences were analyzed by One-way ANOVA (Holm-Sidak test). Different small letters indicate statistic significant differences.
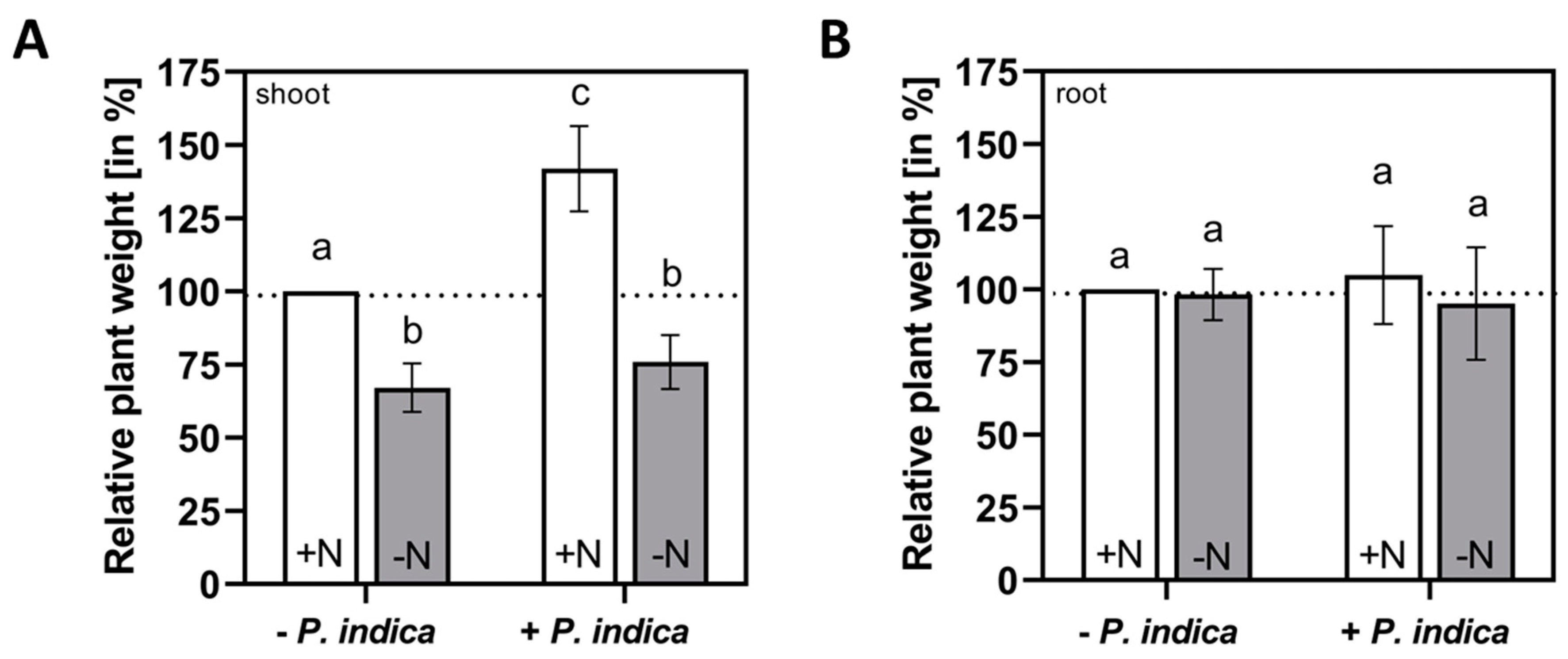
Figure 2.
Total N in the shoots of uncolonized (white) and colonized (grey) Arabidopsis seedlings. (A) Total N in the shoots of seedlings grown with or without P. indica on either full or N-limiting conditions for 5 days. % N was determined in dried material of 20 shoots. (B) 15N accumulation in the shoots of seedlings grown with or without P. indica on either full or N-limiting conditions. 15N was determined in dried material of 20 shoots. Based on 3 independent experiments, error bars are SEs. Statistic significant differences were analyzed by One-way ANOVA (Holm-Sidak test). Different small letters indicate statistic significant differences.
Figure 2.
Total N in the shoots of uncolonized (white) and colonized (grey) Arabidopsis seedlings. (A) Total N in the shoots of seedlings grown with or without P. indica on either full or N-limiting conditions for 5 days. % N was determined in dried material of 20 shoots. (B) 15N accumulation in the shoots of seedlings grown with or without P. indica on either full or N-limiting conditions. 15N was determined in dried material of 20 shoots. Based on 3 independent experiments, error bars are SEs. Statistic significant differences were analyzed by One-way ANOVA (Holm-Sidak test). Different small letters indicate statistic significant differences.
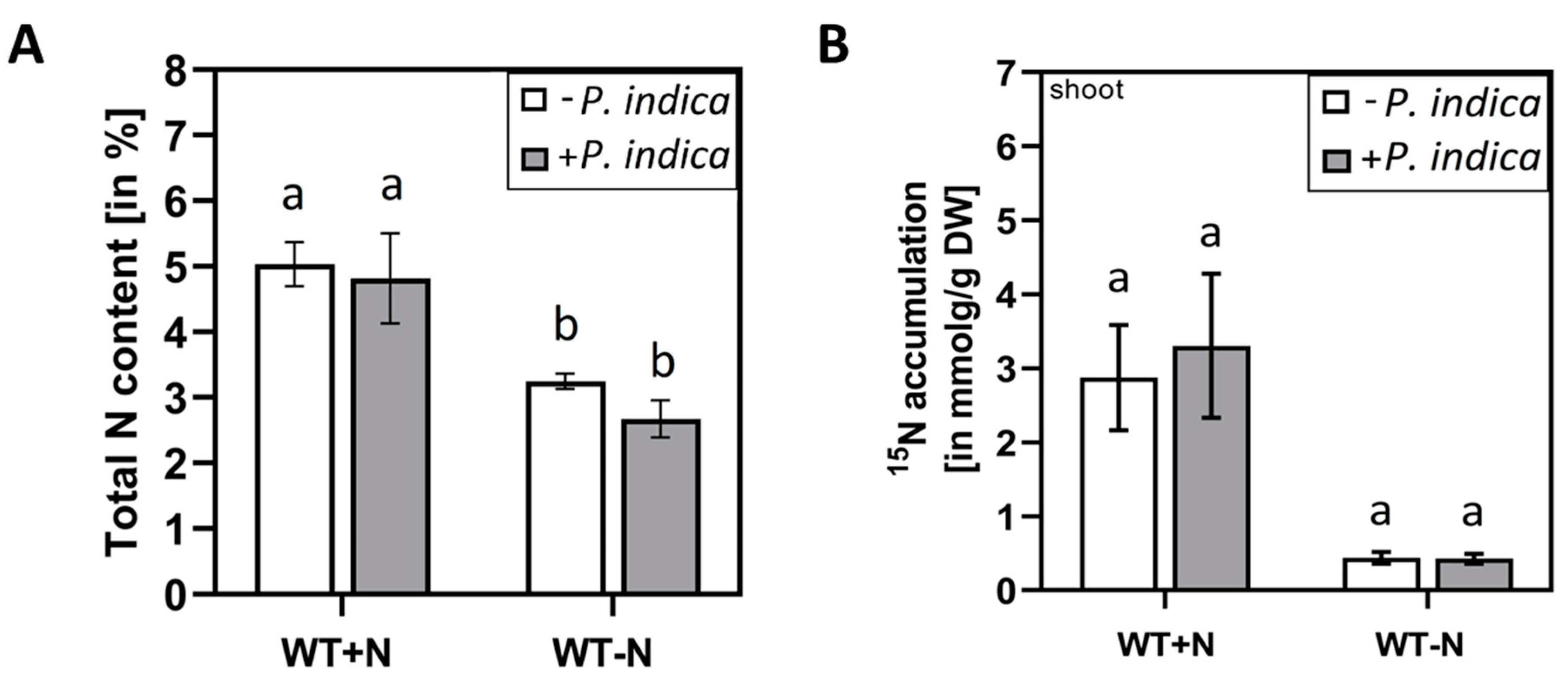
Figure 3.
(A) Experimental set-up of sterile Arabidopsis seedlings co-cultivated with 15N-labelled P. indica. To obtain labelled fungal material, fungal plugs grown on KM plate were transferred to a modified liquid N-free KM medium supplemented with 10g/L ISOGRO®-15N and incubated for 14 days in a well plate. The fungal material was separated from the medium, washed carefully with PNM-N and cut into 5x5 mm pieces. The fungus was placed onto the nylon membranes with 1 cm distance to the roots. The co-cultures were incubated for 14 days. For details, cf. Material and Methods. (B) 15N label in the shoots of seedlings, which were exposed to the 15N-labelled hyphae on either full (white) or N-limited (grey) media. The accumulation of 15N was determined in dried leaf material of 20 colonized shoots, 14 days after the beginning of the co-culture. For experimental details, cf. Methods and Material. Based on 3 independent experiments, bars represent SEs. Ns, not significant, which was analyzed by t-test on ranks (Mann-Whitney test).
Figure 3.
(A) Experimental set-up of sterile Arabidopsis seedlings co-cultivated with 15N-labelled P. indica. To obtain labelled fungal material, fungal plugs grown on KM plate were transferred to a modified liquid N-free KM medium supplemented with 10g/L ISOGRO®-15N and incubated for 14 days in a well plate. The fungal material was separated from the medium, washed carefully with PNM-N and cut into 5x5 mm pieces. The fungus was placed onto the nylon membranes with 1 cm distance to the roots. The co-cultures were incubated for 14 days. For details, cf. Material and Methods. (B) 15N label in the shoots of seedlings, which were exposed to the 15N-labelled hyphae on either full (white) or N-limited (grey) media. The accumulation of 15N was determined in dried leaf material of 20 colonized shoots, 14 days after the beginning of the co-culture. For experimental details, cf. Methods and Material. Based on 3 independent experiments, bars represent SEs. Ns, not significant, which was analyzed by t-test on ranks (Mann-Whitney test).
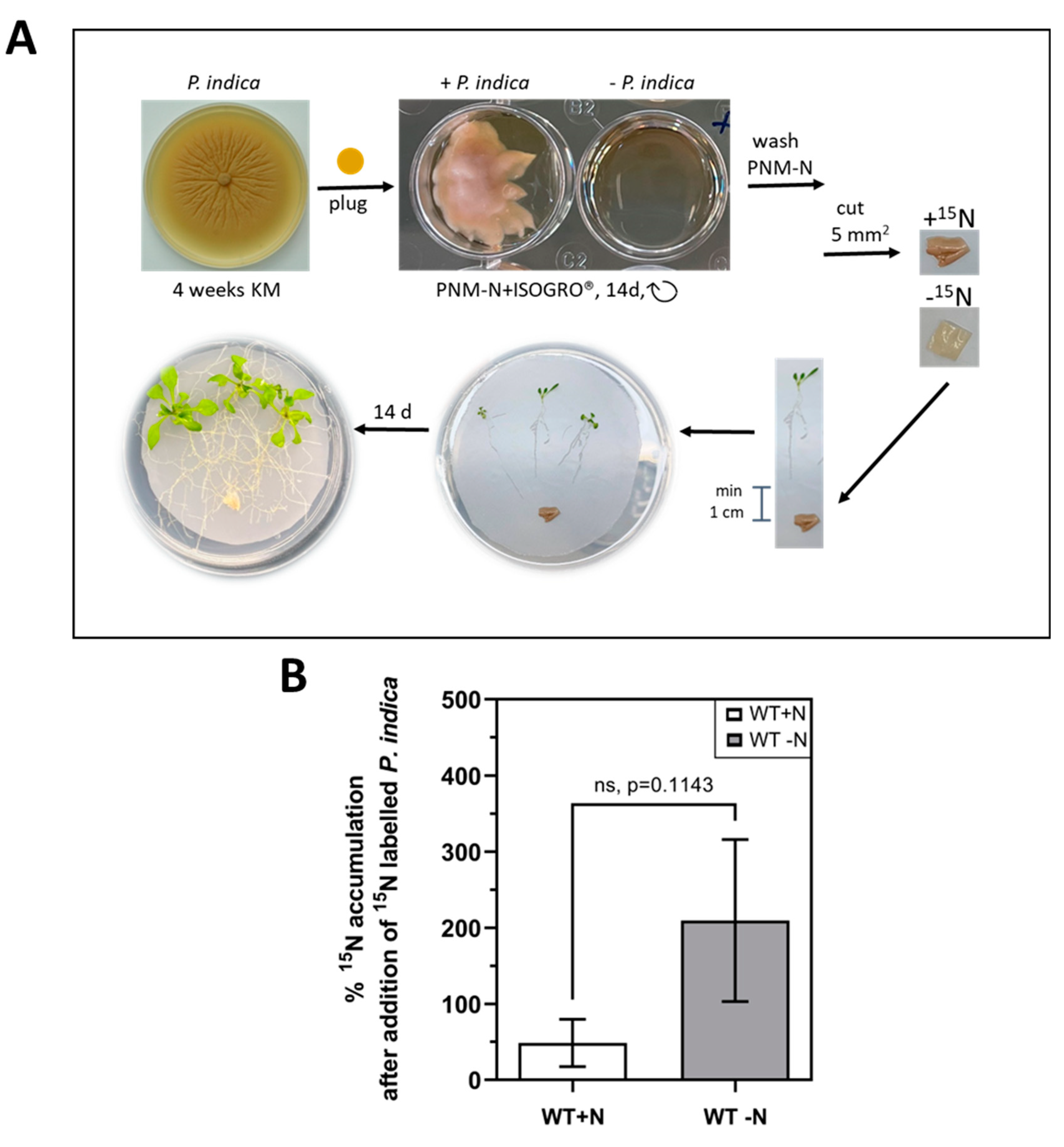
Figure 4.
Selected differentially accumulated metabolites (DAMs) in Arabidopsis shoots. Values are gives as the ratio of the (relative) content in N-limiting to N-sufficient growth condition as measured by GC-MS profiling. Data from 3 independent cultures on pools of 20 plantlets. The mean value is indicated. .
Figure 4.
Selected differentially accumulated metabolites (DAMs) in Arabidopsis shoots. Values are gives as the ratio of the (relative) content in N-limiting to N-sufficient growth condition as measured by GC-MS profiling. Data from 3 independent cultures on pools of 20 plantlets. The mean value is indicated. .
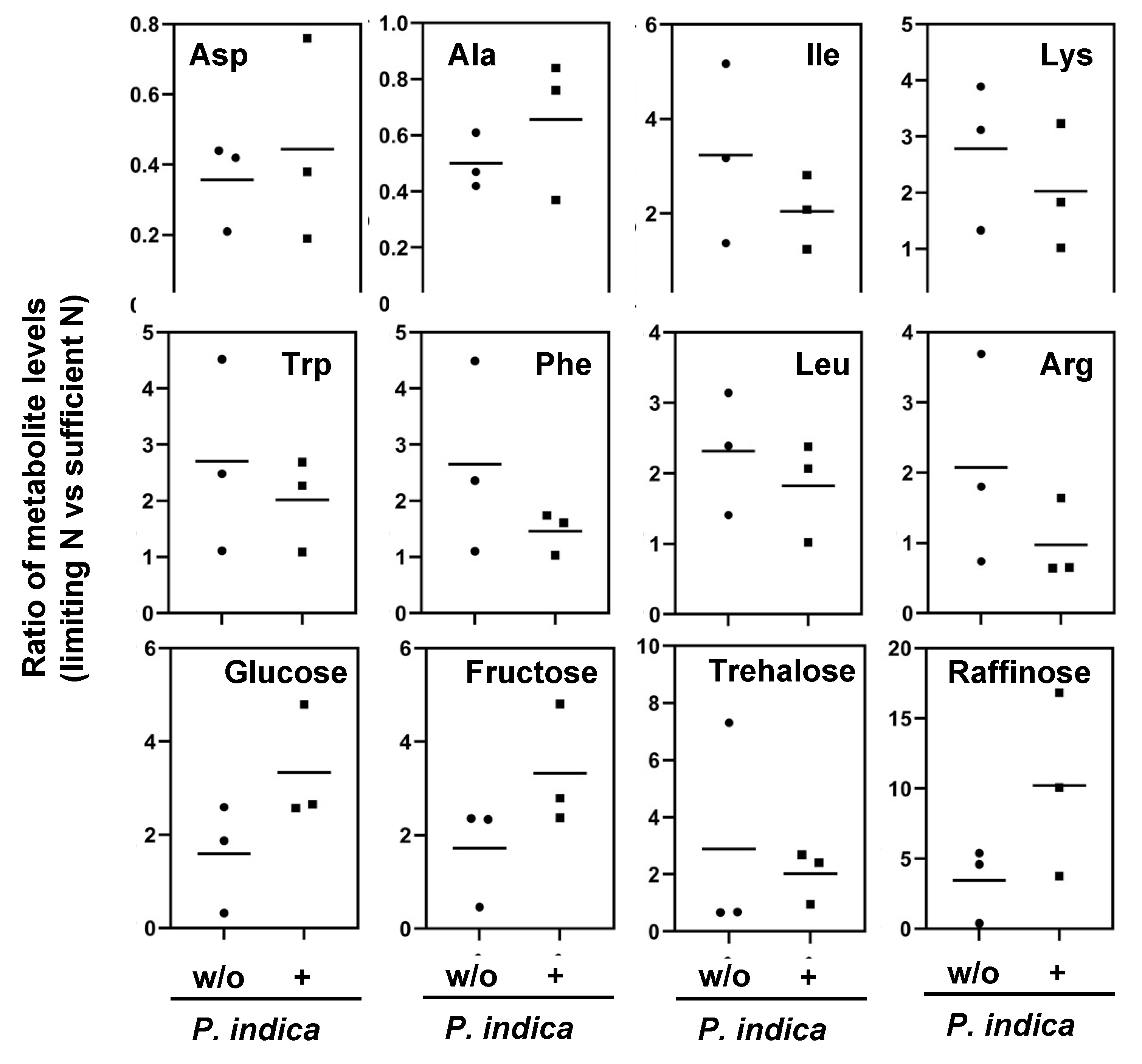

Table 1.
Differentially accumulated metabolites (DAMs) in Arabidopsis shoots. DAMs regulated by N limitation in Arabidopsis thaliana rosettes without or with P. indica colonization. Values are gives as the ratio of the (relative) content in N-limiting to N-sufficient growth condition as measured by GC-MS profiling. Data are means +/- SE of 3 replicates from independent cultures on pools of 20 plantlets. A) amino acids; B) soluble carbohydrates. The gradual color scale is indicated.
Table 1.
Differentially accumulated metabolites (DAMs) in Arabidopsis shoots. DAMs regulated by N limitation in Arabidopsis thaliana rosettes without or with P. indica colonization. Values are gives as the ratio of the (relative) content in N-limiting to N-sufficient growth condition as measured by GC-MS profiling. Data are means +/- SE of 3 replicates from independent cultures on pools of 20 plantlets. A) amino acids; B) soluble carbohydrates. The gradual color scale is indicated.
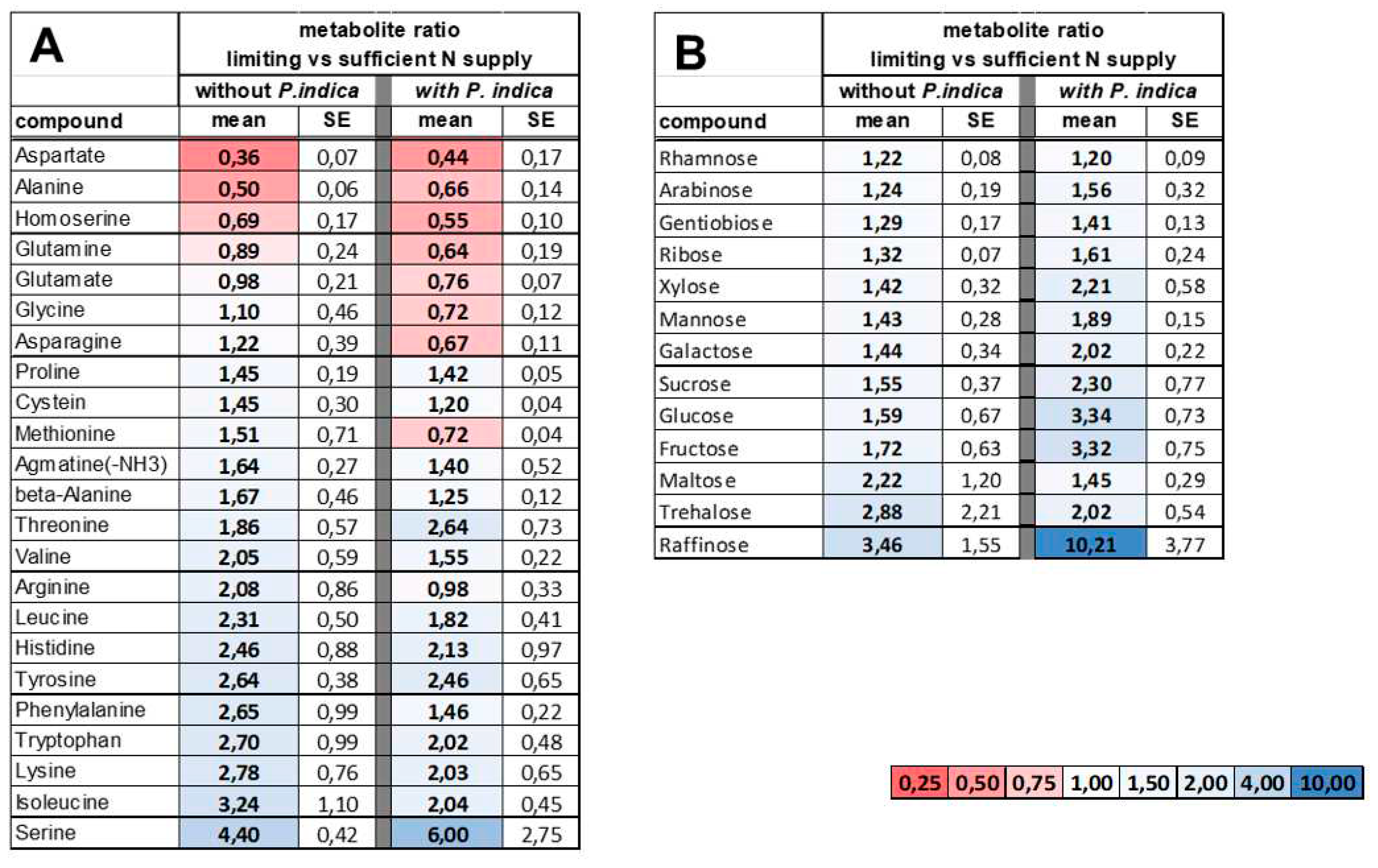 |
Table 2.
Differentially regulated transporters in Arabidopsis. DEGs regulated by nitrogen limitation (-N) in Arabidopsis thaliana tissues without (w/o) or with (w) P. indica colonization. Values are given as log2-fold differential expression identified by RNAseq analysis, p<0.05. x = not differentially expressed compared to full N (+N).
Table 2.
Differentially regulated transporters in Arabidopsis. DEGs regulated by nitrogen limitation (-N) in Arabidopsis thaliana tissues without (w/o) or with (w) P. indica colonization. Values are given as log2-fold differential expression identified by RNAseq analysis, p<0.05. x = not differentially expressed compared to full N (+N).
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



