
Submitted:
12 September 2023
Posted:
14 September 2023
You are already at the latest version
Abstract
Food-grinding occurs in rodents and is influenced by multiple-factors. However, the factors affecting this behavior remain unclear. In this study, we aimed to investigate the effect of food restriction on food grinding by Brandt’s voles (Lasiopodomys brandtii), as well as the potential role of the gut microbiota in this process, through a comparison of the variations between voles with different food supplies. Food restriction reduced the relative amount of ground food to a greater extent than it lowered the relative food consumption, and altered the abundance of Staphylococcus, Aerococcus, Jeotgalicoccus, and Un--s-Clostridiaceae bacterium GM1. Strong correlations between the ground-to-consumed food ratio and the abundance of fecal microbiota were found for Un--s-Clostridiaceae bacterium GM1 and Aerococcus. The content of fecal acetate and propionate for the 7.5 g-food supply group was lower than that for the 15 g-food supply group. Further, the relative amount of ground food and ground-to-consumed food ratio were both positively correlated with the acetate content. Our study indicated that food restriction can effectively inhibit food grinding. Further, Un--s-Clostridiaceae bacterium GM1 abundance, Aerococcus abundance, and acetate content were strongly related to food grinding. Variations in gut microbial abundance and metabolite short-chain fatty acid content induced by food restriction likely promote the inhibition of food grinding. These results could potentially provide guidance for reducing food waste during laboratory rodent maintenance.
Keywords:
food grinding; Brandt’s voles; food restriction; short-chain fatty acids; fecal microbiota
1. Introduction
Food grinding, also known as food spilling or food wasting, is a behavior of captive rodents that involves grinding food into orts that are left uneaten on the floor of cages [1,2], or a behavior of wild herbivorous rodents involving cutting and discarding substantial amounts of grass or herbaceous plant material [3,4]. Food grinding is regarded as a characteristic or obsessive behavior that occurs owing to the tedium of the environment or one that is stimulated by an optimal food-intake strategy, in which rodents choose a portion of food components through grinding to render their net energy intake as large as possible [1,5]. Food grinding is also affected by the hardness of food [1,6], genetic components [4], sleep deprivation [7,8], diet quality (cellulose content), and caloric restriction (food supply) (Cameron and Speakman 2010). However, the factors and mechanisms affecting food-grinding behavior are still poorly understood. Specifically, how the food supply influences food-grinding behavior and its underlying mechanisms are not clear. Understanding the influencing factors and mechanisms of food-grinding behavior is important because this behavior results in a large amount of forage being wasted during laboratory rodent feeding. Ground food, as the leftovers of wild rodents, also plays an important role in grassland ecosystem functioning [3].
In our previous study, we first hypothesized that the gut microbiota could be related to food-grinding behavior and demonstrated that the relative abundances of gut microbiome components and their metabolites are significantly correlated with food-grinding behavior in captive Brandt’s voles (Lasiopodomys brandtii) [9], a small herbivorous mammalian species that exhibits food-wasting behavior, and is found in the grasslands of Inner Mongolia, China [10,11,12,13]. In this study, we reduced the food supply and investigated the response of voles exhibiting high-level food-grinding to the reduction in the food supply. Specially, we assessed the variations in their body mass, consumed and ground food, ground-to-consumed food ratio, fecal short-chain fatty acids (SCFAs), fecal microbial alpha and beta diversity, and the predicted functions of the fecal microbiota, as well as the correlations among these parameters. The ultimate aim was to uncover the effect of food supply variations on food-grinding behavior and further determine the relation between the gut microbiota and this behavior. Our findings provide guidance for reducing food waste during laboratory rodent rearing and provide new insights into the mechanisms through which the gut microbiota affect food-grinding behavior. This will ultimately broaden our understanding of the effects of vegetation biomass on food-wasting behavior in wild rodents.
2. Materials and Methods
2.1. Animals and Housing
The environmental conditions for breeding Brandt’s voles (captured from the grasslands of Xilinhot, China) at the College of Bioscience and Biotechnology, Yangzhou University, China, were as follows: air temperature, 22 ± 1 °C; photoperiod, 12 L:12 D (light period: 06:00–18:00); and relative humidity, 50 ± 5%. All voles were offered water ad libitum and were freely fed rodent pellet chow (Yizheng Animal Biotechnology Co., LTD, Yangzhou, China). At 21 days of age, young voles were weaned and separately housed in plastic cages until they were 60-days-old, which represents the adult stage for this species (Wan et al. 2002). The food ground by each vole, from 21 to 60 days of age, was recorded.
2.2. Experimental Design
At 60 days of age, 23 male voles, including voles that ground both large and small amounts of food according to the record, were randomly chosen to perform the 15 g-daily food supply experiment for 2 weeks. In each cage, an iron mesh with 6 × 6 mm holes was placed 1 cm above the cage floor, filtering the ground chow to the cage floor, to prevent the voles from ingesting the ground chow. Of the 23 male voles, six that ground a higher average daily amount of food were marked as the high-level food-grinding group based on the upper 27% rule [14]. Thereafter, we conducted a food restriction experiment using the voles in this group, in which the chow supply was reduced from 15 to 7.5 g for 2 weeks, representing a final value below the average daily chow intake used for the 15 g-food supply experiment. Daily amounts of consumed and ground food were recorded, and body mass was measured every 3 days in both the 15- and 7.5 g-food supply groups. Fresh fecal pellets were collected from each vole twice during the last week of each experiment and placed in sterile tubes, which were then immediately frozen in liquid nitrogen and stored at −70 °C. The daily ground food was dried in an oven set at 40 °C, to a constant weight, and then weighed. Calculations of daily food consumption were conducted as described by Shen, Shi, Gu, Wei, Yang and Dai [9]. The average daily amounts of consumed and ground food for each experiment were calculated separately. Relative food consumption, relative ground food, and the ratio of ground to consumed food were calculated as described by Shen, Shi, Gu, Wei, Yang and Dai [9]. The change in relative ground food was obtained as the difference between the relative ground food in the 7.5 g-food supply group and that in the 15 g-food supply group divided by that in the 15 g-food supply group. The same formula was used to calculate the change in relative food consumption. The difference in body mass between days 14 and 1 divided by that on day 1 for both the 15- and 7.5 g- food supply experiments was regarded as the body mass growth rate. All protocol and procedures were approved by the Animal Care and Use Committee of the Faculty of Veterinary Medicine at the Yangzhou University (No. NSFC2020-SKXY-6).
2.2.1. 16S rRNA Gene Sequencing and Bioinformatic Analysis of Fecal Microbiota
Two fecal samples collected during the second week of each experiment from each vole were mixed together as the fecal sample of the individual and then sent to Novogene Co. Ltd., Beijing, China, for DNA extraction and 16S rRNA gene sequencing of the fecal microbiota, according to the procedures of Dai, et al. [15]. Uparse software (v7.0.1001) were used to analyze the sequence [16]. The sequence pairs were identified as different bacterial operational taxonomic units (OTUs) according to a minimum identity threshold of 97%. Taxonomic information regarding OTU sequences was annotated referring to the RDP database (11_4 release) according to the method of the Mothur algorithm (Threshold:0.8–1) [17]. Sequence data are available at the NIH Sequence Read Archive under Bioproject ID PRJNA868862. Alpha and beta diversity analyses were performed as described by Dai, Chen, Liu, Liu, Jiang, Xu, Wang, Yang and Wei [15]. The observed species (OBSP), Chao1, abundance-based coverage estimator (ACE), and Shannon and Simpson indices were used to determine the alpha diversity of the fecal microbiota. All index calculations were performed using Quantitative Insights into Microbial Ecology (QIIME, Version 1.7.0). Dominant biomarkers in fecal microbiota for the 15- and 7.5 g-food supply groups were determined based on the linear discriminant analysis (LDA) effect size (LEfSe; LDA score > 3) using the online LEfSe program (http://huttenhower.sph.harvard.edu/galaxy/) [15].
2.2.2. Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Prediction of Fecal Microbiota
The OTUs were clustered using QIIME 2 (version 2023.5) referring to the Greengene data-resource (version GG 13.5) [18] based on the OTU data of the 16S rRNA gene sequencing. Then, the biological functions of the fecal microbiota of voles for the 15- and 7.5 g-food supply groups were predicted using the PICRUSt program (http://huttenhower.sph.harvard.edu/galaxy/), according to the information of clustered OTUs, and annotated using the KEGG pathway database (Dai et al. 2022).
2.2.3. SCFA Assay
The content of seven SCFAs in the feces of voles, including acetate, propionate, isobutyrate, butyrate, isovalerate, valerate, and caproate, were identified and quantified by headspace gas chromatography (Agilent 7890A-7697A, Agilent Technologies, Böblingen, Germany) based on a protocol detailed by Shen, Shi, Gu, Wei, Yang and Dai [9].
2.2.4. Statistical Analysis
We used the non-parametric Wilcoxon signed-rank test in SPSS Statistics (version 22; IBM Corp., Armonk, NY, USA) to investigate the differences in the number of total tags, taxon tags and OTUs, OTU biomarkers, alpha diversity indices, SCFA contents, KEGG pathway enrichment, body mass, body mass growth rate, relative food consumption, relative ground food, and ground-to-consumed food ratios in food-grinding voles between the 15- and 7.5 g-food supply groups, as well as the differences between changes in the relative ground food and relative food consumption. Variations in the beta diversity of the fecal microbiota were tested by performing PERMANOVA (permutational multivariate analysis of variance), with Bray–Curtis distance matrix analysis conducted using the nested adonis function in the “vegan” package in R ver. 4.0.4. Spearman’s rank correlations among parameters were determined using R ver. 4.0.4. Correlations were considered significant when the false discovery rate-adjusted P-value was < 0.05. The level of statistical significance was determined at P < 0.05.
3. Results
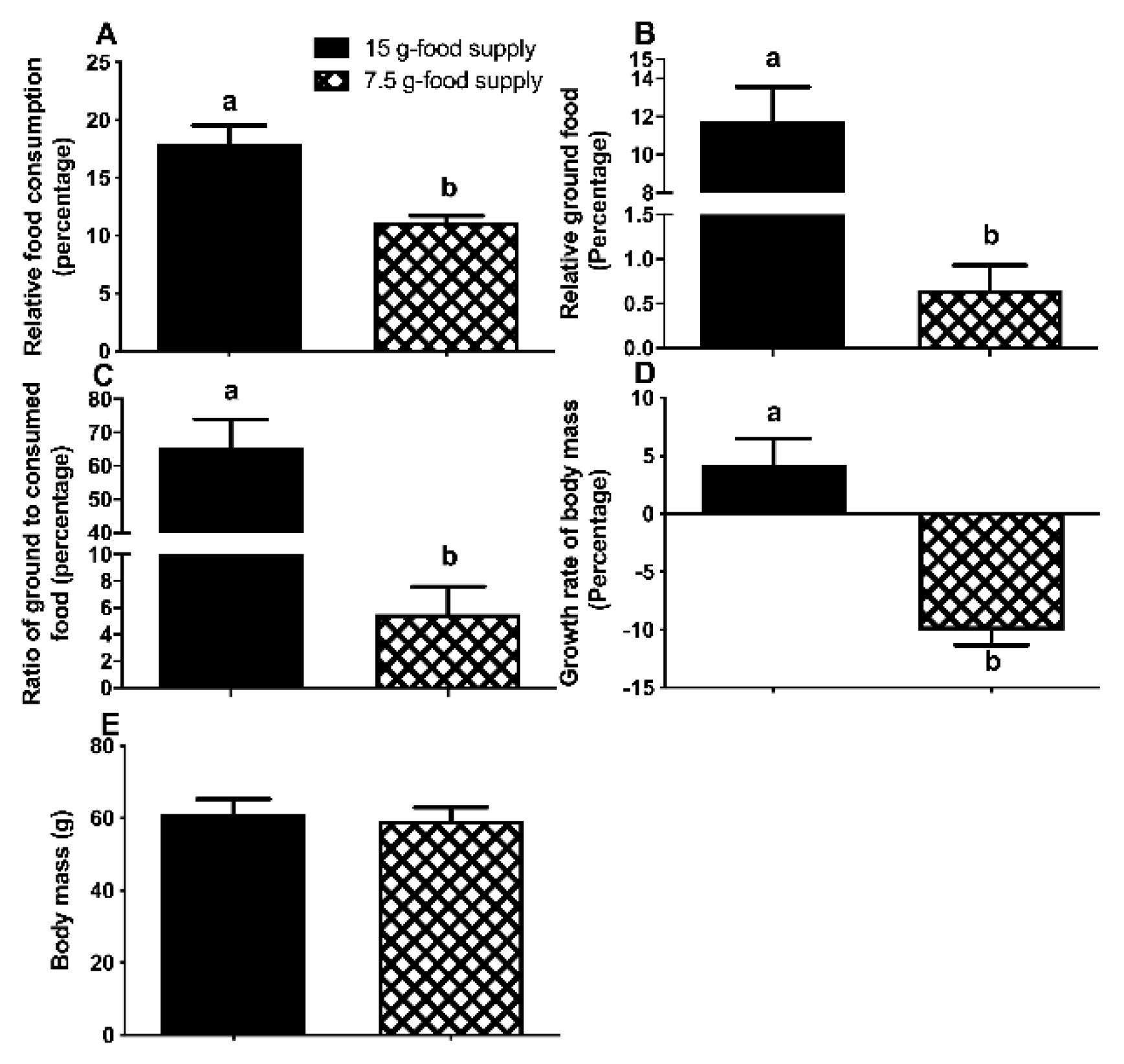
The relative food consumption and relative amount of ground food, the ratio of ground to consumed food, and the body mass growth rate significantly decreased for the high-level food-grinding voles when the food supply was reduced from 15 to 7.5 g (P = 0.028, for all cases; Figure 1A-C, E). However, body weight did not sharply differ between the 15- and 7.5 g-food supply groups (P = 0.116; Figure 1D). The change in relative ground food was greater than the change in the relative food consumption (P = 0.028) when the food supply was reduced from 15 to 7.5 g (Figure 2).
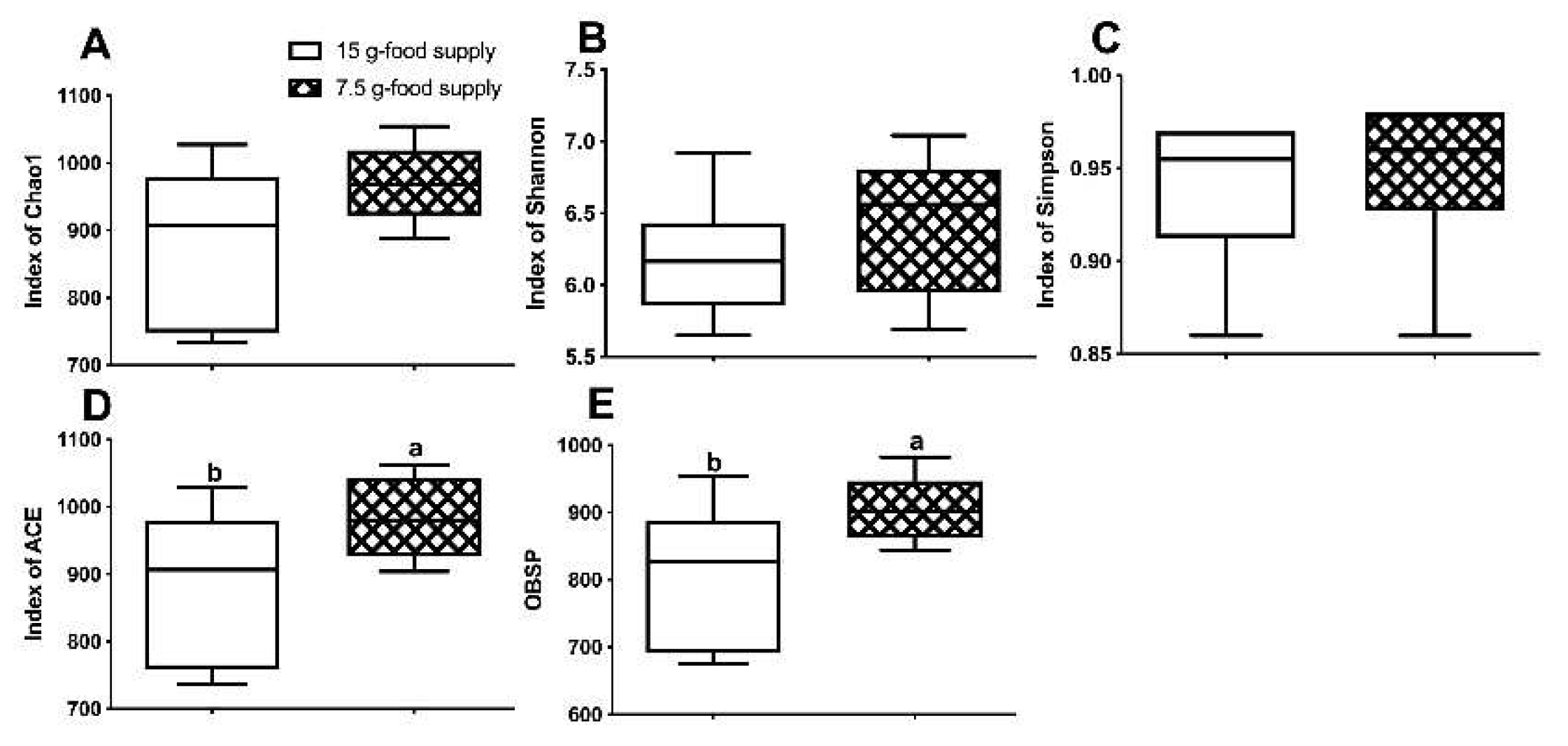
In both the 15- and 7.5 g-food supply groups, the values of the average Goods coverage were as high as 99.75%, indicating that the16S rDNA sequencing method could identify the vast majority of taxa among the microbiota present in the fecal samples (Table 1). The numbers of total tags and taxon tags for the 7.5 g-food supply group were not significantly different from those for the 15 g-food supply group (P = 0.116 and 0.917, respectively; Table 1). However, the number of OTUs for the 15 g-food supply group was significantly lower than that for the 7.5 g-food supply group (P = 0.028; Table 1). Although no obvious differences between the two food supply groups were detected with respect to the Chao1 (P = 0.116), Shannon (P = 0.249), or Simpson (P = 0.753) indices (Figure 3A, D-E), the ACE index (P = 0.046) and OBSP values (P = 0.028) (Figure 3B-C) for the 15 g-food supply group were lower than those for the 7.5 g-food supply group. The beta diversity of the fecal microbiota did not sharply vary after the food supply was reduced to 7.5 g (F = 1.070, P = 0.34).
Figure 1.
Differences of relative food consumption (A), relative ground food (B), food ground to consumption ratio (C), body mass (D), and growth rate of body mass (E) between the 15 g- and 7.5 g-food supply groups of Brandt’s voles. Error bars indicate standard errors. Different letters connect bars with significant differences at P < 0.05 (n = 6).
Figure 1.
Differences of relative food consumption (A), relative ground food (B), food ground to consumption ratio (C), body mass (D), and growth rate of body mass (E) between the 15 g- and 7.5 g-food supply groups of Brandt’s voles. Error bars indicate standard errors. Different letters connect bars with significant differences at P < 0.05 (n = 6).
The biomarkers for the Bacilli class, Bacillales and Lactobacillales orders, Staphylococcaceae and Aerococcaceae families, and Staphylococcus, Aerococcus, and Jeotgalicoccus genera (Figure 4A-B) were enriched for the 15 g-food supply group. The biomarkers for the Flavobacteriia and Betaproteobacteria classes, Flavobacteriales and Burkholderiales orders, Flavobacteriaceae family, and Un--s-Clostridiaceae bacterium GM1 genus (Figure 4A-B) were enriched for the 7.5 g-food supply group. The relative abundances of Bacilli (P = 0.028), Bacillales (P = 0.028), Lactobacillales (P = 0.046), Staphylococcaceae (P = 0.028), Aerococcaceae (P = 0.028), Staphylococcus (P = 0.028), Aerococcus (P = 0.028), and Jeotgalicoccus (P = 0.028) for the 15 g-food supply group were all higher than those for the 7.5 g-food supply group (Figure 5A-D). Moreover, the relative abundances of Flavobacteriia (P = 0.028), Betaproteobacteria (P = 0.028), Flavobacteriales (P = 0.028), Burkholderiales (P = 0.028), Flavobacteriaceae (P = 0.028), and Un--s-Clostridiaceae bacterium GM1 (P = 0.039) were all higher for the 7.5 g-food supply group than for the 15 g-food supply group (Figure 5A-D).
Figure 2.
Differences between the changes of relative food consumption and relative ground food in Brandt’s vole when food supply reduced from 15 g to 7.5 g. Error bars indicate standard errors. Different letters connect bars with significant differences at P < 0.05 (n = 6).
Figure 2.
Differences between the changes of relative food consumption and relative ground food in Brandt’s vole when food supply reduced from 15 g to 7.5 g. Error bars indicate standard errors. Different letters connect bars with significant differences at P < 0.05 (n = 6).
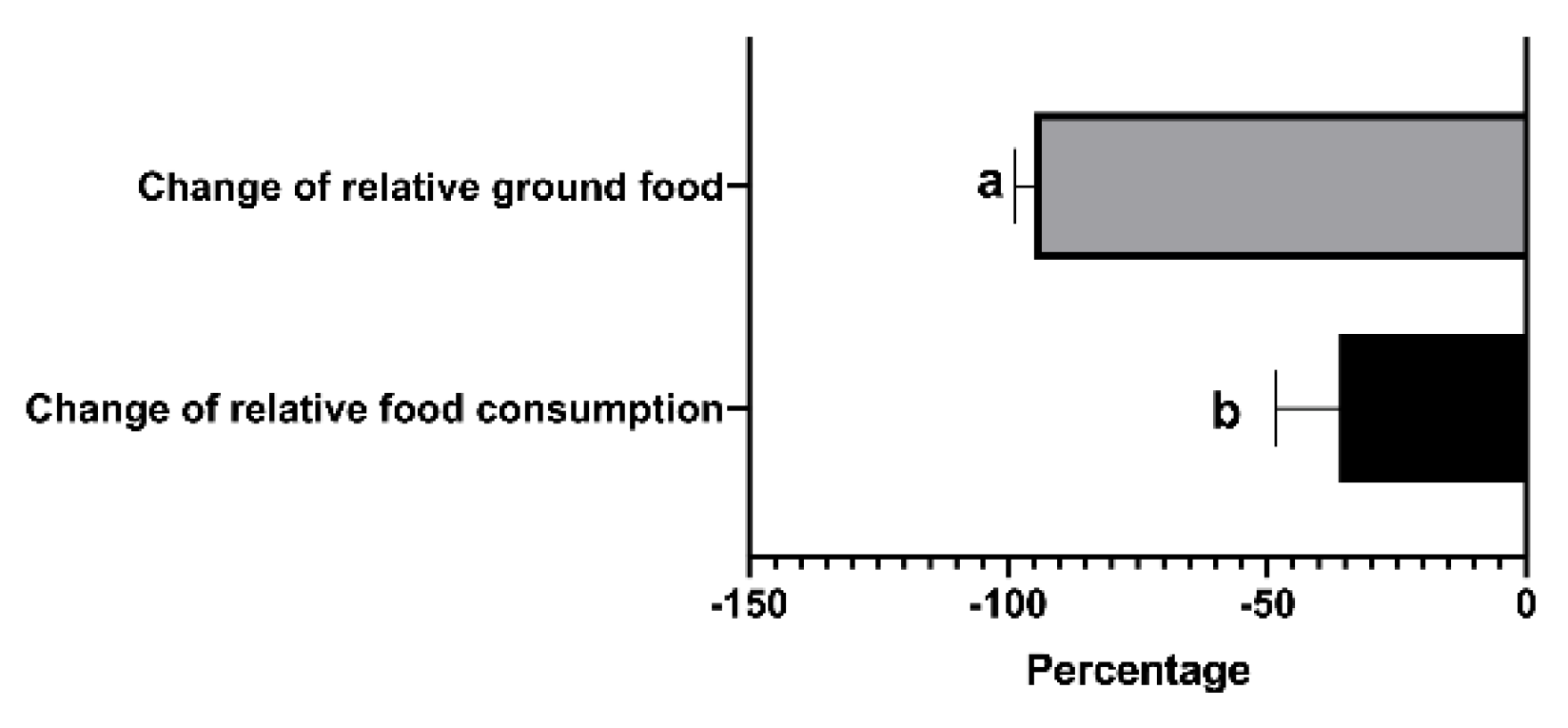
Figure 3.
Differences in the fecal microbiota composition between the 15 g- and 7.5 g-food supply groups of Brandt’s voles. (A) Chao1; (B) abundance-based coverage estimator (ACE); (C) observed species (OBSP); (D) Shannon index; (E) Simpson index. Error bars indicate standard errors. Different letters connect bars with significant differences at P < 0.05 (n = 6).
Figure 3.
Differences in the fecal microbiota composition between the 15 g- and 7.5 g-food supply groups of Brandt’s voles. (A) Chao1; (B) abundance-based coverage estimator (ACE); (C) observed species (OBSP); (D) Shannon index; (E) Simpson index. Error bars indicate standard errors. Different letters connect bars with significant differences at P < 0.05 (n = 6).
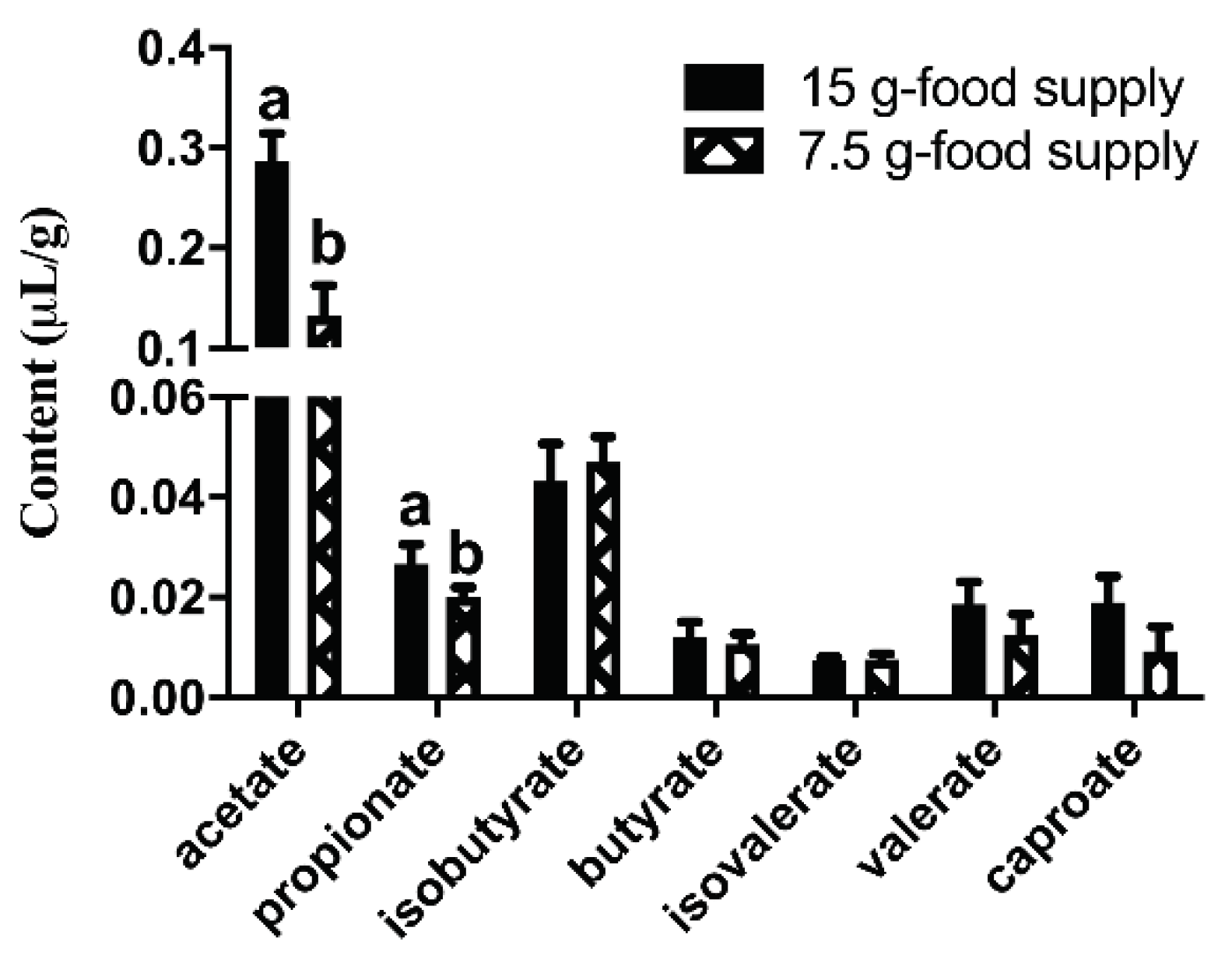
The KEGG pathways associated with Staphylococcus aureus infection, the ubiquitin system, and alpha-linolenic acid metabolism (P = 0.046, 0.028, and 0.028, respectively) were more enriched for the 15 g-food supply group compared to that for the 7.5 g-food supply group (Figure 6). Further, the acetate and propionate content were lower for the 7.5 g-food supply group than for the 15 g-food supply group (P = 0.028 and 0.042, respectively; Figure 7). Meanwhile, the isobutyrate, butyrate, isovalerate, valerate, and caproate content did not significantly differ between the 15 g- and 7.5 g-food supply groups (P = 0.600, 0.416, 0.581, 0.172, and 0.173, respectively; Figure 7).
Figure 4.
Biomarker of taxa with statistically significant differences in relative abundance selected in the fecal microbiota between the 15 g- and 7.5 g-food supply groups of Brandt’s voles (n = 6). (A) Histogram displaying the differential taxa using the linear discriminant analysis (LDA) effect size (LEfSe) analysis with a LDA score significant threshold > 3; (B) Cladogram displaying the phylogenetic position of differential taxa.
Figure 4.
Biomarker of taxa with statistically significant differences in relative abundance selected in the fecal microbiota between the 15 g- and 7.5 g-food supply groups of Brandt’s voles (n = 6). (A) Histogram displaying the differential taxa using the linear discriminant analysis (LDA) effect size (LEfSe) analysis with a LDA score significant threshold > 3; (B) Cladogram displaying the phylogenetic position of differential taxa.
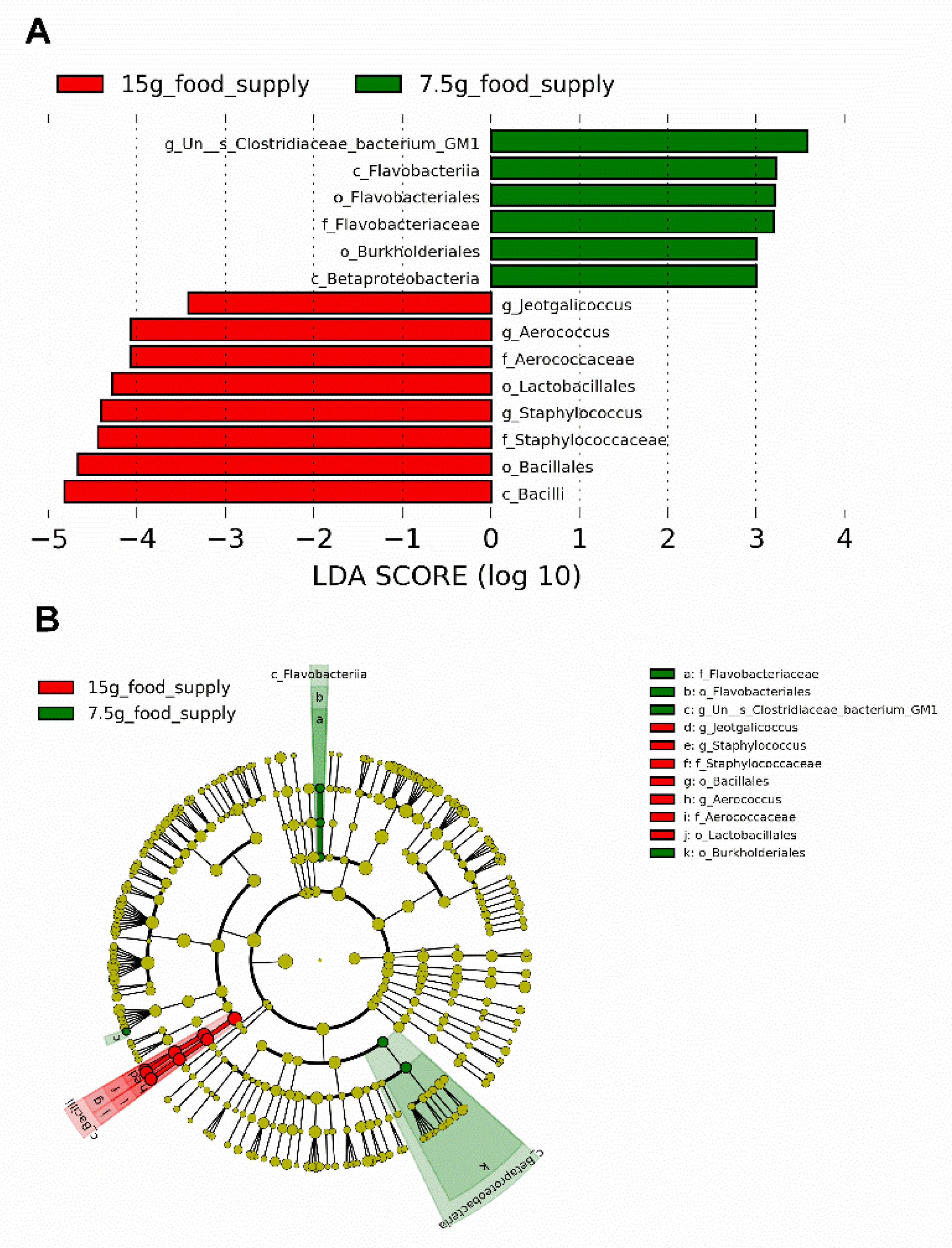
Figure 5.
Differences in taxa relative abundances of fecal microbiota at different taxonomic levels between the 15 g- and 7.5 g-food supply groups of Brandt’s voles. (A) class level; (B) order level; (C) family level; (D) genus level. Error bars indicate standard errors. Different letters connect bars with significant differences at P < 0.05 (n = 6).
Figure 5.
Differences in taxa relative abundances of fecal microbiota at different taxonomic levels between the 15 g- and 7.5 g-food supply groups of Brandt’s voles. (A) class level; (B) order level; (C) family level; (D) genus level. Error bars indicate standard errors. Different letters connect bars with significant differences at P < 0.05 (n = 6).
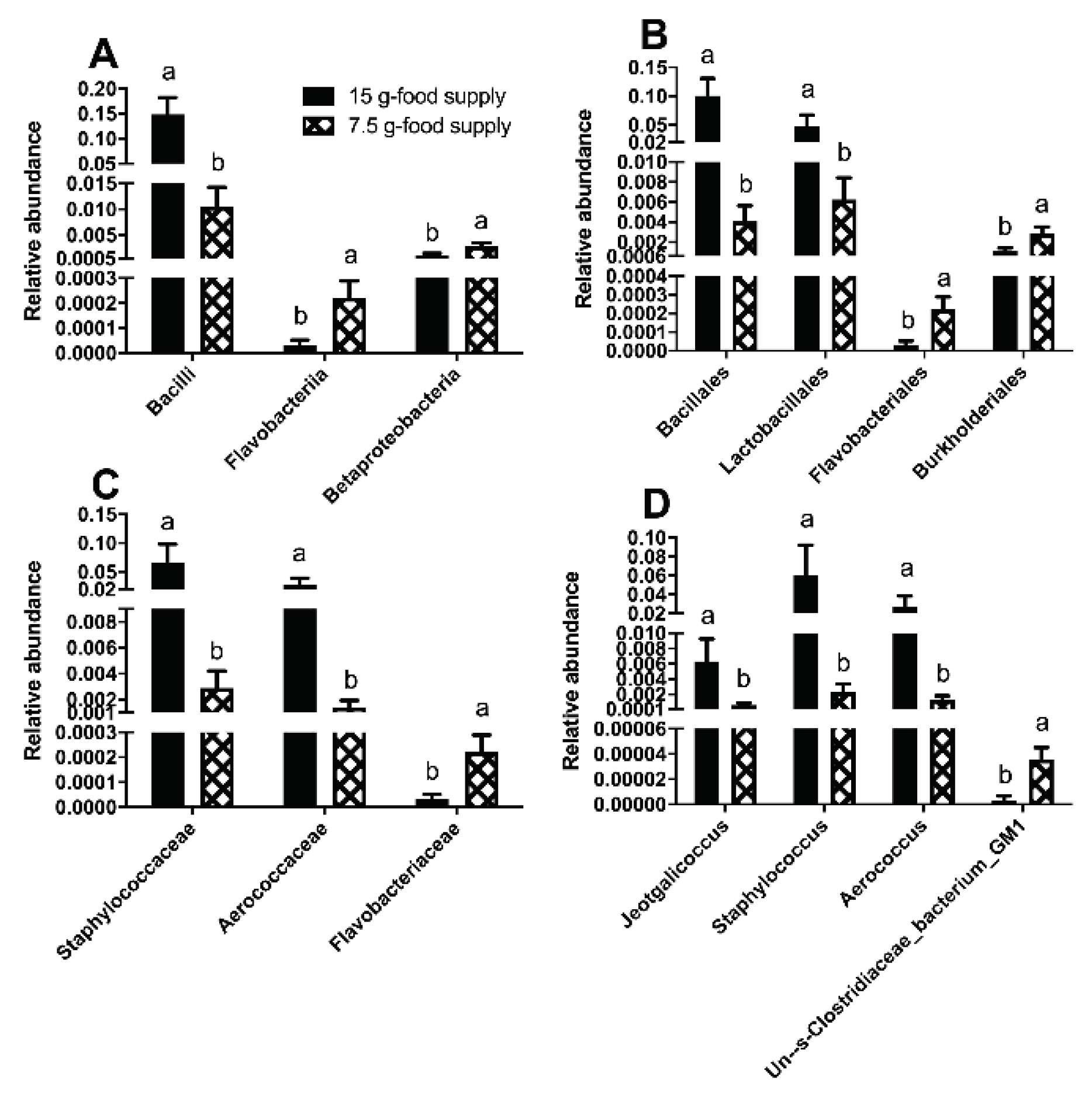
Figure 6.
Differences in the abundances of fecal microbial KEGG pathways between the 15 g- and 7.5 g-food supply groups of Brandt’s voles. Error bars indicate standard errors. Different letters connect bars with significant differences at P < 0.05 (n = 6).
Figure 6.
Differences in the abundances of fecal microbial KEGG pathways between the 15 g- and 7.5 g-food supply groups of Brandt’s voles. Error bars indicate standard errors. Different letters connect bars with significant differences at P < 0.05 (n = 6).
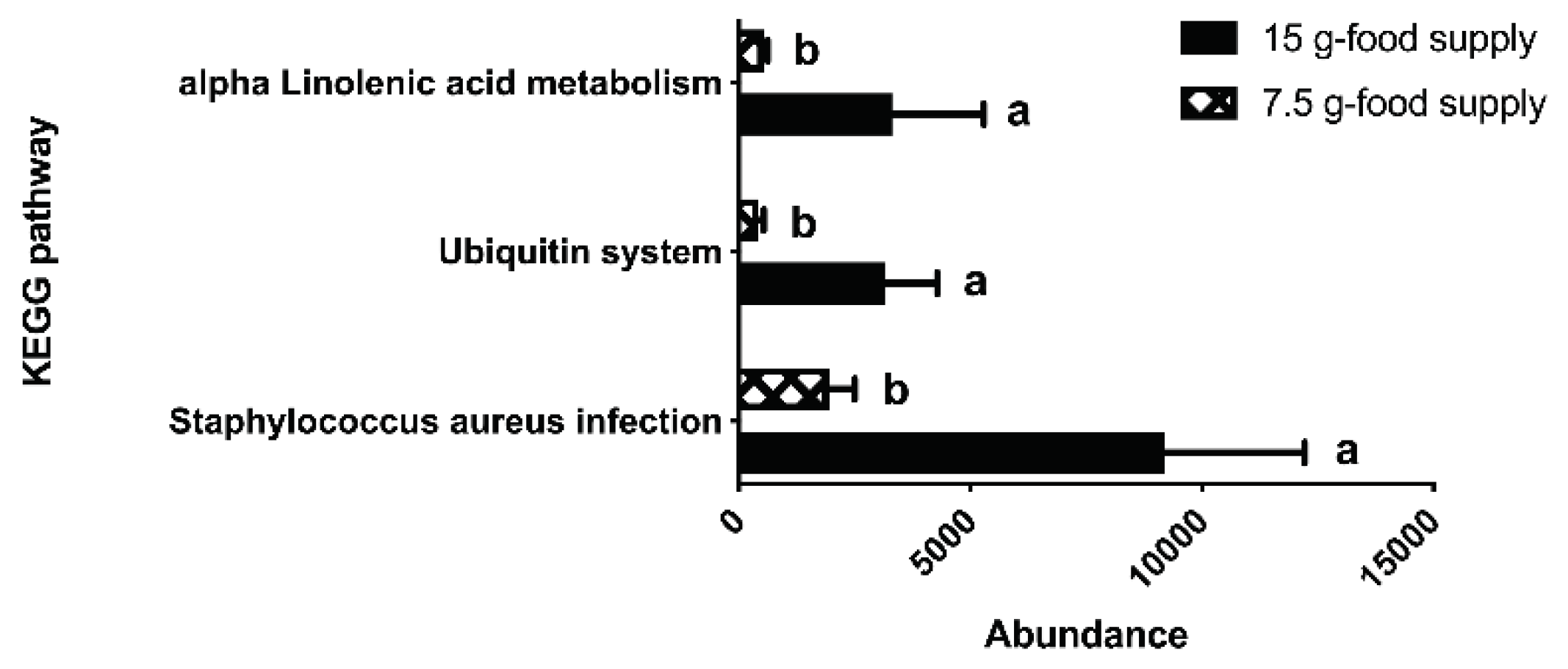
Figure 7.
Difference of fecal short-chain fatty acids (SCFAs: acetate, propionate, isobutyrate, butyrate, isovalerate, valerate, and caproate) content between the 15 g- and 7.5 g-food supply groups of Brandt’s voles. Error bars indicate standard errors. Different letters connect bars with significant differences at P < 0.05 (n = 6).
Figure 7.
Difference of fecal short-chain fatty acids (SCFAs: acetate, propionate, isobutyrate, butyrate, isovalerate, valerate, and caproate) content between the 15 g- and 7.5 g-food supply groups of Brandt’s voles. Error bars indicate standard errors. Different letters connect bars with significant differences at P < 0.05 (n = 6).
The body mass growth rate was positively correlated with relative food consumption, relative amount of ground food, and alpha-linolenic acid metabolism (P = 0.004, 0.022, and 0.037, respectively), and negatively correlated with the relative abundance of the Betaproteobacteria class and Burkholderiales order (P = 0.037 and 0.028, respectively; Figure 8). Relative food consumption was positively correlated with the relative amount of ground food, ground-to-consumed food ratio, acetate and caproate contents, relative abundances of the Bacilli class, Bacillales order, Staphylococcaceae family, Staphylococcus genus, and pathway enrichment of the ubiquitin system and alpha-linolenic acid metabolism (P = 0.008, 0.041, 0.015, 0.050, 0.038, 0.011, 0.038, 0.035, 0.011, and 0.004, respectively; Figure 8). Relative ground food was positively correlated with the ground-to-consumed food ratio (P < 0.001), acetate content, relative abundances of the Bacillales order, Aerococcaceae family, Aerococcus and Jeotgalicoccus genera, and pathway enrichment of alpha-linolenic acid metabolism (P = 0.011, 0.033, 0.039, 0.042, 0.049, and 0.034, respectively; Figure 8), but negatively correlated with the relative abundance of Un-s-Clostridiaceae bacterium GM1 (P = 0.038). The ratio of ground to consumed food was positively correlated with acetate content and the relative abundances of the Aerococcaceae family and Aerococcus genus (P = 0.026, 0.035, and 0.027, respectively), but negatively correlated with the Un-s-Clostridiaceae bacterium GM1 genus (P = 0.027) (Figure 8). The OBSP was positively correlated with the relative abundances of Betaproteobacteria and Burkholderiales (P = 0.035 and 0.048, respectively; Figure 8). Meanwhile, acetate content was positively correlated with the relative abundances of the Bacillales order, Aerococcaceae family, and Aerococcus genus (P = 0.021, 0.038, and 0.041, respectively), and pathway enrichment of alpha-linolenic acid metabolism (P = 0.024) (Figure 8). Moreover, the relative abundances of the Betaproteobacteria class and Burkholderiales order were negatively correlated with the valerate (P = 0.013 and 0.017, respectively) and caproate (P = 0.030 and 0.037, respectively) content (Figure 8), whereas the acetate (P = 0.002) and valerate (P = 0.025) content was positively correlated with the caproate content. Correlations among the abundances of microbiota biomarkers were either negative or positive (Figure 8). Pathway enrichment related to S. aureus infection, the ubiquitin system, and alpha-linolenic acid metabolism was also positively or negatively correlated with biomarkers of the microbiota (Figure 8).
4. Discussion
In this study, we investigated the effect of food supply changes on food grinding in male Brandt’s voles and its potential underlying mechanisms with respect to the gut microbiota. When the food supply for Brandt’s voles was restricted to 7.5 g, which was below the average daily food intake, the body mass growth rate, food consumption, amount of ground food, and the ground-to-consumed food ratio decreased. Some alpha diversity indices and the abundances of some fecal microbiota, along with the content of some SCFAs, varied; however, the fecal microbial structure did not change after reducing the food supply. Although body mass did not differ remarkably between the 15- and 7.5 g-supply groups when the food supply was reduced to 7.5 g, food consumption decreased, and the body mass growth rate was negative and significantly lower than that observed when the food supply was 15 g. A positive correlation was detected between the relative food consumption and body mass growth rate. This indicates that food restriction (7.5 g) can reduce food consumption and restrict vole growth. The ratio of ground to consumed food also decreased sharply, consistent with the higher amplitude of variation in relative ground food than in relative food consumption, indicating that the quantity of ground food decreased more than food consumption under a reduced food supply. Ort production in mice decreases to zero when they are offered 80% of their food intake [1]. Therefore, we concluded that the food-grinding behavior could be influenced by the quantity of food supplied. This suggests that when vegetation in grasslands is plentiful, food waste by wild Brandt’s voles is more extensive. Moreover, the inhibitive effect of food restrictions on food grinding could be explained by the hypothesis of an optimal food intake strategy [19] with food components having high energy per gram selected through grinding, to render the energy intake as large as possible [1]. When the food supply was reduced to less than the average food intake and the body mass growth rate was negative, food grinding would further reduce the available food to levels which could not meet the daily energy demand. Additionally, food grinding is an energy-cost behavior. Thus, no advantage would be gained regarding food selectivity or food grinding. Therefore, voles would reduce their food-grinding behavior and ingest as much food as possible to maximize energy intake and save energy. In our previous study, no significant correlation was detected between food consumption and relative ground food [9], whereas in this study, a positive correlation was observed. This inconsistency might be attributed to the negative effects of food restriction on food consumption and grinding. Relative ground food was positively correlated with the body mass growth rate, and the ground-to-consumed food ratio was nearly significantly and positively correlated with this parameter (P = 0.057), suggesting that food-grinding behavior might have other beneficial effects for voles, other than maximizing energy intake, which requires further research. The reason behind the body weight not being significantly different between the 15- and 7.5 g-food supply groups is that for the 15 g-food supply group, the body weights of voles increased from day 1 to day 14, whereas for the 7.5 g-food supply group, the body weights decreased from day 1 to day 14, and the food restriction experiment was conducted immediately following the 15 g-food supply experiment. Thus, the mean body weights were not obviously different between the 15- and 7.5 g-food supply groups. This also resulted in the lack of a pronounced correlation between the body weight and ground food or the ground-to-consumed food ratio.
In this study, the ACE index of the gut microbiota was high when the food supply was decreased, whereas the Shannon and Simpson indices did not change significantly, which is consistent with previously observed results for Chinese individuals after fasting [20]. Increases in the numbers of OTUs as well as the OBSP and ACE indices, suggested that food restriction enhances the number of species in the gut microbiota of Brandt’s voles. The increase in the number of gut bacterial species could be due to the enrichment of biomarkers for Betaproteobacteria and Burkholderiales caused by the positive correlations between OBSP and the relative abundances of these taxa. The acetate content was also lower for the 7.5 g-food supply group than for the 15 g-food supply group and was positively correlated with the ground food and ground-to-consumed food ratio, supporting our speculation that acetate could promote food-grinding behavior [9]. Therefore, the decreased acetate content might have restrained food-grinding behavior in this study. Assessing the ratio of ground to consumed food eliminated the influence of food intake on ground food. Given the strong negative or positive correlations between the ground-to-consumed food ratio and the Un--s-Clostridiaceae bacterium GM1 and Aerococcus genera between the 15- and 7.5 g-food supply groups, it is likely that these two genera could contribute to the regulation of food-grinding behavior, with the genus Un--s-Clostridiaceae bacterium GM1 inhibiting and the genus Aerococcus promoting it. The genera Aerococcus and Un--s-Clostridiaceae bacterium GM1 were the common biomarkers observed, based on two LEfSe analyses, that could differentiate the high- and low-level food-grinding groups in our previous study [9], as well as the 15- and 7.5 g-food supply groups in this study. This indicates a strong relationship between the Aerococcus or Un--s-Clostridiaceae bacterium GM1 genera and food-grinding behavior, which supports our previous speculation that gut microbiota participate in the regulation of food-grinding behavior [9]. The positive correlation between Aerococcus and the acetate content indicates that the effect of this genus on food-grinding behavior might occur through alterations to the acetate content. Therefore, we inferred that changes in the abundance of the gut microbiome components and their metabolite SCFAs due to food restriction would be helpful in inhibiting food grinding in Brandt’s voles.
An increase in S. aureus has been observed for obese individuals [21]. Further, reductions in the body weight, Staphylococcus abundance in the rectum, and Staphylococcaceae levels in the cecum have been observed for Kunming mice [22]. In this study, the abundance of the genus Staphylococcus, along with an increase in the body mass, was consistently higher for the 15 g-food supply group. Staphylococcaceae species are pathogens [23], and S. aureus infects a variety of tissues, organs, and systems in humans [24,25,26]. The pathway associated with S. aureus infection was more enriched for the 15 g-food supply group than for the 7.5 g-food supply group owing to the abundance of the genus Staphylococcus and the family Staphylococcaceae. Therefore, it is likely that food restriction can help reduce the abundance of pathogenic Staphylococcaceae species. Alpha-linolenic acid is an antibacterial substance that inhibits S. aureus [27,28]. Moreover, autophagy, mediated by ubiquitin receptors, is essential for zebrafish resistance to S. aureus [29]. Therefore, the enrichment of pathways related to the ubiquitin system and alpha-linolenic acid metabolism for the 15 g-food supply group could be associated with resistance to Staphylococcus bacteria. In Rex rabbits with an increased weight, the concomitant enrichment of pathways related to alpha-linolenic acid metabolism, and increased S. aureus colonization among the fecal microbiota was observed [30]. Further, strongly negative correlations between the abundances of taxa within Bacilli and Flavobacteriia indicate that these bacterial classes might restrain each other in Brandt’s voles.
In this study, differences in the amount of ground food, the ground-to-consumed food ratio, the abundances and diversity of fecal microbiota components, and the metabolite content between voles with different food supplies were observed. Based on these findings, we speculated on the effects of food restriction on food-grinding behavior, as well as the correlation between the gut microbiota, in addition to their metabolites, and food-grinding behavior. This study, along with our previous research [9], preliminarily clarifies the factors influencing food-grinding behavior and the mechanisms underlying this characteristic from the perspective of the gut microbiota. However, we could not distinguish whether the direct effect of food restriction, owing to the insufficient energy supply, or indirect effect of food restriction, via the gut microbiota, on food-grinding behavior was predominantly important. Food restriction in association with gut microbiota transplantation experiments should help to address this ambiguity. Additionally, the precise gut microbiota-associated mechanisms underlying food-grinding behavior and the functions of food-grinding behavior in voles require further research.
5. Conclusions
Our study demonstrated that food restriction could reduce the amount of ground food to a greater extent than its effect on food consumption, increase the numbers of certain gut microbiota species, and alter the abundance of the gut microbiome and its metabolites. The Aerococcus and Un--s-Clostridiaceae bacterium GM1 genera could be important for the regulation of food-grinding behavior, and acetate might be a key metabolite that mediates this behavior. Thus, variations in the abundance of the gut microbiome components and their metabolite SCFAs, induced by food restriction, could contribute to the inhibition of food grinding in Brandt’s voles. This study suggests that reducing the food supply could effectively inhibit food grinding during laboratory rodent feeding, further supporting the hypothesis that food grinding could be motivated by an optimal food-intake strategy and partially verifying the hypothesis that the gut microbiota might be related to food-grinding behavior. These results could potentially provide guidance for reducing food waste during laboratory rodent maintenance and promote deeper research on the gut microbial mechanism of food-grinding behavior and the potential role of food-grinding behavior.
Author Contributions
Conceptualization, X.D.; Methodology, X.D.; Software, X.D.; Validation, S.M.Y.; Formal Analysis, X.D.; Investigation, Y.X.H., Q.Y.S., H.T. and L.Z.C.; Resources, S.M.Y. and W.H.W.; Data Curation, Y.X.H. and X.D.; Writing – Original Draft Preparation, X.D.; Writing – Review & Editing, X.X.; Visualization, X.D. and F.P.Y.; Supervision, F.P.Y., S.M.Y. and W.H.W.; Project Administration, S.M.Y.; Funding Acquisition, X.D., S.M.Y. and W.H.W.
Data availability statement
The sequence data are available at the NIH Sequence Read Archive with Bioproject ID PRJNA868862.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (grant number: 31971418, 31770422, 32370438) and the Priority Academic Program Development of Jiangsu Higher Education Institutions. We would like to thank Editage (www.editage.cn) for English language editing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cameron, K.M.; Speakman, J.R. The extent and function of 'food grinding' in the laboratory mouse (Mus musculus). Lab. Anim. 2010, 44, 298-304. [CrossRef]
- Starr, M.E.; Saito, H. Age-related increase in food spilling by laboratory mice may lead to significant overestimation of actual food consumption: implications for studies on dietary restriction, metabolism, and dose calculations. J. Gerontol. A Biol. Sci. Med. Sci. 2012, 67, 1043-1048. [CrossRef]
- Tertil, R. Impact of the Common Vole, Microtus arvalis (Pallas) on Winter Wheat and Alfalfa Crops. EPPO Bull. 1977, 7, 317-339. [CrossRef]
- Koteja, P.; Carter, P.A.; Swallow, J.G.; Garland, T., Jr. Food wasting by house mice: variation among individuals, families, and genetic lines. Physiol. Behav. 2003, 80, 375-383. [CrossRef]
- Peterson, J.; Wunder, B.A. Food Sorting by Collared Lemmings (Dicrostonyx groenlandicus) and Prairie Voles (Microtus ochrogaster): A Cautionary Note for Digestibility Studies. Comp. Biochem. Physiol. A: Physiol. 1997, 116, 119-124. [CrossRef]
- Ford, D.J. Influence of diet pellet hardness and particle size on food utilization by mice, rats and hamsters. Lab. Anim. 1977, 11, 241-246. [CrossRef]
- Martins, P.J.F.; Nobrega, J.N.; Tufik, S.; D'Almeida, V. Sleep deprivation-induced gnawing—relationship to changes in feeding behavior in rats. Physiol. behav. 2008, 93, 229-234. [CrossRef]
- Esumi, L.A.; Palma, B.D.; Gomes, A.C.; Tufik, S.; Hipólide, D.C. Inflammatory markers are associated with inhibitory avoidance memory deficit induced by sleep deprivation in rats. Behav. Brain Res. 2011, 221, 7-12. [CrossRef]
- Shen, Q.; Shi, J.; Gu, K.; Wei, W.; Yang, S.; Dai, X. Relationship between food grinding and gut microbiota in Brandt’s voles. Can. J. Zool. 2023, 00, 1-12, . [CrossRef]
- Xie, X.M.; Sun, R.Y.; Fang, J.M. The mating system and reproduction of Brandt’s voles (Microtus brandtii). Acta Zool. Sin. 1994, 40, 262-265.
- Shi, D.Z.; Hai, S.Z.; Lu, D.; Liu, X.F. The structure and order in colony of Brandt’s vole. Acta Theriol. Sin. 1999, 19, 48-55.
- Wan, X.R.; Wang, M.J.; Wang, G.H.; Liu, W.; Zhong, W.Q. The reproductive parameters in the marked populations of Brandt’s vole. Acta Theriol. Sin. 2002, 22, 116-122.
- Li, G.; Yin, B.; Li, J.; Wang, J.; Wei, W.; Bolnick, D.I.; Wan, X.; Zhu, B.; Zhang, Z. Host-microbiota interaction helps to explain the bottom-up effects of climate change on a small rodent species. ISME J. 2020, 14, 1795-1808. [CrossRef]
- Kelley, T.L. The selection of upper and lower groups for the validation of test items. J. Educ. Psychol. 1939, 30, 17-24.
- Dai, X.; Chen, L.; Liu, M.; Liu, Y.; Jiang, S.; Xu, T.; Wang, A.; Yang, S.; Wei, W. Effect of 6-Methoxybenzoxazolinone on the Cecal Microbiota of Adult Male Brandt's Vole. Front. Microbiol. 2022, 13, 847073. [CrossRef]
- Edgar, R.C. UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996-998. [CrossRef]
- Campos, P.M.; Darwish, N.; Shao, J.; Proszkowiec-Weglarz, M. Research Note: Choice of microbiota database affects data analysis and interpretation in chicken cecal microbiota. Poult. Sci. 2022, 101, 101971. [CrossRef]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814-821. [CrossRef]
- Pritchett-Corning, K.R.; Keefe, R.; Garner, J.P.; Gaskill, B.N. Can seeds help mice with the daily grind? Lab. Anim. 2013, 47, 312-315. [CrossRef]
- Ali, I.; Liu, K.; Long, D.; Faisal, S.; Hilal, M.G.; Ali, I.; Huang, X.; Long, R. Ramadan Fasting Leads to Shifts in Human Gut Microbiota Structured by Dietary Composition. Front. Microbiol. 2021, 12, 642999. [CrossRef]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes 2018, 9, 308-325. [CrossRef]
- Cheng, S.; Mao, H.; Ruan, Y.; Wu, C.; Xu, Z.; Hu, G.; Guo, X.; Zhang, C.; Cao, H.; Liu, P. Copper Changes Intestinal Microbiota of the Cecum and Rectum in Female Mice by 16S rRNA Gene Sequencing. Biol. Trace Elem. Res. 2020, 193, 445-455. [CrossRef]
- Ruzauskas, M.; Armalyte, J.; Lastauskiene, E.; Siugzdiniene, R.; Klimiene, I.; Mockeliunas, R.; Bartkiene, E. Microbial and Antimicrobial Resistance Profiles of Microbiota in Common Carps (Cyprinus carpio) from Aquacultured and Wild Fish Populations. Animals (Basel) 2021, 11, 929. [CrossRef]
- Narita, K.; Asano, K.; Nakane, A. IL-17A plays an important role in protection induced by vaccination with fibronectin-binding domain of fibronectin-binding protein A against Staphylococcus aureus infection. Med. Microbiol. Immunol. 2017, 206, 225-234. [CrossRef]
- Stokes, W.; Parkins, M.D.; Parfitt, E.C.T.; Ruiz, J.C.; Mugford, G.; Gregson, D.B. Incidence and Outcomes of Staphylococcus aureus Bacteriuria: A Population-based Study. Clin. Infect. Dis. 2019, 69, 963-969. [CrossRef]
- Sethi, S.; Chakraborty, T. Role of TLR- / NLR-signaling and the associated cytokines involved in recruitment of neutrophils in murine models of Staphylococcus aureus infection. Virulence 2011, 2, 316-328. [CrossRef]
- A Petropoulos, S.; Fernandes, A.; Dias, M.I.; Pereira, C.; Calhelha, R.C.; Chrysargyris, A.; Tzortzakis, N.; Ivanov, M.; D Sokovic, M.; Barros, L.; et al. Chemical Composition and Plant Growth of Centaurea raphanina subsp. mixta Plants Cultivated under Saline Conditions. Molecules 2020, 25, 2204. [CrossRef]
- Sado-Kamdem, S.L.; Vannini, L.; Guerzoni, M.E. Effect of alpha-linolenic, capric and lauric acid on the fatty acid biosynthesis in Staphylococcus aureus. Int. J. Food Microbiol. 2009, 129, 288-294. [CrossRef]
- Munoz-Sanchez, S.; van der Vaart, M.; Meijer, A.H. Autophagy and Lc3-Associated Phagocytosis in Zebrafish Models of Bacterial Infections. Cells 2020, 9, 2372. [CrossRef]
- Zeng, B.; Han, S.; Wang, P.; Wen, B.; Jian, W.; Guo, W.; Yu, Z.; Du, D.; Fu, X.; Kong, F.; et al. The bacterial communities associated with fecal types and body weight of rex rabbits. Sci. Rep. 2015, 5, 9342. [CrossRef]
Figure 8.
Spearman’s rank correlations among parameters in the 15 g- and 7.5 g-food supply groups of Brandt’s voles. Note: *, **, and *** mean false discovery rate-adjusted P-value was < 0.05, 0.01, and 0.001, respectively (n = 6).
Figure 8.
Spearman’s rank correlations among parameters in the 15 g- and 7.5 g-food supply groups of Brandt’s voles. Note: *, **, and *** mean false discovery rate-adjusted P-value was < 0.05, 0.01, and 0.001, respectively (n = 6).
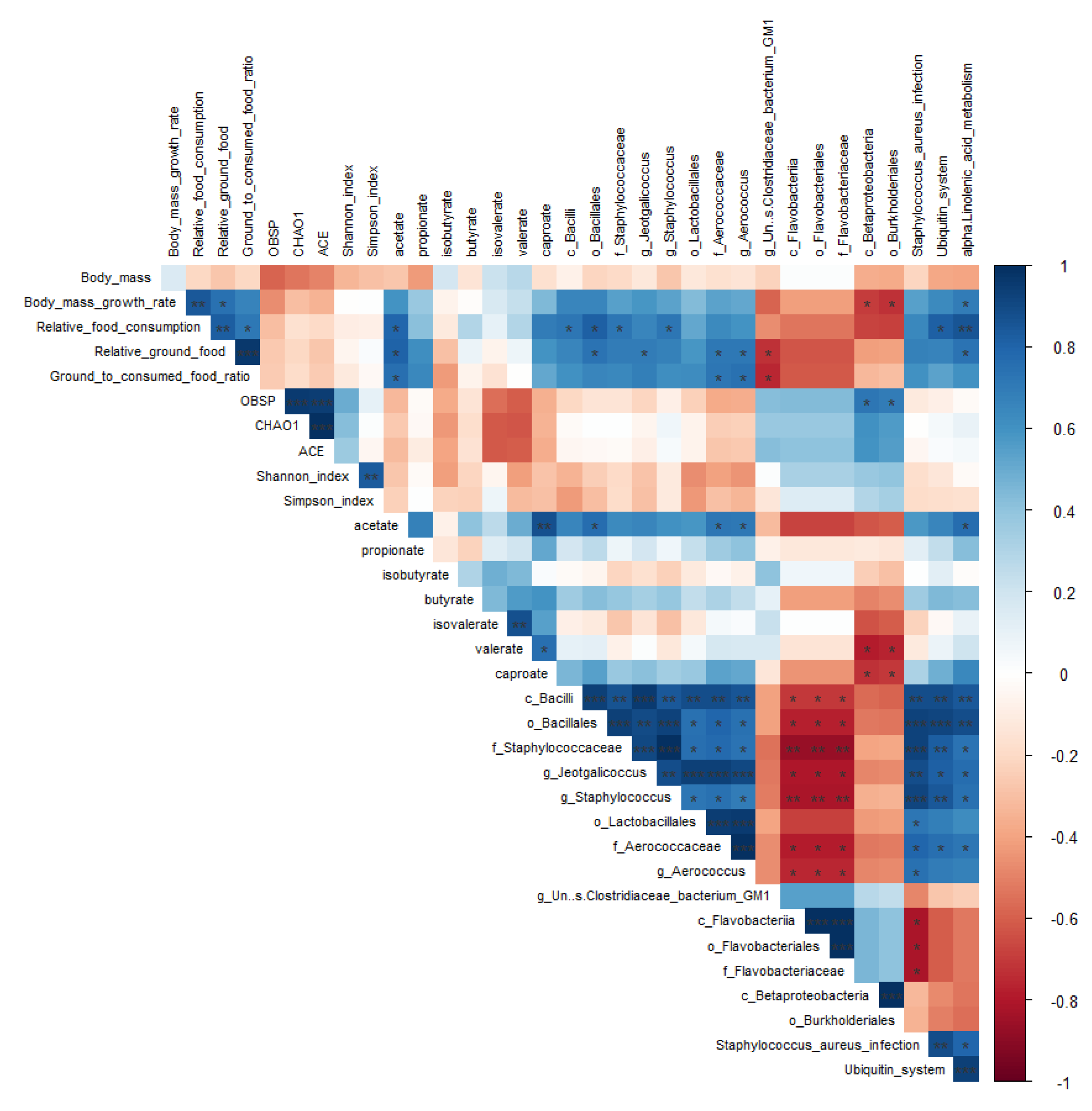

Table 1.
Differences of total tags, taxon tags, operational taxonomic units (OTUs), and Goods coverage in 16S rRNA libraries of fecal microbiota in the 15 g- and 7.5 g-food supply groups of Brandt’s voles (mean ±SE) (n = 6).
Table 1.
Differences of total tags, taxon tags, operational taxonomic units (OTUs), and Goods coverage in 16S rRNA libraries of fecal microbiota in the 15 g- and 7.5 g-food supply groups of Brandt’s voles (mean ±SE) (n = 6).
| Groups | Total tags | Taxon tags | OTU numbers | Goods coverage |
|---|---|---|---|---|
| 15 g-food supply | 63,501±1,070 | 57,234±1,017 | 877±47b | 99.77±0.02 |
| 7.5 g-food supply | 66,282±1567 | 57,704±2,233 | 981±22a | 99.75±0.02 |
Note: different letters mean significant differences at P < 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



