
Submitted:
13 September 2023
Posted:
14 September 2023
Read the latest preprint version here
Abstract
Overexpression of human epidermal growth factor receptor 2 (HER2) in breast and gastric cancers is an important target for monoclonal antibody (mAb) therapy such as trastuzumab. All therapeutic mAbs, including anti-HER2 mAbs, exhibit adverse effects probably due to the recognition of antigens expressed in normal cells. Therefore, tumor-selective or specific mAbs have been desired to reduce adverse effects. In this study, we provide a strategy for the selection of cancer-specific mAb against HER2. We screened more than 200 of anti-HER2 mAbs obtained by our laboratory and established a novel cancer-specific anti-HER2 antibody, H2Mab-250 (IgG1, kappa). H2Mab-250 reacted with HER2-positive breast cancer BT-474 and SK-BR-3 cells. Importantly, H2Mab-250 never showed reactivity to non-transformed normal epithelial cells (HaCaT and MCF 10A) and immortalized normal epithelial cells in flow cytometry. In contrast, most anti-HER2 mAbs including H2Mab-119 (IgG1, kappa) reacted with both cancer and normal epithelial cells. The epitope mapping revealed that H2Mab-250 recognized the domain VI of HER2 and the Trp614 mainly contributes to the recognition by H2Mab-250. In immunohistochemical analysis, H2Mab-250 exhibited a superior reactivity to HER2-positive breast cancer section compared to H2Mab-119. Importantly, H2Mab-250 never showed any reactivity to normal tissues by immunohistochemical analysis. The strategy to select cancer-specific mAbs would contribute to the development of novel antibodies and modalities for cancer therapy.
Keywords:
HER2
; cancer-specific monoclonal antibody
; screening
; epitope
; flow cytometry
1. Introduction
Human epidermal growth factor receptor 2 (HER2) is included in the receptor tyrosine kinase family of human epidermal growth factor receptor (EGFR). To activate the downstream signaling, HER2 must either form heterodimers with other HER members and their specific ligands or self-assemble into ligand-independent homodimers when overexpressed [1]. The HER2 overexpression is observed in approximately 20% of breast cancers [2] and 20% of gastric cancers [3] associated with higher rates of recurrence, poor prognosis, and shorter overall survival. A monoclonal antibody (mAb) against HER2, trastuzumab, exhibited an anti-proliferating effect in vitro and a potent antitumor effect in vivo [4,5]. The addition of trastuzumab to chemotherapy improves objective response rates, progression-free survival, and overall survival in HER2-positive breast cancer patients with metastasis [6]. Trastuzumab has become the standard treatment for HER2-positive breast cancers [7] and HER2-positive gastric cancers [8]. For more than 20 years, trastuzumab has been the most effective therapy for HER2-positive breast cancer [9].
All anti-HER2 therapeutic mAbs exhibit cardiotoxicity, a major adverse effect [10]. Routine cardiac monitoring is required for cardiotoxic surveillance in clinic [10]. Furthermore, mice lacking ErbB2 (ortholog of HER2) displayed embryonic lethal due to the dysfunctions associated with a lack of cardiac trabeculae [11]. Ventricular-restricted ErbB2-deficient mice showed the features of dilated cardiomyopathy [12]. These results indicated that HER2 plays an important role in normal heart development and homeostasis. Therefore, more selective anti-HER2 mAbs against tumors have been desired to reduce heart failure.
We previously established anti-HER2 mAbs, H2Mab-77 (IgG1, kappa) [13], H2Mab-119 (IgG1, kappa) [14], and H2Mab-139 (IgG1, kappa) [15] by the immunization of HER2 ectodomain produced by cancer cells. During the development, we have established a variety of anti-HER2 mAbs. In this study, we performed screening to select the cancer-specific mAb (CasMab) as previously described [16,17,18], and successfully developed a cancer-specific HER2 mAb.
2. Materials and Methods
2.1. Cell lines
Chinese hamster ovary (CHO)-K1, BT-474, SK-BR-3, MDA-MB-468, MCF 10A, hTERT TIGKs, HBEC3-KT, hTERT-HME1, were RPTEC/TERT1 were obtained from the American Type Culture Collection (ATCC). HaCaT was purchased from Cell Lines Service GmbH. hTCEpi, hTEC/SVTERT24-B, and HCEC-1CT were purchased from EVERCYTE.
The cDNA of HER2 (wild type; WT) and deletion mutants (dN218, dN342, and dN511) were cloned into the pCAG/nPA16 vector. A HER2 point mutant (W614A) and HER2 WT were cloned into the pCAG/nPA-cRAPMAP vector. CHO-K1 cells were transfected with the above vectors using a Neon transfection system (Thermo Fisher Scientific Inc.). A few days after transfection, PA tag-positive cells were sorted by a cell sorter (SH800; Sony Corp.) using NZ-1, which was originally developed as an anti-human PDPN mAb [19]. Finally, CHO/HER2 and CHO/HER2 (dN218, dN342, and dN511) cell lines were established.
CHO-K1, CHO/HER2 (WT, deletion, and point mutants) were cultured in Roswell Park Memorial Institute (RPMI)-1640 medium (Nacalai Tesque, Inc.), and BT-474, SK-BR-3, MDA-MB-468, HEK293T, and HaCaT (keratinocyte) were cultured in Dulbecco’s Modified Eagle Medium (DMEM) medium (Nacalai Tesque, Inc.), supplemented with 10% heat-inactivated fetal bovine serum (FBS; Thermo Fisher Scientific Inc.), 100 units/ml of penicillin, 100 μg/ml streptomycin, and 0.25 μg/ml amphotericin B (Nacalai Tesque, Inc.). Mammary epithelial cell line, MCF 10A was cultured in Mammary Epithelial Cell Basal Medium BulletKitTM (Lonza) supplemented with 100 ng/ml cholera toxin (Sigma-Aldrich Corp.).
Immortalized normal epithelial cell lines were maintained, as follows; hTERT TIGKs, Dermal Cell Basal Medium and Keratinocyte Growth Kit (ATCC); HBEC3-KT, Airway Epithelial Cell Basal Medium and Bronchial Epithelial Cell Growth Kit (ATCC); hTERT-HME1, Mammary Epithelial Cell Basal Medium BulletKitTM without GA-1000 (Lonza); hTCEpi, KGMTM-2 BulletKitTM (Lonza); hTEC/SVTERT24-B, OptiPROTM SFM and GlutaMAXTM-I (Gibco); RPTEC/TERT1, DMEM/F-12 and hTERT Immortalized RPTEC Growth Kit with supplement A and B (ATCC); HCEC-1CT, DMEM / M199 (4:1, Gibco), 2 % Cosmic Calf Serum (Hyclone), 20 ng/ml hEGF (Sigma-Aldrich Corp.), 10 μg/ml insulin (Sigma-Aldrich Corp.), 2 μg/ml apo-transferrin (Sigma-Aldrich Corp.), 5 nM sodium-selenite (Sigma-Aldrich Corp.), 1 μg/ml hydrocortisone (Sigma-Aldrich Corp.).
All cell lines were cultured at 37°C in a humidified atmosphere with 5% CO2 and 95% air.
2.2. CasMab screening
We previously established anti-HER2 mAbs including H2Mab-119 [14] by the immunization of HER2 ectodomain (HER2ec) derived from glioblastoma LN229 or HER2-overexpressed LN229. We finally established 278 clones of hybridoma and performed the flow cytometry-based screening of CasMab using the supernatants.
2.3. Recombinant mAb production
To generate recombinant H2Mab-250 and H2Mab-119, their VH cDNAs and the CH cDNA of mouse IgG1 were cloned into the pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation). The VL cDNAs and CL cDNA of the mouse kappa light chain were also cloned into the pCAG-Neo vector (FUJIFILM Wako Pure Chemical Corporation). The vectors were transfected into ExpiCHO-S cells, and recombinant H2Mab-250 and H2Mab-119 were purified using Ab-Capcher (ProteNova).
2.4. Flow cytometry
Cells were collected using 0.25% trypsin and 1 mM ethylenediamine tetraacetic acid (EDTA; Nacalai Tesque, Inc.). The cells (1 × 105 cells/sample) were treated with H2Mab-250, H2Mab-119, or blocking buffer [control; 0.1% bovine serum albumin (BSA) in phosphate-buffered saline (PBS)] for 30 min at 4˚C. Next, the cells were treated with Alexa Fluor 488-conjugated anti-mouse IgG (1:2,000; Cell Signaling Technology) for 30 min at 4˚C. EC800 or SA3800 Cell Analyzer (Sony Corp) was used to collect the fluorescence data, which were analyzed using FlowJo (BD Biosciences).
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
Synthesized peptides covering the HER2 extracellular domain IV and point mutant peptides were synthesized by Sigma-Aldrich Corp. The peptides (10 µg/mL) were immobilized on Nunc Maxisorp 96-well immunoplates (Thermo Fisher Scientific Inc.). Plate washing was performed with PBS containing 0.05% (v/v) Tween 20 (PBST; Nacalai Tesque, Inc.). After blocking with 1% (w/v) BSA in PBST, H2Mab-250 (10 µg/mL) was added to each well. Then, the wells were further incubated with peroxidase-conjugated anti-mouse immunoglobulins (1:2000 dilution; Agilent Technologies, Inc). One-Step Ultra TMB (Thermo Fisher Scientific Inc.) was used for enzymatic reactions. An iMark microplate reader (Bio-Rad Laboratories, Inc.) was used to measure the optical density at 655 nm.
2.6. Determination of KD via Surface Plasmon Resonance (SPR)
Measurement of KD between H2Mab-250 and the HER2 peptides was performed using SPR. H2Mab-250 was immobilized on the sensor chip CM5 according to the manufacturer’s protocol by Cytiva. H2Mab-250 (10 μg/mL in acetate buffer (pH 4.0); Cytiva) was immobilized using an amine coupling reaction. The surface of the flow cell 2 of the sensor chip CM5 was treated with 1-ethyl-3-(3-dimethylaminopropyl)-carbodiimide and N-hydroxysuccinimide (NHS), followed by the injection of H2Mab-250. The KD between H2Mab-250 and the peptides was determined using Biacore X100 (Cytiva). A single cycle kinetics method was used to measure the binding signals. The data were analyzed by 1:1 binding kinetics to determine the association rate constant (ka) and dissociation rate constant (kd) and KD using Biacore X100 evaluation software (Cytiva). The affinity constant (KA) at equilibrium was calculated as 1/KD.
2.7. Immunohistochemical Analysis
Formalin-fixed paraffin-embedded (FFPE) tissue of HER2-positive breast cancer was obtained from Sendai Medical Center [13]. Informed consent for sample procurement and subsequent data analyses was obtained from the patient or the patient’s guardian at Sendai Medical Center. Normal tissues were purchased from BioChain Institute or Cybrdi. The tissue sections were autoclaved in citrate buffer (pH 6.0; Nichirei Biosciences) for 20 min. The blocking was performed using SuperBlock T20 (Thermo Fisher Scientific Inc). The sections were incubated with H2Mab-250 (1, 0.5, or 0.1 μg/mL) and H2Mab-119 (0.5 or 0.1 μg/mL), and then treated with the EnVision+ Kit for mouse (Agilent Technologies, Inc). The chromogenic reaction was performed using 3,3′-diaminobenzidine tetrahydrochloride (DAB; Agilent Technologies, Inc). Hematoxylin (FUJIFILM Wako Pure Chemical Corporation) was used for counterstaining. A Leica DMD108 (Leica Microsystems) was used to obtain images and examine the sections.
3. Results
3.1. Selection of H2Mab-250 possessing the cancer-specific HER2 recognition
Using recombinant HER2ec derived from glioblastoma LN229 or HER2-overexpressed LN229 as antigens, we developed 278 clones of anti-HER2 mAbs. We further screened the reactivity to HER2-positive breast cancers (BT-474 and SK-BR-3) and non-transformed normal epithelial cells (HaCaT and MCF 10A) using flow cytometry (Figure 1A). Although most of the clones including H2Mab-119 (IgG1, kappa) reacted with both, a clone H2Mab-250 (IgG1, kappa) reacted with cancer cells, but not with normal epithelial cells. After producing the recombinant antibody, we further confirmed the reactivity. As shown in Figure 1B, H2Mab-250 reacted with CHO/HER2, HER2-positive BT-474 and SK-BR-3 cells, but not with triple-negative MDA-MB-468 cells. H2Mab-250 did not react with HaCaT and MCF 10A cells. In contrast, H2Mab-119 showed reactivity to both cancer and normal epithelial cells.
We next investigated the difference in the reactivity to immortalized normal epithelial cells, including hTERT TIGKs (gingiva), HBEC3-KT (lung bronchus), hTERT-HME1 (mammary gland), hTCEpi (corneal), hTEC/SVTERT24-B (thymus), RPTEC/TERT1 (kidney proximal tubule), and HCEC-1CT (colon). Although H2Mab-119 showed reactivity to all immortalized normal epithelial cells, H2Mab-250 did not react with those normal cells (Figure 1C). These results indicated that H2Mab-250 possesses cancer-specific reactivity against HER2.
3.2. Epitope identification for H2Mab-250
To determine the H2Mab-250 epitope, we examined the reactivity to CHO/HER2 (WT) and the N-terminal HER2 deletion mutants (dN218, dN342, and dN511)-expressed CHO-K1 cells (Figure 2A, left). H2Mab-250 reacted with dN218, dN342, dN511, and HER2 (WT). In contrast, H2Mab-119 reacted with only WT, but not with dN218, dN342, and dN511. Because HER2 (WT) and the deletion mutants possess PA16 tag at the N-terminus, we could confirm all expression on the cell surface by anti-PA16 tag mAb, NZ-1 (Figure 2A, right). These results suggest that H2Mab-250 and H2Mab-119 recognize the domain IV and domain I, respectively.
To further assess the H2Mab-250 epitope, we performed ELISA using synthetic peptides that cover HER2 domain IV. As shown in Figure 2B, H2Mab-250 reacted with HER2 domain IV peptide, amino acids 630–622, 613–632, but not with 593–612, 623–642, and 633–652, indicating that H2Mab-250 recognizes the 613–622 of HER2 domain IV. We further used alanine-substituted peptides of the 603–622 in HER2 domain IV. A potent reduction of the reactivity was observed in the W614A peptide (Figure 2C). We confirmed that the reactivity of H2Mab-250 completely disappeared in CHO/HER2 W614A cells in flow cytometry (Figure 2D).
We also measured the dissociation constant (KD) of H2Mab-250 with the alanine-substituted peptides of HER2 domain IV (603–622) using Biacore X100 (Table 1). The affinity constant (KA) at equilibrium was calculated as 1/KD (Figure 2E). Compared to the KA of the 603–622 (WT) peptide, decreased KA values were observed from the 613–617 region, suggesting that the 613–617 region is involved in the binding to H2Mab-250. A remarkable reduction was measured in the W614A peptide. These results indicated that Trp614 mainly contributes to the recognition by H2Mab-250.
3.3. Immunohistochemical analysis of H2Mab-250 in breast cancer and normal epithelium
We next performed immunohistochemistry to examine the reactivity of H2Mab-250 to normal and tumor tissue. In contrast to flow cytometry, H2Mab-250 exhibited more potent reactivity to the HER2-positive breast cancer section than H2Mab-119 (Figure 3A). Because all anti-HER2 therapeutic mAbs exhibit cardiotoxicity, a major adverse effect [10], we investigated the reactivity of H2Mab-250 to a normal heart. We could not detect the reactivity even in higher concentrations of H2Mab-250 (1 µg/mL) (Figure 3B). Finally, the reactivity of H2Mab-250 to other normal tissues was investigated. As shown in Figure 3C, we could not observe any reactivity to normal tissues, including breast, stomach, lung, colon, kidney, and esophagus.
4. Discussion
In this study, we developed a cancer-specific mAb targeting HER2. H2Mab-250 can recognize breast cancer cells, but not normal cells in flow cytometry (Figure 1) and immunohistochemistry (Figure 3). We also identified the H2Mab-250 epitope sequence (613-IWKFP-617) by SPR analysis (Figure 2). Furthermore, we found that H2Mab-250 never reacted with CHO/HER2 W614A in flow cytometry (Figure 2), suggesting that Trp614 plays a central role in recognition by H2Mab-250 in living cells. Although H2Mab-250 possesses a high affinity to epitope-containing peptide (603–622) in SPR analysis, the recognition by flow cytometry was low compared with that of H2Mab-119 (Figure 2). In contrast, H2Mab-250 exhibited a higher reactivity than H2Mab-119 in the immunohistochemical analysis of breast cancer (Figure 3). There is a possibility that the epitope sequence is partially exposed in cancer cells, but not in normal cells in living cells. We should reveal the mechanism of recognition by H2Mab-250 in future studies. Furthermore, the strategy of CasMab selection would contribute to the development of novel mAbs against a variety of antigens.
For the clinical treatment of metastatic breast cancer, trastuzumab is administered for patients with HER2-overexpressing tumors, which are defined by strong and complete IHC membranous staining of more than 10% of cells (IHC 3+) and/or in situ hybridization (ISH)-amplified. Furthermore, trastuzumab-based antibody-drug conjugates (ADCs) such as trastuzumab-deruxtecan (T-DXd) have been evaluated in various clinical trials. Based on the studies, T-DXd has been approved in not only HER2-positive breast cancer [20,21], but also HER2-mutant lung cancer [22] and HER2-low (IHC 1+ or IHC 2+ / ISH-non-amplified) advanced breast cancer [23]. Because approximately half of all breast cancers are classifiable as HER2-low [24], a greater number of patients may benefit from T-DXd therapy. Meanwhile, cardiotoxicity is the most significant toxicity associated with T-DXd [25]. Further studies are essential to evaluate in vivo toxicities of H2Mab-250.
Chimeric antigen receptor (CAR)-T cell therapy against HER2 has been evaluated in clinical studies [24]. Although the optimal epitope for HER2-targeting CAR-T has not been evaluated, it is worthwhile to investigate the cancer specificity of H2Mab-250 scFv and the efficacy of CAR-T against HER2-positive tumors.
Author Contributions
H.S. and M.K.K. performed the experiments. M.K.K. and Y.K. designed the experiments. H.S., M.K.K., and Y.K. analyzed the data. H.S. and Y.K. wrote the manuscript. All authors have read and agreed to the manuscript.
Funding
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP23ama121008 (to Y.K.), JP23am0401013 (to Y.K.), 23bm1123027h0001 (to Y.K.), JP23ck0106730 (to Y.K.), JP18am0301010 (to Y.K.), and JP21am0101078 (to Y.K.).
Data Availability Statement
The data presented in this study are available in the article and supplementary material.
Conflicts of Interest
The authors have no conflicts of interest to declare.
References
- Yarden, Y.; Sliwkowski, M.X. Untangling the ErbB signalling network. Nat Rev Mol Cell Biol 2001, 2, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Slamon, D.J.; Clark, G.M.; Wong, S.G.; Levin, W.J.; Ullrich, A.; McGuire, W.L. Human breast cancer: Correlation of relapse and survival with amplification of the HER-2/neu oncogene. Science 1987, 235, 177–182. [Google Scholar] [CrossRef]
- Van Cutsem, E.; Bang, Y.J.; Feng-Yi, F.; Xu, J.M.; Lee, K.W.; Jiao, S.C.; Chong, J.L.; López-Sanchez, R.I.; Price, T.; Gladkov, O.; et al. HER2 screening data from ToGA: Targeting HER2 in gastric and gastroesophageal junction cancer. Gastric Cancer 2015, 18, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.S.; Mason, K.; Ramyar, K.X.; Stanley, A.M.; Gabelli, S.B.; Denney, D.W., Jr.; Leahy, D.J. Structure of the extracellular region of HER2 alone and in complex with the Herceptin Fab. Nature 2003, 421, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Tsao, L.C.; Force, J.; Hartman, Z.C. Mechanisms of Therapeutic Antitumor Monoclonal Antibodies. Cancer Res 2021, 81, 4641–4651. [Google Scholar] [CrossRef]
- Essadi, I.; Benbrahim, Z.; Kaakoua, M.; Reverdy, T.; Corbaux, P.; Freyer, G. HER2-Positive Metastatic Breast Cancer: Available Treatments and Current Developments. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef]
- Slamon, D.J.; Leyland-Jones, B.; Shak, S.; Fuchs, H.; Paton, V.; Bajamonde, A.; Fleming, T.; Eiermann, W.; Wolter, J.; Pegram, M.; et al. Use of chemotherapy plus a monoclonal antibody against HER2 for metastatic breast cancer that overexpresses HER2. N Engl J Med 2001, 344, 783–792. [Google Scholar] [CrossRef]
- Bang, Y.J.; Van Cutsem, E.; Feyereislova, A.; Chung, H.C.; Shen, L.; Sawaki, A.; Lordick, F.; Ohtsu, A.; Omuro, Y.; Satoh, T.; et al. Trastuzumab in combination with chemotherapy versus chemotherapy alone for treatment of HER2-positive advanced gastric or gastro-oesophageal junction cancer (ToGA): A phase 3, open-label, randomised controlled trial. Lancet 2010, 376, 687–697. [Google Scholar] [CrossRef]
- Maadi, H.; Soheilifar, M.H.; Choi, W.S.; Moshtaghian, A.; Wang, Z. Trastuzumab Mechanism of Action; 20 Years of Research to Unravel a Dilemma. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Copeland-Halperin, R.S.; Liu, J.E.; Yu, A.F. Cardiotoxicity of HER2-targeted therapies. Curr Opin Cardiol 2019, 34, 451–458. [Google Scholar] [CrossRef]
- Lee, K.F.; Simon, H.; Chen, H.; Bates, B.; Hung, M.C.; Hauser, C. Requirement for neuregulin receptor erbB2 in neural and cardiac development. Nature 1995, 378, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Crone, S.A.; Zhao, Y.Y.; Fan, L.; Gu, Y.; Minamisawa, S.; Liu, Y.; Peterson, K.L.; Chen, J.; Kahn, R.; Condorelli, G.; et al. ErbB2 is essential in the prevention of dilated cardiomyopathy. Nat Med 2002, 8, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Itai, S.; Fujii, Y.; Kaneko, M.K.; Yamada, S.; Nakamura, T.; Yanaka, M.; Saidoh, N.; Chang, Y.W.; Handa, S.; Takahashi, M.; et al. H2Mab-77 is a Sensitive and Specific Anti-HER2 Monoclonal Antibody Against Breast Cancer. Monoclon Antib Immunodiagn Immunother 2017, 36, 143–148. [Google Scholar] [CrossRef]
- Yamada, S.; Itai, S.; Nakamura, T.; Chang, Y.W.; Harada, H.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Establishment of H(2)Mab-119, an Anti-Human Epidermal Growth Factor Receptor 2 Monoclonal Antibody, Against Pancreatic Cancer. Monoclon Antib Immunodiagn Immunother 2017, 36, 287–290. [Google Scholar] [CrossRef]
- Kaneko, M.K.; Yamada, S.; Itai, S.; Kato, Y. Development of an Anti-HER2 Monoclonal Antibody H2Mab-139 Against Colon Cancer. Monoclon Antib Immunodiagn Immunother 2018, 37, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.K.; Ohishi, T.; Kawada, M.; Kato, Y. A cancer-specific anti-podocalyxin monoclonal antibody (60-mG(2a)-f) exerts antitumor effects in mouse xenograft models of pancreatic carcinoma. Biochem Biophys Rep 2020, 24, 100826. [Google Scholar] [CrossRef]
- Yamada, S.; Ogasawara, S.; Kaneko, M.K.; Kato, Y. LpMab-23: A Cancer-Specific Monoclonal Antibody Against Human Podoplanin. Monoclon Antib Immunodiagn Immunother 2017, 36, 72–76. [Google Scholar] [CrossRef]
- Kato, Y.; Kaneko, M.K. A cancer-specific monoclonal antibody recognizes the aberrantly glycosylated podoplanin. Sci Rep 2014, 4, 5924. [Google Scholar] [CrossRef]
- Kato, Y.; Kaneko, M.K.; Kuno, A.; Uchiyama, N.; Amano, K.; Chiba, Y.; Hasegawa, Y.; Hirabayashi, J.; Narimatsu, H.; Mishima, K.; et al. Inhibition of tumor cell-induced platelet aggregation using a novel anti-podoplanin antibody reacting with its platelet-aggregation-stimulating domain. Biochem Biophys Res Commun 2006, 349, 1301–1307. [Google Scholar] [CrossRef]
- Modi, S.; Saura, C.; Yamashita, T.; Park, Y.H.; Kim, S.B.; Tamura, K.; Andre, F.; Iwata, H.; Ito, Y.; Tsurutani, J.; et al. Trastuzumab Deruxtecan in Previously Treated HER2-Positive Breast Cancer. N Engl J Med 2020, 382, 610–621. [Google Scholar] [CrossRef]
- Shitara, K.; Bang, Y.J.; Iwasa, S.; Sugimoto, N.; Ryu, M.H.; Sakai, D.; Chung, H.C.; Kawakami, H.; Yabusaki, H.; Lee, J.; et al. Trastuzumab Deruxtecan in Previously Treated HER2-Positive Gastric Cancer. N Engl J Med 2020, 382, 2419–2430. [Google Scholar] [CrossRef] [PubMed]
- Li, B.T.; Smit, E.F.; Goto, Y.; Nakagawa, K.; Udagawa, H.; Mazières, J.; Nagasaka, M.; Bazhenova, L.; Saltos, A.N.; Felip, E.; et al. Trastuzumab Deruxtecan in HER2-Mutant Non-Small-Cell Lung Cancer. N Engl J Med 2022, 386, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Modi, S.; Jacot, W.; Yamashita, T.; Sohn, J.; Vidal, M.; Tokunaga, E.; Tsurutani, J.; Ueno, N.T.; Prat, A.; Chae, Y.S.; et al. Trastuzumab Deruxtecan in Previously Treated HER2-Low Advanced Breast Cancer. N Engl J Med 2022, 387, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Mercogliano, M.F.; Bruni, S.; Mauro, F.L.; Schillaci, R. Emerging Targeted Therapies for HER2-Positive Breast Cancer. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef]
- Soares, L.R.; Vilbert, M.; Rosa, V.D.L.; Oliveira, J.L.; Deus, M.M.; Freitas-Junior, R. Incidence of interstitial lung disease and cardiotoxicity with trastuzumab deruxtecan in breast cancer patients: A systematic review and single-arm meta-analysis. ESMO Open 2023, 8, 101613. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Selection of H2Mab-250, a CasMab against HER2. (A) A scheme of CasMab selection from hybridoma clones. (B) Flow cytometry using H2Mab-250 (Red line) and H2Mab-119 (Blue line) against CHO-K1, CHO/HER2, HER2-positive breast cancers (BT-474 and SK-BR-3), a triple-negative breast cancer (MDA-MB-468), and non-transformed normal epithelial cells (HaCaT and MCF 10A). (C) Flow cytometry using H2Mab-250 (Red line) and H2Mab-119 (Blue line) against immortalized normal epithelial cells including hTERT TIGKs (gingiva), HBEC3-KT (lung bronchus), hTERT-HME1 (mammary gland), hTCEpi (corneal), hTEC/SVTERT24-B (thymus), RPTEC/TERT1 (kidney proximal tubule), and HCEC-1CT (colon). The black line represents the negative control (blocking buffer).
Figure 1.
Selection of H2Mab-250, a CasMab against HER2. (A) A scheme of CasMab selection from hybridoma clones. (B) Flow cytometry using H2Mab-250 (Red line) and H2Mab-119 (Blue line) against CHO-K1, CHO/HER2, HER2-positive breast cancers (BT-474 and SK-BR-3), a triple-negative breast cancer (MDA-MB-468), and non-transformed normal epithelial cells (HaCaT and MCF 10A). (C) Flow cytometry using H2Mab-250 (Red line) and H2Mab-119 (Blue line) against immortalized normal epithelial cells including hTERT TIGKs (gingiva), HBEC3-KT (lung bronchus), hTERT-HME1 (mammary gland), hTCEpi (corneal), hTEC/SVTERT24-B (thymus), RPTEC/TERT1 (kidney proximal tubule), and HCEC-1CT (colon). The black line represents the negative control (blocking buffer).
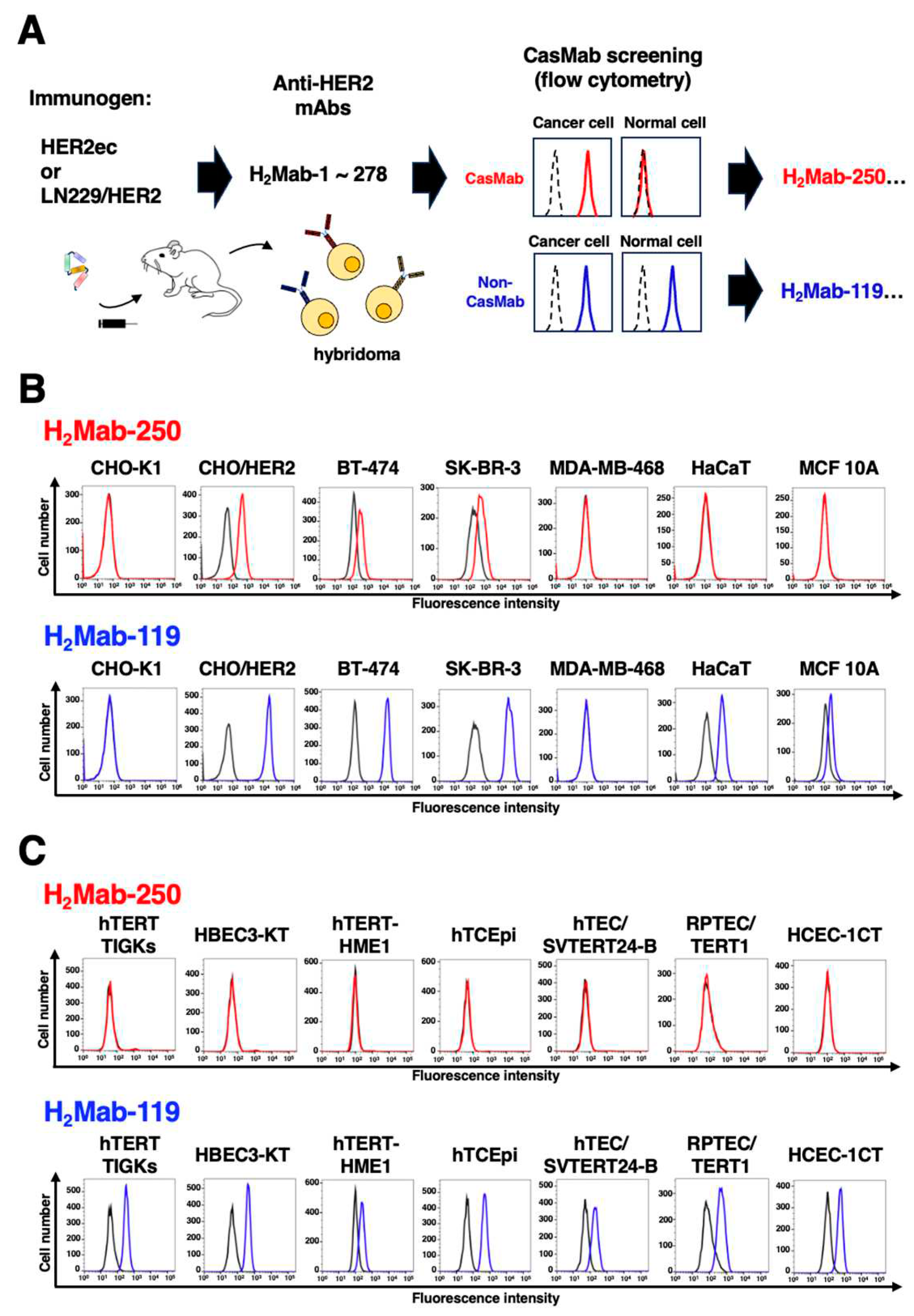
Figure 2.
Epitope identification for H2Mab-250. (A) Epitope determination of H2Mab-250 and H2Mab-119 using flow cytometry. The schematic representation of HER2 and the deletion mutants (left). Flow cytometry using H2Mab-250 (Red line) and H2Mab-119 (Blue line) against CHO/HER2 (WT and deletion mutants). The cell surface expression was confirmed by an anti-PA tag mAb, NZ-1 (Green). The black line represents the negative control (blocking buffer). (B and C) Determination of H2Mab-250 epitope by ELISA. Five synthesized peptides that cover the HER2 domain IV (B), alanine-substituted peptides of HER2 domain IV (603–622) (C), HER2ec, or buffer control (NC) were immobilized on immunoplates. The plates were incubated with H2Mab-250, followed by incubation with peroxidase-conjugated anti-mouse immunoglobulins. Optical density was measured at 655 nm. (D) Flow cytometry using H2Mab-250 against CHO/HER2 (WT and W614A). The cell surface expression was confirmed by an anti-PA tag mAb, NZ-1. The black line represents the negative control (blocking buffer). (E) Surface Plasmon Resonance analysis between H2Mab-250 and HER2 domain IV (603–622) peptides. The affinity constant (KA) at equilibrium was calculated as 1/KD.
Figure 2.
Epitope identification for H2Mab-250. (A) Epitope determination of H2Mab-250 and H2Mab-119 using flow cytometry. The schematic representation of HER2 and the deletion mutants (left). Flow cytometry using H2Mab-250 (Red line) and H2Mab-119 (Blue line) against CHO/HER2 (WT and deletion mutants). The cell surface expression was confirmed by an anti-PA tag mAb, NZ-1 (Green). The black line represents the negative control (blocking buffer). (B and C) Determination of H2Mab-250 epitope by ELISA. Five synthesized peptides that cover the HER2 domain IV (B), alanine-substituted peptides of HER2 domain IV (603–622) (C), HER2ec, or buffer control (NC) were immobilized on immunoplates. The plates were incubated with H2Mab-250, followed by incubation with peroxidase-conjugated anti-mouse immunoglobulins. Optical density was measured at 655 nm. (D) Flow cytometry using H2Mab-250 against CHO/HER2 (WT and W614A). The cell surface expression was confirmed by an anti-PA tag mAb, NZ-1. The black line represents the negative control (blocking buffer). (E) Surface Plasmon Resonance analysis between H2Mab-250 and HER2 domain IV (603–622) peptides. The affinity constant (KA) at equilibrium was calculated as 1/KD.
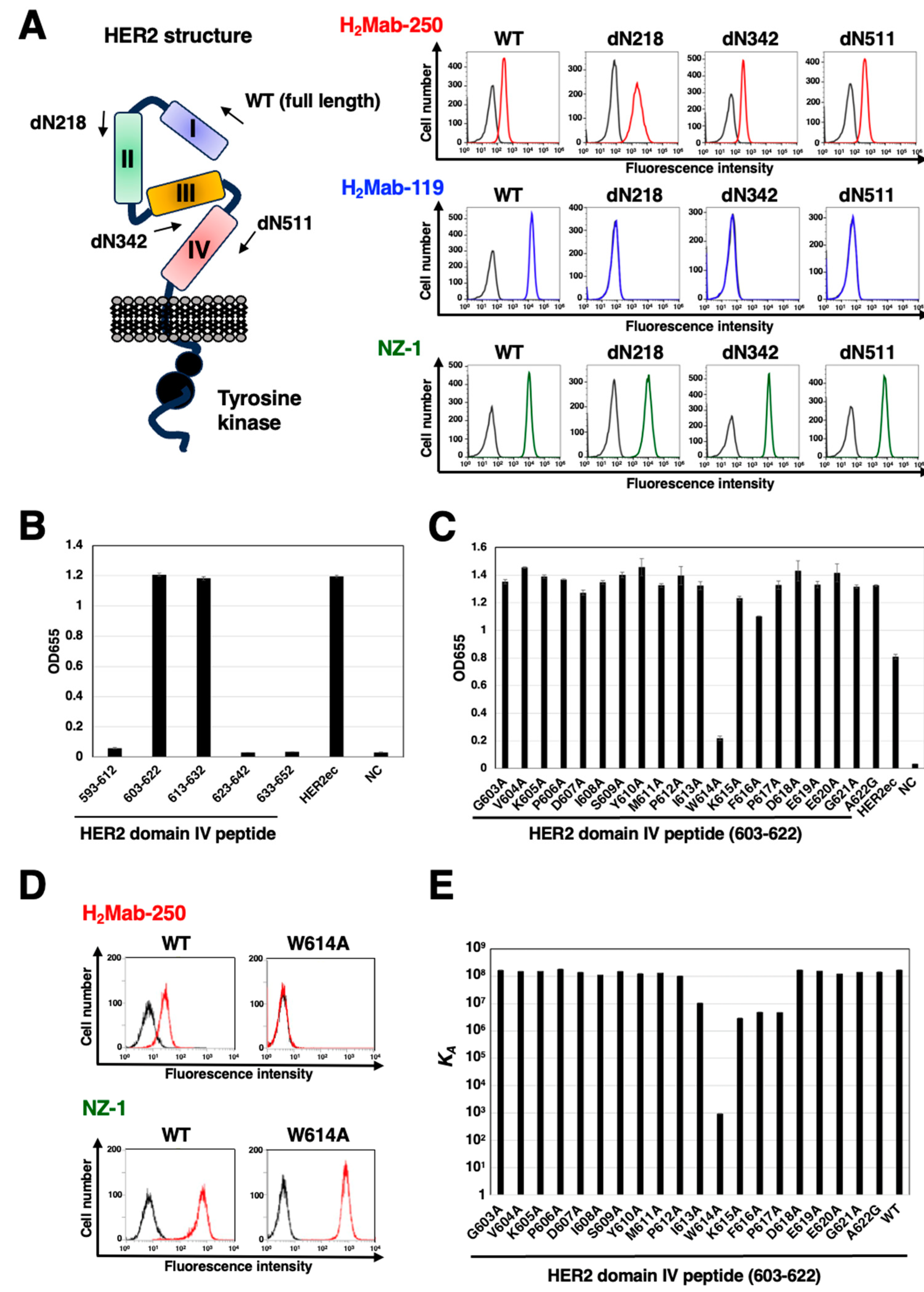
Figure 3.
Immunohistochemical analysis of H2Mab-250 in breast cancer and normal epithelium. (A) The HER2-positive breast cancer sections were treated with H2Mab-250 or H2Mab-119 (0.1 or 0.5 µg/mL). (B) A normal heart section was treated with H2Mab-250 (1 µg/mL). (C) Sections of normal breast, stomach, lung, colon, kidney, and esophagus were treated with H2Mab-250 (0.1 µg/mL). The sections were then treated with the Envision+ kit. The chromogenic reaction was performed using DAB, and the sections were counterstained with hematoxylin. Scale bar = 100 µm.
Figure 3.
Immunohistochemical analysis of H2Mab-250 in breast cancer and normal epithelium. (A) The HER2-positive breast cancer sections were treated with H2Mab-250 or H2Mab-119 (0.1 or 0.5 µg/mL). (B) A normal heart section was treated with H2Mab-250 (1 µg/mL). (C) Sections of normal breast, stomach, lung, colon, kidney, and esophagus were treated with H2Mab-250 (0.1 µg/mL). The sections were then treated with the Envision+ kit. The chromogenic reaction was performed using DAB, and the sections were counterstained with hematoxylin. Scale bar = 100 µm.
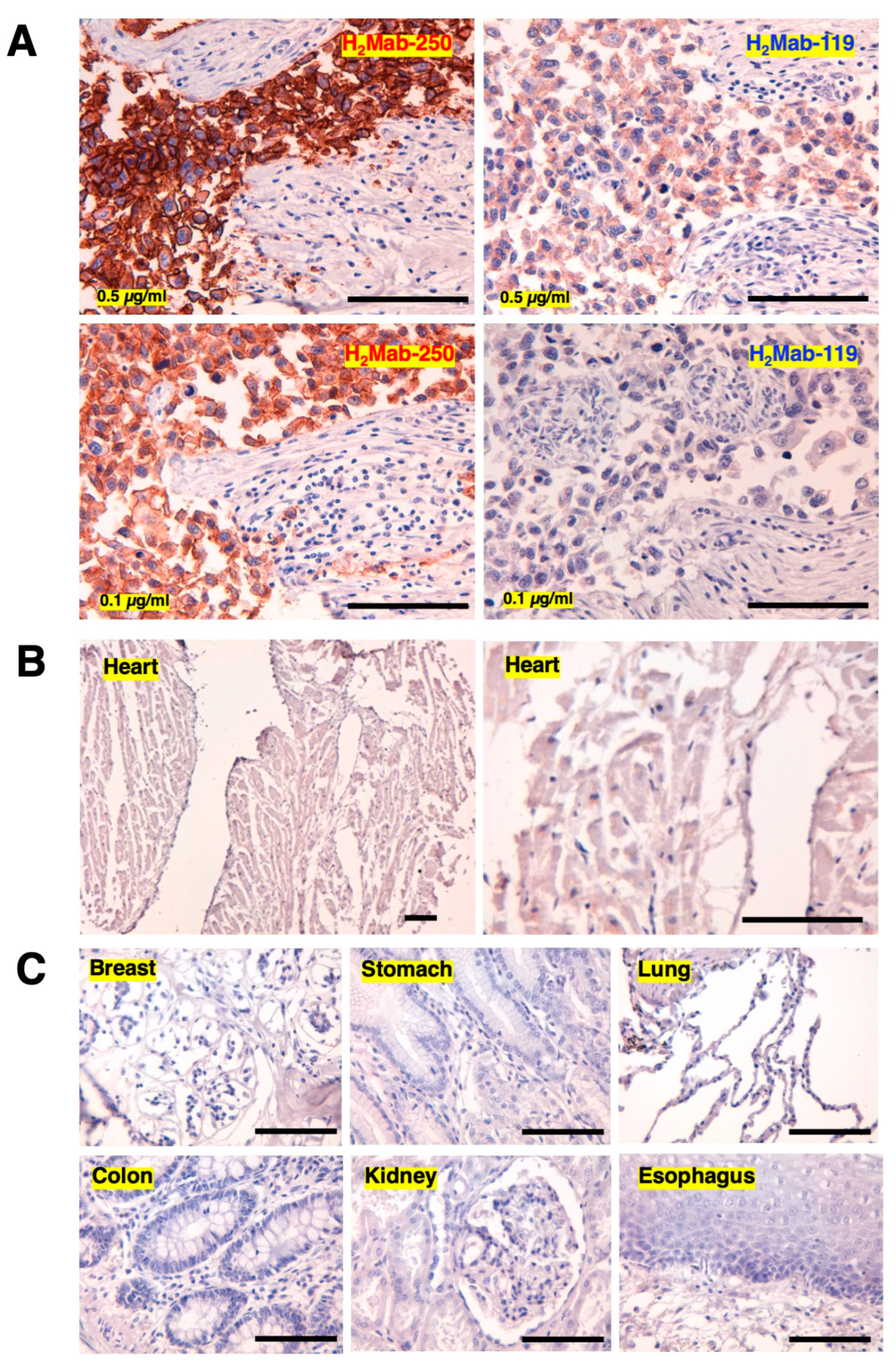

Table 1.
Identification of H2Mab-250 epitope using point mutants by Biacore.
| Peptide | Sequence | KD (M) |
|---|---|---|
| 603-622 | GVKPDLSYMPIWKFPDEEGA | 5.8 × 10-9 |
| G603A | AVKPDLSYMPIWKFPDEEGA | 5.9 × 10-9 |
| V604A | GAKPDLSYMPIWKFPDEEGA | 6.5 × 10-9 |
| K605A | GVAPDLSYMPIWKFPDEEGA | 6.5 × 10-9 |
| P606A | GVKADLSYMPIWKFPDEEGA | 5.3 × 10-9 |
| D607A | GVKPALSYMPIWKFPDEEGA | 7.1 × 10-9 |
| L608A | GVKPDASYMPIWKFPDEEGA | 8.8 × 10-9 |
| S609A | GVKPDLAYMPIWKFPDEEGA | 6.5 × 10-9 |
| Y610A | GVKPDLSAMPIWKFPDEEGA | 7.9 × 10-9 |
| M611A | GVKPDLSYAPIWKFPDEEGA | 7.5 × 10-9 |
| P612A | GVKPDLSYMAIWKFPDEEGA | 9.5 × 10-9 |
| I613A | GVKPDLSYMPAWKFPDEEGA | 9.4 × 10-8 |
| W614A | GVKPDLSYMPIAKFPDEEGA | 1.1 × 10-3 |
| K615A | GVKPDLSYMPIWAFPDEEGA | 3.4 × 10-7 |
| F616A | GVKPDLSYMPIWKAPDEEGA | 2.0 × 10-7 |
| P617A | GVKPDLSYMPIWKFADEEGA | 2.1 × 10-7 |
| D618A | GVKPDLSYMPIWKFPAEEGA | 5.8 × 10-9 |
| E619A | GVKPDLSYMPIWKFPDAEGA | 6.3 × 10-9 |
| E620A | GVKPDLSYMPIWKFPDEAGA | 8.0 × 10-9 |
| G621A | GVKPDLSYMPIWKFPDEEAA | 6.9 × 10-9 |
| A622G | GVKPDLSYMPIWKFPDEEGG | 6.9 × 10-9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



