
Submitted:
12 September 2023
Posted:
15 September 2023
You are already at the latest version
Abstract
In recent years, the interaction between adiponectin (APN), a versatile fat-associated adipokines, and eye disorders has become a captivating focus of interest. Unveiling the intricate relationship between APN and its receptors (AdipoRs) with aging eye disorders has emerged as a fascinating frontier in medical research. This review article delves into this captivating connection, illuminating the hidden influence of adipocytokine, APN on retinal health. This comprehensive review critically examines the latest findings and breakthroughs that underscore the pivotal roles of APN/AdipoRs signaling in maintaining ocular homeostasis and protecting against eye ailments. Here, we meticulously explore the intriguing mechanisms by which APN protein influences retinal function, lens clarity, and overall visual acuity. Drawing from an extensive array of cutting-edge studies, the article highlights APN's multifaceted functions, ranging from anti-inflammatory properties and oxidative stress reduction to angiogenic regulation within retinal and macula tissues. The involvement of APN/AdipoRs in mediating these effects opens up novel avenues for potential therapeutic interventions targeting prevalent aging eye conditions. Moreover, this review unravels the interplay between APN signaling pathways and age-related macular degeneration (AMD). The single-cell RNA seq results validate the expression of both the receptor isoform (AdipoR1/R2) in retinal cells. The transcriptomic analysis showed lower expression of AdipoR1/2 in dry AMD pathogenesis compared to healthy subjects. The inhibitory peptide (APN1) demonstrated over 75% suppression of CNV, whereas the control peptide did not exert any inhibitory effect on CNV. The elucidation of these relationships fosters a deeper under-standing of adipose tissue's profound influence on ocular health, presenting new prospects for personalized treatments and preventative measures. In conclusion, "Obesity Hormone's Role in Aging Eyes: Linking Adiponectin/AdipoRs to AMD" provides a captivating journey into the enthralling world of APN intertwining the realms of adipose biology and aged ophthalmology.
Keywords:
Obesity
; Adiponectin (APN)
; Adiponectin receptors (AdipoRs)
; Age-Related Macular Degeneration (AMD)
; Dry AMD
; Wet AMD
; Choroidal neovascularization (CNV)
; Angiogenesis
; Inflammation
1. Introduction
Adiponectin (APN), a hormone linked to obesity regulation weighing around 30 kDa, is produced by the ADIPOQ gene and primarily originates from white adipose tissue (WAT). APN play a crucial role in various essential bodily functions, such as managing glucose and fatty acid metabolism [1,2]. It also contributes to maintaining glucose and lipid balance [3], overall energy regulation [4], immune responses [5], and the effects of aging and metabolism [6,7,8]. Notably, APN extends its safeguarding influence on ocular tissue [8,9]. The retina, a metabolically active component often outpacing even the brain's metabolic rate, triggers blood vessel growth and regression due to its high-energy needs. APN has also demonstrated protective effects against multiple retinal disorders, including diabetic retinopathy (DR) [9], choroidal neovascularization (CNV) arising from age-related macular degeneration (AMD) [10], and other retinal conditions [11].
Neovascular AMD is a complex retinal condition where an individual's genetic disposition is influenced by the effects of age and environmental stressors. These factors cascade a series of signaling pathways that involve inflammation, oxidation, and/or angiogenesis within the retinal pigment epithelial (RPE) cells and choroidal endothelial cells (CECs). Ultimately, this process results in vision loss due to the advancement of CNV. Navigating the world of AMD, we encounter its dual personas: the wet and dry types. Unveiling its dramatic impact, the wet form takes the spotlight, emerging as a chief instigator of irreversible blindness and the complete eclipse of central vision among the elderly. The hallmark of this drama is CNV. In contrast, the dry type, though not synonymous with total blindness, casts a shadow over central vision, posing challenges in reading, driving, and perceiving the world around. As the curtain rises on advanced stages, dry AMD can take a perilous turn, progressing into geographic atrophy (GA) or even evolving into its wet counterpart, both orchestrating a symphony of severe vision loss.
Initiating a cascade of events, retinal hypoxia triggers an upsurge in metabolic demands, setting off signaling pathways that endeavor to tap into new vascular resources, ultimately culminating in the eye's neovascularization [10]. Literature hints at a shift in the balance of two predominant circulating adipokines, APN, and leptin, pivotal players in metabolic modulation across diverse tissues. This dynamic duo might play a role in driving the progression of neovascular eye conditions [9]. Further insights point to the heightened release of leptin, a hormone originating from adipocytes, as a harbinger of disrupted energy equilibrium, increased oxidative stress on vascular endothelial cells (ECs), and consequent dysfunction of these cells, ultimately contributing to retinopathy [12]. In parallel, another metabolic influencer, primarily sourced from adipocytes, called 'APN,' joins the orchestra of metabolic irregularities in the retina. The levels of circulating APN are intricately tied to DR [13,14], the development and advancement of premature retinopathy [15], and age-related macular degeneration [16]. This correlation is underscored by research, including studies involving laser-induced choroidal neovascularization [17,18] and a rodent model of oxygen-induced proliferative retinopathy, where higher circulating APN levels correlate with suppression of pathological vascular proliferation [19]. Embarking on a journey through ongoing investigations and pertinent studies, we delve into this burgeoning realm of APN/AdipoRs, dissecting their roles in the intricate landscape of retinal neovascular disorders.
2. Unveiling Adiponectin/AdipoRs Physiological Roles
2.1. Unlocking the Mysteries of APN/AdipoRs: A Journey Through Discovery, Structure, and Circulation Forms
During the mid-1990s, APN's discovery unfolded as a harmonized effort across four research laboratories. This revelation led to its christening with various appellations, encompassing the 30 kilo Dalton (KDa) adipocyte complement-related protein (Acrp30), ADIPOQ, Adipose Most Abundant Gene Transcript 1 (apM1), and the enigmatic 28 kDa Gelatin Binding Protein (GBP28) [20,21,22,23]. In contrast, the identification of APN receptors, denoted as AdipoRs, underwent a protracted gestation period, finally coming to fruition in 2003 through pioneering work by Yamauchi et al. This milestone was achieved by selectively extracting two closely related membrane-bound receptors, AdipoR1 and AdipoR2, from human skeletal muscle using expression cloning vectors [25]. Within the realm of scholarly contributions, it is imperative that extensive datasets, housed within publicly accessible repositories, adhere to transparent principles by disclosing their residence alongside the vital accession numbers. In circumstances where these codes are pending at submission, acknowledgment of their impending availability is permissible, contingent upon their full disclosure prior to the commencement of the publication process.
The APN gene finds its residence on chromosome 3q27, stretching across a span of 16 kilo base pairs (kbp). Encoded within the human APN gene is a 244 amino acid (aa) secretory peptide (with a 247 aa counterpart for the mouse ortholog), clocking in at approximately 30 KDa. The foundational architecture of APN is constructed with a Carboxy (C)-terminal globular domain paired with an Amino (N)-terminal collagen-like domain [26]. Both human and mouse AdipoR1 comprise 375 aa in their protein structure, with a forecasted molecular mass of 42.4 kDa. Meanwhile, the human AdipoR2 protein consists of 299 aa (and mouse AdipoR2, 311 aa), bearing a predicted molecular mass of 35.4 kDa. Remarkably, AdipoR1 and AdipoR2, despite originating from separate genetic sources, exhibit a sequence identity of 67%. These receptor isoforms adopt the form of integral membrane proteins, where the N-terminus faces internally, and the C-terminus is positioned externally—a distinctive arrangement that diverges from the topology and role of other recognized G protein–coupled receptors (GPCRs) [27]. The research underscores the variability in the binding affinity of these receptors towards different forms of APN. Notably, AdipoR1 displays a heightened affinity for globular APNs, whereas AdipoR2 exhibits an intermediate affinity for both globular and full-length APNs. In a pattern of ubiquity, APN and AdipoRs manifest their presence across diverse tissues [28]. In addition to the dynamic duo of AdipoR1 and AdipoR2, APN extends its influence through interaction with the receptor T-cadherin, although, at present, its role seems less pivotal in comparison to AdipoRs [29].
The obesity-related peptide APN assumes a complex structural configuration and circulates within the bloodstream in three distinct molecular forms: a low molecular weight (LMW) trimer, a middle molecular weight (MMW) hexamer, and a high molecular weight (HMW) oligomer. These diverse APN variants exhibit varying levels of biological activity, with HMW APN being identified as the biologically active iteration of this hormone [30]. Interestingly, in specific scenarios, the HMW form has demonstrated superior insulin-sensitizing properties when compared to other LMW forms. In addition to its intricate structural diversity, APN undergoes glycosylation, a crucial post-translational modification necessary for maintaining its functionality. Its concentration in circulation typically falls within the range of 3 µg/ml to 30 µg/ml in both humans and rodents, making it one of the most abundant adipokines present in the plasma [31,32].
2.2. Tissue Distribution, Mechanism, Physiological and Pathophysiological Relevance of APN/AdipoRs Pathway
Primarily originating from adipocytes, APN is expressed in various locations and imparts favorable influences on several metabolically active organs and cell types [33,34]. These include liver parenchymal cells, such as hepatocytes [35], skeletal muscle and myocytes [36], the brain [37], blood vessels [38], reproductive organs in both males and females [6,39], as well as ocular tissues [9,40]. AdipoR1 and AdipoR2 exhibit broad and abundant expression, not limited to skeletal muscle and liver tissues but also extending to macrophages [41], the hypothalamus [42], white adipose tissue (WAT) [43], reproductive tissues [44,45], and the retina [46]. In both in vitro and in vivo studies, AdipoRs have emerged as pivotal mediators of APN signaling [8]. APN engages with its two renowned, distinct cell-surface receptor variants, namely AdipoR1 and AdipoR2 [24]. Furthermore, AdipoR1 plays a more prominent role in initiating the AMP-activated protein kinase (AMPK) pathway, leading to the inhibition of hepatic glucose production and an increase in fatty acid oxidation. On the other hand, AdipoR2 is primarily associated with activating the peroxisome proliferator-activated receptor alpha (PPARα) nuclear receptor pathways, which in turn promote fatty acid oxidation and mitigate tissue inflammation and oxidative stress [47]. In metabolic organs targeted by insulin, such as the liver and skeletal muscle, the expression of AdipoRs significantly increases during fasting conditions compared to refed conditions in rodent models. Additionally, in vitro studies have revealed that insulin reduces the expression of AdipoRs through the phosphoinositide 3-kinase/FoxO1-dependent pathway [48].
The levels of APN in circulation and the presence of AdipoR1/R2 expression in metabolically active organs remain lower in obese and diabetic individuals as compared to healthy and lean individuals [49,50,51]. Previous study has revealed sexual dimorphism in adiponectin (APN) synthesis, demonstrating that males exhibit lower APN levels than their female counterparts. Furthermore, there is a significant reduction in APN concentrations among obese patients with Type 2 diabetes (T2D) and infertility [52]. In most cases, insulin acts as a stimulating factor, while tumor necrosis factor-alpha (TNF-α) serves as an inhibitor of APN signaling and secretion [53]. Furthermore, APN is recognized for its specific actions in regulating metabolism and developing insulin sensitivity. Additionally, it exerts a controlled influence on inflammatory responses by mitigating the production and functional activity of tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6) within macrophages through the inhibition of NF-κB activation, as elaborated earlier [54].
APN also plays a role in managing multiple physiological processes, including glucose utilization, lipid biosynthesis, energy homeostasis, and inflammatory, and retinal function [15,55]. Lack of APN secretion and expression globally or locally leads to insulin resistance, glucose intolerance, and hyperlipidemia in rodents [56,57]. APN along with other adipocytokine plays a primary pathophysiological function in the interaction between metabolism and reproduction and may be associated with the detrimental effect of aging on male reproductive actions [7,39]. The use of APN supplementation could potentially serve as a crucial therapeutic approach for addressing reproductive disorders associated with obesity, such as male and female infertility [6,58].
In vitro knockdown experiments involving the ADIPOQ gene in skeletal muscles demonstrate the pivotal function of AdipoR1 in orchestrating various processes, including but not limited to β-oxidation, PPAR activation, glucose uptake, and AMPK activation [59]. AdipoR1 plays a role in elevating the phosphorylation of AMPK protein within the liver, consequently affecting the gluconeogenesis process. Concurrently, AdipoR2 is accountable for the removal of reactive oxygen species (ROS) and the initiation of nuclear receptor PPAR activation, along with the downstream modulation of target genes associated with β-oxidation. The emergence of Nonalcoholic Fatty Liver Disease (NAFLD) and steatohepatitis (NASH) accompanied by fibrosis and inflammation in obese rats fed a high-fat/high-cholesterol diet highlights the involvement of AdipoRs in regulating hepatic fatty acid metabolism. The NASH condition led to a remarkable reduction in the expression of both AdipoR1/R2 isoforms. This decrease was linked to lower levels of PPARα and AMPKα 1/2. Furthermore, the specific tissues played a crucial role in determining these effects. AdipoR1 in the liver played a role in activating AMPK, while AdipoR2 was actively engaged in activating PPARα, resulting in heightened insulin sensitivity [61]. Due to its ability to establish communication with other organs and lipid retention capacity, APN play an indispensable role in the maintenance of lipid and glucose homeostasis. In one study, overexpression of adiponectin/AdipoRs showed numerous beneficial effects such as reduction of visceral fat and amelioration of inflammatory responses and hepatic fibrosis [62].
3. Unlocking the Potential of APN/AdipoRs as Metabolic Regulators in Retinal Diseases
Recent studies have investigated the presence of APN in ocular tissues [46,63], with a particular focus on its role in conditions such as DR, retinopathy of prematurity, the preservation of photoreceptor integrity, retinitis pigmentosa, hypoxia-induced retinal neovascularization, and AMD within the context of ocular pathophysiology. As previously discussed, it's important to note that the retina is among the most metabolically demanding tissues in the human body, and photoreceptors, in particular, house a greater number of mitochondria compared to any other cell type [64]. The retina is supplied with essential nutrients and oxygen through its vascular network. Premature loss of these blood vessels can induce hypoxia and insufficiency of energy substrates, both of which are recognized as pivotal factors in instigating angiogenesis within retinal tissue. Hypoxia leads to a reduction in the activity of prolyl hydroxylase, an enzyme well known for its capacity to rapidly degrade the hypoxia-inducible factor (HIF)-1 protein under normal oxygen conditions. Elevated levels of HIF-1 protein, in turn, initiate the expression of angiogenic factors, most notably vascular endothelial growth factor A (VEGFA). It is worth noting that specific metabolic pathways, independent of HIF-1, can also influence the regulation of VEGFA expression [65]. VEGFA plays a pivotal role in promoting the proliferation of blood vessels, a crucial response aimed at restoring oxygen and energy substrate supply to the retina. However, these newly formed blood vessels often exhibit structural abnormalities that may potentially damage the delicate retinal tissue [66], and in severe cases, this can progress to blindness.
Neurodegenerative eye diseases often manifest with symptoms such as hazy, blurred, or distorted vision. The activation of the APN/AdipoRs signaling pathway has demonstrated significant neuroprotective potential, offering promise for ameliorating these conditions and enhancing visual function. Notably, both APN and AdipoRs are found within various retinal cells. When APN binds to AdipoRs and subsequently triggers downstream molecular pathways, it exerts its therapeutic effects, with detectable expression in the retina. While adipose tissues predominantly secrete APN [43], it's worth mentioning that the retina [46] and brain [37] can also locally produce this protein. Furthermore, APN readily traverse the bloodstream and efficiently cross the blood-brain barrier. Numerous pathophysiological conditions, such as elevated glucose levels, dyslipidemia, and disruptions in mitochondrial function, have the potential to disrupt retinal functions and contribute to retinal vascular disorders [67]. A pivotal driver of glycolysis, specifically the isoform 3 of 6-Phosphofructo-2-kinase/fructose-2,6-bisphosphatase, plays a vital role in the regulation of angiogenesis [68]. Perturbations in the activity of glucose metabolic enzymes within the polyol pathway can also assume a protective role for the retina, safeguarding it against retinal dysfunction and abnormal blood vessel growth [69,70]. Furthermore, an alternative approach to curbing vessel sprouting involves inhibiting the rate-limiting enzyme in the fatty acid oxidation process, namely, carnitine palmitoyl transferase 1 [71].
Several studies have explored the protective function of APN/AdipoRs pathways, and one noteworthy investigation was conducted among individuals diagnosed with T2D in Japan. Utilizing laser Doppler velocimetry, the study observed that in males, there was a positive correlation between blood APN levels and retinal blood vessel diameter, as well as retinal blood velocity and flow. However, such a correlation was not observed in females. Achieving a better balance in one's lifestyle or utilizing medications that lead to an elevation in plasma APN levels may unveil a promising avenue for the development of innovative therapeutic strategies in diabetes treatment [72]. It's worth noting that high glucose levels have been identified as the primary predisposing factor for angiogenesis in DR. Findings from a previous study have elucidated the role of APN in the dysregulated autophagy process and retinal angiogenesis. Additionally, APN has exhibited a protective effect against high glucose-induced damage to RF/6A cells. Furthermore, it has been shown to mitigate high glucose-induced angiogenesis in RF/6A cells by inhibiting the autophagy pathway [73].
4. Current Understanding of the Pathophysiological Role of APN/AdipoRs in Neovascular AMD
Neovascular AMD is the primary factor in legal blindness in the elderly. This progressive eye disease affects the macula (also called macula lutea), a yellow-pigmented area of the retina, containing color-sensitive rods which is vital for sharp, central vision. Evidence from the literature suggests a crucial role of APN in ameliorating neovascularization in AMD [8,17]. Mice with laser-induced neovascularization in the choroid serve as a model to simulate various inflammatory responses associated with AMD. The growing body of research indicates that Adiponectin (APN) may emerge as a novel and highly promising therapeutic target for addressing angiogenesis associated with Diabetic Retinopathy (DR). More specifically, it has the potential to significantly reduce the development of new blood vessels in the retina, a process known as retinal neovascularization, in primary human cell cultures of retinal microvascular endothelial cells (ECs), umbilical vein macrovascular ECs, and choroidal ECs. This beneficial effect is closely linked to its ability to interfere with the function of the VEGF, a key driver of angiogenesis in DR.
Importantly, the most closely related counterpart of APN, a cytokine called C1q/TNF-related protein-9, has shown the capability to protect the integrity of the blood-retinal barrier (BRB). Preserving the BRB serves to reduce the inflammatory response observed in diabetic db/db mice affected by DR. In addition to its potential applications in treating DR, therapies centered around APN/AdipoRs may hold promise in addressing the vision-threatening consequences of AMD. AMD encompasses two prevalent phenotypes: Dry AMD and Wet AMD. This condition is characterized by the accumulation of drusen, which are composed of a mixture of proteins, fats, minerals, and other debris, forming spherical structures bound tightly to proteins. Over time, drusen inflict damage on the retina, leading to irreversible changes in retinal cells. Dry AMD can progress to Wet AMD, characterized by the development of new blood vessels originating from the choroid. These vessels invade the Retinal Pigment Epithelium (RPE) and subretinal space, ultimately culminating in central vision loss. Mallordo and colleagues have postulated the distinct role of APN in ocular diseases, highlighting its inhibitory effects on the proliferation and migration of RPE cells [76]. Furthermore, Osada and his team demonstrated the consequences of AdipoR1 deletion on abnormal lipid metabolism within the retina, as well as retinal neurodegeneration, using AdipoR1 knockout mice. Their research, utilizing in situ hybridization, revealed robust AdipoR1 mRNA expression in the photoreceptor inner segment (PIS) and faint staining in the inner retinal layers in 4-week-old control mouse retinas. The expression of AdipoR1 in the retina appears to play a crucial role in inducing the elongase of very long-chain fatty acids (ELOVL2), a potentially essential step in providing an adequate supply of docosahexaenoic acid (DHA) necessary for the proper functioning and survival of photoreceptor cells [77].
Potentially, the expression of AdipoR1/R2 in single-cell RNA-seq datasets related to AMD can help researchers better understand the possible mechanism of APN/AdipoRs signaling in the retinal disease, including its impact on inflammation, angiogenesis, and specific retinal cell populations. This knowledge may ultimately contribute to the development of more effective treatments for AMD. To date, none of the literature on single-cell RNAseq presented the expression of APN/AdipoRs in the various retinal cell populations, which emphasizes us to enumerate the APN pathway genes during AMD pathogenesis. The single-cell RNA sequencing data for dry AMD was obtained from the GEO database (https://www.ncbi.nlm.nih.gov/geo) under accession number GSE221042. We conducted an in-depth analysis of cellular heterogeneity and landscape using the Seurat package [78] by plotting UMAP (Uniform Manifold Approximation and Projection). Furthermore, we identified distinct cell types using the corresponding marker genes (Figure 1A and B) as previously reported by Kuchroo et al. [79]. By employing the marker genes, we were able to distinguish various types of neuronal cells, such as retinal ganglion cells, horizontal cells, bipolar cells, rod photoreceptors, cone photoreceptors, and amacrine cells, in addition to uncovering infrequent non-neuronal cell types, such as microglia, astrocytes, and Müller glia. To understand the cellular expression of APN and its receptors and how the AdipoR1/R2 changes during AMD pathogenesis, we performed a focused features dot plotting (Figure 1C and D). Our results demonstrate that expression of both the receptor isoform (AdipoR1/R2) showed lower expression in dry AMD pathogenesis compared to healthy subjects. Furthermore, we checked the expression of other vascular cell subpopulations. Our analysis revealed the elevation of all angiogenesis marker genes VEGFA in Müller glia cells and AMD-associated genes HTRA1 in the horizontal cells during dry AMD pathogenesis. Dry AMD is characterized by the activation of intrinsic immune cells within the retina, specifically microglia cells, Müller cells, retinal pigment epithelial (RPE) cells, and macrophages. Under dry AMD pathophysiological conditions, Muller cells are involved in retinal angiogenesis [80]. Notably, drusen, a hallmark feature of dry AMD, contains a plethora of pro-inflammatory factors, including apolipoprotein E, coagulation proteins, acute phase proteins, immunoglobulin G (IgG), complement components, and activators [81]. This indicates that local inflammation plays a pivotal role in the early pathogenesis of AMD.
In their study, Bushra and colleagues noted that APN played an inhibitory role in the adhesion of endothelial cells (ECs) and the organization of the extracellular matrix. Simultaneously, this led to an enhancement in the barrier function, effectively mitigating the damage induced by high glucose levels in human microvascular retinal endothelial cells (HMRECs) [82]. In another study using a diabetic mouse model induced by STZ, researchers examined the impact of APN on the early development of vascular system damage in the retina. The immunofluorescence findings revealed that APN localized in the vascular endothelium of retinal arterioles in a T-cadherin-dependent manner, which progressively declined as diabetes progressed. This decline in retinal APN expression was concurrent with early signs of DR, characterized by increased vessel permeability. Importantly, treatment with dapagliflozin, a selective sodium-glucose co-transporter 2 inhibitor, aimed at lowering glucose levels, effectively prevented this reduction in retinal APN/AdipoRs system expression [83]. Furthermore, the study found that a deficiency in APN resulted in pronounced vascular permeability during relatively short-term hyperglycemia. This was accompanied by a significant increase in vascular cellular adhesion molecule-1 (VCAM-1) and a decrease in claudin-5 expression in the retinal endothelium [83,84].
We think for the purpose of sustained APN1 activity, it can be pegylated for clinical use. Several different doses of APN1 can be utilized in the rodent model, to mimic some insight into appropriate human dosing. Once we know the binding mechanism of APN1 to its receptors, like AdipoR1, by our experiments or from a literature search, we can design a blocker or antagonist to proceed further to find out whether this blocker or antagonist will also block APN1 binding. It is already known where APN binds to AdipoR1 but not APN1 (which we have designed) (28, 55, 60). Currently, we are unsure how well APN1 inhibition of wet AMD will do in humans, we expect it will be less invasive as an eye drop or may require fewer injections with greater intervals between treatment visits compared to current drugs available on the market today.
Mice lacking the very low-density lipoprotein receptor knockout (Vldlr KO) exhibit pathological retinal angiomatous proliferation, a condition that also affects people with AMD. Omega 3-long-chain-polyunsaturated-fatty-acid (ω3-LCPUFA), a supplementary ingredient added to the food in the form of docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) is known to suppress laser-induced CNV in controlled mice whereas this suppression is abolished in the APN KO mice. Besides, in the retinas of Vldlr KO mice the ω3-LCPUFA can also increase AdipoR1 expression and inhibit neovascularization. The clinical and experimental evidence suggests that ω3-LCPUFA-rich food may serves as protective role for AMD patients. Another study illustrated that the extent of CNV can be reduced with the use of APN peptide I utilizing laser-induced neovascularization in a choroid mice model [15,16,85]. APN is an anti-inflammatory protein and has more than 75% homology with complement protein C1q. APN1 peptide was derived from the globular region of this protein. APN1 inhibited CNV by more than 75% when injected sub-retinal compared to the control peptide (Figure 2). CNV was analyzed by confocal microscopy by measuring newly formed green vessels. Newly formed vessels were stained green by perfusion of FITC-Dextran. Image analysis was performed using the ImageJ program.
We have designed and synthesized several peptides. APN1, APN2, APN3 and Control peptide. After scanning all the peptides, APN1 inhibited CNV by more than 75%. APN2 and APN3 inhibited CNV by 50% and 40% respectively. The control peptide did not inhibit CNV at all. Therefore, we used APN1 and Control peptide for our experiments (Figure 3). We think APN1 may be a better alternative for the most frequently used Wet AMD treatments. Anti-VEGF treatments can cause hemorrhage, require frequent injections to patients, and are very expensive for patients. APN1 is not an anti-VEGF treatment, it binds to Adiponectin receptor one then through CAMP it inhibits CNV development [86,87].
We think APN1 can be pegylated for clinical use this will keep the APN around for a long time. In the mouse model, we used several doses, the best dose was 20 μg/Kg, and this will give us some idea about the dose to be used in humans. Once, we know how APN binds to its receptors, AdipoR1 by our experiments or from a literature search, we will design a blocker or antagonist to proceed further to know whether this blocker or antagonist will also block APN binding. It is already known where APN binds to AdipoR1 but not APN which we have designed [28,29,37,55,60]. The wet form of AMD is characterized by an excessive growth of new blood vessels around the macula, a region of the retina. This abnormal process is called CNV, which can lead to the development of leaky blood vessels. Therefore, the vision loss associated with wet AMD tends to be more severe compared to the dry form. Recent research indicates that a peptide known as Adiponectin Peptide 1 (APNp1) has demonstrated the ability to decelerate the advancement of CNV.
5. Exercise, APN/AdipoRs signaling, and neovascular AMD
Physical exercise has the potential to excite the systemic and localized production of APN, offering a protective effect in various ocular conditions, including but not limited to DR, AMD, RP, glaucoma, and light-induced retinal degeneration [88]. Engaging in daily physical exercise seems to provide a safeguard against the development of neovascular AMD. Recent studies observed that AIM2/NLRP2 inflammasome (predominantly expressed in microglia) was most abundantly expressed in the mouse retina-choroid complex. They found that the expression of AIM2 was significantly induced during CNV pathogenesis which was significantly attenuated by treadmill training exercise in rodent study [88]. Physical exercise mitigates neovascular AMD by suppressing the AIM2 inflammasome in myeloid cells. Moreover, earlier proteomic data directed that physical exercise facilitated the secretion of APN, from fat to circulation, which in turn, led to a decrease in ROS-induced DNA damage and the inhibition of AIM2 inflammasome activation in myeloid cells within eyes affected by choroidal neovascularization (CNV), and this effect was facilitated through the AMPK-p47phox pathway. Findings from earlier studies suggested that in sedentary AdipoQ-deficient (APNKO) mice, there was a tendency for an enhanced leakage area and choroidal neovascularization (CNV) volume when compared to sedentary wild-type mice. Furthermore, this difference became more pronounced in exercised mice, implying that APN plays an essential role in the positive effects observed in exercised mice. Notably, while treadmill exercise training significantly reduced the leakage area and CNV volume in wild-type mice, this protective effect was not statistically significant in APNKO mice [88]. These results substantiate the essential role of APN/AdipoRs in mediating the protective benefits of exercise.
6. Adiponectin's antioxidative properties in ocular tissues
Oxidative stress assumes a pivotal role in the pathogenesis of age-related ocular diseases, notably age-related macular degeneration (AMD), cataracts, and glaucoma. The advancing age is associated with a declining capacity for antioxidant defense mechanisms, leading to an accumulation of reactive oxygen species (ROS) within diverse ocular cell types. This surge in ROS levels precipitates oxidative damage, a hallmark of age-related ocular pathologies [89]. APN hormone has garnered increasing attention for its anti-oxidative properties within the ocular tissues. Emerging research has demonstrated that APN exerts a multifaceted anti-oxidative influence on retinal structures, particularly in the context of ocular diseases such as AMD and DR. APN appears to modulate oxidative stress by downregulating ROS generation, mitigating lipid peroxidation, and bolstering antioxidant defense mechanisms. Additionally, APN's capacity to enhance ECs function and mitigate inflammation plays a pivotal role in its anti-oxidative effects, as inflammation and oxidative stress are often intertwined in ocular pathologies [90]. The intricate interplay between APN and oxidative stress pathways in retinal cells presents a promising avenue for future investigations and therapeutic interventions aimed at preserving visual health.
7. Unlocking Clarity: Adiponectin's Transformative Quest from Fat Reserves to Optical Resilience
The multifaceted obesity hormone APN has evolved from being initially perceived as a critical regulator of adipose tissue metabolism to a multifunctional molecule with far-reaching effects across various physiological domains. Beyond its contributions to glucose homeostasis, lipid metabolism, and insulin sensitivity, recent research has unveiled APN's involvement in unexpected domains, such as optical resilience. Intriguingly, APN appear to exert a protective influence on vision especially retinal function, a feat that illuminates its versatile nature. This newfound role is rooted in its ability to modulate inflammatory responses, maintain vascular integrity, and mitigate oxidative stress within the intricate microenvironment of the eye. By suppressing the production of inflammatory cytokines, such as TNF-α and interleukin-6 (IL-6), APN helps to create a less hostile environment for retinal cells [83]. Moreover, its vasodilatory properties contribute to improved blood flow, which is crucial for maintaining the highly vascularized retina's health. Additionally, APN's antioxidant effects further shield the delicate retinal structures from oxidative damage. This newfound role in safeguarding the delicate structures of the eye underscores the profound and far-reaching impact of APN on overall health. As we delve deeper into the mechanisms underpinning its actions, the journey of APN from adipose reserves to retinal resilience continues to illuminate novel avenues for therapeutic interventions and a more comprehensive understanding of the intricate interplay between metabolic and physiological processes.
8. Conclusion
In conclusion, "From Fat to Sight" provides a captivating journey into the enthralling world of APN/AdipoR1/2, intertwining the realms of adipose biology and ophthalmology. This innovative review not only piques the curiosity of researchers but also holds promise in revolutionizing future approaches to combating eye disorders.
8.1. Future directions for research and clinical applications
Multifarious beneficial effects of APN/AdipoRs signaling have been exerted in numerous cell types, such as insulin-sensitizing, anti-inflammatory actions, anti-atherosclerotic, anti-carcinogenic, and antiproliferative effects. Since APN has an ameliorating function on insulin resistance, diabetes, and aging, a reduced APN level is considered to play a vital role in the pathophysiology of retinal diseases and is associated with the possibility of developing diabetes-associated DR and AMD-associated neovascularization. The research to prove the significant roles of AdipoR1 and AdipoR2 gained momentum due to the cloning of these two adiponectin receptors, confirming their requisition for binding of APN and subsequently its glucose-lowering effect. Furthermore, the activation of APN could lead to the activation of AMPK/SIRT1/PGC-1α and nuclear receptors PPARs through AdipoRs signaling. Screening of low molecular compounds for AdipoR1/R2 agonists using myriad therapeutic approaches along with other potential therapeutic regimes/approaches could be devised utilizing the 3D conformational analysis of AdipoRs. In fact, AdipoRs agonists are optimized based on 3D analysis of AdipoRs agonist-AdipoRs conformation to develop efficacious, safe, and premium class drugs for treating visually disabling diseases. Future research should focus on clarifying AdipoR1/2 and targeting its agonist to develop novel anti-aging and anti-diabetic drugs, all while facilitating both the concept of molecular mechanisms of APN activity and obesity-related and other metabolic disorders.
Furthermore, AMD, DR, and retinopathy of prematurity are all associated with altered circulating APN/AdipoRs function or APN variant distributions. Experimentally, APN inhibits retinal and CNV defects. As a key glucose and lipid modulator, APNs may re-establish metabolic balance. Intervention with ω-3 LCPUFA and derivatives of fibric acid enhances levels of APN in the blood. Exercise may exert a positive production of APN systemically as well as locally and it plays a protective role in several eye diseases, such as DR, AMD, RP, glaucoma, and light-induced retinal degeneration. Additional planned studies are needed to further investigate and clarify the role of APN/AdipoRs in neovascular AMD as well as the underlying molecular mechanisms to better understand both the experimental and clinical impact of this pathway.
In this comprehensive review, we aim to provide a deeper understanding of the intricate relationship between adiponectin and aged eye disorders. By elucidating the molecular mechanisms, clinical associations, and therapeutic implications, we hope to inspire further research and innovative strategies for the prevention, diagnosis, and treatment of ocular diseases. The remarkable connection between fat, in the form of adiponectin, and t654r4dexzsight opens exciting avenues for advancing eye health and improving patient outcomes.
Author Contributions
Mayank Choubey and Puran Bora conceived the idea. Mayank Choubey and Puran Bora have written the manuscript, Nalini Bora edited wrote in parts. Munichandra B Tirumalasetty has performed single-cell RNA seq analysis.
Funding
This research received no external funding. This work was funded by grants from the University of Arkansas for Medical Sciences, Little Rock, AR, 72205.
Institutional Review Board Statement
The Institutional Animal Care and Use Committee (IACUC) at the University of Arkansas approved this study for Medical Sciences, Little Rock, AR. ID Puran Bora 4008. No human subjects were used in this study.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to acknowledge the University of Arkansas for Medical Sciences for all the funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Straub, L.G.; Scherer, P.E. Metabolic Messengers: adiponectin. Nat Metab. 2019, 1, 334–339. [Google Scholar] [CrossRef]
- Wang ZV, Scherer PE. Adiponectin, the past two decades. J Mol Cell Biol. 2016, 8, 93–100. [CrossRef]
- Tao, C.; Sifuentes, A.; Holland, W. L. Regulation of glucose and lipid homeostasis by adiponectin: effects on hepatocytes, pancreatic β cells and adipocytes. Best practice & research. Clinical endocrinology & metabolism, 2014, 28, 43–58. [Google Scholar] [CrossRef]
- Lee, B.; Shao, J. Adiponectin and energy homeostasis. Rev Endocr Metab Disord. 2014, 15, 149–156 https://. [Google Scholar] [CrossRef] [PubMed]
- Fantuzzi, G. Adiponectin in inflammatory and immune-mediated diseases. Cytokine. 2013, 64, 1–10 https://. [Google Scholar] [CrossRef]
- Choubey, M.; Ranjan, A.; Krishna, A. Adiponectin/AdipoRs signaling as a key player in testicular aging and associated metabolic disorders. Vitam Horm. 2021; 115, 611-634. [CrossRef]
- Singh, A.; Choubey, M.; Bora, P.; and Krishna, A. Adiponectin and Chemerin: Contrary Adipokines in Regulating Reproduction and Metabolic Disorders. Reproductive sciences, 2018, 25, 1462–1473. [Google Scholar] [CrossRef]
- Choubey, M.; Bora, P. Emerging Role of Adiponectin/AdipoRs Signaling in Choroidal Neovascularization, Age-Related Macular Degeneration, and Diabetic Retinopathy. Biomolecules. 2023, 13, 982. [Google Scholar] [CrossRef]
- Fu, Z.; Gong, Y.; Löfqvist, C. ; Hellström,A. ; Smith, L.E. Review: adiponectin in retinopathy. Biochim Biophys Acta. 2016, 1862, 1392–1400 https://. [Google Scholar] [CrossRef]
- Campochiaro, P.A. Ocular neovascularization. J Mol Med. 2013, 91, 311–321 https://. [Google Scholar] [CrossRef]
- Kuo, J.Z. ; Guo X, Klein R, Klein BE, Genter P, Roll K, Hai Y, Goodarzi MO, Rotter JI, Chen YD, Ipp E. Adiponectin, Insulin Sensitivity and Diabetic Retinopathy in Latinos With Type 2 Diabetes. J Clin Endocrinol Metab. 2015, 100, 3348–55 https://. [Google Scholar] [CrossRef]
- Rahmouni, K.; Haynes, W.G. Endothelial effects of leptin: Implications in health and diseases. Curr Diab Rep. 2005, 5, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.I; Sonmez, A.; Acikel, C.; et al. Adiponectin may play a part in the pathogenesis of diabetic retinopathy. Eur J Endocrinol. 2004, 151, 135–140 https://. [Google Scholar] [CrossRef]
- Fan, X. , Wu, Q. , Li, Y., Hao, Y., Ning, N., Kang, Z., Cui, Y., Liu, R., & Han, L. Association between adiponectin concentrations and diabetic retinopathy in patients with type 2 diabetes: a meta-analysis. Chinese medical journal. 2014, 127, 765–771. [Google Scholar]
- Fu, Z.; Löfqvist, C.A.; Liegl, R.; et al. Photoreceptor glucose metabolism determines normal retinal vascular growth. EMBO Mol Med. 2018, 10, 76–90 https://. [Google Scholar] [CrossRef]
- Iwabu, M.; Okada-Iwabu, M.; Yamauchi, T. Adiponectin/adiponectin receptor in disease and aging. npj Aging Mech Dis. 2015, 1, 15013. [Google Scholar] [CrossRef] [PubMed]
- Bora, P.S.; Kaliappan, S.; Lyzogubov, V. V, et al. Expression of adiponectin in choroidal tissue and inhibition of laser-induced choroidal neovascularization by adiponectin. FEBS Lett. 2007, 581, 1977–1982 https://. [Google Scholar] [CrossRef]
- Lyzogubov, V.V.; Tytarenko, R.G.; Bora, N.S.; Bora, P.S. Inhibitory role of adiponectin peptide I on rat choroidal neovascularization. Biochim Biophys Acta. 2012, 1823, 1264–1272 https://. [Google Scholar] [CrossRef]
- Higuchi A, Ohashi K, Kihara S, Walsh K, Ouchi N. Adiponectin suppresses pathological microvessel formation in retina through modulation of tumor necrosis factor-alpha expression. Circ Res. 2009, 104, 1058–1065 https://. [CrossRef]
- Scherer, P.E.; Williams, S.; Fogliano, M.; Baldini, G.; Lodish, H.F. A novel serum protein similar to C1q, produced exclusively in adipocytes. J Biol Chem, 1995, 270, 26746–26749 https://. [Google Scholar] [CrossRef] [PubMed]
- Hu, E.; Liang, P.; Spiegelman, B.M. AdipoQ is a novel adipose-specific gene dysregulated in obesity. J Biol Chem. 1996, 271, 10697–10703 https://. [Google Scholar] [CrossRef] [PubMed]
- Maeda, K.; Okubo, K.; Shimomura, I.; Funahashi, T.; Matsuzawa, Y.; Matsubara, K. cDNA cloning and expression of a novel adipose specific collagen-like factor, apM1 (AdiPose Most abundant Gene transcript 1). Biochem Biophys Res Commun. 1996, 221, 286–289 https://. [Google Scholar] [CrossRef]
- Nakano, Y. ; Tobe, T,; Choi-Miura, N. H.; Mazda, T.; Tomita, M. Isolation and characterization of GBP28, a novel gelatin-binding protein purified from human plasma. J Biochem. 1996, 120, 803–812 https://. [Google Scholar] [CrossRef]
- Yamauchi T, Kamon J, Ito Y, Tsuchida A, Yokomizo T, Kita S, Sugiyama T, Miyagishi M, Hara K, Tsunoda M, Murakami K, Ohteki T, Uchida S, Takekawa S, Waki H, Tsuno NH, Shibata Y, Terauchi Y, Froguel P, Tobe K, Koyasu S, Taira K, Kitamura T, Shimizu T, Nagai R, Kadowaki T. Cloning of adiponectin receptors that mediate antidiabetic metabolic effects. Nature. 2003, 423, 762–769. [CrossRef]
- Yamauchi T, Nio Y, Maki T, Kobayashi M, Takazawa T, Iwabu M, Okada-Iwabu M, Kawamoto S, Kubota N, Kubota T, Ito Y, Kamon J, Tsuchida A, Kumagai K, Kozono H, Hada Y, Ogata H, Tokuyama K, Tsunoda M, Ide T, Murakami K, Awazawa M, Takamoto I, Froguel P, Hara K, Tobe K, Nagai R, Ueki K, Kadowaki T. Targeted disruption of AdipoR1 and AdipoR2 causes abrogation of adiponectin binding and metabolic actions. Nat Med. 2007, 13, 332–339. [CrossRef]
- Ruan, H.; Dong, L.Q. Adiponectin signaling and function in insulin target tissues. J Mol Cell Biol. 2016, 8, 101–109 https://. [Google Scholar] [CrossRef]
- Tang, Y.T.; Hu, T.; Arterburn, M.; et al. PAQR proteins: a novel membrane receptor family defined by an ancient 7-transmembrane pass motif. J Mol Evol. 2005, 61, 372–380 https://. [Google Scholar] [CrossRef]
- Kadowaki, T.; Yamauchi, T. Adiponectin and Adiponectin Receptors, Endocrine Reviews. 2005; 26. [Google Scholar] [CrossRef]
- Capeau, J. The story of adiponectin and its receptors AdipoR1 and R2: to follow. J Hepatol. 2007, 47, 736–738 https://. [Google Scholar] [CrossRef]
- Zhu, N.; Pankow, J.S.; Ballantyne, C.M.; et al. High-molecular-weight adiponectin and the risk of type 2 diabetes in the ARIC study. J Clin Endocrinol Metab. 2010, 95, 5097–5104 https://. [Google Scholar] [CrossRef]
- Arita, Y.; Kihara, S.; Ouchi, N.; et al. Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. Biochem Biophys Res Commun. 1999, 257, 79–83 https://. [Google Scholar] [CrossRef] [PubMed]
- Sieminska, L.; Marek, B.; Kos-Kudla, B.; et al. Serum adiponectin in women with polycystic ovarian syndrome and its relation to clinical, metabolic and endocrine parameters. J Endocrinol Invest. 2004, 27, 528–534 https://. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Waki, H.; et al. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat Med. 2001, 7, 941–946 https://. [Google Scholar] [CrossRef]
- Whitehead, J.P.; Richards, A.A.; Hickman, I.J.; Macdonald, G.A.; Prins, J.B. Adiponectin--a key adipokine in the metabolic syndrome. Diabetes, obesity & metabolism. 2006, 8, 264–280. [Google Scholar] [CrossRef]
- Yoda-Murakami, M.; Taniguchi, M.; Takahashi, K.; Kawamata, S.; Saito, K.; Choi-Miura, N.H.; Tomita, M. Change in expression of GBP28/adiponectin in carbon tetrachloride-administrated mouse liver. Biochemical and biophysical research communications, 2001, 285, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Delaigle, A.M.; Senou, M.; Guiot, Y.; Many, M.C.; Brichard, S.M. Induction of adiponectin in skeletal muscle of type 2 diabetic mice: In vivo and in vitro studies. Diabetologia, 2006, 49, 1311–1323. [Google Scholar] [CrossRef]
- Thundyil, J.; Pavlovski, D.; Sobey, C.G.; Arumugam, T.V. Adiponectin receptor signalling in the brain. British journal of pharmacology, 2012, 165, 313–327. [Google Scholar] [CrossRef]
- Hopkins, T.A.; Ouchi, N.; Shibata, R.; Walsh, K. Adiponectin actions in the cardiovascular system. Cardiovasc Res. 2007, 74, 11–18 https://. [Google Scholar] [CrossRef]
- Choubey, M.; Ranjan, A.; Bora, P.S.; Baltazar, F.; Martin, L.J.; Krishna, A. Role of adiponectin as a modulator of testicular function during aging in mice. Biochim Biophys Acta Mol Basis Dis. 2019, 1865, 413–427. [Google Scholar] [CrossRef]
- Li, H.Y.; Hong, X.; Cao, Q.Q.; So, K.F. Adiponectin, exercise and eye diseases. International review of neurobiology, 2019, 147, 281–294. [Google Scholar] [CrossRef]
- Luo, N.; Liu, J.; Chung, B.H.; Yang, Q.; Klein, R.L.; Garvey, W.T.; Fu, Y. Macrophage adiponectin expression improves insulin sensitivity and protects against inflammation and atherosclerosis. Diabetes. 2010, 59, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Kubota, N.; Yano, W.; Kubota, T.; Yamauchi, T.; Itoh, S.; Kumagai, H.; Kozono, H.; Takamoto, I.; Okamoto, S.; Shiuchi, T.; Suzuki, R. Satoh, H. ; Tsuchida, A.; Moroi, M.; Sugi, K.; Noda, T.; Ebinuma, H.; Ueta, Y.; Kondo, T.; Araki, E.; Ezaki, O.; Nagai, R.; Tobe, K.; Terauchi, Y.; Ueki, K.; Minokoshi, Y.; Kadowaki, T. Adiponectin stimulates AMP-activated protein kinase in the hypothalamus and increases food intake. Cell metabolism. 2007, 6, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; van de Wall, E.; Laplante, M.; Azzara, A.; Trujillo, M.E.; Hofmann, S.M.; Schraw, T.; Durand, J.L.; Li, H.; Li, G.; Jelicks, L.A.; Mehler, M.F.; Hui, D.Y.; Deshaies, Y.; Shulman, G.I.; Schwartz, G.J.; Scherer, P.E. Obesity-associated improvements in metabolic profile through expansion of adipose tissue. The Journal of clinical investigation. 2007, 117, 2621–2637. [Google Scholar] [CrossRef] [PubMed]
- Choubey, M.; Ranjan, A.; Bora, P.S.; Baltazar, F.; and Krishna, A. Direct actions of adiponectin on changes in reproductive, metabolic, and anti-oxidative enzymes status in the testis of adult mice. Gen Comp Endocrinol, 2019, 279, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Choubey, M.; Ranjan, A.; Bora, P.S.; and Krishna, A. Protective role of adiponectin against testicular impairment in high-fat diet/streptozotocin-induced type 2 diabetic mice. 2020, Biochimie, 168, 41–52. [CrossRef]
- Lin, T.; Qiu, Y.; Liu, Y.; Mohan, R.; Li, Q.; Lei, B. Expression of adiponectin and its receptors in type 1 diabetes mellitus in human and mouse retinas. Mol Vis. 2013, 19, 1769–78. [Google Scholar]
- Matsunami, T.; Sato, Y.; Ariga, S.; Sato, T.; Kashimura, H.; Hasegawa, Y.; Yukawa, M. Regulation of oxidative stress and inflammation by hepatic adiponectin receptor 2 in an animal model of nonalcoholic steatohepatitis. International journal of clinical and experimental pathology. 2010, 3, 472–481. [Google Scholar] [PubMed]
- Tsuchida, A.; Yamauchi, T.; Ito, Y.; Hada, Y.; Maki, T.; Takekawa, S.; Kamon, J.; Kobayashi, M.; Suzuki, R.; Hara, K.; Kubota, N.; Terauchi, Y.; Froguel, P.; Nakae, J.; Kasuga, M.; Accili, D.; Tobe, K.; Ueki, K.; Nagai, R.; Kadowaki, T. Insulin/Foxo1 pathway regulates expression levels of adiponectin receptors and adiponectin sensitivity. The Journal of biological chemistry. 2004, 279, 30817–30822. [Google Scholar] [CrossRef]
- Nigro, E. , Scudiero, O., Monaco, M. L., Palmieri, A., Mazzarella, G., Costagliola, C., Bianco, A., & Daniele, A. New insight into adiponectin role in obesity and obesity-related diseases. BioMed research international, 2014, 658913. [CrossRef]
- Gavrila, A.; Chan, J. L.; Yiannakouris, N.; Kontogianni, M.; Miller, L.C.; Orlova, C.; Mantzoros, C.S. Serum adiponectin levels are inversely associated with overall and central fat distribution but are not directly regulated by acute fasting or leptin administration in humans: cross-sectional and interventional studies. The Journal of clinical endocrinology and metabolism, 2003, 88, 4823–4831. [Google Scholar] [CrossRef]
- Escobar-Morreale, H.F.; Villuendas, G.; Botella-Carretero, J.I.; Alvarez-Blasco, F.; Sanchón, R.; Luque-Ramírez, M.; San Millán, J.L. Adiponectin and resistin in PCOS: a clinical, biochemical and molecular genetic study. Human reproduction. 2006, 21, 2257–2265. [Google Scholar] [CrossRef]
- Genchi, V.A.; Rossi, E.; Lauriola, C.; D’Oria, R.; Palma, G.; Borrelli, A.; Caccioppoli, C.; Giorgino, F.; Cignarelli, A. Adipose Tissue Dysfunction and Obesity-Related Male Hypogonadism. Int. J. Mol. Sci. 2022, 23, 8194. [Google Scholar] [CrossRef]
- Degawa-Yamauchi, M.; Moss, K.A.; Bovenkerk, J.E.; Shankar, S.S.; Morrison, C.L.; Lelliott, C.J.; Vidal-Puig, A.; Jones, R.; Considine, R.V. Regulation of adiponectin expression in human adipocytes: effects of adiposity, glucocorticoids, and tumor necrosis factor alpha. Obesity research. 2005, 13, 662–669. [Google Scholar] [CrossRef]
- Ouchi, N.; Kenneth, W. Adiponectin as an anti-inflammatory factor. Clinica chimica acta. 2007, 380, 24–30. [Google Scholar] [CrossRef]
- Kadowaki, T.; Yamauchi, T. Adiponectin and adiponectin receptors. Endocrine reviews. 2005, 26, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Kubota, N.; Terauchi, Y.; Yamauchi, T.; Kubota, T.; Moroi, M.; Matsui, J.; Eto, K.; Yamashita, T.; Kamon, J.; Satoh, H.; Yano, W.; Froguel, P.; Nagai, R.; Kimura, S.; Kadowaki, T.; Noda, T. Disruption of adiponectin causes insulin resistance and neointimal formation. The Journal of biological chemistry. 2002, 277, 25863–25866. [Google Scholar] [CrossRef]
- Maeda, N.; Shimomura, I.; Kishida, K.; Nishizawa, H.; Matsuda, M.; Nagaretani, H.; Furuyama, N.; Kondo, H.; Takahashi, M.; Arita, Y.; Komuro, R.; Ouchi, N.; Kihara, S.; Tochino, Y.; Okutomi, K.; Horie, M.; Takeda, S.; Aoyama, T.; Funahashi, T.; Matsuzawa, Y. Diet-induced insulin resistance in mice lacking adiponectin/ACRP30. Nature medicine. 2002, 8, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Barbe, A.; Bongrani, A.; Mellouk, N.; Estienne, A. , Kurowska, P. ; Grandhaye, J.; Elfassy, Y.; Levy, R.; Rak, A.; Froment, P.; Dupont, J. Mechanisms of Adiponectin Action in Fertility: An Overview from Gametogenesis to Gestation in Humans and Animal Models in Normal and Pathological Conditions. International journal of molecular sciences. 2019, 20, 1526. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.V.; Scherer, P.E. Adiponectin, cardiovascular function, and hypertension. Hypertension. 2008, 51, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Kikani, C.K.; Riojas, R.A.; Langlais, P.; Wang, L.; Ramos, F.J.; Fang, Q.; Christ-Roberts, C.Y.; Hong, J.Y.; Kim, R.Y.; Liu, F.; Dong, L.Q. APPL1 binds to adiponectin receptors and mediates adiponectin signalling and function. Nature cell biology. 2006, 8, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Park, C.W. Mechanisms of Adiponectin Action: Implication of Adiponectin Receptor Agonism in Diabetic Kidney Disease. International journal of molecular sciences. 2019, 20, 1782. [Google Scholar] [CrossRef]
- Xu, H.; Zhao, Q.; Song, N.; Yan, Z.; Lin, R.; Wu, S.; Jiang, L.; Hong, S.; Xie, J.; Zhou, H.; et al. AdipoR1/AdipoR2 dual agonist recovers nonalcoholic steatohepatitis and related fibrosis via endoplasmic reticulum-mitochondria axis. Nat Commun. 2020, 11, 5807. [Google Scholar] [CrossRef]
- Li, Z.; Woo, J.M.; Chung, S.W.; Kwon, M.Y.; Choi, J.S.; Oh, H.J.; Yoon, K.C. Therapeutic effect of topical adiponectin in a mouse model of desiccating stress-induced dry eye. Investigative ophthalmology & visual science. 2013, 54, 155–162. [Google Scholar] [CrossRef]
- Liu, H.; Prokosch, V. Energy Metabolism in the Inner Retina in Health and Glaucoma. International journal of molecular sciences. 2021, 22, 3689. [Google Scholar] [CrossRef]
- Mizukami, Y.; Li, J.; Zhang, X.; Zimmer, M.A.; Iliopoulos, O.; Chung, D.C. Hypoxia-inducible factor-1-independent regulation of vascular endothelial growth factor by hypoxia in colon cancer. Cancer research. 2004, 64, 1765–1772. [Google Scholar] [CrossRef]
- Nakahara, T.; Hoshino, M.; Hoshino, S.; Mori, A.; Sakamoto, K.; Ishii, K. Structural and functional changes in retinal vasculature induced by retinal ischemia-reperfusion in rats. Experimental eye research. 2015, 135, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Ferrington, D.A.; Fisher, C.R.; Kowluru, R.A. Mitochondrial Defects Drive Degenerative Retinal Diseases. Trends in molecular medicine. 2020, 26, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; An, X.; Guo, X.; Habtetsion, T.G.; Wang, Y.; Xu, X.; Kandala, S.; Li, Q.; Li, H.; Zhang, C.; et al. Endothelial PFKFB3 plays a critical role in angiogenesis. Arteriosclerosis, thrombosis, and vascular biology. 2014, 34, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Safi, S.Z.; Qvist, R.; Kumar, S.; Batumalaie, K.; Ismail, I.S. Molecular mechanisms of diabetic retinopathy, general preventive strategies, and novel therapeutic targets. BioMed research international. 2014, 801269. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.Y.; Yiang, G.T.; Lai, T.T.; Li, C.J. The Oxidative Stress and Mitochondrial Dysfunction during the Pathogenesis of Diabetic Retinopathy. Oxidative medicine and cellular longevity. 2018, 3420187. [Google Scholar] [CrossRef]
- Wong, B.W.; Marsch, E.; Treps, L.; Baes, M.; Carmeliet, P. Endothelial cell metabolism in health and disease: impact of hypoxia. The EMBO journal. 2017, 36, 2187–2203. [Google Scholar] [CrossRef]
- Omae, T.; Nagaoka, T.; Yoshida, A. Relationship Between Retinal Blood Flow and Serum Adiponectin Concentrations in Patients With Type 2 Diabetes Mellitus. Investigative ophthalmology & visual science. 2015, 56, 4143–4149. [Google Scholar] [CrossRef]
- Li, R.; Du, J.; Yao, Y.; Yao, G.; Wang, X. Adiponectin inhibits high glucose-induced angiogenesis via inhibiting autophagy in RF/6A cells. J Cell Physiol. 2019, 234, 20566–20576. [Google Scholar] [CrossRef]
- Palanisamy, K.; Nareshkumar, R.N.; Sivagurunathan, S.; Raman, R.; Sulochana, K.N.; Chidambaram, S. Anti-angiogenic effect of adiponectin in human primary microvascular and macrovascular endothelial cells. Microvascular research. 2019, 122, 136–145. [Google Scholar] [CrossRef]
- Srinivasan, V.; Sulochana, K.N. Effect of adiponectin on expression of vascular endothelial growth factor and pigment epithelium-derived factor: An in vitro study. The Indian Journal of Pharmacology. 2015, 47, 117–120. [Google Scholar] [CrossRef]
- Mallardo, M.; Costagliola, C.; Nigro, E.; Daniele, A. AdipoRon negatively regulates proliferation and migration of ARPE-19 human retinal pigment epithelial cells. Peptides, 2021, 146, 170676. [Google Scholar] [CrossRef] [PubMed]
- Osada, H.; Toda, E.; Homma, K.; Guzman, NA.; Nagai, N.; Ogawa, M.; Negishi, K.; Arita, M.; Tsubota, K.; Ozawa, Y. ADIPOR1 deficiency-induced suppression of retinal ELOVL2 and docosahexaenoic acid levels during photoreceptor degeneration and visual loss. Cell Death Dis. 2021, 12, 458. [Google Scholar] [CrossRef]
- Butler A, Hoffman P, Smibert P, Papalexi E, Satija R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat Biotechnol. 2018 Jun;36(5):411-420. [CrossRef]
- Kuchroo M, DiStasio M, Song E, Calapkulu E, Zhang L, Ige M, Sheth AH, Majdoubi A, Menon M, Tong A, Godavarthi A, Xing Y, Gigante S, Steach H, Huang J, Huguet G, Narain J, You K, Mourgkos G, Dhodapkar RM, Hirn MJ, Rieck B, Wolf G, Krishnaswamy S, Hafler BP. Single-cell analysis reveals inflammatory interactions driving macular degeneration. Nat Commun. 2023 ;14(1):2589. /: https, 5 May. [CrossRef]
- Wu, K.H; Madigan, M.C.; Billson, F.A.; Penfold, P.L. Differential expression of GFAP in early v late AMD: a quantitative analysis. The British journal of ophthalmology, 2003, 87, 1159–1166. [Google Scholar] [CrossRef]
- van Lookeren Campagne, M.; LeCouter, J.; Yaspan, B.L.; Ye, W. Mechanisms of age-related macular degeneration and therapeutic opportunities. J Pathol, 2014, 232, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Bushra, S.; Al-Sadeq, DW.; Bari, R.; Sahara, A.; Fadel, A.; Rizk, N. Adiponectin Ameliorates Hyperglycemia-Induced Retinal Endothelial Dysfunction, Highlighting Pathways, Regulators, and Networks. J Inflamm Res. 2022, 15, 3135–3166. [Google Scholar] [CrossRef]
- Sakaue TA, Fujishima Y, Fukushima Y, Tsugawa-Shimizu Y, Fukuda S, Kita S, Nishizawa H, Ranscht B, Nishida K, Maeda N, Shimomura I. Adiponectin accumulation in the retinal vascular endothelium and its possible role in preventing early diabetic microvascular damage. Sci Rep. 2022, 12, 4159. [CrossRef]
- Kaur, G.; Sharma, D.; Bisen, S.; Mukhopadhyay, C.S.; Gurdziel, K.; Singh, N.K. Vascular cell-adhesion molecule 1 (VCAM-1) regulates JunB-mediated IL-8/CXCL1 expression and pathological neovascularization. Communications biology. [CrossRef]
- Fu, Z.; Liegl, R.; Wang, Z.; Gong, Y.; Liu, C.H.; Sun, Y.; Cakir, B.; Burnim, S.B.; Meng, S.S.; Löfqvist, C.; et al. Adiponectin Mediates Dietary Omega-3 Long-Chain Polyunsaturated Fatty Acid Protection Against Choroidal Neovascularization in Mice. Investigative ophthalmology & visual science. 2017, 58, 3862–3870. [Google Scholar] [CrossRef]
- Logan, C.; Lyzogubov, V.; Bora, N.; Bora, P. Role of Adiponectin Peptide I (APNp1) in Age-Related Macular Degeneration. Biomolecules. 2022, 12, 1232. [Google Scholar] [CrossRef]
- Bora, P.S. New Discoveries in Retinal Cell Degeneration and Retinal Diseases. Biomolecules, 2023, 13, 1121. [Google Scholar] [CrossRef]
- Cui, B.; Guo, X.; Zhou, W.; Zhang, X.; He, K.; Bai, T.; Lin, D.; Wei-Zhang, S.; Zhao, Y.; Liu, S.; Zhou, H.; Wang, Q.; Yao, X.; Shi, Y.; Xie, R.; Dong, X.; Lei, Y.; Du, M.; Chang, Y.; Xu, H.; Yan, H. Exercise alleviates neovascular age-related macular degeneration by inhibiting AIM2 inflammasome in myeloid cells. Metabolism: clinical and experimental, 2023, 144, 155584. [Google Scholar] [CrossRef] [PubMed]
- 89. Kushwah, N;, Bora, K.; Maurya, M.; Pavlovich, M.C.; & Chen, J. Oxidative Stress and Antioxidants in Age-Related Macular Degeneration. Antioxidants (Basel, Switzerland), 2023; 12, 1379. [CrossRef]
- Deng, H. , Ai, M., Cao, Y., Cai, L., Guo, X., Yang, X., Yi, G., & Fu, M. Potential Protective Function of Adiponectin in Diabetic Retinopathy. Ophthalmology and therapy, 2023; 12, 1519–1534. [Google Scholar] [CrossRef]
Figure 1.
Single-cell UMAP Visualization depicting the cellular landscape of Healthy (A) and Dry Age-Related Macular Degenerated Eye (B). The expression of AdipoR1 and AdipoR2 genes in various retinal cell populations of Healthy (C) and diseased dry AMD (D). Data was analyzed using the software, Seurat v4.1.1 implemented in R v4.2.1.
Figure 1.
Single-cell UMAP Visualization depicting the cellular landscape of Healthy (A) and Dry Age-Related Macular Degenerated Eye (B). The expression of AdipoR1 and AdipoR2 genes in various retinal cell populations of Healthy (C) and diseased dry AMD (D). Data was analyzed using the software, Seurat v4.1.1 implemented in R v4.2.1.
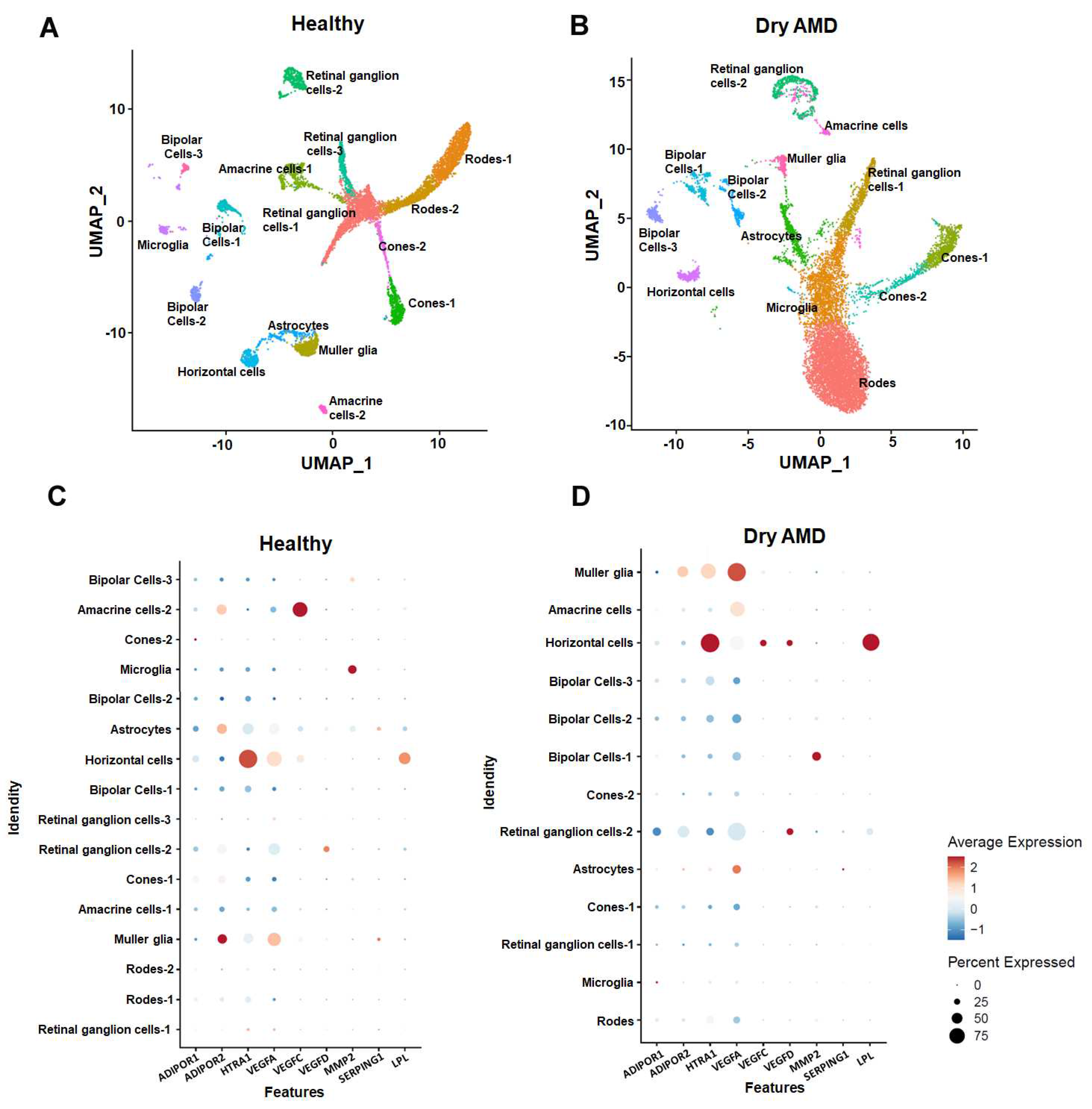
Figure 2.
Laser-induced CNV in mice. Four laser spots were placed in each mouse eye. Animals were sacrificed on day 7 (CNV fully matures on day seven in this model), confocal pictures were taken after making flat mounts. The green color indicates new vessels formed from choroid (Figure 2A). Phospho-Buffer Saline (PBS) treated mice did not show any green color indicating no CNV formation from choroid (Figure 2B).
Figure 2.
Laser-induced CNV in mice. Four laser spots were placed in each mouse eye. Animals were sacrificed on day 7 (CNV fully matures on day seven in this model), confocal pictures were taken after making flat mounts. The green color indicates new vessels formed from choroid (Figure 2A). Phospho-Buffer Saline (PBS) treated mice did not show any green color indicating no CNV formation from choroid (Figure 2B).
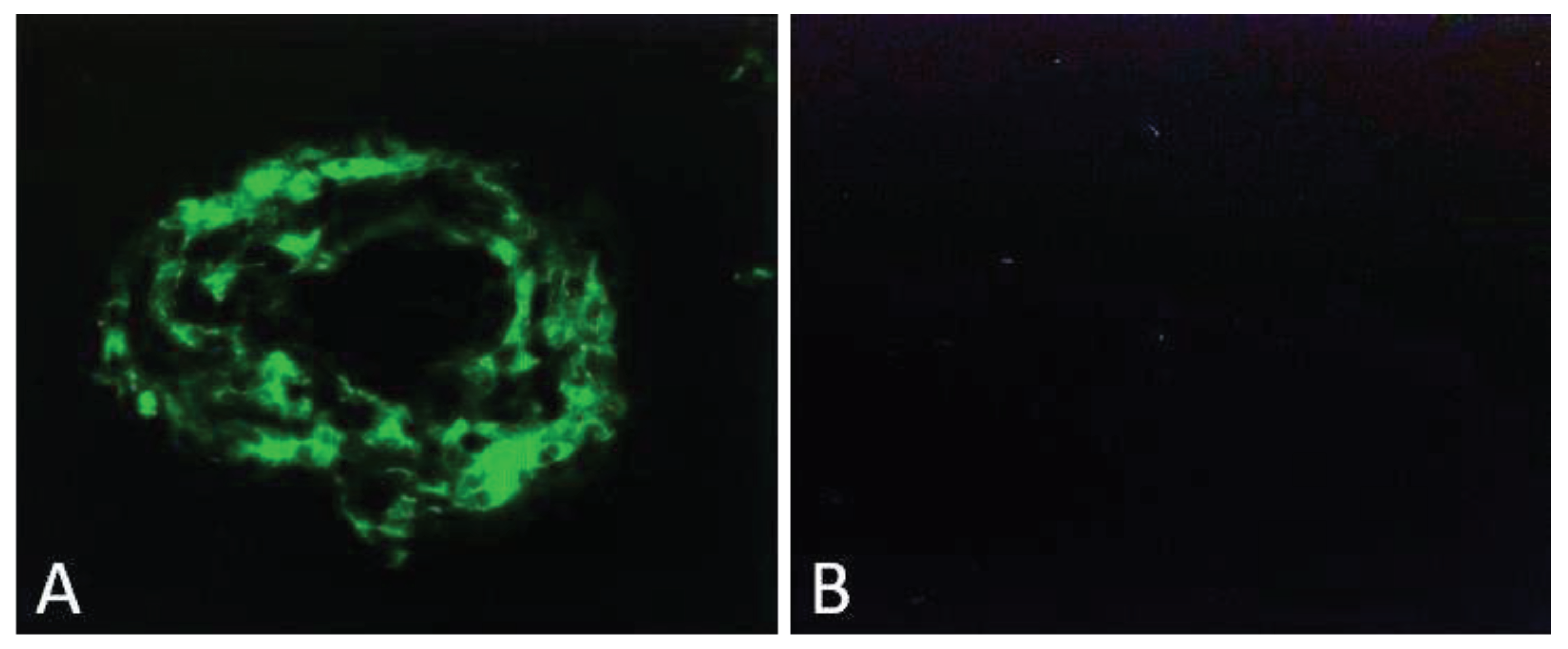
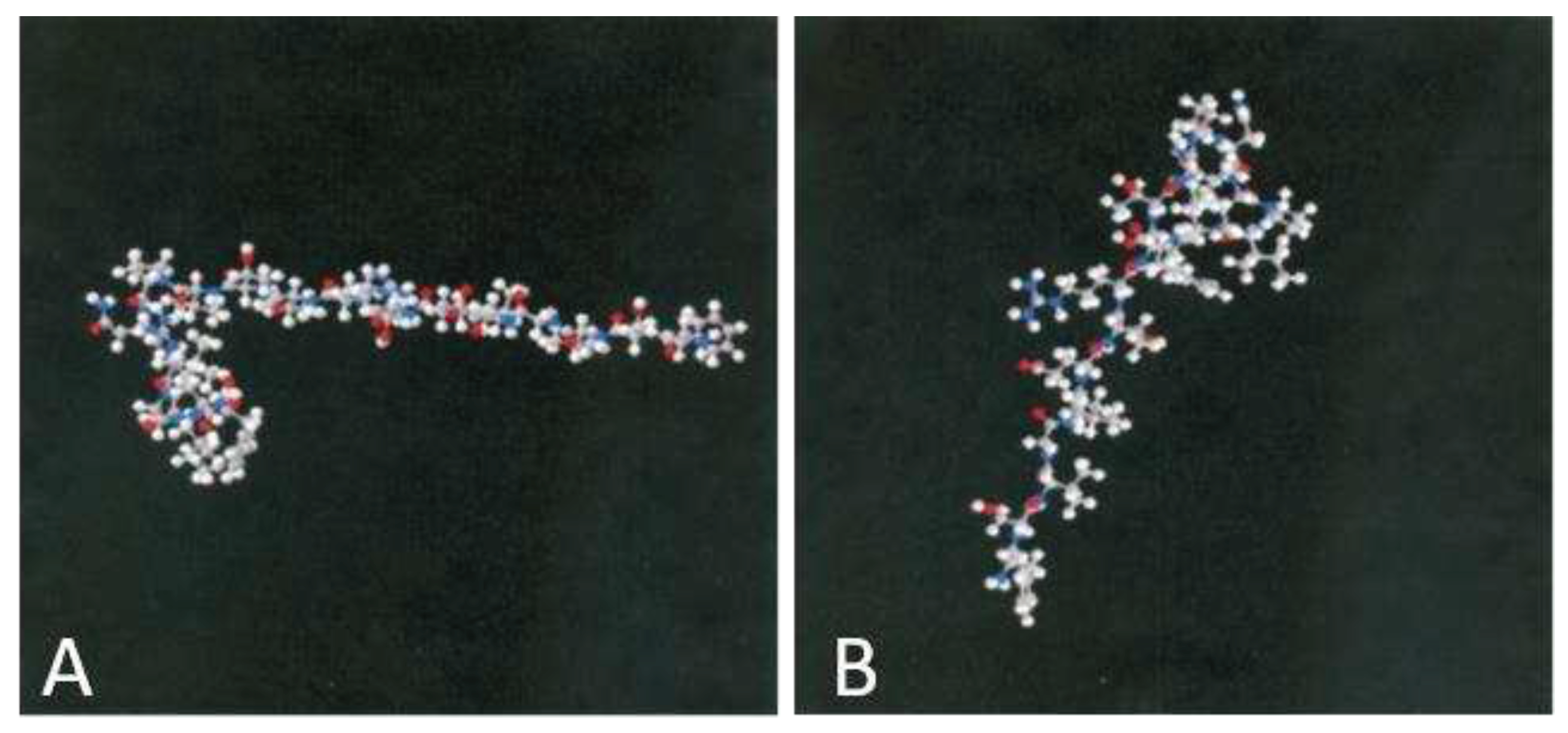
Figure 3.
Three-dimensional (3D) structure of Control Peptide. (A) FSVGLETRVTVPNVPIRF and APN1 inhibitory peptide (B) KDKAVLFTYDQYQEKNVD. All modeling calculations were performed using the SYBYL program, and the structure of the peptide was built using the ‘build protein’ tool in SYBYL. The lowest energy conformer for the peptide was calculated to be 43.145 hartrees (17).
Figure 3.
Three-dimensional (3D) structure of Control Peptide. (A) FSVGLETRVTVPNVPIRF and APN1 inhibitory peptide (B) KDKAVLFTYDQYQEKNVD. All modeling calculations were performed using the SYBYL program, and the structure of the peptide was built using the ‘build protein’ tool in SYBYL. The lowest energy conformer for the peptide was calculated to be 43.145 hartrees (17).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



