Submitted:
18 September 2023
Posted:
19 September 2023
You are already at the latest version
Abstract
Pulmonary infections with Mycobacterium avium occur in susceptible individuals following exposure to the bacterium in the environment, where it often persists in biofilms. Many methods have been used to generate biofilms of M. avium, and it is unknown whether different approaches generate similar structures and cell phenotypes. To make a parallel comparison of in vitro biofilm ultrastructure, extracellular matrix (ECM) composition, and drug susceptibility of biofilm resident bacteria, we used two published methods to generate M. avium biofilms: four week incubation in M63 medium or 24 hour exposure to dithiothreitol (DTT). Scanning electron microscopy revealed differences in biofilm ultrastructure between the two methods, including variation in appearance of ECM materials and morphology of resident cells, while light microscopy and staining with calcofluor white indicated that both biofilms contained polysaccharides characteristic of cellulose. Measuring susceptibility of biofilms to degradation by enzymes suggested differences in structurally important ECM molecules, with DTT biofilms having important protein and, to a lesser extent, cellulose components, and M63 biofilms having moderate protein, cellulose, and DNA components. Both biofilms conferred resistance to the bactericidal effects of amikacin and clarithromycin, with resident cells being killed at >10-fold lower rates than planktonic cells at almost all concentrations. Though these comparisons indicate differences in biofilm responses by M. avium under differing conditions, they also suggest common features of biofilm formation including cellulose production and antimicrobial resistance.
Keywords:
Mycobacterium avium
; biofilm
; resistance
; extracellular matrix
1. Introduction
Mycobacterium avium subs. hominissuis is an opportunistic pathogen, causing pulmonary infections in people whose respiratory systems that have been damaged due to injuries or diseases such as cystic fibrosis, bronchiectasis, chronic obstructive pulmonary disease, or emphysema, people who are immunocompromised, and, increasingly, in elderly people without predisposing conditions, especially elderly women with slender physique [1]. Infections with M. avium occur worldwide, and increasing incidence has been reported in resource rich countries where longitudinal epidemiological studies have been possible [2,3,4]. Like many other species of nontuberculous mycobacteria (NTM), M. avium is commonly found in numerous natural and engineered habitats, including marginal niches where few species are able to survive [5]. These include household reservoirs, such as showerheads, hot tubs, and humidifiers, that place people in frequent contact with M. avium and which have been shown to provide a route for infection via inhalation of aerosols [6,7,8,9,10]. M. avium is able to persist in plumbing systems and other habitats by biofilm formation, a process in which the exceptionally hydrophobic mycobacterial cells preferentially adhere to a surface or to particles in suspension, then undergo sessile growth and secrete an extracellular matrix (ECM) [11]. Growth of M. avium in biofilms has been observed to cause increased resistance of the bacterium to water treatment applications [12], disinfectants [13,14], and antimicrobial drugs [15,16]. These resistance phenotypes have led to the hypothesis that biofilm growth might condition M. avium to succeed as an opportunistic pathogen and/or that biofilm growth in vivo might explain the recalcitrance of M. avium pulmonary infections to therapy [17].
A limitation to our understanding of M. avium biofilms is the fact that studies have employed a variety of methods to cause biofilm formation in vitro, and these methods vary in many aspects, including temperature, nutrient and ion availability, substrate material for biofilm attachment, presence or absence of chemical stressors, pH, and presence or absence of flow. Most observations of M. avium biofilms have been made using only one method, and this raises uncertainty about whether observations made using one method are characteristic of M. avium biofilms broadly or whether they might be caused by conditions specific to that method. A side-by-side comparison of biofilms generated by multiple methods will help delineate which biofilm phenotypes are characteristic of M. avium biofilm formation mechanisms generally, and which phenotypes appear in response to specific conditions.
To enable a parallel comparison of M. avium biofilms formed in vitro, we selected two previously published methods for producing surface attached biofilms which varied in media composition and time required for biofilm maturation. In the first method, cells of M. avium were incubated in a previously described M63-based biofilm medium for 4 weeks to allow biofilm development over an extended period [18,19,20]. The other method used dithiothreitol (DTT)-induced thiol reductive stress to trigger rapid biofilm development, with biofilm formation occurring in one day [16,21,22]. In this study, we present a comparison of the ultrastructure appearance, biofilm ECM composition, and drug susceptibility of biofilm resident cells of M. avium grown in these two biofilm models.
2. Materials and Methods
2.1. Bacterial strains and media
Mycobacterium avium subs. hominissuis strains A5 and 2151 were originally isolated from the blood of an AIDS patient with disseminated mycobacterial infection and from the sputum of a patient with pulmonary mycobacterial infection, respectively. These strains were cultured on Middlebrooks 7H10 agar (BD, Franklin Lakes, NJ, USA) containing 10% oleic acid-albumin-dextrose-catalase (OADC; BD, Franklin Lakes, NJ, USA) supplement at 37℃ and in Middlebrooks 7H9 broth (BD, Franklin Lakes, NJ, USA) containing 10% OADC at 37℃ with constant agitation.
2.2. Biofilm culture conditions
Both biofilm methods studied were adapted for biofilm growth on multiwell polystyrene plates (unless otherwise noted) at 37℃ and stationary conditions. Growth of biofilms in M63 media was based on previous studies [18,19]. M. avium strains were grown in liquid culture to an optical density 600 nm (OD600) of approximately 0.8, then pelleted and resuspended in biofilm media to an OD600 = 0.2. The biofilm medium consisted of M63 medium (United States Biological, Salem, MA, USA) supplemented with 2% glucose, 0.5% Casamino Acids, 1 mM MgSO4, and 0.7 mM CaCl2. An appropriate volume (1 ml in a 24-well plate well, or equivalent scaled to the surface area of the bottom of other vessels) was dispensed into polystyrene plates, and plates were sealed with parafilm and incubated at 37℃ for four weeks. PBS was added to interwell spaces to prevent evaporation. Biofilm formation using dithiothreitol (DTT) was based on previous studies [17,21]. M. avium strains were grown in liquid culture to OD600 of approximately 0.8, then pelleted and resuspended in 7H9 broth + 5% OADC to an OD600 = 0.8. DTT (Roche, Basel, Switzerland) was added to a final concentration of 6 mM. An appropriate volume (1 ml in a 24-well plate well, or equivalent) was dispensed into polystyrene plates, and plates were incubated at 37°C for 24 hours.
2.3. Scanning electron microscopy
Biofilms were grown as described above, with the addition of circular glass coverslips to the bottom of the multiwell plate wells. Following biofilm formation, the biofilm media supernatants were removed and the biofilms were rinsed with water. Coverslips with adherent biofilms were removed from the multiwell plates, and the samples were dried overnight. Planktonic samples were prepared by resuspending M. avium grown in liquid culture in PBS, then allowing the bacterial suspension to dry on circular coverslips overnight. Then, the samples were fixed with 2% glutaraldehyde in 0.1 M sodium cacodylate (pH 7.5) for 1 hour. Samples were then rinsed with 0.1 M sodium cacodylate (pH 7.5), fixed in 1% osmium tetroxide, then rinsed again three times with buffer. The samples were then dehydrated using a graded ethanol series, before drying with a critical-point dryer. The dried samples were mounted on SEM stubs and sputter coated with iridium to a thickness of 5µm. Samples were imaged at the Notre Dame Integrated Imaging Facility using a Magellan 400 XHR scanning electron microscope (FEI, Hillsboro, OR, USA).
2.4. Calcofuor white staining
Biofilms were grown as described above, with volumes adjusted for 4-well chamber slides. Bacteria grown in liquid culture, then resuspended in PBS were added to 4-well chamber slides and allowed to dry in the wells overnight. Biofilm media supernatants were removed and the biofilms were rinsed with PBS, then all samples were fixed in 10% formalin for 1 hour. Then they were rinsed three times with PBS, followed by staining with calcofluor white (3 µg/ml; Sigma Aldrich, St. Louis, MO, USA) for 30 minutes. Samples were rinsed again, stained with Auramine M (BD, Franklin Lakes, NJ, USA) for 15 minutes, then destained with acid alcohol and rinsed with water. Finally, samples were mounted using mounting media (Thermo Fisher, Waltham, MA, USA) and imaged using an inverted fluorescent microscope (Zeiss, Oberkochen, Germany).
2.5. Biofilm enzymatic degradation assay
Biofilms were grown in 24-well polystyrene dishes, as described above. Biofilm media supernatants were removed and the biofilms were rinsed with phosphate-buffered saline (PBS). Then enzyme reaction buffers with or without enzymes were added to biofilm containing wells in triplicate, and the biofilms were incubated at 37℃ for 6 hours. Enzymes and buffers were formulated as follows: 0.1 mg/ml proteinase K (VWR, Radnor, PA, USA) in 30mM Tris-HCl (pH 8.0), 1 mg/ml Cellulysin Cellulase (MilliporeSigma, Burlington, MA, USA) in 50mM citrate buffer (pH 4.0), and 10 U/ml DNase I (Roche, Basel, Switzerland) in 200 mM Tris-HCl (pH 8.4), 20 mM MgCl2, 500 mM KCl. Following incubation, the enzyme or buffer supernatants were removed from the biofilms, and the biofilms were stained with 0.1% crystal violet for 10 minutes, rinsed twice with water, then allowed to dry overnight. To quantify the residual biomass, the crystal violet was solubilized in 30% acetic acid for 10 minutes with agitation and quantified by measuring the OD600 of the solution.
2.6. Antimicrobial susceptibility
Biofilms were grown as described above, with volumes adjusted for 48-well polystyrene plates. Planktonic cultures were inoculated in normal growth media at an OD600 = 0.1. Amikacin (Alfa Aesar, Haverhill, MA, USA) or clarithromycin (Sigma Aldrich, St. Louis, MO, USA) dilutions were prepared in the growth media corresponding to the different growth conditions (7H9 or M63), applied to the biofilm or planktonic bacteria, then incubated for 4 days at 37℃. Then the culture supernatants were removed, samples were resuspended in PBS, and biofilms were disrupted by scraping and pipetting in PBS+0.5% Tween 80, and a 30 minute incubation in a sonic bath (Thermo Fisher, Waltham, MA, USA). Serial dilutions of bacteria were made in PBS and plated on 7H10 agar +10% OADC to quantify colony forming units (CFU) surviving. Colonies were counted after 10 days of incubation at 37℃.
3. Results
3.1. Biofilm models display differences in ultrastructure cell
The general ultrastructure appearance of biofilms formed by each in vitro method were observed using scanning electron microscopy. Biofilms and planktonic samples of two strains of M. avium (A5 and 2151) were included in each comparison to observed strain-specific effects. The strains in this study represent different serovariants, each expressing a distinct serotype specific glycopeptidolipid (GPL): A5 is a serotype 4 strain and 2151 is a serotype 2 strain. The different methods of in vitro biofilm formation resulted in biofilms with distinct ultrastructure appearances that were similar between strains within a given method (Figure 1). Induction of biofilm formation with DTT gave rise to biofilms with relatively homogenous appearances, with few bacteria visible under a blanket of extracellular material. In contrast, biofilm formation in M63 medium resulted in biofilms with heterogeneous extracellular material appearing as sheets, threads, and granules and the many visible cells had heterogeneous morphologies including elongation and branching (Figure 1).
3.2. Calcofluor white staining suggests the presence of cellulose in biofilm ECM in both models
Previous studies characterizing the DTT induced biofilm model (Chakraborty et al., 2021) demonstrated the presence of cellulose in that model with multiple mycobacterial species, including M. avium. Cellulose was also identified by one study in the ECM of M. smegmatis biofilms formed in M63 media (Van Wyk et al., 2017). To compare the presence of polysaccharides with β(1→4) linked D-glucose units (characteristic of cellulose) in specimens from both biofilm models and planktonic bacteria, samples were stained with calcofluor white and counterstained with Auramine M to visualize the mycobacterial cell membrane. Relative to the planktonic bacteria controls, where no calcofluor white staining was detected, increased staining was observed in both biofilm models for each strain (Figure 2). In the DTT biofilm model, biofilms of M. avium strain A5 seemed to stain more strongly than strain 2151, suggesting possible strain specific differences in biofilm ECM composition. In all biofilm samples, calcofluor white staining colocalized closely with the Auramine M mycobacterial cell wall stain.
3.3. Biofilm models differ in structurally important ECM molecules
To further compare characteristics of the ECM in each biofilm model, we sought to infer structurally important ECM molecules by exposing biofilms to DNase I, Proteinase K, and Cellulase to cause targeted degradation of ECM molecules that have been identified in previous studies (Rose et al., 2015; Chakraborty et al., 2021). In each biofilm model, the two M. avium strains assayed displayed similar patterns of susceptibility to degradation by the different enzymes, suggesting that the structurally important ECM molecules are similar between strains within a given model (Figure 3). Biofilms generated using the DTT model were significantly degraded by Proteinase K, resulting in 87.3% and 92.2% reductions in biomass in strains A5 and 2151, respectively. Degradation by Cellulase was moderate, though variable between replicates, resulting in 45% and 38% reductions in biomass in strains A5 and 2151, respectively. DNase caused no degradation in DTT biofilms of strain 2151 biofilms, and a slight (7%) reduction in biomass in DTT biofilms of strain A5 (Figure 3). Biofilms grown in M63 biofilm media were moderately susceptible to degradation by each enzyme tested, suggesting heterogenous ECM composition, and biofilms of strain A5 exhibited higher degradation by each enzyme than biofilms of strain 2151. For strain A5 and 2151 biofilms, DNase caused biomass reductions of 42% and 18%, Proteinase K caused biomass reductions of 34% and 19%, and Cellulase caused biomass reductions of 40% and 24%, respectively (Figure 3).
3.4. Biofilm resident M. avium is resistant to killing by antimicrobial drugs
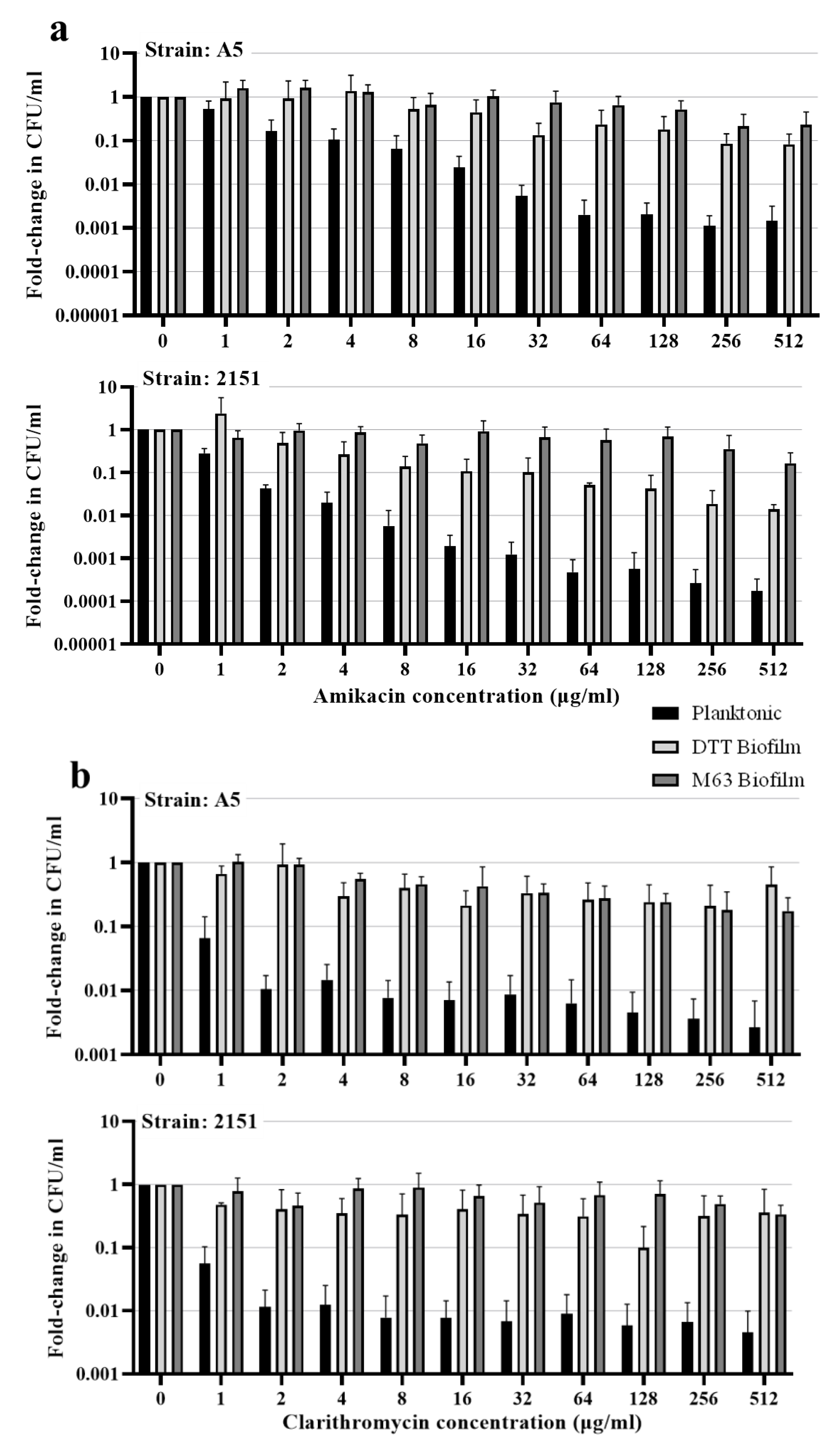
To assess resistance to killing by two drugs commonly prescribed to treat M. avium pulmonary disease, clarithromycin and amikacin, we exposed intact biofilms or planktonic cultures of both strains of M. avium to a range of concentrations of each drug for four days. In both strains, each biofilm model increased resistance of bacteria to killing by either drug (Figure 4). Amikacin killed >90% of planktonic M. avium A5 at 8 μg/ml and >99% at 32 μg/ml. Planktonic M. avium 2151 was more sensitive to killing by amikacin, with >90% killed at 2 μg/ml, >99% killed at 8 μg/ml, and >99.9% killed at 64 μg/ml. In contrast, when bacteria were grown in DTT biofilms both strains required 32x higher concentrations of amikacin (256 μg/ml in A5 and 64 μg/ml in 2151) for >90% killing, and in neither strain were >99% of bacteria killed even at the highest drug concentration tested (512 μg/ml) (Figure 4a). Resistance to killing was even higher in M63 biofilms, where the highest concentration of amikacin (512 μg/ml) did not kill >90% of M. avium of either strain in M63 biofilms (Figure 4a). Resistance to clarithromycin in both biofilm models followed a similar pattern. In both strains, clarithromycin killed >90% of planktonic bacteria at 1 μg/ml and >99% at 8 μg/ml (Figure 4b). However, in both biofilm models, the drug failed to kill 90% of bacteria of either strain at any concentration tested, with one exception. In the 2151 DTT biofilm >90% of bacteria were killed at a clarithromycin concentration of 128 μg/ml; however, this is likely not a true indication of drug susceptibility, as clarithromycin failed to kill >90% at drug higher concentrations tested (256 μg/ml and 512 μg/ml) (Figure 4b). An alternative explanation for this observation could be decreased solubility of the drug at higher concentrations, though evidence of this was not seen in the other clarithromycin treated samples.
4. Discussion
In this study, we compared two methods for growing M. avium biofilms in vitro, which cause biofilm formation by markedly different stimuli. The M63 biofilm media, four week incubation method likely models the gradual process of biofilm formation that occurs in the environment [18], while the DTT induced, one day incubation method models conditions the opportunistic pathogen might encounter in a potential host [16,21,22]. These methods resulted in biofilms with apparent ultrastructure differences, as observed by SEM. Similar to SEM observations of DTT induced biofilms of M. tuberculosis [21], the DTT induced biofilms of both strains of M. avium were characterized by an homogenous extracellular matrix concealing the cells within the biofilm. In contrast, the M63 incubated biofilms had extracellular materials with a variety of appearances and many visible cells with variable phenotypes, including elongation and branching. This heterogenous ultrastructure appearance has similarities to previous imaging of M. avium biofilms grown with similarly extended incubation times [23,24]. Observation of abnormal bacterial cell morphologies raises the possibility that aberrations from the normal cell division processes may occur when M. avium resides in a biofilm for weeks or months. Previous studies in M. smegmatis observed that both depletion and overexpression of the tubulin-like protein FtsZ, which is responsible for septum formation in mycobacteria, results in cells with abnormal elongation and branching phenotypes, and this protein could serve as a starting point for further study into the unusual morphologies observed [25,26,27].
Consistent with the apparent differences in ultrastructure between models, enzymatic degradation of biofilms with enzymes specific to ECM molecules identified in previous studies indicated differences in structurally important molecules between the models. In particular, the DTT biofilm model resulted in biofilms that were readily degraded by Proteinase K, in agreement with the finding of a previous study in M. tuberculosis [21], indicating a structurally essential protein component in the ECM of those biofilms, while M63 biofilms were degraded more moderately by Proteinase K. Since the presence of albumin in the culture media is necessary for DTT stimulated biofilm formation [22], it is possible that this is the major source of protein in the biofilm ECM in that model. The models also differed in susceptibility to DNase degradation: biofilms formed in M63 media were moderately degraded by DNase (with strain A5 biofilms degraded to a larger extent that strain 2151 biofilms), while biofilms formed by DTT stimulation had little to no susceptibility to degradation by the enzyme, consistent with previous characterization of the DTT biofilm model in M. tuberculosis [21]. Previously, contribution of extracellular DNA (eDNA) to the M. avium biofilm ECM was characterized in studies using a biofilm model based on a seven day incubation of bacteria in Hank’s Balanced Salt Solution [23,28], where the authors observed strain specific differences in abundance of extracellular DNA produced by M. avium, with strain A5 producing more eDNA than the other strains studied, in agreement with our observation that strain A5 M63 biofilms had greater susceptibility to DNase I degradation than strain 2151 M63 biofilms.
Notable similarities were also observed in our comparison of the biofilm ECM in the two models studied. Staining for β(1→4) linked D-glucose polysaccharides with calcofluor white suggested that both biofilm models produced biofilms with cellulose content in the ECM. Taken with the observation of moderate biofilm degradation by Cellulase in both models, these findings suggest that production of cellulose may be a biofilm formation behavior employed by M. avium generally. In our study, we observed more modest degradation of the DTT biofilm by cellulase than a previous study [21], where DTT biofilms of M. tuberculosis were almost totally disrupted by the enzyme, possibly indicating species specific differences between M. avium and M. tuberculosis. Production of cellulose by M. avium and mycobacteria more broadly was discovered only recently [21], and the genetic basis for this capability is unknown, as mycobacteria lack the classical cellulose synthase gene found in most bacteria [20,29]. In addition to cellulose in the mycobacterial biofilm ECM, M. tuberculosis has been observed to produce cellulose in the context of pulmonary infection [16]. Intriguingly, a recent study characterized a glycoside hydrolase family 6 (GH6) cellulase enzyme in M. smegmatis with high amino acid similarity to the M. tuberculosis cellulase CelA (88% similarity) and to a M. avium GH6 enzyme (82% similarity) [20]. In that study, the cellulase enzyme was not used by M. smegmatis for metabolism of cellulose, as is the case in other cellulolytic bacteria, but was found to prevent biofilm formation when overexpressed and to disrupt the biofilm ECM of the bacterium when added exogenously. This suggests the possibility that this cellulase could have a functional role in biofilm modeling by reverting bacteria in a biofilm back into the planktonic state, similar to a behavior observed previously in the cellulolytic bacterium Acetivibrio thermocellus [20,30]. Our observation of cellulose in biofilms formed by M. avium in response to different stimuli adds further motivation to fully characterize the mechanism of cellulose production in mycobacteria, the stimuli that trigger cellulose production, the role of mycobacterial cellulase enzymes, and the relevance of these processes for both biofilm and in vivo persistence.
The biofilm models were also similar in their conferral of resistance to killing by clarithromycin and amikacin. Cells residing in either biofilm model were killed at >10-fold lower rates than planktonic cells at almost all concentrations tested. This is in general agreement with other studies of each model, where increased resistance has been observed previously: M. avium and other NTM in DTT induced biofilms have been shown to be resistant to bedaquiline [16], and cells of M. smegmatis grown in M63 biofilms have been shown to have increased resistance to rifampin [20]. There are multiple potential explanations for the resistance observed in each model, and the differences in media and incubation time likely lead to some differences between models in the mechanisms contributing. However, it is possible that, despite the differences in the models, there may be common mechanisms of increased resistance in biofilm resident M. avium. Studies of M. avium grown in other models have demonstrated that bacteria grown in biofilms initially retain high levels of resistance to killing by antimicrobial drugs and disinfectants when resuspended, then lose resistance after 24 hours of growth in liquid culture, suggesting that changes in gene expression play a role in resistance [13,31]. Therefore, future studies comparing overall gene expression of bacteria residing in different biofilm models may offer leads toward discovering adaptations in gene expression leading to resistance that are common to M. avium biofilms formed under differing conditions.
The design of this study, in which we compared aspects of M. avium biofilm appearance, ECM composition, and resident cell phenotypes between two biofilm models, should serve as a template for expanded parallel analyses that could include additional biofilm models and employ additional methods to characterize the biofilms and their constituent cells. Characterization and comparison of the involvement of membrane lipids in M. avium biofilms would be particularly informative, as studies of these molecules in the context of biofilms have been carried out almost entirely in other mycobacterial species. For instance, M. tuberculosis and M. smegmatis pellicle (liquid-air interface) biofilms contain short chain mycolic acids in the ECM [32,33,34,35], and these have not been assayed in biofilms of other species. Additionally, M. avium and several other NTM express glycopeptidolipids (GPLs) which are necessary for biofilm formation [18,19,36], may have altered expression under biofilm conditions [37], and also appear to be immunologically important in the context of exposure and infection [38]. Further characterization of the GPL content and localization in M. avium biofilms is necessary to determine the specific role that these molecules play in biofilm formation.
Author Contributions
W.R.M. and J.S.S contributed to conceptualization and methodology. W.R.M. carried out the experiments, analyses, and data visualization. W.R.M. prepared the original draft of the manuscript and carried out revisions under the critical guidance of J.S.S. J.S.S. was responsible for supervision, project administration, and funding acquisition. Both authors have read and agreed to the published version of the manuscript.
Funding
JSS is funded by National Institute of Heath, grant number R21 AI168662
Institutional Review Board Statement
N/A
Data Availability Statement
Data from this study can be found as part of the manuscript.
Acknowledgments
We are grateful for the technical expertise and guidance from members of the University of Notre Dame Integrated Imaging Facility, especially Sarah Chapman and Tatyana Orlova. We are also grateful to Felipe Santiago Tirado and Ana Flores Mireles for use of their Zeiss inverted fluorescent microscope.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chalmers, J.D.; Aksamit, T.; Carvalho, A.C.C.; Rendon, A.; Franco, I. Non-Tuberculous Mycobacterial Pulmonary Infections. Pulmonology, 2018, 24, 120–131. [Google Scholar] [CrossRef]
- Hoefsloot, W.; Van Ingen, J.; Andrejak, C.; Ängeby, K.; Bauriaud, R.; Bemer, P.; Beylis, N.; Boeree, M.J.; Cacho, J.; Chihota, V.; Chimara, E.; Churchyard, G.; Cias, R.; Daza, R.; Daley, C.L.; Dekhuijzen, P.N.R.; Domingo, D.; Drobniewski, F.; Esteban, J.; Fauville-Dufaux, M.; Folkvardsen, D.B.; Gibbons, N.; Gómez-Mampaso, E.; Gonzalez, R.; Hoffmann, H.; Hsueh, P.R.; Indra, A.; Jagielski, T.; Jamieson, F.; Jankovic, M.; Jong, E.; Keane, J.; Koh, W.J.; Lange, B.; Leao, S.; Macedo, R.; Mannsåker, T.; Marras, T.K.; Maugein, J.; Milburn, H.J.; Mlinkó, T.; Morcillo, N.; Morimoto, K.; Papaventsis, D.; Palenque, E.; Paez-Peña, M.; Piersimoni, C.; Polanová, M.; Rastogi, N.; Richter, E.; Ruiz-Serrano, M.J.; Silva, A.; Da Silva, M.P.; Simsek, H.; Van Soolingen, D.; Szabó, N.; Thomson, R.; Fernandez, T.T.; Tortoli, E.; Totten, S.E.; Tyrrell, G.; Vasankari, T.; Villar, M.; Walkiewicz, R.; Winthrop, K.L.; Wagner, D. The Geographic Diversity of Nontuberculous Mycobacteria Isolated from Pulmonary Samples: An NTM-NET Collaborative Study. Eur. Respir. J. 2013, 42, 1604–1613. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Upadhyay, V. Epidemiology, Diagnosis & Treatment of Non-Tuberculous Mycobacterial Diseases. Indian Journal of Medical Research, 2020, 152, 185–226. [Google Scholar] [CrossRef]
- Prevots, D.R.; Marras, T.K. Epidemiology of Human Pulmonary Infection with Nontuberculous Mycobacteria a Review. Clinics in Chest Medicine, 2015, 36, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Falkinham, III, J.O. Surrounded by Mycobacteria: Nontuberculous Mycobacteria in the Human Environment. J. Appl. Microbiol., 2009, 107, 356–367. [CrossRef] [PubMed]
- Tzou, C.L.; Dirac, M.A.; Becker, A.L.; Beck, N.K.; Weigel, K.M.; Meschke, J.S.; Cangelosi, G.A. Association between Mycobacterium Avium Complex Pulmonary Disease and Mycobacteria in Home Water and Soil a Case-Control Study. Ann. Am. Thorac. Soc., 2020, 17, 57–62. [Google Scholar] [CrossRef]
- Falkinham, J.O.; Iseman, M.D.; de Haas, P.; van Soolingen, D. Mycobacterium Avium in a Shower Linked to Pulmonary Disease. J. Water Health, 2008, 6, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Mangione, E.J.; Huitt, G.; Lenaway, D.; Beebe, J.; Bailey, A.; Figoski, M.; Rau, M.P.; Albrecht, K.D.; Yakrus, M.A. Nontuberculous Mycobacterial Disease Following Hot Tub Exposure. Emerg. Infect. Dis., 2001, 7, 1039–1042. [Google Scholar] [CrossRef]
- Nishiuchi, Y.; Maekura, R.; Kitada, S.; Tamaru, A.; Taguri, T.; Kira, Y.; Hiraga, T.; Hirotani, A.; Yoshimura, K.; Miki, M.; Ito, M. The Recovery of Mycobacterium Avium-Intracellulare Complex (MAC) from the Residential Bathrooms of Patients with Pulmonary MAC. Clin. Infect. Dis., 2007, 45, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, L.A.; Falkinham, J.O. Aerosolization of Mycobacterium Avium and Mycobacterium Abscessus from a Household Ultrasonic Humidifier. J. Med. Microbiol., 2018, 67, 1491–1495. [Google Scholar] [CrossRef]
- Falkinham, J. Mycobacterium Avium Complex: Adherence as a Way of Life. AIMS Microbiol., 2018, 4, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Norton, C.D.; LeChevallier, M.W.; Falkinham, J.O. Survival of Mycobacterium Avium in a Model Distribution System. Water Res., 2004, 38, 1457–1466. [Google Scholar] [CrossRef]
- Steed, K.A.; Falkinham, J.O. Effect of Growth in Biofilms on Chlorine Susceptibility of Mycobacterium Avium and Mycobacterium Intracellulare. Appl. Environ. Microbiol., 2006, 72, 4007–4011. [Google Scholar] [CrossRef]
- Carter, G.; Wu, M.; Drummond, D.C.; Bermudez, L.E. Characterization of Biofilm Formation by Clinical Isolates of Mycobacterium Avium. J. Med. Microbiol., 2003, 52, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Mcnabe, M.; Tennant, R.; Danelishvili, L.; Young, L.; Bermudez, L.E. Mycobacterium Avium Ssp. Hominissuis Biofilm Is Composed of Distinct Phenotypes and Influenced by the Presence of Antimicrobials. Clin. Microbiol. Infect., 2011, 17, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, P.; Bajeli, S.; Kaushal, D.; Radotra, B.D.; Kumar, A. Biofilm Formation in the Lung Contributes to Virulence and Drug Tolerance of Mycobacterium Tuberculosis. Nat. Commun., 2021, 12. [Google Scholar] [CrossRef]
- Chakraborty, P.; Kumar, A. The Extracellular Matrix of Mycobacterial Biofilms: Could We Shorten the Treatment of Mycobacterial Infections? Microbial Cell, 2019, 6, 105–122. [Google Scholar] [CrossRef]
- Freeman, R.; Geier, H.; Weigel, K.M.; Do, J.; Ford, T.E.; Cangelosi, G.A. Roles for Cell Wall Glycopeptidolipid in Surface Adherence and Planktonic Dispersal of Mycobacterium Avium. Appl. Environ. Microbiol., 2006, 72, 7554–7558. [Google Scholar] [CrossRef]
- Recht, J.; Kolter, R. Glycopeptidolipid Acetylation Affects Sliding Motility and Biofilm Formation in Mycobacterium Smegmatis. J. Bacteriol., 2001, 183, 5718–5724. [Google Scholar] [CrossRef]
- Van Wyk, N.; Navarro, D.; Blaise, M.; Berrin, J.-G.; Henrissat, B.; Drancourt, M.; Kremer, L. Characterization of a Mycobacterial Cellulase and Its Impact on Biofilm-and Drug-Induced Cellulose Production. Glycobiology, 2017, 27, 392–399. [Google Scholar] [CrossRef]
- Trivedi, A.; Mavi, P.S.; Bhatt, D.; Kumar, A. Thiol Reductive Stress Induces Cellulose-Anchored Biofilm Formation in Mycobacterium Tuberculosis. Nat. Commun., 2016, 7. [Google Scholar] [CrossRef]
- Mavi, P.S.; Singh, S.; Kumar, A. Media Component Bovine Serum Albumin Facilitates the Formation of Mycobacterial Biofilms in Response to Reductive Stress. BMC Microbiol., 2023, 23, 111. [Google Scholar] [CrossRef] [PubMed]
- Rose, S.J.; Babrak, L.M.; Bermudez, L.E. Mycobacterium Avium Possesses Extracellular DNA That Contributes to Biofilm Formation, Structural Integrity, and Tolerance to Antibiotics. PLoS One, 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Nishiuchi, Y. Ultrastructure of the Mycobacterium Avium Subsp. Hominissuis Biofilm. Microbes Environ., 2021, 36, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dziadek, J.; Rutherford, S.A.; Madiraju, M. V.; Atkinson, M.A.L.; Rajagopalan, M. Conditional Expression of Mycobacterium Smegmatis FtsZ, an Essential Cell Division Gene. Microbiology, 2003, 149, 1593–1603. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Carette, X.; Potluri, L.P.; Sharp, J.D.; Xu, R.; Prisic, S.; Husson, R.N. Investigating Essential Gene Function in Mycobacterium Tuberculosis Using an Efficient CRISPR Interference System. Nucleic Acids Res., 2016, 44. [Google Scholar] [CrossRef]
- Hett, E.C.; Rubin, E.J. Bacterial Growth and Cell Division: A Mycobacterial Perspective. Microbiol. Mol. Biol. Rev., 2008, 72, 126–156. [Google Scholar] [CrossRef]
- Rose, S.J.; Bermudez, L.E. Identification of Bicarbonate as a Trigger and Genes Involved with Extracellular DNA Export in Mycobacterial Biofilms. MBio, 2016, 7. [Google Scholar] [CrossRef]
- Römling, U.; Galperin, M.Y. Bacterial Cellulose Biosynthesis: Diversity of Operons, Subunits, Products, and Functions. Trends in Microbiology, 2015, 23, 545–557. [Google Scholar] [CrossRef]
- Dumitrache, A.D.; Wolfaardt, G.; Allen, G.; Liss, S.N.; Lynd, L.R. Form and Function of Clostridium Thermocellum Biofilms. Appl. Environ. Microbiol., 2013, 79, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Falkinham, J.O. Growth in Catheter Biofilms and Antibiotic Resistance of Mycobacterium Avium. J. Med. Microbiol., 2007, 56, 250–254. [Google Scholar] [CrossRef]
- Ojha, A.; Anand, M.; Bhatt, A.; Kremer, L.; Jacobs, W.R.; Hatfull, G.F. GroEL1: A Dedicated Chaperone Involved in Mycolic Acid Biosynthesis during Biofilm Formation in Mycobacteria. Cell, 2005, 123, 861–873. [Google Scholar] [CrossRef]
- Ojha, A.; Hatfull, G.F. The Role of Iron in Mycobacterium Smegmatis Biofilm Formation: The Exochelin Siderophore Is Essential in Limiting Iron Conditions for Biofilm Formation but Not for Planktonic Growth. Mol. Microbiol., 2007, 66, 468–483. [Google Scholar] [CrossRef]
- Ojha, A.K.; Baughn, A.D.; Sambandan, D.; Hsu, T.; Trivelli, X.; Guerardel, Y.; Alahari, A.; Kremer, L.; Jacobs, W.R.; Hatfull, G.F. Growth of Mycobacterium Tuberculosis Biofilms Containing Free Mycolic Acids and Harbouring Drug-Tolerant Bacteria. Mol. Microbiol., 2008, 69, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Ojha, A.K.; Trivelli, X.; Guerardel, Y.; Kremer, L.; Hatfull, G.F. Enzymatic Hydrolysis of Trehalose Dimycolate Releases Free Mycolic Acids during Mycobacterial Growth in Biofilms. J. Biol. Chem., 2010, 285, 17380–17389. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Danelishvili, L.; Wu, M.; MacNab, M.; Bermudez, L.E. Mycobacterium Avium Genes Associated with the Ability to Form a Biofilm. Appl. Environ. Microbiol., 2006, 72, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Maya-Hoyos, M.; Leguizamón, J.; Mariño-Ramírez, L.; Soto, C.Y. Sliding Motility, Biofilm Formation, and Glycopeptidolipid Production in Mycobacterium Colombiense Strains. Biomed Res. Int., 2015, 2015, 419549–10. [Google Scholar] [CrossRef] [PubMed]
- Schorey, J.S.; Sweet, L. The Mycobacterial Glycopeptidolipids: Structure, Function, and Their Role in Pathogenesis. Glycobiology, 2008, 18, 832–841. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Ultrastructure appearance differs between DTT and M63 M. avium biofilm models. SEM was carried out on M. avium planktonic or biofilm samples grown on glass coverslips. (a) Representative images for each biofilm and planktonic condition for each M. avium strain at 10,000X magnification. (b) 30,000X magnification showing ECM detail of select representative images from each biofilm model.
Figure 1.
Ultrastructure appearance differs between DTT and M63 M. avium biofilm models. SEM was carried out on M. avium planktonic or biofilm samples grown on glass coverslips. (a) Representative images for each biofilm and planktonic condition for each M. avium strain at 10,000X magnification. (b) 30,000X magnification showing ECM detail of select representative images from each biofilm model.

Figure 2.
DTT and M63 biofilms both contain cellulose like polysaccharides. Biofilm or planktonic specimens of two M. avium strains (A5 and 2151) were grown and fixed in chamber slides, then stained with calcofluor white (3 μg/ml) to detect the β(1→4) linked D-glucose polysaccharides. Samples were then rinsed and stained with Auramine M for detection of the mycobacterial cell envelope, then destained with acid alcohol and rinsed with water. Representative images are shown, visualized at 1000X magnification on a Zeiss inverted fluorescence microscope.
Figure 2.
DTT and M63 biofilms both contain cellulose like polysaccharides. Biofilm or planktonic specimens of two M. avium strains (A5 and 2151) were grown and fixed in chamber slides, then stained with calcofluor white (3 μg/ml) to detect the β(1→4) linked D-glucose polysaccharides. Samples were then rinsed and stained with Auramine M for detection of the mycobacterial cell envelope, then destained with acid alcohol and rinsed with water. Representative images are shown, visualized at 1000X magnification on a Zeiss inverted fluorescence microscope.
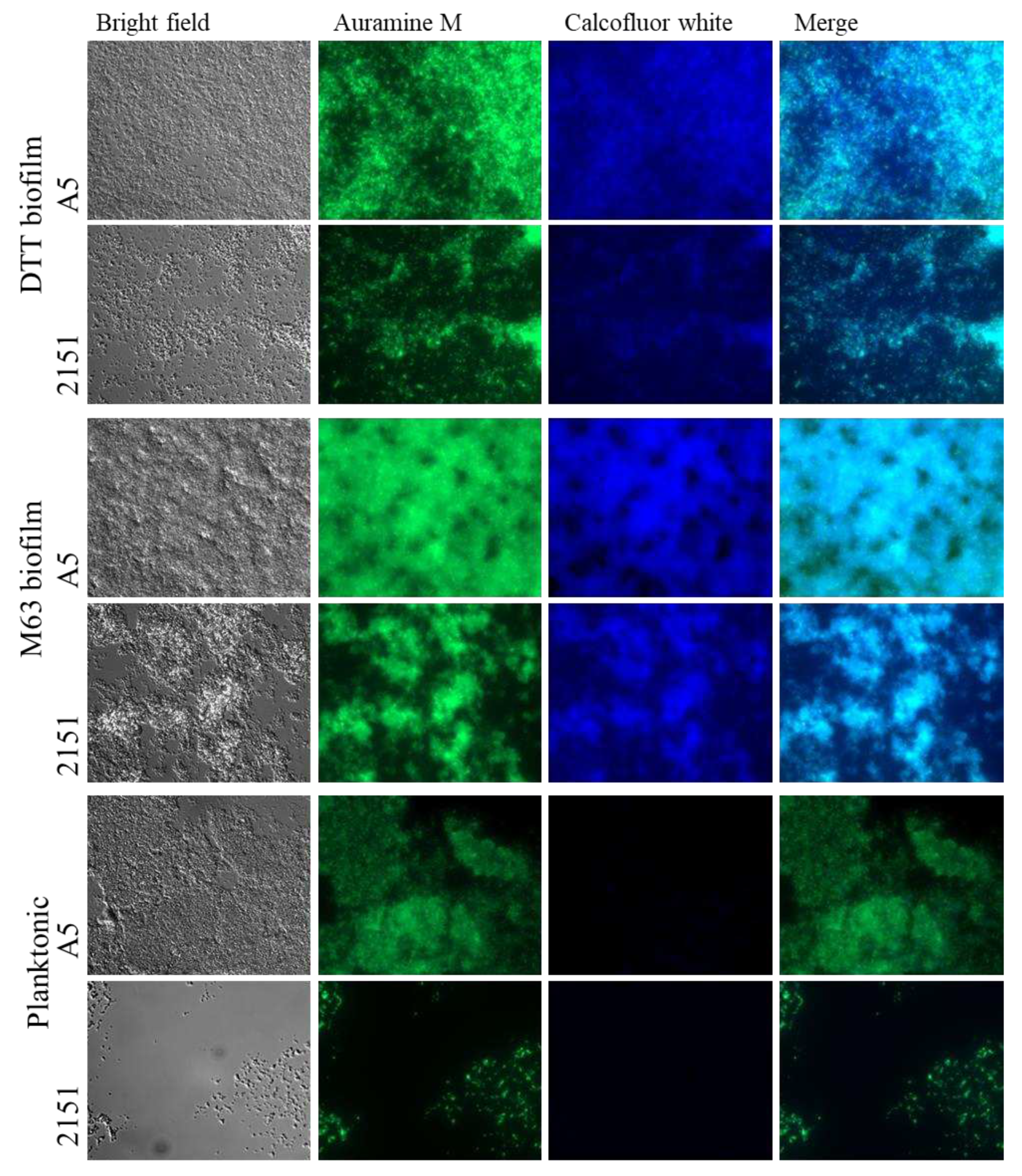
Figure 3.
M63 and DTT biofilms have differing susceptibility to enzyme degradation. Biofilms were treated with enzymes or enzyme reaction buffer for 6 hours, then stained for 10 minutes with 0.1% crystal violet, and rinsed twice with water. The stained biomass was dried overnight, then the crystal violet was solubilized in 30% acetic acid and the OD600 was quantified. Fold-change in biofilm biomass was calculated as the ratio of enzyme treated biofilm to paired buffer treated biofilm crystal violet staining, and the plot shows the average and standard deviation of 3-4 biological replicates. Statistics: one sample t-test comparing enzyme treated biofilms to respective buffer treated controls: * p<0.05, *** p<0.001. .
Figure 3.
M63 and DTT biofilms have differing susceptibility to enzyme degradation. Biofilms were treated with enzymes or enzyme reaction buffer for 6 hours, then stained for 10 minutes with 0.1% crystal violet, and rinsed twice with water. The stained biomass was dried overnight, then the crystal violet was solubilized in 30% acetic acid and the OD600 was quantified. Fold-change in biofilm biomass was calculated as the ratio of enzyme treated biofilm to paired buffer treated biofilm crystal violet staining, and the plot shows the average and standard deviation of 3-4 biological replicates. Statistics: one sample t-test comparing enzyme treated biofilms to respective buffer treated controls: * p<0.05, *** p<0.001. .
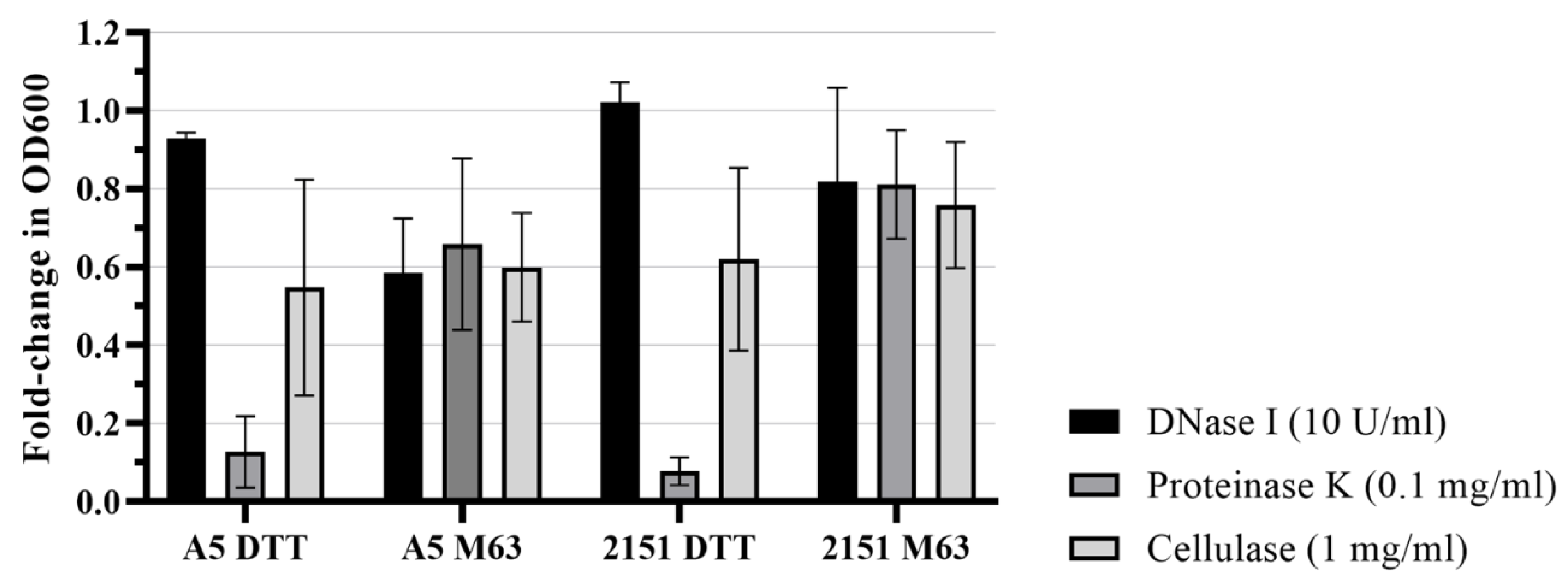
Figure 4.
Growth of M. avium in biofilms confers resistance to antimycobacterial drugs. M. avium strains A5 and 2151 grown in planktonic culture, DTT biofilm, or M63 biofilm were exposed to varying concentrations of (a) amikacin or (b) clarithromycin for 4 days. After 4 days, supernatants were removed and biofilms were disrupted by scraping, pipetting, and vortexing, and dilutions of each sample were plated on 7H10 agar +10% OADC to quantify CFU. For each condition, counts were normalized to the untreated (0μg/ml) count. The plot shows average fold-change and standard deviation for 3-4 biological replicates.
Figure 4.
Growth of M. avium in biofilms confers resistance to antimycobacterial drugs. M. avium strains A5 and 2151 grown in planktonic culture, DTT biofilm, or M63 biofilm were exposed to varying concentrations of (a) amikacin or (b) clarithromycin for 4 days. After 4 days, supernatants were removed and biofilms were disrupted by scraping, pipetting, and vortexing, and dilutions of each sample were plated on 7H10 agar +10% OADC to quantify CFU. For each condition, counts were normalized to the untreated (0μg/ml) count. The plot shows average fold-change and standard deviation for 3-4 biological replicates.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



