
Submitted:
12 September 2023
Posted:
14 September 2023
You are already at the latest version
Abstract
Abstract: A multitude of biotic and abiotic stress factors do harm to plants by bringing about diseases and inhibiting normal growth and development. As an omnipotent signaling molecule, salicylic acid (SA) plays crucial roles in plants' tolerance responses to both biotic and abiotic stresses, thereby maintaining plants’ normal growth and improving yields under stresses. In view of this, this paper mainly discusses the role of SA in both biotic stresses, and abiotic stresses of plants. SA regulates the expression of genes involved in defense signaling pathways, thus enhancing plants’ immunity. In addition, SA mitigates the negative effects of abiotic stresses, and acts as a signaling molecule to induce the expression of stress-responsive genes and the synthesis of stress-related proteins. In addition, SA also improves certain yield-related photosynthetic indexes, thereby enhancing crop yield under stresses. On the other hand, SA acts with other signaling molecules in regulating plants’ growth and improving tolerances under stresses. This paper reviews recent advances in SA’s roles in plant stress tolerances, so as to provide theoretical references for further studies concerning the decryption of molecular mechanisms for SA’s roles and the improvement of crop management under stresses.
Keywords:
salicylic acid
; biotic stresses
; abiotic stresses
; tolerance
; immunity
; yield
1. Introduction
A wide range of stress factors in the plant growth environment, such as pathogen infection, drought, salt stress, etc., have a significant impact on plant survival and development [1,2,3,4]. In order to adapt to these stresses, plants have evolved a series of tolerance mechanisms involving the regulation of multiple cell signaling pathways [4,5,6,7]. Among them, salicylic acid plays an important role in plant stress tolerance [8,9,10]. This article will discuss the research progress of salicylic acid’s involvement in the cell signaling pathways of plants in combating stress.
It is generally held that salicylic acid (SA) plays vital roles in plant stress tolerance. SA’s main roles in plant tolerance to stresses can be summarized as following: the activation of plant defense mechanisms [2,11,12,13,14], the improvement of photosynthesis[15,16,17,18,19,20,21,22] , the regulation of plant normal growth and development[8,19,23,24,25,26,27,28,29,30], and the enhancement of plant tolerance to drought[9,11,31,32,33,34,35,36], cold[37,38,39], heat[36,39,40,41,42], salt[43,44,45,46,47,48] and heavy metal stresses[49,50,51,52].
It was widely reported and reviewed that salicylic acid (SA) directly participates in the activation of plant defense mechanisms [2,11,12,13,14]. During its activation of relevant plant defense mechanism, SA can induce the biosynthesis of such stress-resistant molecules as (including SOD, POD and CAT) [11,20,53,54,55,56,57,58,59], active anti-pathogenic substances[1,60,61,62,63,64,65], and anti-fungal substances[66], thereby enhancing plants’ resistances or tolerances to biotic stresses caused by pathogens, fungi and pests.
It was also generally reported and reviewed that salicylic acid (SA) signaling pathways act to increase chlorophyll contents in plant leaves and help to improve photosynthetic efficiencies [15,16,17] , thereby significantly enhancing crop yield and other yield-related physiological indexes under various stresses[8,67,68,69].
SA is involved in the regulation of normal growth and development of plants [8,19,23,24,25,26,27,28,29,30]. For example, SA can promote the differentiation of blossom buds, thus directly enhancing flowering [70,71]. On the other hand, SA can also improve number and quality of flowers. In addition, SA also exerts modulatory effects on seedlings by regulating seed dormancy and growth [72], thereby making them more vigorous or less susceptible to stresses [11,31,52,73,74,75,76]. New research has shown that SA plays multiple roles in enhancing plant stress resistances [32,38,43,73,77]. Such roles of SA can be categorized into the following aspects: role of in plant immune response[13,25,78,79,80], role of salicylic acid in plant antioxidative defense [11,20,53,54,55,56,57,58,59], role of salicylic acid in plant salt tolerance [43,44,45,47,48,81,82,83,84], role of salicylic acid in plant drought resistance[2,11,32,33,34,35,36,71,85,86,87].
A wide range of studies have found that salicylic acid is an important signaling molecule in the immune response of plants to biotic and abiotic stress [88]. It can activate the expression of a wide range of disease resistance genes[89], regulate cell wall strengthening [90], and promote hormone production to enhance plant resistance to diseases [5,37,38,49,74,91,92,93]. In addition, SA can enhance plant resistance to oxidative stress. For example, research has also shown that salicylic acid can activate the expression of antioxidative enzymes, thereby enhancing the plant's ability to eliminate reactive oxygen species and reduce cellular oxidative damage [94].
SA is an essential signaling molecule that triggers responses to a range of abiotic stressors, including cold, heat, drought, salt, heavy metals, and so on, and it also participates in developmental signaling pathways essential for normal plant growth, such as flowering and senescence [95,96].
Recent studies have found that salicylic acid can enhance plant tolerance to salt stress[43,47,82,84,97]. Salicylic acid can regulate ion balance [83], maintain cell membrane integrity and stability, and modulate the antioxidative system to enhance the plant's response to salt stress. In addition, other relevant studies have also demonstrated the important regulatory role of salicylic acid in plant drought resistance [11,32,33,71,87,98]. Therefore, salicylic acid can enhance plant drought resistance by regulating root growth, improving leaf water use efficiency, stabilizing cell membranes, and modulating the antioxidative enzyme system.
In a word, salicylic acid, as an important plant signaling molecule, plays a crucial regulatory role in plant stress resistance. As further research into the molecular mechanisms of salicylic acid continues, we will continue to gain deeper understandings of plant stress tolerances. Overall, this paper highlights the significance of SA in enhancing plant resilience against both abiotic and biotic stresses, thereby promoting their survival and productivity in challenging environmental conditions (Figure 1). Further studies in this field can contribute to the development of effective strategies for crop improvement and stress management.
2. Biosynthetic pathway of salicylic acid in plants
Two distinct pathways are involved in the biosynthesis of salicylic acid (SA) in plants: the isochorismate synthase (ICS) pathway and the phenylalanine ammonia-lyase (PAL) pathway [25,78,99,100,101,102]. The ICS pathway was primarily elucidated using Arabidopsis mutants that were deficient in pathogen-induced SA biosynthesis, while the PAL pathway has been found to contribute to SA biosynthesis in tobacco [78].
Research has proposed and reviewed that the levels of SA in plants are regulated by both positive and negative feedback. Under positive feedback regulation, genes involved in SA biosynthesis (ICS1/EDS5/PBS3) and N-hydroxypipecolic acid (NHP) biosynthesis (ALD1/SARD4/FMO1) are coordinately regulated by the immune transcription factors SARD1 and CBP60g. Activation of these factors leads to increased SA and NHP biosynthesis. Upstream of SARD1 and CBP60g, TGA1/TGA4 and GTL1 act as positive regulators, while WRKY70 and CAMTA1/2/3 act as negative regulators of gene transcription related to SA and NHP biosynthesis [78,79]. Additionally, calcium ions (Ca2+) may play a dual role in regulating SA and NHP biosynthesis. They positively modulate the activities of calcium-dependent protein kinases (CPKs) and CBP60g, while negatively regulating the activities of CBP60a and CAMTA1/2/3. However, the exact mechanisms in this process are not yet fully understood [78].
3. The role of salicylic acid in plants’ tolerance responses to biotic stresses
Plants, including crops, are constantly exposed to various biotic stresses [103], such as pathogens (e.g., bacteria, fungi, oomycetes, nematodes), arthropods (particularly insects), birds, mammals, and competitive plants (especially weeds) [104].
In recent decade, especially since 2021, numerous peer-reviewed review and research articles concerning plants’ tolerance responses to biotic stresses have been published, and some articles published from 2021 to 2023 and selected from four database sites, including Sciencedirect, Web of Science, Scopus, and MDPI, are listed in Table 1. From Table 1, it can be seen that these selected articles mainly deal with such biotic stress factors as bacteria, fungi, and insects.
It was reported that SA has the ability to activate various antioxidant enzymes, PR genes, SAR pathways, and even trigger programmed cell death (PCD) [3].
It is generally accepted that SA-dependent signaling processes are involved in systemic acquired resistance (SAR) in plants, leading to the synthesis of pathogenesis-related (PR) proteins in response to pathogen infections. It was also reviewed that SA acts as a vital endogenous signal for SAR, contributing to the activation of defense responses against biotrophic pathogens and enhancing plant resistance to multiple pathogens [4]. On the other hand, it was also considered that SA is involved in the activation of multiple pathways, including MAPK, CDPK, and other protein kinases, thus promoting the biosynthesis of secondary metabolites in plants, which are crucial in plant-pathogen interactions [6].
Vañó et al. proposed that SA, as a crucial phytohormone with a special role in plant defense responses to phytopathogens, is targeted by many clubroot pathogens, which actively attempt to suppress SA biosynthesis, accumulation, or downstream function. During host colonization, various phytopathogens may also modulate phytohormones directly involved in host defense responses, including SA, jasmonic acid (JA), and ethylene (ETH) [60]. Wang et al. reported that in apple, SA significantly upregulates NPR1 and PR1 and enhances endogenous salicylic acid levels. An E2H-based coating initiates the salicylic acid signaling pathway and induces an increase in resistance-related genes and enzymes [66].
In another study by Wang and Dong, it was discovered that the SA signaling pathway controls the synthesis of antimicrobial PR (pathogenesis-related) proteins, which are a critical mechanism against bacterial pathogens of plants[105]. Furthermore, Orozco-Mosqueda et al. proposed that bacterial pathogen infections elicit the secretion of SA-dependent PRs, which directly contributes to the acquisition of systemic acquired resistance (SAR) via the SA signaling pathway [106].
In table grapes, it has been shown that complex interplays among phytohormones and plant secondary metabolites, such as abscisic acid (ABA), indole-3-acetic acid (IAA), SA, and JA, play a crucial role in the fruit ripening and immune response or defense against necrotrophic fungal pathogens [107].
According to Tripathi et al., SA triggers systemic acquired resistance (SAR) in plants, leading to enhanced resistance against various pathogens [108]. Moreover, functional SA analogs have been found to induce plants' defensive responses to pathogens by activating SA-mediated signaling pathways [108,109].
A combination of water deficit and nutrient deprivation can also cause certain damages comparable to those caused by pathogens. For example, it was also reported that in house leek (Sempervivum tectorum L.) individuals, the lack of visual pathogens, the content of SA is strongly positively correlated with leaf hydration and relative water content. Therefore, it is possible that ABA and SA may interact antagonistically to modulate certain aspects related to morphological changes induced by water deficit [110].
As for SA’s roles in plant immunity, it was proposed that salicylic acid (SA) plays a critical role in plant immune signaling, contributing to the elicitation of the hypersensitive response (HR) and the activation of systemic acquired resistance (SAR) [111]. Additionally, it was also reported that SA triggers the hypersensitive response (HR) and systemic acquired resistance (SAR). SA synthesis rapidly follows pathogen detection through receptors such as PRRs and NBLRRs, leading to changes in cellular redox status and subsequent expression of immune-related genes. [112].
Apart from its roles in plant immunity, SA can also simultaneously exert certain effects on plants’ tolerances to abiotic stresses by acting together with other molecules under biotic stresses. For instance, it was reported that biomolecular condensate (BMC) assembly can be triggered by the accumulation of stress-related metabolites, including SA, or changes in cellular redox state. Small molecules like SA can bind and modify the intramolecular interactions of proteins, thus participating into plants’ signaling responses to both biotic and abiotic stresses [113].
4. The role of salicylic acid in plants’ tolerance responses to abiotic stresses
In recent decade, especially since 2021, numerous peer-reviewed review and research articles relating to plants’ tolerance responses to abiotic stresses have been published, and some articles published from 2021 to 2023 and selected from four database sites, including MDPI, Sciencedirect, Web of Science, and Scopus, are listed in Table 2. From Table 2, it can be concluded that mainly focus on such abiotic stress factors as drought, salt, heat, cold and heavy metals.
4.1. The role of salicylic acid in drought stress
It is generally held that the following harms can be caused to plants and crops by drought stress: water deficiency, hindered transpiration, wilting and withering, nutrient imbalance, disrupted physiological metabolism, decreased yield, and increased biotic stresses.
During drought conditions, the supply of water in the soil decreases, making it difficult for plants to absorb enough water for normal growth and development, resulting in water deficiency [114].
As a consequence of biotic stresses, under drought conditions, plants become more susceptible to pest and disease infestation, as their defense mechanisms are weakened. So, the harms caused by drought stress to plants and crops include water deficiency, hindered transpiration, wilting and withering, nutrient imbalance, disrupted physiological metabolism, decreased yield, and increased biotic stress. These harms significantly impact plant growth, development, crop yield, and quality.
As a crucial phytohormone, SA has been found to alleviate plant drought stress to some extent [85,87,98]. SA’s main functions in drought stress alleviation include: promoting seedling growth, enhancing plant antioxidant capacity, regulating plant water balance, promoting the expression of stress-related genes in plants, and regulating plant physiological metabolism. For example, it was reported that priming wheat seeds with 50µM SA has been found to enhance drought stress tolerance by upregulating antioxidant defense and glyoxalase systems, thus ensuring better wheat seedling establishment [11]. SA at a concentration of 0.5 mM effectively mitigates the negative effects of drought stress on Sardari ecotypes of winter wheat. It improves photosynthetic performance, maintains membrane permeability, induces stress proteins, and enhances the activity of antioxidant enzymes [18].
In addition, under drought conditions, SA can improve important physiological indicators in wheat, including seed germination rate, plumule length, radicle length, root length, and shoot length. The application of SA to wheat leaves can also enhance seedling vigor and indirectly improve water use efficiency by increasing levels of potassium (K+), calcium (Ca2+), and magnesium (Mg2+) in the roots. Additionally, SA can enhance chlorophyll content and membrane stability index (MSI), thereby boosting overall photosynthetic efficiency and improving yield and yield-related traits in drought-stressed wheat plants [34]. SA can also act as a positive regulator of drought tolerance by modulating antioxidant defense and reactive oxygen species (ROS) scavenging responses [35].
On the other hand, SA can also exert certain effects on plants’ tolerance responses to drought in combination with other stresses. For example, it was reported that under combined drought and heat stresses, SA levels were correlated with increased amino acid levels in certain citrus plants [36]. It was also reported that during the flowering stage, a physiological concentration of SA (1 mM) is significantly effective against salt stress. During the mature vegetative stage, proline accumulates under SA treatment, which is useful in developing NaCl-induced drought stress tolerance [44].
SA participates in the regulation of plant physiological metabolic processes, such as regulating stomatal opening and closure, thereby alleviating the adverse effects of drought stress on plants. For example, a study indicated that in rice, salicylic acid regulates stomatal aperture through the OsWRKY45-reactive oxygen pathway under salt and drought stresses. [115](Table.2)
4.2. The role of salicylic acid in salt stress
Among the factors limiting crop production, soil salinization is a major environmental challenge. For example, according to Fu et al. phytohormones such as ABA, auxin, CK, BRs, JA, GA, SA, and ETH play certain roles in enhancing crop salt tolerance [43].
The harm of salt stress to plants includes the following aspects: penetrability-related damages, imbalance of water balance, disruption of ion balance, impaired nutrient absorption, damages to photosynthesis, and oxidative stresses [42,43,47,82,84,97]. In summary, salt stress causes cellular dehydration, disruption of ion balance, impairment of nutrient absorption, and damage to photosynthesis, resulting in inhibited plant growth and development.
SA is an endogenous substance produced by plants, and it plays a role in alleviating salt stress in plants. There are several main mechanisms by which salicylic acid alleviates plant salt stress: ion balance regulation, antioxidant effects, and hormone level regulation. For example, Hundare et al. reported that treatment with SA (a foliar spray of 0.5 mM salicylic acid) enhances the contents of chlorophyll a, chlorophyll b, total chlorophyll, and carotenoids, while reducing chlorophyllase activity under salt stress conditions. Additionally, SA treatment further increases the activities of antioxidative enzymes, including superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD), which are induced by NaCl stress [24]. According to Shaki et al., exogenous SA improves the response of safflower to salinity by increasing glycine betaine, total soluble protein, carbohydrates, chlorophylls, carotenoids, flavonoid, and anthocyanin contents. SA application under saline conditions decreases the levels of proline, indicating successful acclimatization of these plants to saline conditions [45]. According to Jini and Joseph, SA application significantly increases the decreased rates of germination and growth (in terms of shoot and root lengths) caused by salt stress. The treatment of SA to the high and low saline soils enhances the growth, yield, and nutrient values of rice. SA application also reduces the accumulation of Na+ and Cl- ions caused by salt stress and at the same time increases the activities of antioxidant enzymes [46]. On the other hand, according to Ali et al., both individual and combined applications of PGPR and SA alleviate the negative effects of salinity and improve all the measured plant attributes. The response of PGPR + SA is significant in enhancing the shoot and root dry weights, relative water contents, chlorophyll a and b contents, and grain yield of maize under higher salinity level [47].
In terms of specific effects, SA can promote the growth and development of plant roots, enhance plant stress resistance, increase chlorophyll content and photosynthetic efficiency in leaves, and reduce transpiration and water loss [48,83,98]. Additionally, salicylic acid can promote plant physiological metabolism [36,94,96,102,111,117], thus increasing plant disease resistance and improving salt and drought tolerance. Overall, SA helps plants adapt to salt stress environments and mitigate the damage caused by salt stress. Other mechanisms by which salicylic acid enhances plant salt tolerance are mainly as follows: activation of stress response pathways [96,118,119,120,121,122], induction of heat shock protein synthesis [40], and regulation of the expression of stress response genes [103,123].
As an endogenous substance in plants, SA can enhance the plant's resistance by activating drought and salt stress response pathways when subjected to external stresses such as drought and salt stress. SA can activate protein kinases and kinase cascades, promote the expression of stress-resistant genes, and thus increase the plant's tolerance [15,116].
In summary, salicylic acid enhances plant salt tolerance or salt resistance through molecular biological mechanisms such as activating drought and salt stress response pathways, promoting the synthesis of protective enzymes, regulating ion channel activity, inducing modulating the expression of stress response genes.
4.3. The role of salicylic acid in heavy metal stress
Numerous studies in plant physiology and molecular biology have indicated that both exogenous and endogenous SA exert alleviative effects on plants' responses to various abiotic stresses, with a particular focus on heavy metal stresses. This effect is achieved through direct or indirect enhancement of their resistances or tolerances [31,49,50,51,52]. Pollutions caused by heavy metals are involved in a wide range of ions, such as Pb2+, Cd2+, Hg2+, Cr2+, Cu2+ and so on. Heavy metal pollutions exert toxic effects on plants mainly through the following aspects: inhibition of plants normal growth and development, suppression of plants’ absorption and utilization of nutrition, disruption of cellular structures and functions, accumulation of heavy metal ions in plants, and other toxic effects [49,50,51].
Heavy metal ions can exert potent inhibitory effects on plants’ growth and development by disrupting physiological processes concerning metabolism, and suppressing photosynthesis, respiration and transpiration. Heavy metal ions can disrupt plants’ absorption and utilization of nutrition. Some heavy metal ions can directly compete with the nutritious elements essentially needed by plants, thereby making it hard for plants to absorb and utilize nutrition and limiting their normal growth. For example, according to Ur Rahman et al., the exogenous application of SA to plants induces acclimatization impacts by improving resistance and leading to tolerance against heavy metal stress. This is achieved through the activation of stress signaling hormonal pathways and the overexpression of stress-related genes and enzymes [49]. According to Saini et al., the exogenous application of SA has been reported to restore heavy metal-induced damage to various photosynthetic traits in plants, thereby improving their stress tolerance. Another major mechanism involved in SA-regulated heavy metal stress tolerance in plants is the modulation of ROS detoxification machinery to protect the plants against oxidative stress. SA has also been documented to modulate the levels of non-enzymatic antioxidant metabolites, including proline, upon exposure to heavy metal stress [51].
It was widely reported and reviewed that SA exerts certain alleviative effects on the toxicity of heavy metals. SA can enhance the activities of antioxidant enzymes (including SOD, POD and CAT), thus lowering the contents of intracellular and intercellular reactive oxygen species (ROS) and reducing oxidative damages caused by heavy metal ions in plants. According to Nivedha et al., SA has been used as an analgesic, anti-inflammatory, and antipyretic agent, and it has also shown a mitigating effect on cadmium toxicity. SA has been reported to reduce heat stress-induced ROS production and its subsequent oxidative damage [57]. According to Jia et al., exogenous application of SA has been found to significantly reduce cadmium (Cd) accumulation in tomato plants and alter its distribution. SA pretreatment enhances cell wall polysaccharide synthesis and related gene expression, leading to cell wall thickening and blocking Cd penetration. Additionally, SA decreases pectin methylesterase activity and content, reducing cell wall Cd accumulation and altering the Cd partition ratio [90].
Although SA at certain concentrations exerts some alleviative effects on stresses by heavy metal ions, SA at high concentrations also brings about some toxic effects to plants. Therefore, appropriate SA applications into the enhancement of plants’ tolerances to stresses should be taken into considerations in terms of both SA concentrations and patterns for applications.
4.4. The role of salicylic acid in high temperature stress
High temperatures or heat can have significant harmful effects on plant growth and development [2,21]. The main harms of high temperature stress on plants include the following several aspects: impairment and reduction of photosynthesis, increased respiration, water stress, cell membrane damage, DNA damage, protein denaturation, reduced nutrient uptake, delayed flowering and fruit set, increased susceptibility to diseases and pests, and other physiological and biochemical changes.
High temperature stress can also lead to a series of physiological and biochemical changes within the plant, such as imbalances in the antioxidant system, abnormal levels of endogenous hormones, and accumulation of reactive oxygen species, affecting the plant's adaptability to the environment. Therefore, it is crucial for farmers and gardeners to monitor and mitigate these effects to ensure optimal plant health and productivity.
SA is an important endogenous signaling molecule in plants and crops, which plays a role in alleviating heat stress [33,74,82,86,87]. Its molecular biological mechanisms mainly include the following aspects: regulation of reactive oxygen species (ROS) homeostasis, accumulation of antioxidants [58,85], SA’s involvement in epigenetic regulation, and SA’s participation in hormonal signaling [74,87].
SA can maintain the cellular ROS homeostasis by regulating the accumulation and clearance of reactive oxygen species. Under heat stress conditions, the production of ROS increases significantly, leading to oxidative damage in cells. However, salicylic acid can effectively remove ROS and alleviate cell damage. Heat stress induces the synthesis and accumulation of antioxidants in plants, enhancing the cell's antioxidant capacity. Salicylic acid can promote the synthesis and accumulation of antioxidants by regulating the expression of related genes, thereby enhancing the plant's resistance to heat stress [11,82,86,87].
It is generally considered that SA plays a regulatory role in mitigating the damage caused by high temperature stress. It acts through various molecular mechanisms, including enhancing the antioxidant system, regulating the synthesis of resistance proteins, modulating signal transduction pathways, and regulating ion balance.SA alleviates high temperature stress in plants through mechanisms that enhance antioxidant capacity, increase resistance protein synthesis, regulate signal transduction pathways, and maintain ion balance.
These mechanisms collectively enable plants to better cope with the damage caused by high temperatures. According to Balfagón et al., under combined drought and heat stresses, SA levels were correlated with increased amino acid levels in certain citrus plants [36]. According to Rasheed et al., both NO and SA protect plants against heat stress by enhancing sulfur assimilation, reducing oxidative stress, and increasing the activity of antioxidant enzymes. Supplementation of NO or SA along with sulfur under heat stress recovers losses, improves photosynthesis and growth [42]. According to Li et al., under heat stress, transcripts of several salicylic acid biosynthesis and signaling genes, as well as heat stress-responsive genes, were significantly elevated in SO2-pretreated maize seedlings. This increase in endogenous SA levels activated the antioxidant machinery and strengthened the stress defense system, improving the thermotolerance of the seedlings [73].
It should be noted that as an endogenous signaling molecule, the mechanism of action of SA is very complex and is regulated by multiple environmental and genetic factors. Different plants and crops may also exhibit variations in their response to salicylic acid. Therefore, further research is needed to gain a deeper understanding of the molecular biological mechanisms of SA in alleviating heat or high temperature stresses.
4.5. The role of salicylic acid in low temperature stress
Low temperature extremes (cold stress, including chilling 0–15℃ and freezing<0℃ temperatures) limit plant growth and development and severely affect plant physiology and biochemical and molecular processes [38].
It is widely considered that SA plays vital roles in plant stress tolerance to a wide range of abiotic stresses, including low-temperature or cold stress [37,38,39,118]. For instance, SA can enhance plants’ resistance to cold, and mitigate plants’ injuries caused by low-temperature environment. In response to cold stress, SA can regulate plants’ metabolism and induce the biosynthesis of cold-resistant proteins, thus improving plants tolerance to low-temperature stresses. According to Seo et al., SA plays a crucial role in regulating chilling tolerance in plants. It controls ROS levels by regulating antioxidant metabolism during chilling stress. SA treatment enhances antioxidant enzymes, thereby increasing chilling tolerance. However, a high concentration of SA may lead to the accumulation of hydrogen peroxide, which reduces chilling tolerance [37]. According to Raza et al.,SA has been found to regulate seed germination rate, early plant responses, protein pattern in apoplasts, and enhance antioxidative system, growth and photosynthetic attributes, net photosynthetic rate, and SOD, CAT, and POD activities under cold stress [38].
It is generally considered that SA is a crucial plant hormone that plays a significant regulatory role in the response of plants to low temperature stress. Its mechanisms of action include the following: promotion of activation of the antioxidant system, preservation of cell membrane stability, regulation of gene expression, and promotion of low temperature signal production.
In summary, the mechanisms by which salicylic acid alleviates plant cold and freezing stress involve the synthesis of cold-resistant proteins, regulation of signal transduction pathways, ROS scavenging, and modulation of hormone synthesis and signaling. These mechanisms interact and work together to enhance the plant's adaptability to low temperature stress.
4.6. The role of salicylic acid in other abiotic stresses
Apart from SA’s roles in the abovementioned abiotic stresses, SA also plays a certain part in plants’ tolerance responses to other biotic stresses, such as arsenic stress [19], UV radiation-induced oxidative stress [57], waterlogging stress [94], and strontium stress [127] (Table 2, Table 3). According to Khan et al., application of SA promotes photosynthesis and growth in plants, with or without arsenic stress, by improving plant defense systems and reducing oxidative stress through crosstalk with ethylene (ETH) and nitric oxide (NO) [19]. According to Nivedha et al., the mechanisms of radio-protective activity of salicylic acid-rich fractions were ROS inhibition and lipid peroxidation inhibition [57]. According to Hasanuzzaman et al., exogenous application of SA and kinetin (KN) reduces the content of malondialdehyde (MDA), hydrogen peroxide (H2O2), proline, and leaf electrolyte leakage under waterlogging stress. Additionally, supplementation with SA and KN enhances the activity of various antioxidant enzymes, such as APX, MDHAR, DHAR, GR, CAT, GST, GPX, POD, Gly I, and Gly II, under waterlogging stress. Both exogenous SA and KN effectively improve ROS metabolism and enhance waterlogging stress tolerance in soybean by enhancing antioxidant defense and upregulating the glyoxalase system [94]. According to Pyo et al., under strontium stress, the expression of pathogenesis-related protein genes, including PR1, PR2, and PR5, has been found to be related to SA in response to stress caused by strontium [127].
5. Interaction of salicylic acid with other molecules in plants’ defense responses
The interaction between salicylic acid and other small molecules or plant hormones can enhance or weaken the resistance of plants through various molecular biology mechanisms. Here are some possible mechanisms: SA’s regulation of the biosynthesis of other phytohormones, SA’s regulation of hormone signal transduction pathways, SA’s regulation of the expression of hormone-responsive genes, and competitions between SA and its interacting hormones.
SA may regulate the resistance of plants by modulating the synthesis of other plant hormones [12,110]. For example, salicylic acid has been found to inhibit the synthesis of ethylene and the methylation of jasmonic acid (JA), thereby reducing the plant's response to adversity [12,19,56,125,126,127].
SA can intervene in the signal transduction pathways of other plant hormones, thereby enhancing or weakening the plant's resistance to stress [56,70,127]. For example, SA can regulate the plant's resistance by activating or inhibiting key components of hormone signal transduction pathways [125,128].
SA is a universally recognized signaling molecule that is considered a pivotal stress hormone in plants. It plays a crucial role in both normal plant growth and stress tolerance. Numerous studies have demonstrated that SA's involvement in plants' tolerance to abiotic and biotic stress is a vital mechanism for their normal growth, defense against unfavorable environmental factors and pathogens, and even survival. For instance, Yang et.al reviewed that SA has been found to confer abiotic stress tolerance in horticultural crops by acting as a signaling molecule that triggers various physiological and morphological responses [129].
SA and other plant hormones may compete to activate or inhibit shared components or signal transduction pathways, thereby regulating the plant's resistance to stress. This competition may result in dominant or complementary effects of hormone signals, affecting the plant's response to adversity.
SA is also an important signaling molecule for triggering responses to a myriad of abiotic stresses, including cold, heat, drought, ultraviolet, heavy metals, and so on, and participates in developmental signaling pathways that control and regulate many physiological and biochemical processes in normal plant growth, development, flowering, ripening and ageing [8,27,28,29,53,54,58,59,72,91,96].
For example, according to Das et al., Zinc-chitosan-salicylic acid (ZCS) nanoparticles have the ability to improve morpho-physiological features, photosynthetic pigment status, osmotic status, osmoprotectant synthesis, ROS scavenging enzyme activity, membrane integrity, cellular protection, and yield increment during drought stress [33]. According to Malko et al., the use of multiple chemicals, including SA, Zn, PAs, SNP, etc., with pre- and post-emergence treatment, has enhanced drought tolerance and maintained the normal physiology of wheat under changing climates [34]. According to Lafuente and Romero, Citrus fruits’ heat-induced cross-adaptation to chilling is regulated by phytohormones such as JA, JA-related metabolites, SA, IAA, and ABA [39]. According to Wei et al., SA and auxin regulate the transcripts of MeHsf8-MeHSP90.9 module in an antagonistic manner. MeHSP90.9 interacts with MeSRS1 and MeWRKY20 to activate SA biosynthesis while inhibiting auxin biosynthesis, providing a mechanistic understanding of MeHSP90.9 co-chaperones in plant immunity [40]. According to Afzal et al., SA and MLE (moringa leaf extract) treatments recorded the maximum number of productive tillers and biological yield. SA treatment maximally enhanced free proline levels and protein content in wheat cultivars [41]. According to Rasheed et al., both NO and SA protect plants against heat stress by enhancing sulfur assimilation, reducing oxidative stress, and increasing the activity of antioxidant enzymes. Supplementation of NO or SA along with sulfur under heat stress recovers losses, improves photosynthesis and growth [42]. According to Mutlu-Durak et al., although SA can enhance salinity tolerance, the beneficial effects observed with willow extract applications cannot be explained by just their SA content. The same benefits could not be obtained with the exogenous application of pure SA [48]. According to Khalil et al., SA, alone and in combination with kinetin or calcium, has been found to improve growth traits, photosynthetic pigment, carbohydrate contents, and nitrogenous constituents in Phaseolus vulgaris plants. Additionally, plants cultivated from seeds soaked in salicylic acid in combination with kinetin or calcium showed enhanced activities of antioxidant enzymes and proline accumulation under nickel and/or lead stress [50]. According to Thepbandit et al., the use of SA-Ricemate as a foliar spray at certain concentrations can reduce the severity of bacterial leaf blight disease by 71%. In field conditions, SA-Ricemate significantly reduces disease severity by 78% and increases total grain yield [67]. According to Kotapati et al., supplementation with both SA and SNP significantly reduced the toxic effect of Ni and increased root and shoot length, chlorophyll content, dry mass, and mineral concentration in Ni-treated plants. Exogenous application of SA or SNP, specifically the combination of SA and SNP, protects finger millet plants from oxidative stress observed under Ni treatment [76]. According to Talaat and Hanafy, co-application of SA and spermine (SPM) can be a superior method for reducing salt toxicity in sustainable agricultural systems. Exogenously applied salicylic acid and/or SPM relieved the adverse effects caused by salt stress and significantly improved wheat growth and production by inducing higher photosynthetic pigment content, nutrient acquisition, ionic homeostasis, osmolyte accumulation, and protein content [82,84].
According to Munsif et al., co-application of SA and K+ under drought stress conditions significantly mitigates negative effects induced by drought stress. This combined treatment has shown to improve grain yield and enhance water use efficiency, particularly under mild and severe drought stress, respectively. Co-application of SA and K+ helps regulate osmotic and metabolic processes, stabilize cell components, and ultimately overcome the detrimental impacts of drought stress [85]. According to Alotaibi et al., co-applying essential plant nutrients with SA has proven to be a feasible, profitable, and user-friendly strategy for mitigating the negative effects of deficit irrigation stress. This approach has also demonstrated further improvements in the growth and production of wheat under normal irrigation conditions [86]. According to Zhang et al., MYB transcription factors play important roles in various physiological activities of plants, including phytohormone signal transduction and disease resistance. Transcripts of MiMYB5, -35, -36, and -54 have shown positive responses to early treatments of SA, methyl jasmonate (MeJA), and hydrogen peroxide (H2O2), suggesting their involvement in these signaling pathways [88]. According to Li et al., in plant immunity, the hormones SA and jasmonic acid (JA) play mutually antagonistic roles in resistance against biotrophic and necrotrophic pathogens, respectively. Transgenic plants with a synthetic dual SA- and JA-responsive promoter controlling the expression of an antimicrobial peptide exhibit enhanced resistance to a wide range of pathogens [93].
According to Kumar and Giridhar, exogenous application of SA and MeJA has been shown to alleviate the maturation-triggered repression of N-methyltransferase genes (NMTs). This suggests a potential cross-talk between signaling cascades involved in the regulation of NMT gene expression during maturation and the overexpression induced by SA and MeJA treatment in the mature endosperm [128].
In summary, the interaction between SA and other plant hormones enhances or weakens the plant’s tolerance through mechanisms such as regulation of hormone synthesis, signal transduction pathways, gene expression, and hormone competition. The specific details of these mechanisms will depend on the plant species, type of stress, and the context of hormone interactions.

Table 1.
Salicylic Acid’s Roles in Plant Biotic Stress Responses.
| Stress Factors | Plant Species or Categorizations | SA’s Roles or Other Signaling Molecules’ Effects on SA | References |
| mycotoxin Fumonisin B1 | Arabidopsis and various crops | Activation of various antioxidant enzymes, PR genes, SAR pathways, and PCD | [3] |
| multiple pathogens | general plants | Activation of SAR pathways, defense response against biotrophic pathogens | [4] |
| multiple biotic stresses | general plants | Activation of MAPK, CDPK and other PKs; enhancement of SM biosynthesis | [6] |
| Pectobacterium carotovorum | Chinese cabbage | Possibly no participation into the defense response to Pectobacterium carotovorum infection | [12] |
| pathogens | Arabidopsis | Modulation of SA biosynthesis by SABC1’s role as a molecular switch in balancing plant defense and growth | [25] |
| pathogens | cassava | Activation of SA biosynthesis by MeHSP90.9’s interaction with MeSRS1 and MeWRKY20 | [40] |
| bacterial pathogens | jujube | Indicator of antimicrobial activity | [55] |
| pathogens | cruciferous crops | Disruption of SA biosynthesis and accumulation by biotrophic pathogens to hijack plant cellular processes | [60] |
| insect herbivoresand pathogens | general plants | Defense responses of SA induction to pathogens or aphid feeding | [61] |
| pathogens | Arabidopsis thaliana | Role of T. hamatum as an inter-plant communicator as a consequence of antagonism between JA and SA | [62] |
| fungal pathogens | apple | Significant upregulation of NPR1 and PR1; enhancement of endogenous SA; initiation of SA signaling pathway by E2H-based coating | [66] |
| bacterial pathogens | rice | Reduction of disease severity; significant increment of total grain yield | [67] |
| weeds | crops | Alteration of crop growth by weeds through increment of SA signaling process | [68] |
| pathogens | wheat | Drastic change of contents of compounds involved in biosynthesis and metabolism of SA in roots | [74] |
| pathogens | general plants | Possible regulation of SA synthesis by cAMP level; stomatal closure by SA, NO and cGMP | [92] |
| pathogens | tobacco and Arabidopsis | Antagonism of SA and JA in resistance to biotrophic and necrotrophic pathogens | [93] |
| pathogens | Arabidopsis, poplar | Pathogen-stimulated SA synthesis; SA-mediated defense; SA’s interplay with other hormones | [99] |
| pathogens | general plants | Basal defense and amplification of local immune responses; establishment of SAR; regulation of plant immunity | [102] |
| bacterial pathogens | kiwifruit vines | Induction of SA-pathway defense genes by postharvest ASM | [103] |
| pathogens | general plants | Establishment of plant immunity; elicitation of defense responses; establishment of both local and systemic resistance against various pathogens | [104] |
| pathogens | general plants | Elicitation of SAR through signals involving SA | [106] |
| pathogens | table grape | Correlation of SA accumulation and degree of anthracnose disease resistance and symptoms; resistance induction by SA level increase | [107] |
| pathogens | general plants | Elicitation of HR; activation of SAR; responses of SA synthesis to pathogens via receptors | [112] |
| pathogens | strawberry | Effects of FaSnRK1α on SA content through regulation of FaPAL1 and FaPAL2 expressions; increment of pathogen-resistance | [119] |
| pathogens | rice | Rice susceptibility caused by proliferation of bacterial pathogens facilitated by compromised SA signaling | [121] |
| pathogens | grapevines | GA’s role as a component of the grapevine SA-dependent response | [123] |
| pests | grapevines | Reduced injury by JA and SA application; reduction of number of eggs laid by D. suzukii females in JA and SA-treated plants | [125] |
Abbreviations: SA (salicylic acid), PR genes (pathogen-related genes), SAR (systemic acquired resistance), PCD (programmed cell death), SM (secondary metabolites), PK (protein kinase), MAPK (mitogen-activated protein kinase), CDPK (calcium dependent protein kinase), SABC1 (salicylic acid biogenesis controller 1), JA (jasmonic acid), PR1 (pathogenesis-related genes 1), NPR1 (nonexpressor of pathogenesis-related genes 1), cAMP (cyclic adenosine monophosphate), NO (nitric oxide), cGMP (cyclic guanosine monophosphate), ASM (acibenzolar-S-methyl), HR (hypersensitive response), GA (gibberellins).
Table 2.
Salicylic Acid’s Roles in Plant Abiotic Stress Responses .
| Stress Factors | Plant Species or Categorizations | SA’s Roles or Other Signaling Molecules’ Effects on SA | References |
| drought | wheat | Elicitation of drought stress tolerance; better seedling establishment | [11] |
| wounding | Arabidopsis and maize | Mediation of glutamate receptors in regeneration; anti-correlation between increased SA response and regeneration in older tissue | [14] |
| drought | winter wheat | Effective alleviation of drought stress | [18] |
| arsenic stress | rice | Promotion of photosynthesis and growth through crosstalk with ETH and NO | [19] |
| fomesafen toxicity | sugar beet | Alleviation of fomesafen stress, improvement of some photosynthetic indexes in leaves, maintenance of cell membrane integrity, and amelioration of adverse effects of fomesafen on seedling growth | [20] |
| high temperature | maize | Reduction of decrease in dry matter and leaf area index; alleviation of chloroplast ultrastructure disruption, and decrease in photosynthetic rate and chlorophyll content; promotion of antioxidant enzyme activity and counteraction of increase in JA and ABA contents | [21] |
| salt | ginger | Enhancement of chlorophyll and carotenoid contents; further increase in NaCl-induced antioxidative enzymatic activities | [24] |
| drought | canola | Development of stress tolerance and improvement of plant growth by co-application of Pseudomonas putida and salicylic acid | [32] |
| drought | wheat | Amelioration of physiological and photosynthetic features; improvement of osmotic status; activation of ROS scavenging enzymes; promotion of yield increment | [33] |
| drought | wheat | Refinement of drought tolerance; maintenance of normal physiology | [34] |
| drought | general plants | Positive regulation of drought tolerance phenotype | [35] |
| combined drought and heat stresses | citrus plants | Correlation of SA levels with the increase of amino acid levels | [36] |
| cold | pepper | Regulation of antioxidant metabolism to control ROS; enhancement of antioxidant enzymes to increase chilling tolerance | [37] |
| cold | Arabidopsis and crop plants | Regulation of seed germination rate, early plant responses, protein pattern in apoplasts; enhancement of antioxidative system, growth and photosynthetic attributes | [38] |
| heat, cold | citrus plants | Involvement in fruits’ heat-induced cross-adaptation to chilling | [39] |
| heat | wheat | Protection against heat stress; reduction of oxidative stress; increment of antioxidant enzyme activities; improvement of photosynthesis and growth | [42] |
| salt | cereal crops | Enhancement of salt tolerance | [43] |
| salt, drought | chilli | Significant effects against salt stress; elicitation of proline accumulation for the development NaCl-induced drought stress tolerance | [44] |
| salt | maize | Alleviation of salinity; significant enhancement of grain yield | [47] |
| salt | maize | Enhancement of salinity tolerance | [48] |
| heavy metals | general plants | Induction of acclimatization impacts; improvement of tolerance against heavy metal stress | [49] |
| heavy metals | common bean | Improvement of growth traits and photosynthetic indexes; enhancement of antioxidant enzyme activities and proline accumulation | [50] |
| heavy metals | general plants | Improvement of stress tolerance; modulation of ROS detoxification machinery to protect plants against HM induced oxidative stress; modulation of non-enzymatic antioxidant metabolites levels | [51] |
| drought | Camelliaoleifera | Decline of SA level during drought stress; Exhibition of stronger antioxidant capacity, water regulation ability, and drought stress protection by drought treated group | [71] |
| heat | maize | Significant elevation of transcripts of SA biosynthesis and signaling, and heat stress-responsive genes in SO2-pretreated seedlings; improvement of seedlings’ thermotolerance | [73] |
| salt | sugarcane | Effective mitigation of adverse impact; facilitation of better germination and early seedling growth; maintenance of relative water content, membrane stability and overall plant growth | [77] |
| salt | wheat | Alleviation of plant growth inhibition; increment of N, P, and K+ acquisition; enhancement of endogenous SA and SPM levels by exogenous SA and/or SPM applications | [82] |
| salt | St John’s wort | Improvement of growth | [83] |
| salt | wheat | Salt stress alleviation by SA and/or SPM; significant improvement of growth and production | [84] |
| drought | wheat | Substantial reduction of drought influence, and enhancement of grain yield and water use efficiency by co-application of K+ and SA | [85] |
| dry climatic conditions | wheat | Attenuation of deficit irrigation, and improvement of growth and production by co-application of essential plant nutrients and SA | [86] |
| drought | wheat | Regulation of stress response | [87] |
| cadmium | tomato | Significant reduction of cell wall Cd accumulation; changes of Cd distribution | [90] |
| waterlogging stress | soybean | Reduction of some physiological indexes by SA and KN; enhancement of antioxidant defense by SA and KN; effective improvement of ROS metabolism and waterlogging stress tolerance | [94] |
| salt | cashew | Attenuation of salt stress; increment of photosynthetic pigment synthesis | [97] |
| drought | oregano | Improvement of PSII efficiency under moderate drought stress | [98] |
| water deficit and nutrient deprivation | Sempervivum tectorum L | Strong positive correlation between SA and some physiological indexes; possible antagonistic modulation of ABA and SA on water deficit-induced morphological changes | [110] |
| water deficiency | potato | Mitigation of oxidative damage; alleviation of water deficiency stress | [114] |
| salt, drought | rice | Regulation of stomatal aperture through the OsWRKY45-reactive oxygen pathway; regulation of adaptation to soil salinity and drought stress | [115] |
| abiotic stresses | Watermelon | Regulation of some ClBBX genes’ transcription for possible growth and development regulation | [118] |
| abiotic stresses | horticultural crops | Provision of abiotic stress tolerance; elicitation of various physiological and morphological responses to stress; regulation of stress-responsive genes’ expression; direct interaction with various hormones, proteins and enzymes involved in abiotic stress tolerance | [129] |
Abbreviations: SA (salicylic acid), ETH (ethylene), NO (nitric oxide), JA (jasmonic acid), ABA (abscisic acid), ROS (reactive oxygen species), HM (heavy metals), SPM (spermine), Cd (cadmium), KN (kinetin), PSII (photosystem II), ClBBX(Citrullus lanatus B-BOX).
Table 3.
Salicylic Acid’s Roles in Plant Biotic + Abiotic Stress Responses.
| Stress Factors | Plant Species or Categorizations | SA’s Roles or Other Signaling Molecules’ Effects on SA | References |
| multiple stresses | maize | Expression of SM biosynthesis pathway | [2] |
| multiple stresses | general plants | SA’s interplays with MT; cross-talks between MT, ETH, and SA in plants’ pathogen resistance; SA’s induction of plants’ defense responses against pathogens | [5] |
| multiple stresses | general plants | Freezing tolerance establishment; improvement of PSII stability; increased chlorophyll content and improved yield of rice plants with bacterial blight disease | [22] |
| multiple stresses | general plants | Partly independent role of SA and MeSA’s involvement in the development of SAR | [23] |
| environmental stresses | sorghum | SA’s participation in sorghum plants’ responses to abiotic and biotic stresses | [26] |
| abiotic and biotic stresses | Mango | Positive responses of MiMYB transcription factor genes to early treatments of SA, MeJA, and H2O2 | [88] |
| abiotic and biotic stresses | general plants | Evolution of MT, auxin and SA from a common precursor, chorsimate | [109] |
| pathogens, heat | Arabidopsis | Restoration of SA production for plant immunity by optimized CBP60g expression at elevated growth temperature | [111] |
| abiotic and biotic stresses | general plants | Elicitation of BMC assembly by accumulation of SA; modification of proteins’ intramolecular interactions by SA binding | [113] |
| strontium stress, biotic stress | Arabidopsis | Relationship between expression of some PR genes and SA’s response to Sr stress | [127] |
Abbreviations: SA (salicylic acid), SM (secondary metabolites), MT (melatonin), ETH (ethylene), PSII (photosystem II), MeSA (methyl salicylate), SAR (systemic acquired resistance), BMC (biomolecular condensate), Sr (strontium).
6. Conclusions and Perspective
In recent decade, especially since 2021, numerous peer-reviewed review and research articles concerning plants’ tolerance responses to both biotic and abiotic stresses have been published, and some articles published from 2021 to 2023 and selected from four database sites, including Sciencedirect, Web of Science, Scopus, and MDPI, are listed in Table 3. From Table 3, it can be seen that these articles mainly deal with plants stressed simultaneously by multiple biotic and abiotic stresses, which are of vital importance in crop improvement and stress management for greater yield.
It is widely known to alleviate plant stress tolerance at the molecular level through various mechanisms. For instance, SA can induce defense genes in plants by activating the expression of a wide range of defense-related genes, including pathogenesis-related (PR) genes, which encode proteins involved in plant defense responses. PR proteins strengthen the plant's immune system and confer resistance to pathogens or pests.
SA can also enhance antioxidant capacity. SA boosts the production of antioxidant enzymes, such as superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD). These enzymes help neutralize reactive oxygen species (ROS) generated during stress conditions, thus reducing oxidative damage and improving stress tolerance. Meanwhile, SA can regulate other phytohormones. For example, SA interacts with other phytohormones, such as jasmonic acid (JA) and ethylene, to modulate plant stress responses. It often acts synergistically or antagonistically with these hormones, depending on the stress type. This crosstalk helps plants fine-tune their defense strategies to combat specific stresses effectively.
In addition, SA primes plants’ immune system by pre-activating defense mechanisms before stress onset. This priming induces a faster and stronger defense response when a stressor is encountered, providing a higher level of stress tolerance or resistance. It is also generally held that SA can activate systemic acquired resistance (SAR), because SA is a key player in SAR, a long-lasting defense response that occurs throughout the entire plant system following localized pathogen attack. SAR confers enhanced resistance to a broad range of pathogens by activating defense responses in uninfected parts of the plant.
On the other hand, stomatal behavior, especially stomatal closure, is generally considered as important physiological indexes for measuring plants’ tolerances. It is generally held that SA can modulate stomatal behavior, influence stomatal closure, thus reducing water loss from the plant under stress conditions. This modulation helps conserve water and maintain plant hydration during drought or other stressors[92,115].
Meanwhile, a wealth of research at the molecular level on SA signaling pathways has indicated that in plants, SA-SABP complexes formed by SA conjugating with SABPs (SA binding proteins) spread cellular messages to secondary messengers such as H2O2 and Ca2+. These messages are amplified by self-feedback mechanisms and transduced, triggering cellular responses and activating a cascade of transcription factors, including NPR1, TGA, and WRKY. This, in turn, promotes the interaction among these factors and the expression of pathogenesis-related (PR) genes that confer systemic acquired resistance (SAR) to plants.
For example, Zhu et al. demonstrated that localized infection by an avirulent pathogen leads to an increase in SA levels, which then activates NPR1 (nonexpressor of pathogenesis-related genes 1) to induce systemic acquired resistance (SAR) and promote cell survival to prevent the spread of cell death signals. In response to varying levels of SA, two other SA receptors, NPR3 and NPR4, act together to maintain optimal NPR1 levels by degrading NPR1, thereby constraining cell death signals to localized levels. Additionally, methyl-SA (MeSA) functions as a mobile signaling molecule for SAR, and activation of SAR confers resistance to the entire plant against secondary infection by a virulent pathogen in systemic leaves [104].
Contrarily, Ding et al. proposed that as SA receptors participating in SA-mediated signaling pathways for plant immunity, NPR1 plays an opposing role to that performed by NPR3 and NPR4 [79] .
Overall, salicylic acid’s alleviative effects on plants' stress tolerance or resistance at the molecular level involve gene regulation, modulation of phytohormonal responses, strengthening antioxidant defense capacity, immune system priming, activation of systemic acquired resistance, and regulation of stomatal behavior. These mechanisms collectively contribute to enhanced stress tolerance and improved plant health.
Numerous studies have also found that SA, as an endogenous signaling molecule in plants, can regulate plant stress tolerance under environmental stress. Salicylic acid enhances plant resilience by modulating the activity of a series of antioxidant enzymes, regulating plant growth and development, and controlling gene expression. Researches showed that salicylic acid can activate the plant's immune response by regulating the expression of a series of disease-resistant genes, thereby enhancing the plant's ability to resist diseases. For example, Luo et al. reported that FaSnRK1α regulates the expression of FaPAL1 and FaPAL2, affecting the content of SA. This regulation, along with the interaction with the FaWRKY33.2 transcription factor, increases the resistance of strawberry fruit to Botrytis cinerea through the SA signaling pathway [119]. Hui et al. reported that bacterial pathogens Xoo and Xoc have been found to activate the expression of miR156 and miR529, which subsequently target and cleave the mRNAs of three self-interacting proteins, namely OsSPL7, OsSPL14, and OsSPL17. The downregulation of these three OsSPLs leads to insufficient activation of their downstream target genes, OsAOS2 and OsNPR1. As a result, the levels of jasmonic acid (JA) decrease, compromising the salicylic acid (SA) signaling pathway. This compromised SA signaling facilitates the proliferation of Xoo or Xoc, ultimately causing susceptibility in rice plants [121].
It is generally held that SA also plays an important regulatory role in plant growth and development. The latest researches have indicated that salicylic acid can regulate seed germination, seedling growth, and plant establishment.
It was reviewed and demonstrated that SA is involved in stress-induced programmed cell death (PCD) in plants. Researches indicate that SA can induce apoptosis in plant cells, thereby enhancing plants' defense against adverse conditions.
Overall, research on the involvement of salicylic acid in plant stress resistance demonstrates its importance in regulating plant stress resistance, immune system, growth and development, and stress-induced programmed cell death. These research findings contribute to the understanding of the physiological regulation mechanisms of salicylic acid in plants and provide a theoretical basis for further improving plant stress resistance.
As an important plant signaling molecule, SA plays crucial regulatory roles in plant stress tolerance. Through regulation of multiple cell signaling pathways, it is involved in plant responses to diseases, drought, and salt stress. In-depth researches on the cell signaling pathways involving SA will help us better understand the mechanisms of plant stress responses, providing a theoretical basis for enhancing plant adaptability and genetic improvement of crops.
Genomics and proteomics are important tools for understanding the role of SA in alleviating plant stress. Genomics techniques, such as RNA-seq and microarray technology, can identify stress-regulated genes and examine how salicylic acid affects their expression. On the other hand, proteomic studies utilizing protein mass spectrometry can analyze the changes in the plant proteome under stress conditions regulated by SA. By comparing the protein expression profiles of salicylic acid-treated and control groups, proteins regulated by salicylic acid can be identified. These findings can further reveal the mechanisms of plant stress response and the involvement of salicylic acid.
In addition, bioinformatics analysis is crucial in processing the data generated by genomics and proteomics. Integrating these datasets allows for a deeper understanding of how salicylic acid alleviates plant stress, including the activation of specific pathways and involvement in signaling cascades. For instance, it enables the identification of salicylic acid-related regulatory factors and the prediction of stress-responsive genes regulated by salicylic acid.
It is worth noting that the role of SA in plant tolerance responses can vary depending on the specific stressor, plant species, and other factors. Additionally, the exact mechanisms involved in SA-mediated stress responses are still being investigated, and further research is needed to fully understand its complexities.
In a word, in the future, genomics and proteomics will continue to be indispensable in studying the mitigation of plant stress by SA, because they can pinpoint the genes and proteins associated with SA and elucidate the mechanisms through which it regulates stress response.
Author Contributions
W.S. and H.S. planned content and wrote manuscript. Other authors(A.Z.;L.F.;Y.J.) collect relevant reference materials for writing this review. All authors have read and agreed to the published version of the manuscript.
Funding
The work has been supported in part by Double-hundred Talent Plan of Jiangsu Province,China.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ali, S.; Ganai, B.A.; Kamili, A.N.; Bhat, A.A.; Mir, Z.A.; Bhat, J.A.; Tyagi, A.; Islam, S.T.; Mushtaq, M.; Yadav, P.; et al. Pathogenesis-related proteins and peptides as promising tools for engineering plants with multiple stress tolerance. Microbiological Research 2018, 212-213, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Arias, C.C.; Ramírez-Godoy, A.; Restrepo-Díaz, H. Influence of drought, high temperatures, and/or defense against arthropod herbivory on the production of secondary metabolites in maize plants. A review. Current Plant Biology 2022, 32. [Google Scholar] [CrossRef]
- Iqbal, N.; Czékus, Z.; Poór, P.; Ördög, A. Plant defence mechanisms against mycotoxin Fumonisin B1. Chemico-Biological Interactions 2021, 343. [Google Scholar] [CrossRef] [PubMed]
- Khoshru, B.; Mitra, D.; Joshi, K.; Adhikari, P.; Rion, M.S.I.; Fadiji, A.E.; Alizadeh, M.; Priyadarshini, A.; Senapati, A.; Sarikhani, M.R.; et al. Decrypting the multi-functional biological activators and inducers of defense responses against biotic stresses in plants. Heliyon 2023, 9. [Google Scholar] [CrossRef]
- Khan, M.S.S.; Ahmed, S.; Ikram, A.u.; Hannan, F.; Yasin, M.U.; Wang, J.; Zhao, B.; Islam, F.; Chen, J. Phytomelatonin: A key regulator of redox and phytohormones signaling against biotic/abiotic stresses. Redox Biology 2023, 64. [Google Scholar] [CrossRef]
- Anjali; Kumar, S. ; Korra, T.; Thakur, R.; Arutselvan, R.; Kashyap, A.S.; Nehela, Y.; Chaplygin, V.; Minkina, T.; Keswani, C. Role of plant secondary metabolites in defence and transcriptional regulation in response to biotic stress. Plant Stress 2023, 8. [Google Scholar] [CrossRef]
- Bürger, M.; Chory, J. Stressed Out About Hormones: How Plants Orchestrate Immunity. Cell Host & Microbe 2019, 26, 163–172. [Google Scholar] [CrossRef]
- Aminifard, M.H.; Jorkesh, A.; Fallahi, H.-R.; Moslemi, F.S. Influences of benzyl adenine and salicylic acid and on growth, yield, and biochemical characteristics of coriander (Coriandrum sativum L.). South African Journal of Botany 2020, 132, 299–303. [Google Scholar] [CrossRef]
- Antonić, D.; Milošević, S.; Cingel, A.; Lojić, M.; Trifunović-Momčilov, M.; Petrić, M.; Subotić, A.; Simonović, A. Effects of exogenous salicylic acid on Impatiens walleriana L. grown in vitro under polyethylene glycol-imposed drought. South African Journal of Botany 2016, 105, 226–233. [Google Scholar] [CrossRef]
- Attaran, E.; He, S.Y. The Long-Sought-After Salicylic Acid Receptors. Molecular Plant 2012, 5, 971–973. [Google Scholar] [CrossRef]
- Alam, M.U.; Fujita, M.; Nahar, K.; Rahman, A.; Anee, T.I.; Masud, A.A.C.; Amin, A.K.M.R.; Hasanuzzaman, M. Seed priming upregulates antioxidant defense and glyoxalase systems to conferring simulated drought tolerance in wheat seedlings. Plant Stress 2022, 6. [Google Scholar] [CrossRef]
- Chen, C.-l.; Yuan, F.; Li, X.-y.; Ma, R.-c.; Xie, H. Jasmonic acid and ethylene signaling pathways participate in the defense response of Chinese cabbage to Pectobacterium carotovorum infection. Journal of Integrative Agriculture 2021, 20, 1314–1326. [Google Scholar] [CrossRef]
- Cui, H.; Qiu, J.; Zhou, Y.; Bhandari, D.D.; Zhao, C.; Bautor, J.; Parker, J.E. Antagonism of Transcription Factor MYC2 by EDS1/PAD4 Complexes Bolsters Salicylic Acid Defense in Arabidopsis Effector-Triggered Immunity. Molecular Plant 2018, 11, 1053–1066. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Coronado, M.; Dias Araujo, P.C.; Ip, P.-L.; Nunes, C.O.; Rahni, R.; Wudick, M.M.; Lizzio, M.A.; Feijó, J.A.; Birnbaum, K.D. Plant glutamate receptors mediate a bet-hedging strategy between regeneration and defense. Developmental Cell 2022, 57, 451–465.e456. [Google Scholar] [CrossRef]
- Gawroński, P.; Witoń, D.; Vashutina, K.; Bederska, M.; Betliński, B.; Rusaczonek, A.; Karpiński, S. Mitogen-Activated Protein Kinase 4 Is a Salicylic Acid-Independent Regulator of Growth But Not of Photosynthesis in Arabidopsis. Molecular Plant 2014, 7, 1151–1166. [Google Scholar] [CrossRef]
- Nazar, R.; Umar, S.; Khan, N.A.; Sareer, O. Salicylic acid supplementation improves photosynthesis and growth in mustard through changes in proline accumulation and ethylene formation under drought stress. South African Journal of Botany 2015, 98, 84–94. [Google Scholar] [CrossRef]
- Atia, M.A.M.; Abdeldaym, E.A.; Abdelsattar, M.; Ibrahim, D.S.S.; Saleh, I.; Elwahab, M.A.; Osman, G.H.; Arif, I.A.; Abdelaziz, M.E. Piriformospora indica promotes cucumber tolerance against Root-knot nematode by modulating photosynthesis and innate responsive genes. Saudi Journal of Biological Sciences 2020, 27, 279–287. [Google Scholar] [CrossRef]
- Khalvandi, M.; Siosemardeh, A.; Roohi, E.; Keramati, S. Salicylic acid alleviated the effect of drought stress on photosynthetic characteristics and leaf protein pattern in winter wheat. Heliyon 2021, 7. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Jahan, B.; AlAjmi, M.F.; Rehman, M.T.; Iqbal, N.; Irfan, M.; Sehar, Z.; Khan, N.A. Crosstalk of plant growth regulators protects photosynthetic performance from arsenic damage by modulating defense systems in rice. Ecotoxicology and Environmental Safety 2021, 222. [Google Scholar] [CrossRef]
- Li, X.; Riaz, M.; Song, B.; Liang, X.; Liu, H. Exogenous salicylic acid alleviates fomesafen toxicity by improving photosynthetic characteristics and antioxidant defense system in sugar beet. Ecotoxicology and Environmental Safety 2022, 238. [Google Scholar] [CrossRef]
- Li, Y.; Han, X.; Ren, H.; Zhao, B.; Zhang, J.; Ren, B.; Gao, H.; Liu, P. Exogenous SA or 6-BA maintains photosynthetic activity in maize leaves under high temperature stress. The Crop Journal 2023, 11, 605–617. [Google Scholar] [CrossRef]
- Sherin, G.; Aswathi, K.P.R.; Puthur, J.T. Photosynthetic functions in plants subjected to stresses are positively influenced by priming. Plant Stress 2022, 4. [Google Scholar] [CrossRef]
- Gondor, O.K.; Pál, M.; Janda, T.; Szalai, G. The role of methyl salicylate in plant growth under stress conditions. Journal of Plant Physiology 2022, 277. [Google Scholar] [CrossRef] [PubMed]
- Hundare, A.; Joshi, V.; Joshi, N. Salicylic acid attenuates salinity-induced growth inhibition in in vitro raised ginger (Zingiber officinale Roscoe) plantlets by regulating ionic balance and antioxidative system. Plant Stress 2022, 4. [Google Scholar] [CrossRef]
- Liu, N.; Xu, Y.; Li, Q.; Cao, Y.; Yang, D.; Liu, S.; Wang, X.; Mi, Y.; Liu, Y.; Ding, C.; et al. A lncRNA fine-tunes salicylic acid biosynthesis to balance plant immunity and growth. Cell Host & Microbe 2022, 30, 1124–1138.e1128. [Google Scholar] [CrossRef]
- Lone, R.; Hassan, N.; Bashir, B.; Rohela, G.K.; Malla, N.A. Role of growth elicitors and microbes in stress management and sustainable production of Sorghum. Plant Stress 2023, 9. [Google Scholar] [CrossRef]
- Mogal, C.S.; Solanki, V.H.; Kansara, R.V.; Jha, S.; Singh, S.; Parekh, V.B.; Rajkumar, B.K. UHPLC-MS/MS and QRT-PCR profiling of PGP agents and Rhizobium spp. of induced phytohormones for growth promotion in mungbean (var. Co4). Heliyon 2022, 8. [Google Scholar] [CrossRef]
- Tan, S.; Abas, M.; Verstraeten, I.; Glanc, M.; Molnár, G.; Hajný, J.; Lasák, P.; Petřík, I.; Russinova, E.; Petrášek, J.; et al. Salicylic Acid Targets Protein Phosphatase 2A to Attenuate Growth in Plants. Current Biology 2020, 30, 381–395.e388. [Google Scholar] [CrossRef]
- Zahid, A.; Yike, G.; Kubik, S.; Fozia; Ramzan, M. ; Sardar, H.; Akram, M.T.; Khatana, M.A.; Shabbir, S.; Alharbi, S.A.; et al. Plant growth regulators modulate the growth, physiology, and flower quality in rose (Rosa hybirda). Journal of King Saud University - Science 2021, 33. [Google Scholar] [CrossRef]
- Zhi, J.-H.; Zhang, D.; Huang, W.-J.; Chen, Q.-L.; Wang, J.-J.; Zheng, Q.-Q. Effects of compound growth regulators on the anatomy of Jujube Leaf and Fruit. Saudi Journal of Biological Sciences 2020, 27, 3660–3668. [Google Scholar] [CrossRef]
- Song, W.Y.; Peng, S.P.; Shao, C.Y.; Shao, H.B.; Yang, H.C. Ethylene glycol tetra-acetic acid and salicylic acid improve anti-oxidative ability of maize seedling leaves under heavy-metal and polyethylene glycol 6000-simulated drought stress. Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology 2014, 148, 96–108. [Google Scholar] [CrossRef]
- Tanveer, S.; Akhtar, N.; Ilyas, N.; Sayyed, R.Z.; Fitriatin, B.N.; Perveen, K.; Bukhari, N.A. Interactive effects of Pseudomonas putida and salicylic acid for mitigating drought tolerance in canola (Brassica napus L.). Heliyon 2023, 9. [Google Scholar] [CrossRef] [PubMed]
- Das, D.; Bisht, K.; Chauhan, A.; Gautam, S.; Jaiswal, J.P.; Salvi, P.; Lohani, P. Morpho-physiological and biochemical responses in wheat foliar sprayed with zinc-chitosan-salicylic acid nanoparticles during drought stress. Plant Nano Biology 2023, 4. [Google Scholar] [CrossRef]
- Malko, M.M.; Khanzada, A.; Wang, X.; Samo, A.; Li, Q.; Jiang, D.; Cai, J. Chemical treatment refines drought tolerance in wheat and its implications in changing climate: A review. Plant Stress 2022, 6. [Google Scholar] [CrossRef]
- Singh, P.K.; Indoliya, Y.; Agrawal, L.; Awasthi, S.; Deeba, F.; Dwivedi, S.; Chakrabarty, D.; Shirke, P.A.; Pandey, V.; Singh, N.; et al. Genomic and proteomic responses to drought stress and biotechnological interventions for enhanced drought tolerance in plants. Current Plant Biology 2022, 29. [Google Scholar] [CrossRef]
- Balfagón, D.; Rambla, J.L.; Granell, A.; Arbona, V.; Gómez-Cadenas, A. Grafting improves tolerance to combined drought and heat stresses by modifying metabolism in citrus scion. Environmental and Experimental Botany 2022, 195. [Google Scholar] [CrossRef]
- Seo, J.; Lee, J.G.; Kang, B.-C.; Lim, S.; Lee, E.J. Indole-3-acetic acid, a hormone potentially involved in chilling-induced seed browning of pepper (Capsicum annuum L.) fruit during cold storage. Postharvest Biology and Technology 2023, 199. [Google Scholar] [CrossRef]
- Raza, A.; Charagh, S.; Najafi-Kakavand, S.; Abbas, S.; Shoaib, Y.; Anwar, S.; Sharifi, S.; Lu, G.; Siddique, K.H.M. Role of phytohormones in regulating cold stress tolerance: Physiological and molecular approaches for developing cold-smart crop plants. Plant Stress 2023, 8. [Google Scholar] [CrossRef]
- Lafuente, M.T.; Romero, P. Hormone profiling and heat-induced tolerance to cold stress in citrus fruit. Postharvest Biology and Technology 2022, 194. [Google Scholar] [CrossRef]
- Wei, Y.; Zhu, B.; Liu, W.; Cheng, X.; Lin, D.; He, C.; Shi, H. Heat shock protein 90 co-chaperone modules fine-tune the antagonistic interaction between salicylic acid and auxin biosynthesis in cassava. Cell Reports 2021, 34. [Google Scholar] [CrossRef]
- Afzal, I.; Akram, M.W.; Rehman, H.U.; Rashid, S.; Basra, S.M.A. Moringa leaf and sorghum water extracts and salicylic acid to alleviate impacts of heat stress in wheat. South African Journal of Botany 2020, 129, 169–174. [Google Scholar] [CrossRef]
- Rasheed, F.; Mir, I.R.; Sehar, Z.; Fatma, M.; Gautam, H.; Khan, S.; Anjum, N.A.; Masood, A.; Sofo, A.; Khan, N.A. Nitric Oxide and Salicylic Acid Regulate Glutathione and Ethylene Production to Enhance Heat Stress Acclimation in Wheat Involving Sulfur Assimilation. Plants (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Khan, M.N.; Yan, J.; Hong, X.; Zhao, F.; Chen, L.; Ma, H.; Li, Y.; Li, J.; Wu, H. Mechanisms of nanomaterials for improving plant salt tolerance. Crop and Environment 2023, 2, 92–99. [Google Scholar] [CrossRef]
- Kumar, S.; Abass Ahanger, M.; Alshaya, H.; Latief Jan, B.; Yerramilli, V. Salicylic acid mitigates salt induced toxicity through the modifications of biochemical attributes and some key antioxidants in capsicum annuum. Saudi Journal of Biological Sciences 2022, 29, 1337–1347. [Google Scholar] [CrossRef] [PubMed]
- Shaki, F.; Maboud, H.E.; Niknam, V. Growth enhancement and salt tolerance of Safflower ( Carthamus tinctorius L.), by salicylic acid. Current Plant Biology 2018, 13, 16–22. [Google Scholar] [CrossRef]
- Jini, D.; Joseph, B. Physiological Mechanism of Salicylic Acid for Alleviation of Salt Stress in Rice. Rice Science 2017, 24, 97–108. [Google Scholar] [CrossRef]
- Ali, Q.; Ahmad, M.; Kamran, M.; Ashraf, S.; Shabaan, M.; Babar, B.H.; Zulfiqar, U.; Haider, F.U.; Ali, M.A.; Elshikh, M.S. Synergistic Effects of Rhizobacteria and Salicylic Acid on Maize Salt-Stress Tolerance. Plants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Mutlu-Durak, H.; Arikan, Y.; Kutman, B.Y. Willow (Salix babylonica) Extracts Can Act as Biostimulants for Enhancing Salinity Tolerance of Maize Grown in Soilless Culture. Plants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Ur Rahman, S.; Li, Y.; Hussain, S.; Hussain, B.; Khan, W.-u.-D.; Riaz, L.; Nadeem Ashraf, M.; Athar Khaliq, M.; Du, Z.; Cheng, H. Role of phytohormones in heavy metal tolerance in plants: A review. Ecological Indicators 2023, 146. [Google Scholar] [CrossRef]
- Khalil, R.; Haroun, S.; Bassyoini, F.; Nagah, A.; Yusuf, M. Salicylic acid in combination with kinetin or calcium ameliorates heavy metal stress in Phaseolus vulgaris plant. Journal of Agriculture and Food Research 2021, 5. [Google Scholar] [CrossRef]
- Saini, S.; Kaur, N.; Pati, P.K. Phytohormones: Key players in the modulation of heavy metal stress tolerance in plants. Ecotoxicology and Environmental Safety 2021, 223. [Google Scholar] [CrossRef] [PubMed]
- Song, W.-Y.; Yang, H.-C.; Shao, H.-B.; Zheng, A.-Z.; Brestic, M. The Alleviative Effects of Salicylic Acid on the Activities of Catalase and Superoxide Dismutase in Malting Barley (Hordeum uhulgareL.) Seedling Leaves Stressed by Heavy Metals. CLEAN - Soil, Air, Water 2014, 42, 88–97. [Google Scholar] [CrossRef]
- Hernanz, D.; Jara-Palacios, M.J.; Santos, J.L.; Gómez Pajuelo, A.; Heredia, F.J.; Terrab, A. The profile of phenolic compounds by HPLC-MS in Spanish oak (Quercus) honeydew honey and their relationships with color and antioxidant activity. Lwt 2023, 180. [Google Scholar] [CrossRef]
- Singh, S. Salicylic acid elicitation improves antioxidant activity of spinach leaves by increasing phenolic content and enzyme levels. Food Chemistry Advances 2023, 2. [Google Scholar] [CrossRef]
- Aldhanhani, A.R.H.; Ahmed, Z.F.R.; Tzortzakis, N.; Singh, Z. Maturity stage at harvest influences antioxidant phytochemicals and antibacterial activity of jujube fruit (Ziziphus mauritiana Lamk. and Ziziphus spina-christi L.). Annals of Agricultural Sciences 2022, 67, 196–203. [Google Scholar] [CrossRef]
- Dey, A.; Nandy, S.; Nongdam, P.; Tikendra, L.; Mukherjee, A.; Mukherjee, S.; Pandey, D.K. Methyl jasmonate and salicylic acid elicit indole alkaloid production and modulate antioxidant defence and biocidal properties in Rauvolfia serpentina Benth. ex Kurz. in vitro cultures. South African Journal of Botany 2020, 135, 1–17. [Google Scholar] [CrossRef]
- Nivedha, K.; Sivasakthi, S.; Prakash, A.; Devipriya, N.; Vadivel, V. In vitro studies on antioxidant and cyto-protective activities of polyphenol-rich fraction isolated from Mangifera indica leaf. South African Journal of Botany 2020, 130, 396–406. [Google Scholar] [CrossRef]
- Sytar, O. Phenolic acids in the inflorescences of different varieties of buckwheat and their antioxidant activity. Journal of King Saud University - Science 2015, 27, 136–142. [Google Scholar] [CrossRef]
- Ali, A.M.A.; El-Nour, M.E.M.; Yagi, S.M. Total phenolic and flavonoid contents and antioxidant activity of ginger (Zingiber officinale Rosc.) rhizome, callus and callus treated with some elicitors. Journal of Genetic Engineering and Biotechnology 2018, 16, 677–682. [Google Scholar] [CrossRef]
- Vañó, M.S.; Nourimand, M.; MacLean, A.; Pérez-López, E. Getting to the root of a club – Understanding developmental manipulation by the clubroot pathogen. Seminars in Cell & Developmental Biology 2023, 148-149, 22–32. [Google Scholar] [CrossRef]
- Musaqaf, N.; Lyngs Jørgensen, H.J.; Sigsgaard, L. Plant resistance induced by hemipterans — Effects on insect herbivores and pathogens. Crop Protection 2023, 163. [Google Scholar] [CrossRef]
- Poveda, J.; Rodríguez, V.M.; Abilleira, R.; Velasco, P. Trichoderma hamatum can act as an inter-plant communicator of foliar pathogen infections by colonizing the roots of nearby plants: A new inter-plant “wired communication”. Plant Science 2023, 330. [Google Scholar] [CrossRef] [PubMed]
- Medina-Puche, L.; Tan, H.; Dogra, V.; Wu, M.; Rosas-Diaz, T.; Wang, L.; Ding, X.; Zhang, D.; Fu, X.; Kim, C.; et al. A Defense Pathway Linking Plasma Membrane and Chloroplasts and Co-opted by Pathogens. Cell 2020, 182, 1109–1124.e1125. [Google Scholar] [CrossRef] [PubMed]
- Qi, G.; Chen, J.; Chang, M.; Chen, H.; Hall, K.; Korin, J.; Liu, F.; Wang, D.; Fu, Z.Q. Pandemonium Breaks Out: Disruption of Salicylic Acid-Mediated Defense by Plant Pathogens. Molecular Plant 2018, 11, 1427–1439. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-d.; Bi, W.-s.; Gao, J.; Yu, X.-m.; Wang, H.-y.; Liu, D.-q. Systemic acquired resistance, NPR1, and pathogenesis-related genes in wheat and barley. Journal of Integrative Agriculture 2018, 17, 2468–2477. [Google Scholar] [CrossRef]
- Wang, X.; Fu, M.; Qu, X.; Liu, J.; Bu, J.; Feng, S.; Zhao, H.; Jiao, W.; Sun, F. (E)-2-Hexenal-based coating induced acquired resistance in apple and its antifungal effects against Penicillium expansum. Lwt 2022, 163. [Google Scholar] [CrossRef]
- Thepbandit, W.; Papathoti, N.K.; Daddam, J.R.; Hoang, N.H.; Le Thanh, T.; Saengchan, C.; Buensanteai, K. In vitro and in silico studies of salicylic acid on systemic induced resistance against bacterial leaf blight disease and enhancement of crop yield. Journal of Integrative Agriculture 2023, 22, 170–184. [Google Scholar] [CrossRef]
- Horvath, D.P.; Clay, S.A.; Swanton, C.J.; Anderson, J.V.; Chao, W.S. Weed-induced crop yield loss: a new paradigm and new challenges. Trends in Plant Science 2023, 28, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Yadav, T.; Kumar, A.; Yadav, R.K.; Yadav, G.; Kumar, R.; Kushwaha, M. Salicylic acid and thiourea mitigate the salinity and drought stress on physiological traits governing yield in pearl millet- wheat. Saudi Journal of Biological Sciences 2020, 27, 2010–2017. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Larruscain, D.; Krüger, M.; Abeyawardana, O.A.J.; Belz, C.; Dobrev, P.I.; Vaňková, R.; Eliášová, K.; Vondráková, Z.; Juříček, M.; Štorchová, H. The high concentrations of abscisic, jasmonic, and salicylic acids produced under long days do not accelerate flowering in Chenopodium ficifolium 459. Plant Science 2022, 320. [Google Scholar] [CrossRef]
- Guo, P.R.; Wu, L.L.; Wang, Y.; Liu, D.; Li, J.A. Effects of Drought Stress on the Morphological Structure and Flower Organ Physiological Characteristics of Camellia oleifera Flower Buds. Plants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Sedláková, V.; Zeljković, S.; Štefelová, N.; Smýkal, P.; Hanáček, P. Phenylpropanoid Content of Chickpea Seed Coats in Relation to Seed Dormancy. Plants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Li, M.; Wang, M.; Chen, J.; Wu, J.; Xia, Z. Sulfur dioxide improves the thermotolerance of maize seedlings by regulating salicylic acid biosynthesis. Ecotoxicology and Environmental Safety 2023, 254. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Tian, X.; Wang, W.; Xu, X.; Su, Y.; Yang, J.; Duan, S.; Li, J.; Xin, M.; Peng, H.; et al. Changes in concentrations and transcripts of plant hormones in wheat seedling roots in response to Fusarium crown rot. The Crop Journal 2023. [Google Scholar] [CrossRef]
- Rehman, H.u.; Iqbal, H.; Basra, S.M.A.; Afzal, I.; Farooq, M.; Wakeel, A.; Wang, N. Seed priming improves early seedling vigor, growth and productivity of spring maize. Journal of Integrative Agriculture 2015, 14, 1745–1754. [Google Scholar] [CrossRef]
- Kotapati, K.V.; Palaka, B.K.; Ampasala, D.R. Alleviation of nickel toxicity in finger millet ( Eleusine coracana L.) germinating seedlings by exogenous application of salicylic acid and nitric oxide. The Crop Journal 2017, 5, 240–250. [Google Scholar] [CrossRef]
- Apon, T.A.; Ahmed, S.F.; Bony, Z.F.; Chowdhury, M.R.; Asha, J.F.; Biswas, A. Sett priming with salicylic acid improves salinity tolerance of sugarcane (Saccharum officinarum L.) during early stages of crop development. Heliyon 2023, 9. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Wang, Y.; Li, X.; Zhang, Y. Biosynthesis and Regulation of Salicylic Acid and N-Hydroxypipecolic Acid in Plant Immunity. Molecular Plant 2020, 13, 31–41. [Google Scholar] [CrossRef]
- Ding, Y.; Sun, T.; Ao, K.; Peng, Y.; Zhang, Y.; Li, X.; Zhang, Y. Opposite Roles of Salicylic Acid Receptors NPR1 and NPR3/NPR4 in Transcriptional Regulation of Plant Immunity. Cell 2018, 173, 1454–1467.e1415. [Google Scholar] [CrossRef]
- Chen, H.; Chen, J.; Li, M.; Chang, M.; Xu, K.; Shang, Z.; Zhao, Y.; Palmer, I.; Zhang, Y.; McGill, J.; et al. A Bacterial Type III Effector Targets the Master Regulator of Salicylic Acid Signaling, NPR1, to Subvert Plant Immunity. Cell Host & Microbe 2017, 22, 777–788.e777. [Google Scholar] [CrossRef]
- Anaya, F.; Fghire, R.; Wahbi, S.; Loutfi, K. Influence of salicylic acid on seed germination of Vicia faba L. under salt stress. Journal of the Saudi Society of Agricultural Sciences 2018, 17, 1–8. [Google Scholar] [CrossRef]
- Talaat, N.B.; Hanafy, A.M.A. Spermine-Salicylic Acid Interplay Restrains Salt Toxicity in Wheat (Triticum aestivum L.). Plants (Basel) 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.H.; Adhikari, A.; Imran, M.; Lee, D.S.; Lee, C.Y.; Kang, S.M.; Lee, I.J. Exogenous SA Applications Alleviate Salinity Stress via Physiological and Biochemical changes in St John's Wort Plants. Plants (Basel) 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Talaat, N.B.; Hanafy, A.M.A. Plant Growth Stimulators Improve Two Wheat Cultivars Salt-Tolerance: Insights into Their Physiological and Nutritional Responses. Plants (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Munsif, F.; Shah, T.; Arif, M.; Jehangir, M.; Afridi, M.Z.; Ahmad, I.; Jan, B.L.; Alansi, S. Combined effect of salicylic acid and potassium mitigates drought stress through the modulation of physio-biochemical attributes and key antioxidants in wheat. Saudi Journal of Biological Sciences 2022, 29. [Google Scholar] [CrossRef]
- Alotaibi, M.; El-Hendawy, S.; Mohammed, N.; Alsamin, B.; Al-Suhaibani, N.; Refay, Y. Effects of Salicylic Acid and Macro- and Micronutrients through Foliar and Soil Applications on the Agronomic Performance, Physiological Attributes, and Water Productivity of Wheat under Normal and Limited Irrigation in Dry Climatic Conditions. Plants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Duvnjak, J.; Lončarić, A.; Brkljačić, L.; Šamec, D.; Šarčević, H.; Salopek-Sondi, B.; Španić, V. Morpho-Physiological and Hormonal Response of Winter Wheat Varieties to Drought Stress at Stem Elongation and Anthesis Stages. Plants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, Z.; Luo, R.; Sun, Y.; Yang, C.; Li, X.; Gao, A.; Pu, J. Genome-Wide Characterization, Identification and Expression Profile of MYB Transcription Factor Gene Family during Abiotic and Biotic Stresses in Mango (Mangifera indica). Plants (Basel) 2022, 11. [Google Scholar] [CrossRef]
- Wang, W.-M.; Ma, X.-F.; Zhang, Y.; Luo, M.-C.; Wang, G.-L.; Bellizzi, M.; Xiong, X.-Y.; Xiao, S.-Y. PAPP2C Interacts with the Atypical Disease Resistance Protein RPW8.2 and Negatively Regulates Salicylic Acid-Dependent Defense Responses in Arabidopsis. Molecular Plant 2012, 5, 1125–1137. [Google Scholar] [CrossRef]
- Jia, H.; Wang, X.; Wei, T.; Wang, M.; Liu, X.; Hua, L.; Ren, X.; Guo, J.; Li, J. Exogenous salicylic acid regulates cell wall polysaccharides synthesis and pectin methylation to reduce Cd accumulation of tomato. Ecotoxicology and Environmental Safety 2021, 207. [Google Scholar] [CrossRef]
- Upadhyay, R.K.; Motyka, V.; Pokorna, E.; Dobrev, P.I.; Lacek, J.; Shao, J.; Lewers, K.S.; Mattoo, A.K. Comprehensive profiling of endogenous phytohormones and expression analysis of 1-aminocyclopropane-1-carboxylic acid synthase gene family during fruit development and ripening in octoploid strawberry (Fragaria× ananassa). Plant Physiology and Biochemistry 2023, 196, 186–196. [Google Scholar] [CrossRef]
- Świeżawska-Boniecka, B.; Szmidt-Jaworska, A. Phytohormones and cyclic nucleotides - Long-awaited couples? Journal of Plant Physiology 2023, 286. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Niu, G.; Fan, Y.; Liu, W.; Wu, Q.; Yu, C.; Wang, J.; Xiao, Y.; Hou, L.; Jin, D.; et al. Synthetic dual hormone-responsive promoters enable engineering of plants with broad-spectrum resistance. Plant Communications 2023, 4. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Ahmed, N.; Saha, T.; Rahman, M.; Rahman, K.; Alam, M.M.; Rohman, M.M.; Nahar, K. Exogenous salicylic acid and kinetin modulate reactive oxygen species metabolism and glyoxalase system to confer waterlogging stress tolerance in soybean (Glycine max L.). Plant Stress 2022, 3. [Google Scholar] [CrossRef]
- Widhalm, Joshua R. ; Dudareva, N. A Familiar Ring to It: Biosynthesis of Plant Benzoic Acids. Molecular Plant 2015, 8, 83–97. [Google Scholar] [CrossRef]
- Lihavainen, J.; Šimura, J.; Bag, P.; Fataftah, N.; Robinson, K.M.; Delhomme, N.; Novák, O.; Ljung, K.; Jansson, S. Salicylic acid metabolism and signalling coordinate senescence initiation in aspen in nature. Nature communications 2023, 14, 4288. [Google Scholar] [CrossRef]
- Arruda, T.F.L.; Lima, G.S.; Silva, A.; Azevedo, C.A.V.; Souza, A.R.; Soares, L.; Gheyi, H.R.; Lima, V.L.A.; Fernandes, P.D.; Silva, F.A.D.; et al. Salicylic Acid as a Salt Stress Mitigator on Chlorophyll Fluorescence, Photosynthetic Pigments, and Growth of Precocious-Dwarf Cashew in the Post-Grafting Phase. Plants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Moustakas, M.; Sperdouli, I.; Moustaka, J.; Şaş, B.; İşgören, S.; Morales, F. Mechanistic Insights on Salicylic Acid Mediated Enhancement of Photosystem II Function in Oregano Seedlings Subjected to Moderate Drought Stress. Plants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Ullah, C.; Chen, Y.-H.; Ortega, M.A.; Tsai, C.-J. The diversity of salicylic acid biosynthesis and defense signaling in plants: Knowledge gaps and future opportunities. Current Opinion in Plant Biology 2023, 72. [Google Scholar] [CrossRef]
- Rekhter, D.; Lüdke, D.; Ding, Y.; Feussner, K.; Zienkiewicz, K.; Lipka, V.; Wiermer, M.; Zhang, Y.; Feussner, I. Isochorismate-derived biosynthesis of the plant stress hormone salicylic acid. Science 2019, 365, 498–502. [Google Scholar] [CrossRef]
- Lefevere, H.; Bauters, L.; Gheysen, G. Salicylic Acid Biosynthesis in Plants. Front Plant Sci 2020, 11, 338. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yang, J.; Li, X.; Zhang, Y. Salicylic Acid: Biosynthesis and Signaling. Annu Rev Plant Biol 2021, 72, 761–791. [Google Scholar] [CrossRef]
- Reglinski, T.; Vanneste, J.L.; Schipper, M.M.; Cornish, D.A.; Yu, J.; Oldham, J.M.; Fehlmann, C.; Parry, F.; Hedderley, D. Postharvest Application of Acibenzolar-S-Methyl Activates Salicylic Acid Pathway Genes in Kiwifruit Vines. Plants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Zhu, F.; Cao, M.-Y.; Zhang, Q.-P.; Mohan, R.; Schar, J.; Mitchell, M.; Chen, H.; Liu, F.; Wang, D.; Fu, Z.Q. Join the green team: Inducers of plant immunity in the plant disease sustainable control toolbox. Journal of Advanced Research 2023. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Dong, X. A Highway for War and Peace: The Secretory Pathway in Plant–Microbe Interactions. Molecular Plant 2011, 4, 581–587. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.d.C.; Fadiji, A.E.; Babalola, O.O.; Santoyo, G. Bacterial elicitors of the plant immune system: An overview and the way forward. Plant Stress 2023, 7. [Google Scholar] [CrossRef]
- Kgang, I.E.; Klein, A.; Husselmann, L.; Nkomo, A.; Mathabe, P.M.K.; Belay, Z.A.; Caleb, O.J. Bioassays and proteomics as early detection tools in postharvest management of table grapes (Vitis vinifera L.) diseases – A Review. Food Bioscience 2023, 53. [Google Scholar] [CrossRef]
- Tripathi, D.; Raikhy, G.; Kumar, D. Chemical elicitors of systemic acquired resistance—Salicylic acid and its functional analogs. Current Plant Biology 2019, 17, 48–59. [Google Scholar] [CrossRef]
- Tripathi, G.D.; Javed, Z.; Mishra, M.; Fasake, V.; Dashora, K. Phytomelatonin in stress management in agriculture. Heliyon 2021, 7. [Google Scholar] [CrossRef]
- Villadangos, S.; Munné-Bosch, S. Acclimation to a combination of water deficit and nutrient deprivation through simultaneous increases in abscisic acid and bioactive jasmonates in the succulent plant Sempervivum tectorum L. Journal of Plant Physiology 2023, 287. [Google Scholar] [CrossRef]
- Kim, J.H.; Castroverde, C.D.M.; Huang, S.; Li, C.; Hilleary, R.; Seroka, A.; Sohrabi, R.; Medina-Yerena, D.; Huot, B.; Wang, J.; et al. Increasing the resilience of plant immunity to a warming climate. Nature 2022, 607, 339–344. [Google Scholar] [CrossRef]
- Borrowman, S.; Kapuganti, J.G.; Loake, G.J. Expanding roles for S-nitrosylation in the regulation of plant immunity. Free Radical Biology and Medicine 2023, 194, 357–368. [Google Scholar] [CrossRef]
- Maruri-Lopez, I.; Chodasiewicz, M. Involvement of small molecules and metabolites in regulation of biomolecular condensate properties. Current Opinion in Plant Biology 2023, 74. [Google Scholar] [CrossRef]
- Acevedo, A.F.G.; Lacerda, V.R.; da Silva Gomes, J.W.; Avilez, A.A.; Sarria, S.D.; Broetto, F.; Vieites, R.L.; de Souza Guimarães, M.L.C. Foliar salicylic acid application to mitigate the effect of water deficiency on potato (Solanum tuberosum L.). Plant Stress 2023, 7. [Google Scholar] [CrossRef]
- Xu, L.; Zhao, H.; Wang, J.; Wang, X.; Jia, X.; Wang, L.; Xu, Z.; Li, R.; Jiang, K.; Chen, Z.; et al. AIM1-dependent high basal salicylic acid accumulation modulates stomatal aperture in rice. The New phytologist 2023. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, Z.; Huang, L.; Ouyang, Y.; Wang, Z.; Wu, S.; Ye, W.; Yu, B.; Zhang, Y.; Yang, C.; et al. Salicylic acid attenuates brassinosteroid signaling via protein de-S-acylation. The EMBO journal 2023, e112998. [Google Scholar] [CrossRef] [PubMed]
- Fàbregas, N.; Fernie, A.R. The interface of central metabolism with hormone signaling in plants. Current Biology 2021, 31, R1535–R1548. [Google Scholar] [CrossRef]
- Wang, X.; Guo, H.; Jin, Z.; Ding, Y.; Guo, M. Comprehensive Characterization of B-Box Zinc Finger Genes in Citrullus lanatus and Their Response to Hormone and Abiotic Stresses. Plants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Luo, J.; Yu, W.; Xiao, Y.; Zhang, Y.; Peng, F. FaSnRK1α mediates salicylic acid pathways to enhance strawberry resistance to Botrytis cinerea. Horticultural Plant Journal 2023. [Google Scholar] [CrossRef]
- Liu, J.; Xu, X.; Zhou, J.; Sun, G.; Li, Z.; Zhai, L.; Wang, J.; Ma, R.; Zhao, D.; Jiang, R.; et al. Phenolic acids in Panax ginseng inhibit melanin production through bidirectional regulation of melanin synthase transcription via different signaling pathways. Journal of Ginseng Research 2023. [Google Scholar] [CrossRef]
- Hui, S.; Ke, Y.; Chen, D.; Wang, L.; Li, Q.; Yuan, M. Rice microRNA156/529-SQUAMOSA PROMOTER BINDING PROTEIN-LIKE7/14/17 modules regulate defenses against bacteria. Plant physiology 2023. [Google Scholar] [CrossRef]
- Kaizhuan, X.; Xiaohui, M.; Yingheng, W.; Jinlan, W.; Yidong, W.; Qiuhua, C.; Hua’an, X.; Jianfu, Z. Transcript Profiling Reveals Abscisic Acid, Salicylic Acid and Jasmonic-Isoleucine Pathways Involved in High Regenerative Capacities of Immature Embryos Compared with Mature Seeds in japonica Rice. Rice Science 2018, 25, 227–234. [Google Scholar] [CrossRef]
- Nutricati, E.; De Pascali, M.; Negro, C.; Bianco, P.A.; Quaglino, F.; Passera, A.; Pierro, R.; Marcone, C.; Panattoni, A.; Sabella, E.; et al. Signaling Cross-Talk between Salicylic and Gentisic Acid in the 'Candidatus Phytoplasma Solani' Interaction with Sangiovese Vines. Plants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Lee, S.Y.; Damodaran, P.N.; Roh, K.S. Influence of salicylic acid on rubisco and rubisco activase in tobacco plant grown under sodium chloride in vitro. Saudi Journal of Biological Sciences 2014, 21, 417–426. [Google Scholar] [CrossRef]
- Hussain, B.; War, A.R.; Pfeiffer, D.G. Jasmonic acid and salicylic acid induced defensive response in wine grapes against Drosophila suzukii (Diptera: Drosophilidae). Heliyon 2023, 9. [Google Scholar] [CrossRef]
- ul Haq, A.; Lone, M.L.; Farooq, S.; Parveen, S.; Altaf, F.; Tahir, I.; Kaushik, P.; El-Serehy, H.A. Efficacy of salicylic acid in modulating physiological andbiochemical mechanisms to improve postharvest longevity in cut spikes of Consolida ajacis (L.) Schur. Saudi Journal of Biological Sciences 2022, 29, 713–720. [Google Scholar] [CrossRef]
- Pyo, Y.; Moon, H.; Nugroho, A.B.D.; Yang, S.W.; Jung, I.L.; Kim, D.-H. Transcriptome analysis revealed that jasmonic acid biosynthesis/signaling is involved in plant response to Strontium stress. Ecotoxicology and Environmental Safety 2022, 237. [Google Scholar] [CrossRef]
- Kumar, A.; Giridhar, P. Salicylic acid and methyljasmonate restore the transcription of caffeine biosynthetic N-methyltransferases from a transcription inhibition noticed during late endosperm maturation in coffee. Plant Gene 2015, 4, 38–44. [Google Scholar] [CrossRef]
- Yang, H.; Fang, R.; Luo, L.; Yang, W.; Huang, Q.; Yang, C.; Hui, W.; Gong, W.; Wang, J. Uncovering the mechanisms of salicylic acid-mediated abiotic stress tolerance in horticultural crops. Frontiers in Plant Science 2023. [Google Scholar] [CrossRef]
Figure 1.
A diagram of salicylic acid’s roles in plant stress responses.
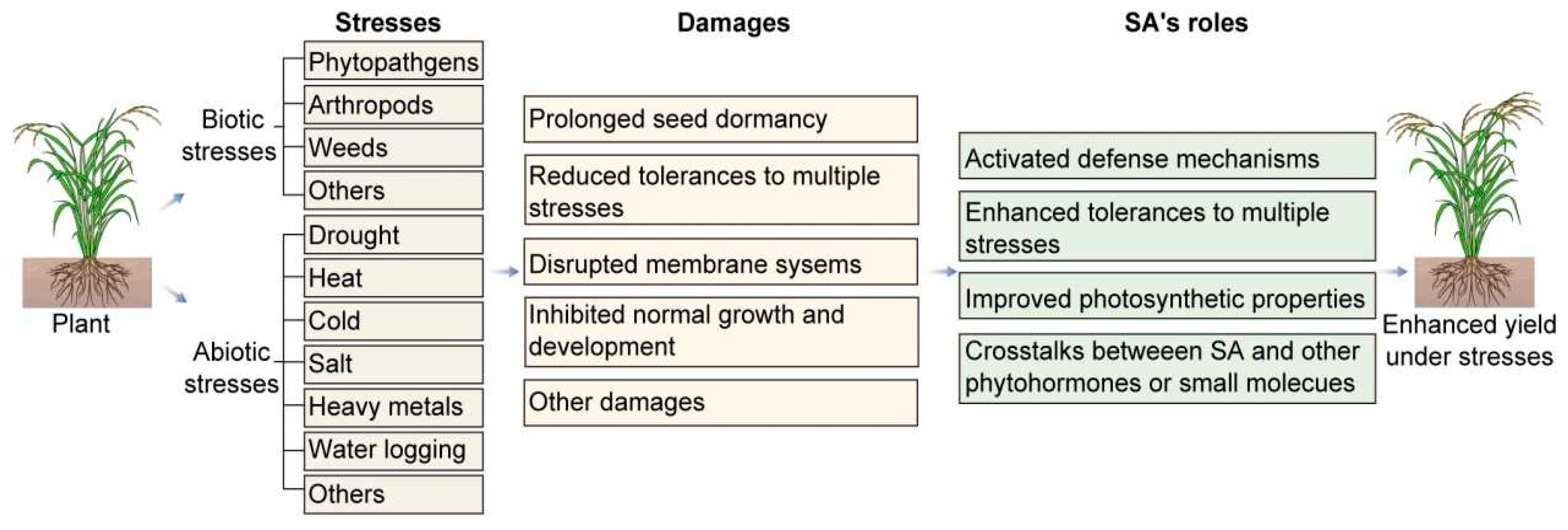
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



