
Submitted:
15 September 2023
Posted:
19 September 2023
You are already at the latest version
Abstract
Vanilla pompona is the third most important commercial species of the genus Vanilla, but its morphometric variation is little known. In orchids, infraspecific variation is expressed in the labellum. This study had the objective of analyzing the floral variation of V. pompona collections from localities in Veracruz, Puebla, Jalisco and Oaxaca, Mexico. During the flowering period, we obtained 55 collections of V. pompona flowers, and in the dissected labellum of each flower 54 lines and 7 angles were measured and portrayed with the ImageJ program. With the data obtained, we performed an analysis of variance, a principal components analysis, and a cluster analysis. The results showed significant differences among the collections and localities in the means of the lines of the basal, middle, and apical regions of the labellum. Also, six morphotypes were obtained, and we found that variation was associated with factors such as the environment, geographic barriers, and pollinator pressure. Moreover, we observed variation in flowering dates in one locality of Veracruz. We concluded that there is infraspecific variation within and between the collection localities of V pompona of the Atlantic region (Puebla and Veracruz) and the Pacific region (Jalisco and Oaxaca), Mexico.
Keywords:
orchids
; morphometry
; vanilla
; infraespecific variation
1. Introduction
Vanilla pompona Schiede and Vanilla planifolia Andrews are two relevant species of the genetic reservoir of the genus Vanilla because of their economic importance [1,2,3,4]. V. pompona pertains to the family Orchidaceae, subfamily Vanilloideae, tribe Vanilleae, genus Vanilla, species V. Pompona [5,6], subspecies pompona, grandiflora and pittieri [4]. V. pompona is a robust plant of broad leaves, resistant to diseases, such as anthracnosis (Collectotrichum sp.) and root rot (Fusarium oxysporum Schltdl) [7], and tolerant to xerophyte climates [4,5], with a flowering period from April to June [4]. This species is distributed in several humid subtropical forests (deciduous, evergreen, gallery forests) and in pine and oak forests in warm climates [8,9]. Currently, V. pompona is an endangered species because its fragmented populations are decreasing [9]. However, there are reports that the processed fruits are used in the cosmetic and perfume industries [1] and in the preparation extracts for flavoring [10].
In Mexico, some morphological differences in the flower have been registered in two biological populations of V. pompona. The first population is characterized by flowers with extended segments of large dimensions, located in northern Oaxaca and in Veracruz, on the slope facing the Gulf of Mexico. The second population is characterized by semi-closed flowers of smaller dimensions and is located on the Mexican Pacific coast [5].
Morphological traits of plants are used to analyze variation within and between species, particularly reproductive morphological traits, such as flowers, which are more stable than morphological traits of vegetative organs [11,12]. The V. pompona flower is made up of sepals, petals, labellum, penicillate callus, column, stigma, and one ovary [4]. In orchids, the labellum is a modified petal [13], which owes its existence and aspect to the presence of specific genes involved in the evolutionary floral development of this plant family [14,15]. It functions as a platform for insects to land on and to attract pollinators [16]. The labellum, as a reproductive structure of orchids, is less susceptible to the environment [11], and for this reason, it is used to identify species within this family [4,17] and to analyze infraspecific variation [18,19]. For the genus Vanilla, flowers are used to differentiate leafless species in the Southwestern Indian Ocean region, considering differences in the shape of the labellum [20]. Infraspecific variation has also been evaluated by characterizing the labellum shape of species such as V. planifolia [21] and V. pompona [22]. In the Mesoamerican region, V. planifolia variation is identified by four morphotypes in Oaxaca [23], four morphotypes in the San Luis Potosí Huastec region, Mexico [21], and five morphotypes in the Hidalgo Huastec region, Mexico [24]. For V. pompona, floral morphological variation of the labellum is still being studied; to date, there is only one study that reports four morphotypes in collections from localities in Oaxaca. This variation is explained by the constant selection pressure exerted by pollinators together with the environment [22].
Genetic diversity or genetic variation within species, referring to populations and individuals that comprise them [25], is one of the forms of biodiversity that should be conserved [26]. Knowledge and understanding of the variation within species are vital for the conservation of ecosystems and for the social and cultural wellbeing of man [27,28].
It is not known whether clonal reproduction, disappearance of the habitat where the species is distributed, such as deciduous tropical forests, gallery forests, and tropical pine and oak forests [9], and problems of genetic erosion [29] have caused low genetic variation in V. pompona. Recently, studies have been conducted to determine genetic variation [30,31] and floral morphological variation [21,22,23,26] of the most used Vanilla species, in a search for different stable populations that would increase genetic variation [3,31]. However, for V. pompona, detailed information on genotypes and phenotypes that make up the primary genetic pool and its interaction with the environment is limited. Its cultivation is recent, and thus, both traditional and technological knowledge of its cultivation and its infraspecific variation is also limited.
Plantations with incipient traditional management located close to natural forests maintain structural diversity, endemic species, and regional native species [32,33], contributing to conservation of adjacent ecosystems [27]. Perennial plants, such as V. pompona, can grow in agroforestry systems where the trees that provide support and shade for this species maintain biogeochemical cycles and help to preserve the local landscape and biodiversity [34]. For example, agroforestry systems where V. planifolia is cultivated in northeastern Madagascar, are recognized as important areas for the conservation of birds [35]. For this reason, traditional Vanilla systems could be considered a determining factor in the preservation of this genetic resource [3,12,31].
In the study of orchid variation, it is important to use an integrated approach, with classic taxonomy [20] as well as morphometric analysis of the labellum [19,21,23], relating it with biotic factors (pollinators) and abiotic factors (environment). Because variation of V. pompona on the Mexican Altantic slope or the Pacific Coast is unknown, this study aims to demonstrate that variation of the flower labellum of V. pompona exists. Also, it will contribute knowledge on the floral labellum variation related to V. pompona flowering dates. In orchids, biotic and abiotic factors, such as temperature and light, have been reported to influence the flowering period [36,37]. For this reason, the objective of this study was to determine morphological variation of the V. pompona labellum in collections from localities of Veracruz, Puebla, Oaxaca, and Jalisco. We believe that this knowledge is useful for conservation, management, and use of this species.
2. Materials and methods
2.1. Geographic location
In this study, V. pompona flowers were collected in the states of Jalisco, Oaxaca, Veracruz and Puebla, Mexico (Figure 1, Table 1).
At each of the sampling sites, we obtained climate type, mean annual precipitation (mm), mean annual temperature (°C), soil moisture regime, and ecological zone [38] (Table 2).
Geographic localities of Mexico where Vanilla pompona flowers were collected.
2.2. Study area
Vanilla pompona Schiede flowers were sampled during the flowering period (March-May) in the localities of Cazuelas, Paso de Barriles, La Pasadita, Cabo Corrientes, Hidalgo, Morelos, Primavera, Santa María Chimalapa, Pluma Hidalgo and Reyes de Vallarta. These are the places where we had the opportunity to collect flowers since there is not much information on where specimens of this species are found. Also, the collections we obtained could represent part of the V. pompona variation that exists in Mexico (Table 1)
2.3. Material vegetal
We obtained 55 collections of V. pompona flowers during the flowering period (March-May) in the years 2019 to 2022. Each collection had three to five replications, and we obtained a total of 192 flowers with pollinia and no observable damage (Figure 2). In the locality of Cazuelas, Veracruz, at the suggestion and empirical knowledge of the owner of the lot, the flowers were collected on different dates of the year. The clones were named according to the flowering period: early flowering clone (March-May), continuous flowering clone (March-October), and late flowering clone (June-August).
2.4. Morphological characterization of the labellum
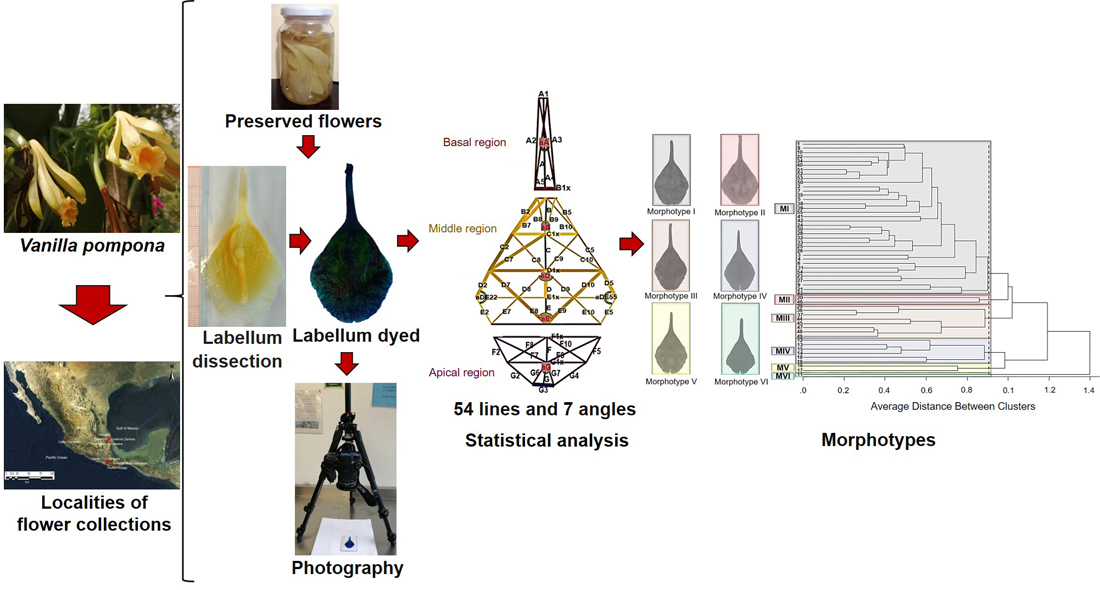
Evaluation of the variation in the flower labellum was based on the technique of Catling [18] and Hernández-Ruíz et al. [22]. First, the flower was dissected, and the labellum was extended on a glass surface. The labellum was then impregnated with methylene blue (0.08%) and photographed with a Sony reflex alpha 65v camera, fixed on a tripod and equipped with a Sony DT macro lens 30 mm F/2.8 SAM. The images were processed with the ImageJ program to generate 54 lines and 7 angles and to measure the lines and angles.
To generate the lines and angles, the labellum was divided into three regions, following the protocol of Hernández-Ruíz et al. [22]. The basal region was formed by lines A, A1, A2, A3, A4, A5, B1x and the angle aA (Figure 3a).
The middle region contained lines B, B2, B5, B7, B8, B9, B10, C, C1x, C2, C5, C7, C8, C9, C10, D, D1x, D2, D5, D7, D8, D9, D10, E, E1x, E2, E5, E7, E8, E9, and E10, while the angles were aB, aD, aDE22, aDE55 and aE (Figure 3b). The apical region was made up of lines F, F1x, F2, F5, F7, F8, F9, F10, G, G1x, G2, G3, G4, G6, G7, and angle aG, and to obtain the length of the labellum, lines A, B, C, D, E, F and G were added (Figure 3c).
2.5. Statistical analyses
With the measurements of the 54 lines and seven angles of the labellum of the 55 collections of V. pompona obtained from different localities, we performed an analysis of variance (ANOVA). The collections were the source of variation. Fifty-five collections (treatments) were analyzed, each with different numbers of replications (labella) under a model equivalent to the completely randomized unbalanced design (PROC GLM; SAS, 2002). With the 61 traits (lines and angles) defined previously for V. pompona [22], a multivariate principal components analysis (PCA) and a cluster analysis were performed [39]. The principal components analysis was conducted based on the matrix of correlations between the selected variables and the cluster analysis with a mean distance between clusters, using the statistical software SAS version 9.0 [40].
3. Results
3.1. Characterization of the labellum
The analysis of variation showed significant differences in the 61 variables evaluated on the V. pompona labellum. For example, with line A (3.65 ***), which is at the base of the labellum and line C1x (4.88 ***), which is on the apical part of the labellum, we found differences in the average measurements of the labellum between collections. The coefficients of variation of the lines were, in general, low; exceptions were lines A1 (10.63) and G3 (12.84%) and angle aG (10.80%). Even so, we detected variation in the measurements of the labella of the collected flowers (Table 3). Therefore, the results indicated that there are differences in the means of the lines of the basal, middle and apical regions of the labellum among collections and localities.
3.2. Distribution of the variation
To identify the variation of the measurements of the 54 lines and 7 angles in the V. pompona flower labellum, we used a principal components analysis. The dispersion of the 55 collections represented by the first three principal components together explained 75% of the accumulated variation. The first principal component (PC1) explained 43% of the general variation and was associated with labellum length (0.187) (Table 4). For this reason, this variable determined part of the morphological variation of the labellum of V. pompona specimens found at the collection sites.
The second principal component (PC2) expressed 20% of the total variation and was associated with the variables aD (0.205), E7 (0.196), E10 (0.196), F1x (0.209), F2 (0.191), F7 (0.211), F9 (0.195), F10 (0.205) and G7 (0.187). The third principal component (PC3) defined 12% of the variation, explained by the variables D (0.264), D2 (0.274), D5 (0.275), D7 (0.208), D8 (0.214), D9 (0.205), D10 (0.217), E2 (0.251), E5 (0.249), and aE (-0.188) (Table 4). Thus, the variables that explain PC2 and PC3 contribute to the shape, width and amplitude of the V. pompona labellum.
In accord with the first three principal components, the spatial distribution of the evaluated collections revealed six groups (six morphotypes) (Figure 4). The distribution of the collections based on principal component 1 (PC1) placed the collections with the longest labella on the positive axis from the center upward (Groups I, II, III). The rest of the groups (Groups IV, V, VI) are on the negative axis, from -5 to -20.
Principal component 2 (PC2) concentrated the collections that have the shortest lines in the apical region of the labellum (E7, E10, F1x, F2, F7, F9, F10 and G7) on the negative axis (Groups IV, V and VI), while collections with longer and wider edges on the lateral lobes and wider labellum were placed in the callus region (aD) on the positive axis (Groups I, II, and III).
Principal component 3 (PC3) was represented by the collections with smaller amplitude (aE) of the middle region of the labellum and short lines in the middle region of the labellum (D, D2, D5, D7, D8, D9, D10, E2 and E5). These collections were placed on the negative axis (Groups IV, V and VI). Likewise, the collections with acute angles of 88-89° in the middle region of the labellum (aE) and some lines of the middle region of the labellum (D, D2, D5, D7, D8, D9, D10, E2 and E5), which define the shape and size of the labellum, were placed on the positive axis (Groups I, II and III).
We identified six morphological groups (morphotypes) in function of the labellum.
Group I included collections 1, 8, 10, 22, 34, 40, 51, 52, 53, 50, 2, 7, 35, 5, 38, 39, 55, 41, 42, 24, 30, 28, 32, 33, 25, 26, 3, 4, 6, 31, 54, 23, 27, 9, 21 and 43. These collections are from the localities of Cazuelas, Papantla; Barriles, Gutiérrez Zamora, state of Veracruz, and from Cabo Corrientes, state of Jalisco. Among these collections are early, continuous, and late flowering clones.
Group II was made up of collections 20 and 46 of the locality Cazuelas and are continuous flowering clones, which begin flowering in March and continue into October.
Group III included collections 29, 36, 37, 44, 45, 47, 48, 49 from La Pasadita, Tihuatlan, Veracruz; Reyes de Vallarta, Tuzamapan de Galeana, Puebla, and Cabo Corrientes, Jalisco. This group also includes collections from Cazuelas land Papantla, Veracruz, that are early-flowering (March-May).
Group IV was represented by collections 12, 13, 14, 15, 18 and 19 from Hidalgo, Morelos, and Primavera, municipality of Santa Cruz Itundujia and Pluma Hidalgo, state of Oaxaca.
Group V included collections 16 and 17 from the municipality of Santa María Chimalapa, Oaxaca.
Group VI was made up of only collection 11 from the municipality of Santa María Chimalapa, Oaxaca.
3.3. Grouping diversity
The cluster analysis, with a distance between clusters of 0.9, confirmed the identification of the six groups defined by the principal components analysis. These groups were denominated morphotypes (Figure 5).
Morphotype I (MI) included 36 collections that were distinguished by an irregular pentagon shape and a rounded shape of the lateral lobes. Total length of the labellum was 76.00 mm and its angles in the middle region were 146° and 146°. The lengths of each region were the following: basal region (B1x) 4.88 mm, middle region (D1x) 33.22 mm, and apical region (F1x) 32.90 mm.
Morphotype II (MII) was represented by collections 20 and 46 from the locality of Cazuelas. This morphotype was characterized by a longer labellum (81.44 mm) and by rounded lateral lobes that significantly differentiate them from the apical lobe of the labellum. The labellum lengths, by region, were basal region (B1x) 4.94 mm, middle region (D1x) 34.50 mm, and apical region (F1x) 36.33 mm. Amplitude of the middle labellum region was 147° and 149°.
Morphotype III (MIII) included the collections 29, 36, 37, 44, 45, 47, 48 and 49. These collections were distinguished by the rhomboid shape of their labellum; length was 80.74 mm and amplitude of the middle region was 149° and 149°. The length of each region of the labellum were the following: basal region (B1x) 4.49 mm, middle region (D1x) 33.72 mm, and apical region (F1x) 29.99 mm.
Morphotype IV (MIV) was represented by collections 12, 13, 14, 15, 18 and 19, which had oval-shaped lateral lobes. This shape was defined by a labellum 68.28 mm long and angles of the middle region of 146° and 142°. They had the following lengths of each region of the labellum: basal region (B1x) 5.31 mm, middle region (D1x) 31.45, and apical region (F1x) 30.85 mm.
Morphotype V (MV) was made up of collections 16 and 17, which had a shape with marked lateral lobes, compared with the lobes of the apical region. Labellum length was 71.34 mm, and the angles of the lateral lobes were 140° and 149°. The lengths of each region were the following: basal region (B1x) 4.44 mm, middle region (D1x) 29.85 mm, and the apical region (F1x) 26.28 mm.
Morphotype VI (MVI) was represented by collection 11, which was differentiated from the other morphotypes by a shorter labellum (60.65 mm) and smaller amplitude in the angles of the lateral lobes (129° and 137°). The different regions of the labellum were defined by the following lengths: basal region (B1x) 4.69 mm, middle region (D1x) 26.47 mm, and apical region (F1x) 22.34 mm.
4. Discussion
4.1. Characterization of the V. pompona labellum
The analysis of variance indicated that there were differences in the means of the lines in the basal, middle, and apical regions of the labellum and of the lengths of the labellum among collections and localities. This agrees with Hernández-Ruiz et al. [22], who report that for V. pompona from Oaxaca, Mexico, the differences were highly significant in 58 of the variables evaluated on the labellum.
The coefficients of variation of the lines and angles evaluated oscillated between 3.29 and 12.84%, percentages that are lower than those found in other studies on orchid variables [21,23]. These results show that the labellum is an important structure of the vanilla flower in the analysis of variation and has been used to analyze infraspecific morphological variation in orchids [19,41]. Also, the labellum has been used to differentiate between subspecies, as reported for Vanilla pompona Schiede subsp. pompona, for which one of the main characteristics that distinguishes this subspecies was the length of the labellum (50 to 80 mm), while the length of Vanilla pompona subsp. grandiflora Lindl. was more than 80 mm [4].
4.2. Distribution of the variation
Dispersion of the labella of the V. pompona collections from the Gulf of Mexico slop and from the Pacific coast was represented by the first three principal components, which together explained 75% of the accumulated variation, a percentage that is lower than the study conducted by Hernández Ruíz et al. [22], who detected 81% of the variation in V. pompona collected only in Oaxaca, suggesting that the variation in floral traits of the first three components of this study is due to the use of collections from localities in different states of Mexico.
The horizontal length of the labellum, the variable that determines the size of the flowers, separated the PC1 groups and explained most of the variation (43%). This contrasts with Hernández-Ruíz et al. [22], who state that, for the populations of V. pompona from Oaxaca, the PC1 is associated with the variation in shape and size of some sections of the middle region of the labellum and with a variable of the apical region of the labellum.
The variables of the apical region of the labellum and one angle of the middle region differentiated the PC2 groups. This was similar to the variables reported that defined the PC2 groups from the V. pompona populations of Oaxaca, the apical region together with the dimension of the basal region [22]. For PC3, the morphological variables that separated the groups were those of the middle region of the labellum, which was likewise one of the variables that influenced the definition of groups of the third component in V. pompona collections from Oaxaca [22].
In our study, the variables that explain PC2 and PC3 determined the shape, size and amplitude of lateral and apical lobes of the V. pompona labellum from localities of Veracruz, Jalisco, Puebla, and Oaxaca. In studies on V. planifolia, with the PCA, it was found that PC2 and PC3 were explained by variables that form part of the labellum shape and size of the middle lobes, aperture, and size of the lateral lobes labellum [23], also in the shape of the labellum in the apical region and amplitude of the lobes [21]. Thus, in the V. pompona collections evaluated in our study, the middle and apical regions and the length the labellum explain the variation. According to studies conducted on variation of the labellum of the genus Vanilla [21,22,23,24], the variables used are suitable since, despite differences in the measures, shapes, and size of the labellum in V. pompona and V. planifolia, these traits allow analysis of the infraspecific variation and the variation between species.
4.3. Morphotype grouping
Variation of the V. pompona labellum grouped in six morphotypes by PCA and cluster analysis indicates that these morphotypes represent the variation that exists in the localities of Veracruz, Jalisco, Puebla, and Oaxaca.
Morphotype I is represented by collections from localities of Veracruz and Jalisco. The collections from Veracruz are influenced by the slope of the Gulf of Mexico on the Atlantic side, while Cabo Corrientes is found on the Pacific coast on the other side of Mexico. However, despite their geographic separation by the Sierra Madre Oriental and the Sierra Madre Occidental [42] both have warm subhumid climate (Table 2) and are located on the same latitude. This could indicate that environmental and geographic characteristics have an influence in the fact that the collections from Veracruz and Jalisco are the same labellum morphotype (Figure 5).
The collections from Cazuelas, Papantla, Veracruz, are morphotype II, which stands out because the labellum (81.44 mm) is longer than that of the other morphotypes. This coincides with Soto-Arenas [5], who reported that V. pompona is made up of two population sets in Mexico, one in Veracruz on the Gulf of Mexico slope, characterized by its flower with extended segments of large dimensions.
Morphotype III is the widest and comprises the collections from Cabo Corrientes, Jalisco, and Papantla, Tihuatlán, Veracruz, and Tuzamapan de Galeana, Puebla. The localities of the collections from Veracruz and Puebla belong to the humid tropical ecological zone, while Cabo Corrientes belongs to the sub-humid tropical ecological zone, but the climate is warm subhumid, like that of Veracruz (Table 2), and allows them to share the same labellum morphotype.
Morphotypes IV, V, and VI of the Oaxaca localities have smaller labella. This coincides with Soto-Arenas [5], who mention that the second V. pompona population is found on the Mexican Pacific coast and is characterized by smaller semi-closed flowers, while Hernández-Ruíz et al. [22] report similar findings in the study of floral morphological variation in the Oaxaca populations, where the morphotype CAZ is found. This morphotype was the smallest labellum.
Morphotypes IV, V, and VI are found on the Mexican Pacific coast, and localities where the flowers were collected are in the Sierra Madre del Sur [42], which facilitates isolation of biological communities and separates ecological zones [43]. Moreover, the collection sites are found in the humid temperate, subhumid, and humid tropical zone (Table 2), which explains the variation found in Oaxaca. Also, a study of the V. pompona labellum from of Oaxaca reports four morphotypes and the variation found was due to the mountain complex belonging to the Sierra Madre del Sur, which separates the ecological zones where the species is found and functions as a geographic barrier [22], while for V. planifolia the study reports that some characteristics of the upper and apical region of the labellum are related to environmental factors [23].
Specifically, morphotype VI from Oaxaca has a horizontally narrower labellum (60.65 mm) and is different from morphotype II from Veracruz, with a horizontal length of 81.44 mm. Morphotype VI is found on the Pacific coast and is in the Sierra Madre del Sur [42], while morphotype II is found on the Gulf of Mexico slope and belongs to the Sierra Madre Oriental [44]. These mountain chains function as geographic barriers and could partly explain the genetic diversity and speciation in orchids [45]. Moreover, it has been documented that the Gulf of Mexico has greater influence in the warm subhumid climate [43] where morphotype II is found, while morphotype VI is found in a warm humid climate (Table 2). It should be considered that the longer labellum of morphotype II could be influenced by the slope of the Gulf of Mexico since it is more humid than the Pacific coast [46]. V. pompona is considered a wild plant because it has not undergone the process of domestication that leads to changes in plant morphology [47]. Another cause of variation in the V. pompona labellum could be the environment, caused by the geographic location different from the collection sites. In Vanilla, the environment where the plants are found plays a highly important role in terms of availability of resources such as water, carbon, and nutrients, and only the apical region of the labellum can be modified by the availability of these resources [23].
Variation into the six labellum morphotypes of V. pompona identified in this study may also have been in part caused by pressure from pollinators or by genetic drift, which has been reported for other orchids [48]. An example of pressure from pollinators is reported for Vanilla palmarum Salzm. ex Lindl., whose flowers have no aroma, and the labellum underwent a modification to guide the beak of the hummingbird Amazilia fimbriata Gmelin to the nectar chamber [49]. V. pompona has a reward strategy for attracting pollinators offering both nectar and fragrance [50]. One of the specific pollinators of this plant is the male of the species Eulaema cingulata Fabricius [51].
The collections that form morphotypes I, II, and III are early-flowering (March-May), continuous-flowering (March-October), and late-flowering (June-August) floral labella from the locality of Cazuelas, Papantla, Veracruz. Studies have reported that V. pompona flowering in Mexico is April to June [4], but in V. pompona populations in Costa Rica and Peru they found two flowering periods, one January to February and another in September [51]. In V. pompona populations in the state of Maranhão, Brazil, flowering occurs in July and August [52], and in the Amazonian wetlands of Madre de Dios, Peru, the V. pompona subsp. grandiflora populations have two flowering periods that can vary depending on their geographic location [53], responding to specific environmental conditions, such as temperature and light during the flowering period [36,37]. It is interesting in Mexico that in a collection locality (established approximately 10 years ago) in Veracruz, specimens of V. pompona can be found with different flowering dates.
This study contributes knowledge on the variation of the V. pompona labellum in the states of Veracruz, Puebla, Jalisco and Oaxaca. The knowledge generated will help producers to better use the plant by selecting which plants to cultivate with different flowering dates. Growers will also be able to decide which specimens to plant according to the morphotype that is found near the area where they live. Moreover, the information contributes to conservation of genetic diversity. However, more studies are needed on genetic variation and with floral ecology of the V. pompona germplasm in Mexico.
5. Conclusions
In localities of the states of Veracruz, Puebla, Oaxaca and Jalisco, Mexico, we obtained six morphotypes of the V. pompona labellum. The variation existing in the labella of this species is possibly associated with pollinator pressure and the presence of geographic barriers, which can generate genetic drift. Because V. pompona is a wild plant, environment plays a highly important role in the variation of the species, and plant diversity is related to phenological-environmental factors since in the locality of Cazuelas, Papantla, Veracruz, plants were found with different flowering dates. Therefore, infraspecific variation exists in the V. pompona germplasm.
Author Contributions
Conceptualization, A.D.-A., C.V.-A. and B.E.H.-C., methodology, A.D.-A., C.V.-A. and B.E.H.-C.; validation, A.D.-A., C.V.-A., B.E.H.-C., A.B.-G.; J.H.-R. and M.D.L.A.-G.; investigation, C.V.-A and A.D.-A.; resources, A.D.-A. and B.E.H.-C.; writing—original draft preparation, C.V.-A. and B.E.H.-C.; writing—review and editing, A.B.-G.; J.H.-R. and M.D.L.A.-G. All authors have read and agreed to the published version of the manuscript.
Funding
This study is product of the doctoral dissertation of the first author, who is grateful to the Consejo Nacional de Humanidades Ciencias y Tecnologías (CONAHCyT) for grant number 594733 in support of her doctoral studies.
Institutional Review Board Statement
“Not applicable” for studies not involving humans or animals.
Data Availability Statement
Not applicable.
Acknowledgments
We thank the people in the collection localities in the states of Puebla, Veracruz, Oaxaca and Jalisco, Mexico, for their support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ehlers, D.; Pfister, M. Compounds of vanillons (Vanilla pompona Schiede). Essent. Oil Res. 1997, 9, 427–431. [Google Scholar] [CrossRef]
- Ranadive, A.S.; Havkinfrenkel, D.; Belanger, F.C. Quality control of vanilla beans and extracts. Handbook of vanilla science and technology 2011, 139–161. [Google Scholar] [CrossRef]
- Salazar-Rojas, V.M.; Herrera-Cabrera, B.E.; Delgado-Alvarado, A.; Soto-Hernández, M.; Castillo-González, F.; Cobos-Peralta, M. Chemotypical variation in Vanilla planifolia Jack. (Orchidaceae) from the Puebla-Veracruz Totonacapan region. Genet. Resour. Crop Evol. 2012, 59, 875–887. [Google Scholar] [CrossRef]
- Soto, M.A.; Dressler, R.L. A revision of the Mexican and Central American species of Vanilla Plumier ex Miller with a characterization of their ITS region of the nuclear ribosomal DNA. Lankesteriana 2010, 9, 285–354. [Google Scholar]
- Soto-Arenas, M. Filogeografía y Recursos Genéticos de las Vainillas de México. Informe Final SNIB-CONABIO Proyecto No. J101. Instituto Chinoín, A.C. Herbario de la Asociación Mexicana de Orquideología, A.C. México. 1999; pp. 1–106.
- Cameron, K.M. Vanilloid Orchids: Systematics and Evolution. In Vanilla. Medicinal and Aromatic Plants-Industrial Profiles; Odoux, E., Grisoni, M., Eds.; CRC Press Taylor and Francis Group: Boca Raton, Florida, 2010. [Google Scholar]
- Hernández-Hernández, J., Lubinsky, P. Vanilla Diseases. In Handbook of Vanilla Science and Technology; En Havkin- Frenkel, D., Belanger, F.C., Eds.; Wiley-Blackwell: 2011; pp. 26–38.
- Soto-Arenas, M.A. Recopilación y Análisis de la Información Existente sobre las Especies Mexicanas del Género Vanilla. Reporte Intermedio. CONACYT. México. 2009; 76 pp.
- Herrera-Cabrera, B.E.; Hernández, M.; Vega, M.; Wegier, A. Vanilla pompona (amended version of 2017 assessment). The IUCN Red List of Threatened Species 2020: 2020. e.T105878897A173977322. 1058. [Google Scholar] [CrossRef]
- Crymes, A.R., Salazer, I.V., Vásquez, J.V., Cubas, J.W.R., Labajos, H.V., Berdak, K.A. Patent No. US20200068817A1. 2020. Available online: https://patents.google.com/patent/US20200068817A1/en.
- Podolsky, R.H.; Holtsford, T.P. Population structure of morphological traits in Clarkia dudleyana. I. Comparison of FST between allozymes and morphological traits. Genetics 1995, 140, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Cabrera, B.E.; Trejo-Miranda, J.; Delgado-Alvarado, A. Conocimiento Tradicional, Predictores Climáticos y Diversidad Genética: Fitoindicadores, observaciones astronómicas y diversidad genética de haba en la agricultura. LAP LAMBERT Academic Publishing. 2010; 64 pp.
- Rudall, P.J.; Bateman, R.M. Roles of synorganisation, zygomorphy and heterotopy in floral evolution: The gynostemium and labellum of orchids and other lilioid monocots. Biol. Rev. 2002, 77, 403–441. [Google Scholar] [CrossRef]
- Pan, Z.J.; Chen, Y.Y.; Du, J.S.; Chen, Y.Y.; Chung, M.C.; Tsai, W.C.; Chen, H.H.; et al. Flower development of Phalaenopsis orchid involves functionally divergent SEPALLATA-like genes. New Phytol. 2014, 202, 1024–1042. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.F.; Hsu, W.H.; Lee, Y.I.; Mao, W.T.; Yang, J.Y.; Li, J.Y.; Yang, C.H. Model for perianth formation in orchids. Nat. Plants. 2015, 1, 1–8. [Google Scholar] [CrossRef]
- Gravendeel, B.; Dirks-Mulder, A. Floral development: Lip formation in orchids unravelled. Nat. plants. 2015, 1, 1–2. [Google Scholar] [CrossRef]
- Davies, K.L.; Stpiczyńska, M. Labellar micromorphology of two euglossine-pollinated orchid genera; Scuticaria Lindl. and Dichaea Lindl. and Dichaea Lindl. Ann. Bot. 2008, 102, 805–824. [Google Scholar] [CrossRef]
- Catling, P.M. Malaxis salazarii, a new species from Mexico and northern Mesoamerica. Orquidea 1990, 12, 93–104. [Google Scholar]
- Salazar-Rojas, V.M.; Herrera-Cabrera, B.E.; Soto-Arenas, M.Á.; Castillo-González, F. Morphological variation in Laelia anceps subsp. dawsonii f. chilapensis Soto-Arenas Orchidaceae in traditional home gardens of Chilapa, Guerrero, Mexico. Genet. Resour. Crop. Evol. 2010, 57, 543–552. [Google Scholar] [CrossRef]
- Andriamihaja, C.F.; Ramarosandratana, A.V.; Grisoni, M.; Jeannoda, V.; Besse, P. The leafless Vanilla species-complex from the South-West Indian Ocean Region: A taxonomic puzzle and a model for orchid evolution and conservation research. Diversity 2020, 12, 1–25. [Google Scholar] [CrossRef]
- Lima-Morales, M.; Herrera-Cabrera, B.E.; Delgado-Alvarado, A. Intraspecific variation of Vanilla planifolia (Orchidaceae) in the Huasteca region, San Luis Potosí, Mexico: Morphometry of floral labellum. Plant Syst. Evol. 2021, 307, 1–11. [Google Scholar] [CrossRef]
- Hernández-Ruíz, J.; Herrera-Cabrera, B.E.; Delgado-Alvarado, A. Variación morfológica del labelo de Vanilla pompona (Orchidaceae) en Oaxaca, México. Rev. Mex. Biodiversidad 2019, 90, 1–9. [Google Scholar] [CrossRef]
- Hernández-Ruíz, J.; Delgado-Alvarado, A.; Salazar-Rojas, V.M.; Herrera-Cabrera, B.E. Morphological variation of the labellum of Vanilla planifolia Andrews (Orchidaceae) in Oaxaca, Mexico. Rev. Fac. Cienc. Agrar. UNCuyo 2020, 52, 160–175. [Google Scholar]
- Maceda, A.; Delgado-Alvarado, A.; Salazar-Rojas, V.M.; Herrera-Cabrera, B.E. Vanilla planifolia Andrews (Orchidaceae): Labellum Variation and Potential Distribution in Hidalgo, Mexico. Diversity 2023, 15, 678. [Google Scholar] [CrossRef]
- Heywood, V.H.; Baste, I. Introducing biodiversity. In. Heywood, V.H and Watson, R.T (Eds). Global Biodiversity Assessment. University Press. Cambridge. 1995; pp 1-19.
- McNeely, J.A.; Miller, K.R.; Reid, W.V.; Mittermeier, R.A.; Werner, T.B. Conserving the world´s biological diversity. IUCN, World Resources Institute, Conservation International, WWF-US, World Bank, Washington. 1990; p. 193.
- Gliessman, S.R. Agroecología: Procesos ecológicos en agricultura sostenible. Catie. Costa Rica. 2002; pp 229-249.
- Piñero, D.; Caballero-Mellado, J.; Cabrera-Toledo, D.; Canteros, C.E.; Casas, A.; Castañeda, A.; Zúñiga, G.; et al. La diversidad genética como instrumento para la conservación y el aprovechamiento de la biodiversidad: Estudios en especies mexicanas, en Capital natural de México. Conocimiento actual de la biodiversidad. Conabio, México. 2008, 1, 437–494.
- Minoo, D.; Jayakumar, V.N.; Veena, S.S.; Vimala, J.; Basha, A.; Saji, K.V.; Peter, K.V. Genetic variations and interrelationships in Vanilla planifolia and few related species as expressed by RAPD polymorphism. Genet. Resour. Crop Evol. 2007, 55, 459–470. [Google Scholar] [CrossRef]
- Bory, S.; Catrice, O.; Brown, S.; Leitch, I.J.; Gigant, R.; Chiroleu, F.; Grisoni, M.; Duval, M.F.; Besse, P. Natural polyploidy in Vanilla planifolia (Orchidaceae). Genome 2008, 51, 816–826. [Google Scholar] [CrossRef]
- Herrera-Cabrera, B.E.; Salazar-Rojas, V.M.; Delgado-Alvarado, A.; Campos-Contreras, J.; Cervantes-Vargas, J. Use and conservation of Vanilla planifolia J. in the Totonacapan region, México. Eur. J. Environ. Sci 2012, 2, 43–50. [Google Scholar] [CrossRef]
- Hending, D.; Andrianiaina, A.; Maxfield, P.; Rakotomalala, Z.; Cotton, S. Floral species richness, structural diversity and conservation value of vanilla agroecosystems in Madagascar. Afr. J. Ecol. 2020, 58, 100–111. [Google Scholar] [CrossRef]
- Espinoza-Pérez, J.; Díaz-Bautista, M.; Barrales-Cureño, H.J.; Herrera-Cabrera, B.E.; Sandoval-Quintero, M.A.; Juárez-Bernabe; Reyes, C. Floristic biodiversity in Vanilla planifolia agroecosystems in the Totonacapan region of Mexico. Biocell 2019, 43, 440–452. [Google Scholar]
- Soto-Arenas, M. La vainilla: Retos y perspectivas de su cultivo. Biodiversitas 2006, 66, 1–9. [Google Scholar]
- Martin, D.A.; Andriafanomezantsoa, R.; Dröge, S.; Osen, K.; Rakotomalala, E.; Wurz, A.; Kreft, H. Bird diversity and endemism along a land-use gradient in Madagascar: The conservation value of vanilla agroforests. BioTROPICA. 2020, 53, 179–190. [Google Scholar] [CrossRef]
- Ordóñez-Blanco, J.C.; Parrado-Rosselli, Á. Relación fenología-clima de cuatro especies de orquídeas en un bosque altoandino de Colombia. Lankesteriana 2017, 17, 1–15. [Google Scholar] [CrossRef]
- Schaik, C.P.; Terborgh, J.W.; Wright, S.J. The phenology of tropical forests: Adaptive significance and consequences for primary consumers. Annu. Rev. Ecol. Syst. 1993, 24, 353–377. [Google Scholar] [CrossRef]
- CONABIO (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad).. Portal de Geoinformación. 2012. (Consultado 04 febrero 2023). Disponible en linea. Available online: http://www.conabio.gob.mx/informacion/gis/.
- Sneath, P.H.A.; Sokal, R.R. Numerical taxonomy. The principles and practices of numerical classification. San Francisco: W. H. Freeman and Co. 1973.
- SAS (Statistical Analysis Systems). SAS/STAT Users guide, version 9. SAS Institute Inc., North Carolina 14. 2002; pp 421.
- Ketjarun, K.; Traiperm, P.; Suddee, S.; Watthana, S.; Gale, S.W. Labellar anatomy of the Nervilia plicata complex (Orchidaceae: Epidendroideae) in tropical Asia. Kew Bull. 2019, 74, 1–13. [Google Scholar] [CrossRef]
- González-Elizondo, M.S.; González-Elizondo, M.; Tena-Flores, J.A.; Ruacho-González, L.; López-Enríquez, I.L. Vegetación de la sierra madre occidental, México: Una síntesis. Acta Bot. Mex. 2012, 100, 351–403. [Google Scholar] [CrossRef]
- Challenger, A. Utilización y conservación de los ecosistemas terrestres de México: Pasado, presente y futuro. (primera edición). Ciudad de México: Conabio/ Instituto de Biología-UNAM/ Agrupación Sierra Madre S.C. 1988; pp. 269–292.
- Suárez-Mota, M.E.; Villaseñor, J.L.; López-Mata, L. Dominios climáticos de la Sierra Madre Oriental y su relación con la diversidad florística. Rev. Mex. biodiversidad 2017, 88, 224–233. [Google Scholar] [CrossRef]
- Han, L.X.; Jin, Y.; Zhang, J.L.; Li, X.L.; Chung, M.Y.; Herrando-Moraira, S.; Kawahara, T.; Yukawa, T.; Chung, S.W.; Chung, J.M.; Kim, Y.D.; López-Pujol, J.; Chung, M.G.; Tian, H.Z. Phylogeography of the endangered orchids Cypripedium japonicum and Cypripedium formosanum in East Asia: Deep divergence at infra-and interspecific levels. TAXON 2022, 71, 733–757. [Google Scholar] [CrossRef]
- Rzedowski, J. Vegetación de México. Editorial Limusa, México. 1978; p. 432.
- Evans, L.T. The Domestication of Crop Plants In: Crop evolution, adaptation and yield, Cambridge University Press. Cambridge. 1996; pp. 62–112.
- Mondragón-Palomino, M.; Theißen, G. Why are orchid flowers so diverse? Reduction of evolutionary constraints by paralogues of class B floral homeotic genes. Ann. Bot. 2009, 104, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Pansarin, E.R.; Ferreira, A.W.C. Evolutionary disruption in the pollination system of Vanilla (Orchidaceae). Plant Biology 2022, 24, 157–167. [Google Scholar] [CrossRef]
- Pansarin, E.R. Non-species-specific pollen transfer and double-reward production in euglossine-pollinated Vanilla. Plant Biology 2023, 25, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Watteyn, C.; Scaccabarozzi, D.; Muys, B.; Van der Schueren, N.; Van Meerbeek, K.; Guizar Amador, M.F.; Karremans, A.P.; et al. Trick or treat? Pollinator attraction in Vanilla pompona (Orchidaceae). BioTROPICA 2021, 54, 268–274. [Google Scholar] [CrossRef]
- Ferreira, A.W.C.; de Oliveira, M.S.; Silva, E.O.; Campos, D.S.; Pansarin, E.R.; Guarçoni, E.A.E. Vanilla bahiana Hoehne and Vanilla pompona Schiede (Orchidaceae, Vanilloideae): Two new records from Maranhão state, Brazil. Check List. 2017, 13, 1131. [Google Scholar] [CrossRef]
- Householder, E.; Janovec, J.; Mozambite, A.B.; Maceda, J.H.; Wells, J.; Valega, R.; Christenson, E.; et al. Diversity, natural history, and conservation of Vanilla (Orchidaceae) in Amazonian wetlands of Madre de Dios, Peru. J. Bot. Res. Inst. Texas. 2010, 4, 227–243. [Google Scholar]
Figure 1.
Geographical location of the collection sites of Vanilla pompona flowers in Mexico. The red dots show the collection sites: Cabo Corrientes (Jalisco), Santa Cruz Itundujia, Pluma Hidalgo and Santa María Chimalapa, (Oaxaca), Tuzamapan de Galeana (Puebla), Papantla, Tihuatlán and Gutiérrez Zamora (Veracruz). .
Figure 1.
Geographical location of the collection sites of Vanilla pompona flowers in Mexico. The red dots show the collection sites: Cabo Corrientes (Jalisco), Santa Cruz Itundujia, Pluma Hidalgo and Santa María Chimalapa, (Oaxaca), Tuzamapan de Galeana (Puebla), Papantla, Tihuatlán and Gutiérrez Zamora (Veracruz). .

Figure 2.
Phenological stages of Vanilla pompona; flowering in April and mature fruits in December.

Figure 3.
Vanilla pompona flower, labellum separated and dyed, fragmentation of the labellum into a. basal region, b. middle region, c. apical region.
Figure 3.
Vanilla pompona flower, labellum separated and dyed, fragmentation of the labellum into a. basal region, b. middle region, c. apical region.
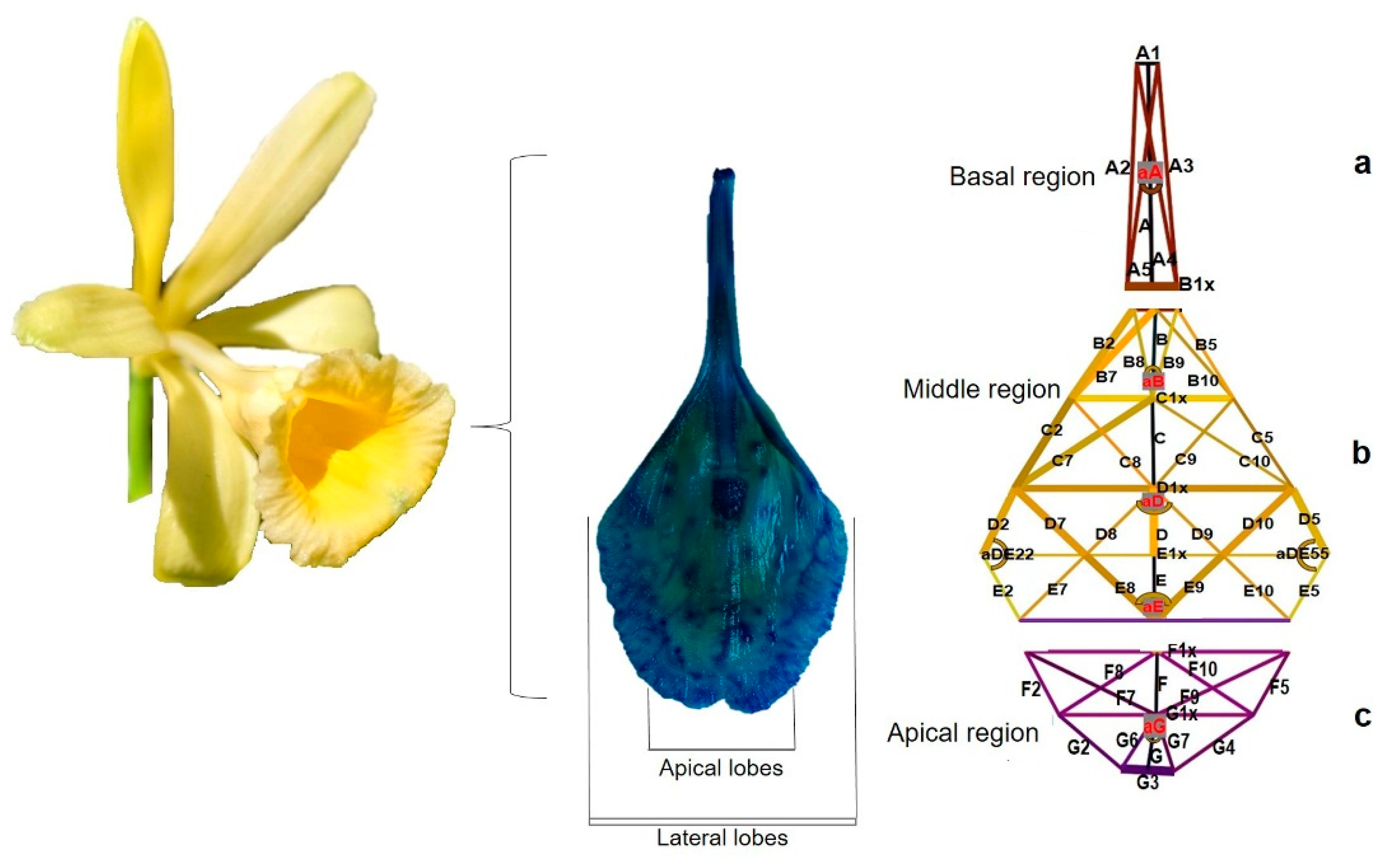
Figure 4.
(A) Variables that define each of the first three principal components. (B) Dispersion of 55 collections of the flower labellum of Vanilla pompona based on the first three principal components, from localities of Veracruz, Jalisco, Puebla, and Oaxaca, Mexico.
Figure 4.
(A) Variables that define each of the first three principal components. (B) Dispersion of 55 collections of the flower labellum of Vanilla pompona based on the first three principal components, from localities of Veracruz, Jalisco, Puebla, and Oaxaca, Mexico.
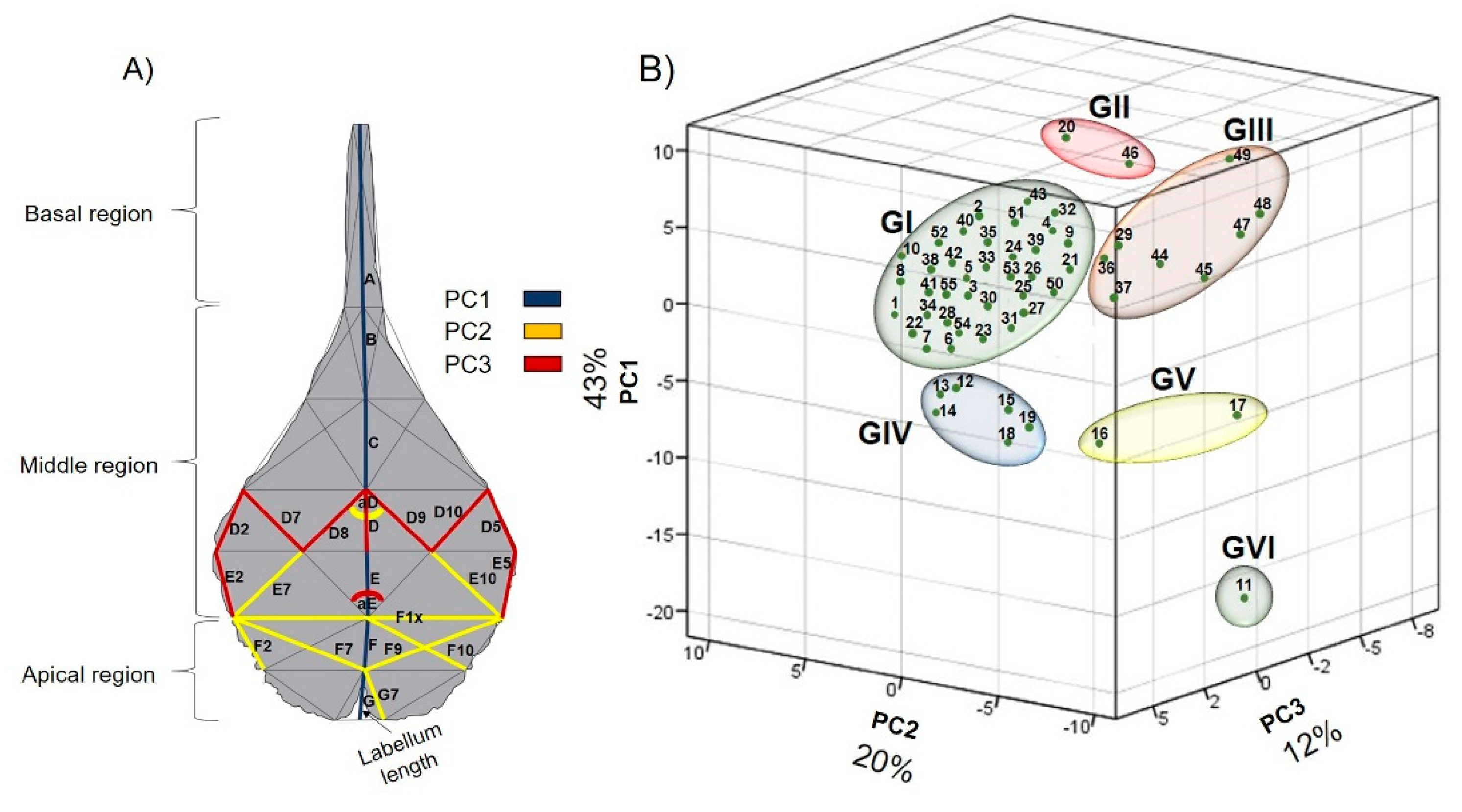
Figure 5.
Dendrogram of labellum morphotypes of Vanilla pompona from localities of Veracruz, Jalisco, Puebla and Oaxaca, Mexico. Based on the average of 61 variables and on grouping by similarity distance.
Figure 5.
Dendrogram of labellum morphotypes of Vanilla pompona from localities of Veracruz, Jalisco, Puebla and Oaxaca, Mexico. Based on the average of 61 variables and on grouping by similarity distance.


Table 1.
Localities where Vanilla pompona were sampled in several states of Mexico.
| State | Municipality | Locality | Altitude (m) |
|---|---|---|---|
| Veracruz | Papantla | Cazuelas | 69 |
| Gutiérrez Zamora | Paso de Barriles | 29 | |
| Tihuatlán | La Pasadita | 159 | |
| Puebla | Tuzamapan de Galeana | Reyes de Vallarta | 365 |
| Jalisco | Cabo Corrientes | Cabo Corrientes | 379 |
| Oaxaca | Sta. Cruz Itundujia | Hidalgo | 1075 |
| Morelos | 1668 | ||
| Primavera | 651 | ||
| Sta. Ma. Chimalapa | Sta. Ma. Chimalapa | 362 | |
| Pluma Hidalgo | Pluma Hidalgo | 1010 |
Table 2.
Environmental variables of the Vanilla pompona flower collection localities.
| Locality | Climate | Mean annual precipitation (mm) | Mean annual temperature (°C) | Soil moisture regime | Ecological zone |
|---|---|---|---|---|---|
| Cazuelas | Warm subhumid | 800-1200 | >22 | Ustic | Humid tropical |
| Paso de Barriles |
Warm subhumid | 1500-2000 | >22 | Udic type II | Humid tropical |
| La Pasadita | Warm subhumid | 1200-1500 | >22 | Ustic | Humid tropical |
| Reyes de Vallarta |
Warm humid | 2500-4000 | >22 | Udic type I | Humid tropical |
| Cabo Corrientes |
Warm subhumid | 1500-2000 | >22 | Xeric | Subhumid tropical |
| Hidalgo | Semi-warm subhumid | 2000-2500 | >22 | Xeric | Subhumid temperate |
| Morelos | Semi-warm subhumid | 2000-2500 | >18 | Xeric | Humid temperate |
| Primavera | Semi-warm subhumid | 2000-2500 | >22 | Xeric | Subhumid temperate |
| Sta. Ma. Chimalapa |
Warm humid | 1500-2000 | >22 | Ustic | Humid tropical |
| Pluma Hidalgo |
Warm subhumid | 1500-2000 | >22 | Ustic | Humid temperate |
Udic type I = 330 to 365 days of rain; Udic type II = 270 to 330 days of rain; Ustic = 180 t0 270 days of rain; Xeric = 90 to 180 days of rain [38].
Table 3.
Coefficients of variation and mean squares of the 61 variables of the Vanilla pompona labella analyzed.
Table 3.
Coefficients of variation and mean squares of the 61 variables of the Vanilla pompona labella analyzed.
| Variable | Mean (mm) | Coefficient of variation | Mean squares | ||||
|---|---|---|---|---|---|---|---|
| Error | Locality | Collection | |||||
| I. Labellum basal region | |||||||
| A | 23.33 | 3.80 | 0.79 | 34.08 | *** | 3.64 | *** |
| A1 | 2.18 | 10.63 | 0.05 | 1.53 | *** | 0.30 | *** |
| A2 | 23.37 | 3.85 | 0.81 | 33.58 | *** | 3.64 | *** |
| A3 | 23.38 | 3.82 | 0.80 | 33.17 | *** | 3.57 | *** |
| A4 | 23.59 | 4.15 | 0.96 | 34.14 | *** | 3.61 | *** |
| A5 | 23.57 | 4.13 | 0.95 | 34.86 | *** | 3.73 | *** |
| aA | 17.17 | 8.19 | 1.98 | 50.91 | *** | 5.15 | *** |
| B1x | 4.87 | 8.03 | 0.15 | 3.09 | *** | 0.51 | *** |
| II. Labellum middle region | |||||||
| B | 11.63 | 4.75 | 0.31 | 8.64 | *** | 0.93 | *** |
| B2 | 13.25 | 4.72 | 0.39 | 9.58 | *** | 1.14 | *** |
| B5 | 13.40 | 4.72 | 0.40 | 8.06 | *** | 1.12 | *** |
| B7 | 14.57 | 4.58 | 0.45 | 8.69 | *** | 1.15 | *** |
| B8 | 11.92 | 4.54 | 0.29 | 7.36 | *** | 0.83 | *** |
| B9 | 11.87 | 4.57 | 0.29 | 8.09 | *** | 0.97 | *** |
| B10 | 14.76 | 4.56 | 0.45 | 6.77 | *** | 1.16 | *** |
| aB | 23.67 | 8.16 | 3.73 | 153.82 | *** | 16.61 | *** |
| C | 11.62 | 4.78 | 0.31 | 8.65 | *** | 0.93 | *** |
| C1x | 17.78 | 7.07 | 1.58 | 18.58 | *** | 4.88 | *** |
| C2 | 13.86 | 5.38 | 0.56 | 13.41 | *** | 1.57 | *** |
| C5 | 13.95 | 5.42 | 0.57 | 9.71 | *** | 1.96 | *** |
| C7 | 20.08 | 5.26 | 1.11 | 20.64 | *** | 2.26 | ** |
| C8 | 14.69 | 4.65 | 0.47 | 5.90 | *** | 1.02 | ** |
| C9 | 14.62 | 4.51 | 0.44 | 9.45 | *** | 1.17 | *** |
| C10 | 20.23 | 5.00 | 1.02 | 14.39 | *** | 2.97 | *** |
| D | 8.35 | 7.37 | 0.38 | 5.29 | *** | 1.57 | *** |
| D1x | 32.90 | 5.95 | 3.84 | 57.94 | *** | 9.48 | *** |
| D2 | 8.74 | 6.87 | 0.36 | 4.43 | *** | 1.25 | *** |
| D5 | 8.77 | 6.74 | 0.35 | 4.31 | *** | 1.28 | *** |
| D7 | 11.84 | 7.37 | 0.76 | 11.70 | *** | 2.37 | *** |
| D8 | 11.61 | 5.66 | 0.43 | 6.49 | *** | 0.93 | ** |
| D9 | 11.64 | 5.25 | 0.37 | 6.61 | *** | 0.84 | ** |
| D10 | 11.84 | 7.27 | 0.74 | 11.37 | *** | 2.53 | *** |
| aD | 88.25 | 4.53 | 16.00 | 230.14 | *** | 102.01 | *** |
| aDE22 | 146.00 | 4.32 | 39.71 | 321.71 | *** | 118.57 | *** |
| aDE55 | 145.98 | 4.47 | 42.55 | 348.61 | *** | 120.61 | *** |
| E | 8.12 | 6.39 | 0.27 | 4.75 | *** | 0.78 | *** |
| E1x | 37.45 | 4.67 | 3.05 | 78.02 | *** | 6.58 | ** |
| E2 | 8.65 | 6.33 | 0.30 | 3.59 | *** | 0.92 | *** |
| E5 | 8.63 | 6.37 | 0.30 | 4.25 | *** | 1.03 | *** |
| E7 | 11.37 | 5.98 | 0.46 | 14.08 | *** | 1.49 | *** |
| E8 | 11.47 | 4.53 | 0.27 | 7.35 | *** | 0.51 | ** |
| E9 | 11.43 | 4.65 | 0.28 | 7.81 | *** | 0.54 | ** |
| E10 | 11.41 | 6.32 | 0.52 | 13.18 | *** | 1.48 | *** |
| aE | 89.69 | 4.25 | 14.55 | 76.97 | *** | 62.19 | *** |
| III. Labellum apical region | |||||||
| F | 6.22 | 7.45 | 0.22 | 5.62 | *** | 0.70 | *** |
| F1x | 32.02 | 5.06 | 2.62 | 96.25 | *** | 11.02 | *** |
| F2 | 7.25 | 7.82 | 0.32 | 5.76 | *** | 0.94 | *** |
| F5 | 7.20 | 8.11 | 0.34 | 6.49 | *** | 0.88 | *** |
| F7 | 17.20 | 5.64 | 0.94 | 28.32 | *** | 3.39 | *** |
| F8 | 13.82 | 6.07 | 0.70 | 21.96 | *** | 2.64 | *** |
| F9 | 13.92 | 6.27 | 0.76 | 19.98 | *** | 3.15 | *** |
| F10 | 17.13 | 5.33 | 0.83 | 28.96 | *** | 2.92 | *** |
| G | 6.25 | 9.31 | 0.34 | 5.55 | *** | 1.23 | *** |
| G1x | 24.77 | 6.14 | 2.31 | 65.83 | *** | 10.69 | *** |
| G2 | 11.30 | 9.46 | 1.14 | 17.49 | *** | 4.06 | *** |
| G3 | 6.16 | 12.84 | 0.63 | 22.79 | *** | 3.90 | *** |
| G4 | 11.20 | 9.40 | 1.11 | 26.84 | *** | 4.78 | *** |
| G6 | 6.92 | 9.75 | 0.46 | 7.23 | *** | 1.53 | *** |
| G7 | 6.98 | 9.60 | 0.45 | 5.43 | *** | 1.19 | *** |
| aG | 51.99 | 10.80 | 31.55 | 1168.06 | *** | 221.56 | *** |
| Labellum length | 75.47 | 3.29 | 6.17 | 286.36 | *** | 21.27 | *** |
***: p ≤ 0.0001 highly significant, **: p≤0.005 moderately significant.
Table 4.
Eigenvalues, eigenvectors, and accumulated proportion of the variation explained by each variable in the first three dimensions of the characterization of 55 collections of Vanilla pompona.
Table 4.
Eigenvalues, eigenvectors, and accumulated proportion of the variation explained by each variable in the first three dimensions of the characterization of 55 collections of Vanilla pompona.
| Variable | Principal component (PC) | Variable | principal Component (PC) | |||||
|---|---|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | |||
| I. Labellum basal region | D9 | 0.150 | -0.047 | 0.205 | ||||
| A | 0.156 | -0.123 | -0.133 | D10 | 0.097 | -0.135 | 0.217 | |
| A1 | 0.101 | -0.055 | -0.026 | aD | 0.011 | 0.205 | -0.232 | |
| A2 | 0.156 | -0.123 | -0.134 | aDE22 | 0.047 | -0.055 | -0.052 | |
| A3 | 0.156 | -0.122 | -0.134 | aDE55 | 0.034 | -0.092 | -0.075 | |
| A4 | 0.159 | -0.121 | -0.130 | E | 0.129 | 0.092 | 0.180 | |
| A5 | 0.158 | -0.122 | -0.130 | E1x | 0.139 | 0.137 | 0.084 | |
| aA | -0.062 | 0.083 | 0.107 | E2 | 0.117 | 0.025 | 0.251 | |
| B1x | -0.025 | 0.073 | 0.069 | E5 | 0.117 | 0.038 | 0.249 | |
| II. Labellum middle region | E7 | 0.125 | 0.196 | 0.029 | ||||
| B | 0.157 | -0.118 | -0.133 | E8 | 0.161 | 0.124 | 0.115 | |
| B2 | 0.168 | -0.116 | -0.065 | E9 | 0.159 | 0.125 | 0.123 | |
| B5 | 0.158 | -0.113 | -0.066 | E10 | 0.126 | 0.196 | 0.021 | |
| B7 | 0.165 | -0.101 | -0.033 | aE | -0.006 | 0.017 | -0.188 | |
| B8 | 0.156 | -0.119 | -0.132 | III. Labellum apical region | ||||
| B9 | 0.159 | -0.115 | -0.128 | F | 0.121 | 0.182 | 0.047 | |
| B10 | 0.151 | -0.099 | -0.026 | F1x | 0.125 | 0.209 | -0.058 | |
| aB | -0.093 | 0.104 | 0.110 | F2 | 0.110 | 0.191 | 0.065 | |
| C | 0.157 | -0.118 | -0.133 | F5 | 0.109 | 0.175 | 0.046 | |
| C1x | 0.064 | -0.013 | 0.146 | F7 | 0.125 | 0.211 | -0.036 | |
| C2 | 0.166 | -0.098 | -0.088 | F8 | 0.132 | 0.186 | -0.067 | |
| C5 | 0.153 | -0.080 | -0.091 | F9 | 0.125 | 0.195 | -0.050 | |
| C7 | 0.170 | -0.077 | 0.020 | F10 | 0.129 | 0.205 | -0.050 | |
| C8 | 0.160 | -0.096 | -0.033 | G | 0.113 | 0.176 | -0.087 | |
| C9 | 0.161 | -0.105 | -0.029 | G1x | 0.126 | 0.185 | -0.084 | |
| C10 | 0.149 | -0.054 | 0.020 | G2 | 0.092 | 0.148 | -0.066 | |
| D | 0.095 | -0.137 | 0.264 | G3 | 0.034 | 0.111 | 0.040 | |
| D1x | 0.132 | -0.025 | 0.092 | G4 | 0.122 | 0.119 | -0.097 | |
| D2 | 0.110 | -0.084 | 0.274 | G6 | 0.104 | 0.181 | -0.064 | |
| D5 | 0.109 | -0.077 | 0.275 | G7 | 0.105 | 0.187 | -0.056 | |
| D7 | 0.101 | -0.143 | 0.208 | aG | -0.027 | 0.027 | 0.072 | |
| D8 | 0.148 | -0.042 | 0.214 | Long_lab | 0.187 | -0.044 | -0.029 | |
| Eigenvalues | 26.126 | 12.174 | 7.401 | |||||
| Proportion | 0.43 | 0.20 | 0.12 | |||||
| Accumulated | 0.43 | 0.63 | 0.75 | |||||
Values in bold type indicate variables that have the most influence in each principal component.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



