
Submitted:
13 September 2023
Posted:
14 September 2023
You are already at the latest version
Abstract
The objective of the study was to analyse the antimicrobial susceptibility trends of Spanish porcine bacteria to quinolones, cephalosporins and polymyxins. Isolates of Actinobacillus pleuropneumoniae, Pasteurella multocida, and Escherichia coli were isolated from sick pigs from 2019 to 2022. Antimicrobial susceptibility test was determined by minimal inhibitory concentration (MIC) following internationally accepted methodology. MIC categorization was based on distributing the range of MIC values in four categories being category one the most susceptible (lowest MIC value) and four the less susceptible one (highest MIC value). Moreover, clinical susceptibility (susceptible/resistant) was also determined according to CLSI and EUCAST clinical breakpoints. A logistic and multinomial logistic regression model was used to analyze the susceptibility data for dichotomized and categorized MIC data, respectively for any pair of antimicrobial/microorganism. In general terms, the antimicrobial susceptibility of pig bacteria to these antimicrobials remained stable or increased in the last four years in Spain. In the case of A. pleuropneumoniae and quinolones, it was observed a significant temporal trend where isolates from 2020 had significantly increased odds of being more susceptible than isolates from 2019. In the case of E. coli and polymyxins, a significant temporal trend was observed where isolates from 2020 and 2021 had significantly increased odds of being more susceptible than isolates from 2019 and 2020, respectively. Finally, a significant odd of being less susceptible was only observed for cephalosporins and E. coli for 2020 versus 2019, stagnating for the rest of study period. These results provide sound data on critical important antimicrobials in swine medicine.
Keywords:
trend analysis
; antimicrobial susceptibility
; porcine pathogens
1. Introduction
Antimicrobial resistance (AMR) threatens the successful treatment of bacterial infections not only in human but also in animal health [1,2]. The use of antimicrobials (AB) in humans and animals is a driver in the increase of AMR in bacterial populations, even following guidelines for prudent use of AB [3,4]. This risk significantly increases with the misuse of these drugs [5]. Moreover, the AMR reservoir of bacteria from livestock has been increasingly investigated for its potential to transfer AMR to humans via direct contact, the environment and contaminated food [6]. Nevertheless, the extent of this transmission remains uncertain due to the enormous complexity of the AMR epidemiology involving animals, environment, and humans [7,8,9]. Nevertheless, policy makers, in the European Union (EU), have developed legislation to monitor and regulate the antimicrobial use in animals with the goal to decrease AMR burden in humans in the long run [10,11]. However, the global effect of these actions, regarding the reduction of AMR at the human-animal-environment interface, is still under investigation, and very few scientific studies have shown encouraging results, limited to some antimicrobials such as colistin [12,13] probable due that AMU is one key driver for AMR but other socio-economic factors should be also taken into account in AMR epidemiology as recently assessed [14]. On the other hand, this long-term reduction of AB consumption in veterinary medicine could seriously hamper the care of animals and generate severe welfare issues if animals are not treated with the right antimicrobial when it is really needed.
The current EU legislation regarding antimicrobials [10] have focused special attention to restrict as much as possible the use of last resource antimicrobials (3rd and 4th generation cephalosporins, polymyxins and quinolones) in animals following the recommendations addressed by the European Medicine Agency in 2019 [15]. Thus, these last resource AB can only be used when no other options belonging to less risky categories (C and D) for AMR are available to treat animals [15]. However, up to date, most of the long-term surveillance data available are only from healthy animals that may not reflect the situation in veterinary bacterial pathogens [16]. Thus, the European Food Safety Authority (EFSA) coordinates a mandatory active monitoring of AMR in zoonotic (Salmonella spp and Campylobacter spp), indicator bacteria (Escherichia coli) and extended-spectrum-cephalosporin-resistant and carbapenemase-producing E. coli from healthy food-producing animals (cattle, poultry, pigs) at slaughter and meat following European directives [17,18]. On the other hand, a coordinated and harmonized strategy for AMR monitoring in diseased animals has just started at European level [19] to fill the gap for AMR data in pathogens from diseased animals. Thus, updated information will be generated to guide antimicrobial stewardship initiatives such as treatment guidelines, and to guide policymakers in regulating veterinary antibiotic use [20].
The use of antimicrobials with therapeutic or metaphylactic purpose in pigs may be necessary to control the relevant pathogens involved in respiratory and enteric disorders, contributing to most of the pig antimicrobial consumption [21,22,23]. Thus, porcine respiratory disease complex (PRDC) and post-weaning diarrhoea (PWD) are some of the most challenging diseases affecting the pig industry worldwide [24,25]. PRDC is a syndrome that results from a combination of infectious (bacteria and viruses) and non-infectious factors. Actinobacillus pleuropneumoniae (APP), Pasteurella multocida, Mycoplasma hyopneumoniae, and Bordetella bronchiseptica are the most common bacterial agents involved [26]. On the other hand, Escherichia coli is the main causative agent of PWD, affecting piglets after weaning. PWD is characterized by a profuse diarrhea, dehydration, significant mortality, and loss of body weight in surviving pigs [27,28,29]. When clinical signs appear, prescription of antimicrobials is in many cases the only solution to control the spread of the PRDC and PWD within the herd [21,22,29,30,31]. Thus, it may be necessary to use last resource antimicrobials if no other option is available according to an antimicrobial stewardship program [3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32]. It must be highlighted that during the last four years, the sales of last resource antimicrobials in European´s livestock are between 0.2 and 2.8% of the total sales of antimicrobials [33], suggesting that the bacterial populations are hardly exposed to these family of drugs across Europe.
An important aspect of dealing with the AMR crisis is surveillance [34], which provides susceptibility data allowing to act more effectively when necessary. Another goal of AMR surveillance is to analyze the temporal trends of AMR patterns for early warning of potential threats and decipher the impact of policies in animals regarding the use of antimicrobials in the long term. Unfortunately, there is scarce knowledge on the antimicrobial susceptibility profiles of veterinary bacterial pathogens in Europe due to a lack of coordinated strategy between member states [35]. The objective of this study was to describe and analyze the temporal trends during the last four years of last resource antimicrobials in Spanish porcine pathogens as a suitable model for other countries, considering the low consumption of these drugs in Spain compared with the total antimicrobial consumption (3-4,1%) and the consistent decrease in the total antimicrobial use in livestock [33].
2. Results
2.1. Bacteria isolation
From January 2019 to 2022, 1,827 samples were received from isowean, wean-to-finish and fattening farms suffering from clinical respiratory disease associated with PRDC. Additionally, 3,813 samples were received from sow, isowean and wean-to-finishing farms suffering clinical sings compatible with PWD. Only one isolate was included by farm across the study to avoid redundancy and overrepresentation of bacterial clones. In the case of sow farms, the samples were obtained from their nursery facility. Bacterial isolation for respiratory pathogens (A. pleuropneumoniae, P. multocida and B. bronchiseptica) was successful in 80% (1,461/1,827) of the cases, furthermore in 20% of the samples, more than one bacterial species were isolated. Bacterial isolation of E. coli was successful in 79.3% (3,024/3,813) of the samples associated to enteric disorders. Finally, in 5% of the enteric samples, it was possible to isolate more than one bacterial species, generally Salmonella spp. The number of isolates collected for each bacterium during the study period is detailed in Table 1. Thus, for A. pleuropneumoniae , E. coli and P. multocida there were at least 100 isolates isolated each year and, therefore they were included in the statistical analysis.
2.2. Distribution of MIC by antimicrobial and microorganism across the years
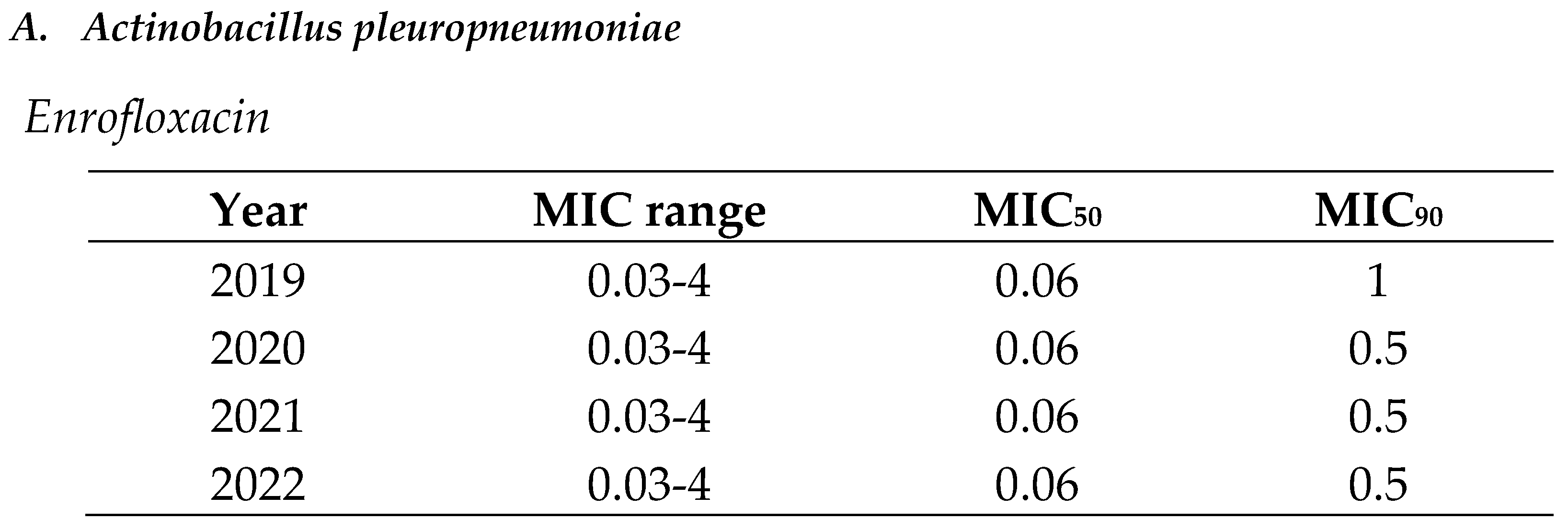
MIC distributions (MIC range, MIC50 and MIC90) are showed from Table 2, Table 3 and Table 4 for A. pleuropneumoniae, P. multocida and E. coli to quinolones (enrofloxacin and marbofloxacin), cephalosporins (ceftiofur and cefquinome) and polymyxins (colistin) during the study period.
In the case of quinolones, there were isolates with low and extremely high MIC values in the same distribution (MIC range of 0.03-4) for all the bacterial pathogens, but the MIC90 was lower for respiratory pathogens (A. pleuropneumoniae and P. multocida) than for digestive ones (E. coli) across the study period. Moreover, MIC90 remained stable across the study period for all the bacterial pathogens or slightly decreased in the case of APP (Table 2).

Table 2.
Minimum inhibitory concentration (MIC) distribution values of (A) Actinobacillus pleuropneumoniae, (B) Pasteurella multocida and (C) Escherichia coli to quinolones (enrofloxacin and marbofloxacin) from 2019 to 2022 in Spain.
Table 2.
Minimum inhibitory concentration (MIC) distribution values of (A) Actinobacillus pleuropneumoniae, (B) Pasteurella multocida and (C) Escherichia coli to quinolones (enrofloxacin and marbofloxacin) from 2019 to 2022 in Spain.
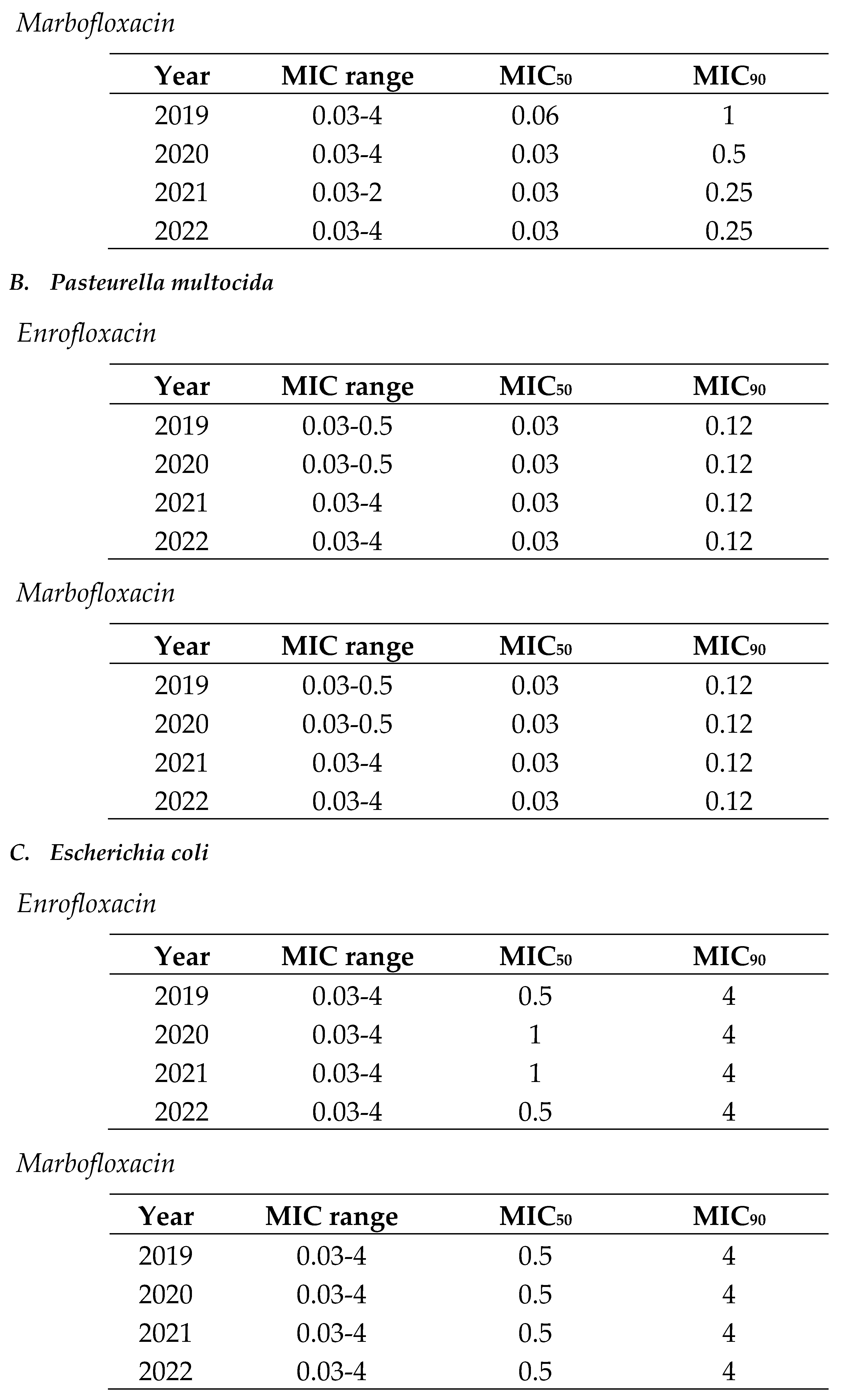
In the case of cephalosporins, the MIC range for respiratory pathogens (0.06-1) was smaller than for digestive ones (0.06-8) (Table 3 and Table 4). Thus, there were E. coli isolates with low and extremely high MIC values in the same distribution. Moreover, MIC90 was also lower for respiratory pathogens (A. pleuropneumoniae and P. multocida) than for digestive ones (E. coli). In both cases, MIC90 remained stable across the study period (Table 3 and Table 4).
Table 3.
Minimum inhibitory concentration (MIC) distribution values of Actinobacillus pleuropneumoniae and Pasteurella multocida to ceftiofur from 2019 to 2022 in Spain.
Table 3.
Minimum inhibitory concentration (MIC) distribution values of Actinobacillus pleuropneumoniae and Pasteurella multocida to ceftiofur from 2019 to 2022 in Spain.
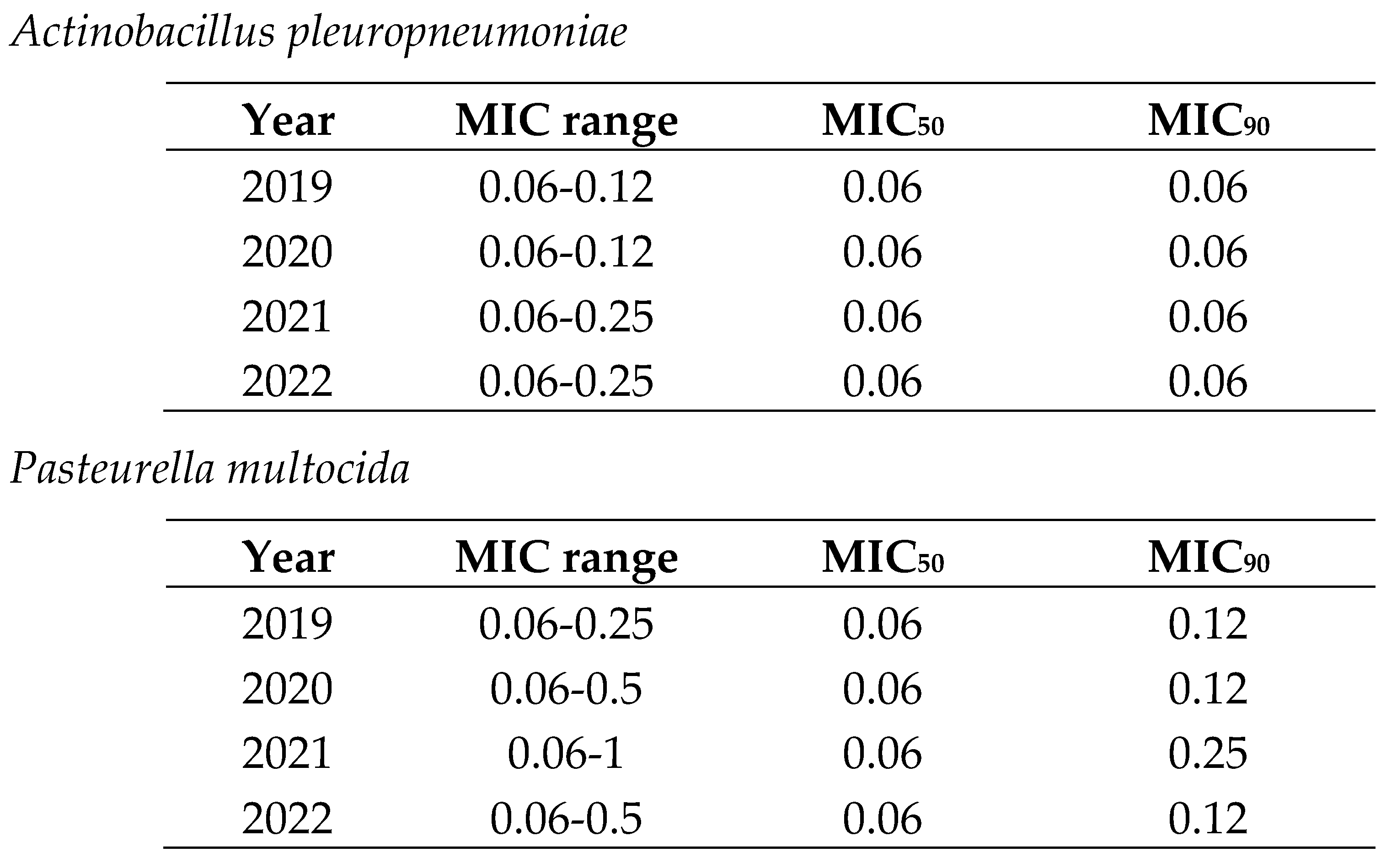
Table 4.
Minimum inhibitory concentration (MIC) distribution values of Escherichia coli to cephalosporins (ceftiofur and cefquinome) from 2019 to 2022 in Spain.
Table 4.
Minimum inhibitory concentration (MIC) distribution values of Escherichia coli to cephalosporins (ceftiofur and cefquinome) from 2019 to 2022 in Spain.
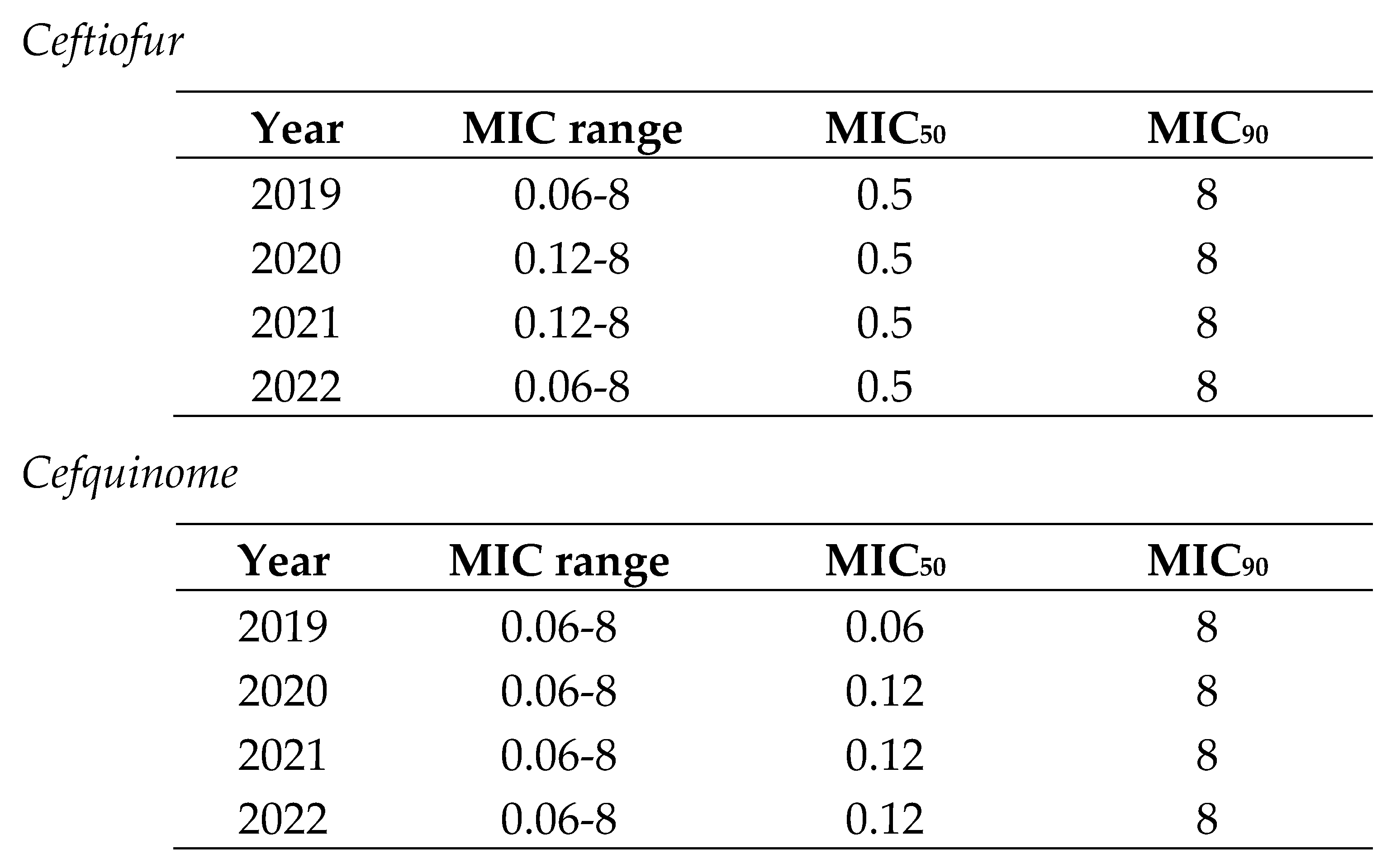
In the case of polymyxins, MIC90 sustainably decreased from 2019 to 2022 but MIC range remained similar during the study period (Table 5).
2.3. Logistic and multinominal model for quinolones
After statistical analysis using dichotomized (susceptible/resistant) and categorized MIC data (Figure 1 and Figure 2), not significant temporal trends were observed for susceptibility to enrofloxacin in E. coli and P. multocida (p>0.05). Contrarily, for A. pleuropneumoniae, a significant temporal trend (p=0.002) was detected for this antimicrobial. Isolates from 2020 had significantly increased odds of being more susceptible to enrofloxacin than isolates from 2019 comparing MIC category 1 versus 3 and 1 versus 4. Moreover, isolates from 2020 (Table 6 and Figure 2) had also increased odds of being more susceptible than isolates from 2019 using dichotomized MIC data (susceptible/resistant, p=0.0002).
In the case of P. multocida (Figure 2 and Figure 3), no temporal trend for susceptibility to marbofloxacin was observed (p>0.05). However, in the case of A. pleuropneumoniae, a significant temporal trend (p<0.0001) was detected where isolates from 2020 had significantly increased odds of being more susceptible than isolates from 2019 comparing MIC category 1 versus 3 and 1 versus 4 (Table 7). Thus, isolates from 2021 had significantly decreased odds of being more susceptible than isolates from 2020 comparing MIC category 1 versus 2. However, when using dichotomized MIC data between 2020 and 2019 significant temporal trend was observed for this bacteria-drug combination (Table 7 and Figure 2). Finally, in the case of E. coli and marbofloxacin, a significant temporal trend (p<0.0001) was also observed, where isolates from 2020 had significantly increased odds of being more susceptible than isolates from 2019 comparing MIC category 1 versus 2 but they had significantly decreased odds of being more susceptible when comparing MIC category 1 versus 3 between these years (Table 8). Interestingly, it was not observed any significant trend using dichotomized data for this drug/microorganism combination (Table 8 and Figure 2).
Figure 2.
Percentage of susceptible isolates by year for enrofloxacin (A) and marbofloxacin (B) of Actinobacillus pleuropneumoniae (circle), Pasteurella multocida (square) and Escherichia coli (diamond), using CLSI and EUCAST clinical breakpoints as detailed in the material and method section.
Figure 2.
Percentage of susceptible isolates by year for enrofloxacin (A) and marbofloxacin (B) of Actinobacillus pleuropneumoniae (circle), Pasteurella multocida (square) and Escherichia coli (diamond), using CLSI and EUCAST clinical breakpoints as detailed in the material and method section.
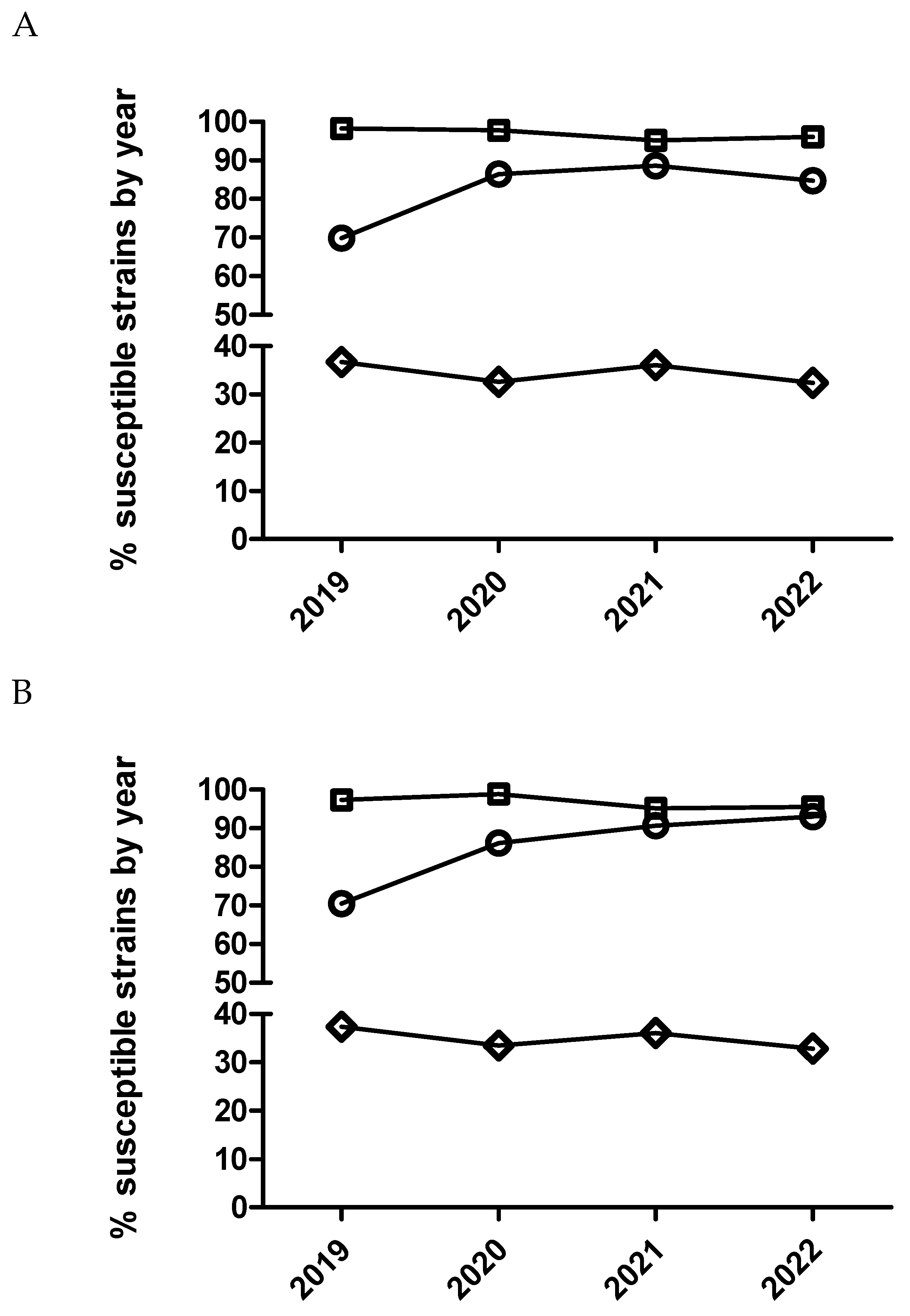
Figure 3.
Percentage of Escherichia coli (A), Actinobacillus pleuropneumoniae (B) and Pasteurella multocida (C) isolates belonging to antimicrobial susceptibility category 1 (circle), category 2 (square), category 3 (diamond) and category 4 (triangle) for marbofloxacin being category 1 the most susceptible (lowest MIC values) and category 4 the less susceptible one (highest MIC value).
Figure 3.
Percentage of Escherichia coli (A), Actinobacillus pleuropneumoniae (B) and Pasteurella multocida (C) isolates belonging to antimicrobial susceptibility category 1 (circle), category 2 (square), category 3 (diamond) and category 4 (triangle) for marbofloxacin being category 1 the most susceptible (lowest MIC values) and category 4 the less susceptible one (highest MIC value).
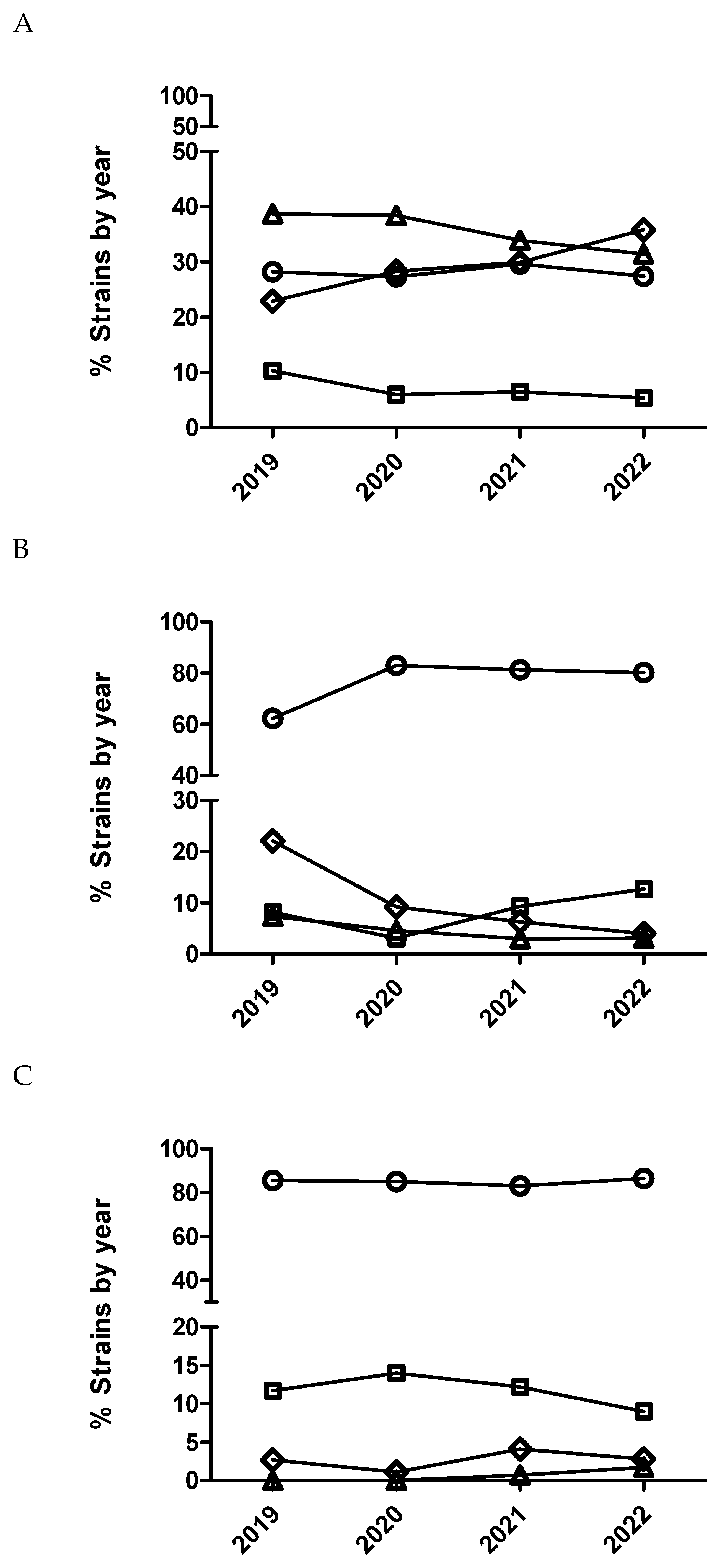
Table 7.
The adjusted odds ratio (95% confidence interval) describing the annual variation in susceptibility of A. pleuropneumoniae isolates to marbofloxacin using the logistic and multinominal regression model. The number of A. pleuropneumoniae isolates by year is detailed in Table 1.
Table 7.
The adjusted odds ratio (95% confidence interval) describing the annual variation in susceptibility of A. pleuropneumoniae isolates to marbofloxacin using the logistic and multinominal regression model. The number of A. pleuropneumoniae isolates by year is detailed in Table 1.
| Logistic analysis (susceptible/resistant) |
Multinominal analysis (MIC outcome categories being compared) | |||
|---|---|---|---|---|
| Predictor variable | NA | 1 vs 2 | 1 vs 3 | 1 vs 4 |
| Year | P<0.0001 | P<0.0001 | ||
| 20 vs 19 | 2.6 (1.5-4.6) | NS | 3.2 (2.1-4.8) | 2.2 (1.1-3.9) |
| 21 vs 20 | NS | 0.38 (0.18-0.69) | NS | NS |
| 22 vs 21 | NS | NS | NS | NS |
NS means not significant (p>0.05). NA means not applicable.
Table 8.
The adjusted odds ratio (95% confidence interval) describing the annual variation in susceptibility of E. coli isolates to marbofloxacin using the logistic and multinominal regression model. The number of E. coli isolates by year is detailed in Table 1.
Table 8.
The adjusted odds ratio (95% confidence interval) describing the annual variation in susceptibility of E. coli isolates to marbofloxacin using the logistic and multinominal regression model. The number of E. coli isolates by year is detailed in Table 1.
| Logistic analysis (susceptible/resistant) |
Multinominal analysis (MIC outcome categories being compared) | |||
|---|---|---|---|---|
| Predictor variable | NA | 1 vs 2 | 1 vs 3 | 1 vs 4 |
| Year | NS | P<0.0001 | ||
| 20 vs 19 | NS | 1.5 (1.2-1.9) | 0.8 (0.7-0.9) | NS |
| 21 vs 20 | NS | NS | NS | NS |
| 22 vs 21 | NS | NS | NS | NS |
NS means not significant (p>0.05). NA means not applicable.
2.4. Logistic and multinominal model for 3rd and 4th cephalosporins
The multinominal regression analysis for E. coli identified significant annual variation in susceptibility to ceftiofur (Figure 4). Thus, E. coli isolates from 2020 had significantly decreased odds of being more susceptible than isolates from 2019 comparing all the MIC categories (1 versus 2, 1 versus 3 and 1 versus 4) (Table 9). However, E. coli isolates from 2021 had significantly increased odds of being more susceptible than isolates from 2020 comparing MIC category 1 versus 4. Using dichotomized data, E. coli isolates from 2020 had also significant odds of being less susceptible than isolates from 2019 (Table 9 and Figure 5). In the case of E. coli and cefquinome (4th generation cephalosporin), no significant temporal trend in antimicrobial susceptibility (p>0.05) was detected using the multinominal regression analysis (Figure 6) whereas, the dichotomized analyses, showed that E. coli isolates from 2020 had significantly decreased odds (0.70-(0.52-0.96)) of being more susceptible than isolates from 2019 (Figure 5).
Figure 4.
Percentage of Escherichia coli (A), Actinobacillus pleuropneumoniae (B) and Pasteurella multocida (C) isolates belonging to antimicrobial susceptibility category 1 (circle), category 2 (square), category 3 (diamond) and category 4 (triangle) for ceftiofur being category 1 the most susceptible (lowest MIC values) and category 4 the less susceptible one (highest MIC value).
Figure 4.
Percentage of Escherichia coli (A), Actinobacillus pleuropneumoniae (B) and Pasteurella multocida (C) isolates belonging to antimicrobial susceptibility category 1 (circle), category 2 (square), category 3 (diamond) and category 4 (triangle) for ceftiofur being category 1 the most susceptible (lowest MIC values) and category 4 the less susceptible one (highest MIC value).
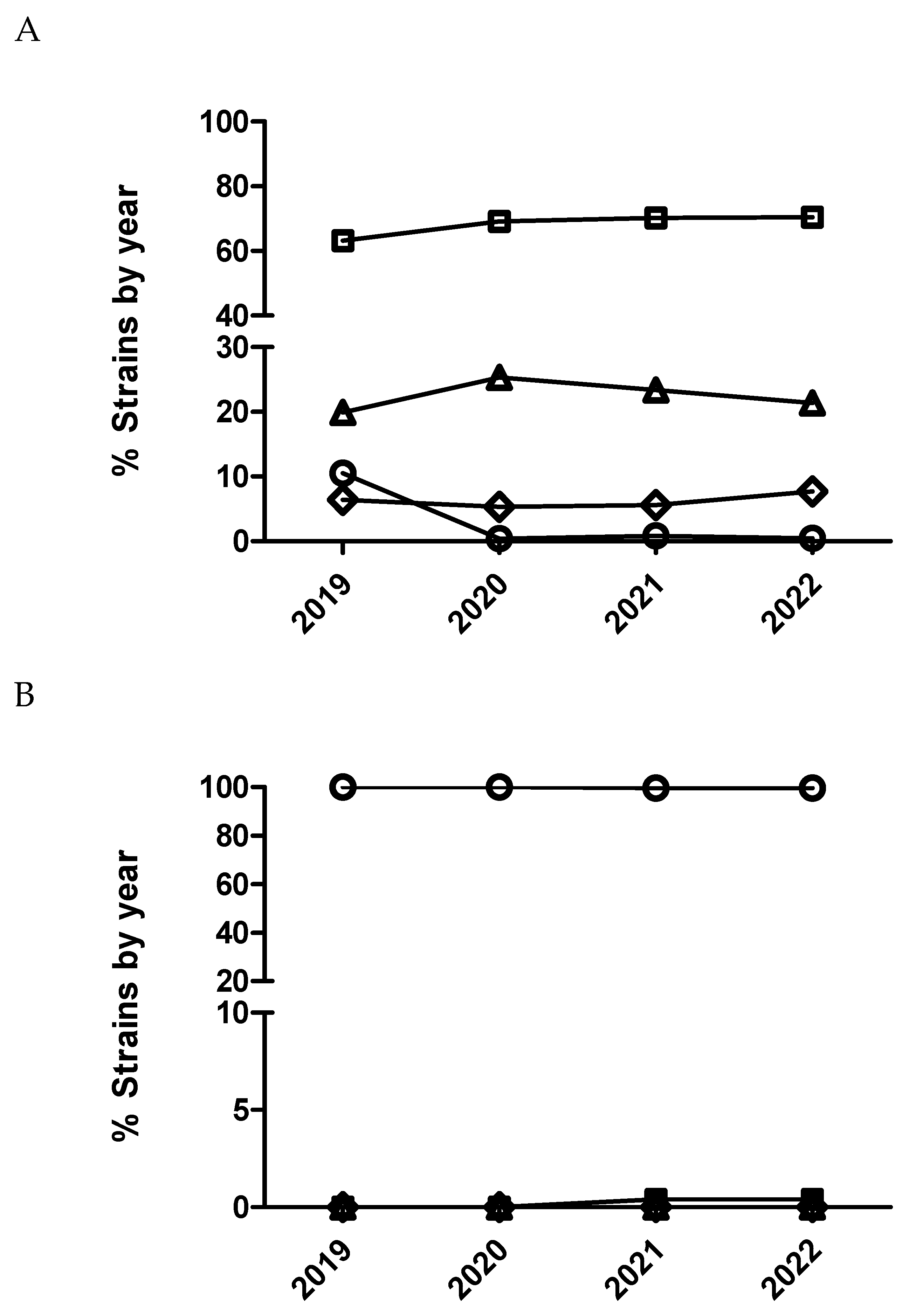
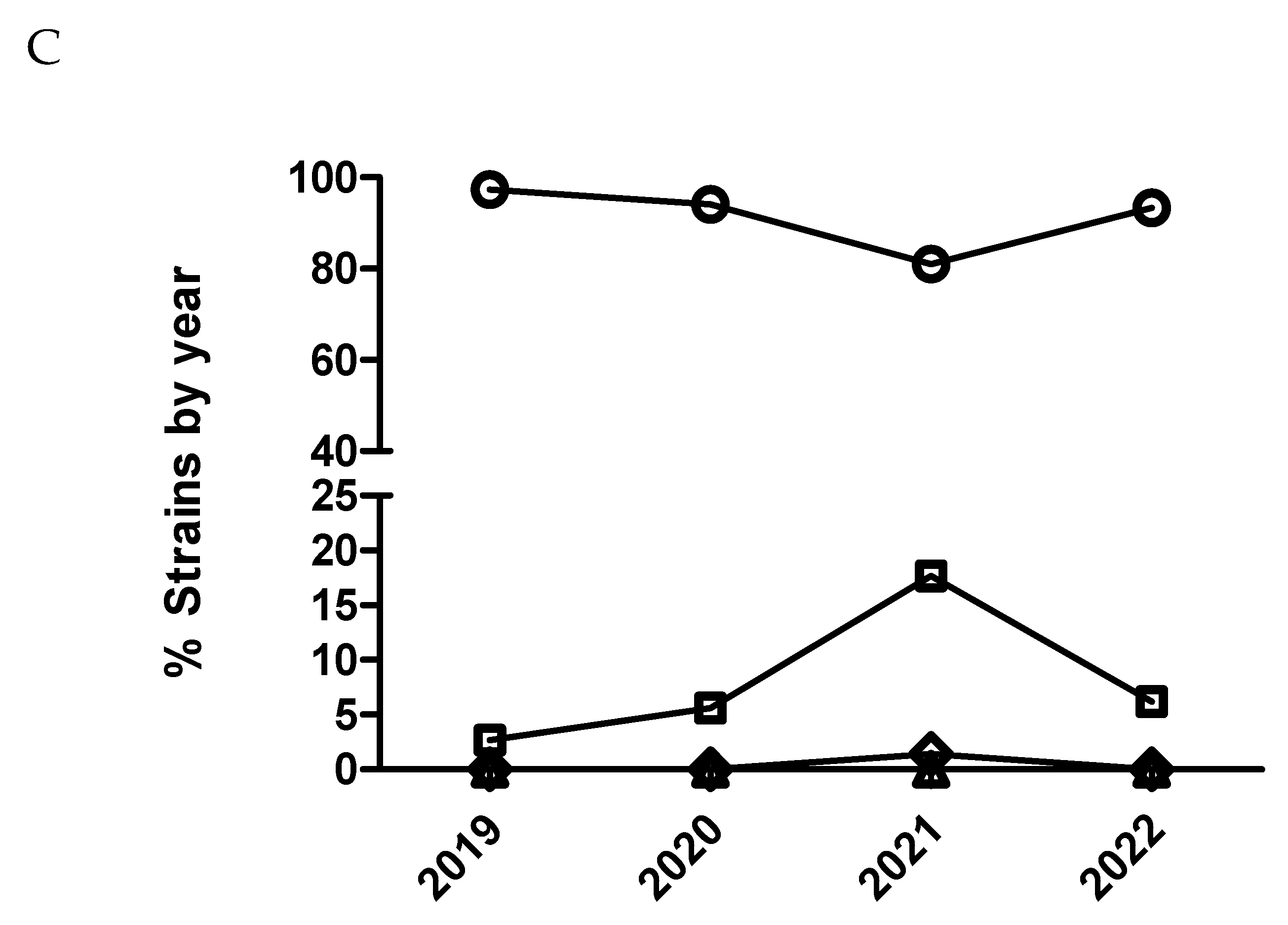
Table 9.
The adjusted odds ratio (95% confidence interval) describing the annual variation in susceptibility of E. coli isolates to ceftiofur using the logistic and multinominal regression model. The number of E. coli isolates by year is detailed in Table 1.
Table 9.
The adjusted odds ratio (95% confidence interval) describing the annual variation in susceptibility of E. coli isolates to ceftiofur using the logistic and multinominal regression model. The number of E. coli isolates by year is detailed in Table 1.
| Logistic analysis (susceptible/resistant) |
Multinominal analysis (MIC outcome categories being compared) | |||
|---|---|---|---|---|
| Predictor variable | NA | 1 vs 2 | 1 vs 3 | 1 vs 4 |
| Year | P=0.15 | P<0.0001 | ||
| 20 vs 19 | 0.73 (0.55-0.98) | 0.10 (0.06-0.16) | 0.11(0.06-0.19) | 0.10 (0.05-0.15) |
| 21 vs 20 | NS | NS | NS | 3.1 (1.2-12.4) |
| 22 vs 21 | NS | NS | NS | NS |
NS means not significant (p>0.05). NA means not applicable.
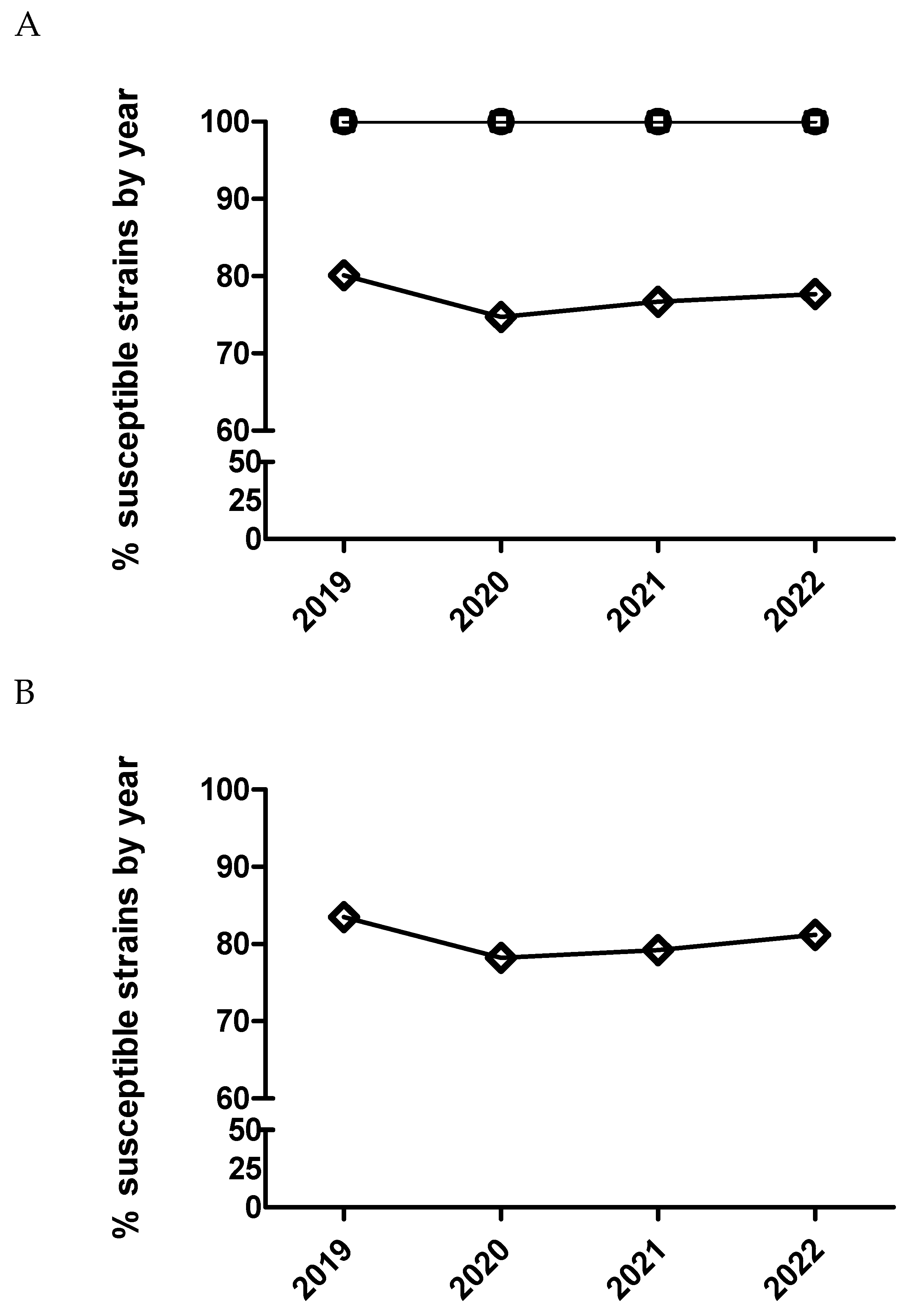
Figure 5.
Percentage of susceptible isolates by year for ceftiofur (A) and cefquinome (B) of Actinobacillus pleuropneumoniae (circle), Pasteurella multocida (square) and Escherichia coli (diamond), using CLSI and EUCAST clinical breakpoints as detailed in the material and method section.
Figure 5.
Percentage of susceptible isolates by year for ceftiofur (A) and cefquinome (B) of Actinobacillus pleuropneumoniae (circle), Pasteurella multocida (square) and Escherichia coli (diamond), using CLSI and EUCAST clinical breakpoints as detailed in the material and method section.
In the case of A. pleuropneumoniae, the percentage of isolates belonging to category 1 for ceftiofur was close to 100% across the study period without observing any temporal trend (p>0.05) either with dichotomized or categorized MIC data. In the case of P. multocida and ceftiofur (Figure 4), a significant temporal trend in antimicrobial susceptibility was observed during the study period (p<0.05). Thus, isolates from 2020 had significantly decreased odds of being more susceptible than isolates from 2019 comparing MIC category 1 versus 2 (Figure 4 and Table 10). However, isolates from 2022 had significantly increased odds of being more susceptible than isolates from 2021 comparing MIC category 1 versus 2 (Table 10). Interestingly, not significant differences were observed using dichotomized data (susceptible/resistant) for this combination of drug/microorganism (Figure 5).
Table 10.
The adjusted odds ratio (95% confidence interval) describing the annual variation in susceptibility of P. multocida isolates to ceftiofur using the logistic and multinominal regression model. The number of P. multocida isolates by year is detailed in Table 1.
Table 10.
The adjusted odds ratio (95% confidence interval) describing the annual variation in susceptibility of P. multocida isolates to ceftiofur using the logistic and multinominal regression model. The number of P. multocida isolates by year is detailed in Table 1.
| Logistic analysis (susceptible/resistant) |
Multinominal analysis (MIC outcome categories being compared) | |||
|---|---|---|---|---|
| Predictor variable | NA | 1 vs 2 | 1 vs 3 | 1 vs 4 |
| Year | NS | p=0.0002 | ||
| 20 vs 19 | NS | 0.39 (0.13-0.88) | NS | NA |
| 21 vs 20 | NS | NS | NS | NA |
| 22 vs 21 | NS | 3.1 (1.8-5.3) | NS | NA |
NS means not significant (p>0.05). NA means not applicable. In this case, there is no isolates belonging to MIC category 4 (the less susceptible).
Figure 6.
Percentage of Escherichia coli isolates belonging to antimicrobial susceptibility category 1 (open circle), category 2 (open squares), category 3 (open diamond) and category 4 (open triangle) for cefquinome being category 1 the most susceptible (lowest MIC values) and category 4 the less susceptible one (highest MIC value).
Figure 6.
Percentage of Escherichia coli isolates belonging to antimicrobial susceptibility category 1 (open circle), category 2 (open squares), category 3 (open diamond) and category 4 (open triangle) for cefquinome being category 1 the most susceptible (lowest MIC values) and category 4 the less susceptible one (highest MIC value).
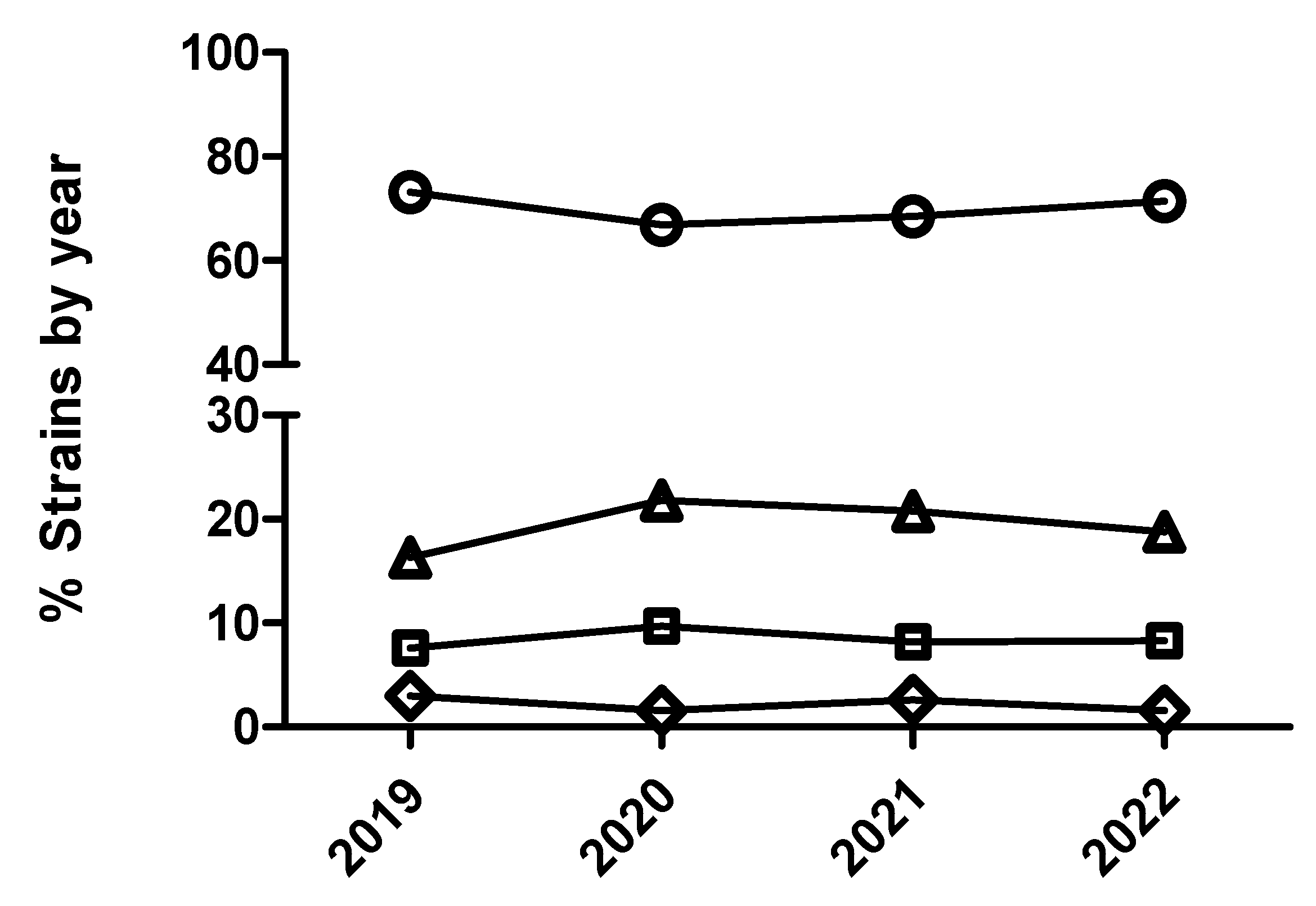
2.5. Logistic and multinominal model for polymyxins
Only E. coli was tested against this antimicrobial. A significant temporal trend (p<0.0001) was detected, where isolates from 2020 had significantly increased odds of being more susceptible than isolates from 2019 comparing MIC category 1 versus 2 and 3. This same result was also observed with isolates from 2021 versus isolates from 2020 but only comparing MIC category 1 versus 2 (Figure 7 and Table 11). On the other hand, isolates from 2022 had significantly decreased odds of being more susceptible than isolates from 2021 comparing MIC category 1 versus the rest of categories (Figure 7B and Table 11). However, using dichotomized MIC data, only isolates from 2020 had significant increased odds of being more susceptible than isolates from 2019 (Figure 8 and Table 11).
3. Discussion
Antimicrobial susceptibility is usually measured by the minimum inhibitory concentration (MIC), which is the lowest concentration that stops in vitro growth of the targeted bacteria using microdilution methods in veterinary laboratories. Modelling the MIC values is challenging since these types of data are interval-censored and ordinal [36,37]. One approach to deal with these data is to dichotomize the MIC values into two categories, resistant (R) and susceptible (S) using established clinical breakpoints or epidemiological cut-off values (ECOFF), followed by logistic regression [38,39]. However, this is not an ideal approach since there is a loss of quantitative information from the MIC values when they are dichotomized [36,40]. Other critical point to dichotomize the MIC values into R and S categories is the existence of accepted clinical breakpoints to obtain comparable results between different studies. In the case of pig respiratory pathogens, there are a reasonable amount of internationally accepted clinical breakpoints, but this is not the case for pig enteric pathogens. Moreover, EUCAST ECOFFs are missing for 45.3% (MIC) and 76.9% (disk diffusion) of bacterial species in the veterinary field [41]. Since we work with clinical cases, it was decided to interpret our MIC results using clinical breakpoints instead of ECOFFs. Therefore, we can monitor the antimicrobial susceptibility pattern for different antibiotics, but we cannot monitor resistance in bacterial populations as suggested by the EARS-VET surveillance network [41]. Moreover, our study is based on clinical cases (passive collection) whose representativeness of the general animal population is unknown [42]. Considering the limited information available for some antibiotic-microorganism pair, we have extrapolated clinical breakpoints available for quinolones and cephalosporins and respiratory pathogens [43,44] to enteric ones, and we have used the clinical breakpoint for colistin and E. coli from humans [45]. This approach seems reasonable to study the antimicrobial susceptibility temporal trends for all the porcine pathogens, but it has not allowed extrapolating directly these findings to clinical efficacy in pigs, especially for digestive pathogens. Despite these limitations, we consider that our data provide robust information about the evolution of the antimicrobial susceptibility pattern of the main pig pathogens in Spain during the study period.
The qualitative categorization into S and R, does not allow to determine dynamics of bacterial population, in particular wild type populations approaching the clinical breakpoint. This is especially important for cases of decrease susceptibility to antimicrobials associated to punctual mutations, like fluoroquinolones and E. coli, where increase in the MIC is associated with chromosomic mutations in the quinolone resistance determining regions [46]. MIC outcome data could be more appropriately modelled using statistical models other than logistic regression such as Cox proportional hazards, multinomial logistic, ordinal logistic, linear and tobit regression models [36,37,38,40,47]. In this case, we have used a multinominal logistic model based on distributing the range of MIC values into four categories (from 1 to 4) that include two MIC values in each category, being category 1 the most susceptible (lowest MIC value) and 4 the less susceptible (highest MIC value), as suggested by other authors with a similar database for E. coli [48]. Finally, the antimicrobial panel was selected to represent commonly used compounds for the treatment of pig diseases in practice [31,32], and not focused on monitoring antimicrobial resistance in surveillance programs. This is a clear limitation of this study since antimicrobials tested herein were not the same for all the porcine pathogens.
Our data clearly showed a different pattern in the evolution of antimicrobial susceptibility for each combination of drug and microorganism. However, in the case of both fluoroquinolones, marbofloxacin and enrofloxacin, in combination with A. pleuropneumoniae, the proportion of isolates susceptible to each of the antimicrobials was practically the same. Similarly occurred for P. multocida, indicating that testing one of those fluoroquinolones in these two pathogens would be sufficient to test for this antimicrobial family [32]. Contrarily, data on susceptibility obtained for E. coli in combination with ceftiofur, could not be extrapolated to cefquinome as it has been also previously suggested by other authors [49]. This is not surprising as cefquinome has been reported not useful in separating isolates with extended spectrum betalactamases or plasmidic AmpC from the cephalosporin-susceptible isolates [50]. These results reinforced that the evolution of antimicrobial susceptibility must be studied in a case-by-case situation where generalization for drug families and bacteria is not possible as described previously [32]. Finally, one interesting line of research could be studying the evolution mechanisms shaping the maintenance of antibiotic resistance in pig pathogens as carried out by Durao et al [51] but it is out of the scope of this paper.
In general terms, pig pathogens involved in respiratory diseases analyzed herein appeared to remain susceptible or tended to increase susceptibility to critical antimicrobials over the study period. For E. coli, there was also a tendency to increase susceptibility for most antimicrobials, except for ceftiofur, where there was a significant decrease in susceptibility for MIC category 1 from 2019 to 2020. Taken together, results obtained using dichotomized versus categorized MIC data were generally similar for all the pairs of drug/microorganism combinations with some exceptions, where categorized MIC was more sensitive detecting slight changes in antimicrobial susceptibility patterns (i.e. cefquinome and marbofloxacin in combination with E. coli). Finally, for the combination colistin with E. coli, by using dichotomized MIC data, a dramatic increase in susceptibility to colistin from 2019 to 2021 was observed, with slight decrease in 2022. This is interesting since there was a voluntary reduction in the sales of colistin in pig production in Spain from 34.9 mg/PCU to 3 mg/PCU between 2015 and 2018, which could explain these results, but we do not have figures of colistin consumption by farm and a sound study linking consumption with antimicrobial susceptibility cannot be carried out with our database. Still, by using dichotomized MIC data (S and R, Figure 8) this decrease in susceptibility observed for the year 2022 was not detected, suggesting that categorized MIC data may be more sensible that dichotomized to detect slight changes in antimicrobial susceptibility pattern. Despite not achieving enough sample size to have robust data, the evolution of antimicrobial susceptibility for Salmonella spp and colistin is very close to the observed tendency for E. coli and colistin (additional Figure S1). It must be highlighted that both bacteria are in the same ecological niche.
In Spain, the antimicrobial susceptibility for last resource antimicrobials in pig pathogens remained stable or increased in the last four years. These are sound results in terms of preserving the efficacy of critical important antimicrobials and minimizing the burden and spread of resistance from farm to fork.
4. Materials and Methods
4.1. Clinical samples
Between January 2019 and December 2022, samples were taken from diseased or recently deceased pigs from farms across Spain showing acute clinical signs of respiratory tract infections or pigs showing diarrhea. None of these animals had been exposed to antimicrobial treatment for, at least, 15 days prior sampling. Thus, the sampled animals were between 3 and 24 weeks old showing overt respiratory symptoms with or without depression and/or hyperthermia (>39.8ºC). For each clinical case, samples of lungs of two recently deceased pigs (<12 hours) were submitted under refrigeration to the laboratory. If no recently dead pigs were suitable for sampling, at least, two animals with acute respiratory signs were humanely sacrificed and lung samples were drawn. On the other hand, for piglets showing PWD, the sampled animals were between 3 and 12 weeks old showing clinical symptoms of the disease. Intestinal content obtained from humanly euthanized animals or watery diarrhea from sick pigs were obtained. In both cases, the samples were submitted under refrigeration to the laboratory and processed during the following 24 hours after collection. Only one isolate was included by farm across the study to avoid redundancy and overrepresentation of bacterial clones.
4.2. Bacterial isolation and identification
Clinical specimens were cultured aseptically onto blood agar (Columbia agar with 5% Sheep blood, 254005 BD), chocolate agar (GC II agar with IsoVitaleX, 254060, BD or blood Agar No. 2 Base, 257011, BD) and MacConkey agar (4016702, Biolife Italiana Srl) and incubated at 35–37°C in aerobic conditions with 5-10% CO2 for 24–48 hours to address the isolation of respiratory bacterial pathogens. Finally, for the isolation of digestive pathogens, specimens were cultured aseptically onto blood agar, MacConkey agar and Xylose-Lysine-Desoxycholate Agar (XLD, CM0469, Oxoid). The plates were incubated at 35–37°C in aerobic conditions for 24 hours.
Identification of isolates for respiratory pathogens and enteric pathogens was carried out by matrix assisted laser desorption ionization-time of flight (MALDI-TOF Biotyper System, Bruker Daltonics, Bremen, Germany) as previously described (25). Individual isolates were stored at -80°C in brain heart infusion (CM1135, Oxoid) with 30% of glycerol (G9012, Sigma-aldrich).
4.3. Antimicrobial susceptibility testing
Antimicrobial susceptibility testing was determined using minimum inhibitory concentration (MIC) value for each combination of bacterial species and antimicrobial tested. Thus, MIC was performed in accordance with the recommendations presented by the Clinical and Laboratory Standards Institute [31,32] in a customized 96-well microtitre plate (Sensititre, Trek diagnostic Systems Inc., East Grinstead, UK) containing a total of 12 and 8 antibiotics/concentrations for respiratory and digestive pathogens, respectively. The antimicrobials tested for swine respiratory pathogens belong to category D [15]: Sulfamethoxazole/trimethoprim, doxycycline, oxytetracycline and amoxicillin; Category C: Florfenicol, tiamulin, tulathromycin, tildipirosin and tilmicosin and category B: Ceftiofur, enrofloxacin and marbofloxacin. On the other hand, the antimicrobials tested for swine enteric pathogens belong to category D: Sulfamethoxazole/trimethoprim and spectinomycin; Category C: florfenicol, apramycin, gentamycin, neomycin and amoxicillin/clavulanic acid and category B: ceftiofur, cefquinome enrofloxacin, marbofloxacin and colistin.
Bacteria were thawed, cultured on chocolate agar or blood agar, and incubated at 35-37ºC in aerobiosis (or with 5-10% CO2 for APP) for 18-24h. Three to five colonies were picked and emulsified in demineralized water (or Cation Adjusted Muëller-Hinton Broth (CAMHB) for APP) to obtain a turbidity of 0.5 McFarland standard (Sensititre™ nephelometer V3011). Suspensions were further diluted in CAMHB for E. coli, CAMHB or CAMHB with 2.5-5% Lysed Horse Blood for P. multocida and Veterinary Fastidious Medium (VFM) or Mueller Hinton Fastidious broth with Yeast (MHF-Y) for APP to reach a final inoculum concentration of 5x105 cfu/ml. Then, the Sensititre panel was reconstituted by adding 100μl/well of the inoculum. Plates containing E. coli isolates were incubated at 35 ± 2ºC for 16-20h, P. multocida isolates were incubated at 35 ± 2ºC for 18-24h. In the case of APP isolates, plates were covered with a perforated seal and incubated at 35 ± 2ºC with 5-10% CO2 for 20-24h.
The antibiotic panels were read manually using Sensititre™ Vizion (V2021) and the MIC value was established as the lowest drug concentration inhibiting visible growth. For each isolate tested, a colony count and a purity check were performed following CLSI and manufacturer recommendations. Moreover, quality control strains were also included. Thus, Actinobacillus pleuropneumoniae (ATCC 27090™), Escherichia coli (ATCC 25922™), Streptococcus pneumoniae (ATCC 49619™) and Enterococcus faecalis (ATCC 29212™) were included as quality control following CLSI recommendations [31,32]. The MICs of the quality control strains had to be within acceptable CLSI ranges to accept the results obtained in the laboratory.
4.5. Statistical methods
All the data analysis was carried out with JMP®, Version 13 (SAS Institute Inc., Cary, NC, USA, 1989–2019). Descriptive statistics (MIC range, MIC50 and MIC90) were performed to summarize the distribution of the isolates within each MIC category. The number of categories was based on distributing the range of MIC values in four categories (from one to four) that include two MIC values for category, being category one the most susceptible (lowest MIC value) and category four the less susceptible (highest MIC value). The range of concentrations tested were 0,06-8, 0,03-4, 0,25-32 g/mL for 3rd and 4th cephalosporins, quinolones and polymyxins, respectively. Moreover, clinical susceptibility (susceptible/resistant for each isolate) was determined according to CLSI clinical breakpoints for APP, P. multocida, and E. coli for quinolones and cephalosporins and EUCAST guidelines for colistin in the case of E. coli, respectively [43,44,45] (Table 12).
A logistic (susceptible/resistant for each isolate) and multinomial logistic regression model (four MIC categories) was used to analyze the susceptibility data for the antimicrobials from year 2019 to 2022, only for those pairs of antimicrobial/microorganisms if at least 100 isolates were available for each year, as recommended by De Jong et al (2022) [16]. Susceptible/resistant and categorized MIC data (MIC category 1, 2, 3 and 4) were used for logistic and multinominal logistic regression model, respectively as dependent variables, and the year as independent one. Thus, year of sampling was categorized by individual years and modelled as a hierarchical indicator variable, where for each year the preceding year was used as the referent [52]. The final multinomial model was executed with outcome category 1 as the base referent category (the most susceptible one). The model assumptions and goodness-of-fit were evaluated as appropriate for these models [52]. Thus, the level of significance used to reject the null hypothesis was p ≤ 0.05.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Percentage of Salmonella spp strains belonging to antimicrobial susceptibility category 1 (open circle), category 2 (open squares), category 3 (open diamond) and category 4 (open triangle) for colistin being category 1 the most susceptible (lowest MIC value) and category 4 the less susceptible one (highest MIC value).
Author Contributions
Conceptualization, A.V., L.M-G and L.F.; methodology, A.V., E.N., V.E.-T. and J.B .; formal analysis, L.F.; writing—original draft preparation, L.F., A.V, L.M.-G. and A.V.; writ-ing—review and editing, L.F., L.M.-G. and A.V.; funding acquisition, E.N. and V.E.-T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable because this study does not involve humans or animals. All the samples were sent for diagnosis due to clinical cases.
Informed Consent Statement
Written informed consent has been obtained from veterinarians to publish this paper. This informed consent is signed whereas filling the information about the case.
Data Availability Statement
The data presented in this study are available on reasonable request from the corresponding author. The data are not publicly available due to confidentiality issues related with clinical cases.
Acknowledgments
This study was carried out with the support of the Porcine Sanitation Group of Lleida, Spain (Grup de Sanejament Porcí- GSP) in collaboration with swine practitioners working in the field.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aarestrup, F.M.; Wegener, H.C.; Collignon, P. Resistance in bacteria of the food chain: epidemiology and control strategies. Expert Rev Anti Infect Ther 2008, 6, 733–750. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. 2015. Global action plan on antimicrobial resistance. World Health Organization. Available online: https://apps.who.int/iris/handle/10665/193736 (accessed on 1 September 2023).
- Guidelines for the Prudent Use of Antimicrobials in Veterinary Medicine. (2015/C299/04). 2015. Available online: https://ec.europa.eu/health/sites/health/files/antimicrobial_resistance/docs/2015_prudent_use_guidelines_en.pdf (accessed on 1 September 2023).
- Magnusson, U.; Sternberg, S.; Eklund, G.; Rozstalnyy, A. Prudent and efficient use of antimicrobials in pigs and poultry. FAO Animal Production and Health Manual. Available online: https://www.fao.org/3/ca6729en/CA6729EN.pdf (accessed on 1 September 2023).
- Holmes, A.H.; Moore, L.S.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Guerin, P.J.; Piddoc, L.J. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.M.; Beller, E.; Glasziou, P.; Clark, J.; Ranakusuma, R.W.; Byambasuren, O.; Bakhit, M.; Page, S.W.; Trott, D.; Mar, C.D. Is antimicrobial administration to food animals a direct threat to human health? A rapid systematic review. Int J Antimicrob Agents 2018, 52, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, B.; Paterson, D.L.; Mollinger, J.L.; Rogers, B.A. Do human extraintestinal Escherichia coli infections resistant to expanded-spectrum cephalosporins originate from food-producing animals? A systematic review. Clin Infect Dis 2015, 60, 439–452. [Google Scholar] [CrossRef]
- Mughini-Gras, L.; Dorado-García, A.; van Duijkeren, E.; van den Bunt, G.; Dierikx, C.M.; Bonten, M.J.M.; Bootsma, M.C.J.; Schmitt, H.; Hald, T.; Evers, E.G.; et al. ESBL Attribution Consortium. Attributable sources of community-acquired carriage of Escherichia coli containing β-lactam antibiotic resistance genes: a population-based modelling study. Lancet Planet Health 2019, 3, e357–e369. [Google Scholar] [CrossRef] [PubMed]
- Martak, D.; Guther, J.; Verschuuren, T.D.; Valot, B.; Conzelmann, N.; Bunk, S.; Riccio, M.E.; Salamanca, E.; Meunier, A.; Henriot, C.P.; et al. Populations of extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella pneumoniae are different in human-polluted environment and food items: a multicentre European study. Clin Microbiol Infect 2022, 28, e7–e447. [Google Scholar] [CrossRef]
- Regulation (EU). 2019/6 of the European Parliament and of the Council of 11 December 2018 on Veterinary Medicinal Products and Repealing Directive 2001/82/EC (Text with EEA Relevance). 2019. Available online: https://eur-lex.europa.eu/eli/reg/2019/6/oj (accessed on 1 September 2023).
- Rhouma, M.; Soufi, L.; Cenatus, S.; Archambault, M.; Butaye, P. Current Insights Regarding the Role of Farm Animals in the Spread of Antimicrobial Resistance from a One Health Perspective. Vet Sci 2022, 9, 480. [Google Scholar] [CrossRef]
- Xia, X.; Wang, Z.; Fu, Y.; Du, X.D.; Gao, B.; Zhou, Y.; He, J.; Wang, Y.; Shen, J.; Jiang, H.; et al. Association of colistin residues and manure treatment with the abundance of mcr-1 gene in swine feedlots. Environ Int 2019, 127, 361–370. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, C.; Zhang, R.; Chen, Y.; Shen, Y.; Hu, F.; Liu, D.; Lu, J.; Guo, Y.; Xia, X.; et al. Changes in colistin resistance and mcr-1 abundance in Escherichia coli of animal and human origins following the ban of colistin-positive additives in China: an epidemiological comparative study. Lancet Infect Dis 2020, 20, 1161–1171. [Google Scholar] [CrossRef]
- Allel, K.; Day, L.; Hamilton, A.; Lin, L.; Furuya-Kanamori, L.; Moore, C.E.; Van Boeckel, T.; Laxminarayan, R.; Yakob, L. Global antimicrobial-resistance drivers: an ecological country-level study at the human-animal interface. Lancet Planet Health 2023, 7, e291–e303. [Google Scholar] [CrossRef]
- EMA AMEG 2019. Answer to the request from the European Commission for updating the scientific advice on the impact on public health and animal health of the use of antibiotics in animals—Categorization of antimicrobials. 2019. Available online: https://www.ema.europa.eu/en/documents/other/answer-request-european-commission-updating-scientific-advice-impact-public-health-animal-health-use_en.pdf (accessed on 1 September 2023).
- De Jong, A.; El Garch, F.; Hocquet, D.; Prenger-Berninghoff, E.; Dewulf, J.; Migura-Garcia, L.; Perrin-Guyomard, A.; Veldman, K.T.; Janosi, S.; Skarzynska, M.; et al. European-wide antimicrobial resistance monitoring in commensal Escherichia coli isolated from healthy food animals between 2004 and 2018. J Antimicrob Chemother. 2022, 7, dkac318. [Google Scholar] [CrossRef] [PubMed]
- Dir. Directive 2003/99/EC of The European parliament and of the council of 17 november 2003 on the monitoring of zoonoses and zoonotic agents, amending Council Decision 90/424/EEC and repealing Council Directive 92/117/EEC Official Journal of the European Communities, 325/31, 12.12.2003 (2003/99/EC, 2003).
- Dir. Directive 2020/1729/EC of The European parliament and of the council of 17 November 2020 on the monitoring and reporting of antimicrobial resistance in zoonotic and commensal bacteria and repealing Implementing Decision 2013/652/EU, 387/8, 19.11.2020 (2020/1729/EC). 17 November.
- Lagrange, J.; Amat, J.P.; Ballesteros, C.; Damborg, P.; Grönthal, T.; Haenni, M.; Jouy, E.; Kaspar, H.; Kenny, K.; Klein, B.; et al. Pilot testing the EARS-Vet surveillance network for antibiotic resistance in bacterial pathogens from animals in the EU/EEA. Front Microbiol. 2023, 14, 1188423. [Google Scholar] [CrossRef] [PubMed]
- Mader, R.; Damborg, P.; Amat, J.P.; Bengtsson, B.; Bourély, C.; Broens, E.M.; Busani, L.; Crespo-Robledo, P.; Filippitzi, M.E.; Fitzgerald, W.; et al. EU-JAMRAI. Building the European Antimicrobial Resistance Sur-veillance network in veterinary medicine (EARS-Vet). Euro Surveill. 2021, 26, 2001359. [Google Scholar] [CrossRef]
- Nielsen, S.S.; Bicout, D.J.; Calistri, P.; Canali, E.; Drewe, J.A.; Garin-Bastuji, B.; Gonzales Rojas, J.L.; Gortazar Schmidt, C.; Herskin, M.; et al.; EFSA Panel on Animal Health and Welfare (AHAW) Ad hoc method for the assessment of animal diseases caused by bacteria resistant to antimicrobials. EFSA J. 2021, 19, e06645. [Google Scholar] [PubMed]
- Nielsen, S.S.; Bicout, D.J.; Calistri, P.; Canali, E.; Drewe, J.A.; Garin-Bastuji, B.; Gonzales Rojas, J.L.; Gortazar Schmidt, C.; Herskin, M.; Michel, V.; et al.; EFSA Panel on Animal Health and Welfare (AHAW) Assessment of animal diseases caused by bacteria resistant to antimicrobials: Swine. EFSA J. 2021, 9, e07113. [Google Scholar]
- Holmer, I.; Salomonsen, C.M.; Jorsal, S.E.; Astrup, L.B.; Jensen, V.F.; Høg, B.B.; Pedersen, K. Antibiotic resistance in porcine pathogenic bacteria and relation to antibiotic usage. BMC Vet Res. 2019, 15, 449. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, J.T.; Tokach, M.D.; Dritz, S.S.; DeRouchey, J.M.; Woodworth, J.C.; Goodband, R.D.; Henry, S.C. Postweaning mortality in commercial swine production II: Review of infectious contributing factors. Transl. Anim. Sci. 2020, 4, txaa052. [Google Scholar] [CrossRef]
- Laird, T.J.; Abraham, S.; Jordan, D.; Pluske, J.R.; Hampson, D.J.; Trott, D.J.; O’Dea, M. Porcine enterotoxigenic Escherichia coli: Antimicrobial resistance and development of microbial-based alternative control strategies. Vet. Microbiol. 2012, 258, 109117. [Google Scholar] [CrossRef]
- Saade, G.; Deblanc, C.; Bougon, J.; Marois-Créhan, C.; Fablet, C.; Auray, G.; Belloc, C.; Leblanc-Maridor, M.; Gagnon, C.A.; et al. Coinfections and their molecular consequences in the porcine respiratory tract. Vet. Res. 2020, 51, 80. [Google Scholar] [CrossRef]
- Fairbrother, J.M.; Nadeau, E.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: an update on bacterial types, pathogenesis, and prevention strategies. Anim Health Res Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef]
- Rhouma, M.; Fairbrother, J.M.; Beaudry, F.; Letellier, A. Post weaning diarrhea in pigs: risk factors and non-colistin-based control strategies. Acta Vet Scand. 2017, 59, 31. [Google Scholar] [CrossRef] [PubMed]
- Luppi, A. Swine enteric colibacillosis: diagnosis, therapy, and antimicrobial resistance. Porcine Health Manag. 2017, 3, 16. [Google Scholar] [CrossRef] [PubMed]
- Vilaró, A.; Novell, E.; Enrique-Tarancón, V.; Baliellas, J.; Vilalta, C.; Martinez, S.; Fraile, L. Antimicrobial Susceptibility Pattern of Porcine Respiratory Bacteria in Spain. Antibiotics 2020, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Vilaró, A.; Novell, E.; Enrique-Tarancon, V.; Baliellas, J.; Migura-García, L.; Fraile, L. Antimicrobial Susceptibility Testing of Porcine Bacterial Pathogens: Investigating the Prospect of Testing a Representative Drug for Each Antimicrobial Family. Antibiotics 2022, 11, 638. [Google Scholar] [CrossRef] [PubMed]
- Vilaró, A.; Novell, E.; Enrique-Tarancón, V.; Baliellas, J.; Allué, E.; Fraile, L. Antimicrobial Stewardship for Respiratory Pathogens in Swine. Antibiotics 2020, 9, 727. [Google Scholar] [CrossRef]
- European Medicines Agency (EMA). ESVAC Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2021; EMA: Amsterdam, The Netherlands, 2021. [Google Scholar]
- World Health Organization (2021) Comprehensive Review of the WHO Global Action Plan on Antimicrobial Resistance. Available online: https://www.who.int/publications/m/item/comprehensive-review-of-the-who-global-action-plan-on-antimicrobial-resistance (accessed on 1 September 2023).
- Mader, R.; Damborg, P.; Amat, J.P.; Bengtsson, B.; Bourély, C.; Broens, E.M.; Busani, L.; Crespo-Robledo, P.; Filippitzi, M.E.; Fitzgerald, W.; et al. EU-JAMRAI. Building the European Antimicrobial Resistance Sur-veillance network in veterinary medicine (EARS-Vet). Euro Surveill. 2021, 26, 2001359. [Google Scholar] [CrossRef]
- Aerts, M.; Faes, C.; Nysen, R. EFSA Scientific report: development of statistical methods for the evaluation of data on antimicrobial resistance in bacterial isolates from animals and food. EFSA J. 2011, 8, 1–77. [Google Scholar] [CrossRef]
- Otto, S.J.G. Antimicrobial resistance of human Campylobacter jejuni infections from Saskatchewan. Population medicine. Doctoral thesis. University of Guelph. 2011. Pp 1-259.
- Bjork, K.E.; Kopral, C.A.; Wagner, B.A.; Dargatz, D.A. Comparison of mixed effects models of antimicrobial resistance metrics of livestock and poultry Salmonella isolates from a national monitoring system. Prev Vet Med. 2015, 122, 265–272. [Google Scholar] [CrossRef]
- Hanon, J.B.; Jaspers, S.; Butaye, P.; Wattiau, P.; Méroc, E.; Aerts, M.; Imberechts, H.; Vermeersch, K.; Van der Stede, Y. A trend analysis of antimicrobial resistance in commensal Escherichia coli from several livestock species in Belgium (2011-2014). Prev Vet Med. 2015, 122, 443–452. [Google Scholar] [CrossRef]
- Zawack, K.; Li, M.; Booth, J.G.; Love, W.; Lanzas, C.; Gröhn, Y.T. Monitoring Antimicrobial Resistance in the Food Supply Chain and Its Implications for FDA Policy Initiatives. Antimicrob Agents Chemother. 2016, 60, 5302–5311. [Google Scholar] [CrossRef]
- Lagrange, J.; Amat, J.P.; Ballesteros, C.; Damborg, P.; Grönthal, T.; Haenni, M.; Jouy, E.; Kaspar, H.; Kenny, K.; Klein, B.; et al. Pilot testing the EARS-Vet surveillance network for antibiotic resistance in bacterial pathogens from animals in the EU/EEA. Front Microbiol. 2023, 14, 1188423. [Google Scholar] [CrossRef] [PubMed]
- Mader, R.; Muñoz Madero, C.; Aasmäe, B.; Bourély, C.; Broens, E.M.; Busani, L.; Callens, B.; Collineau, L.; Crespo-Robledo, P.; Damborg, P.; et al. Review and Analysis of National Monitoring Systems for Antimicrobial Resistance in Animal Bacterial Pathogens in Europe: A Basis for the Development of the European Antimicrobial Resistance Surveillance Network in Veterinary Medicine (EARS-Vet). Front Microbiol. 2022, 13, 838490. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals. 5th ed. CLSI standard VET01. Wayne, PA: Clinical laboratory institute; 2018.
- Clinical and Laboratory Standards Institute (CLSI) Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals. 5th ed. Clinical and Laboratory Standards Institute (CLSI); Wayne, PA, USA: 2021. VET01S.
- EUCAST. Breakpoint tables for interpretation of MICs and zone diameters, version 9.0. 2019.
- Cavaco, L.M.; Frimodt-Møller, N.; Hasman, H.; Guardabassi, L.; Nielsen, L.; Aarestrup, F.M. Prevalence of quinolone resistance mechanisms and associations to minimum inhibitory concentrations in quinolone-resistant Escherichia coli isolated from humans and swine in Denmark. Microb Drug Resist. 2008, 14, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Stegeman, J.A.; Vernooij, J.C.; Khalifa, O.A.; Van den Broek, J.; Mevius, D.J. Establishing the change in antibiotic resistance of Enterococcus faecium strains isolated from Dutch broilers by logistic regression and survival analysis. Prev Vet Med. 2006, 74, 56–66. [Google Scholar] [CrossRef]
- MacKinnon, M.C.; Pearl, D.L.; Carson, C.A.; Parmley, E.J.; McEwen, S.A. A comparison of modelling options to assess annual variation in susceptibility of generic Escherichia coli isolates to ceftiofur, ampicillin and nalidixic acid from retail chicken meat in Canada. Prev Vet Med. 2018, 160, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Teale, C.; Borriello, P. A proposed scheme for the monitoring of antibiotic resistance in veterinary pathogens of food animals in the UK. Vet Rec 2021, 189, e201. [Google Scholar] [CrossRef]
- Aarestrup, F.M.; Hasman, H.; Veldman, K.; Mevius, D. Evaluation of eight different cephalosporins for detection of cephalosporin resistance in Salmonella enterica and Escherichia coli. Microb Drug Resist. 2010, 16, 253–256. [Google Scholar] [CrossRef]
- Durão, P.; Balbontín, R.; Gordo, I. Evolutionary Mechanisms Shaping the Maintenance of Antibiotic Resistance. Trends Microbiol. 2018, 26, 677–691. [Google Scholar] [CrossRef]
- Dohoo, I.; Martin, W.; Stryhn, H. 2014. Veterinary Epidemiology research. VER Inc., Charlottetown, Prince Eduward Island.
Figure 1.
Percentage of Escherichia coli (A), Actinobacillus pleuropneumoniae (B) and Pasteurella multocida (C) isolates belonging to antimicrobial susceptibility category 1 (circle), category 2 (square), category 3 (diamond) and category 4 (triangle) for enrofloxacin being category 1 the most susceptible (lowest MIC values) and category 4 the less susceptible one (highest MIC value).
Figure 1.
Percentage of Escherichia coli (A), Actinobacillus pleuropneumoniae (B) and Pasteurella multocida (C) isolates belonging to antimicrobial susceptibility category 1 (circle), category 2 (square), category 3 (diamond) and category 4 (triangle) for enrofloxacin being category 1 the most susceptible (lowest MIC values) and category 4 the less susceptible one (highest MIC value).
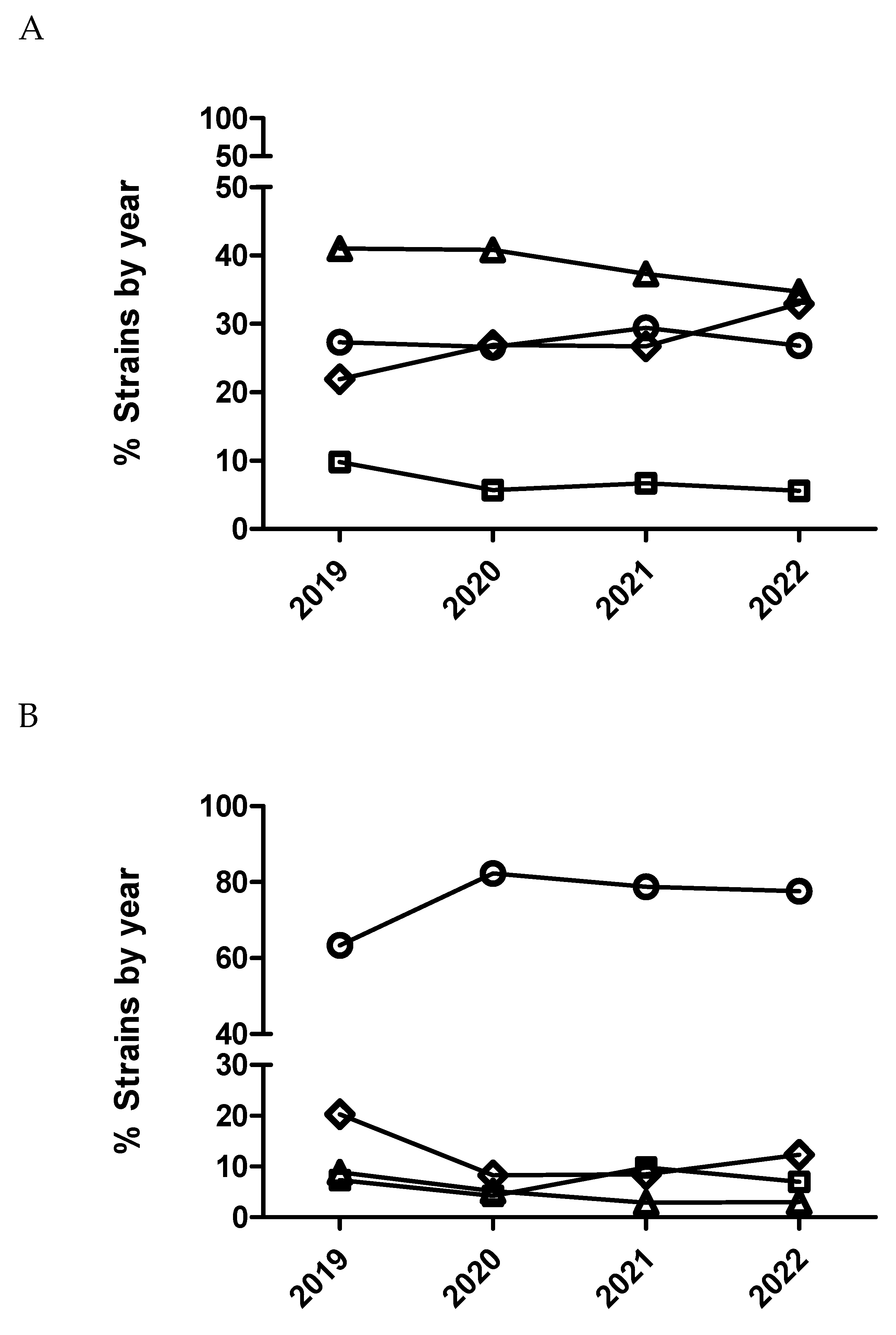
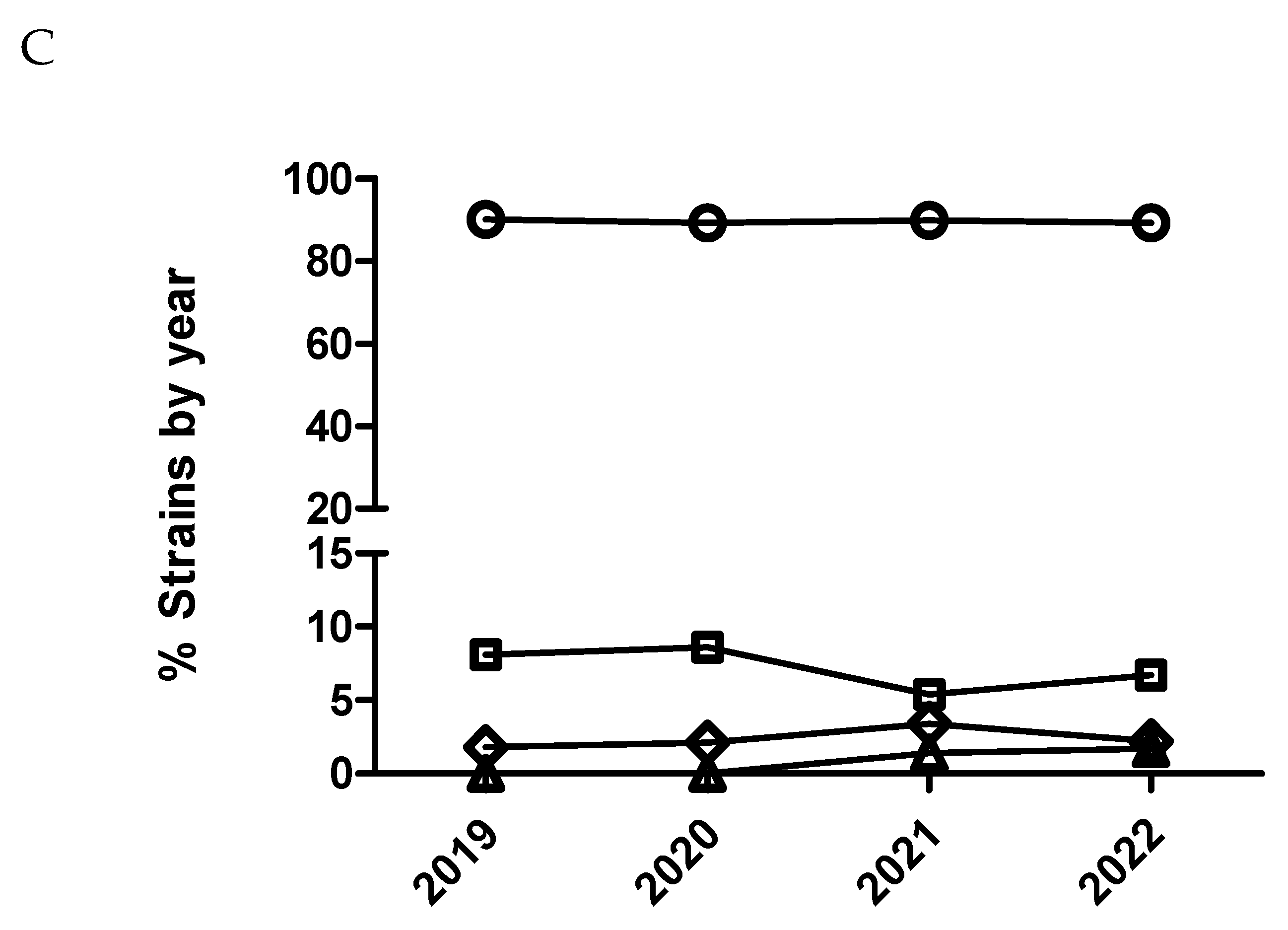
Figure 7.
Percentage of Escherichia coli isolates belonging to antimicrobial susceptibility category 1 (circle), category 2 (square), category 3 (diamond) and category 4 (triangle) for colistin being category 1 the most susceptible (lowest MIC values) and category 4 the less susceptible one (highest MIC value). It has been represented with two different scales (A and B) for the Y axis to have more detail for the categories with low percentage of isolates.
Figure 7.
Percentage of Escherichia coli isolates belonging to antimicrobial susceptibility category 1 (circle), category 2 (square), category 3 (diamond) and category 4 (triangle) for colistin being category 1 the most susceptible (lowest MIC values) and category 4 the less susceptible one (highest MIC value). It has been represented with two different scales (A and B) for the Y axis to have more detail for the categories with low percentage of isolates.
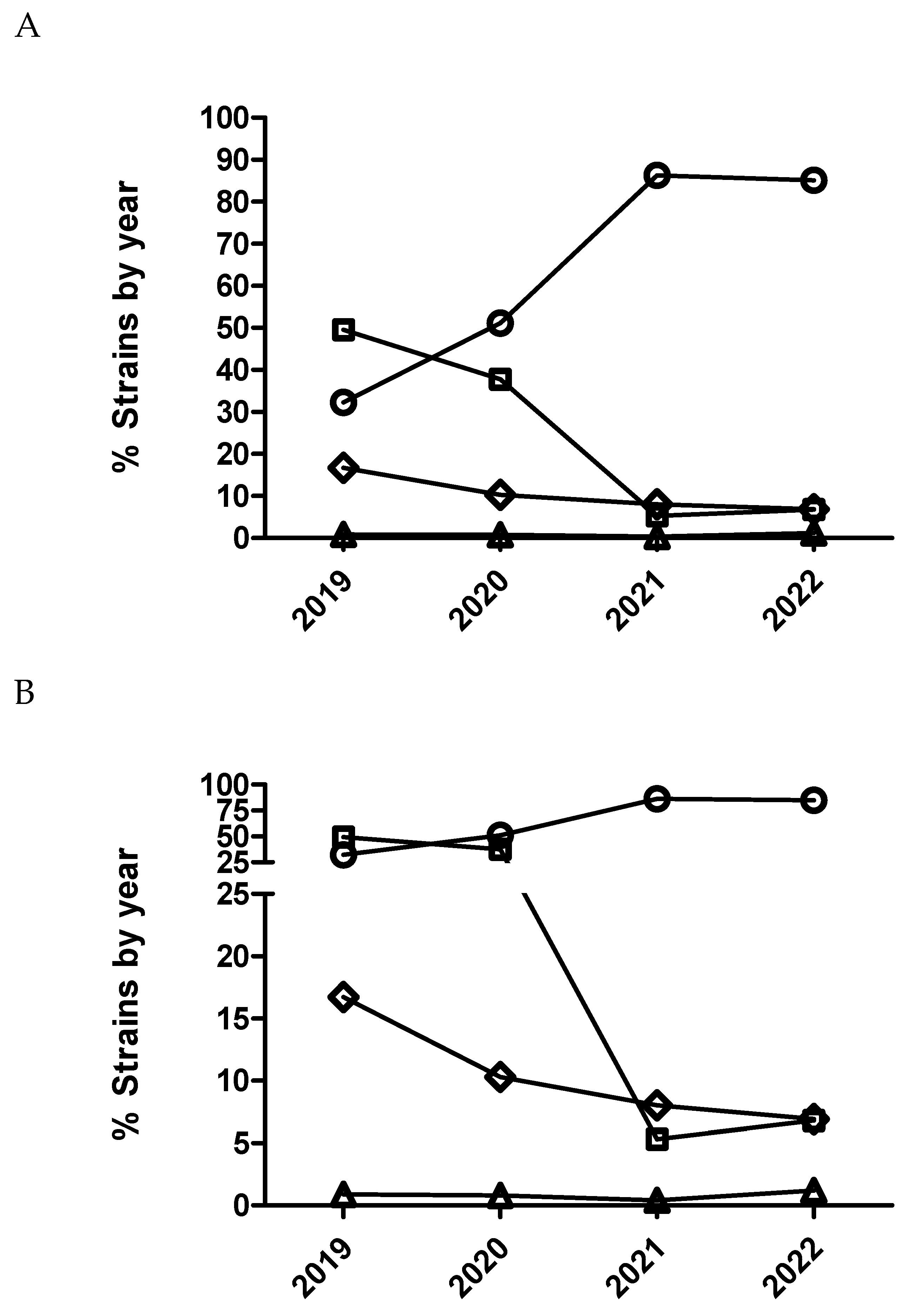
Figure 8.
Percentage of susceptible isolates by year for colistin of Escherichia coli (diamond), using CLSI and EUCAST clinical breakpoints as detailed in the material and method section.
Figure 8.
Percentage of susceptible isolates by year for colistin of Escherichia coli (diamond), using CLSI and EUCAST clinical breakpoints as detailed in the material and method section.
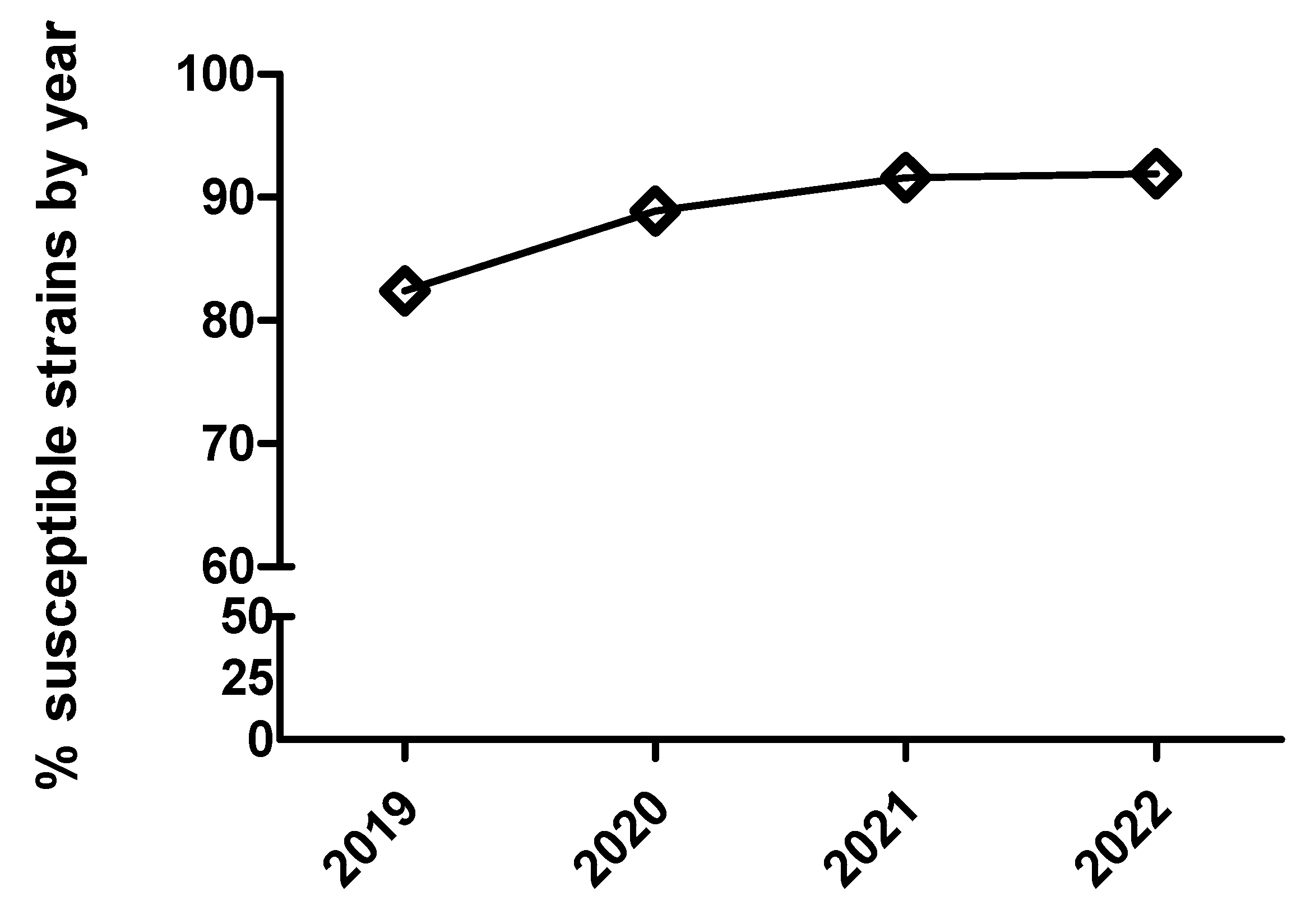
Table 1.
Number of Actinobacillus pleuropneumoniae (APP), Pasteurella multocida, Bordetella bronchiseptica, Escherichia coli and Salmonella spp isolates isolated during the studies period (2019-2022).
Table 1.
Number of Actinobacillus pleuropneumoniae (APP), Pasteurella multocida, Bordetella bronchiseptica, Escherichia coli and Salmonella spp isolates isolated during the studies period (2019-2022).
| Pathogen | 2019 | 2020 | 2021 | 2022 |
|---|---|---|---|---|
| APP | 123 | 195 | 237 | 228 |
| P. multocida | 111 | 100 | 147 | 178 |
| B. bronchiseptica | 24 | 21 | 44 | 53 |
| E. coli | 563 | 512 | 735 | 1,082 |
| Salmonella spp | 18 | 28 | 34 | 52 |
Table 5.
Minimum inhibitory concentration (MIC) distribution values of Escherichia coli to polymyxins (colistin) from 2019 to 2022 in Spain.
Table 5.
Minimum inhibitory concentration (MIC) distribution values of Escherichia coli to polymyxins (colistin) from 2019 to 2022 in Spain.
| Year | MIC range | MIC50 | MIC90 |
|---|---|---|---|
| 2019 | 0.5-32 | 1 | 1 |
| 2020 | 0.5-16 | 0.5 | 1 |
| 2021 | 0.5-16 | 0.5 | 0.5 |
| 2022 | 0.5-16 | 0.5 | 0.5 |
Table 6.
The adjusted odds ratio (95% confidence interval) describing the annual variation in susceptibility of A. pleuropneumoniae isolates to enrofloxacin using the logistic and multinominal regression model. The number of A. pleuropneumoniae isolates by year is detailed in Table 1.
Table 6.
The adjusted odds ratio (95% confidence interval) describing the annual variation in susceptibility of A. pleuropneumoniae isolates to enrofloxacin using the logistic and multinominal regression model. The number of A. pleuropneumoniae isolates by year is detailed in Table 1.
| Logistic analysis (susceptible/resistant) |
Multinominal analysis (MIC outcome categories being compared) | |||
|---|---|---|---|---|
| Predictor variable | NA | 1 vs 2 | 1 vs 3 | 1 vs 4 |
| Year | P=0.0002 | P=0.002 | ||
| 20 vs 19 | 2.7 (1.6-4.8) | NS | 2.1 (1.4-3.1) | 2.3 (1.3-4.1) |
| 21 vs 20 | NS | NS | NS | NS |
| 22 vs 21 | NS | NS | NS | NS |
NS means not significant (p>0.05). NA means not applicable.
Table 11.
The adjusted odds ratio (95% confidence interval) describing the annual variation in susceptibility of E. coli isolates to colistin using the logistic and multinominal regression model. The number of E. coli isolates by year is detailed in Table 1.
Table 11.
The adjusted odds ratio (95% confidence interval) describing the annual variation in susceptibility of E. coli isolates to colistin using the logistic and multinominal regression model. The number of E. coli isolates by year is detailed in Table 1.
| Logistic analysis (susceptible/resistant) |
Multinominal analysis (MIC outcome categories being compared) | |||
|---|---|---|---|---|
| Predictor variable | NA | 1 vs 2 | 1 vs 3 | 1 vs 4 |
| Year | P<0.0001 | P<0.0001 | ||
| 20 vs 19 | 1.7 (1.2-2.4) | 5.5 (4.6-6.9) | 3 (2.4-3.8) | NS |
| 21 vs 20 | NS | 2.7 (2.3-3.3) | NS | NS |
| 22 vs 21 | NS | 0.23 (0.17-0.29) | 0.55 (0.44-0.70) | 0.37 (0.12-0.86) |
NS means not significant (p>0.05). NA means not applicable.
Table 12.
Clinical breakpoints (susceptible/resistant for each isolate) used according to CLSI clinical breakpoints for Actinobacillus pleuropneumoniae (APP), Pasteurella multocida (PM), and Escherichia coli (EC) for quinolones and cephalosporins and EUCAST guidelines for colistin in the case of E. coli, respectively.
Table 12.
Clinical breakpoints (susceptible/resistant for each isolate) used according to CLSI clinical breakpoints for Actinobacillus pleuropneumoniae (APP), Pasteurella multocida (PM), and Escherichia coli (EC) for quinolones and cephalosporins and EUCAST guidelines for colistin in the case of E. coli, respectively.
| Antimicrobial | APP | PM | EC |
| Enrofloxacin | <0.25 | <0.25 | <0.25* |
| Marbofloxacin | <0.25 | <0.25 | <0.25* |
| Ceftiofur | <2 | <2 | <2* |
| Cefquinome | NA | NA | <2* |
| Colistin | NA | NA | <2 |
NA --- Not applicable for this study. *Extrapolated from respiratory to digestive pathogens.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



