
Submitted:
16 September 2023
Posted:
18 September 2023
You are already at the latest version
Abstract
Mucin glycans are important component of the mucus barrier and vital defence against physical and chemical damage as well as pathogens. Abnormal expression of mucin glycans can lead to disease, especially cancer. Here, we first summarize the main types of glycosylation on mucins and the mechanisms by which abnormal mucin glycans occur. We next describe the role of ab-normal mucin glycans in cancer. Finally, we describe MUC1-based antibodies, vaccines, radio-pharmaceuticals, and CAR-T therapies using the best characterized MUC1 as an example.
Keywords:
mucin
; glycosylation
; cancer
; MUC1
; targeted therapy
1. Introduction
Mucins are a class of high molecular weight glycoproteins with molecular weights typically ranging from 0.2 to 10 million Dalton[1], which are mainly synthesized by goblet cells and combines with inorganic salts suspended in water to form mucus. They cover the surfaces of the respiratory, digestive, gastrointestinal and genitourinary tracts, protecting epithelial cells from infection, dehydration, and physical or chemical damage, providing protection and lubrication for the epithelial surface. They are widely expressed in the body and are associated with many physiological and pathological processes[2]. So far, there are 20 mucins in human body have been discovered, including the secreted mucins MUC2, MUC5AC, MUC5B, MUC6, MUC19, MUC7, and MUC9, of which MUC2 is the main form of secreted mucin[3,4], as well as transmembrane mucins including MUC1, MUC3A , MUC3B, MUC4, MUC11, MUC12, MUC13, MUC15, MUC16, MUC17, MUC20, MUC21 and MUC22[5]. In addition, there are some proteins that belong to atypical or mucin-like protein molecules, atypical mucins including MUC10, MUC14, and MUC18. Mucin-like protein molecules are exist in parasites, viruses and fungi, Herpes virus has a mucin region, and Toxoplasma Gondii have mucin-like structural domains, and Candida albicans has mucin-like proteins. Similar to the structure and function of mucins, mucin-like protein molecules have structural domains rich in Pro, Thr and Ser, which can undergo extensive O-glycosylation and also act as barriers to protect cell membranes[6,7]. For example, the Msb2 glycoprotein of Candida albicans produces a mucosal layer to protect cells. This protein is considered a functional analogue of mammalian MUC1/MUC2[8]. Transmembrane mucins have a C-terminal cytoplasmic tail, a transmembrane region and an extracellular portion. Characterized by the SEA (sea urchin spermatoglycan, enterokinase and agrin) structural domain and the VWD (vascular haemophilic factor D) structural domain. Unlike transmembrane mucins, secreted mucins lack transmembrane structural domains and exist in a secreted form[9]. Typical secreted mucins consist of a VWD domain rich in N-terminal cysteine, followed by a C-terminal "CK" domain. The N-terminal participates in polymerization through intermolecular disulfide bonds, while the C-terminal "CK" domain participates in monomer dimerization[10] (Figure 1). Both types of mucins have a highly glycosylated protein core, also known as the PTS domain, with multiple tandem repeats rich in Pro, Ser or Thr in the PTS domain. The size and number of tandem repeat sequences in different mucins, hence they are called variable number tandem repeats (VNTR), which means that there may be significant differences in size between individuals or individual mucins[1]. Secreted mucins are responsible for conferring viscoelasticity to epithelial tissues, and transmembrane mucins are involved in maintaining epithelial cell polarity and cell signaling[11]. In addition, the expression patterns of mucin genes in the respiratory, digestive, and reproductive tracts are complex and strictly regulated. The expression of each mucin has organ and cell specificity[12]. Available studies have shown that the mucin components of the lungs are mainly MUC5AC and MUC5B, MUC5B is essential for cilia motility, while MUC5AC is more responsive to environmental or infectious factors, and elevated concentrations of MUC5AC may contribute to the development of chronic obstructive pulmonary disease (COPD)[13]. Terada found by immunohistochemistry that normal gastric mucosa specifically expresses MUC2, MUC5AC and MUC6, but never MUC1, colorectal crypt epithelial cells highly express MUC2, but never MUC1, MUC5AC and MUC6[14]. In the female reproductive tract, the major transmembrane mucins are MUC1 and MUC4, and the major secreted mucins are MUC5B and MUC5AC[15]. These evidences suggest that the expression of mucins indeed organ and cell specific.
As mentioned earlier, mucin possesses a highly glycosylated PTS domain, which has a large and dense glycan chain structure. It is similar to a “bottle brush”, which allows carbohydrates to make up 80% of the total mass of the mucin. These glycan chains consist of a large number of O-glycans, and we refer to mucins and the glycans on them collectively as mucin glycans. Mucin glycans are beneficial for maintaining a high viscosity state (Figure 2). For example, in the stomach, high viscosity helps to lubricate indigestible lumen contents and accelerate gastric emptying. At the same time, they can protect epithelial cells from dehydration and mechanical forces during the passage of lumen contents[16]. However, abnormally high viscosity can also lead to disease, and it has been shown that abnormally high mucin viscosity leads to increased concentration of gallbladder bile and possible formation of gallstones[17].
It should be noted that mucins encoded by the same gene may have different structures due to different glycosylation patterns. Meanwhile, their expression levels and glycan patterns may vary depending on the organization and species, which may lead to the occurrence of diseases[18]. This may be due to changes in the topology, function and expression of individual glycosyltransferases and their molecular chaperones leading to glycan loss, which results in the production of aberrant glycan chains on mucins[19]. Existing studies have shown that alterations in intestinal mucin glycans are associated with a variety of intestinal disorders, and that they can lead to dysbiosis, which usually occurs in the early stages of enterocolitis[20]. Alterations in O-glycans on MUC2 have been observed in ulcerative colitis (UC), which refers to an increase in the number of truncated O-glycans Tn antigen (GalNAcα1-O-Ser/Thr), STn antigen (Neu5Acα2-6GalNAcα1-O-Ser/Thr) and a decrease in the number of complex O-glycans on MUC2. These abnormal mucin glycans will further aggravate UC[21]. In addition, during the development of cancer, the glycosylation pattern of mucins also changes. This abnormal glycosylation pattern can alter cell function and participate in cancer cell proliferation, invasion, metastasis and angiogenesis[19]. For example, the mucins of normal breast epithelial cells contain a mixture of O-glycans, most of which are core 2 structure (GlcNAcβ1-6Galβ1-3GalNAcα1-O-Ser/Thr) , but in breast cancer (BC) there is a decrease in core 2 structure and an increase in sialyl Lweis x (NeuAcα2-3Galβ1-4 [Fucα1-3] GlcNAc, SLex) antigen, which can lead to enhanced the adhesion of BCcells to endothelial cells[22].
2. Major Types of Glycosylation on Mucins
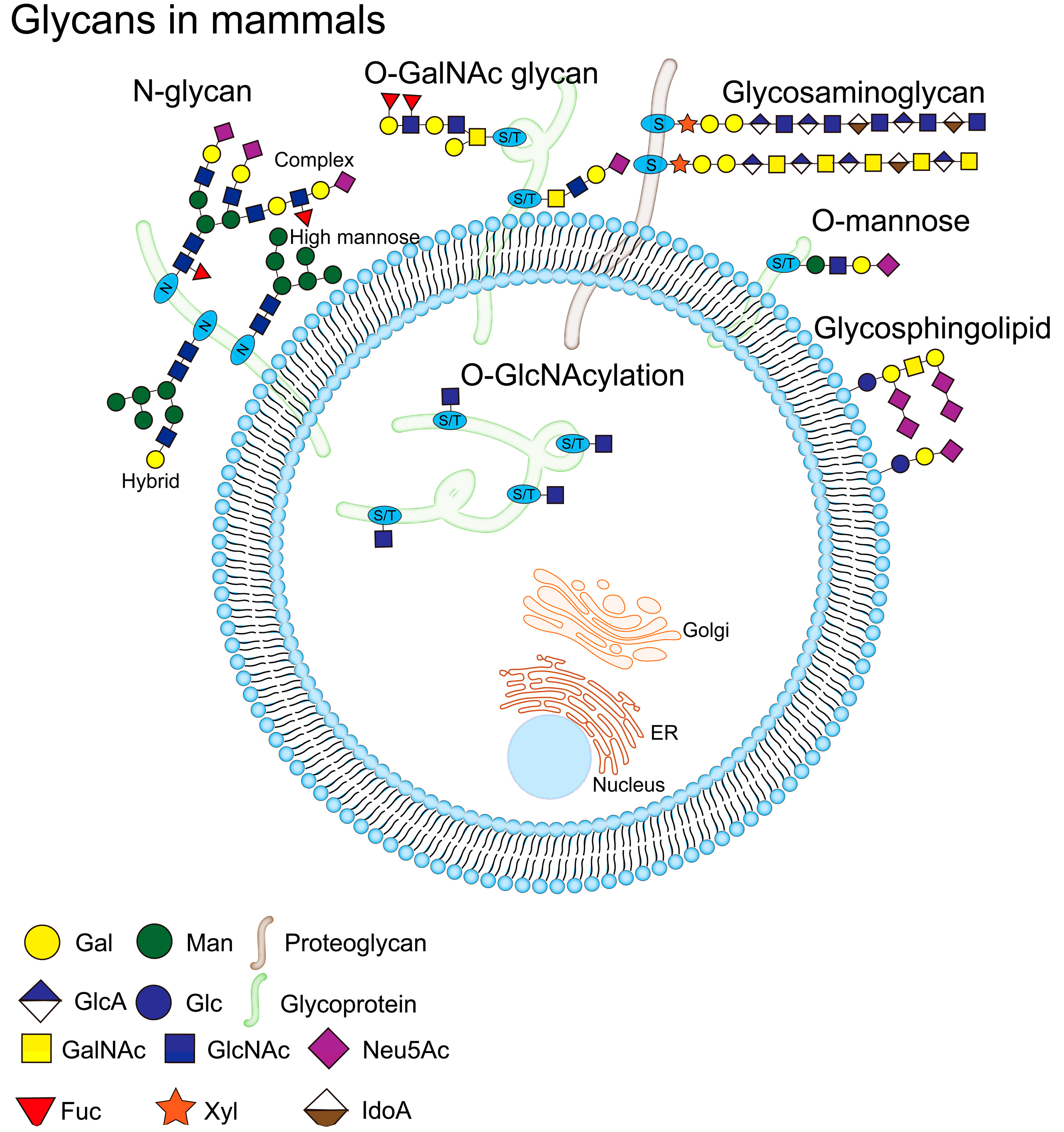
The complexity of mucin structures is due to their long polypeptide chains and various post-translational modifications (PTM), such as glycosylation, sulfation and phosphorylation[11]. Among them, glycosylation is one of the main PTM that define these mucins and affect their function, which mainly occurring in the endoplasmic reticulum (ER) and Golgi apparatus[23]. There are differences in the site of occurrence of each type of glycosylation, O-GalNAc glycosylation (also called mucin-type O-glycosylation) mainly occurs in the Golgi apparatus. N-glycosylation is initiated in the ER and further processed and terminated in the Golgi apparatus. Other types of glycosylation, including O-fucose, O-glucose, O-galactose, O-N-acetylglucosamine (O-GlcNAc), O-mannose, and O-xylose (proteoglycan) are occur in ER, while O-GlcNAcylation occurs in the cytoplasm,and their eventual localizations in the cells are also different [24,25,26,27,28] (Figure 3). Glycans are composed of ten monosaccharides: fucose (Fuc), galactose (Gal), glucose (Glc), N-acetylglucosamine (GlcNAc), N-acetylgalactosamine (GalNAc), glucuronic acid (GlcA), iduronic acid (IdoA), sialic acid (Sia), mannose (Man) and xylose (Xyl), which are linked by α or β bonding to form linear or branched structures[29,30,31]. Sia can provide a negative charge to the protein molecule, which introduces spatial repulsion in the side chains of the glycan chain, which in turn imparts rigid structural chains to the mucin, and these structures maximize the extension of the glycan chain[32]. Fuc can confer more neutral charge properties to mucins, which can maintain high viscosity of mucins by regulating the interaction of water molecules (or other charged ions) with mucins[33].
The glycosylation on mucin is mainly O-GalNAc glycosylation (hereinafter referred to as O-glycosylation) and N-glycosylation[34]. These two types of glycosylation may occur simultaneously on mucin, depending on the amino acid residues on the mucin. N-glycosylation connects carbohydrates to the amide nitrogen of Asn, while O-glycosylation connects carbohydrates to the hydroxyl groups of Ser/Thr[35,36]. Compared to N-glycosylation, O-glycosylation is the main glycosylation modification on mucins[37]. The processes of N-glycosylation and O-glycosylation are interconnected, and it has been found that when N-glycosylation is blocked, O-glycosylation of sucrose isomaltase and intestinal dipeptidyl peptidase IV is also inhibited[38]. In addition, there are some shared carbohydrate antigens between N-glycosylation and O-glycosylation. For example, the typical tumour-associated antigens sialyl Lewis a (NeuAcα2-3Galβ1-3 [Fucα1-4] GlcNAc, SLea) and SLex, which can appear on both N-glycans and O-glycans[39].
Figure 3.
Glycosylation is one of the processes of the PTS, glycosylation occurs mainly in the ER and the Golgi, and there is a difference on the site of occurrence of each type of glycosylation. O-GalNAc glycosylation occurs mainly in the Golgi. N-glycosylation is initiated in the ER and processed and terminated in the Golgi. O-fucose, O-glucose, O-galactose, O-N-acetylglucosamine (O-GlcNAc), O-mannose and O-xylose (proteoglycan) all occur in the ER, and O-GlcNAcylation occurs in the cytoplasm. Mammalian cell surfaces are covered with a dense array of glycoproteins, including N-glycans (high mannose, hybrid and complex N-glycan) and O-GalNAc glycans, etc. O-GlcNAcylation is present in cytoplasmic, nuclear and mitochondrial glycoproteins. The glycan structure is represented according to the Symbolic Nomenclature for Glycans (SNFG) format[40].
Figure 3.
Glycosylation is one of the processes of the PTS, glycosylation occurs mainly in the ER and the Golgi, and there is a difference on the site of occurrence of each type of glycosylation. O-GalNAc glycosylation occurs mainly in the Golgi. N-glycosylation is initiated in the ER and processed and terminated in the Golgi. O-fucose, O-glucose, O-galactose, O-N-acetylglucosamine (O-GlcNAc), O-mannose and O-xylose (proteoglycan) all occur in the ER, and O-GlcNAcylation occurs in the cytoplasm. Mammalian cell surfaces are covered with a dense array of glycoproteins, including N-glycans (high mannose, hybrid and complex N-glycan) and O-GalNAc glycans, etc. O-GlcNAcylation is present in cytoplasmic, nuclear and mitochondrial glycoproteins. The glycan structure is represented according to the Symbolic Nomenclature for Glycans (SNFG) format[40].
2.1. O-Glycosylation
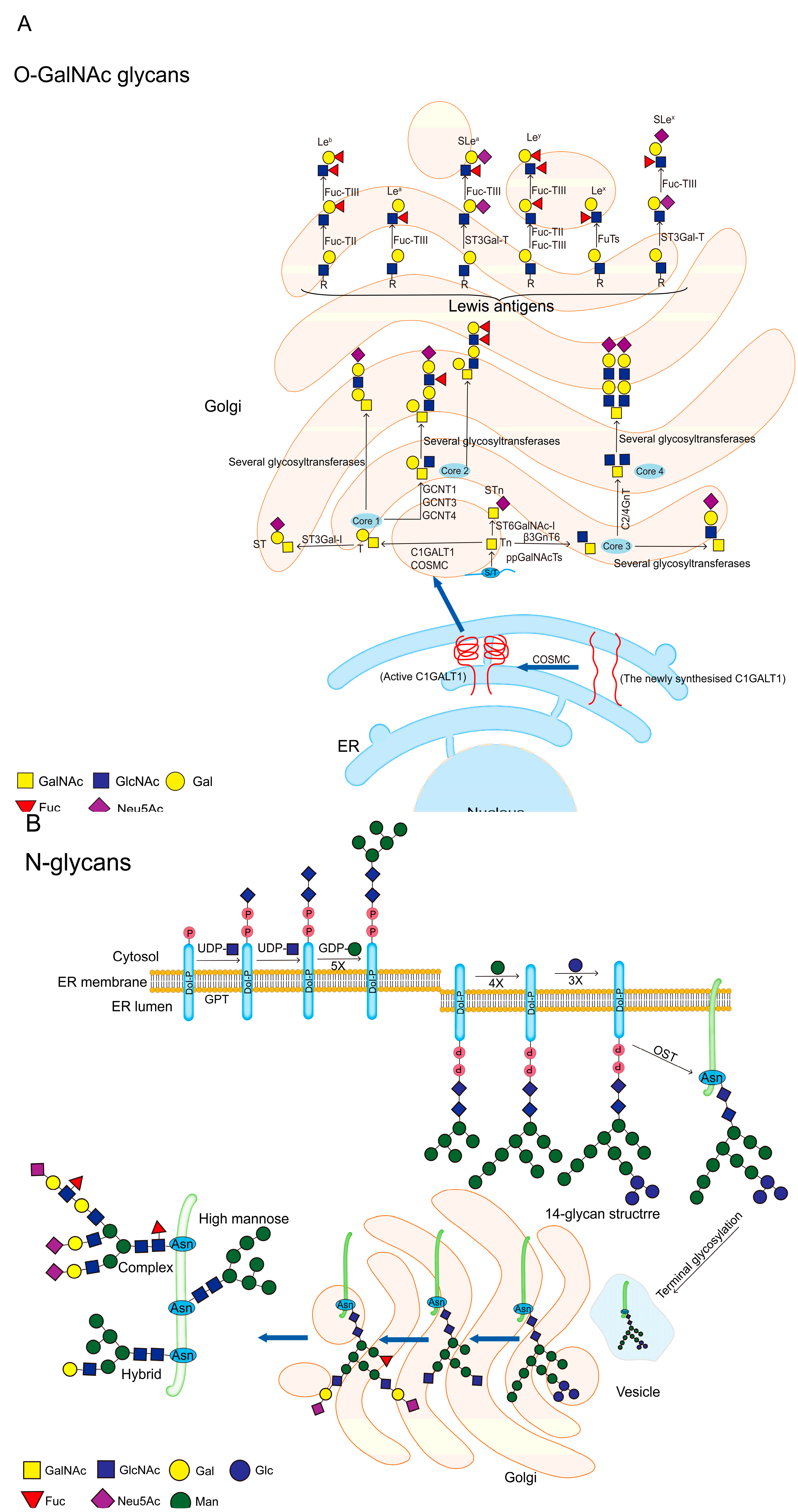
More than 80% of the glycan chains on mucin are composed of O-glycans. The first step of O-glycosylation is to add GalNAc from UDP-GalNAc to the Ser/Thr residue of the mucin PTS domain under the action of the peptide transferases family (ppGalNAcTs) to form the initial structure Tn antigen. There are 19 ppGalNAcTs family members in mice, 20 in mammals and 10 in drosophila, the expression of these enzymes is substrate and tissue specific[10]. Downward extension of the Tn antigen can produce eight different core structures, with core 1-4 are the four main core structures that are widely expressed in vivo. Expression of other core structures 5-8 are very limited, and expression of core 7 is not found in humans[35]. In detail, Tn antigen is synthesized into core 1 structure (also called T antigen, Galβ1-3GalNAcα1-O-Ser/Thr) by the action of core 1β1,3-galactosyltransferase (C1GALT1). In mammals, C1GALT1 requires the action of the molecular chaperone COSMC to fulfil its normal biological role[41]. COSMC is an ER-localized chaperone that interacts with newly synthesized C1GALT1 to promote correct folding of C1GALT1 and keeping C1GALT1 active[42]. Tn antigen can also be synthesized by core 3β1,3-N-acetylglucosaminyl transferase (β3GnT6) to form a core 3 structure (GlcNAcβ1-3GalNAcα1-O-Ser/Thr), or by sialyl transferase ST6GalNAc-I to synthesise STn antigen. T antigen and core 3 structures can be further branched to core 2 or core 4 structures by adding other sugar groups, and the four core structures are usually further extended and covered by terminal residues[43]. These residues include Fuc (Fucα1-2, -3 or -4) and Sia (NeuAc(Gc)α2-3 or -6) [9] (Figure 4A).The most common structure on mucins is the sialylated core 1 structure (also called ST antigen, NeuAcα2-3Galβ1-3 [NeuAcα2-6]+/-GalNAcα1-O-Ser/Thr), as well as the sialylated core 2 structure (NeuAcα2-3Galβ1-3[NeuAcα2-3Galβ1-3/4GlcNAcβ1-6] GalNAcα1-O-Ser/Thr) [35].
Normal initiation and extension of O-glycosylation on mucins affects a variety of biological processes, including cellular aspects (targeted transport of glycoproteins), molecular aspects (protein conformation, protein hydrolysis resistance) and aspects of cellular communication (cell-cell and cell-substrate interactions)[44]. On the one hand, O-glycans on mucins can inhibit the virulence of pathogens, for example, O-glycans on the salivary mucin MUC5B can inhibit the virulence of the oral pathogen Streptococcus mutans[45]. On the other hand, O-glycans on mucins are essential for maintaining the normal state of the eyes, and O-glycans on mucins are involved in maintaining the highly extended and rigid structure of mucins, which enables mucin and its O-glycan to maintain the sugar calyx structure at the top of the eye surface[46]. Moreover, O-glycans on mucin can interact with galactose lectins, for example, O-glycans on MUC16 and MUC1 interact with galactose lectins -1 (Gal-1) and -3 (Gal-3) to promote the formation of protective lattices in the cornea[47,48]. Abnormal O-glycosylation of mucins also leads to congenital disorders of glycosylation (CDG). So far, most CDGs are caused by defects in the coding region or spliceosomes of glycosyltransferases, which can cause abnormal truncated structures (Tn, STn, T, ST) or extended structures (SLex, SLea) on glycan chains[49,50].
Finally, normal biosynthesis of O-glycans on mucins is essential for cancer development. Taking MUC1 as an example, it is clinically called carbohydrate antigen (CA153). As early as May 1996, the American Society of Clinical Oncology (ASCO) evaluated MUC1 and designated it as a tumor marker for BC[51]. MUC1 expression and its aberrant O-glycosylation are frequently observed in epithelial cancers, and this aberrant O-glycosylation mainly refers to the increased expression of truncated O-glycans as well as aberrant extended structures, which play a key role in the occurrence, progression, and metastasis of cancer[52]. Expression of aberrant ST structures on MUC1 can promote cancer cell growth, and the immune system seems to play a role in this[53]. Existing studies have shown that the loss of O-glycans derived from core 1 can lead to dysregulation of MUC1 expression, damage to the gastric mucus layer, and altered gastric acid balance, leading to the occurrence of gastritis and gastric cancer (GC) [54]. Subsequently, studies found that MUC1 can promote GC metastasis, is associated with poor prognosis, and appears to be involved in the progression of diffuse gastric cancer (DGC) [55,56].
2.2. N-Glycosylation
In summary, the biological properties of mucins are associated with O-glycosylation. In turn, their N-glycosylation is important in the processing and biological properties of mucins[38]. N-glycosylation is initiated by the transfer of GlcNAc from UDP-GlcNAc to Dol-P lipid molecules on the ER membrane to form Dol-P-P-GlcNAc, this process mediated by UDP-N-acetylglucosamine-dolichyl-phosphate N-acetylglucosaminephosphotransferase (DAPGT1/GPT). Then, UDP-GlcNAc and 5 GDP-Man residues are sequentially linked by specific glycosyltransferases to generate the Man5GlcNAc2-P-P-Dol structure. This structure flips into the ER lumen and proceeds to add 4 Man and 3 Glc to produce the Glc3Man9GlcNAc2-P-P-Dol structure (14-glycan structure), the precursor of N-glycans. The 14-glycan structure is transferred by oligosaccharyl transferase (OST) to asparagine residues in the N-glycosylation site (Asn-X-Ser/Thr) of the nascent protein[36]. After that the 14-glycan structure is encapsulated by vesicles and enters the Golgi for terminal glycosylation, which involves removing mannose and adding other sugar groups. After terminal glycosylation, N-glycosylated proteins are divided into three main types, high mannose, hybrid and complex N-glycan[57] (Figure 4B).
N-glycosylation is important for the dimerization and multimerization of mucins, which can provide stability to mucins by promoting folding and inhibiting degradation, and is essential for mucin folding, sorting and secretion[38,58]. The presence of N-glycans at N9 site on MUC2 helps to maintain the folding rate of MUC2 monomers, which is slow enough to allow for the structural maturity and stability of MUC2 dimers. This means that the formation of MUC2 dimers cannot be separated from the N-glycosylation of MUC2[59]. Asker et al. found that inhibition of N-glycosylation of MUC2 with tunicamycin resulted in a reduction of dimers and oligomers of MUC2[60]. In addition, mucins and their N-glycans are also important for maintaining the normal state of the eyes. Taniguchi et al. found that the expression of MUC16 was downregulated and the function of the glycocalyx barrier was disrupted after inhibiting N-glycosylation in human corneal epithelial cells with tunicamycin. Notably, N-glycans in human corneal epithelial cells are mainly complex structures, which are galactose lectin ligands. N-glycans on MUC16 can promote the binding of MUC16 and Gal-3. And compared to O-glycans, Gal-3 has a higher affinity for N-glycans, indicating that complex N-glycans are the preferred ligand for galactose lectins[58].
Finally, abnormal N-glycosylation of mucins is also a mark of cancer[61]. Expression of highly branched β1-6 N-linked glycans is significantly up-regulated in various cancer cells compared to normal cells, which is associated with cancer cell proliferation, invasion and metastasis[62]. N-glycans can promote the binding of MUC16 and glycosylphosphatidylinositol anchored glycoprotein mesothelin, thereby promoting peritoneal metastasis of ovarian cancer (OV) [63]. Although N-glycosylation on mucins has been less studied in carcinogenesis, the role of N-glycosylation on other proteins in carcinogenesis has been widely reported. N-glycosylation of Asn294 and Asn454 of Mer tyrosine kinase (MerTK) promotes the proliferation of hepatocellular carcinoma (HCC) by stabilising the expression of MerTK[64]. N-glycosylation at Asn157 site of CD82 regulates colon cancer (COAD) transfer and adhesion by inhibiting epithelial mesenchymal transition (EMT) [65].
Figure 4.
O-GalNAc glycans and N-glycans. (a) The first step in the synthesis of O-GalNAc glycans is the attachment of UDP-GalNAc to Ser/Thr residues of the protein in the presence of ppGalNAcTs to form the initial structure GalNAcα1-O-Ser/Thr (Tn antigen). Tn antigen is synthesized through the core 1 structure (T antigen) by C1GALT1. C1GALT1 needs to be rendered active by COSMC in the ER, and active C1GALT1 exists as a dimer. Tn antigens can also synthesize core 3 structures via β3GnT6, and core 1 and core 3 structures can be further synthesized into core 2 or core 4 structures. STn antigens are produced by the addition of sialic acid to the α1-6 linkage of the GalNAc residue of Tn via ST6GalNAc-I. These core structures are usually further extended and terminated with sialic and fucose residues. Finally, terminal Lewis antigens includes Lea, Leb, Slea, Lex, Ley and Slex,ect; (b) N-glycosylation is initiated by transferring GlcNAc to Dol-P at the ER membrane to form Dol-P-P-GlcNAc, a process mediated by GPT. The 5 GDP-Man residues are then sequentially ligated on by specific glycosyltransferases to generate the Man5GlcNAc2-P-P-Dol structure. This structure is flipped into the ER lumen and the addition of 4 Man and 3 Glc continues to generate the 14-glycan structure, the precursor of the N-glycan. 14-glycan structure is transferred by OST to the Asn residue of the protein. The 14-glycan structure is then vesicle-wrapped into the Golgi for terminal glycosylation, which involves the removal of mannose and the addition of other sugar groups. After terminal glycosylation, N-glycosylated proteins are divided into three main types, high mannose, hybrid and complex. The glycan structure is represented according to the SNFG format[40].
Figure 4.
O-GalNAc glycans and N-glycans. (a) The first step in the synthesis of O-GalNAc glycans is the attachment of UDP-GalNAc to Ser/Thr residues of the protein in the presence of ppGalNAcTs to form the initial structure GalNAcα1-O-Ser/Thr (Tn antigen). Tn antigen is synthesized through the core 1 structure (T antigen) by C1GALT1. C1GALT1 needs to be rendered active by COSMC in the ER, and active C1GALT1 exists as a dimer. Tn antigens can also synthesize core 3 structures via β3GnT6, and core 1 and core 3 structures can be further synthesized into core 2 or core 4 structures. STn antigens are produced by the addition of sialic acid to the α1-6 linkage of the GalNAc residue of Tn via ST6GalNAc-I. These core structures are usually further extended and terminated with sialic and fucose residues. Finally, terminal Lewis antigens includes Lea, Leb, Slea, Lex, Ley and Slex,ect; (b) N-glycosylation is initiated by transferring GlcNAc to Dol-P at the ER membrane to form Dol-P-P-GlcNAc, a process mediated by GPT. The 5 GDP-Man residues are then sequentially ligated on by specific glycosyltransferases to generate the Man5GlcNAc2-P-P-Dol structure. This structure is flipped into the ER lumen and the addition of 4 Man and 3 Glc continues to generate the 14-glycan structure, the precursor of the N-glycan. 14-glycan structure is transferred by OST to the Asn residue of the protein. The 14-glycan structure is then vesicle-wrapped into the Golgi for terminal glycosylation, which involves the removal of mannose and the addition of other sugar groups. After terminal glycosylation, N-glycosylated proteins are divided into three main types, high mannose, hybrid and complex. The glycan structure is represented according to the SNFG format[40].
3. The Mechanisms of Abnormal Mucin Glycans Production
As mentioned earlier, normal glycosylation of mucin is particularly important for the maintenance of body health. When abnormal glycosylation occurs, it not only affects the expression of mucins, but also affects their biological functions. There are several main factors that contribute to the occurrence of abnormal glycosylation on mucins, including the alterations in the activity and localization of glycosyltransferases, alterations in the pH of Golgi, and the efficiency of nucleotide transport proteins[66]. Each of these mechanisms will be described in turn below.
3.1. The Activities and Localizations of Glycosyltransferases
The activity of glycosyltransferase and its localization in Golgi apparatus are the basis for normal glycosylation synthesis, which determines the structure and quantity of O-glycans[19]. Abnormal expression and activity of glycosyltransferases have been shown to be a factor in cancer development[49]. For example, GALNT7 can modify the O-glycosylation of prostate cancer (PRAD) cells and promote their proliferation[67]. Notably, O-glycosyltransferase activity can depend on N-glycosylation. Glycosyltransferases lacking N-glycosylation may misfold and aggregate in the ER. Prorok Hamon et al. found that FUT7 has two N-glycosylation sites at N81 and N291, changes in N-glycosylation at both sites can significantly decrease FUT7 activity[68]. Subsequently, Ruggiero et al. reported that ST3Gal-II has two N-glycosylation sites at N92 and N211. When N-glycosylation is absent at N211 site, ST3Gal-II undergoes misfolding, stalls in the ER and loses activity[69]. These abnormal glycosyltransferases further affect the biological process of glycosylation.
The localization of glycosyltransferase on the Golgi apparatus is an important factor in controlling glycan biosynthesis. The enzymes that synthesize core 1 and core 2 are mainly located in the cis Golgi apparatus, while the enzymes that synthesize terminal structures are mainly located in the trans Golgi apparatus[19]. The activity of the glycosyltransferase also changes when the glycosyltransferase is transferred from the cis Golgi to trans Golgi. Researchers transferred C2GnT (mainly located at cis Golgi apparatus) to trans Golgi apparatus and analyzed it. The results showed that the expression of core 2 branched oligosaccharides was drastically reduced after the transfer of C2GnT to the trans Golgi apparatus[70]. On the other hand, if the first glycosyl (GalNAc) is added later than the interval where the elongase is located, then GalNAc cannot be used as a glycosylation substrate[71].
3.2. Changes of Golgi pH
It is well known that the acidic pH of the Golgi lumen is essential for the correct glycosylation, transport and sorting of proteins and lipids in the organelle[72]. Elevated Golgi pH appears to be directly related to the expression of T antigens in cancer cells, and it may also affect the localization and distribution of glycosyltransferases in the Golgi apparatus[73]. Axelsson et al. stimulated HeLa and LS174T cells with ammonium chloride (NH4Cl) or proton pump inhibitor (BafA1) and found that they effectively neutralized pH, which resulted in the relocation of GALNT1 and GALNT2 from the ER to the Golgi apparatus[74]. Changes in Golgi pH can lead to incorrect synthesis of O-glycans and N-glycans[73]. There is evidence that stimulation of fibroblast COS-7 with pH gradient dissipating drugs (BafA1, chloroquine or NH4Cl) leads to an increase in Golgi pH and up-regulation of intracellular T antigen expression[71]. Treating colorectal cancer cell line LS174T with BafA1 promotes sulfation of mucin and upregulates the expression of T antigen[75].
3.3. Efficiency of Nucleotide Transporters
Before a glycosylation reaction can occur in eukaryotes, the activated sugar group must be translocated to the Golgi or ER, and only there can the sugar group be used as a substrate by glycosyltransferases. This is a task performed by a family of nucleotide sugar transporter proteins (NSTs) [76]. Mutations in NSTs or inefficient transport are associated with impaired glycosylation. For example, salic acid is transported into the Golgi as CMP-sialic acid (CMP-Sia), which is carried out by the CMP-Sia transporter protein (CST). CST mutations can lead to incomplete sialylation and thus abnormal glycosylation[77]. CST can transport CMP-Sia and UDP-Galactose (UDP-Gal). Mutations at the Tyr214 and Ser216 sites on CST can lead to a loss of its transport activity towards CMP-Sia, but do not affect its transport activity towards UDP-Gal. That is, Tyr214 and Ser216 on CST are essential for CMP-Sia transporter[78].
4. The Role of Abnormal Mucin Glycans in Cancer Development
Glycosylation is a highly dynamic process that produces significant changes during cancer development. Compared to normal cells, cancer cells express more branched N-glycans, higher levels of fucose, sialic acid glycans and truncated O-glycans. This rich and aberrant glycosylation is now widely regarded as one of the unique marks of cancer[61]. Under physiological conditions, mucin glycans are involved in the composition of the mucus barrier, they are the first responders of epithelial cells to mechanical or chemical damage, helping to maintain environmental balance within the body[79]. However, their role in protecting and repairing epithelial cells turns into a sharp edge in the carcinogenesis process. Abnormal mucin glycans are involved in the occurrence and development of cancer, and their role in cancer cell proliferation, adhesion, and invasion has been fully demonstrated. Here, we focus on the most studied MUC1 as an example. The expression of MUC1 is generally upregulated in most cancer cells, and the upregulated MUC1 is mainly in the plasma membrane and cytoplasm of cancer cells[80]. In addition to elevated expression, MUC1 on cancer cells undergoes aberrant glycosylation[81]. Below we will summarize the mechanisms by which aberrant mucin glycans contribute to cancer development.
4.1. Proliferative Capacity
MUC1 on normal breast epithelial cells mainly carries the core 2 structure. During the development of BC, core 2 structure on MUC1 is absent and core 1 structure is increased. In order to determine whether this change will promote the proliferation of BC cells in vivo, Mungul et al. constructed MUC1 cell lines expressing core 1 and core 2, respectively. The results showed that cells expressing core 1 structure proliferated faster than cells expressing core 2 structure[82]. Moreover, it has been well established that C1GALT1/GALNT6 can regulate the O-glycan structure on MUC1 and activate the MUC1-C/β-catenin signaling pathway to promote the proliferation of BC cell lines[83,84]. On the other hand, in PRAD, upregulation of the expression of glycosyltransferase GCNT1 leads to aberrant expression of SLex on MUC1, but whether this aberrant O-glycosylation is involved in prostate carcinogenesis has not been clearly stated[85]. Researchers have characterized MUC1 with O-glycans, and subsequently found that in prostate cancer, the O-glycans on MUC1 are mainly core 2 structure, followed by fucosylated types of core 2 structure. Overexpression of MUC1 did not affect the proliferation of PRAD cells, but may promote PRAD metastasis. This suggests that the pro-carcinogenic mechanism of MUC1 in prostate cancer may be different from other epithelial cancers such as BC, COAD and pancreatic cancer (PAAD) [86].
4.2. Loss of Adhesion
Adhesion molecules (CAM) are a class of proteins located on the surface of cells that mediate cell-to-cell or cell-to-extracellular matrix (ECM) contact and binding. As a form of information exchange between cells, they can undergo adhesion by recognizing specific adhesion receptors[87]. Adhesion molecules consist of four main families (integrins, cadherins, immunoglobulins and selectins). Downregulation of cadherin, upregulation of integrins, immunoglobulins and selectins leads to loss of adhesion of cancer cells from the primary site, thus promoting metastasis of cancer cells[88]. The O-glycan structure on mucins is one of the most important factors in cell adhesion or anti adhesion, and it is mainly changes in SLea and SLex that lead to loss of adhesion in cancer cells[11] (Figure 5). SLea and SLex on MUC1 cover adhesion molecules on the surface of cancer cells, thus causing cancer cells to lose adhesion, which has been confirmed in different cancers[89]. Rodriguez et al. found that MUC1 on COAD cells carries SLea and SLex, and that SLea and SLex modify MUC1 into an epitope of E-selectin, which may be one of the molecular mechanisms by which MUC1 promotes COAD metastasis[90]. Solatycka et al. found that overexpression of MUC1 in BC cells resulted in upregulation of T antigen expression and deletion of SLex expression, and overexpression of MUC1 altered the adhesion ability of breast cancer cells[91]. Park et al. found that overexpression of MUC1 in breast cancer cells induced abnormalities in cell adhesion molecules (β-catenin and E-cadherin), resulting in an anti-adhesion effect[92]. In cigarette smoke induced pneumonia, abnormal glycosylation on MUC1 can promote E-cadherin degradation, thereby promoting EMT[93].
4.3. Cancer Metastasis
Most cancer patient deaths result from cancer metastasis, and clinically upregulated SLex expression is usually associated with poor prognosis in cancer patients[94]. In addition, sialic acid glycans on MUC1 can bind to lectins to promote adhesion of cancer cells to vascular endothelial cells or promote the production of tumor microenvironment, which is conducive to cancer progression and metastasis[61,95]. Cell surface lectins that can bind to MUC1 are divided into three major classes (C-type, S-type and I-type lectins)[52]. The interaction of MUC1 with lectins is enhanced when there is an abnormal increase in SLea and SLex on MUC1, which promotes the aggregation of cancer cells and their metastasis to distal sites[52,96]. When lectin (Siglec-9) binds to the ST structure on MUC1 of myeloma cells, it promotes the formation of the tumor microenvironment and thus metastasis[53,97]. In addition, MUC1 is also a natural ligand for Gal-3, and the interaction of Gal-3 with MUC1 promotes the adhesion of already disseminated cancer cells to the endothelium of blood vessels, thus metastasis[98]. Finally, a possible mechanism by which MUC1 promotes cancer cell metastasis is related to anoikis. Piyush et al. proposed in 2017 that truncated O-glycans on MUC1 can enhance the anti anoikis properties of cancer cells, leading to loss of cancer cell adhesion and metastasis[99].
4.4. Cancer Immune Escape
Tumor immune escape is essential to ensure the survival of cancer cells, which is defined as the escape of cancer cells from recognition and attack by the immune system through various mechanisms. Tumor immune escape is induced by a number of factors, including loss or alteration of cancer antigenic substances, weakening of cancer immunogenicity, epigenetic changes in cancer cells, changes in intracellular signaling pathways in cancer cells, etc[100]. A large amount of evidence suggests that upregulated mucins in most adenocarcinoma cells are the main shielding medium for cancer cells to evade immune system surveillance, and abnormal glycosylation of mucins can help cancer cells evade immune system surveillance. This is mostly mediated by lectins, which affect the recognition of tumor antigens by the immune system by binding to aberrant mucin glycans[101].
SLea and SLex on MUC1 not only play a role in cancer cell adhesion and metastasis, but also compete with selectin ligands on leukocytes, interfering with their recognition and clearance of cancer cells, thereby helping cancer cells escape immune system surveillance[89]. Furthermore, when lectins are recruited to highly sialylated cancer cells, they prevent NK cells from recognizing cancer cells and block their initial immune response to cancer cells[52]. Overaccumulation of Tn and STn on MUC1 is particularly important for tumor cell immune escape. Moreover, aberrant glycans on mucins modulate the susceptibility to NK cell mediated antibody dependent cytotoxicity (ADCC) and cytotoxic T lymphocyte (CTL) mediated killing[102].
4.5. Carcinogenic Pathogens
As of 2018, the four most important of the 11 infectious agents defined by the International Agency for Research on Cancer (IARC) as Class I carcinogens are Helicobacter pylori (Hp), high-risk human papillomavirus (HPV), hepatitis B virus (HBV) and hepatitis C virus (HCV) [103]. Abnormal mucin glycans may be either favourable or unfavourable to pathogens infecting the body, as they themselves can affect the resistance of host[104,105]. It has been shown that abnormal mucin glycans can provide a favourable environment for pathogens in the mucus layer during pathogen infection, thus increasing the survival of pathogens[106].
Infection with Hp is the greatest risk factor for GC[107]. It was shown that knockout of fucosyltransferase 2 (Fut2) resulted in downregulation of the expression of α1,2-fucosylated structures and upregulation of SLea expression on MUC5AC. This aberrant fucosylation on MUC5AC would help Hp to colonize the gastric mucosa[108]. In addition, Skoog et al. analyzed the O-glycans on gastric mucin infected and uninfected with Hp, respectively. It was found that after infection with Hp, the O-glycan structure on gastric mucin became more numerous and complex[109]. HPV can lead to the occurrence of cervical cancer (CESC). Solórzano et al. conducted O-glycan analysis of mucins in CESC infected with HPV, and the results showed that HPV infection can cause abnormal fucosylation of mucins, which can affect the invasion of CESC cells[110]. Ahmad et al. found that during HCV/HBV infection, the binding of O-glycans at the Ser204 site of insulin-like growth factor to cellular growth factor is reduced, which can lead to the occurrence of HCC[111].
Speaking of oncogenic pathogens here it is important to mention Epstein-Barr virus (EBV), which is the first human virus directly associated with cancer and has been linked to the pathogenesis of Burkitt's lymphoma (BL), Hodgkin's lymphoma (HL), non-Hodgkin's lymphoma (NHL) and nasopharyngeal carcinoma (NPC). Not only that, it is also associated with epithelial malignancies such as GC and BC[112]. EBV can upregulate the expression of core 2β-1,6-acetylglucosaminyltransferase (GCNT3) by affecting the NF-κB signaling pathway, which promotes the proliferation and migration of GC cells[113]. In addition, EBV latent membrane protein 1 (LMP1) can activate STAT signaling by regulating MUC1 expression, which ultimately promotes cancer invasion and metastasis[114]. Unfortunately, relevant studies are currently limited to interactions between EBV and aberrant glycosylation or mucin expression, whether EBV can interact with glycosylation on mucins is currently unknown. In summary, it is worth considering whether EBV can cause cancer by regulating glycosylation on mucins.
5. MUC1-Based Cancer Diagnosis and Targeted Therapy
Aberrant glycosylation may induce a range of different cancer associated epitopes. These epitopes can be divided into three categories, starting with truncated O-glycans (T, Tn and STn antigens), terminal structures of glycoproteins and lipids (SLea and SLex) [50,115,116]. Abnormal expression of mucins is often observed during the development of cancer[116]. Therefore, using abnormal mucin glycans on cancer cells as an immunotherapy strategy can provide a basis for the diagnosis and treatment of cancer[117].
MUC1 was initially detected by monoclonal antibodies (mAbs) targeting human milk fat globules (HMFG) [118]. It is the first cloned mucin and the best characterized mucin to date[119,120]. The N-terminal structural domain of MUC1 has variable number tandem repeat (VNTR) on which glycosylation occurs, and each VNTR consists of 20 amino acids (GVTSAPDTRPAPGSTAPPAH). The PDTR sequence is core of MUC1, and is also defined as the unique extracellular domain of MUC1[121]. The PDTRP sequence can be recognized by monoclonal antibodies and appears to be the immunodominant epitope[122,123]. MUC1 has always been considered a target for cancer therapy and diagnosis because it is upregulated and frequently aberrantly glycosylated in most adenocarcinomas[120]. Recently, in a ranking of 75 tumor antigens based on characteristics such as therapeutic function, immunogenicity, oncogenicity and specificity, MUC1 received the second best rating (after WT1), underlining its potential for medical research and vaccine development[124]. Here, we describe the antibodies, radiopharmaceuticals, vaccines, and CAR-T cell therapies based on MUC1 and summarize them in a table (Table1).
5.1. Antibodies
Antibodies against aberrant glycosylation on MUC1 (MUC1-Tn, MUC1-STn) have been demonstrated in BC, OV and PRAD[125]. As early as 1987, Burchell et al. designed a mAb (SM3) against the PDTRP sequence on MUC1, which responded to 91% of BC, but showed little response to benign breast tumor, normal resting, pregnant or breastfeeding mammary glands[126,127]. Subsequently, Rokhlin et al. in 1998 designed mAb (5E10) targeting the extracellular structural domain of MUC1 [128]. The specificity of 5E10 is relatively good, and it can react with benign and malignant PRAD, but has no reactivity to normal prostate tissue[128]. In addition, humanized antibody (5E5) directed against the MUC1-Tn/STn epitope specifically recognizes the GSTAP sequence of MUC1 and its immunoreactivity against MUC1-Tn is better, it can activate NK cell mediated antibody-dependent cellular cytotoxicity (ADCC) process in vitro [129,130,131]. To this day, various antibodies have been created against different structural domains of MUC1, and they have been clinically successful. However, antibodies against the VNTR epitope have not yet yielded results in clinical trials, which have failed mainly due to the shedding of the VNTR epitope and its release into the circulation, thus preventing the binding of antibodies to the structural domain of MUC1 on the surface of cancer cells[132].
Therefore, in recent years, most attention has been focused on designing antibodies or antibody conjugated drugs targeting other domains of MUC1. Recently, a mAb (TAB004) specifically targeting the extracellular structural domain of human MUC1 has been developed. It is reported that upon treatment of PAAD cells with TAB004, the binding of TAB004 to MUC1 induced endoplasmic reticulum stress and anoikis in the PAAD cells, which resulted in an inhibition of the proliferative capacity of the PAAD cells. When TAB004 was combined with 5-FU, it enhanced the survival of PAAD mice[133]. Wu et al. developed a mAb targeting MUC1-C (hMUC1), which recognized recombinant MUC1 as well as natural MUC1-C in BC cells, and hMUC1 significantly inhibited BC cell proliferation in vivo [134]. Subsequently, they developed a humanized MUC1 antibody (HzMUC1) targeting the interaction region between MUC1-N and MUC1-C. They coupled HzMUC1 with monomethyloristatin (MMAE) to generate an antibody drug coupling (ADC), which can inhibit the growth of PAAD cells by inducing PAAD cell cycle arrest and apoptosis[135].
5.2. Radiopharmaceuticals
Despite the high specificity and affinity of mAbs, they have some drawbacks such as high immunogenicity, limited penetration into tumor tissue, large size and long plasma half-life. And effective strategy to reduce these drawbacks is to use radioactive elements to label mAbs, which is also known as radiopharmaceuticals[136]. Radiopharmaceuticals consist of two parts, namely radioactive elements and carriers. The carrier is a class of biologically active molecules, typically antibodies, peptides, and aptamers[137].
The specific antibody (PR81) of MUC1 has a high affinity for breast cancer, so Salouti et al. radiolabeled PR81 with 99mTc, which improved the stability and immune reactivity of PR81 in vitro [138]. In addition, the anti-MUC1 aptamer was labelled with 99mTc to generate a labelled drug delivery system (DDS), which was subsequently used for in vivo imaging of triple-negative breast cancer (TNBC). The results showed that the DDS had a high tumor uptake (5%) and with great in vivo imaging properties[139]. Jammaz et al. radiolabeled the MUC1-FA-SFB hybrid conjugate with 18F, and the labeled conjugate (MUC1-FA - [18F] SFB) has good affinity and specificity for BC. In addition, MUC1-FA - [18F] SFB may be a PET imaging probe for BC detection and prognosis monitoring[140]. Therefore, there is great potential for labelling antibodies or aptamers to MUC1 with radionuclides, which could enhance the therapeutic effect of drugs for cancer treatment[141]. Compared to traditional treatment methods, Radiopharmaceutical therapy (RPT) is the least toxic and more targeted form of effective cancer treatment[142,143]. Although RPT has been shown to be an effective cancer treatment and has been clinically studied for over 40 years, it has not become part of the cancer treatment armamentarium like other therapies[142]. One possible reason is that the targeted pathways of these drugs are not involved in the formation of the malignant phenotype of cancer[144].
5.3. Vaccines
MUC1 is also a promising target for vaccine development, which can induce an immune response against MUC1[145]. Recently, MUC1 was rated by the American Cancer Institute Working Group as one of the most promising cancer vaccine targeted antigens in clinical practice[124]. Vaccines targeting MUC1 include subunit, DNA, viral, dendritic cell (DC) and glycopeptide vaccines[146]. The B subunit of Vibrio cholerae toxin (CTB) has great potential as a carrier for subunit vaccines. Pinkhasov et al. ligated the VNTR region of MUC1 with CTB to form a subunit vaccine. The vaccine was then delivered orally to tumor-bearing mice, and they found that the vaccine effectively inhibited tumor growth in the mice[147]. The DNA vaccine developed by Rong et al. (pcDNA3.1 VNTR) can enhance MUC1 induced CTL activity, inhibit tumor growth in mice, and prolong their survival time[148]. Quoix et al. developed a viral vaccine (TG4010), is a recombinant viral vaccine targeting MUC1, which significantly improves the survival rate of non-small cell lung cancer (NSCLC) patients with TG4010 treatment[149]. In addition, targeted MUC1 vaccines based on dendritic cells (DC) have broad therapeutic prospects. Studies have shown that this vaccine can induce anti-tumor immune responses, thereby prolonging the survival of NSCLC patients[150]. Finally, MUC1 glycopeptide vaccine targeting MUC1-Tn and MUC1-STn have been synthesized, which is used to diagnose BC and PAAD. However, its clinical performance is still being evaluated in phase II/III clinical trials[151].
With the deepening of anti-tumor immune mechanisms, MUC1 based vaccines have become a major concern in the clinical diagnosis and treatment of cancer. Although cancer vaccines targeting MUC1 have not yet been successfully used in the clinic. However, it must be emphasized that cancer vaccines targeting MUC1 have great anti-tumor potential when further refined and evaluated in clinical trials[146].
5.4. CAR-T
CAR-T is a type of T cell that can recognize a specific antigen and has the function of killing cancer cells when injected into the patient's body. This tumor targeted therapy method is called CAR-T therapy[152]. CAR-T therapy has made promising breakthroughs in the treatment of hematological malignancies, but its therapeutic role in solid tumors is limited[153]. Carl H June’s team in 2016 designed a new CAR-T cell targeting MUC1-Tn. This CAR-T cell inhibits the growth of MUC1-Tn positive cancer cells in the mouse model of leukemia and pancreatic cancer[154]. Then they designed CAR-T cells targeting c-Met and evaluated the safety and feasibility of c-Met-CAR T cells for the treatment of metastatic breast cancer in a phase 0 clinical trial, they found extensive tumor necrosis at the injection site, tumor cell debris and good tolerability[155]. Recent studies have indicated that MUC1-Tn is a potential therapeutic target for intrahepatic cholangiocarcinoma (ICC), and that CAR-T cells targeting MUC1-Tn can specifically eliminate MUC1-Tn-positive ICC cells but not MUC1-Tn-negative ICC cells in vitro and in vivo, which may be a novel therapeutic strategy for ICC[154,156]. Meanwhile, CAR-T cells targeting MUC1 (MUC28z CAR-T) exhibited specific cytotoxicity against TNBC, and after recognizing MUC1 on TNBC cells, the MUC28z CAR-T cells markedly inhibited the growth of TNBC cells with minimal damage to normal breast epithelial cells[153].
In fact, the efficacy of CAR-T therapy in solid tumors has always been poor, which may be due to the tumor microenvironment inhibiting the action of CAR-T cells as well as the incomplete activation of T cell function. Therefore, in order to improve the efficacy of CAR-T cells against solid tumors, Zhang et al. constructed a new CAR-T cell in 2020. They used the JAK-STAT structural domain to provide cytokine signals to CAR-T cells. This enhanced CAR-T cell has stronger cytotoxicity and more significant inhibitory effect on esophageal cancer (EC)[157] . In the same year, Shao et al. designed an inverted chimeric cytokine receptor (ICR), which consists of the extracellular structural domain of TGF-β and the intracellular structural domain of the IL-7 receptor, co-expressed on CAR-T cells. It can target prostate specific membrane antigen (PSMA), which exhibits superior anti-tumor ability and prolongs the survival time of PRAD mice[158]. Based on the above studies, CAR-T cells targeting MUC1 are also expected to make more progress. In the future, we may also be able to design enhanced CAR-T cells targeting MUC1.

Table 1.
MUC1-based antibodies, radiopharmaceuticals, vaccines and CAR-T therapies are covered in the text.
Table 1.
MUC1-based antibodies, radiopharmaceuticals, vaccines and CAR-T therapies are covered in the text.
| Type of therapy | Designation | Antigenic epitope/target | Type of cancer | Ref. |
|---|---|---|---|---|
|
Antibodies |
SM3 5E10 5E5 TAB004 hMUC1 HzMUC1 |
PDTRP MUC1-N MUC1-Tn/STn MUC1(STAPPAHGV) MUC1-C MUC1-N and MUC1-C |
Breast cancer Prostate cancer Breast cancer Pancreatic cancer Breast cancer Pancreatic cancer |
[126,127] [128] [129,131] [133] [134] [135] |
|
Radiopharmac-euticals |
99mTc-HYNIC-PR81 DDS MUC1-FA- [18F] SFB |
MUC1(AVGLSPDGSRGV) MUC1 MUC1 |
Breast cancer Triple-negative breast cancer Breast cancer |
[138] [139] [140] |
|
Vaccines |
CTB-MUC1 pcDNA3.1-VNTR TG4010 DC-based vaccine MUC1-glycopeptide vaccines |
VNTR VNTR MUC1 MUC1 MUC1-Tn/STn |
Breast cancer Pancreatic cancer Non-small-cell lung cancer Non-small-cell lung cancer Breast cancer, Pancreatic cancer |
[147] [148] [149] [150] [151] |
|
CAR-T |
Anti-MUC1-Tn-CAR-T Anti-MUC1-Tn-CAR-T MUC28z CAR-T Enhanced MUC1-CAR-T ICR |
MUC1-Tn MUC1-Tn MUC1 MUC1 --- |
Leukemia, pancreatic cancer Intrahepatic cholangiocarcinoma Triple-negative breast cancer Esophageal cancer Prostate cancer |
[154] [156] [153] [157] [158] |
6. Conclusions
Mucin glycans are involved in the composition of the mucus barrier. The health of the body and homeostasis of the internal environment are dependent on the normal expression of mucin glycans. Abnormal mucin glycan is a significant marker of cancer cells, the phenomenon noted by researchers as early as 1952[159]. Since then, more and more researchers have devoted themselves to the effect of mucin glycans on the development of tumors. To this day, the mechanisms by which abnormal mucin glycans lead to cancer development have been widely reported. On this basis, targeting mucin glycans turns into a new strategy for cancer diagnosis and treatment. Taking the best characterized MUC1 as an example, researchers have developed antibodies, radiopharmaceuticals, vaccines, and CAR-T cell therapies against MUC1. There is encouraging news that MUC1 targeting CAR-T cells has been successfully used in clinical trials for the treatment of advanced NSCLC and BC[160]. As we continue to explore the pathogenic mechanisms by which mucin glycans regulate cancer progression, the combination of targeted mucin glycans with existing cancer treatment options may have a significant impact on cancer treatment.
Author Contributions
L.S., Y.Z. (Yuhan Zhang), J.Z. and Y.Z. (Yuecheng Zhang) conceived and wrote the article; L.S., Y.Z. (Yuhan Zhang), W.L., J.Z. and Y.Z. (Yuecheng Zhang) reviewed and edited the article. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Behera, S.K.; Praharaj, A.B.; Dehury, B.; Negi, S. Exploring the role and diversity of mucins in health and disease with special insight into non-communicable diseases. Glycoconj. J. 2015, 32, 575–613. [CrossRef]
- Wi, D.-H.; Cha, J.-H.; Jung, Y.-S. Mucin in cancer: a stealth cloak for cancer cells. BMB Rep. 2021, 54, 344–355. [CrossRef]
- Van Seuningen, P. Pigny, M. Perrais, N. Porchet, J.P. Aubert, Transcriptional regulation of the 11p15 mucin genes. Towards new biological tools in human therapy, in inflammatory diseases and cancer?, Frontiers in bioscience : a journal and virtual library 6 (2001) D1216-34. [CrossRef]
- Tailford, L.E.; Crost, E.H.; Kavanaugh, D.; Juge, N. Mucin glycan foraging in the human gut microbiome. Front. Genet. 2015, 6, 81–81. [CrossRef]
- Jonckheere, N.; Vincent, A.; Neve, B.; Van Seuningen, I. Mucin expression, epigenetic regulation and patient survival: A toolkit of prognostic biomarkers in epithelial cancers. Biochim. et Biophys. Acta (BBA) - Rev. Cancer 2021, 1876, 188538. [CrossRef]
- S. Pinzón Martín, P.H. Seeberger, D. Varón Silva, Mucins and Pathogenic Mucin-Like Molecules Are Immunomodulators During Infection and Targets for Diagnostics and Vaccines, Frontiers in chemistry 7 (2019) 710. [CrossRef]
- Buscaglia, C.A.; Campo, V.A.; Frasch, A.C.C.; Di Noia, J.M. Trypanosoma cruzi surface mucins: host-dependent coat diversity. Nat. Rev. Genet. 2006, 4, 229–236. [CrossRef]
- Szafranski-Schneider, E.; Swidergall, M.; Cottier, F.; Tielker, D.; Román, E.; Pla, J.; Ernst, J.F. Msb2 Shedding Protects Candida albicans against Antimicrobial Peptides. PLOS Pathog. 2012, 8, e1002501. [CrossRef]
- G.C. Hansson, Mucins and the Microbiome, Annual review of biochemistry 89 (2020) 769-793. [CrossRef]
- Syed, Z.A.; Zhang, L.; Hagen, K.G.T. In vivo models of mucin biosynthesis and function. Adv. Drug Deliv. Rev. 2022, 184, 114182–114182. [CrossRef]
- Hollingsworth, M.A.; Swanson, B.J. Mucins in cancer: protection and control of the cell surface. Nat. Rev. Cancer 2004, 4, 45–60. [CrossRef]
- Jonckheere, N.; Van Seuningen, I. The membrane-bound mucins: From cell signalling to transcriptional regulation and expression in epithelial cancers. Biochimie 2010, 92, 1–11. [CrossRef]
- G. Radicioni, A. Ceppe, A.A. Ford, N.E. Alexis, R.G. Barr, E.R. Bleecker, S.A. Christenson, C.B. Cooper, M.K. Han, N.N. Hansel, A.T. Hastie, E.A. Hoffman, R.E. Kanner, F.J. Martinez, E. Ozkan, R. Paine, 3rd, P.G. Woodruff, W.K. O'Neal, R.C. Boucher, M. Kesimer, Airway mucin MUC5AC and MUC5B concentrations and the initiation and progression of chronic obstructive pulmonary disease: an analysis of the SPIROMICS cohort, The Lancet. Respiratory medicine 9(11) (2021) 1241-1254. [CrossRef]
- Terada, T. An immunohistochemical study of primary signet-ring cell carcinoma of the stomach and colorectum: II. Expression of MUC1, MUC2, MUC5AC, and MUC6 in normal mucosa and in 42 cases.. Int. J. Clin. Exp. Pathol. 2013, 6, 613–21.
- Burcham, L.R.; Bath, J.R.; Werlang, C.A.; Lyon, L.M.; Liu, N.; Evans, C.; Ribbeck, K.; Doran, K.S. Role of MUC5B during Group B Streptococcal Vaginal Colonization. Mbio 2022, 13, e0003922. [CrossRef]
- Johansson, M.E.V.; Sjövall, H.; Hansson, G.C. The gastrointestinal mucus system in health and disease. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 352–361. [CrossRef]
- D. Jüngst, A. Niemeyer, I. Müller, B. Zündt, G. Meyer, M. Wilhelmi, R. del Pozo, Mucin and phospholipids determine viscosity of gallbladder bile in patients with gallstones, World journal of gastroenterology 7(2) (2001) 203-7. [CrossRef]
- Kohout, V.R.; Wardzala, C.L.; Kramer, J.R. Synthesis and biomedical applications of mucin mimic materials. Adv. Drug Deliv. Rev. 2022, 191, 114540. [CrossRef]
- Brockhausen, Mucin-type O-glycans in human colon and breast cancer: glycodynamics and functions, EMBO reports 7(6) (2006) 599-604. [CrossRef]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [CrossRef]
- Larsson, J.M.H.; Karlsson, H.; Crespo, J.G.; Johansson, M.E.V.; Eklund, L.; Sjövall, H.; Hansson, G.C. Altered O-glycosylation profile of MUC2 mucin occurs in active ulcerative colitis and is associated with increased inflammation. Inflamm. Bowel Dis. 2011, 17, 2299–2307. [CrossRef]
- Matsuura, N.; Narita, T.; Hiraiwa, N.; Hiraiwa, M.; Murai, H.; Iwase, T.; Funahashi, H.; Imai, T.; Takagi, H.; Kannagi, R. Gene expression of fucosyl- and sialyl-transferases which synthesize sialyl Lewisx, the carbohydrate ligands for E-selectin, in human breast cancer.. Int. J. Oncol. 1998, 12, 1157–1221. [CrossRef]
- Varki, Biological roles of glycans, Glycobiology 27(1) (2017) 3-49. [CrossRef]
- B.M. Harvey, R.S. Haltiwanger, Regulation of Notch Function by O-Glycosylation, Advances in experimental medicine and biology 1066 (2018) 59-78. [CrossRef]
- B.C. Holdener, R.S. Haltiwanger, Protein O-fucosylation: structure and function, Current opinion in structural biology 56 (2019) 78-86. [CrossRef]
- Larsen, I.S.B.; Narimatsu, Y.; Clausen, H.; Joshi, H.J.; Halim, A. Multiple distinct O-Mannosylation pathways in eukaryotes. Curr. Opin. Struct. Biol. 2019, 56, 171–178. [CrossRef]
- Mereiter, S.; Balmaña, M.; Campos, D.; Gomes, J.; Reis, C.A. Glycosylation in the Era of Cancer-Targeted Therapy: Where Are We Heading? Cancer Cell 2019, 36, 6–16. [CrossRef]
- Yu, H.; Takeuchi, H. Protein O-glucosylation: another essential role of glucose in biology. Curr. Opin. Struct. Biol. 2019, 56, 64–71. [CrossRef]
- Schjoldager, K.T.; Narimatsu, Y.; Joshi, H.J.; Clausen, H. Global view of human protein glycosylation pathways and functions. Nat. Rev. Mol. Cell Biol. 2020, 21, 729–749. [CrossRef]
- Cummings, R.D. The repertoire of glycan determinants in the human glycome. Mol. Biosyst. 2009, 5, 1087–1104. [CrossRef]
- A.V. Nairn, W.S. York, K. Harris, E.M. Hall, J.M. Pierce, K.W. Moremen, Regulation of glycan structures in animal tissues: transcript profiling of glycan-related genes, The Journal of biological chemistry 283(25) (2008) 17298-313. [CrossRef]
- L.A. Tabak, In defense of the oral cavity: structure, biosynthesis, and function of salivary mucins, Annual review of physiology 57 (1995) 547-64.
- R.A. Cone, Barrier properties of mucus, Advanced drug delivery reviews 61(2) (2009) 75-85. [CrossRef]
- Chugh, S.; Gnanapragassam, V.S.; Jain, M.; Rachagani, S.; Ponnusamy, M.P.; Batra, S.K. Pathobiological implications of mucin glycans in cancer: Sweet poison and novel targets. Biochim. et Biophys. Acta (BBA) - Rev. Cancer 2015, 1856, 211–225. [CrossRef]
- in: A. Varki, R.D. Cummings, J.D. Esko, P. Stanley, G.W. Hart, M. Aebi, D. Mohnen, T. Kinoshita, N.H. Packer, J.H. Prestegard, R.L. Schnaar, P.H. Seeberger (Eds.), Essentials of Glycobiology, Cold Spring Harbor Laboratory Press Copyright © 2022 by the Consortium of Glycobiology Editors, La Jolla, California. Published by Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York. All rights reserved., Cold Spring Harbor (NY), 2022.
- R. Kornfeld, S. Kornfeld, Assembly of asparagine-linked oligosaccharides, Annual review of biochemistry 54 (1985) 631-64. [CrossRef]
- P. Van den Steen, P.M. Rudd, R.A. Dwek, G. Opdenakker, Concepts and principles of O-linked glycosylation, Critical reviews in biochemistry and molecular biology 33(3) (1998) 151-208. [CrossRef]
- Parry, S.; Hanisch, F.G.; Leir, S.-H.; Sutton-Smith, M.; Morris, H.R.; Dell, A.; Harris, A. N-Glycosylation of the MUC1 mucin in epithelial cells and secretions. Glycobiology 2006, 16, 623–634. [CrossRef]
- Blanas, N.M. Sahasrabudhe, E. Rodríguez, Y. van Kooyk, S.J. van Vliet, Fucosylated Antigens in Cancer: An Alliance toward Tumor Progression, Metastasis, and Resistance to Chemotherapy, Frontiers in oncology 8 (2018) 39. [CrossRef]
- Varki, A.; Cummings, R.D.; Aebi, M.; Packer, N.H.; Seeberger, P.H.; Esko, J.D.; Stanley, P.; Hart, G.; Darvill, A.; Kinoshita, T.; et al. Symbol Nomenclature for Graphical Representations of Glycans. Glycobiology 2015, 25, 1323–1324. [CrossRef]
- T. Ju, R.D. Cummings, A unique molecular chaperone Cosmc required for activity of the mammalian core 1 beta 3-galactosyltransferase, Proceedings of the National Academy of Sciences of the United States of America 99(26) (2002) 16613-8. [CrossRef]
- T. Ju, R.P. Aryal, M.R. Kudelka, Y. Wang, R.D. Cummings, The Cosmc connection to the Tn antigen in cancer, Cancer biomarkers : section A of Disease markers 14(1) (2014) 63-81. [CrossRef]
- Bergstrom, K.S.B.; Xia, L. Mucin-type O-glycans and their roles in intestinal homeostasis. Glycobiology 2013, 23, 1026–1037. [CrossRef]
- Breloy, I.; Hanisch, F.-G. Functional Roles of O-Glycosylation. Molecules 2018, 23, 3063. [CrossRef]
- Werlang, C.A.; Chen, W.G.; Aoki, K.; Wheeler, K.M.; Tymm, C.; Mileti, C.J.; Burgos, A.C.; Kim, K.; Tiemeyer, M.; Ribbeck, K. Mucin O-glycans suppress quorum-sensing pathways and genetic transformation in Streptococcus mutans. Nat. Microbiol. 2021, 6, 574–583. [CrossRef]
- B. Nichols, C.R. Dawson, B. Togni, Surface features of the conjunctiva and cornea, Investigative ophthalmology & visual science 24(5) (1983) 570-6.
- Argüeso, P.; Guzman-Aranguez, A.; Mantelli, F.; Cao, Z.; Ricciuto, J.; Panjwani, N. Association of Cell Surface Mucins with Galectin-3 Contributes to the Ocular Surface Epithelial Barrier. J. Biol. Chem. 2009, 284, 23037–23045. [CrossRef]
- Seelenmeyer, C.; Wegehingel, S.; Lechner, J.; Nickel, W. The cancer antigen CA125 represents a novel counter receptor for galectin-1. J. Cell Sci. 2003, 116, 1305–1318. [CrossRef]
- Brockhausen, Pathways of O-glycan biosynthesis in cancer cells, Biochimica et biophysica acta 1473(1) (1999) 67-95. [CrossRef]
- S. Hakomori, Glycosylation defining cancer malignancy: new wine in an old bottle, Proceedings of the National Academy of Sciences of the United States of America 99(16) (2002) 10231-3. [CrossRef]
- D.F. Hayes, Serum (circulating) tumor markers for breast cancer, Recent results in cancer research. Fortschritte der Krebsforschung. Progres dans les recherches sur le cancer 140 (1996) 101-13.
- Beckwith, D.M.; Cudic, M. Tumor-associated O-glycans of MUC1: Carriers of the glyco-code and targets for cancer vaccine design. Semin. Immunol. 2020, 47, 101389–101389. [CrossRef]
- Beatson, R.; Tajadura-Ortega, V.; Achkova, D.; Picco, G.; Tsourouktsoglou, T.-D.; Klausing, S.; Hillier, M.; Maher, D.A.J.; Noll, S.K.T.; Crocker, P.R.; et al. The mucin MUC1 modulates the tumor immunological microenvironment through engagement of the lectin Siglec-9. Nat. Immunol. 2016, 17, 1273–1281. [CrossRef]
- Liu, F.; Fu, J.; Bergstrom, K.; Shan, X.; McDaniel, J.M.; McGee, S.; Bai, X.; Chen, W.; Xia, L. Core 1–derived mucin-type O-glycosylation protects against spontaneous gastritis and gastric cancer. J. Exp. Med. 2019, 217. [CrossRef]
- Baldus, S.E.; Zirbes, T.K.; Engel, S.; Hanisch, F.-G.; Mönig, S.P.; Lorenzen, J.; Glossmann, J.; Fromm, S.; Thiele, J.; Pichlmaier, H.; et al. Correlation of the immunohistochemical reactivity of mucin peptide cores MUC1 and MUC2 with the histopathological subtype and prognosis of gastric carcinomas. Int. J. Cancer 1998, 79, 133–138. [CrossRef]
- Carvalho, F.; Seruca, R.; David, L.; Amorim, A.; Seixas, M.; Bennett, E.; Clausen, H.; Sobrinho-Simoes, M. MUC1 gene polymorphism and gastric cancer–an epidemiological study. Glycoconj. J. 1997, 14, 107–111. [CrossRef]
- Malaby, H.L.H.; Kobertz, W.R. Molecular determinants of co- and post-translational N-glycosylation of type I transmembrane peptides. Biochem. J. 2013, 453, 427–434. [CrossRef]
- Taniguchi, T.; Woodward, A.M.; Magnelli, P.; McColgan, N.M.; Lehoux, S.; Jacobo, S.M.P.; Mauris, J.; Argüeso, P. N-Glycosylation affects the stability and barrier function of the MUC16 mucin. J. Biol. Chem. 2017, 292, 11079–11090. [CrossRef]
- Bell, S.L.; Xu, G.; Khatri, I.A.; Wang, R.; Rahman, S.; Forstner, J.F. N-linked oligosaccharides play a role in disulphide-dependent dimerization of intestinal mucin Muc2. Biochem. J. 2003, 373, 893–900. [CrossRef]
- Asker, N.; Axelsson, M.A.B.; Olofsson, S.-O.; Hansson, G.C. Dimerization of the Human MUC2 Mucin in the Endoplasmic Reticulum Is Followed by a N-Glycosylation-dependent Transfer of the Mono- and Dimers to the Golgi Apparatus. J. Biol. Chem. 1998, 273, 18857–18863. [CrossRef]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [CrossRef]
- T. Lange, S. Ullrich, I. Müller, M.F. Nentwich, K. Stübke, S. Feldhaus, C. Knies, O.J. Hellwinkel, R.L. Vessella, C. Abramjuk, M. Anders, J. Schröder-Schwarz, T. Schlomm, H. Huland, G. Sauter, U. Schumacher, Human prostate cancer in a clinically relevant xenograft mouse model: identification of β(1,6)-branched oligosaccharides as a marker of tumor progression, Clinical cancer research : an official journal of the American Association for Cancer Research 18(5) (2012) 1364-73.
- Gubbels, J.A.; Belisle, J.; Onda, M.; Rancourt, C.; Migneault, M.; Ho, M.; Bera, T.K.; Connor, J.; Sathyanarayana, B.K.; Lee, B.; et al. Mesothelin-MUC16 binding is a high affinity, N-glycan dependent interaction that facilitates peritoneal metastasis of ovarian tumors. Mol. Cancer 2006, 5, 1–50. [CrossRef]
- Liu, Y.; Lan, L.; Li, Y.; Lu, J.; He, L.; Deng, Y.; Fei, M.; Lu, J.-W.; Shangguan, F.; Lu, J.-P.; et al. N-glycosylation stabilizes MerTK and promotes hepatocellular carcinoma tumor growth. Redox Biol. 2022, 54, 102366. [CrossRef]
- Zhu, L.; Chen, Y.; Du, H.; Cong, Y.; Yan, W.; Ma, K.; Huang, X. N-glycosylation of CD82 at Asn157 is required for suppressing migration and invasion by reversing EMT via Wnt/β-catenin pathway in colon cancer. Biochem. Biophys. Res. Commun. 2022, 629, 121–127. [CrossRef]
- Varki, A. Factors controlling the glycosylation potential of the Golgi apparatus. Trends Cell Biol. 1998, 8, 34–40. [CrossRef]
- Scott, E.; Hodgson, K.; Calle, B.; Turner, H.; Cheung, K.; Bermudez, A.; Marques, F.J.G.; Pye, H.; Yo, E.C.; Islam, K.; et al. Upregulation of GALNT7 in prostate cancer modifies O-glycosylation and promotes tumour growth. Oncogene 2023, 42, 926–937. [CrossRef]
- M. Prorok-Hamon, F. Notel, S. Mathieu, C. Langlet, M. Fukuda, A. El-Battari, N-glycans of core2 beta(1,6)-N-acetylglucosaminyltransferase-I (C2GnT-I) but not those of alpha(1,3)-fucosyltransferase-VII (FucT-VII) are required for the synthesis of functional P-selectin glycoprotein ligand-1 (PSGL-1): effects on P-, L- and E-selectin binding, The Biochemical journal 391(Pt 3) (2005) 491-502.
- Ruggiero, F.M.; Vilcaes, A.A.; Iglesias-Bartolomé, R.; Daniotti, J.L. Critical role of evolutionarily conserved glycosylation at Asn211 in the intracellular trafficking and activity of sialyltransferase ST3Gal-II. Biochem. J. 2015, 469, 83–95. [CrossRef]
- Skrincosky, D.; Kain, R.; El-Battari, A.; Exner, M.; Kerjaschki, D.; Fukuda, M. Altered Golgi Localization of Core 2 β-1,6-N-Acetylglucosaminyltransferase Leads to Decreased Synthesis of Branched O-Glycans. J. Biol. Chem. 1997, 272, 22695–22702. [CrossRef]
- G. Egea, C. Francí, G. Gambús, T. Lesuffleur, A. Zweibaum, F.X. Real, cis-Golgi resident proteins and O-glycans are abnormally compartmentalized in the RER of colon cancer cells, Journal of cell science 105 ( Pt 3) (1993) 819-30.
- Rivinoja, A.; Hassinen, A.; Kokkonen, N.; Kauppila, A.; Kellokumpu, S. Elevated Golgi pH impairs terminalN-glycosylation by inducing mislocalization of Golgi glycosyltransferases. J. Cell. Physiol. 2009, 220, 144–154. [CrossRef]
- Rivinoja, A.; Kokkonen, N.; Kellokumpu, I.; Kellokumpu, S. Elevated Golgi pH in breast and colorectal cancer cells correlates with the expression of oncofetal carbohydrate T-antigen. J. Cell. Physiol. 2006, 208, 167–174. [CrossRef]
- Axelsson, M.A.; Karlsson, N.G.; Steel, D.M.; Ouwendijk, J.; Nilsson, T.; Hansson, G.C. Neutralization of pH in the Golgi apparatus causes redistribution of glycosyltransferases and changes in the O-glycosylation of mucins. Glycobiology 2001, 11, 633–644. [CrossRef]
- B.J. Campbell, G.E. Rowe, K. Leiper, J.M. Rhodes, Increasing the intra-Golgi pH of cultured LS174T goblet-differentiated cells mimics the decreased mucin sulfation and increased Thomsen-Friedenreich antigen (Gal beta1-3GalNac alpha-) expression seen in colon cancer, Glycobiology 11(5) (2001) 385-93.
- Hadley, B.; Maggioni, A.; Ashikov, A.; Day, C.J.; Haselhorst, T.; Tiralongo, J. Structure and function of nucleotide sugar transporters: Current progress. Comput. Struct. Biotechnol. J. 2014, 10, 23–32. [CrossRef]
- Nji, E.; Gulati, A.; Qureshi, A.A.; Coincon, M.; Drew, D. Structural basis for the delivery of activated sialic acid into Golgi for sialyation. Nat. Struct. Mol. Biol. 2019, 26, 415–423. [CrossRef]
- Takeshima-Futagami, T.; Sakaguchi, M.; Uehara, E.; Aoki, K.; Ishida, N.; Sanai, Y.; Sugahara, Y.; Kawakita, M. Amino acid residues important for CMP-sialic acid recognition by the CMP-sialic acid transporter: analysis of the substrate specificity of UDP-galactose/CMP-sialic acid transporter chimeras. Glycobiology 2012, 22, 1731–1740. [CrossRef]
- Bhatia, R.; Gautam, S.K.; Cannon, A.; Thompson, C.; Hall, B.R.; Aithal, A.; Banerjee, K.; Jain, M.; Solheim, J.C.; Kumar, S.; et al. Cancer-associated mucins: role in immune modulation and metastasis. Cancer Metastasis Rev. 2019, 38, 223–236. [CrossRef]
- S.J. Gendler, MUC1, the renaissance molecule, Journal of mammary gland biology and neoplasia 6(3) (2001) 339-53.
- Burchell, J.M.; Mungul, A.; Taylor-Papadimitriou, J. O-Linked Glycosylation in the Mammary Gland: Changes that Occur During Malignancy. J. Mammary Gland. Biol. Neoplasia 2001, 6, 355–364. [CrossRef]
- Mungul, A.; Cooper, L.; Brockhausen, I.; Ryder, K.; Mandel, U.; Clausen, H.; Rughetti, A.; Miles, D.W.; Taylor-Papadimitriou, J.; Burchell, J.M. Sialylated core 1 based O-linked glycans enhance the growth rate of mammary carcinoma cells in MUC1 transgenic mice.. Int. J. Oncol. 2004, 25.
- Chou, C.-H.; Huang, M.-J.; Chen, C.-H.; Shyu, M.-K.; Huang, J.; Hung, J.-S.; Huang, C.-S.; Huang, M.-C. Up-regulation of C1GALT1 promotes breast cancer cell growth through MUC1-C signaling pathway. Oncotarget 2015, 6, 6123–6135. [CrossRef]
- Mao, Y.; Zhang, Y.; Fan, S.; Chen, L.; Tang, L.; Chen, X.; Lyu, J. GALNT6 Promotes Tumorigenicity and Metastasis of Breast Cancer Cell via β-catenin/MUC1-C Signaling Pathway. Int. J. Biol. Sci. 2019, 15, 169–182. [CrossRef]
- Z. Chen, Z.G. Gulzar, C.A. St Hill, B. Walcheck, J.D. Brooks, Increased expression of GCNT1 is associated with altered O-glycosylation of PSA, PAP, and MUC1 in human prostate cancers, The Prostate 74(10) (2014) 1059-67.
- Premaratne, P.; Welén, K.; Damber, J.-E.; Hansson, G.C.; Bäckström, M. O-glycosylation of MUC1 mucin in prostate cancer and the effects of its expression on tumor growth in a prostate cancer xenograft model. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2010, 32, 203–213. [CrossRef]
- Honn, K.V.; Tang, D.G. Adhesion molecules and tumor cell interaction with endothelium and subendothelial matrix. Cancer Metastasis Rev. 1992, 11, 353–375. [CrossRef]
- Paschos, K.A.; Canovas, D.; Bird, N.C. The role of cell adhesion molecules in the progression of colorectal cancer and the development of liver metastasis. Cell. Signal. 2009, 21, 665–674. [CrossRef]
- Burdick, M.D.; Harris, A.; Reid, C.J.; Iwamura, T.; Hollingsworth, M.A. Oligosaccharides Expressed on MUC1 Produced by Pancreatic and Colon Tumor Cell Lines. J. Biol. Chem. 1997, 272, 24198–24202. [CrossRef]
- Fernandez-Rodriguez, J.; Dwir, O.; Alon, R.; Hansson, G.C. Tumor cell MUC1 and CD43 are glycosylated differently with sialyl-Lewis a and x epitopes and show variable interactions with E-selectin under physiological flow conditions.. Glycoconj. J. 2001, 18, 925–930. [CrossRef]
- Solatycka, T. Owczarek, F. Piller, V. Piller, B. Pula, L. Wojciech, M. Podhorska-Okolow, P. Dziegiel, M. Ugorski, MUC1 in human and murine mammary carcinoma cells decreases the expression of core 2 β1,6-N-acetylglucosaminyltransferase and β-galactoside α2,3-sialyltransferase, Glycobiology 22(8) (2012) 1042-54.
- J.H. Park, T. Nishidate, K. Kijima, T. Ohashi, K. Takegawa, T. Fujikane, K. Hirata, Y. Nakamura, T. Katagiri, Critical roles of mucin 1 glycosylation by transactivated polypeptide N-acetylgalactosaminyltransferase 6 in mammary carcinogenesis, Cancer research 70(7) (2010) 2759-69.
- Zhang, L.; Gallup, M.; Zlock, L.; Chen, Y.T.F.; E Finkbeiner, W.; A McNamara, N. Pivotal role of MUC1 glycosylation by cigarette smoke in modulating disruption of airway adherens junctions in vitro. J. Pathol. 2014, 234, 60–73. [CrossRef]
- M. Amado, F. Carneiro, M. Seixas, H. Clausen, M. Sobrinho-Simões, Dimeric sialyl-Le(x) expression in gastric carcinoma correlates with venous invasion and poor outcome, Gastroenterology 114(3) (1998) 462-70.
- Rambaruth, N.D.; Dwek, M.V. Cell surface glycan–lectin interactions in tumor metastasis. Acta Histochem. 2011, 113, 591–600. [CrossRef]
- L. Borsig, Selectins in cancer immunity, Glycobiology 28(9) (2018) 648-655.
- Nath, D.; Hartnell, A.; Happerfield, L.; Miles, D.W.; Burchell, J.; Papadimitriou, T.; Crocker, P.R. Macrophage-tumour cell interactions: identification of MUC1 on breast cancer cells as a potential counter-receptor for the macrophage-restricted receptor, sialoadhesin. Immunology 1999, 98, 213–219. [CrossRef]
- Q. Zhao, X. Guo, G.B. Nash, P.C. Stone, J. Hilkens, J.M. Rhodes, L.G. Yu, Circulating galectin-3 promotes metastasis by modifying MUC1 localization on cancer cell surface, Cancer research 69(17) (2009) 6799-806.
- Piyush, T.; Rhodes, J.M.; Yu, L.-G. MUC1 O-glycosylation contributes to anoikis resistance in epithelial cancer cells. Cell Death Discov. 2017, 3, 17044. [CrossRef]
- X. Jiang, J. Wang, X. Deng, F. Xiong, J. Ge, B. Xiang, X. Wu, J. Ma, M. Zhou, X. Li, Y. Li, G. Li, W. Xiong, C. Guo, Z. Zeng, Role of the tumor microenvironment in PD-L1/PD-1-mediated tumor immune escape, Molecular cancer 18(1) (2019) 10.
- Rabinovich, G.A.; Toscano, M.A. Turning 'sweet' on immunity: galectin–glycan interactions in immune tolerance and inflammation. Nat. Rev. Immunol. 2009, 9, 338–352. [CrossRef]
- C.B. Madsen, K. Lavrsen, C. Steentoft, M.B. Vester-Christensen, H. Clausen, H.H. Wandall, A.E. Pedersen, Glycan elongation beyond the mucin associated Tn antigen protects tumor cells from immune-mediated killing, PloS one 8(9) (2013) e72413.
- de Martel, C.; Georges, D.; Bray, F.; Ferlay, J.; Clifford, G.M. Global burden of cancer attributable to infections in 2018: A worldwide incidence analysis. Lancet Glob. Health 2020, 8, e180–e190. [CrossRef]
- Kashyap, P.C.; Marcobal, A.; Ursell, L.K.; Smits, S.A.; Sonnenburg, E.D.; Costello, E.K.; Higginbottom, S.K.; Domino, S.E.; Holmes, S.P.; Relman, D.A.; et al. Genetically dictated change in host mucus carbohydrate landscape exerts a diet-dependent effect on the gut microbiota. Proc. Natl. Acad. Sci. 2013, 110, 17059–17064. [CrossRef]
- Lin, B.; Qing, X.; Liao, J.; Zhuo, K. Role of Protein Glycosylation in Host-Pathogen Interaction. Cells 2020, 9, 1022. [CrossRef]
- Owen, C.D.; Tailford, L.E.; Monaco, S.; Šuligoj, T.; Vaux, L.; Lallement, R.; Khedri, Z.; Yu, H.; Lecointe, K.; Walshaw, J.; et al. Unravelling the specificity and mechanism of sialic acid recognition by the gut symbiont Ruminococcus gnavus. Nat. Commun. 2017, 8, 1–15. [CrossRef]
- D.M. Hardbower, R.M. Peek, Jr., K.T. Wilson, At the Bench: Helicobacter pylori, dysregulated host responses, DNA damage, and gastric cancer, Journal of leukocyte biology 96(2) (2014) 201-12.
- Magalhães, A.; Rossez, Y.; Robbe-Masselot, C.; Maes, E.; Gomes, J.; Shevtsova, A.; Bugaytsova, J.; Borén, T.; Reis, C.A. Muc5ac gastric mucin glycosylation is shaped by FUT2 activity and functionally impacts Helicobacter pylori binding. Sci. Rep. 2016, 6, 25575–25575. [CrossRef]
- Skoog, E.C.; Padra, M.; Åberg, A.; Gideonsson, P.; Obi, I.; Quintana-Hayashi, M.P.; Arnqvist, A.; Lindén, S.K. BabA dependent binding of Helicobacter pylori to human gastric mucins cause aggregation that inhibits proliferation and is regulated via ArsS. Sci. Rep. 2017, 7, srep40656. [CrossRef]
- C. Solórzano, M. Angel Mayoral, M. de los Angeles Carlos, J. Berumen, J. Guevara, F. Raúl Chávez, G. Mendoza-Hernández, C. Agundis, E. Zenteno, Overexpression of glycosylated proteins in cervical cancer recognized by the Machaerocereus eruca agglutinin, Folia histochemica et cytobiologica 50(3) (2012) 398-406.
- W. Ahmad, K. Shabbiri, B. Ijaz, S. Asad, N. Nazar, S. Nazar, K. Fouzia, H. Kausar, S. Gull, M.T. Sarwar, I. Shahid, S. Hassan, Serine 204 phosphorylation and O-β-GlcNAC interplay of IGFBP-6 as therapeutic indicator to regulate IGF-II functions in viral mediated hepatocellular carcinoma, Virology journal 8 (2011) 208.
- M.P. Thompson, R. Kurzrock, Epstein-Barr virus and cancer, Clinical cancer research : an official journal of the American Association for Cancer Research 10(3) (2004) 803-21.
- Liu, J.; Zhang, Y.; Liu, W.; Zhang, Q.; Xiao, H.; Song, H.; Luo, B. MiR-BART1-5p targets core 2β-1,6-acetylglucosaminyltransferase GCNT3 to inhibit cell proliferation and migration in EBV-associated gastric cancer. Virology 2019, 541, 63–74. [CrossRef]
- Kondo, S.; Yoshizaki, T.; Wakisaka, N.; Horikawa, T.; Murono, S.; Jang, K.L.; Joab, I.; Furukawa, M.; Pagano, J.S. MUC1 Induced by Epstein-Barr Virus Latent Membrane Protein 1 Causes Dissociation of the Cell-Matrix Interaction and Cellular Invasiveness via STAT Signaling. J. Virol. 2007, 81, 1554–1562. [CrossRef]
- Springer, G.F. T and Tn, General Carcinoma Autoantigens. Science 1984, 224, 1198–1206. [CrossRef]
- Fuster, M.M.; Esko, J.D. The Sweet and Sour of Cancer: Glycans As Novel Therapeutic Targets. Nat. Rev. Cancer 2005, 5, 526–542. [CrossRef]
- Julien, S.; Picco, G.; Sewell, R.; Vercoutter-Edouart, A.-S.; Tarp, M.; Miles, D.; Clausen, H.; Taylor-Papadimitriou, J.; Burchell, J.M. Sialyl-Tn vaccine induces antibody-mediated tumour protection in a relevant murine model. Br. J. Cancer 2009, 100, 1746–1754. [CrossRef]
- Taylor-Papadimitriou, J.; Peterson, J.A.; Arklie, J.; Burchell, J.; Ceriani, R.L.; Bodmer, W.F. Monoclonal antibodies to epithelium-specific components of the human milk fat globule membrane: Production and reaction with cells in culture. Int. J. Cancer 1981, 28, 17–21. [CrossRef]
- Gendler, S.; Lancaster, C.; Taylor-Papadimitriou, J.; Duhig, T.; Peat, N.; Burchell, J.; Pemberton, L.; Lalani, E.; Wilson, D. Molecular cloning and expression of human tumor-associated polymorphic epithelial mucin.. J. Biol. Chem. 1990, 265, 15286–15293. [CrossRef]
- Tarp, M.A.; Clausen, H. Mucin-type O-glycosylation and its potential use in drug and vaccine development. Biochim. et Biophys. Acta (BBA) - Gen. Subj. 2008, 1780, 546–563. [CrossRef]
- Wilkinson, R.W.; Ross, E.L.; E Lee-MacAry, A.; Laylor, R.; Burchell, J.; Taylor-Papadimitriou, J.; Snary, D. A transgenic mouse model for tumour immunotherapy: induction of an anti-idiotype response to human MUC1. Br. J. Cancer 2000, 83, 1202–1208. [CrossRef]
- Moore, Z. Medarova, A. Potthast, G. Dai, In vivo targeting of underglycosylated MUC-1 tumor antigen using a multimodal imaging probe, Cancer research 64(5) (2004) 1821-7.
- Singh, R.; Bandyopadhyay, D. MUC1: A target molecule for cancer therapy. Cancer Biol. Ther. 2007, 6, 481–486. [CrossRef]
- M.A. Cheever, J.P. Allison, A.S. Ferris, O.J. Finn, B.M. Hastings, T.T. Hecht, I. Mellman, S.A. Prindiville, J.L. Viner, L.M. Weiner, L.M. Matrisian, The prioritization of cancer antigens: a national cancer institute pilot project for the acceleration of translational research, Clinical cancer research : an official journal of the American Association for Cancer Research 15(17) (2009) 5323-37.
- H.H. Wandall, O. Blixt, M.A. Tarp, J.W. Pedersen, E.P. Bennett, U. Mandel, G. Ragupathi, P.O. Livingston, M.A. Hollingsworth, J. Taylor-Papadimitriou, J. Burchell, H. Clausen, Cancer biomarkers defined by autoantibody signatures to aberrant O-glycopeptide epitopes, Cancer research 70(4) (2010) 1306-13.
- Burchell, J.; Gendler, S.; Taylor-Papadimitriou, J.; Girling, A.; Lewis, A.; Millis, R.; Lamport, D. Development and characterization of breast cancer reactive monoclonal antibodies directed to the core protein of the human milk mucin.. Cancer Res. 1987, 47, 5476–82.
- Burchell, J.; Taylor-Papadimitriou, J.; Boshell, M.; Gendler, S.; Duhig, T. A short sequence, within the amino acid tandem repeat of a cancer-associated mucin, contains immunodominant epitopes. Int. J. Cancer 1989, 44, 691–696. [CrossRef]
- Rokhlin, O.W.; Weiner, G.J.; Cohen, M.B. 5E10: a prostate-specific surface-reactive monoclonal antibody. Cancer Lett. 1998, 131, 129–136. [CrossRef]
- Gong, Y.; Klein Wolterink, R.G.; Gulaia, V.; Cloosen, S.; Ehlers, F.A.; Wieten, L.; Graus, Y.F.; Bos, G.M.J.; Germeraad, W.T.W. Defucosylation of Tumor-Specific Humanized Anti-MUC1 Monoclonal Antibody Enhances NK Cell-Mediated Anti-Tumor Cell Cytotoxicity. Cancers 2021, 13, 2579. [CrossRef]
- J. Macías-León, I.A. Bermejo, A. Asín, A. García-García, I. Compañón, E. Jiménez-Moreno, H. Coelho, V. Mangini, I.S. Albuquerque, F. Marcelo, J.L. Asensio, G.J.L. Bernardes, H.J. Joshi, R. Fiammengo, O. Blixt, R. Hurtado-Guerrero, F. Corzana, Structural characterization of an unprecedented lectin-like antitumoral anti-MUC1 antibody, Chemical communications (Cambridge, England) 56(96) (2020) 15137-15140.
- Sørensen, A.L.; Reis, C.A.; Tarp, M.A.; Mandel, U.; Ramachandran, K.; Sankaranarayanan, V.; Schwientek, T.; Graham, R.; Taylor-Papadimitriou, J.; Hollingsworth, M.A.; et al. Chemoenzymatically synthesized multimeric Tn/STn MUC1 glycopeptides elicit cancer-specific anti-MUC1 antibody responses and override tolerance. Glycobiology 2005, 16, 96–107. [CrossRef]
- Maleki, F.; Rezazadeh, F.; Varmira, K. MUC1-Targeted Radiopharmaceuticals in Cancer Imaging and Therapy. Mol. Pharm. 2021, 18, 1842–1861. [CrossRef]
- M. Bose, A. Sanders, C. De, R. Zhou, P. Lala, S. Shwartz, B. Mitra, C. Brouwer, P. Mukherjee, Targeting tumor-associated MUC1 overcomes anoikis-resistance in pancreatic cancer, Translational research : the journal of laboratory and clinical medicine 253 (2023) 41-56.
- Wu, G.; Kim, D.; Kim, J.N.; Park, S.; Maharjan, S.; Koh, H.; Moon, K.; Lee, Y.; Kwon, H.-J. A Mucin1 C-terminal Subunit-directed Monoclonal Antibody Targets Overexpressed Mucin1 in Breast Cancer. Theranostics 2018, 8, 78–91. [CrossRef]
- Wu, G.; Li, L.; Liu, M.; Chen, C.; Wang, G.; Jiang, Z.; Qin, Y.; He, L.; Li, H.; Cao, J.; et al. Therapeutic effect of a MUC1-specific monoclonal antibody-drug conjugates against pancreatic cancer model. Cancer Cell Int. 2022, 22, 1–14. [CrossRef]
- Okarvi, S. Peptide-based radiopharmaceuticals and cytotoxic conjugates: Potential tools against cancer. Cancer Treat. Rev. 2008, 34, 13–26. [CrossRef]
- Bolzati, C.; Refosco, F.; Marchiani, A.; Ruzza, P. 99mTc-Radiolabelled Peptides for Tumour Imaging: Present and Future. Curr. Med. Chem. 2010, 17, 2656–2683. [CrossRef]
- Salouti, M.; Rajabi, H.; Babaei, M.H.; Rasaee, M.J. Breast tumor targeting with 99mTc-HYNIC-PR81 complex as a new biologic radiopharmaceutical. Nucl. Med. Biol. 2008, 35, 763–768. [CrossRef]
- Carmo, F.S.D.; Ricci-Junior, E.; Cerqueira-Coutinho, C.; Albernaz, M.D.S.; Bernardes, E.S.; Missailidis, S.; Santos-Oliveira, R. Anti-MUC1 nano-aptamers for triple-negative breast cancer imaging by single-photon emission computed tomography in inducted animals: initial considerations. Int. J. Nanomed. 2016, ume 12, 53–60. [CrossRef]
- Al Jammaz, I.; Al-Otaibi, B.; Al-Malki, Y.; Abousekhrah, A.; Okarvi, S.M. Fast Fluorine-18 labeling and preclinical evaluation of novel Mucin1 and its Folate hybrid peptide conjugate for targeting breast carcinoma. EJNMMI Radiopharm. Chem. 2021, 6, 1–17. [CrossRef]
- Nabavinia, M.S.; Gholoobi, A.; Charbgoo, F.; Nabavinia, M.; Ramezani, M.; Abnous, K. Anti-MUC1 aptamer: A potential opportunity for cancer treatment. Med. Res. Rev. 2017, 37, 1518–1539. [CrossRef]
- Sgouros, G.; Bodei, L.; McDevitt, M.R.; Nedrow, J.R. Radiopharmaceutical therapy in cancer: clinical advances and challenges. Nat. Rev. Drug Discov. 2020, 19, 589–608. [CrossRef]
- Sun, Q.; Li, J.; Ding, Z.; Liu, Z. Radiopharmaceuticals heat anti-tumor immunity. Theranostics 2023, 13, 767–786. [CrossRef]
- Lin, A.; Giuliano, C.J.; Palladino, A.; John, K.M.; Abramowicz, C.; Yuan, M.L.; Sausville, E.L.; Lukow, D.A.; Liu, L.; Chait, A.R.; et al. Off-target toxicity is a common mechanism of action of cancer drugs undergoing clinical trials. Sci. Transl. Med. 2019, 11, eaaw8412. [CrossRef]
- Lakshminarayanan, V.; Thompson, P.; Wolfert, M.A.; Buskas, T.; Bradley, J.M.; Pathangey, L.B.; Madsen, C.S.; Cohen, P.A.; Gendler, S.J.; Boons, G.-J. Immune recognition of tumor-associated mucin MUC1 is achieved by a fully synthetic aberrantly glycosylated MUC1 tripartite vaccine. Proc. Natl. Acad. Sci. 2011, 109, 261–266. [CrossRef]
- T. Gao, Q. Cen, H. Lei, A review on development of MUC1-based cancer vaccine, Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie 132 (2020) 110888.
- Pinkhasov, J.; Alvarez, M.L.; Pathangey, L.B.; Tinder, T.L.; Mason, H.S.; Walmsley, A.M.; Gendler, S.J.; Mukherjee, P. Analysis of a cholera toxin B subunit (CTB) and human mucin 1 (MUC1) conjugate protein in a MUC1-tolerant mouse model. Cancer Immunol. Immunother. 2010, 59, 1801–1811. [CrossRef]
- Rong, Y.; Jin, D.; Wu, W.; Lou, W.; Wang, D.; Kuang, T.; Ni, X.; Qin, X. Induction of protective and therapeutic anti-pancreatic cancer immunity using a reconstructed MUC1 DNA vaccine. BMC Cancer 2009, 9, 191–191. [CrossRef]
- Quoix, E.; Lena, H.; Losonczy, G.; Forget, F.; Chouaid, C.; Papai, Z.; Gervais, R.; Ottensmeier, C.; Szczesna, A.; Kazarnowicz, A.; et al. TG4010 immunotherapy and first-line chemotherapy for advanced non-small-cell lung cancer (TIME): results from the phase 2b part of a randomised, double-blind, placebo-controlled, phase 2b/3 trial. Lancet Oncol. 2015, 17, 212–223. [CrossRef]
- Teramoto, K.; Ozaki, Y.; Hanaoka, J.; Sawai, S.; Tezuka, N.; Fujino, S.; Daigo, Y.; Kontani, K. Predictive biomarkers and effectiveness of MUC1-targeted dendritic-cell-based vaccine in patients with refractory non-small cell lung cancer. Ther. Adv. Med Oncol. 2016, 9, 147–157. [CrossRef]
- Palitzsch, B.; Gaidzik, N.; Stergiou, N.; Stahn, S.; Hartmann, S.; Gerlitzki, B.; Teusch, N.; Flemming, P.-D.D.P.; Schmitt, E.; Kunz, H. A Synthetic Glycopeptide Vaccine for the Induction of a Monoclonal Antibody that Differentiates between Normal and Tumor Mammary Cells and Enables the Diagnosis of Human Pancreatic Cancer. Angew. Chem. Int. Ed. 2016, 55, 2894–2898. [CrossRef]
- M. Sadelain, R. Brentjens, I. Rivière, The basic principles of chimeric antigen receptor design, Cancer discovery 3(4) (2013) 388-98.
- Zhou, R.; Yazdanifar, M.; Das Roy, L.; Whilding, L.M.; Gavrill, A.; Maher, J.; Mukherjee, P. CAR T Cells Targeting the Tumor MUC1 Glycoprotein Reduce Triple-Negative Breast Cancer Growth. Front. Immunol. 2019, 10, 1149. [CrossRef]
- Stanley, P.; Ad, P.; Rd, S.; Ac, B.; C, S.; U, M.; B, E.; Jd, S.; Td, M.; K, S.; et al. Faculty Opinions recommendation of Engineered CAR T Cells Targeting the Cancer-Associated Tn-Glycoform of the Membrane Mucin MUC1 Control Adenocarcinoma.. 2016, 44. [CrossRef]
- Tchou, J.; Zhao, Y.; Levine, B.L.; Zhang, P.J.; Davis, M.M.; Melenhorst, J.J.; Kulikovskaya, I.; Brennan, A.L.; Liu, X.; Lacey, S.F.; et al. Safety and Efficacy of Intratumoral Injections of Chimeric Antigen Receptor (CAR) T Cells in Metastatic Breast Cancer. Cancer Immunol. Res. 2017, 5, 1152–1161. [CrossRef]
- Mao, L.; Su, S.; Li, J.; Yu, S.; Gong, Y.; Chen, C.; Hu, Z.; Huang, X. Development of Engineered CAR T Cells Targeting Tumor-Associated Glycoforms of MUC1 for the Treatment of Intrahepatic Cholangiocarcinoma. J. Immunother. 2023, 46, 89–95. [CrossRef]
- Zhang, H.; Zhao, H.; He, X.; Xi, F.; Liu, J. JAK-STAT Domain Enhanced MUC1-CAR-T Cells Induced Esophageal Cancer Elimination. Cancer Manag. Res. 2020, ume 12, 9813–9824. [CrossRef]
- Weimin, S.; Abula, A.; Qianghong, D.; Wenguang, W. Chimeric cytokine receptor enhancing PSMA-CAR-T cell-mediated prostate cancer regression. Cancer Biol. Ther. 2020, 21, 570–580. [CrossRef]
- E. Grishman, Histochemical analysis of mucopolysaccharides occurring in mucus-producing tumors; mixed tumors of the parotid gland, colloid carcinomas of the breast, and myxomas, Cancer 5(4) (1952) 700-7.
- Mei, Z.; Zhang, K.; Lam, A.K.; Huang, J.; Qiu, F.; Qiao, B.; Zhang, Y. MUC1 as a target for CAR-T therapy in head and neck squamous cell carinoma. Cancer Med. 2019, 9, 640–652. [CrossRef]
Figure 1.
Transmembrane mucin and secreted mucin. Transmembrane mucin has a C-terminal cytoplasmic tail, a transmembrane domain (TM) and an extracellular structure, a SEA domain, a VWD domain and a PTS domain. Secreted mucin does not have a TM, which consists of a large number of VWD structural domains, followed by PTS structural domain and “CK” domain.
Figure 1.
Transmembrane mucin and secreted mucin. Transmembrane mucin has a C-terminal cytoplasmic tail, a transmembrane domain (TM) and an extracellular structure, a SEA domain, a VWD domain and a PTS domain. Secreted mucin does not have a TM, which consists of a large number of VWD structural domains, followed by PTS structural domain and “CK” domain.
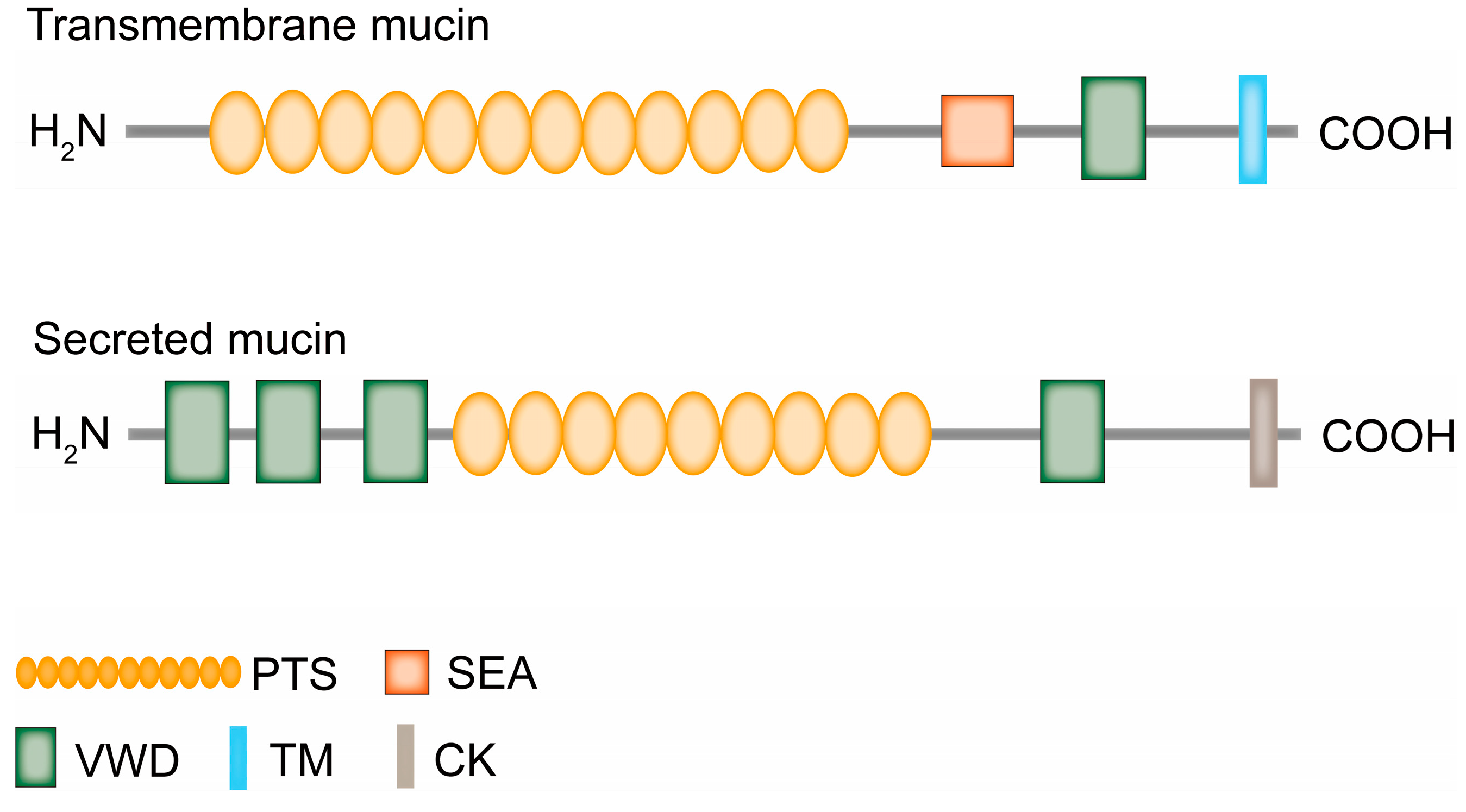
Figure 2.
Glycans in mammals. The PTS domain of mucin is rich in Pro, Ser and Thr residues. There are densely arranged O-glycan scaffolds on the PTS domain, which account for 80% of the total mass of mucin and help maintain a high viscosity state and rigid structure of mucin.
Figure 2.
Glycans in mammals. The PTS domain of mucin is rich in Pro, Ser and Thr residues. There are densely arranged O-glycan scaffolds on the PTS domain, which account for 80% of the total mass of mucin and help maintain a high viscosity state and rigid structure of mucin.
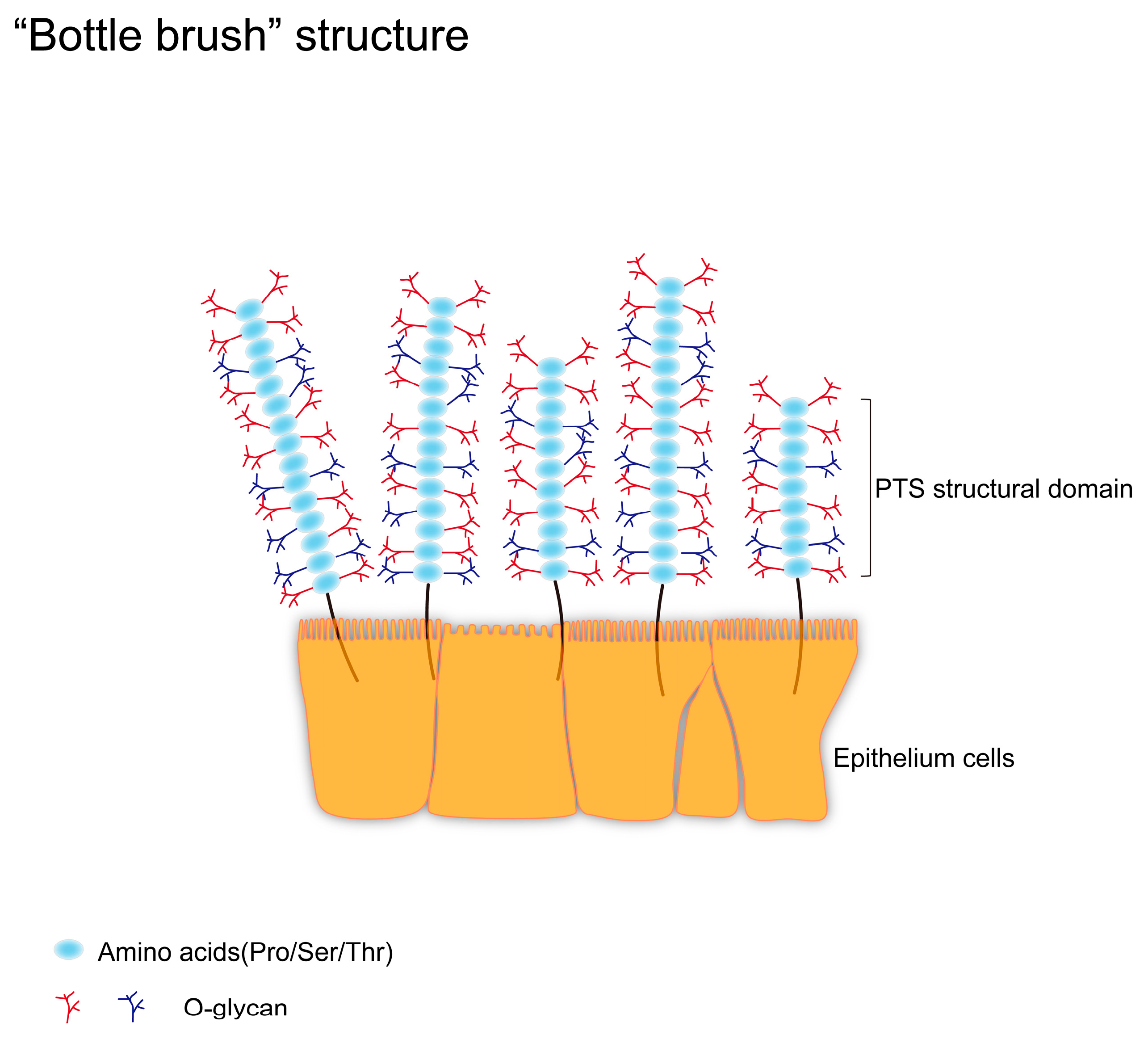
Figure 5.
Glycans in mammals. Normal mucins have intact glycan chains on them that are involved in maintaining the morphology, structure and function of epithelial cells. Abnormal alterations in glycosylation on mucins may lead to cellular carcinogenesis. A large number of truncated O-glycans (Tn, T, ST, STn) and extended O-glycans with core 2 structure (SLea, SLex) are present on cancer cells. Among them, the SLea and SLex structures lead to the detachment of cancer cells from the primary lesion and metastasis. The glycan structure is represented according to the SNFG format[40].
Figure 5.
Glycans in mammals. Normal mucins have intact glycan chains on them that are involved in maintaining the morphology, structure and function of epithelial cells. Abnormal alterations in glycosylation on mucins may lead to cellular carcinogenesis. A large number of truncated O-glycans (Tn, T, ST, STn) and extended O-glycans with core 2 structure (SLea, SLex) are present on cancer cells. Among them, the SLea and SLex structures lead to the detachment of cancer cells from the primary lesion and metastasis. The glycan structure is represented according to the SNFG format[40].
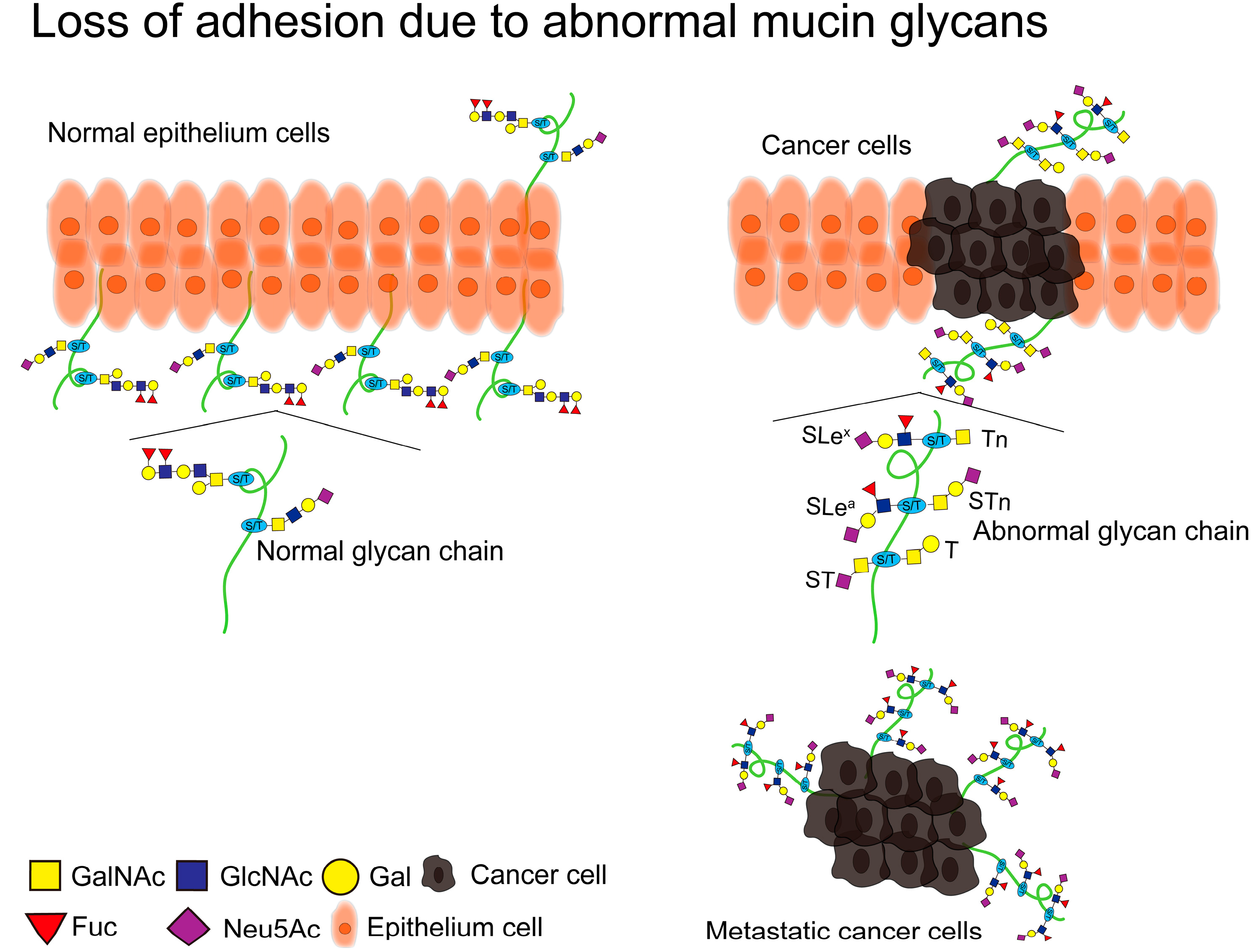
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



