
Submitted:
11 September 2023
Posted:
14 September 2023
You are already at the latest version
Abstract
Yerba maté, an herbal tea derived from Ilex paraguariensis, has previously been reported to be protective against obesity-related and other cardiometabolic disorders. Using high-resolution respirometry and reverse-phase high-performance liquid chromatography, the effects of four weeks of yerba maté consumption on mitochondrial efficiency and cellular redox status in skeletal muscle, adipose, and liver, tissues highly relevant to whole body metabolism, were explored in healthy adult mice. Yerba maté treatment increased mitochondrial oxygen consumption in adipose, but not in other tissues examined. Yerba maté increased ATP concentration in skeletal muscle and decreased ATP concentration in adipose. Combined with observed changes in oxygen consumption, these data yielded a significantly higher ATP:O2, a measure of mitochondrial efficiency, in muscle and a significantly lower ATP:O2 in adipose, which were consistent with yerba maté-induced weight loss. Yerba maté treatment also altered the hepatic glutathione (GSH)/glutathione disulfide (GSSG) redox potential to a more reduced redox state, suggesting potential protective effects against oxidative stress and preservation of cellular function. Together, these data indicate beneficial, tissue-specific effects of yerba maté supplementation on mitochondrial bioenergetics and redox states in healthy mice that are protective against obesity.
Keywords:
Yerba Maté
; Metabolism
; Obesity
; Nutraceuticals
; Mitochondria
1. Introduction
The unabated rise in cardiometabolic disorders such as heart disease, obesity, and type 2 diabetes mellitus necessitates novel approaches to reduce associated risk factors. In recent years, scrutiny has been placed on dietary factors with a particular emphasis on macronutrients (i.e., fat, protein, carbohydrate) [1] and meal frequency [2]. However, a growing body of evidence suggests that these strategies, while certainly relevant, may overlook potential roles of other metabolites in mitigating cardiometabolic disease risk [3,4].
Secondary metabolites are naturally occurring substances produced by plants to make them more competitive in their environments, thus increasing their survival. These metabolites have historically been an important source of medicinal compounds and have been utilized in developing a number of therapeutics for diverse disease states [5]. Yerba maté is an herbal tea derived from dried leaves of the South American Ilex paraguariensis plant and is widely consumed in South American countries such as Brazil, Argentina, Paraguay, and Uruguay. Yerba maté is a particularly rich source of secondary metabolites that have been implicated in improving cardiometabolic health. These compounds include purine alkaloids such as caffeine and theobromine, polyphenols such as phenolic acids and flavonoids, and terpenes such as saponins and carotenoids, as well as numerous minerals and vitamins [5].
While some foods are known sources of secondary metabolites [6], environmental and farming trends have resulted in the steady reduction of these beneficial molecules in the foods we consume [7], increasing the need for obtaining secondary metabolites from supplemental sources. Supplements enriched with these and other secondary metabolites have been shown to improve aspects of cardiovascular [3] and metabolic health [6,8], cognitive function [9], and more [10].
Indeed, yerba maté tea has been shown to favorably alter the lipid profile in humans [11]. Phenolic compounds such as chlorogenic acids, a class of phenol that is uniquely elevated in yerba mate tea [12], have been linked to several beneficial health outcomes such as lipid homeostasis and weight management, cardiovascular protection, and glucose control [13]. Additionally, phenols and flavonoids have been shown to bolster antioxidant defenses [14] and reduce inflammation [15,16], which likely contribute to yerba maté’s protective effects in aging.
Mitochondria are widely considered hubs of cellular metabolism for their essential role in energy production. Impairments in mitochondria have been demonstrated in and have been suggested to drive metabolic disease [17]. Improving mitochondrial health, therefore, has been proposed as a means of mitigating disease. In this study, we explore the effects of a four-week-long exposure to yerba maté on mitochondrial bioenergetics and redox potential within skeletal muscle, adipose, and hepatic tissue and demonstrate yerba maté supplementation has beneficial metabolic effects that vary by tissue. In total, this work adds to a host of evidence demonstrating that whole-body and tissue-specific metabolism is alterable via dietary intervention.
2. Materials and Methods
Animals
Six-month-old male and female C57BL/6 mice were group-housed at 22±1°C, 60%-70% humidity, and were maintained on a 12-hour light-dark cycle with ad libitum access to food. Mice were randomly divided into two groups and given free access to either water (control, CON) or yerba maté (MATÉ, Unicity International) for four weeks. Both groups of mice had ad libitum access to a standard rodent chow (LabDiet 5001) and their respective drinks. Water bottles containing water and yerba maté were made fresh daily after consumption measurements were taken and mice were weighed weekly. Following conditioning, mice were sacrificed and tissues were collected (gastrocnemius skeletal muscle, subcutaneous adipose, and liver), then were either snap frozen and stored at 80°C for ATP quantification or kept on ice for mitochondrial respirometry and GSH/GSSG quantification.
Studies were conducted in accordance with the principles and procedures outlined in the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee at Brigham Young University (protocol #20-0203).
Tissue permeabilization
Approximately 10-20 mg of each tissue were held on ice for mitochondrial respirometry. Samples were transferred to tubes containing 50 µg/ml of saponin in MiR05 respiration buffer [0.5 mM EGTA, 3 mM MgCl2, 60 mM K-lactobionate, 20 mM taurine, 10 mM KH2PO4, 20 mM HEPES, 110 mM sucrose, and g/l BSA (Sigma; A3803) adjusted to pH 7.1].
Mitochondrial respirometry
High-resolution O2 consumption was determined at 37°C in permeabilized muscle fiber bundles, adipose, and liver using the Oroboros O2K Oxygraph with MiR05 respiration buffer, as described previously [18,19,20,21]. Before the addition of sample into respiration chambers, a baseline respiration rate was determined. After the addition of sample, the chambers were briefly hyperoxygenated to ∼250 nmol/ml. Following hyperoxygenation, respiration was determined by all or parts of the following substrate–uncoupler–inhibitor–titration protocol: electron flow through complex I was supported by glutamate + malate (10 and 2 mM, respectively) to determine leak oxygen consumption (GM). Following stabilization, ADP (2.5 mM) was added to determine oxidative phosphorylation capacity (D). Succinate was added (S) for complex I + II electron flow into the Q-junction.
ATP quantification
ATP was quantified from frozen tissue samples with an ATPLite Luminescence Assay kit (Perkin Elmer). Frozen samples were thawed on ice and homogenized in mammalian cell lysis buffer provided by the manufacturer. Homogenates were diluted in water and were transferred to opaque 96-well plates in volumes of100 µl per well. ATPLite lysis buffer was added (50 µl) to each sample and the plates were agitated for 5 min at 700 rpm at room temperature. ATPLite substrate solution was then added (50 µl) to each well. The plates were covered with aluminum foil, agitated for an additional 5 min at 700 rpm at room temperature, and dark-adapted for 10 min. Luminescence was measured with a Victor Nivo Multimode Plate Reader (Perkin Elmer).
Glutathione/glutathione disulfide redox potential analysis
Methods for redox potential analysis have been previously described by Piorczynski, et al. [22]. Isolated tissues were collected in 5% perchloric acid and boric acid (0.2 M) containing γ-glutamylglutamate (10 μM, Sigma-Aldrich) for GSH analysis. Concentrations of GSH and its oxidized form, glutathione disulfide (GSSG), were measured by reverse-phase high-performance liquid chromatography as S-carboxymethyl, N-dansyl derivatives using the internal standard γ-glutamylglutamate for normalization as described previously [23]. Proteins were acid-precipitated and samples centrifuged at 16,000 ×g for 5 min after which the soluble fraction containing free GSH and GSSG was derivatized. Samples were analyzed using an e2695 Separations Module (Waters) fitted with a Supelcosil LC-NH2 5 μm column (Sigma-Aldrich) and detection of peaks was determined using a 2474 FLR Detector (excitation 335 nm and emission 518 nm, Waters). The GSH/GSSG redox potential (Eh) was calculated by the Nernst equation using intracellular GSH and GSSG concentrations [24].
Statistics
Data are presented as the means ±SEM. Differences between the CON and MATÉ means were compared using Student’s t-tests (GraphPad Prism; Microsoft Excel). Significance was determined at p < 0.05 and a trend toward significance at p < 0.15.
3. Results
Yerba maté consumption prevents weight gain in male and female mice
Over the four weeks of conditioning, male and female mice were maintained on a standard rodent diet. CON mice gained a significant amount of weight compared to MATÉ mice, which displayed no significant changes relative to their starting weight. Significant differences in weight were observed between CON and MATÉ mice throughout conditioning beginning at week 2 (Figure 1; week 1, p = 0.444; week 2, p = 0.0432; week 3, p = 0.0189; week 4, p = 0.0118).
Yerba maté consumption increases mitochondrial efficiency in skeletal muscle
High-resolution respirometry performed on permeabilized gastrocnemius muscle fiber bundles indicated a significant increase in mitochondrial oxygen consumption between CON and MATÉ mice (Figure 2A) with the addition of ADP (p = 0.0296) but no significant differences with the addition of glutamate and malate (p = 0.216) or succinate (p = 0.481). ATP quantification indicated a significant increase in ATP concentration in homogenized gastrocnemius with yerba maté consumption (Figure 2B, p = 0.0023). The ratio of gastrocnemius ATP concentration to oxygen consumption (ATP:O2), a measure of mitochondrial efficiency, was also significantly increased with yerba maté treatment compared to control (Figure 2C, p = 0.0332).
To determine changes in intracellular redox potential with MATÉ treatment, concentrations of reduced and oxidized forms of glutathione, GSH and GSSG respectively, were quantified. These analyses yielded a significant increase in GSH concentration (Figure 2D, p = 0.0491) but no significant difference in GSSG concentration (Figure 2E, p = 0.898) or the calculated Eh (Figure 2F, p = 0.2778).
Yerba maté consumption decreases mitochondrial efficiency in white adipose tissue
High-resolution respirometry performed on permeabilized subcutaneous adipose tissue collected from subcutaneous white adipose fat pads revealed significant increases in mitochondrial oxygen consumption rates with yerba maté treatment (Figure 3A) with the addition of glutamate and malate (p = 0.0101), ADP (p = 0.0079), and succinate (p = 0.0367). ATP quantification indicated a significant decrease in ATP production with yerba maté consumption compared to water (Figure 3B, p = 0.0434). ATP:O2 ratios strongly trended toward a significant decrease (Figure 3C, p = 0.050007).
Yerba maté consumption alters hepatic redox potential but not mitochondrial efficiency
High-resolution respirometry performed on permeabilized liver tissue revealed no significant changes in mitochondrial oxygen consumption rate (Figure 4A) between control and yerba mate-treated mice with the addition of glutamate and malate (p = 0.262), ADP (p = 0.297), or succinate (p = 0.882). No changes in ATP production (Figure 4B, p = 0.6141) or ATP:O2 ratio (Figure 4C, p = 0.315) were observed.
Redox analysis revealed a significant increase in GSH concentration (Figure 4D, p = 0.0449), but no significant change in GSSG concentration (Figure 4E, p = 0.334). This yielded a significant decrease in GSH/GSSG Eh (Figure 4F, p = 0.0251), indicating a shift toward a more reduced intracellular environment.
4. Discussion
Previous studies from our lab have demonstrated that diet has the capacity to alter mitochondrial efficiency in adipose [21] and brain tissue [25]. This study adds further support that diet alters mitochondrial bioenergetics. Specifically, these data demonstrate that yerba maté affects mitochondrial efficiency in a tissue-specific manner such that maté consumption increases mitochondrial efficiency within skeletal muscle but decreases mitochondrial efficiency within adipose tissue. We interpreted both of these changes to be metabolically beneficial, as they agreed with our observation that yerba maté consumption protected mice against weight gain on a standard rodent diet (Figure 1). We also observed beneficial tissue-specific shifts in hepatic redox potential (Figure 4F), which further support our positive interpretations.
While CON mice gained a significant amount of weight over the four weeks of conditioning, MATE mice experienced no significant changes compared to their baseline weight (Figure 1). This is consistent with other rodent and human studies that have demonstrated beneficial effects of yerba maté on lipid oxidation and weight management, at large [26,27,28,29] . Indeed, multiple studies have demonstrated anti-obesity effects of yerba maté in rodents fed obesogenic diets. For example, one study fed Swiss mice a high fat diet (HFD) for eight weeks [30]. These mice were then divided into two groups and fed either water (vehicle control) or yerba maté for an additional eight weeks. Obese HFD-fed mice expectedly gained a significant amount of weight. This increase in weight was accompanied by systemic and hepatic insulin resistance and elevations in total cholesterol, high and low-density lipoprotein cholesterol, and triglycerides. Yerba maté treatment attenuated, and in some instances, completely reversed these effects. In a hamster model of diet-induced hyperlipidemia, yerba maté consumption lowered the concentrations of serum lipids by modulating the expression of genes of lipid oxidation and increasing the activities of lipoprotein and hepatic lipases [27]. Another study in Wistar rats maintained on a standard rodent diet supplemented with teas made from raw (not industrially processed) or commercial (industrially processed) maté indicated no significant changes in body weight or plasma lipids over 30 days [31]. However, despite the lack of weight loss, intra-abdominal fat pad mass was significantly reduced in the raw maté group and epididymal fat pad mass was significantly lower in both maté groups, indicating more efficient lipid oxidation. In our study, mice were also maintained on a standard rodent diet. However, we observed significant increase in body weight in CON mice that was not observed in Silva, et al.’s work [31]. Although we did not directly assess changes in the mass of intra-abdominal or epididymal fat pads, the differences in weight we observed are suggestive.
Anti-obesity effects of yerba maté supplements have also been observed in human clinical trials. For example, a double-blind, placebo-controlled study administered capsules of either yerba mate extract or placebo to male and female subjects with obesity for 12 weeks [32]. In this time, they observed significant reductions in body fat mass and percentage, and waist-to-hip ratio without any significant changes in safety parameters. These effects are supported by other trials [33]. Treatment with herbal preparations containing yerba maté, such as YGD (Yerba maté, leaves of Ilex paraguayensis; Guarana, seeds of Paullinia cupana; and Damiana, leaves of Turnera diffusa) have also yielded reductions in body weight and body fat [28,29]. Andersen and Fogh observed a significant delay in gastric emptying after 45 days of YGD treatment taken before each main meal. They concluded this delay likely increased feelings of satiety and contributed to subsequent weight loss [29]. The study by Harrold, et al. explored the effects of YGD and soluble fermentable fiber on appetite and energy intake by feeding participants an energy-controlled breakfast, administered the treatments, and measuring appetite and energy consumption during lunch [28]. They observed significnat reductions in both hunger and energy intake with YGD treatment that was further amplified with the addition of soluble fiber. While reductions in energy intake due to appetite suppression may be a contributing factor to the weight loss we observed in the mice in this study, energy output was the focus of the work presented here, as we assessed changes in metabolic rates of skeletal muscle, adipose, and hepatic tissue.
Here, we quantified changes in mitochondrial oxygen consumption, indicative of oxidative metabolism. We expected to observe differences in their metabolic responses to MATÉ treatment because tissues inherently vary in their metabolic rates and functions. In skeletal muscle, no changes in oxygen consumption were observed with the addition of glutamate and malate, ADP, and succinate, substrates that feed into different complexes of the mitochondrial electron transport system (Figure 2A). However, significant increases in ATP concentration (Figure 2B) and ATP:O2, a measure of mitochondrial efficiency, or ATP produced per unit of oxygen consumed (Figure 2C) were observed. This increase in efficiency indicates a more “coupled” metabolic state in which oxygen consumption is more tightly coupled to energy production. Interestingly, we observed somewhat of an opposite phenomenon in white adipose tissue. Adipose oxygen consumption rates increased with yerba maté treatment, indicating an increase in metabolic rate. While ATP production increased in skeletal muscle, maté administration induced a significant decrease in ATP production in white adipose tissue (Figure 3B). When combined with the changes in oxygen consumption, ATP:O2 ratios strongly trended toward a significant decrease (Figure 3C, p = 0.050007), or a more “uncoupled”, energetically inefficient state.
Although the effect of yerba maté on some aspects of exercise, such as muscle strength, remains controversial [34,35], maté-induced changes in skeletal muscle and adipose mitochondrial bioenergetics have previously been associated with positive health outcomes – improved low and high-intensity exercise performance [36], and increased lipid oxidation and fat loss [37]. Because yerba maté contains a number of bioavailable compounds, it is impossible to discern which compounds were directly responsible for these effects. However, weight loss observed with yerba mate consumption has been previously attributed to the caffeine content of the tea, which has been demonstrated to make up 1-2% of dry leaf weight [38]. As a stimulant, caffeine has been shown to increase basal metabolic rate, improve aerobic performance and muscle oxygen content during exercise tests [39], and increase lipid oxidation [40,41]. Additionally, caffeine has been reported to increase the in vivo and in vitro expression of adipose UCP1 (uncoupling protein 1), a mitochondrial uncoupling protein that releases energy stored in the mitochondrial proton gradient in the form of heat, in brown adipose [42]. Although we did not quantify changes in white adipose UCP1 expression, the combination of our observations that yerba maté decreases white adipose mitochondrial efficiency and induces weight loss suggest mitochondrial uncoupling and loss of energy as heat. This has been shown to induce fat loss in humans and rodents and is consistent with studies that demonstrate yerba maté stimulates mitochondrial biogenesis, thermogenesis, and white adipose “browning” [26]. This browning process makes white adipocytes phenotypically more similar to brown adipocytes, which are characterized by higher mitochondrial content and the expression of UCP1 [43].
Mitochondrial dysfunction and downstream oxidative stress are considered hallmarks of aging [44] and metabolic disorders [45,46]. Cellular homeostasis of oxidizing and reducing equivalents is normally maintained via enzymatic and nonenzymatic antioxidant systems that eliminate ROS and preserve cellular function. However, when the production of reactive oxygen species (ROS) exceeds cellular antioxidant defenses, redox states shift due to an increase in oxidizing equivalents and/or a decrease in reducing equivalents to promote a more oxidizing system. Oxidizing environments promoted oxidative modification of target macromolecules such as DNA, proteins, and lipids and impair cellular function [47]. Glutathione is the most abundant intracellular antioxidant thiol [48]. The GSH/GSSG couple is, therefore, commonly used as a proxy of cellular redox state and oxidative stress due to glutathione’s high concentration and involvement in first line antioxidant defenses and as a direct free radical scavenger [47,49]. Here, we report significant increases in GSH concentration in skeletal muscle (Figure 2D) and liver (Figure 4D) in response to yerba maté treatment, which may indicate a greater antioxidant buffering capacity. However, we only observed a significant change in GSH/GSSG Eh in the liver (Figure 4F). The significant decrease in GSH/GSSG Eh observed in the liver indicates a beneficial shift toward a more reduced environment, suggesting an enhanced cellular homeostatic function. Although we did not directly quantify ROS, the shift in GSH/GSSG is suggestive of decreased oxidative stress, which could be due to a reduction in the production of ROS, an upregulation of antioxidant defenses and free radical scavenging, or possibly both. This would require further investigation; however, our findings are consistent with other studies that report yerba maté attenuates oxidative stress [38,50].
The liver is a central player in whole body energy metabolism for its roles in digestion and carbohydrate, lipid, and amino acid metabolism. The liver also has essential functions in the metabolism and elimination of xenobiotics. As such, antioxidants are necessary for hepatic function. Indeed, the liver is the largest source of GSH, which not only participates in the detoxification of xenobiotics and determines redox status, but has also been shown to regulate other essential cellular functions such as cell proliferation and apoptosis [51]. Hepatic GSH production is impaired in liver diseases such as cholestatic liver injury, nonalcoholic fatty liver disease, and hepatocellular carcinoma as well as metabolic disorders such as obesity, insulin resistance, type 2 diabetes, and cardiovascular diseases [52]. While oxidation of the GSH/GSSG Eh is associated with metabolic pathologies such as obesity, insulin resistance, type 2 diabetes, and cardiovascular disease, reduction of the GSH/GSSG Eh have been associated with hepatoprotection and a greater capacity to buffer oxidative stress. The favorable, pro-reducing shift in redox potential indicates an even greater, enhanced protection against oxidative stress.
5. Conclusions
Although the literature on the effect of yerba maté on some aspects of exercise performance, such as muscle strength remains controversial [34,35], there is a substantial amount of evidence indicating beneficial effects on lipid oxidation and fat loss in rodents and humans [26,28,29]. Together, the data presented here demonstrate metabolically beneficial, tissue-specific effects of yerba maté on healthy mice such that maté increases energy efficiency in skeletal muscle but decreases efficiency in adipose, resulting in significant weight loss. Yerba maté has been repeatedly demonstrated to be safe in humans [32,33,53], and therefore, represents a safe and accessible means of potentially improving metabolic function and protecting against obesity and other metabolic disorders.
Author Contributions
Conceptualization, J.M.H., B.T.B.; Methodology, C.M.W., E.R.S., J.M.H., and B.T.B.; Investigation, C.M.W., E.R.S., J.M.H., P.R.R., J.G.L., N.P.R. and B.T.B.; Formal Analysis, C.M.W., E.R.S. J.M.H., and B.T.B.; Writing—Original Draft Preparation, C.M.W., E.R.S. J.M.H., and B.T.B.; Writing—Review and Editing C.M.W., E.R.S., J.M.H., and B.T.B.; Project Administration, B.T.B.; Supervision, B.T.B.; Validation, C.M.W., E.R.S., J.M.H., and B.T.B.; Visualization, C.M.W., E.R.S., J.M.H., and B.T.B.; and Funding Acquisition, B.T.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by Unicity International (award number R0602556)
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable
Data Availability Statement
Data can be provided by contacting the corresponding author.
Acknowledgments
None.
Conflicts of Interest
B.T.B. receives royalties from the sale of a book about insulin resistance. All other authors declare that they have no conflicts of interest. Funders had no role in the design of the study; in the collection, analyses, or interpretation of the data; in the writing of the manuscript; or in the decision to publish the results.
References
- Shai, I.; Schwarzfuchs, D.; Henkin, Y.; Shahar, D.R.; Witkow, S.; Greenberg, I.; Golan, R.; Fraser, D.; Bolotin, A.; Vardi, H. , et al. Weight loss with a low-carbohydrate, Mediterranean, or low-fat diet. The New England journal of medicine 2008, 359, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, A.T.; Heilbronn, L.K. Metabolic impacts of altering meal frequency and timing - Does when we eat matter? Biochimie 2016, 124, 187–197. [Google Scholar] [CrossRef]
- Plotnick, G.D.; Corretti, M.C.; Vogel, R.A.; Hesslink, R., Jr.; Wise, J.A. Effect of supplemental phytonutrients on impairment of the flow-mediated brachial artery vasoactivity after a single high-fat meal. Journal of the American College of Cardiology 2003, 41, 1744–1749. [Google Scholar] [CrossRef]
- Vasanthi, H.R.; ShriShriMal, N.; Das, D.K. Phytochemicals from plants to combat cardiovascular disease. Curr Med Chem 2012, 19, 2242–2251. [Google Scholar] [CrossRef]
- Drašar, P.A.-O. Plant Secondary Metabolites Used for the Treatment of Diseases and Drug Development. 2022. [Google Scholar] [CrossRef]
- Miao, M.; Jiang, H.; Jiang, B.; Zhang, T.; Cui, S.W.; Jin, Z. Phytonutrients for controlling starch digestion: evaluation of grape skin extract. Food Chem 2014, 145, 205–211. [Google Scholar] [CrossRef]
- Weigel, H.J. Plant quality declines as CO2 levels rise. Elife 2014, 3, e03233. [Google Scholar] [CrossRef]
- Rowley, T.J.t.; Bitner, B.F.; Ray, J.D.; Lathen, D.R.; Smithson, A.T.; Dallon, B.W.; Plowman, C.J.; Bikman, B.T.; Hansen, J.M.; Dorenkott, M.R. , et al. Monomeric cocoa catechins enhance beta-cell function by increasing mitochondrial respiration. The Journal of nutritional biochemistry 2017, 49, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Galli, R.L.; Shukitt-Hale, B.; Youdim, K.A.; Joseph, J.A. Fruit polyphenolics and brain aging: nutritional interventions targeting age-related neuronal and behavioral deficits. Annals of the New York Academy of Sciences 2002, 959, 128–132. [Google Scholar] [CrossRef]
- Beecher, G.R. Phytonutrients' role in metabolism: effects on resistance to degenerative processes. Nutrition reviews 1999, 57, S3–6. [Google Scholar] [CrossRef] [PubMed]
- de Morais, E.C.; Stefanuto, A.; Klein, G.A.; Boaventura, B.C.; de Andrade, F.; Wazlawik, E.; Di Pietro, P.F.; Maraschin, M.; da Silva, E.L. Consumption of yerba mate ( Ilex paraguariensis ) improves serum lipid parameters in healthy dyslipidemic subjects and provides an additional LDL-cholesterol reduction in individuals on statin therapy. Journal of agricultural and food chemistry 2009, 57, 8316–8324. [Google Scholar] [CrossRef] [PubMed]
- Meinhart, A.D.; Damin, F.M.; Caldeirao, L.; da Silveira, T.F.F.; Filho, J.T.; Godoy, H.T. Chlorogenic acid isomer contents in 100 plants commercialized in Brazil. Food Res Int 2017, 99, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Cao, J.; Feng, Q.; Peng, J.; Hu, Y. Roles of chlorogenic Acid on regulating glucose and lipids metabolism: a review. Evid Based Complement Alternat Med 2013, 2013, 801457. [Google Scholar] [CrossRef]
- Kungel, P.; Correa, V.G.; Correa, R.C.G.; Peralta, R.A.; Sokovic, M.; Calhelha, R.C.; Bracht, A.; Ferreira, I.; Peralta, R.M. Antioxidant and antimicrobial activities of a purified polysaccharide from yerba mate (Ilex paraguariensis). Int J Biol Macromol 2018, 114, 1161–1167. [Google Scholar] [CrossRef] [PubMed]
- Arçari, D.P.; Bartchewsky, W.; dos Santos, T.W.; Oliveira, K.A.; DeOliveira, C.C.; Gotardo, É.M.; Pedrazzoli, J.; Gambero, A.; Ferraz, L.F.C.; Carvalho, P.d.O. , et al. Anti-inflammatory effects of yerba maté extract (Ilex paraguariensis) ameliorate insulin resistance in mice with high fat diet-induced obesity. Molecular and Cellular Endocrinology 2011, 335, 110–115. [Google Scholar] [CrossRef]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food chemistry 2019, 299, 125124. [Google Scholar] [CrossRef]
- Bhatti, J.S.; Bhatti, G.K.; Reddy, P.H. Mitochondrial dysfunction and oxidative stress in metabolic disorders—A step towards mitochondria based therapeutic strategies. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease 2017, 1863, 1066–1077. [Google Scholar] [CrossRef]
- Tippetts, T.S.; Winden, D.R.; Swensen, A.C.; Nelson, M.B.; Thatcher, M.O.; Saito, R.R.; Condie, T.B.; Simmons, K.J.; Judd, A.M.; Reynolds, P.R. Cigarette smoke increases cardiomyocyte ceramide accumulation and inhibits mitochondrial respiration. BMC cardiovascular disorders 2014, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.; Tippetts, T.; Anderson, M.; Holub, Z.; Moulton, E.; Swensen, A.; Prince, J.; Bikman, B. Insulin increases ceramide synthesis in skeletal muscle. Journal of diabetes research 2014, 2014. [Google Scholar] [CrossRef]
- Smith, Melissa E. ; Tippetts, Trevor S.; Brassfield, Eric S.; Tucker, Braden J.; Ockey, A.; Swensen, Adam C.; Anthonymuthu, Tamil S.; Washburn, Trevor D.; Kane, Daniel A.; Prince, John T., et al. Mitochondrial fission mediates ceramide-induced metabolic disruption in skeletal muscle. Biochemical Journal 2013, 456, 427–439. [Google Scholar] [CrossRef]
- Walton, C.M.; Jacobsen, S.M.; Dallon, B.W.; Saito, E.R.; Bennett, S.L.H.; Davidson, L.E.; Thomson, D.M.; Hyldahl, R.D.; Bikman, B.T. Ketones elicit distinct alterations in adipose mitochondrial bioenergetics. International Journal of Molecular Sciences 2020, 21, 6255. [Google Scholar] [CrossRef] [PubMed]
- Piorczynski, T.B.; Lapehn, S.; Ringer, K.P.; Allen, S.A.; Johnson, G.A.; Call, K.; Lucas, S.M.; Harris, C.; Hansen, J.M. NRF2 activation inhibits valproic acid-induced neural tube defects in mice. Neurotoxicology and Teratology 2022, 89, 107039. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.P.; Carlson, J.L.; Mody Jr, V.C.; Cai, J.; Lynn, M.J.; Sternberg Jr, P. Redox state of glutathione in human plasma. Free Radical Biology and Medicine 2000, 28, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.P. Redox potential of GSH/GSSG couple: assay and biological significance. In Methods in enzymology; Elsevier, 2002; Vol. 348, pp. 93–112. [Google Scholar]
- Saito, E.R.; Warren, C.E.; Hanegan, C.M.; Larsen, J.G.; du Randt, J.D.; Cannon, M.; Saito, J.Y.; Campbell, R.J.; Kemberling, C.M.; Miller, G.S. A novel ketone-supplemented diet improves recognition memory and hippocampal mitochondrial efficiency in healthy adult mice. Metabolites 2022, 12, 1019. [Google Scholar] [CrossRef]
- Andrade, V.M.d.M.; de Moura, A.F.; da Costa Chaves, K.; da Rocha, C.P.D.; de Andrade, C.B.V.; Trevenzoli, I.H.; Ortiga-Carvalho, T.M.; Barcellos, L.C.; Vaisman, M.; Salerno, V.P. Yerba mate consumption by ovariectomized rats alters white adipose tissue. Molecular and Cellular Endocrinology 2023, 564, 111881. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Long, Y.; Jiang, X.; Liu, Z.; Wang, D.; Zhao, Y.; Li, D.; Sun, B.-l. Beneficial effects of Yerba Mate tea (Ilex paraguariensis) on hyperlipidemia in high-fat-fed hamsters. Experimental Gerontology 2013, 48, 572–578. [Google Scholar] [CrossRef]
- Harrold, J.A.; Hughes, G.M.; O’Shiel, K.; Quinn, E.; Boyland, E.J.; Williams, N.J.; Halford, J.C.G. Acute effects of a herb extract formulation and inulin fibre on appetite, energy intake and food choice. Appetite 2013, 62, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Andersen, T.; Fogh, J. Weight loss and delayed gastric emptying following a South American herbal preparation in overweight patients. Journal of Human Nutrition and Dietetics 2001, 14, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Arçari, D.P.; Bartchewsky, W.; dos Santos, T.W.; Oliveira, K.A.; Funck, A.; Pedrazzoli, J.; de Souza, M.F.F.; Saad, M.J.; Bastos, D.H.M.; Gambero, A. , et al. Antiobesity Effects of yerba maté Extract (Ilex paraguariensis) in High-fat Diet–induced Obese Mice. Obesity 2012, 17, 2127–2133. [Google Scholar] [CrossRef]
- Silva, R.D.A.; Bueno, A.L.S.; Gallon, C.W.; Gomes, L.F.; Kaiser, S.; Pavei, C.; Ortega, G.G.; Kucharski, L.C.; Jahn, M.P. The effect of aqueous extract of gross and commercial yerba mate (Ilex paraguariensis) on intra-abdominal and epididymal fat and glucose levels in male Wistar rats. Fitoterapia 2011, 82, 818–826. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Oh, M.-R.; Kim, M.-G.; Chae, H.-J.; Chae, S.-W. Anti-obesity effects of Yerba Mate (Ilex Paraguariensis): a randomized, double-blind, placebo-controlled clinical trial. BMC Complementary and Alternative Medicine 2015, 15, 338. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Ko, J.; Storni, C.; Song, H.J.; Cho, Y.G. Effect of green mate in overweight volunteers: A randomized placebo-controlled human study. Journal of Functional Foods 2012, 4, 287–293. [Google Scholar] [CrossRef]
- Lobo, P.C.B.; da Silva, D.D.; Pimentel, G.D. Acute Supplementation of Yerba Mate Extract Did Not Change Muscle Strength in Physically Active Men Following the Strength Muscle Test: A Pilot Clinical Trial. Nutrients 2022, 14, 2619. [Google Scholar] [CrossRef] [PubMed]
- Panza, V.P.; Diefenthaeler, F.; Tamborindeguy, A.C.; de Quadros Camargo, C.; de Moura, B.M.; Brunetta, H.S.; Sakugawa, R.L.; de Oliveira, M.V.; de Oliveira Puel, E.; Nunes, E.A. Effects of mate tea consumption on muscle strength and oxidative stress markers after eccentric exercise. British Journal of Nutrition 2016, 115, 1370–1378. [Google Scholar] [CrossRef]
- Krolikowski, T.C.; Borszcz, F.K.; Panza, V.P.; Bevilacqua, L.M.; Nichele, S.; da Silva, E.L.; Amboni, R.D.M.C.; Guglielmo, L.G.A.; Phillips, S.M.; de Lucas, R.D. , et al. The Impact of Pre-Exercise Carbohydrate Meal on the Effects of Yerba Mate Drink on Metabolism, Performance, and Antioxidant Status in Trained Male Cyclists. Sports Medicine - Open 2022, 8, 93. [Google Scholar] [CrossRef]
- Alkhatib, A. Yerba Maté (Illex Paraguariensis) ingestion augments fat oxidation and energy expenditure during exercise at various submaximal intensities. Nutrition & Metabolism 2014, 11, 42. [Google Scholar] [CrossRef]
- Gawron-Gzella, A.A.-O.; Chanaj-Kaczmarek, J.A.-O.; Cielecka-Piontek, J.A.-O. Yerba Mate-A Long but Current History. 2021. [Google Scholar] [CrossRef] [PubMed]
- Ruíz-Moreno, C.; Lara, B.; Brito de Souza, D.; Gutiérrez-Hellín, J.; Romero-Moraleda, B.; Cuéllar-Rayo, Á.; Del Coso, J. Acute caffeine intake increases muscle oxygen saturation during a maximal incremental exercise test. British Journal of Clinical Pharmacology 2020, 86, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Bracco, D.; Ferrarra, J.M.; Arnaud, M.J.; Jequier, E.; Schutz, Y. Effects of caffeine on energy metabolism, heart rate, and methylxanthine metabolism in lean and obese women. American Journal of Physiology-Endocrinology and Metabolism 1995, 269, E671–E678. [Google Scholar] [CrossRef]
- Jo, E.; Lewis, K.L.; Higuera, D.; Hernandez, J.; Osmond, A.D.; Directo, D.J.; Wong, M. Dietary Caffeine and Polyphenol Supplementation Enhances Overall Metabolic Rate and Lipid Oxidation at Rest and After a Bout of Sprint Interval Exercise. The Journal of Strength & Conditioning Research 2016, 30. [Google Scholar]
- Velickovic, K.; Wayne, D.; Leija, H.A.L.; Bloor, I.; Morris, D.E.; Law, J.; Budge, H.; Sacks, H.; Symonds, M.E.; Sottile, V. Caffeine exposure induces browning features in adipose tissue in vitro and in vivo. Scientific Reports 2019, 9, 9104. [Google Scholar] [CrossRef] [PubMed]
- Bartelt, A.; Heeren, J. Adipose tissue browning and metabolic health. Nature Reviews Endocrinology 2014, 10, 24–36. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C.S. Oxidative stress and metabolic disorders: Pathogenesis and therapeutic strategies. Life sciences 2016, 148, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Rotariu, D.; Babes, E.E.; Tit, D.M.; Moisi, M.; Bustea, C.; Stoicescu, M.; Radu, A.-F.; Vesa, C.M.; Behl, T.; Bungau, A.F. Oxidative stress–Complex pathological issues concerning the hallmark of cardiovascular and metabolic disorders. Biomedicine & Pharmacotherapy 2022, 152, 113238. [Google Scholar]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alexandria Journal of Medicine 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Biswas, S.K.; Rahman, I. Environmental toxicity, redox signaling and lung inflammation: the role of glutathione. 2010. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.; Hansen, J.M. Oxidative stress, thiols, and redox profiles. Developmental Toxicology: Methods and Protocols 2012, 325–346. [Google Scholar]
- Nogueira Silva Lima, M.T.; Boulanger, E.; Tessier, F.J.; Takahashi, J.A. Hibiscus, rooibos, and yerba mate for healthy aging: A review on the attenuation of in vitro and in vivo markers related to oxidative stress, glycoxidation, and neurodegeneration. Foods 2022, 11, 1676. [Google Scholar] [CrossRef]
- Lu, S.C. Dysregulation of glutathione synthesis in liver disease. Liver Research 2020, 4, 64–73. [Google Scholar] [CrossRef]
- Ballatori, N.; Krance Sm Fau - Notenboom, S.; Notenboom S Fau - Shi, S.; Shi S Fau - Tieu, K.; Tieu K Fau - Hammond, C.L.; Hammond, C.L. Glutathione dysregulation and the etiology and progression of human diseases. 2009. [Google Scholar] [CrossRef]
- Ulbrich, N.C.M.; do Prado, L.L.; Barbosa, J.Z.; Araujo, E.M.; Poggere, G.; Motta, A.C.V.; Prior, S.A.; Magri, E.; Young, S.D.; Broadley, M.R. Multi-elemental Analysis and Health Risk Assessment of Commercial Yerba Mate from Brazil. Biological Trace Element Research 2022, 200, 1455–1463. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Yerba maté consumption prevents weigh gain in male and female mice. Body weight was measured weekly over the four weeks of yerba maté treatment and normalized to initial weights at week = 0. * p < 0.05.
Figure 1.
Yerba maté consumption prevents weigh gain in male and female mice. Body weight was measured weekly over the four weeks of yerba maté treatment and normalized to initial weights at week = 0. * p < 0.05.
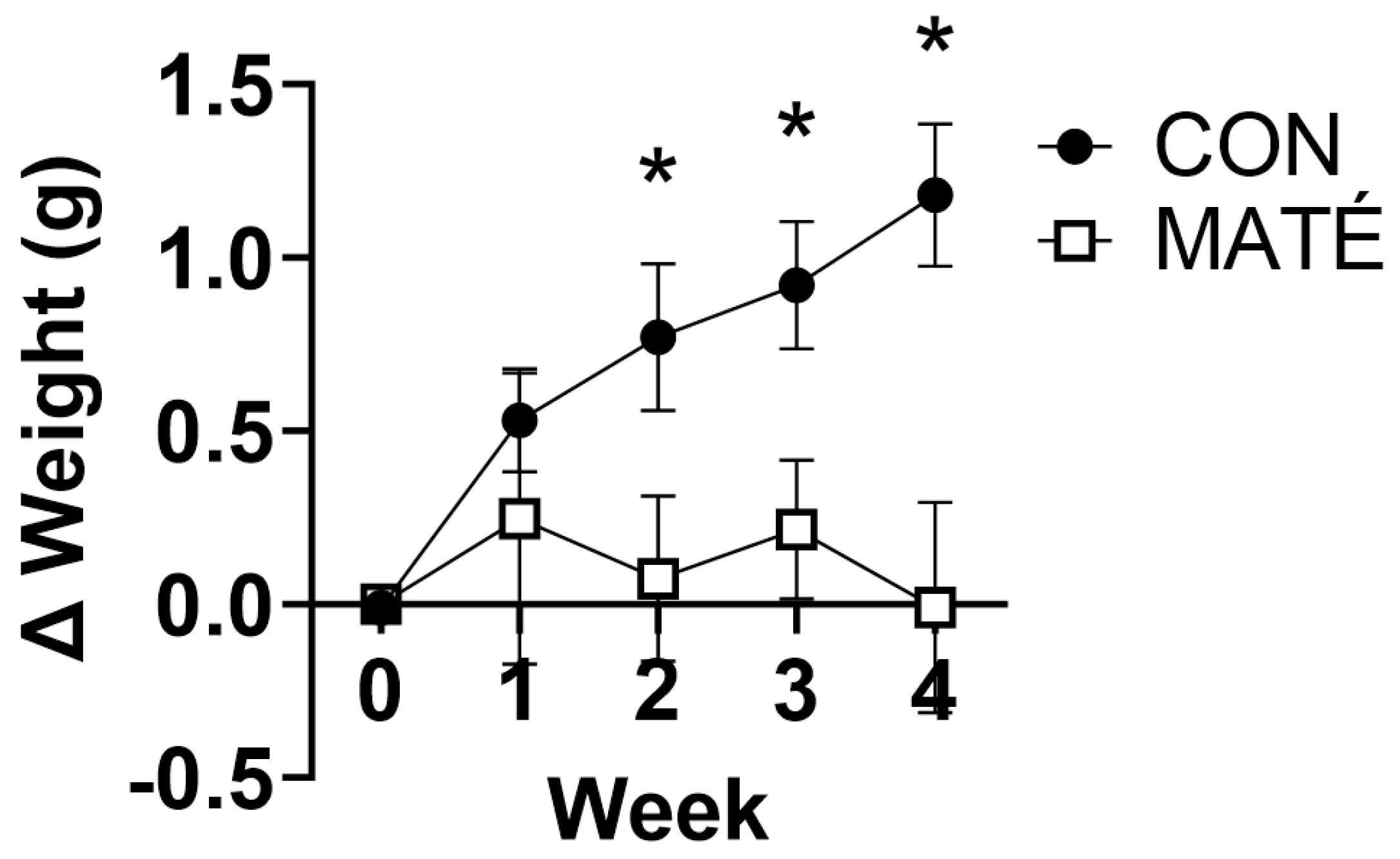
Figure 2.
Yerba maté consumption increases mitochondrial efficiency in skeletal muscle from male and female mice. Rates of mitochondrial oxygen consumption (A) were measured via a substrate–uncoupler–inhibitor–titration protocol from gastrocnemius tissue following four weeks of yerba maté supplementation. The protocol included the addition of glutamate and malate (GM), ADP, succinate (S), and carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone (FCCP). ATP concentration was quantified (B) and ATP:O2 flux ratios (C) were calculated (CON, n = 6; MATÉ, n = 6). Concentrations of reduced (GSH, D) and oxidized (GSSG, E) glutathione were quantified via reverse-phase high-performance liquid chromatography. GSH/GSSG Eh redox potentials were calculated (F; CON, n = 8; MATÉ, n = 8). * p < 0.05, ** p < 0.01.
Figure 2.
Yerba maté consumption increases mitochondrial efficiency in skeletal muscle from male and female mice. Rates of mitochondrial oxygen consumption (A) were measured via a substrate–uncoupler–inhibitor–titration protocol from gastrocnemius tissue following four weeks of yerba maté supplementation. The protocol included the addition of glutamate and malate (GM), ADP, succinate (S), and carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone (FCCP). ATP concentration was quantified (B) and ATP:O2 flux ratios (C) were calculated (CON, n = 6; MATÉ, n = 6). Concentrations of reduced (GSH, D) and oxidized (GSSG, E) glutathione were quantified via reverse-phase high-performance liquid chromatography. GSH/GSSG Eh redox potentials were calculated (F; CON, n = 8; MATÉ, n = 8). * p < 0.05, ** p < 0.01.
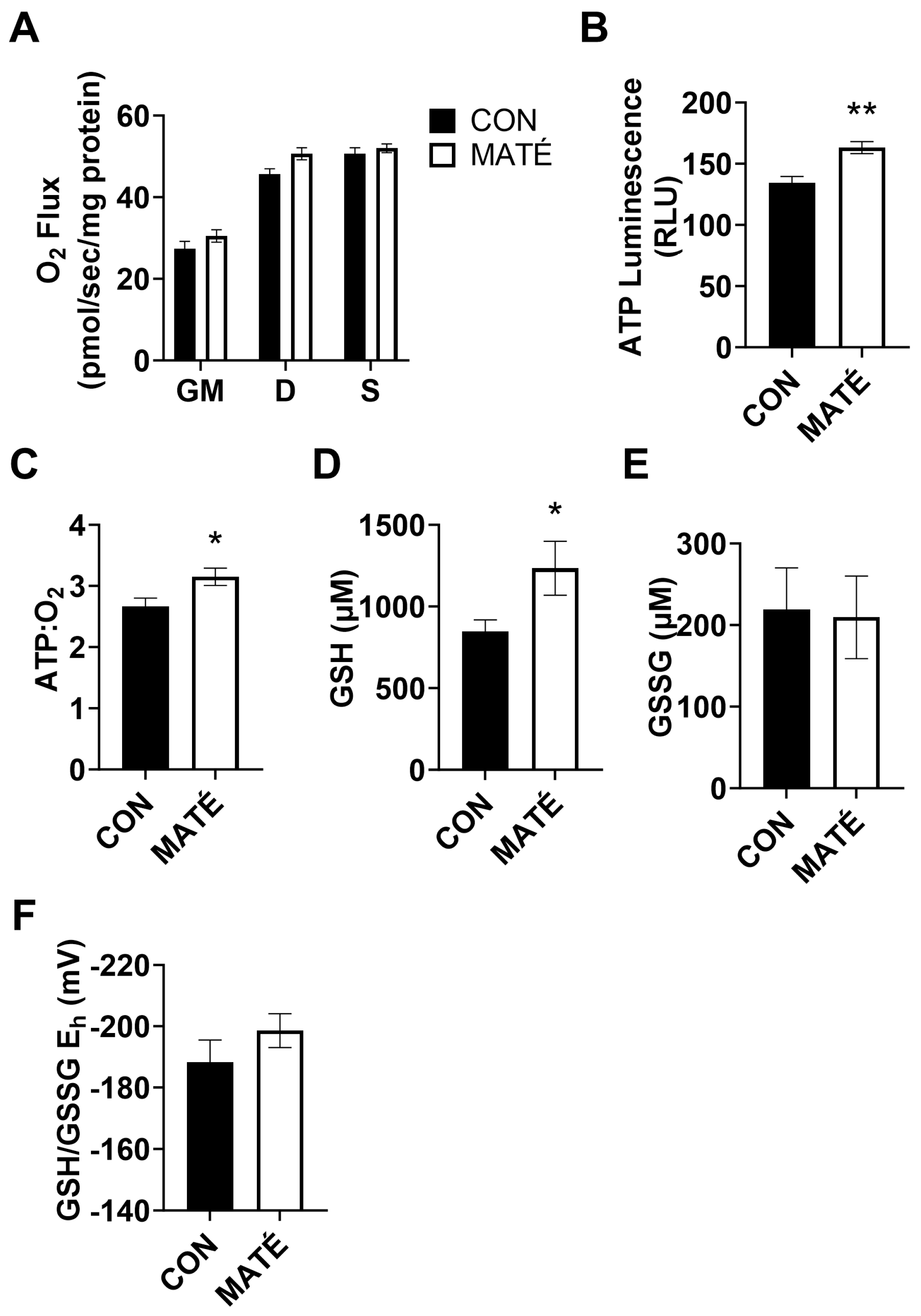
Figure 3.
Yerba maté consumption decreases mitochondrial efficiency in white adipose tissue. Rates of mitochondrial oxygen consumption (A) were measured via a substrate–uncoupler–inhibitor–titration protocol from gastrocnemius tissue following four weeks of yerba maté supplementation. The protocol included the addition of glutamate and malate (GM), ADP, succinate (S), and carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone (FCCP). ATP concentration was quantified (B) and ATP:O2 flux ratios (C) were calculated (CON, n = 6; MATÉ, n = 6). Concentrations of reduced (GSH, D) and oxidized (GSSG, E) glutathione were quantified via reverse-phase high-performance liquid chromatography. GSH/GSSG Eh redox potentials were calculated (F; CON, n = 8; MATÉ, n = 8). * p < 0.05.
Figure 3.
Yerba maté consumption decreases mitochondrial efficiency in white adipose tissue. Rates of mitochondrial oxygen consumption (A) were measured via a substrate–uncoupler–inhibitor–titration protocol from gastrocnemius tissue following four weeks of yerba maté supplementation. The protocol included the addition of glutamate and malate (GM), ADP, succinate (S), and carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone (FCCP). ATP concentration was quantified (B) and ATP:O2 flux ratios (C) were calculated (CON, n = 6; MATÉ, n = 6). Concentrations of reduced (GSH, D) and oxidized (GSSG, E) glutathione were quantified via reverse-phase high-performance liquid chromatography. GSH/GSSG Eh redox potentials were calculated (F; CON, n = 8; MATÉ, n = 8). * p < 0.05.
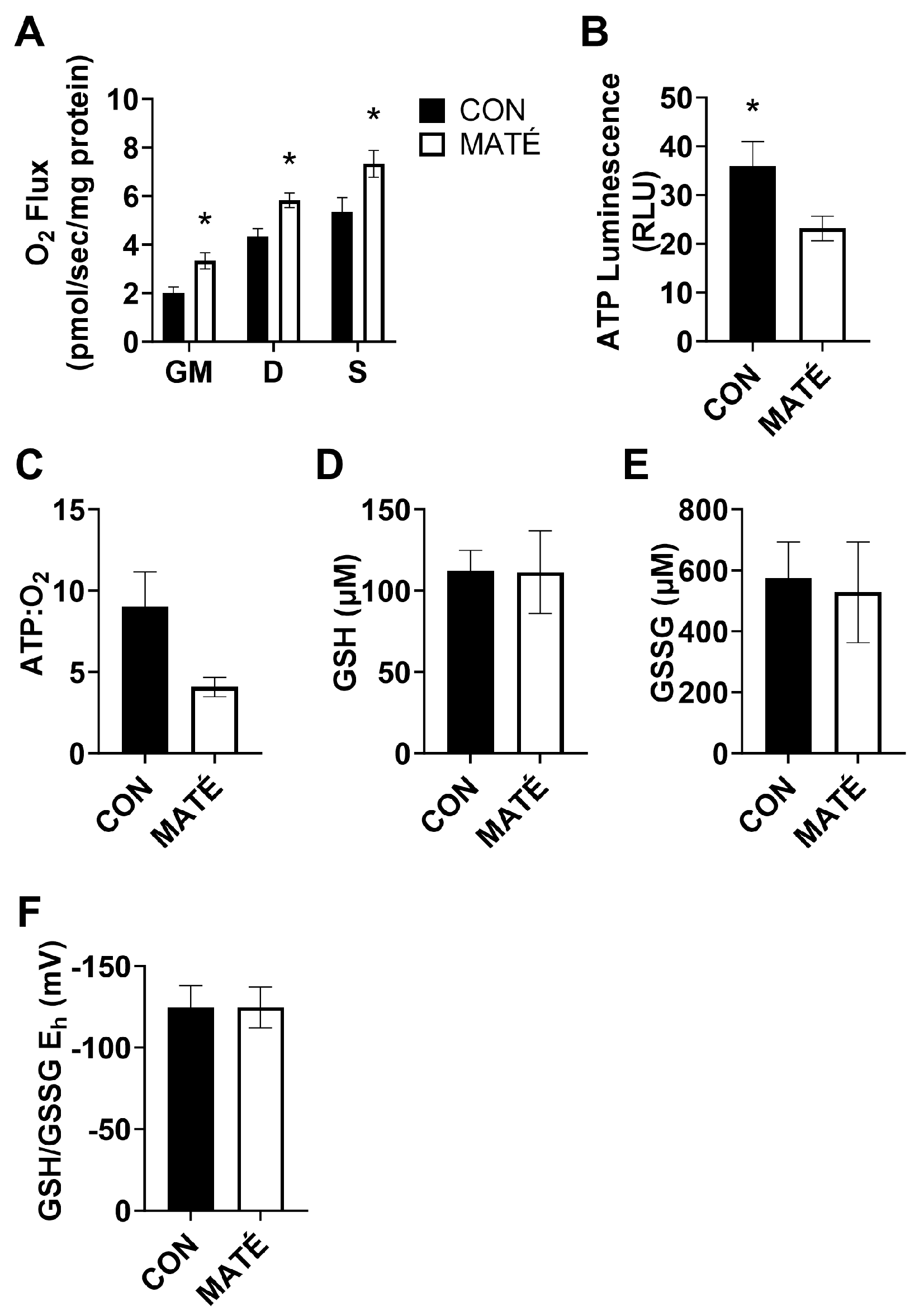
Figure 4.
Yerba maté consumption alters hepatic redox potential but not mitochondrial efficiency. Rates of mitochondrial oxygen consumption (A) were measured via a substrate–uncoupler–inhibitor–titration protocol from gastrocnemius tissue following four weeks of yerba maté supplementation. The protocol included the addition of glutamate and malate (GM), ADP, succinate (S), and carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone (FCCP). ATP concentration was quantified (B) and ATP:O2 flux ratios (C) were calculated (CON, n = 6; MATÉ, n = 6). Concentrations of reduced (GSH, D) and oxidized (GSSG, E) glutathione were quantified via reverse-phase high-performance liquid chromatography. GSH/GSSG Eh redox potentials were calculated (F; CON, n = 8; MATÉ, n = 8). * p < 0.05.
Figure 4.
Yerba maté consumption alters hepatic redox potential but not mitochondrial efficiency. Rates of mitochondrial oxygen consumption (A) were measured via a substrate–uncoupler–inhibitor–titration protocol from gastrocnemius tissue following four weeks of yerba maté supplementation. The protocol included the addition of glutamate and malate (GM), ADP, succinate (S), and carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone (FCCP). ATP concentration was quantified (B) and ATP:O2 flux ratios (C) were calculated (CON, n = 6; MATÉ, n = 6). Concentrations of reduced (GSH, D) and oxidized (GSSG, E) glutathione were quantified via reverse-phase high-performance liquid chromatography. GSH/GSSG Eh redox potentials were calculated (F; CON, n = 8; MATÉ, n = 8). * p < 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



