
Submitted:
14 September 2023
Posted:
15 September 2023
You are already at the latest version
Abstract
Shiga toxin-producing Escherichia coli (STEC) are notorious foodborne pathogens capable of causing severe diarrhea and other life-threatening complications in humans. Cattle remain a major reservoir and asymptomatic carriers of STEC. In reservoir animals, the rectoanal junction (RAJ) serves as the primary colonization site of STEC playing a critical role in its transmission to humans through contaminated food sources. However, the molecular mechanisms behind the adaptation of STEC in the RAJ of the asymptomatic reservoir host and its subsequent infection of human colonic epithelial cells in an event of a foodborne transmission to cause diarrhea remain largely unexplored. This study aimed to unravel the intricate dynamics of STEC O157:H7 serotype in these two distinct host environments: bovine RAJ cells and human colonic epithelial cells, during initial colonization. In this context, comparative transcriptomics analysis was employed to investigate the differential gene expression profiles of STEC O157:H7 during its interaction with these cell types. The bacterial cells were cultured under controlled conditions to simulate the microenvironments of both bovine RAJ and human colonic epithelial cells. Using high-throughput RNA sequencing, we identified key bacterial genes and regulatory pathways that were significantly modulated in response to each specific host environment. Our findings reveal distinct expression patterns of bacterial genes involved in virulence, including adherence, metal iron homeostasis and stress response in STEC O157:H7 grown in bovine RAJ cells as op-posed to human colonic epithelial cells six hours post-infection. Additionally, the comparative analysis highlights the potential role of certain genes in host adaptation and tissue-specific pathogenicity. Furthermore, this study sheds light on the potential factors contributing to the survival and persistence of STEC O157:H7 in the bovine reservoir, and its ability to colonize in-testinal epithelium and cause disease in humans.
Keywords:
adherence
; biofilm formation
; colonic epithelium
; rectoanal junction
; Shiga toxin-producing Escherichia coli
; transcriptomics
; virulence genes
1. Introduction
Shiga toxin-producing Escherichia coli (STEC) are highly virulent foodborne pathogens that pose a significant threat to public health worldwide and responsible for numerous outbreaks of severe gastrointestinal infections [1,2]. Infection with this pathogen can lead to a spectrum of symptoms ranging from mild diarrhea to life-threatening complications, such as hemolytic uremic syndrome (HUS) and hemorrhagic colitis (HC) [2,3]. The major route of transmission to humans is through the consumption of contaminated food, particularly undercooked beef and dairy products [4,5]. While STEC can asymptomatically colonize the gastrointestinal tracts of healthy cattle and other ruminants, the mechanisms by which it adapts to its ruminant reservoir and subsequently causes infections in humans are still not fully understood. In the United States, disease outbreaks often result from the consumption of contaminated bovine food products and fresh produce, which have been tainted with the bacterium due to exposure to bovine waste [6].
The bovine rectoanal junction (RAJ) has emerged as a critical anatomical site for STEC colonization in cattle [7]. The RAJ is a transition zone between the rectum and the anal canal, comprising of columnar and squamous cells, and characterized by a unique microenvironment that favors bacterial colonization and persistence [8,9]. Consequently, the RAJ serves as a major reservoir for STEC, and the contamination of food products can occur through fecal shedding from infected cattle during the slaughtering process [10,11]. The adaptation of STEC to the bovine RAJ microenvironment likely involves the differential regulation of specific genes and virulence factors that facilitate bacterial adherence, survival, and evasion of the host immune system. Conversely, during human infections, STEC encounters a vastly different environment in the colonic epithelial cells of the human gut. The pathogen must overcome numerous host defenses, including the mucus layer, antimicrobial peptides, and the gut microbiota, to colonize and cause disease [12,13]. During this process, STEC is believed to differentially regulate gene expression to promote its survival, colonization and pathogenicity in the human colonic epithelium.
Understanding the molecular mechanisms behind STEC disease pathogenesis is crucial for the development of targeted therapies and preventive measures to combat this dangerous foodborne pathogen. The adherence mechanisms employed by STEC are pivotal in their ability to initiate and establish infections [14]. It is well known that the intimin protein, encoded by eae, is a crucial virulence factor that mediates intimate attachment of STEC by binding to a receptor called Tir on the host cell [15]. This interaction leads to the formation of attaching and effacing (A/E) lesions, characterized by localized destruction of host cell microvilli and intimate bacterial adhesion. However, the adhesins involved in initial adherence of STEC, leading to subsequent intimate adherence, are largely unknown. This is despite the fact that several fimbrial and non-fimbrial adhesins of STEC have been implicated in STEC initial adherence to host cells [16,17,18]. Previous research also suggests the carbon salvation protein Slp might be involved in initial adherence of STEC to human colonic epithelial cells [19]. These adherence mechanisms may not only facilitate bacterial colonization but also enable the pathogens to evade host defenses and deliver their virulence factors, ultimately contributing to the severity of STEC infections.
Transcriptomics, which enables the genome-wide analysis of gene expression patterns, has become an indispensable tool in understanding the intricate interplay between pathogens and their hosts [20]. Comparative transcriptomics analysis allows for the identification of differentially expressed genes and pathways in response to specific environmental conditions, thereby offering valuable insights into the molecular basis of host adaptation and pathogenesis [21]. Despite the potential of transcriptomics to shed light on the mechanisms underlying STEC’s adaptation to different host environments, only limited studies have focused on the comparative transcriptomic analysis of this pathogen grown on bovine RAJ cells and human colonic epithelial cells. In this study, we aimed to address this knowledge gap by conducting a comprehensive comparative transcriptomics analysis of STEC O157:H7, the most studied serotype of STEC, cultivated on both bovine RAJ and human colonic epithelial cells. High-throughput RNA sequencing was utilized to profile the global gene expression patterns of STEC O157:H7 under these two distinct host environments. Through this approach, we intend to identify key genes and regulatory pathways that are differentially expressed in response to the bovine and human host environments.
2. Materials and Methods
2.1. Bacterial Strains
E. coli O157:H7 wild-type (O157 WT) EDL 932 (American Type Culture Collection /ATCC, Manassas, VA, USA), which is positive for stx1, stx2, and eae was used in this study. Bacteria were grown at 37°C in Luria Bertani (LB) (BD, Difco, Franklin Lakes, New Jersey, USA) broth or LB agar plates without antibiotics.
2.2. Cell Culture and Media
The CCD CoN 841 human colonic epithelial cells (ATCC CRL-1790) purchased from the (ATCC, Manassas, VA, USA) were grown in Eagle's Minimum Essential Medium (EMEM; ATCC-30-2003) containing 10% fetal bovine serum (FBS: Atlanta Biologicals, Flowery Branch, GA, USA) in the presence of 5% CO2 at 37°C. RAJ cells, including both squamous and columnar cells, were isolated from healthy cattle using a previously standardized protocol [22,23] and evaluated similarly at 37°C in the presence of 5% CO2 in Dulbecco's Modified Eagle's Medium-High Glucose (DMEM-HG; ATCC-30-2002) containing 10% FBS.
2.3. Bacterial Infection and Adherence
The CCD CoN 841 cells and RAJ cells were tested in tissue culture-treated six-well plates (Corning Life Sciences, Tewksbury, MA, USA), at a confluency/suspension of 106 cells per well, respectively. The cells were rinsed with sterile phosphate-buffered saline (PBS) and fresh cell culture media added 2 to 4 hr prior to infecting the cells with bacteria. To infect the cells, E. coli O157:H7 from overnight culture was washed with phosphate buffered saline (PBS), resuspended in respective cell growth media, and added at a multiplicity of infection (MOI) of 20 per well. Following incubation for six hours, the cells were gently washed three times with sterile PBS to remove unadhered bacteria. Cells with adhered bacteria were collected using a sterile cell scraper and wells were rinsed with 1 mL sterile PBS to ensure collection of all components. Collected samples were centrifuged and pellets were stored in RNAlater RNA stabilization solution (Invitrogen, Waltham, MA, USA).
2.4. Isolation of Bacterial RNA from Infected Cells and mRNA Enrichment
Total RNA was extracted using RiboPure RNA Purification Kit (Invitrogen, Waltham, MA, USA). Bacterial RNA was separated from total RNA by depleting the mammalian RNA using MICROBEnrich Kit (Invitrogen, Waltham, MA, USA). Host depleted samples were further processed for rRNA depletion to enrich for bacterial mRNA using MICROBExpress Bacterial mRNA Enrichment Kit (Invitrogen, Waltham, MA, USA). All steps were carried out according to the manufacturer’s instructions. Quality and concentration of RNA samples in each step was measured using Qubit™ 4.0 Fluorometer (Thermo Fisher Scientific, Wilmington, DE, USA).
2.5. Illumina Library Preparation and RNA Sequencing
Library preparation and RNA sequencing were done on an Illumina platform through Genewiz (South Plainfield, NJ, USA). Briefly, the library preparation was done using NEBNext Ultra RNA Library Prep Kit (Illumina Inc, San Diego, CA, USA) as per manufacturer’s recommendations. Enriched mRNAs were fragmented, first-strand and second-strand cDNA were synthesized, and fragments were end-repaired, adenylated at 3’ ends, and ligated with universal adapters. The mRNA libraries were multiplexed and clustered on one lane of a flow cell for sequencing on HiSeq 2500 (Illumina) platform with 2x100 paired-end (PE) reads. E. coli O157 incubated for six hours in EMEM and DMEM-HG media only without cells, were used as controls.
2.6. Sequence Analysis and Detection of Differential Gene Expression
Quality checking, base trimming, read mapping, normalization, and differential gene expression analysis were performed using the CLC Genomics Workbench version 22.0.3 (QIAGEN Inc., Redwood City, CA, USA). Raw read sequences were imported into the CLC Genomics Workbench and aligned to E. coli O157:H7 strain EDL933 (NCBI RefSeq assembly: GCF_000006665.1) chromosome (NC_002655.2) as well as the associated plasmid pO157 (NC_007414.1). Gene expression was calculated using the RPKM (reads per kilobase of exon model per million mapped reads), and applying the equation, RPKM = number of gene reads/ (mapped reads (millions) x gene length (kb). False discovery rate (FDR) [24] was applied to identify statistically significant alterations in gene expression. To identify differential expression (DE) in two groups (treated vs control), TMM Normalization (Trimmed Mean of M values) described for Whole Transcriptome RNA-seq Technology was applied. Samples were submitted in triplicate, and the results were averaged to obtain fold changes (FC). Three gene filtering criteria were applied to differentially expressed data: FDR ≤ 0.05, FC ≥ 2.0, and p-value ≤ 0.01. Gene enrichment analysis was performed using ShinyGO 0.77 and iDEP 1.1 [25]. Heat maps were created using iDEP 1.1, applying the Euclidean distance with average linkage clustering.
2.7. Statistical Analysis, Software, and Data Preparation
All quantitative assays were performed in triplicate in independent experiments. Statistical analysis was performed with CLC Genomics Workbench, whereas the graphs were plotted using GraphPad Prism version 10.0.1.
2.8. Data Availability
The transcriptomic profile data (both raw and processed) described in this study were deposited in the Gene Expression Omnibus (GEO) database in NCBI, under the accession number GSE240423.
3. Results
3.1. Overall transcriptomics profiles of E. coli O157:H7 cultured with CCD CoN 841 and RAJ cells during initial adherence
To determine the differential gene expression of E. coli O157:H7 during initial adherence to the intestinal epithelia of bovine and human hosts, CCD CoN 841 and RAJ cells were infected with E. coli O157:H7 for six hours. Total RNA with RIN values between 8.7 and 9.5 were enriched for bacterial RNA and depleted of rRNA. The resultant bacterial mRNA was subjected to deep sequencing at 100 million reads coverage. The obtained transcript reads successfully mapped to the E. coli O157:H7 strain EDL933 genome. The percentage coverages of map reads were 87.7 for CCD CoN 841 /O157:H7 and 88.18 for RAJ/O157:H7. For the respective controls, the percent coverages were 80.53 and 83.18 for E. coli O157:H7 grown in EMEM and DMEM-HG, respectively. Diagnostic plots for read count data were generated for analyzing the variations in the mapped library sizes (Figure 1).
3.2. Overall Transcriptomics Differences of E. coli O157:H7 cultured with CCD CoN 841 and RAJ Cells during Initial Adherence
Transcriptome of E. coli O157:H7 was completely different in the human colon cell line compared to the bovine RAJ cells during initial adherence at six hours. Differential expression analysis was performed on CLC Genomics Expression Browser tool, and the results were provided in Supplementary File 1 (E. coli O157:H7 in CCD CoN 841 vs. EMEM media control) and Supplementary File 2 (E. coli O157:H7 in RAJ vs. DMEM-HG media control). The total number of expressed genes of E. coli O157:H7 in CCD CoN 841 cells and RAJ cells were 829 and 1,939, respectively. A total of 83 E. coli O157:H7 genes were uniquely expressed in CCD CoN 841 cells, whereas 1193 genes were uniquely expressed in RAJ cells. Another 746 genes were expressed commonly in both cell types (Figure 2). A total of 417 E. coli O157:H7 genes were upregulated and 412 were downregulated in CCD CoN 841 cells, whereas 1,024 genes were upregulated and 915 were downregulated in RAJ adhered E. coli O157:H7. Differential changes in transcriptomes are represented by volcano plots (Figure 2). Heatmap of expression profiles of all differentially expressed genes are depicted in Supplementary Figure S1. To investigate comparative analysis of transcriptomic shifts, the differentially expressed genes (DEGs) were filtered using criteria such as at least two-fold change, FDR ≤ 0.05 and P-value ≤ 0.01.
3.3. The Enriched Biological Pathways During the Initial Adherence of E. coli O157:H7 in CCD CoN 841 and RAJ cells
The gene enrichment analysis, which categorized the genes into their respective metabolic function, has shed light on the cellular and molecular processes that underlie interactions between E. coli O157:H7 and host cells. Genes involved in lipopolysaccharide biosynthesis, polysaccharide biosynthesis, lipid biosynthesis, and metal ion homeostasis were considerably elevated in E. coli O157:H7 adhered to CCD CoN 841 cells (p-value 0.01 and FDR 0.05). However, processes involved in the production of aromatic and organic compounds as well as heterocycles were suppressed. Whereas, E. coli O157:H7 attached to RAJ cells showed considerable downregulation in metabolic pathways involving antibiotics and secondary metabolites. Both treatments showed similar enrichment for all other cellular metabolic pathways (Figure 3).
3.4. Functional Annotation of E. coli O157:H7 Genes Differentially Expressed in Two Different Host Cell Types
The top 20 genes upregulated in each treatment group (i.e., two host cell types) with their fold expression, subcellular location, and functional category are listed in Table 1. Specifically, 10 E. coli O157:H7 genes implicated in iron acquisition, utilization, and survival were upregulated during initial adherence to human colonic epithelial cell line CCD CoN 841. However, all those genes were down regulated during adherence to bovine RAJ cells (Figure 4a). These included three genes involved in the iron transport locus chu (chuA, chuS, chuY), two genes in the shu heme utilization locus (shuU, shuT), enterobactin synthase multienzyme complex component genes entF, ybdZ involved in the enterobactin biosynthetic process, high-affinity ferric uptake transmembrane transporter fhuE, and the iron uptake and assimilation genes efeB and efeO.
This study unveiled the presence of seven virulence genes, namely espW (Type III secretion system effector), wzb (capsular polysaccharide biosynthesis and assembly), iraD (regulating flagellar synthesis), purC and purD (purine biosynthesis), and narG (nitrate reductase) expressed in E. coli O157:H7 on both cell types. The expression was much higher on bovine cells than on human cells. In contrast, the gene encoding superoxide dismutase, sodB, was upregulated in the bovine context and down regulated in human (Figure 4b).
We also noted 22 genes encoding outer membrane proteins (OMPs), out of which 15 consistently expressed at equal levels in E. coli O157:H7 grown with either bovine or human cells. In contrast, the remaining seven OMP-encoding genes, fhuE, yddB, cirA, ybdZ, eleB, yncE and asr, displayed substantial and notable upregulation, specifically in E. coli O157:H7 grown with human CCD CoN 841 cells (Figure 5a). Biofilm formation and other processes related to lipopolysaccharide metabolism were up-regulated in both treatment groups. These included (i) a group of genes associated with the biosynthesis of colanic acid and a component of the extracellular matrix (wcaA, wcaB, wcaC, wcaD, wcaI, wcaJ, wcaF, wcaL, wcaK, ugd, wzc), (ii) ugd, wcaC, and glmU involved in biosynthesis of UDP-N-acetylglucosamine (UDP-GlcNAc), (iii) lipA (biotin biosynthesis) and fabI (fatty acid beta-oxidation) (Figure 5b), and (iv) yjbG, yjbE, and yjbF involved in extracellular polysaccharide production during biofilm formation (Figure 5c).
The majority of genes related to cellular homeostasis exhibited upregulation in CCD CoN 841-adherent E. coli O157:H7, signifying a concerted effort to maintain equilibrium in the human host. For example, numerous genes associated with iron metabolism and homeostasis, including those encoding the iron transporter proteins (fhuA, fhuD, fhuC, fhuD, and fhuE) were upregulated as compared to E. coli O157:H7 grown with RAJ cells. Similarly, the genes participating in cellular response to DNA protection (fepB), colicin transport (fepA), cellular oxidant detoxification (efeB), and cell redox homeostasis (nrdH) were also upregulated (Figure 5d). Interestingly, we found a deliberate upregulation of genes involved in antibiotic resistance and secondary metabolic processes during E. coli O157:H7 adherence to human CCD841 cells. Specifically, genes linked to the enterobactin transporter and enterobactin synthase components (entA, entB, entF, entH, ybdZ, and ycbX) were activated (Figure 5e). During interaction with the bovine RAJ cells, E. coli O157:H7 genes associated with the glycolytic pathway (Figure 5f), and fimbriae/pilus assembly and regulation such as fimD, fimG, fimA_1, fimA_2, fimA_3, fimH_1, fimZ, fimC_1, fimF, and fimI (Figure 5g) were upregulated in comparison to E. coli O157:H7 adhered to CCD CoN 841. Likewise, 15 eut genes (eutS, eutE, eutQ, eutK, eutC, eutM, eutH, eutF, eutG, eutC, eutB, eutK, eutL, eutT, and eutH), which play pivotal roles in ethanolamine catabolism demonstrated significantly higher upregulation during initial attachment to bovine RAJ cells compared to the CCD CoN 841 cells. Additional four eut genes—namely, eutA, eutE, eutI, and eutP—were expressed solely in association with bovine RAJ cells.
Our comprehensive analysis, after applying the gene filtering criteria, revealed a total of 221 and 436 E. coli O157:H7 genes were upregulated during initial adherence with CCD CoN and RAJ cells, respectively. In addition to these commonly upregulated genes, a set of 22 genes exhibited unique expression with human cells while 155 genes displayed unique expression with bovine cells. A detailed list of noteworthy upregulated and uniquely expressed genes in E. coli O157:H7, during the initial adherence to CCD CoN 841 and RAJ cells, is included in Supplementary Table S1.
4. Discussion
The O157 and non-O157 STEC are prominent foodborne pathogens with global impact, known for inducing severe illnesses in human populations across the world [26,27,28]. STEC finds a significant reservoir in healthy cattle, which serves as an asymptomatic carrier. Initial attachment to intestinal cells is of pivotal importance as it marks a crucial early stage in the development of pathogenicity in humans or asymptomatic colonization of cattle [29,30]. Our research was aimed to enhance the understanding of transcriptional gene regulation occurring during the initial attachment of STEC O157:H7 to both human and bovine intestinal cells. The goal was to shed light onto the current knowledge of STEC asymptomatic colonization in the bovine host versus pathogenesis in the human host.
In the context of E. coli O157:H7 adherence to CCD CoN 841 cells, the enrichment of the lipopolysaccharide, polysaccharide, and lipid biosynthesis genes highlighted that E. coli O157:H7 might strategically manipulate its outer membrane composition to enhance adhesion and potentially establish a stable niche within the host environment within the first six hours of infection [31,32,33]. Furthermore, the enrichment of genes associated with metal ion homeostasis indicates the bacterium's adaptation to the host cell environment, possibly to ensure its survival and effective colonization [34,35]. While the upregulated genes were more noticeable during E. coli adherence to CCD CoN 841 cells, the downregulation of pathways related to antibiotic, drug, and secondary metabolism were particularly intriguing during their adherence to RAJ cells suggesting distinct mechanisms employed by E. coli O157:H7 during adaptation to the reservoir host and the human host. The reduced emphasis on secondary metabolic pathways might reflect the bacterium's shift in resource allocation toward more immediate requirements, such as adherence and colonization of RAJ to establish a long-term carrier state in the bovine host [36,37].
The findings from the differential expression analysis also provide valuable insights into the dynamic response of STEC O157:H7 during infection, illuminating its strategies for iron acquisition, utilization, and overall survival within distinct host environments. Notably, the observed differential regulation of 10 genes associated with iron-related processes presents a compelling glimpse into the bacterium's adaptive mechanisms, when interacting with different host cell types. In the context of attachment to human colonic epithelial cell line CCD CoN 841, the upregulation of these genes highlights the bacterium's heightened demand for iron. Iron is an essential nutrient for bacterial growth, and pathogens often employ sophisticated tactics to secure this critical resource from their host [38]. The substantial upregulation of genes involved in iron transport, heme utilization, enterobactin biosynthesis, and high-affinity ferric uptake underscores E. coli O157:H7's concerted efforts to scavenge and efficiently utilize iron from the human colonic environment. This strategic response could potentially contribute to the bacterium's ability to establish a foothold within the human host and establish conditions conducive to pathogenicity. The five virulence genes chuA, chuS, chuY, shuU, and shuT are particularly interesting because of the complex interplay between their products in relation to iron transport and heme utilization. The chuA gene, which encodes an outer membrane hemoglobin receptor protein crucial for heme uptake, stands out as a pivotal player in the bacterium's quest for iron. Its differential expression could play a key role in the overall iron acquisition strategy of E. coli O157:H7, enabling it to efficiently harvest heme-bound iron from the host environment [39]. This heightened activity of chuA within the human colon might contribute to the bacterium's virulence and ability to cause disease in humans. However, a converse pattern emerged in the bovine RAJ cell infection scenario, where the same set of genes displayed downregulation. This distinct regulatory behavior suggests a tailored approach of E. coli O157:H7, when confronted with the bovine host environment. The observed downregulation of these iron-related genes in bovine RAJ-infected E. coli O157:H7 implies a different iron usage landscape during initial hours in the bovine host, establishing its mutual existence, rather than damaging the host, as is the case within the human host. For example, in a chicken infection model, iron acquisition systems, such as salmochelin and aerobactin played a more significant role in virulence of ExPEC than heme utilization [40]. This might reflect the bacterium's adaptability to varying host contexts.
Infection of human CCD CoN 841 cells by E. coli O157:H7 prompted the upregulation of numerous genes associated with cellular homeostasis, particularly those linked to metal iron homeostasis. This included the significant elevation of iron transporter genes, along with genes involved in DNA protection, colicin transport, cellular oxidant detoxification, and cell redox homeostasis, suggesting a coordinated response to host cell interactions. The iron-related genetic response could reflect the bacterium's adaptation to the host's iron availability, a critical factor for its survival and growth [41,42]. This particularly intriguing observation emerged as we investigated the behavior of E. coli O157:H7 during its adhesion to CCD CoN 841 cells.
The investigation revealed a distinctive set of E. coli O157:H7 virulence genes exhibiting upregulation within the bovine RAJ environment, which might shed valuable insights into the pathogen's adaptation and response depending on the specific host environment. An area that needs further exploration is the upregulation of espW, which encodes a vital component of Type III secretion system (T3SS). The espW is predominantly located within the locus of enterocyte effacement (LEE) pathogenicity island, which allows for intimate attachment of typical STEC, like the E. coli strain we used in this study, to the intestinal epithelium causing characteristic attaching and effacing lesions during infection [43]. Previously, Kudva et al. [44] demonstrated that LEE-encoded proteins do not have a role in STEC adherence to squamous cells at the RAJ, although needed for effective adherence to the columnar cells of the RAJ. The EspW effector is responsible for triggering changes in the arrangement of actin filaments within human host cells [43]. However, the role of EspW in the colonization of E. coli O157:H7 in bovine RAJ cells has yet to be determined. Another gene overly expressed in the context of RAJ cells only was wzb, which is involved in the synthesis and assembly of capsular polysaccharides. Capsular polysaccharides play an important role in evading host immune responses and enhancing the pathogen's ability to colonize and persist within the host [45] and therefore might aid E. coli O157:H7 to establish an overt infection in the bovine host.
The outer membrane proteins (OMPs) facilitate various processes such as biofilm formation, infection of eukaryotic cells, antibiotic resistance, and modulation of immune responses [46]. Therefore, OMPs are essential players in the bacterium's adaptation to its environment and its interactions with the host [47].
The observed differential regulation of 15 OMP-encoding genes in bovine and human intestinal epithelium suggests the adaptability and plasticity of E. coli O157:H7 in tailoring its molecular strategies to suit the requirements of in both hosts during its initial attachment. Additionally, the seven OMP genes (fhuE, yddB, cirA, ybdZ, eleB, yncE, and asr) of E. coli O157:H7 upregulated in association with human CCD841 CoN 841 only. This implies a specialized role of these gene products in the human host environment, but not in the bovine host. Understanding the role of these OMPs might open new avenues for exploring potential targets for interventions and therapies aimed at mitigating the impact of E. coli O157:H7 infections in both animal and human populations. We also observed upregulation of a set of genes (wcaA, wcaB, wcaC, wcaD, wcaI, wcaJ, wcaF, wcaL, wcaK, ugd, wzc) associated with colanic acid biosynthesis in E. coli O157 adhered to both human and bovine cells. Colanic acid, a component of the extracellular matrix, plays a pivotal role in biofilm development, contributing to the structural integrity and stability of the biofilm matrix [48]. Along with biotin biosynthesis, lipolate synthesis and fatty acid beta-oxidation were also elevated in both infection scenarios. These metabolic pathways may provide the necessary energy and components for the biosynthesis of biofilm matrix components.
When the genes differentially expressed were examined closely, E. coli O157:H7 in the RAJ context exhibited substantial overexpression of three genes involved in extracellular polysaccharide production (yjbG, yjbE, and yjbF) during biofilm formation. This finding underscores the bacterium's strategic adaptation to the biofilm lifestyle within the specific bovine RAJ environment, perhaps facilitating its persistence as a colonizer. Extracellular polysaccharides are integral components of the biofilm matrix, contributing to cell-cell adhesion and overall biofilm structure [49]. Genes involved in biosynthesis of UDP-GlcNAc, a fundamental building block in lipopolysaccharide, also exhibited significant upregulation in E. coli O157:H7 attached to bovine RAJ cells. This observation suggests an enhanced metabolic activity related to lipopolysaccharide production, potentially contributing to the modification of the bacterial cell surface and interactions with the environment. Upregulation of genes encoding lipid metabolism, and extracellular polysaccharide production, provides a comprehensive view of the genetic responses underlying biofilm formation in E. coli O157:H7 within both treatment groups. These findings highlight the complex coordination of various metabolic pathways and genetic factors that contribute to the establishment and maintenance of biofilm structures [50,51]. The insights gained from this study pave the way for a deeper understanding of biofilm-related processes and their potential implications for E. coli O157:H7 pathogenicity and survival in diverse host environments.
Notably, E. coli genes associated with antibiotic and secondary metabolic processes, including the key components of the enterobactin transporter and enterobactin synthase system, exhibited deliberate upregulation during adherence to human colonic epithelial cells. The activation of these genes signifies a strategic genetic response to acquire nutrients from the host as adaptive tactics for survival and persistence within the human host environment during pathogenesis [52,53,54].
Another intriguing observation was the upregulation of genes that encode E. coli type-1 fimbriae during initial attachment of E. coli O157:H7 to bovine RAJ cells. Fimbriae and pili are structures present on the surface of bacteria that serve as essential factors for host adhesion, colonization, and infection (55). The fimbriae/pilus assembly genes, including fimD, fimG, and fimC, play pivotal roles in the assembly, structural integrity, and initiation of these adhesive structures. Additionally, fimH, fimZ, fimF, and fimI are regulatory genes that contribute to the coordination of fimbrial expression and the bacterial response to environmental cues [56,57,58]. The significant upregulation of these genes in bovine-adhered E. coli O157:H7 suggests a specialized response and adaptation to the bovine host environment. Bovines are recognized as a major reservoir for E. coli O157:H7, and this bacterium's ability to effectively adhere to the bovine intestinal epithelium is a critical factor for establishing colonization and potential transmission to humans [6]. The observed upregulation likely enhances the bacterium's capacity to bind to bovine cells, thereby increasing its likelihood of successful colonization. In fact, E. coli O157:H7 type-1 fimbrial genes have been reported to play role in the attachment of supershed E. coli O157:H7 to bovine RAJ squamous epithelial cells in a strain dependent manner [59]. Further studies could elucidate whether this enhanced adhesion contributes to increased colonization in bovines, potentially exploiting the new knowledge to mitigate STEC colonization of the bovine RAJ thus reducing human foodborne STEC disease.
The heightened expression of ethanolamine catabolic genes (eut) in E. coli O157:H7 during its initial adherence to bovine intestinal cells as compared to human colonic epithelial cells, also is noteworthy. It is known that ethanolamine plays a multifaceted role in the virulence of E. coli O157:H7 as a nutrient source, and signaling molecule, and influences gene expression contributing to its ability to cause disease and compete with other bacteria in the gut [60,61]. However, the paradox lies in the fact that while E. coli O157:H7 colonizes bovines, it does not cause overt disease in this context [6]. Therefore, our finding of eut gene expression in the context of bovine RAJ cells but not human CCD CoN 841 cells is somewhat contrary to previous reports. Future studies must be directed towards elucidating the role of ethanolamine in initial adherence of STEC to the bovine RAJ rather than causing disease in the human host.
In summary, our findings provide insights into the mechanisms employed by E. coli O157:H7 in human and bovine hosts, following the first six hours of infection. This study provides a foundation for further investigations into the host-pathogen interactions in two different host milieus: leading to disease in one host, whereas colonization and persistence does not result in disease in another host.
5. Conclusions
This study highlights the differences in STEC gene regulation during initial adherence to human CCD CoN 841 cells and bovine RAJ cells, revealing the bacterium's remarkable adaptability to diverse host microenvironments. Notably, our findings identified the differential regulation of virulence factors, metabolic pathways, and other cellular responses, underscoring versatile strategies used by STEC for colonization, survival, and potential pathogenesis. Particularly significant was the deliberate upregulation of genes associated with iron acquisition, metabolic pathways, and antibiotic resistance during adherence to human colonic epithelial cells. This is contrary to their significant down regulation in bovine RAJ cells, suggesting the bacterium's different molecular responses to different host cues. These results contribute to a deeper understanding of E. coli O157:H7's behavior within specific host contexts, as well as provide a foundation for future research aimed at deciphering its colonization and pathogenic mechanisms in different host niches and developing targeted interventions to mitigate its impact on both animal and human health. We have initiated additional research based on our findings, with the goal of reducing STEC colonization of bovine RAJ and shedding, as a preharvest intervention strategy to reduce STEC foodborne outbreaks.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Supplementary File S1: Differential expression analysis of E. coli O157:H7 in CCD CoN 841 vs. EMEM media control performed on CLC Genomics expression browser; Supplementary File S2: Differential expression analysis of E. coli O157:H7 in RAJ vs. DMEM-HG media control performed on CLC Genomics expression browser; Figure S1: Heatmap of expression profiles of all differentially expressed genes; Table S1: List of upregulated and uniquely expressed genes in E. coli O157:H7 during the initial adherence to both CCD CoN 841 and RAJ cells.
Author Contributions
Conceptualization, S.K; methodology, L.E.K., I.T.K. and S.K.; software, L.E.K. and S.K.; validation, L.E.K. and S.K.; formal analysis, L.E.K. and S.K.; investigation, L.E.K., I.T.K. and S.K.; resources, I.T.K. and S.K.; data curation, L.E.K. and S.K.; writing—original draft preparation, L.E.K. and S.K.; writing—review and editing, L.E.K., I.T.K. and S.K.; visualization, L.E.K. and S.K.; supervision, S.K.; project administration, S.K.; funding acquisition, S.K. All authors have read and agreed to the published version of the manuscript.
Funding
ITK was supported by the USDA-ARS CRIS project 5030-32000-225-00D.
Data Availability Statement
The transcriptomic profile (both raw and processed data) described in this study was deposited in Gene Expression Omnibus (GEO) database in NCBI, under the accession number GSE240423.
References
- Rangel, J. M.; Sparling, P. H.; Crowe, C.; Griffin, P. M.; Swerdlow, D. L. Epidemiology of Escherichia coli O157:H7 Outbreaks, United States, 1982–2002. Emerg. Infect. Dis. 2005, 11(4), 603–609. [Google Scholar] [CrossRef]
- Tarr, P. I.; Gordon, C. A.; Chandler, W. L. Shiga-Toxin-Producing Escherichia coli and Haemolytic Uraemic Syndrome. Lancet 2005, 365(9464), 1073–1086. [Google Scholar] [CrossRef]
- Karmali, M. A.; Steele, B. T.; Petric, M.; Lim, C. Sporadic Cases of Haemolytic-Uraemic Syndrome Associated with Faecal Cytotoxin and Cytotoxin-Producing Escherichia coli in Stools. Lancet 1983, 1(8325), 619–620. [Google Scholar] [CrossRef]
- Matthews, L.; Low, J. C.; Gally, D. L.; Pearce, M. C.; Mellor, D. J.; Heesterbeek, J. A.; Chase-Topping, M.; Naylor, S. W.; Shaw, D. J.; Reid, S. W.; Gunn, G. J.; Woolhouse, M. E. Heterogeneous Shedding of Escherichia coli O157 in Cattle and Its Implications for Control. Proc. Natl. Acad. Sci. U. S. A. 2006, 103(3), 547–552. [Google Scholar] [CrossRef]
- Friedrich, A. W.; Bielaszewska, M.; Zhang, W. L.; Pulz, M.; Kuczius, T.; Ammon, A.; Karch, H. Escherichia coli Harboring Shiga Toxin 2 Gene Variants: Frequency and Association with Clinical Symptoms. J. Infect. Dis. 2002, 185(1), 74–84. [Google Scholar] [CrossRef]
- Lim, J. Y.; Yoon, J.; Hovde, C. J. A Brief Overview of Escherichia coli O157:H7 and Its Plasmid O157. J. Microbiol. Biotechnol. 20(1), 5–14. [CrossRef]
- Bosilevac, J. M.; Guerini, M. N.; Kalchayanand, N. , et al. Prevalence and Characterization of Non-O157 Shiga Toxin-Producing Escherichia coli Isolates from Commercial Ground Beef in the United States. Appl. Environ. Microbiol. 2007, 73 (18), 6365–6371.
- Mir, R. A.; Schaut, R. G.; Looft, T.; Allen, H. K.; Sharma, V. K.; Kudva, I. T. Recto-Anal Junction (RAJ) and Fecal Microbiomes of Cattle Experimentally Challenged with Escherichia coli O157:H7. Front. Microbiol. 11, 693. [CrossRef]
- Naylor, S. W.; Low, J. C.; Besser, T. E.; Mahajan, A.; Gunn, G. J.; Pearce, M. C.; McKendrick, I. J.; Smith, D. G.; Gally, D. L. Lymphoid Follicle-Dense Mucosa at the Terminal Rectum Is the Principal Site of Colonization of Enterohemorrhagic Escherichia Coli O157:H7 in the Bovine Host. Infect. Immun. 2003, 71(3), 1505–1512. [Google Scholar] [CrossRef]
- Hancock, D. D.; Besser, T. E.; Rice, D. H.; Ebel, E. D.; Herriott, D. E.; Carpenter, L. V. Multiple Sources of Escherichia coli O157 in Feedlots and Dairy Farms in the Northwestern USA. Prev. Vet. Med. 1998, 35(1), 11–19. [Google Scholar] [CrossRef]
- Hancock, D. D.; Besser, T. E.; Kinsel, M. L. , et al. The Prevalence of Escherichia coli O157. H7 in Dairy and Beef Cattle in Washington State. Epidemiol. Infect. 2001, 127(2), 293–302. [Google Scholar]
- Garmendia, J.; Frankel, G.; Crepin, V. F. Enteropathogenic and Enterohemorrhagic Escherichia Coli Infections: Translocation, Translocation, Translocation. Infect. Immun. 2005, 73(5), 2573–2585. [Google Scholar] [CrossRef]
- Lawley, T. D.; Clare, S.; Walker, A. W. , et al. Targeted Restoration of the Intestinal Microbiota with a Simple, Defined Bacteriotherapy Resolves Relapsing Clostridium difficile Disease in Mice. PLOS Pathog. 2013, 9 (10), e1002995.
- Govindarajan, D. K.; Viswalingam, N.; Meganathan, Y.; Kandaswamy, K. Adherence Patterns of Escherichia coli in the Intestine and Its Role in Pathogenesis. Medicine in Microecology 2020, 5. [Google Scholar] [CrossRef]
- Pakbin, B.; Brück, W. M.; Rossen, J. W. A. Virulence Factors of Enteric Pathogenic Escherichia coli: A Review. Int. J. Mol. Sci. 2021 , 22 (18), 9922. 14 September. [CrossRef]
- Nguyen, Y.; Sperandio, V. Enterohemorrhagic E. coli (EHEC) Pathogenesis. Front. Cell. Infect. Microbiol. 2012 , 2, 90. 12 July. [CrossRef]
- Lai, Y.; Rosenshine, I.; Leong, J. M.; Frankel, G. Intimate Host Attachment: Enteropathogenic and Enterohaemorrhagic Escherichia Coli. Cell. Microbiol. 15(11), 1796–1808. [CrossRef]
- Galán, J. E.; Wolf-Watz, H. Protein Delivery into Eukaryotic Cells by Type III Secretion Machines. Nature 2006, 444(7119), 567–573. [Google Scholar] [CrossRef]
- Fedorchuk, C.; Kudva, I. T.; Kariyawasam, S. The Escherichia coli O157:H7 Carbon Starvation-Inducible Lipoprotein Slp Contributes to Initial Adherence In Vitro via the Human Polymeric Immunoglobulin Receptor. PLOS ONE 14(6), e0216791. [CrossRef]
- Barh, D.; Kumar, A.; Tiwari, S. , et al. A Novel Comparative Genomics Analysis for Common Drug and Vaccine Targets in Corynebacterium pseudotuberculosis and Other CMN Group of Human Pathogens. Chem. Biol. Drug Des. 2013, 81(4), 60–69. [Google Scholar]
- Grin, I.; Linke, D.; Schütz, M. S. Controlling the Message: A Structural Analysis of Protein Sorting to the TpsB/Omp85 Superfamily. Curr. Opin. Struct. Biol. 2014, 29, 1–8. [Google Scholar]
- Kudva, I. T.; Dean-Nystrom, E. A. Bovine Recto-Anal Junction Squamous Epithelial (RSE) Cell Adhesion Assay for Studying Escherichia coli O157 Adherence. J. Appl. Microbiol. 2011, 111(5), 1283–1294. [Google Scholar] [CrossRef]
- Kudva, IT. In vitro adherence patterns of Shigella serogroups to bovine recto-anal junction squamous epithelial (RSE) cells are similar to those of Escherichia coli O157. Foodborne Pathog Dis. 2012;9(4):346-351. [CrossRef]
- Pawitan, Y.; Michiels, S.; Koscielny, S.; Gusnanto, A.; Ploner, A. False Discovery Rate, Sensitivity and Sample Size for Microarray Studies. Bioinformatics 21(13), 3017–3024. [CrossRef]
- Ge, S. X.; Son, E. W.; Yao, R. iDEP: An Integrated Web Application for Differential Expression and Pathway Analysis of RNA-Seq Data. B.M.C. Bioinformatics 2018, 19(1), 534. [Google Scholar] [CrossRef]
- Blankenship, H. M.; Mosci, R. E.; Dietrich, S.; Burgess, E.; Wholehan, J.; McWilliams, K.; Pietrzen, K.; Benko, S.; Gatesy, T.; Rudrik, J. T.; Soehnlen, M.; Manning, S. D. Population Structure and Genetic Diversity of Non-O157 Shiga Toxin-Producing Escherichia coli (STEC) Clinical Isolates from Michigan. Sci. Rep. 2021, 11(1), 4461. [Google Scholar] [CrossRef] [PubMed]
- Majowicz, S. E.; Scallan, E.; Jones-Bitton, A.; Sargeant, J. M.; Stapleton, J.; Angulo, F. J.; Yeung, D. H.; Kirk, M. D. Global Incidence of Human Shiga Toxin-Producing Escherichia coli Infections and Deaths: A Systematic Review and Knowledge Synthesis. Foodborne Pathog. Dis. 11(6), 447–455. [CrossRef] [PubMed]
- Hedican, E. B.; Medus, C.; Besser, J. M.; Juni, B. A.; Koziol, B.; Taylor, C.; Smith, K. E. Characteristics of O157 Versus Non-O157 Shiga Toxin-Producing Escherichia coli Infections in Minnesota, 2000–2006. Clin. Infect. Dis. 2009, 49(3), 358–364. [Google Scholar] [CrossRef]
- Adegbola, R. A. Bacterial Adhesion and Pathogenicity. Afr. J. Med. Med. Sci. 1988, 17(2), 63–69. [Google Scholar] [PubMed]
- Rahal, E. A.; Kazzi, N.; Nassar, F. J.; Matar, G. M. Escherichia coli O157:H7—Clinical Aspects and Novel Treatment Approaches. Front. Cell. Infect. Microbiol. 2012, 2, 138. [Google Scholar] [CrossRef]
- Paquin-Veillette, J.; Dozois, C. M.; Harel, J. The ecological habitat and transmission of Escherichia coli O157:H7. Hekabab SM. F.E.M.S. Microbiol. Lett. 2013, 341(1), 1–12. [Google Scholar] [CrossRef]
- Segura, A.; Bertin, Y.; Durand, A.; Benbakkar, M.; Forano, E. Transcriptional Analysis Reveals Specific Niche Factors and Response to Environmental Stresses of Enterohemorrhagic Escherichia Coli O157:H7 in Bovine Digestive Contents. B.M.C. Microbiol. 2021, 21(1), 284. [Google Scholar] [CrossRef]
- Dresen, M.; Valentin-Weigand, P.; Berhanu Weldearegay, Y. Role of Metabolic Adaptation of Streptococcus suis to Host Niches in Bacterial Fitness and Virulence. Pathogens 2023, 12, 541. [Google Scholar] [CrossRef]
- Torres, A. G.; Payne, S. M. Haem Iron-Transport System in Enterohaemorrhagic Escherichia Coli O157:H7. Mol. Microbiol. 1997, 23(4), 825–833. [Google Scholar] [CrossRef]
- Sheldon, J. R.; Laakso, H. A.; Heinrichs, D. E. Iron Acquisition Strategies of Bacterial Pathogens. Microbiol. Spectr. 2016 April, 4 (2). [CrossRef]
- Seyedsayamdost, M. R. Toward a Global Picture of Bacterial Secondary Metabolism. J. Ind. Microbiol. Biotechnol. 2019 March, 46 (3–4), 301–311. [CrossRef]
- Baral, B.; Akhgari, A.; Metsä-Ketelä, M. Activation of Microbial Secondary Metabolic Pathways: Avenues and Challenges. Synth. Syst. Biotechnol. 2018 , 3 (3), 163–178. 12 September. [CrossRef]
- Hammer, N. D.; Skaar, E. P. Molecular Mechanisms of Staphylococcus aureus Iron Acquisition. Annu. Rev. Microbiol. 2011, 65, 129–147. [Google Scholar] [CrossRef]
- Nagy, G.; Dobrindt, U.; Kupfer, M.; Emödy, L.; Karch, H.; Hacker, J. Expression of Hemin Receptor Molecule ChuA Is Influenced by RfaH in Uropathogenic Escherichia coli strain 536. Infect. Immun. 69(3), 1924–1928. [CrossRef] [PubMed]
- Gao, Q.; Wang, X.; Xu, H.; Xu, Y.; Ling, J.; Zhang, D.; Gao, S.; Liu, X. Roles of Iron Acquisition Systems in Virulence of Extraintestinal Pathogenic Escherichia coli: Salmochelin and Aerobactin Contribute More to Virulence than Heme in a Chicken Infection Model. B.M.C. Microbiol. 2012, 12, 143. [Google Scholar] [CrossRef]
- Chandrangsu, P.; Rensing, C.; Helmann, J. D. Metal Homeostasis and Resistance in Bacteria. Nat. Rev. Microbiol. 15(6), 338–350. [CrossRef] [PubMed]
- Burcham, L. R.; Hill, R. A.; Caulkins, R. C.; Emerson, J. P.; Nanduri, B.; Rosch, J. W.; Fitzkee, N. C.; Thornton, J. A. Streptococcus pneumoniae Metal Homeostasis Alters Cellular Metabolism. Metallomics 2020 , 12 (9), 1416–1427. 23 September. [CrossRef]
- Sandu, P.; Crepin, V. F.; Drechsler, H.; McAinsh, A. D.; Frankel, G.; Berger, C. N. The Enterohemorrhagic Escherichia Coli Effector EspW Triggers Actin Remodeling in a Rac1-Dependent Manner. Infect. Immun. 85(9), e00244–17. [CrossRef] [PubMed]
- Kudva, I. T.; Hovde, C. J.; John, M. Adherence of Non-O157 Shiga Toxin-Producing Escherichia coli to Bovine Recto-Anal Junction Squamous Epithelial Cells Appears to Be Mediated by Mechanisms Distinct from Those Used by O157. Foodborne Pathog. Dis. 10(4), 375–381. [CrossRef]
- Hagelueken, G.; Huang, H.; Mainprize, I. L.; Whitfield, C.; Naismith, J. H. Crystal Structures of Wzb of Escherichia coli and CpsB of Streptococcus pneumoniae, Representatives of Two Families of Tyrosine Phosphatases That Regulate Capsule Assembly. J. Mol. Biol. 2009 , 392 (3), 678–688. 25 September. [CrossRef]
- Nie, D.; Hu, Y.; Chen, Z.; Li, M.; Hou, Z.; Luo, X.; Mao, X.; Xue, X. Outer Membrane Protein A (OmpA) as a Potential Therapeutic Target for Acinetobacter baumannii Infection. J. Biomed. Sci. 2020, 27(1), 26. [Google Scholar] [CrossRef]
- Lin, J.; Huang, S.; Zhang, Q. Outer Membrane Proteins: Key Players for Bacterial Adaptation in Host Niches. Microbes Infect. 4(3), 325–331. [CrossRef]
- Balducci, E.; Papi, F.; Capialbi, D. E.; Del Bino, L. Polysaccharides’ Structures and Functions in Biofilm Architecture of Antimicrobial-Resistant (AMR) Pathogens. Int. J. Mol. Sci. 2023, 24(4), 4030. [Google Scholar] [CrossRef]
- Limoli, D. H.; Jones, C. J.; Wozniak, D. J. Bacterial Extracellular Polysaccharides in Biofilm Formation and Function. Microbiol. Spectr. 2015 June, 3 (3). [CrossRef]
- Davey, M. E.; O’Toole, G. A. Microbial Biofilms: From Ecology to Molecular Genetics. Microbiol. Mol. Biol. Rev. 64(4), 847–867. [CrossRef]
- Wang, R.; Bono, J. L.; Kalchayanand, N.; Shackelford, S.; Harhay, D. M. Biofilm Formation by Shiga Toxin-Producing Escherichia coli O157:H7 and Non-O157 Strains and Their Tolerance to Sanitizers Commonly Used in the Food Processing Environment. J. Food Prot. 2012, 75(8), 1418–1428. [Google Scholar] [CrossRef]
- Raymond, K. N.; Dertz, E. A.; Kim, S. S. Enterobactin: An Archetype for Microbial Iron Transport. Proc. Natl. Acad. Sci. U. S. A. 100(7), 3584–3588. [CrossRef] [PubMed]
- Saha, P.; Xiao, X.; Yeoh, B. S.; Chen, Q.; Katkere, B.; Kirimanjeswara, G. S.; Vijay-Kumar, M. The Bacterial Siderophore Enterobactin Confers Survival Advantage to Salmonella in Macrophages. Gut Microbes 2019, 10(3), 412–423. [Google Scholar] [CrossRef] [PubMed]
- Cassat, J. E.; Skaar, E. P. Iron in Infection and Immunity. Cell Host Microbe 13(5), 509–519. [CrossRef] [PubMed]
- Mandlik, A.; Swierczynski, A.; Das, A.; Ton-That, H. Pili in Gram-Positive Bacteria: Assembly, Involvement in Colonization and Biofilm Development. Trends Microbiol. 16(1), 33–40. [CrossRef]
- Zeiner, S. A.; Dwyer, B. E.; Clegg, S. FimA, FimF, and FimH Are Necessary for Assembly of Type 1 Fimbriae on Salmonella enterica serovar Typhimurium. Infect. Immun. 80(9), 3289–3296. [CrossRef]
- Gahlot, D. K.; Taheri, N.; MacIntyre, S. Diversity in Genetic Regulation of Bacterial Fimbriae Assembled by the Chaperone Usher Pathway. Int. J. Mol. Sci. 24(1), 161. [CrossRef]
- Schwan, W. R. Regulation of fim Genes in Uropathogenic Escherichia coli. World J. Clin. Infect. Dis. 1(1), 17–25. [CrossRef]
- Katani R, Kudva, IT, Srinivasan S, Stasko JB, Schilling M, Li L, Cote R, DebRoy C, Arthur TM, Sokurenko EV, and Kapur V. Strain and host-cell type dependent role of type 1 fimbriae genes in the adherence phenotype of super-shedder strains of Escherichia coli O157:H7. Int. J. Med. Micro. 2021. [CrossRef]
- Kendall, M.M. , Gruber, C., Parker, C., & Sperandio, V. (2012). Ethanolamine Controls Expression of Genes Encoding Components Involved in Interkingdom Signaling and Virulence in Enterohemorrhagic Escherichia coli O157:H7. mBio, 3.
- Rowley CA, Anderson CJ, Kendall MM. Ethanolamine Influences Human Commensal Escherichia coli Growth, Gene Expression, and Competition with Enterohemorrhagic E. coli O157:H7. mBio. 2018 Oct 2;9(5):e01429-18. [CrossRef]
Figure 1.
Diagnostics plots depicting the sequencing depth bias in each treatment. (A and D) Total raw read counts per each library. (B and E) Density plot of regularized log transformed total read counts. (C and F) Differentiation between treated and control samples is readily apparent in the principal component analysis (PCA), which accounts for 88.6 and 91.26% of the observed variation in CDD CoN 841and RAJ infected E. coli O157:H7, respectively, when compared to untreated samples. Total read counts are significantly different among sample groups (p= 1.16e-04) based on ANOVA.
Figure 1.
Diagnostics plots depicting the sequencing depth bias in each treatment. (A and D) Total raw read counts per each library. (B and E) Density plot of regularized log transformed total read counts. (C and F) Differentiation between treated and control samples is readily apparent in the principal component analysis (PCA), which accounts for 88.6 and 91.26% of the observed variation in CDD CoN 841and RAJ infected E. coli O157:H7, respectively, when compared to untreated samples. Total read counts are significantly different among sample groups (p= 1.16e-04) based on ANOVA.
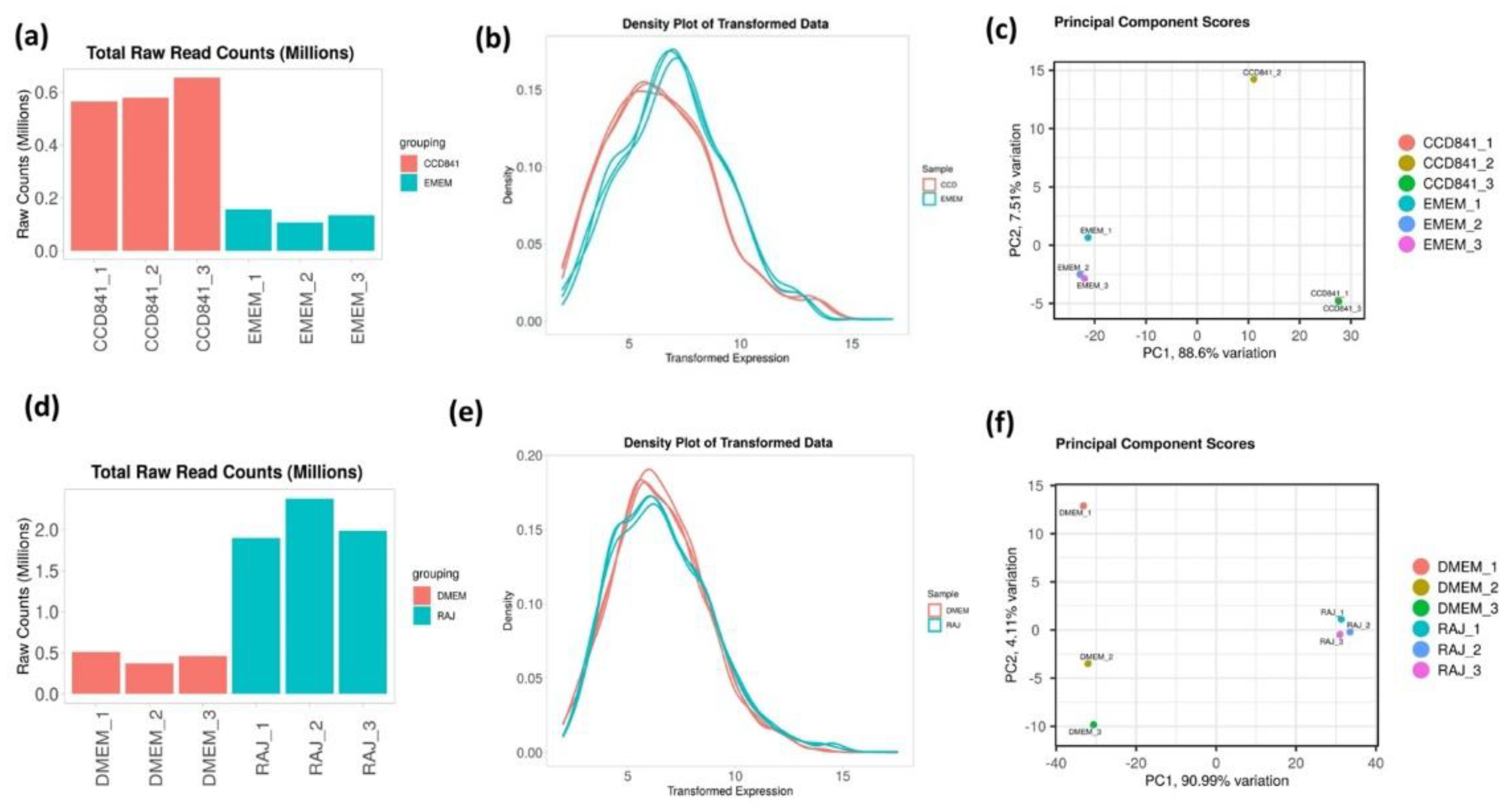
Figure 2.
Differential transcriptome profiles of E. coli O157:H7 in CCD CoN 841 and RAJ cells (total transcriptomic data without filtration). (A) Venn diagram showing unique and differentially expressed genes. Volcano plots showing differentially expressed E. coli O157:H7 genes in CCD CoN 841 cells (B) and RAJ cells (C). In volcano plots, the relationship between p-values and the magnitude of the difference in expression values were noted. X-axis indicates log2 fold change threshold and Y-axis depicts -log10 (p-values).
Figure 2.
Differential transcriptome profiles of E. coli O157:H7 in CCD CoN 841 and RAJ cells (total transcriptomic data without filtration). (A) Venn diagram showing unique and differentially expressed genes. Volcano plots showing differentially expressed E. coli O157:H7 genes in CCD CoN 841 cells (B) and RAJ cells (C). In volcano plots, the relationship between p-values and the magnitude of the difference in expression values were noted. X-axis indicates log2 fold change threshold and Y-axis depicts -log10 (p-values).
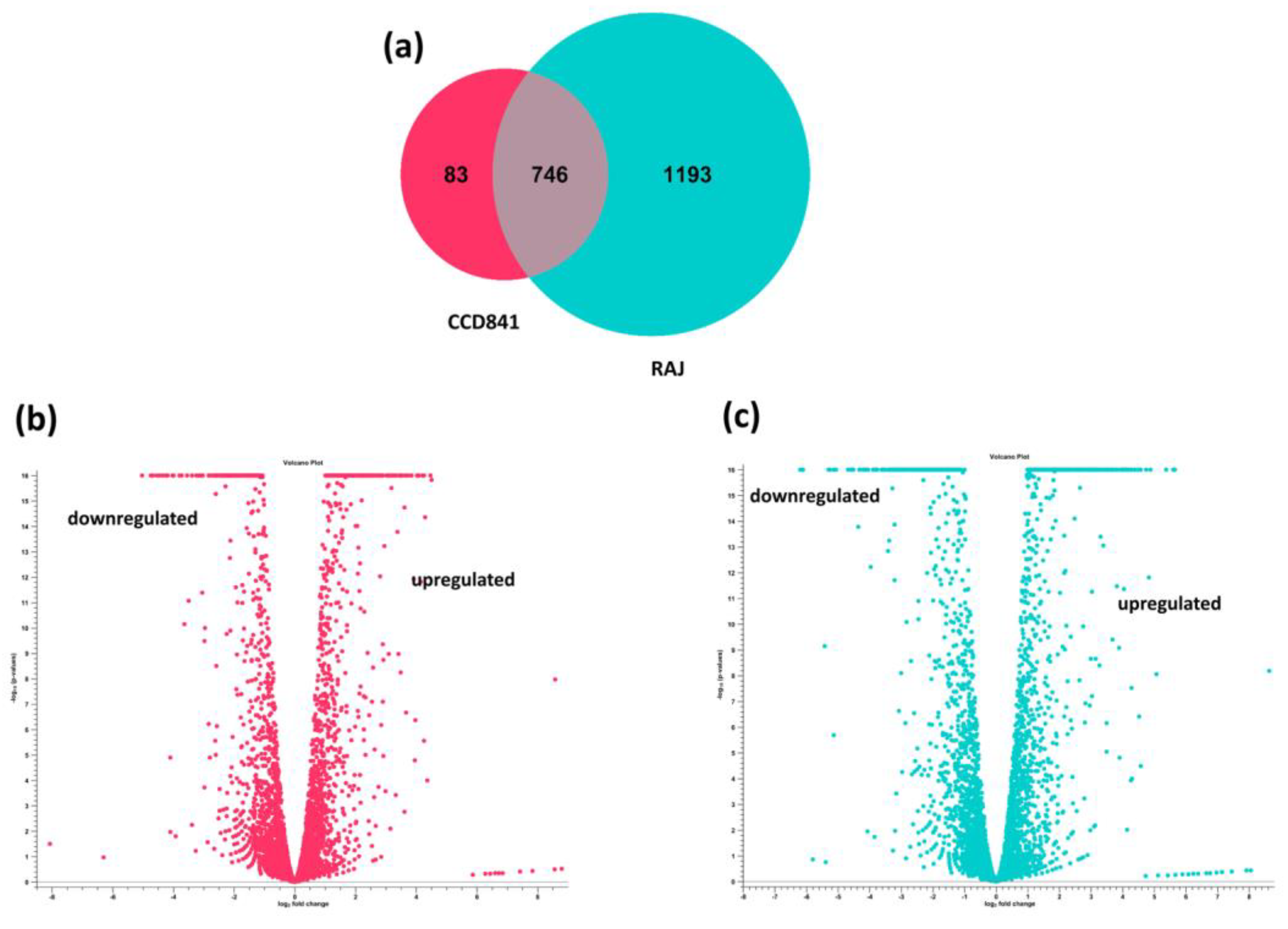
Figure 3.
Enriched functional categories of differentially expressed genes. (A) E. coli O157:H7 attached to CCD CoN 841 cells. (B) E. coli O157:H7 attached to RAJ cells. The x-axis is on log10 (p-value) scale. BP denotes ‘biological process’.
Figure 3.
Enriched functional categories of differentially expressed genes. (A) E. coli O157:H7 attached to CCD CoN 841 cells. (B) E. coli O157:H7 attached to RAJ cells. The x-axis is on log10 (p-value) scale. BP denotes ‘biological process’.
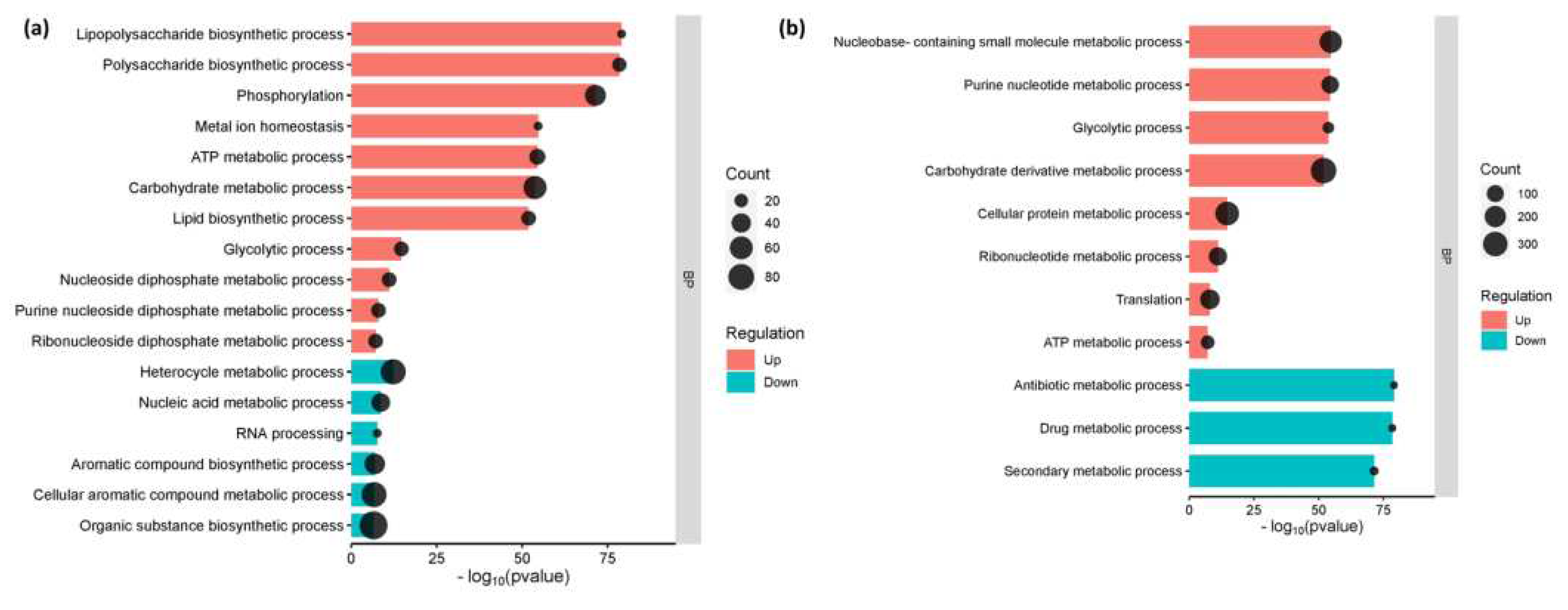
Figure 4.
Expression patterns of differentially expressed genes. (A) Upregulated virulence genes in E. coli O157:H7 adhered to CCD CoN 841. (B) Highly expressed virulence genes in E. coli O157:H7 during RAJ adhesion. Virulence genes were identified in a search with the virulence factor database (VFDB) (http://www.mgc.ac.cn/VFs/main.htm). Gene expression data represent the mean of three replicates ± standard deviation. Please note that some house-keeping genes were also included in the VFDB.
Figure 4.
Expression patterns of differentially expressed genes. (A) Upregulated virulence genes in E. coli O157:H7 adhered to CCD CoN 841. (B) Highly expressed virulence genes in E. coli O157:H7 during RAJ adhesion. Virulence genes were identified in a search with the virulence factor database (VFDB) (http://www.mgc.ac.cn/VFs/main.htm). Gene expression data represent the mean of three replicates ± standard deviation. Please note that some house-keeping genes were also included in the VFDB.
Figure 5.
Heat maps of transcriptional responses of E. coli O157:H7, during the initial adherence of human CCD CoN 841 cells and bovine RAJ cells based on functional analysis. (A) Outer membrane proteins. (B) Lipopolysaccharide metabolic processes. (C) Polysaccharide biosynthesis. (D) Cellular homeostasis. (E) Antibiotic metabolic processes and secondary metabolism. (F) Glycolytic pathway. (G) Cell adhesion or fimbriae/pilus assembly and regulation. Transformed FPKM numbers (normalized read counts) are represented by colors, with green indicating lower expression and red indicating higher expression.
Figure 5.
Heat maps of transcriptional responses of E. coli O157:H7, during the initial adherence of human CCD CoN 841 cells and bovine RAJ cells based on functional analysis. (A) Outer membrane proteins. (B) Lipopolysaccharide metabolic processes. (C) Polysaccharide biosynthesis. (D) Cellular homeostasis. (E) Antibiotic metabolic processes and secondary metabolism. (F) Glycolytic pathway. (G) Cell adhesion or fimbriae/pilus assembly and regulation. Transformed FPKM numbers (normalized read counts) are represented by colors, with green indicating lower expression and red indicating higher expression.
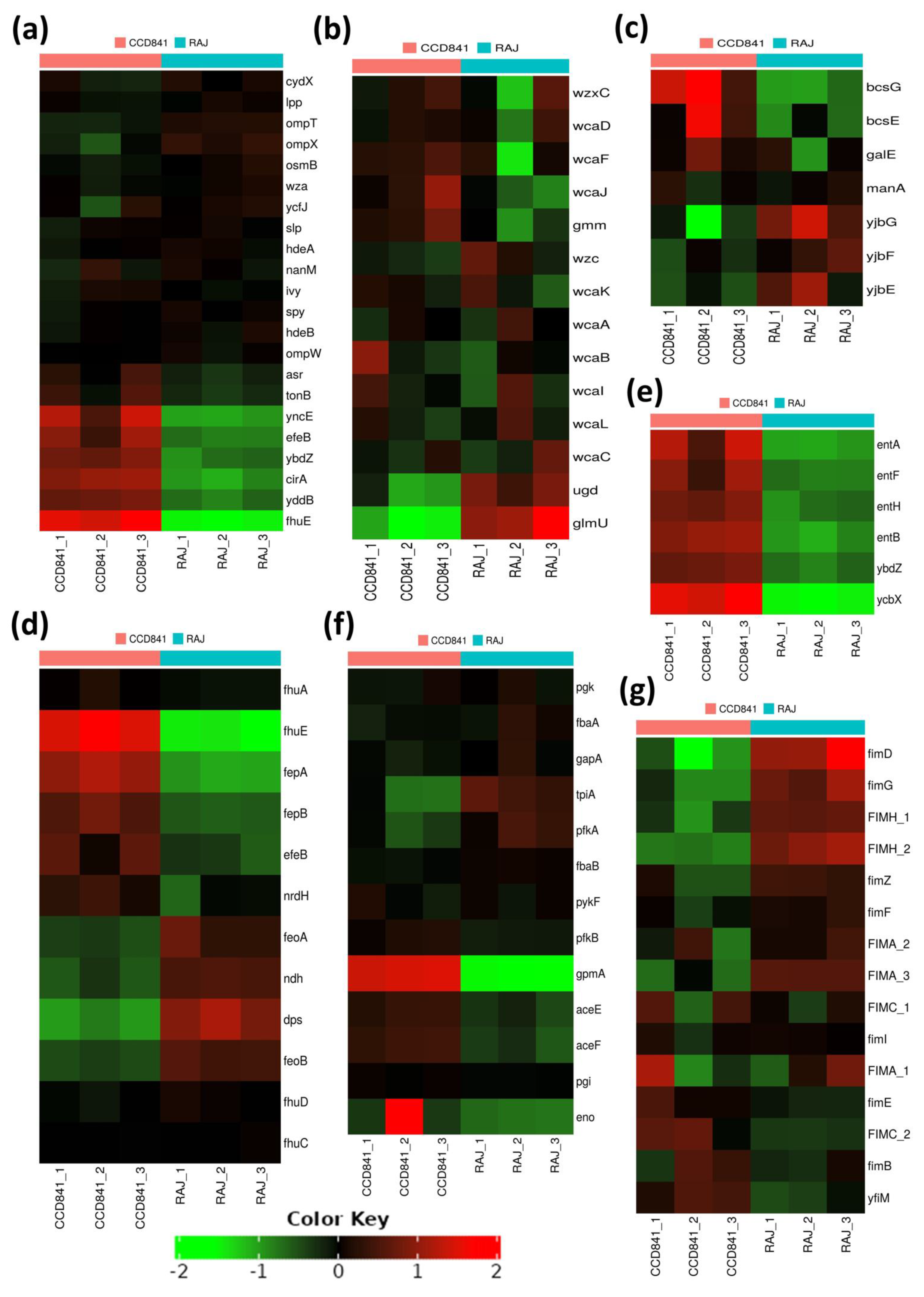

Table 1.
List of top 20 genes upregulated in E. coli O157:H7 during initial adherence to human CCD CoN 841 cells or bovine RAJ cells.
Table 1.
List of top 20 genes upregulated in E. coli O157:H7 during initial adherence to human CCD CoN 841 cells or bovine RAJ cells.
| E. coli O157:H7 adhering to CCD CoN 841 cells | ||||
|---|---|---|---|---|
| Symbol | Log2 FC | Location | Type | Gene Name |
| ndhF | 4.224748 | membrane | dehydrogenase | Type II NADH:quinone oxidoreductase |
| ynfM | 3.996075 | membrane | transporter | Inner membrane transport protein YnfM |
| yagU | 3.99386 | membrane | unknown | Inner membrane protein YagU |
| wcaJ | 3.745511 | membrane | transferase | UDP-glucose: undecaprenyl-phosphate glucose-1-phosphate transferase |
| fhuE | 3.649405 | cell outer membrane | Iron transporter | Hydroxamate siderophore receptor FhuE |
| yjbF | 3.632319 | membrane | Unknown | Uncharacterized lipoprotein YjbF |
| wza | 3.417282 | cell outer membrane | transporter | Putative polysaccharide export protein Wza |
| efeO | 3.411274 | periplasmic space | transporter | Iron uptake system component EfeO |
| fdhF | 3.283415 | cytosol | dehydrogenase | Formate dehydrogenase H |
| feoB | 3.260966 | membrane | transporter | Fe(2+) transporter FeoB |
| wcaD | 3.235622 | membrane | Unknown | Putative colanic acid polymerase |
| slp | 3.206504 | cell outer membrane | Unknown | Outer membrane protein Slp |
| wzc | 3.199261 | membrane | Kinase | Tyrosine-protein kinase wzc |
| adhE | 3.186165 | cytosol | acetaldehyde dehydrogenase | Bifunctional aldehyde-alcohol dehydrogenase AdhE |
| yohK | 3.087307 | membrane | unknown | Inner membrane protein YohK |
| pflB | 3.080942 | cytoplasm | transferase | Formate acetyltransferase 1 |
| hycC | 3.043606 | membrane | dehydrogenase | Formate hydrogenlyase subunit 3 |
| efeB | 3.017162 | periplasmic space | ferrochelatase | Deferrochelatase |
| glnK | 2.907097 | membrane | enzyme regulator | Nitrogen regulatory protein GlnK |
| amtB | 2.852313 | membrane | transporter | Ammonium transporter AmtB |
| E. coli O157:H7 adhering to RAJ cells | ||||
| yohJ | 4.832419 | membrane | unknown | UPF0299 membrane protein YohJ |
| fdhF | 4.422977 | cytosol | dehydrogenase | Formate dehydrogenase H |
| yagU | 4.28355 | membrane | Unknown | Inner membrane protein YagU |
| hycC | 4.249511 | membrane | dehydrogenase | Formate hydrogenlyase subunit 3 |
| yohK | 4.207073 | membrane | unknown | Inner membrane protein YohK |
| ynfM | 4.193856 | membrane | transporter | Inner membrane transport protein YnfM |
| pflB | 4.165908 | cytoplasm | transferase activity | Formate acetyltransferase 1 |
| yjbF | 4.156806 | membrane | unknown | Uncharacterized lipoprotein YjbF |
| hycD | 3.936897 | membrane | unknown | Formate hydrogenlyase subunit 4 |
| wza | 3.806629 | cell outer membrane | transporter | Putative polysaccharide export protein Wza |
| ndh | 3.8042 | membrane | dehydrogenase | Type II NADH:quinone oxidoreductase |
| btsT | 3.674308 | membrane | transporter | Pyruvate/proton symporter BtsT |
| adiC | 3.625606 | membrane | transporter | Arginine/agmatine antiporter |
| wcaJ | 3.532739 | membrane | transferase | UDP-glucose:undecaprenyl-phosphate glucose-1-phosphate transferase |
| hycF | 3.352964 | membrane | dehydrogenase | Formate hydrogenlyase subunit 6 |
| tpiA | 3.309538 | cytoplasm | isomerase | Triosephosphate isomerase |
| gapA | 3.309281 | cytoplasm | dehydrogenase | Glyceraldehyde-3-phosphate dehydrogenase A |
| wzc | 3.280554 | membrane | kinase | Tyrosine-protein kinase wzc |
| ynfF | 3.279045 | periplasmic space | reductase | Probable dimethyl sulfoxide reductase chain YnfF |
| ptsG | 3.210761 | membrane | transporter | PTS system glucose-specific EIICB component |
| slp | 2.011981 | cell outer membrane | unknown | Outer membrane protein Slp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



