
Submitted:
18 September 2023
Posted:
20 September 2023
You are already at the latest version
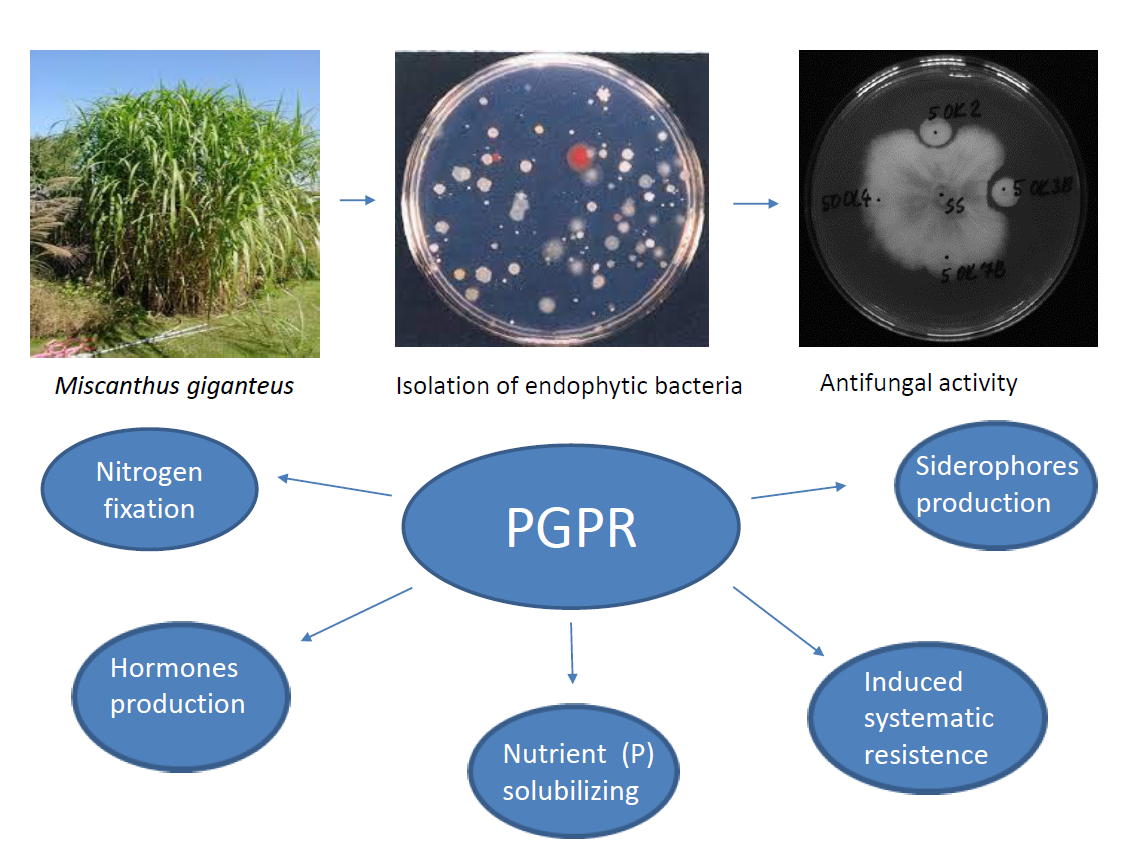
Abstract
Modern technologies can satisfy human needs only with the use of large quantities of fertilizers and pesticides that are harmful to the environment. For this reason, it is possible to develop new technologies for sustainable agriculture. The process could be made by using endophytic microorganisms with a (possible) positive effect on plant vitality. Bacterial endophytes have been reported as plant growth promoters in several kinds of plants under normal and stress conditions. In this study, isolates of bacterial endophytes from the roots and leaves of Miscanthus giganteus plants were tested for the presence of plant growth-promoting properties and their ability to inhibit pathogens of fungal origin. Pantoea ananatis and Pseudomonas libanensis were the predominant bacteria in leaves whereas other pseudomonads prevailed in roots. Selected bacterial isolates were able to solubilize inorganic phosphorus, to fix nitrogen, and to produce IAA, ACC deaminase and siderophore. Leaf bacterial isolate Pantoea ananatis 50 OL 2 had high production of siderophores and weak capabilities for phytohormone production. The root bacterial isolate of Pseudomonas libanensis 5 OK 7A showed the best results of phytohormone production. Four fungal cultures - Fusarium sporotrichioides DBM 3229, Sclerotinia sclerotiorum SS-01, Botrytis cinerea and Sphaerodes fimicola were used to test the antifungal activity and these cultures were selected for their representatives of known pathogenic fungi families, especially for crops. All selected roots endophyte isolates significantly inhibited the pathogenic growth of Fusarium sporotrichoides with a mean inhibition diamater (zone ≥ 5 mm). These results demonstrated that the root endophytic Pseudomonas sp. could be used as biofertilizer for crops.
Keywords:
endophytes
; Miscantus
; antifungal activity
; plant growth promotion properties
1. Introduction
Ornamental grass (Miscantus) is a perennial ryegrass of photosynthetic type C4. It is unique among C4-type plants in that it maintains high photosynthetic activity at lower temperatures and is highly productive even in cold regions. In addition to the energy potential, growing ornamental plants also have a beneficial effect on soil because they increase the amount of organic matter, nutrients and supports the microbial community [1]. In the conditions of Central Europe, diploid Chinese sedge (Miscanthus sinensis Andersson (1855)) or its triploid sterile hybrid with tetraploid sugar sedge (Miscanthus sacchariflorus (Maxim.) Franch.), giant sedge (Miscanthus × giganteus Greef & Deuter ex Hodkinson & Renvoiage) is most often cultivated.
Miscanthus × giganteus is either moderately or highly tolerant of heat, drought, flooding, salinity (below 100 mM), and cool soil temperatures (down to −3.4 °C). Miscanthus grows relatively well in soils contaminated by metals, or by industrial activities in general. Miscanthus × giganteus’ perennial nature, its ability to grow on marginal land, its water efficiency, non-invasiveness, low fertilizer needs, significant carbon sequestration and high yield have sparked significant interest among researchers, with some arguing that it has “ideal” energy crop properties. These properties can be reflected in an interesting microbial endophytic diversity.
The main reason for the growing interest in endophytes is their possible use in biotechnological applications. In agriculture, endophytic microorganisms are mainly used to promote plant growth and bio-control of plant diseases [2]. Until now, most of the microbial products used to promote plant growth and health were from the rhizosphere. The advantage of use of endophytic bacteria that return to the endophytic stage after product application is that they are better protected from biotic and abiotic threats coming from the plant’s surroundings. In addition, endophytes are ecologically adapted to the target niche and therefore can better overcome defensive reactions [3] than microorganisms originating from rhizosphere.
Endophytes usually have antifungal activity but also have antibacterial effects and sometimes they can to suppress the proliferation of nematodes [4]. A microorganism used in biotechnological applications or for biocontrol in agriculture should be stable in changing conditions (pH, temperature, concentration of various ions), to efficiently colonize roots for the even distribution of e.g., antimicrobial agents throughout the root system [5]. Endophytic microorganisms can inhibit the growth of fungal or bacterial pathogens by various mechanisms, including the production of antimicrobial substances, lytic enzymes, or siderophores. These mechanisms are similar to well-researched mechanisms that use rhizosphere bacteria. However, the study of them is more difficult due to the “hidden” way of life of endophytes in plant tissues [6,7,8,9].
The role of individual antimicrobial agents in the control of pathogens can vary with different bacterial endophytes. The degree of antimicrobial synthesis depends on nutritional factors (type of carbon source, trace elements, availability of other nutrients) as well as environmental factors. In the broader sense, we could include environmental protection and other stress conditions in the bio-control mechanisms [3].
The advantage of endophytic bacteria that return to the endophytic stage after product application is that they are better protected from biotic and abiotic threats coming from the plant’s surroundings. In addition, they are ecologically adapted to the target niche and can overcome defensive reactions [3]. Many endophytic bacteria prove in vitro plant-friendly properties, but only some of them maintain these properties in the plant, and only a small number of originally endophytic bacteria prove a significant effect in promoting plant growth or are abble to serve as a bio-control agent in agricultural practice [10]. Endophytic bacteria can also facilitate the phytoremediation of a number of environmental contaminants (e.g., organic pollutants). Endophytes colonize the interior of the plant body and can thus have bigger impact on the host plant than rhizosphere bacteria. Endophytes colonize the interior of the plant and can thus act on the host plant better than rhizobacteria even when the pollutants are degraded. Endophytic bacteria having both biodegradation and growth-promoting properties are better in these effects than bacteria having only one of these properties. One of the protection mechanisms is a formation of ACC deaminase, which can reduce the level of ethylene directly due to the stress conditions or promote plant growth by providing indolylacetic acid [11,12]. Endophyte bacteria increase the availability or supply of major nutrients that can also promote plant growth [8]. Nutrients, such as bound nitrogen, phosphorus or iron, are usually deficient in agricultural soils.
The innovativeness of the presented work lies in the study of unique strains of endophytic microorganisms isolated from ornamental plants for antifungal protection and to support the survival and growth of this energy crop. We hypothesized that since the Miscanthus plant is not frequently susceptible to fungal pathogens, the isolated endophytic bacteria would exhibit antifungal activity.
2. Materials and Methods
2.1. Isolation of Bacterial Endophytes
Endophytic bacteria were isolated using the method described by Schmidt (2017) [13]. Shoots and roots of Miscanthus gigantheus were harvested in a field in Lukavec, Czech Republic (49°33′56″ N, 14°59′26″ E) at summer and five samples of leaves or roots from ten plants preserved in sterile 0.01M MgSO4 max. 2 hours. Plant material was thoroughly washed with tap water to remove any adhering soil/debris, surface sterilized by disinfecting agent for 7 min (5% sodium hypochlorite with addition of 20 µl 0.1% Tween 20), transferred to a 70% ethanol solution for 2 min, rinsed four times with sterile distilled water and cut to small pieces (<0.5 cm). Approximately 2 g (fresh weight) of cut material was homogenized (Ultra-Turrax IKA-T10, max speed, 1 min) by thorough mixing under sterile conditions in 20 mL of sterile 12.5 mM potassium phosphate buffer (pH 7.1). This way prepared homogenate of Miscanthus gigantheus was used for inoculation of plates with diluted tryptic soy agar (5% TSA, 50% TSA) with cycloheximide (0.025 g L-1) and 48 h incubated at 20°C. Grown colonies were used for isolation of bacteria.
Endophytes from roots were labeled by the designation “OK” in their code respectively. Endophytes from leaves were labeled by the designation “OL” in their code, respectively.
2.2. Identification of Bacterial Isolates
Bacterial isolates were identified by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) according to Koubek et al. (2012) [14] using an Autoflex Speed MALDI-TOF/TOF mass spectrometer and MALDI Biotyper 3.1 software (Bruker Daltonik, Germany). With MALDI-TOF MS scores between 2.300-3.000 high is probability of identification at species level; 2.000-2.299 secure identification to genus level, probable identification to species level; 1.700-1.999 probable identification at genus level. Selected bacterial isolated were identified via 16S rRNA gene analysis [21].
2.3. Antifungal Activity of Endophytic Bacteria (Dual Culture Plate Test)
Dual culture plate tests were performed for all isolated bacterial strains. It is carried out on Waksman’s agar adapted for mold growth (D – glucose 7 g.L-1, D – fructose 7 g.L-1, bacteriological agar 15 g.L-1, bacteriological peptone 5 g.L-1, meat extract 5 g.L-1. After sterization KH2PO4 3,92 μM, NaCl 4,02 μM, KCl 1,8 μM, CaCl2 100 μM, MgCl2 134 μM, H3BO3 5 μM, L-ascorbic acid 850 μM, Al2(SO4)3 0,95 μM, K2SO4 0,95 μM, CuSO4 0,25 μM was added. Antifungal activities of isolated endophytic bacteria were tested against four fungal species (Botrytis cinerea, Fusarium sporotrichioides, Sclerotinia sclerotiorum, Sphaerodes fimicola). To evaluate the antagonist effect of different live bacteria, mycelia plugs from the edges of actively growing fungal cultures were placed in the center of Petri dish containing agar. Four bacteria isolates were streaked on the same plates at equal distance from the fungal inoculum. Plates with the fungal plug without bacteria were used as control. Plates were incubated at 25 °C for 72 hours to evaluate the inhibition activity of bacteria on the fungus. Assessment was done with three levels ‘+’ activity proven (zone ≥ 5 mm); ‘-’ activity not proven (no zone); ‘+/-’ ambiguous of fungal growth (zone ≤ 5 mm)
2.4. Detection of ACC-Deaminase Activity
The selected bacterial endophytes were tested for ACC deaminase activity. ACC-deaminase activity was detected with the colorimetric assay of 1-aminocyclopropane-1-carboxylate (ACC) based on ninhydrin reaction to rapidly screen bacteria containing ACC-deaminase from many microbial isolates according to Li et al. [15]. A bacterial isolate causing visibly reduced color intensity at 570 nm reflected in lower absorbance of supernatant compared with that of the non-inoculated nitrogen-defined medium with ACC was regarded as an ACC-utilizing bacterial isolate. The activity was rated: low activity demonstrated OD570nm≤ 0.05, mean activity demonstrated OD570nm 0.5 – 0.1 and high activity demonstrated OD570nm≥ 0.1.
2.5. Siderophore Production
Bacterial strains were screened for the production of siderophores according to Louden et al. [16] on blue agar chrome azurol S (CAS) medium containing CAS and hexadecyltrimethylammonium bromide (HDTMA) as indicators. Tested endophytic bacterial strains were inoculated into the CAS - agar and incubated at 28°C for 24 h. Development of yellowish orange halo around the colonies ware taken as indicator of siderophore production. Assessment was done semi-quantitatively with three levels (low - zone≤ 2 mm, medium - zone 2-5 mm, high - zone ≥5 mm).
2.6. Phosphate Solubilization
The ability to solubilize calcium phosphate was tested with Pikovskaya medium (agar) according to Jasim et al. [17]. The presence of lysis circle around the bacterial colony was observed as indicator of phosphate solubilization Phosphate solubilization was detected by clear zone. Similarly to siderophores, semi-quantitative evaluation (three levels: low - zone≤ 2 mm, medium – zone 2-5 mm, high - zone ≥5 mm) of P-solubilization was used.
2.7. Nitrogenase Activities
Colonies that appeared on the plates were selected and screened for nitrogenase activity on yeast-extract-mannitol-glutamate-succinate-medium (g.L-1: yeast extract, 0.2; K2HPO4, 0.5; MgSO4.7H2O, 0.2; NaCl, 0.1; CaCO3, 0.5; mannitol, 10; sodium glutamate, 0.32; sodium succinate, 1.6; pH 7.2). The strain showed high nitrogenase activity when grown on this medium. The procedure for determining nitrogenase activity on agar and its interpretation is described by Dadarwal et al. [18]. Endophytic bacteria were inoculated into 5 mL of liquid LB medium and incubated at 28 °C with constant shaking for 24 h. 100 µL of the grown bacterial culture were inoculated into 5 mL of liquid medium for testing nitrogenase activity. Cultivation took place for 48 h at 28 °C with constant shaking. Subsequently, the optical density of individual samples was measured at a wavelength of 550 nm. The medium itself for testing nitrogenase activity served as a negative control. The activity was rated: low activity demonstrated OD517nm≤ 0.05, medium activity demonstrated OD517nm 0.5 – 0.1 and high activity demonstrated OD517nm≥ 0.1.
2.8. Phytohormones Detection
Bacterial culture is prepared by cultivation for 48 hours in LB medium and suspension was filtered by 0,2 µm bacterial filter. Indole-3-acetic acid (IAA) and the cytokinins N6-(Δ2-isopentenyl) adenine (iP) and N6-(Δ2-isopentenyl) adenosine (iPR) were quantified in cultural filtrates with ultra-high performance liquid chromatography combined with mass spectrometry (UHPLC–MS/MS). This was done using an Acquity UPLC HSS T3 column (1.8 μm, 2.1 x 100 mm; Waters, Acquity) in a tandem mass spectrometer (AB Sciex, Qtrap 5500; electrospray ionization in positive mode) according to Hajšlová et al. [19]. Blank sterile LB medium served as control.
3. Results
3.1. Isolation of Endophytic Bacteria from Leaves and Roots of the Miscanthus Giganteus
From leaves of the Miscanthus giganteus plant, 24 bacterial isolates (OL labels) were obtained. From the roots of the Miscanthus giganteus, 34 bacterial isolates (OK markings) were obtained. From the 58 isolates, 43 bacterial strains were identified by MALDI TOF MS. With MALDI-TOF MS scores greater than 2.000, a majority of isolates could securely be identified at genus level. Gammaproteobacteria were by far most frequently isolated with Pantoea ananatis (Enterobacteriales) and Pseudomonas libanensis present in leaves, and other members of the genus Pseudomonas residing in roots. With MALDI-TOF MS scores between 2.300-3.000 high probability of identification at species level; 2.000-2.299 secure identification to genus level, probable identification to species level; 1.700-1.999 probable identification at genus level. Selected isolates with the best PGP properties were subjected to 16S rRNA analysis and the isolates were identified as Pseudomonas libanensis, azotoformans, fluorescent and marginalis (Table 1).
3.2. Antifungal Activity of Bacterial Isolates
The comprehensive results of the isolation and characterization of a large group of endophytic microorganisms are presented in article of Schmidt et al. [20]. In our case, we focused on the antifungal activity of already isolated bacteria, with the possibility of their use in organic farming, especially in the protection of crops from Fusarium wilt. The four fungal cultures - Fusarium sporotrichioides DBM 3229, Sclerotinia sclerotiorum SS-01, Botrytis cinerea and Sphaerodes fimicola were used to test the antifungal activity. These cultures were selected because they are representatives of known pathogenic fungi families, especially for crops. Fusarium sporotrichoides cause, for example, the disease of the pastinacea (Pastinaca sativa). This mold is a producer of phytotoxins [21]. Sclerotinia sclerotiorum is a common non-specific soil pathogen affecting more than 400 species of plants, e.g., cabbage, strawberry, lettuce, bean, etc. The symptoms of the attack include plant tissue wetting, coatings and black sculptures on the surface of plants. Botrytis cinerea is a necrotrophic fungi that affects many plant species, its most notable host is grapevine. In general, root isolates exhibit more pronounced antifungal activity than leaf isolates (Figure 1, Table 1 and 2). Six of the root endophytes (50 OK 3, 5 OK 5A, 5 OK 7A - Pseudomonas libanensis, 50 OK 6 – Pseudomonas marginalis, 50 OK 5 - Variovorax paradoxus, 5 OK 3B - Pseudomonas azotoformans) were able to inhibit growth of all 4 tested fungi (Botrytis cinerea, Fusarium sporotrichioides, Sclerotinia sclerotiniorum, Sphaerodes fimicola). Growth of Botrytis cinerea and Fusarium sporotrichioides was not inhibited by root endophyte 5 OK 7D and growth of Sclerotinia sclerotiniorum and Sphaerodes fimicola was not inhibited by root endophyte 50 OK 2. None of the leaf endophyte microorganisms inhibited all tested strains. Isolate 50 OL2 inhibited Botrytis cinerea, Fusarium sporotrichioides, Sclerotinia sclerotiniorum. Antifungal activity is usually not species-specific. Endophytic bacteria in most cases inhibited the growth of Botrytis cinerea and were also very well acting against Fusarium sporotrichoides. On the other hand, growth of the fungus Sphaerodes fimicola was not inhibited by any leaf isolate.

Table 2.
Evaluation of antifungal activity of roots endophyte isolates of Miscanthus giganteus.
| Isolates from plant tissue | Identification | Score* | Botrytis cinerea |
Fusarium sporotri chioides |
Sclerotinia sclerotiniorum | Sphaerodes fimicola |
|---|---|---|---|---|---|---|
| 50 OK 2 | Pseudomonas fluorescent | 2.179 | +/- | + | +/- | - |
| 50 OK 3 | Pseudomonas libanensis | 2.202 | + | + | + | + |
| 50 OK 5 | Variovorax paradoxus | 2.003 | + | + | + | + |
| 50 OK 6 | Pseudomonas marginalis | 2.223 | + | + | + | + |
| 50 OK 7 | Pseudomonas sp. | 2.238 | + | + | + | +/- |
| 50 OK 14A | Pseudomonas sp. | 2.277 | + | + | +/- | +/- |
| 5 OK 3B | Pseudomonas azotoformans | 2.138 | + | + | + | + |
| 5 OK 5A | Pseudomonas sp. | 2.198 | + | + | + | + |
| 5 OK 7A | Pseudomonas libanensis | 2.300 | + | + | + | + |
| 5 OK 7B | Pseudomonas sp. | 2.178 | + | + | - | +/- |
50, isolation from 50% TSA; 5, isolation from 5% TSA; OK, isolates from roots, OL, isolates from leaves‘+’ activity . proven (zone ≥ 5 mm); ‘-’ activity not proven (no zone); ‘+/-’ ambiguous of fungal growth (zone ≤ 5 mm). *2.300-3.000 high probability of identification at species level; 2.000-2.299 secure identification to genus level, probable identification to species level; 1.700-1.999 probable identification at genus level. Isolates identified via 16S rRNA gene analysis are highlighted in bold.
According to the test results of the plant growth promotion properties, 25 samples were selected, in which antifungal activity was tested. There are many works describing the antifungal activity of selected species [22,23] with the only exception, and this is Variovorax paradoxus, which is not much known from this perspective. Three samples, which had the highest antifungal activity under all conditions tested, all bacteria belonging to the group Pseudomonas.
3.3. Plant growth promoting potential of bacterial isolates
The plant growth promoting potential of selected five isolates from plant tissue was characterized in vitro by determining qualitatively and/or quantitatively: ACC deaminase activity, the ability of phosphate solubilization, iron carrier production, nitrogen fixation, and (Table 3) and producing of phytohormones (Table 4). The results shown are a selection of large published studies by Smidt et al. [20].
The observed activities differred significantly even among individual isolates, even though they were identified as the same genera. The deaminase activity was very low, the most striking features related to the production of phosphorus siderophores. The ability to fix nitrogen was monitored for all isolates, with the root isolates to a greater extent than for the leaf isolate. Leaf bacterial isolate 50 OL 2 had the highest antioxidant activity and weak capabilities for phytohormone production. Isolates 50 OL 2, 50 OK 6, 5OK 7A were identified as phosphate solubilizing microbes. According to their ability to solubilize inorganic phosphorus. The root bacterial isolate 5 OK 7A showed the best results of phytohormone production and was the only one produced by the IAA.
4. Discussion
The main task of this work was to screen the properties of naturally occurring strains of endophytic bacteria and their applicability in practice. Several interesting bacterial genera have been isolated. Pantoea ananatis isolated from leaves is a common species with a worldwide occurrence. It is airborne and can be found in soil, water and on plant surfaces as an epiphyte. It is an endophyte for many plant species and may also be a pathogen [23]. Pseudomonas savastanoi is most widely studied as the pathogen of olive trees (Olea europaea L.), where it causes the formation of galls, but has been isolated as an endophyte from many other plant species (e.g., cotton and corn) [24,25]. Bacillus mycoides is another bacterium with worldwide distribution, occurring in water and soil, where it lives saprophytic life. It has long been known for its association with the surface of plant roots, and in recent years was found that it can also be an endophyte of many plant species [10,26]. Most of the root isolates studied in this study belong to the Pseudomonas group - Pseudomonas fluorescens, Ps. extremorientalis, Ps. fluorescens, Ps.libanensis, Ps. antarctica, Ps. azotoformans. The MALDI-TOF MS method is not able to accurately distinguish between individual species in this group and therefore species identification is only indicative. Bacteria from this group can be found almost anywhere in the world, even in plant and animal bodies where they are part of the common microflora but can act as pathogens [27]. Other endophytic bacteria isolated were identified as Variovorax paradoxus, Rhizobium radiobacter and Janthinobacterium lividum [20,25,28], which are the root endophytes of many plant species, Bacillus mycoides and Bacillus weihenstephanensis (both members of Bacillus cereus) are difficult to distinguish by MALDI-TOF MS and Pseudomonas jessenii, a bacterium closely related to the Pseudomonas fluorescens group with which it has a similar incidence [29]. Due to the large number of isolates, only identification was performed using the MALDI TOF MS device, in our case the identification was successful at the genus level, probable identification at the species level. Thus, none of the isolated bacteria are unique to the plant Miscanthus giganteus, but they can be endophytes in other plant species. Analysis of 16S rRNA genes has been used as a complementary to achieve level-identification of bacterial strains that could not be securely identified by proteomic MALDI-TOF MS alone.
When testing the ACC deaminase activity of the isolated Pantoea ananatis bacteria, it showed this ability in three out of five cases. This finding agrees with the published results where only a fraction of strains showed this activity [30,31,32]. For Pseudomonas libanensis, ACC deaminase activity occurred occasionally. This finding agrees with the results published in the literature, where it was shown that some strains may have this type of activity [32]. Among the isolated bacteria of the Pseudomonas group originating from the roots, it was possible to prove ACC deaminase activity in the only single isolate. This is not in direct contradiction to the published results in the literature, although this activity has been demonstrated in the published results, but so low that it is unlikely to be proven with the used method [30,33]. Variovorax paradoxus did not show ACC deaminase when tested, although this activity was described in the literature [34].
Regarding phosphate solubilization, published data show that some strains of the bacterium Pantoea ananatis can show the ability to solubilize phosphate, but this ability does not occur in all strains [31,35]. This finding agrees with the results obtained with our isolates, where this ability occurs only occasionally. None from the isolated Pseudomonas savastanoi bacteria have the ability to solubilize phosphate. Ps. Savastanoi demonstrated this ability, but it always goes from the isolates from the rhizosphere [33]. Pseudomonads from the group Ps. fluorescens were able to solubilize phosphate in most cases, but there were large differences in the degree of solubilization among them [32].
In general, it can be said that isolates from roots are able to solubilize phosphate, with the exceptions on the other hand, because leaf isolates do not have this ability, except for a single sample. This is quite logical, because when the bacterium is associated with the roots, it is a beneficial property that allows the plant to obtain phosphate, whereas above ground, where there is no insoluble phosphate, this property is meaningless.
All isolates of the bacteria Pantoea ananatis, Pseudomonas savastanoi and except one and bacteria from the group Pseudomonas showed the ability to produce siderophores, which result agrees with those in the literature [31,33]. Variovorax paradoxus shows no ability to produce siderophores but, for which this ability is confirmed in most published works [36,37].
All samples were able to fix atmospheric nitrogen. It has been shown in the literature that they may have this ability [35]. The peculiarity was the large differences between individual leaf isolates, and on the contrary almost the same rate of fixation for all root endophytes. However, the origin of this phenomenon may be just coincidental. Although not directly tested for the presence of nitrogenase, Dadarwal et al. [18] proved in their work that all bacteria that were able to grow on the medium for testing nitrogenase activity showed this ability.
The achieved result reveal that all isolated strains produce cytokinin in the form of N6-(2-isopentenyl) adenine (iP), may not be surprising, since N6-(2-isopentenyl) adenosine is a part of the tRNA of all bacteria and eukaryotes, and the formation of iP depends only on the presence of two common enzymes [39,40]. If we stick to the division of strains of the bacterium Pantoea ananatis, as suggested in their work by Kido et al. [40], which is based on the relationship between phytohormone production and pathogenicity, then 3 of the samples probably belong to II. group that produces both auxin and cytokinins but causes no disease and the remaining samples belong to III. group that is characterized by the absence of genes for the production of indolylacetic acid (IAA), but can be pathogenic to some plant species. In Pseudomonas savastanoi, genes for IAA production are mostly present on plasmids, and therefore IAA production is highly variable in this species [41]. In root pseudomonads from the group Pseudomonas, the production of IAA is dependent on a yet to be precisely identified pathway that is tryptophan-dependent [42]. The production of phytohormones is highly variable in different strains of the bacterium Variovorax paradoxus and strains with high productivity are known, as well as strains that do not have the pathway for auxin synthesis at all [42].
The plant Miscanthus giganteus is not often attacked by many pathogens of fungal origin, although more of them are beginning to appear with the spread of its monocultural cultivation [43]. It is possible that the plant itself forms no antifungal agents and relies on fungal pathogen for the protection of endophytic microorganisms having antifungal properties themselves. When comparing all results, two bacterial isolates appear to be the most suitable for practical use: Pantoea ananatis 50 OL 2 and Pseudomonas libanensis 5 OK 7A, which showed both the most prominent growth promoting properties and antifungal activity.
5. Conclusions
Since the sixties of the twentieth century there has been a talk of sustainable forms of agriculture, where in addition to the social and economic point of view, the environmental aspect is emphasized. Methods and procedures have been sought and applied to preserve the environment and the agricultural landscape. One of the possible ways how to promote plant growth and health may be the use of favorable interactions between plants and microorganisms that have been known for over a hundred years. New discoveries in the field of plant microbiome stimulate the development of microbial inoculants, which can be used as biological fertilizers or plant protection agents against stress or disease. The bacteria induced alterations of the plants offer many possibilities for biotechnological medicinal and agricultural applications.
In this study, endophytic bacteria were isolated from roots and leaves of Miscanthus giganteus. Leaf bacterial isolate Pantoea ananatis 50 OL 2 had high production of siderophores and weak capabilities for phytohormone production. The root bacterial isolate Pseudomonas libanensis 5 OK 7A showed the best results of phytohormone production. Endophytic bacteria mainly Pseudomonas strain inhibited the growth of Botrytis cinerea and were also very well acting against Fusarium sporotrichoides. Thus, this root Pseudomonas isolates offers a potential for applications in the areas of agronomy, and could be used as a bio-fertilizer for crops.
Author Contributions
P. L. designed and supervised the experiments; G.K. made identification of bacteria, J.R: made isolation and characterization of endophytic microorganisms, P.L., Z.N., K.D. prepared and checked the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
A financial support by Technology Agency of the Czech Republic, project no. TN020000044.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kahle, P.; Beuch, S.; Boelcke, B.; Leinweber, P.; Schulten, H. R. Cropping of Miscanthus in Central Europe: biomass production and influence on nutrients and soil organic matter. European Journal of Agronomy 2001, 15, 171–184. [Google Scholar]
- Kobayashi, D.Y.; Palumbo, J.D. Bacterial endophytes and their effects on plants and uses in agriculture. In Microbial endophytes; Bacon, C.W., White, J.F., Eds.; Marcel Dekker Inc.: New York, USA, 2000; pp. 199–233. [Google Scholar]
- Mercado-Blanco, J.; Lugtenberg, B.J.J. Biotechnological Applications of Bacterial Endophytes. Current Biotechnology 2014, 3, 60–75. [Google Scholar] [CrossRef]
- Rana, K.L.; Kour, D.; Kaur, T. Endophytic microbes: biodiversity, plant growth-promoting mechanisms and potential applications for agricultural sustainability. Antonie van Leeuwenhoek 2020, 113, 1075–1107. [Google Scholar] [PubMed]
- Gunjal, A.; Waghmode, M.; Patil, N. , B. Endophytes and Their Role in Phytoremediation and Biotransformation Process. In Microbial Biotechnology in Environmental Monitoring and Cleanup, IGI Global. Editor Sharma A., Eds.; Publisher: Engineering Science Reference, Harshey, USA, 2018; pp. 238–251. [Google Scholar]
- Rosenblueth, M.; Martínez–Romero, E. Bacterial endophytes and their interactions with hosts. Molecular Plant-Microbe Interactions 2006, 19, 827–837. [Google Scholar] [PubMed]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 963401. [Google Scholar]
- Gupta, G.; Panwar, J.; Akhtar, M.S.; Jha, P.N. Endophytic nitrogen-fixing bacteria as biofertilizer. Sustainable Agriculture Reviews 2012, 11, 183–221. [Google Scholar]
- Gaiero, J.R.; McCall, C.A.; Thompson, K.A.; Day, N.J.; Best, A.S.; Dunfield, K.E. Inside the root microbiome: Bacterial root endophytes and plant growth promotion. American Journal of Botany 2013, 100, 1738–1750. [Google Scholar]
- Malfanova, N.V. Endophytic bacteria with plant growth promoting and biocontrol abilities. Ph.D. thesis, Leiden University, Leiden, Netherlands, 2013. [Google Scholar]
- Reed, M.L.E.; Glick, B.R. Applications of plant growth-promoting bacteria for plant and soil systems. In Application of Microbial Engineering; Editor Gupta, V.K., Schmoll, M., Mazutti, M.A., Maki M., Tuohy, M.G., Eds., Publisher: CRC Press, Boca Raton, USA, 2013; pp. 181–228. [Google Scholar]
- Afzal, M.; Khan, Q.M.; Sessitsch, A. Endophytic bacteria: Prospects and applications for the phytoremediation of organic pollutants. Chemosphere 2014, 117, 232–242. [Google Scholar]
- Schmidt, C.S.; Lovecká, P.; Mrnka, L.; Vychodilová, A.; Strejček, M.; Fenclová, M.; Demnerová, K. Distinct communities of poplar endophytes on an unpolluted and a risk elements-polluted site and their plant growth promoting potential in vitro. Microbial Ecology 2017, 75, 955–969. [Google Scholar]
- Koubek, J.; Uhlík, O.; Ječná, K.; Junková, P.; Vrkoslavová, J.; Lipov, J.; Kurzawová, V.; Macek, T.; Macková, M. Whole-cell MALDI-TOF: rapid screening method in environmental microbiology. International Biodeterioration and Biodegradation 2012, 69, 82–86. [Google Scholar]
- Li, Z.; Chang, S.; Lin, L.; Li, Y.; An, Q. A colorimetric assay of 1-aminocyclopropane-1-carboxylate (ACC) based on ninhydrin reaction for rapid screening of bacteria containing ACC deaminase. Letters in Applied Microbiology 2011, 53, 178–185. [Google Scholar]
- Louden, B.C.; Haarmann, D.; Lynne, A.M. Use of Blue Agar CAS Assay for Siderophore Detection. Journal of Microbiology and Biology Education 2011, 12, 51–53. [Google Scholar]
- Jasim, B.; Jimtha, C.J.; Jyothis, M.; Radhakrishnan, E.K. Plant growth promoting potential of endophytic bacteria isolated from Piper nigrum. Plant Growth Regulation 2013, 71, 1–11. [Google Scholar]
- Dadarwal, K.R.; Kundu, B.S.; Tauro, P. In vitro and in vivo nitrogenase aktivity of Rhizobium mutants and their symbiotic effectivity. Journal of Biosciences 1981, 3, 117–124. [Google Scholar]
- Hajšlová, J.; Fenclová, M.; Zachariašová, M. Methodology for the rapid screening of isolates of endophytic microorganisms and identification of strains with phytohormonal activity [in Czech]. ISBN 978-80-7080-869-6, 2013.
- Schmidt, C.S.; Mrnka, L.; Frantík, T.; Loveckaá, P.; Vosádka, M. Plant growth promotion of Miscanthus × giganteus by endophytic bacteria and fungi on non-polluted and polluted soils. World Journal of Microbiology and Biotechnology 2018, 34, 48. [Google Scholar] [PubMed]
- Manka, M.; Visconti, A.; Chełkowski, J.; Baottalico, A. Pathogenicity of Fusarium Isolates from Wheat, Rye and Triticale Towards Seedlings and their Ability to Produce Trichothecenes and Zearalenone. Journal of Phytopathology 2008, 113, 24–29. [Google Scholar]
- Loper, J.E.; Gross, H. Genomic analysis of antifungal metabolite production by Pseudomonas fluorescens Pf-5. European Journal of Plant Pathology 2007, 119, 265–278. [Google Scholar]
- Coutinho, T.A.; Venter, S.N. Pantoea ananatis: an unconventional plant pathogen. Molecular Plant Pathology 2009, 10, 325–335. [Google Scholar] [PubMed]
- Ercolani, G.L. Pseudomonas savastanoi and other bacteria colonizing the surface of olive leaves in the field. Journal of the General Microbiology 1978, 109, 245–257. [Google Scholar] [CrossRef]
- McInroy, J.A.; Kloepper, J.W. Survey of indigenous bacterial endophytes from cotton and sweet corn. Plant and Soil 1995, 173, 337–342. [Google Scholar]
- Earl, A.M.; Losick, R.; Kolter, R. Ecology and genomics of Bacillus subtilis. Trends in Microbiology 2008, 16, 269–275. [Google Scholar] [PubMed]
- Scales, B.S.; Dickson, R.P.; LiPuma, J.J.; Huffnagle, G.B. Microbiology, genomics, and clinical significance of the Pseudomonas fluorescens species complex, an unappreciated colonizer of humans. Clinical Microbiology Reviews 2014, 27, 927–948. [Google Scholar] [PubMed]
- Li, Y.H.; Zhu, N.N.; Zhai, Z.H.; Zhang, Q. Endophytic bacterial diversity in roots of Phragmites australis in constructed Beijing Cuihu Wetland (China). FEMS Microbiology Letters 2010, 309, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Andreani, N.A.; Martino, M.E.; Fasolato, L.; Carraro, L.; Montemurro, F.; Mioni, R.; Bordin, P.; Cardazzo, B. Reprint of ’Tracking the blue: A MLST approach to characterise the Pseudomonas fluorescens group´. Food Microbiology 2015, 45, 148–158. [Google Scholar]
- Johnston-Monje, D.; Raizada, M.N. Conservation and diversity of seed associated endophytes in Zea across boundaries of evolution, ethnography and ecology. PLoS ONE 2011, 6. [Google Scholar]
- Verma, P.; Yadav, A.N.; Kazy, S.K.; Saxena, A.K.; Suman, A. Evaluating the diversity and phylogeny of plant growth promoting bacteria associated with wheat (Triticum aestivum) growing in central zone of India. International Journal of Current Microbiology and Applied Sciences 2014, 3, 432–447. [Google Scholar]
- Zhang, Y.; He, L.; Chen, Z.; Wang, Q.; Qian, M.; Sheng, X. Characterization of ACC deaminase-producing endophytic bacteria isolated from copper-tolerant plants and their potential in promoting the growth and copper accumulation of Brassica napus. Chemosphere 2011, 83, 57–62. [Google Scholar]
- Trivedi, P.; Spann, T.; Wang, N. Isolation and characterization of beneficial bacteria associated with citrus roots in Florida. Microbial Ecology 2011, 62, 324–336. [Google Scholar]
- Belimov, A.A.; Safronova, V.I.; Sergeyeva, T.A.; Egorova, T.N.; Matveyeva, V.A.; Tsyganov, V.E.; Borisov; A. Y., Tikhonovich, I.A.; Kluge, C.; Preisfeld, A.; Dietz, K.; Stepanok, V.V. Characterization of plant growth promoting rhizobacteria isolated from polluted soils and containing 1-aminocyclopropane-1-carboxylate deaminase. Canadian Journal of Microbiology 2011, 47, 642–652. [Google Scholar]
- Oliveira, C.A.; Alves, V.M.C.; Marriel, I.E.; Gomes, E.A.; Scotti, M.R. .; Carneiro, N.P.; Guimaraes, C.T.; Schaffert, R.E.; Sá, N.M.H. Phosphate solubilizing microorganisms isolated from rhizosphere of maize cultivated in an oxisol of the Brazilian Cerrado Biome. Soil Biology and Biochemistry 2009, 41, 1782–1787. [Google Scholar] [CrossRef]
- Cornelis, P. Iron uptake and metabolism in pseudomonads. Applied Microbiology and Biotechnology 2010, 86, 1637–1645. [Google Scholar] [PubMed]
- Chen, L.; Dodd, I.C.; Theobald, J.C.; Belimov, A.A.; Davies, W.J. The rhizobacterium Variovorax paradoxus 5C-2, containing ACC deaminase, promotes growth and development of Arabidopsis thaliana via an ethylene-dependent pathway. Journal of Experimental Botany 2013, 64, 1565–1573. [Google Scholar] [PubMed]
- Persson, B.C. , Esberg, B., Ólafso,n Ó., Björk, G.R. Synthesis and function of isopentenyl adenosine derivatives in tRNA. Biochimie 1994, 76, 1152–1160. [Google Scholar] [PubMed]
- Sakakibara, H. Cytokinins: activity, biosynthesis, and translocation. Annual Review of Plant Biology 2006, 57, 431–449. [Google Scholar]
- Kido, K.; Hasegawa, M.; Matsumoto, H.; Kobayashi, M.; Takikawa, Y. Pantoea ananatis strains are differentiated into three groups based on reactions of tobacco and welsh onion and on genetic characteristics. Journal of General Plant Pathology 2010, 76, 208–218. [Google Scholar] [CrossRef]
- Pérez-Martínez, I.; Zhao, Y.; Murillo, J.; Sundin, G.W.; Ramos, C. Global genomic analysis of Pseudomonas savastanoi pv. savastanoi plasmids. Journal of Bacteriology 2008, 190, 625–635. [Google Scholar]
- Kochar, M.; Upadhyay, A.; Srivastava, S. Indole-3-acetic acid biosynthesis in the biocontrol strain Pseudomonas fluorescens Psd and plant growth regulation by hormone overexpression. Research in Microbiology 2011, 162, 426–435. [Google Scholar]
- Stewart, A.; Cromey, M. Identifying disease threats and management practices for bio-energy crops. Current Opinion in Environmental Sustainability 2011, 3, 75–80. [Google Scholar]
Figure 1.
Illustration of the distribution of bacteria and fungi culture on a Petri dish with Waksman‘s agar; red point – fungal culture, green point – bacterial culture.
Figure 1.
Illustration of the distribution of bacteria and fungi culture on a Petri dish with Waksman‘s agar; red point – fungal culture, green point – bacterial culture.
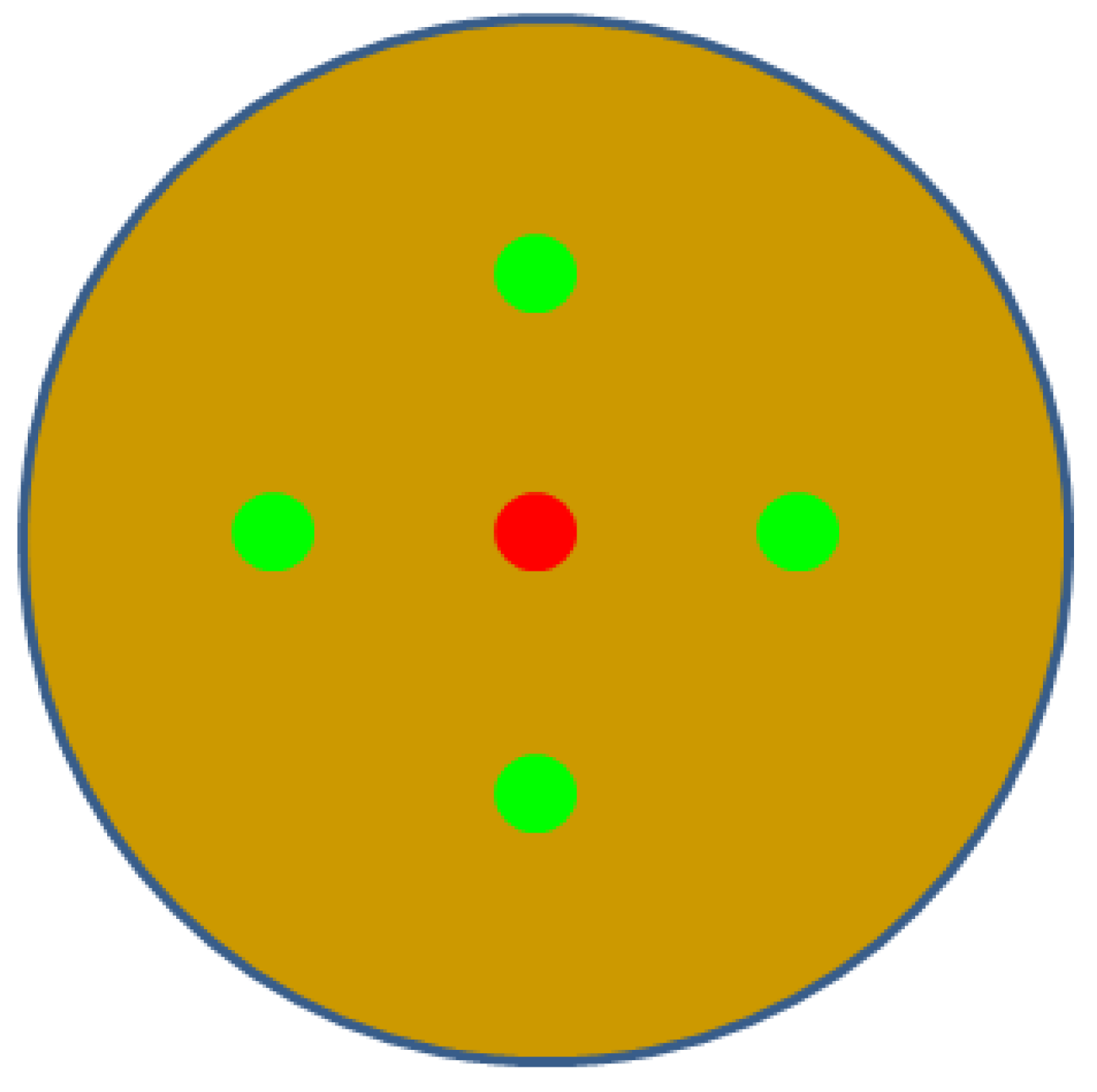
Figure 2.
A - Antifungal activity of selected endophyte (5 OK 7B - Pseudomonas sp., 50 OL 4 - Pseudomonas sp., 50 OK 2 - Pseudomonas fluorescent, 5 OK 3B - Pseudomonas azotoformans on Sclerotinia sclerotiniorum on Waksman‘s agar; B - control growth of Sclerotinia sclerotiniorum (SS).
Figure 2.
A - Antifungal activity of selected endophyte (5 OK 7B - Pseudomonas sp., 50 OL 4 - Pseudomonas sp., 50 OK 2 - Pseudomonas fluorescent, 5 OK 3B - Pseudomonas azotoformans on Sclerotinia sclerotiniorum on Waksman‘s agar; B - control growth of Sclerotinia sclerotiniorum (SS).
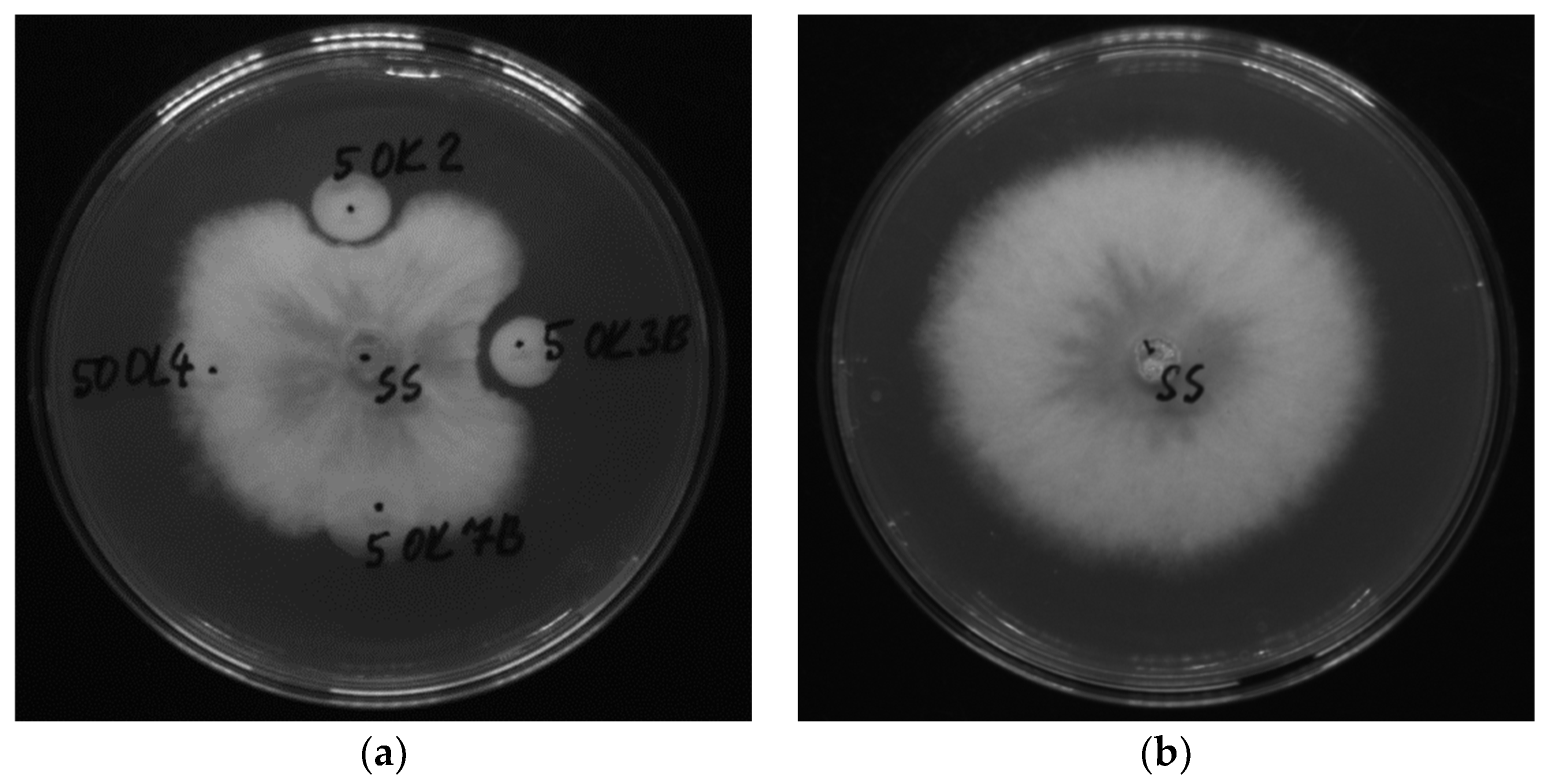
Table 1.
Antifungal activity of endophytes isolated from leaves of Miscanthus giganteus.
| Isolates from plant tissue | Identification | Score* |
Botrytis cinerea |
Fusarium sporotri- chioides |
Sclerotinia scleroti- niorum |
Sphaerodes fimicola |
|---|---|---|---|---|---|---|
| 50 OL 2 | Pantoea ananatis | 2.386 | + | + | + | - |
| 50 OL 4 | Pseudomonas sp. | 2.109 | + | +/- | - | - |
| 50 OL 5 | Pseudomonas sp. | 2.056 | +/- | + | +/- | - |
| 50 OL 6 | Ps. libanensis | 2.117 | + | - | + | - |
| 50 OL 7 | Pseudomonas sp. | 1.990 | + | - | - | - |
| 5 OL 1 | Pantoea ananatis | 2.299 | + | + | +/- | - |
| 5 OL 2 | Pantoea ananatis | 2.381 | + | +/- | + | - |
| 5 OL 3 | Pseudomonas sp. | 2.029 | +/- | + | + | - |
| 5 OL 4 | Pseudomonas sp. | 2.111 | +/- | - | - | - |
| 5 OL 5 | Pseudomonas sp. | 2.095 | +/- | + | - | - |
| 5 OL 6 | Pantoea ananatis | 2.368 | + | +/- | +/- | - |
| 5 OL 7 | Pseudomonas sp. | 2.086 | +/- | + | - | - |
| 5 OL 8 | Pseudomonas sp. | 2.106 | +/- | +/- | - | - |
| 5 OL 11 | Pseudomonas sp. | 2.108 | +/- | - | - | - |
| 5 OL 12 | Pantoea ananatis | 2.277 | + | + | +/- | - |
50, isolation on 50% TSA; 5, isolation on 5% TSA; OL, isolates from leaves. ‘+’ activity proven (zone ≥ 5 mm); ‘-’ activity not proven (no zone); ‘+/-’ ambiguous of fungal growth (zone ≤ 5 mm). *2.300-3.000 high probability of identification at species level; 2.000-2.299 secure identification to genus level, probable identification to species level; 1.700-1.999 probable identification at genus level; Isolates identified via 16S rRNA gene analysis are highlighted in bold.
Table 3.
Plant growth promoting potentials of selected bacterial isolates from leaves and roots of Miscanthus giganteus.
Table 3.
Plant growth promoting potentials of selected bacterial isolates from leaves and roots of Miscanthus giganteus.
| Isolates from plant tissue | Identification | ACCd | P-SOL | SID | NITRO |
|---|---|---|---|---|---|
| 50 OK 3 | Pseudomonas libanensis | + | - | +++ | +++ |
| 50 OK 5 | Variovorax paradoxus | + | - | +++ | + |
| 50 OK 6 | Pseudomonas marginalis | - | + | +++ | ++ |
| 5 OK 7A | Pseudomonas libanensis | - | +++ | + | ++ |
| 50 OL 2 | Pantoea ananatis | + | ++ | +++ | + |
50, isolation of z 50% TSA; 5, isolation from 5% TSA; OK, isolates from roots; OL, isolates from leaves; ACCd, ACC-deaminase: ‘+’ low activity demonstrated (OD570nm≤ 0.05); ‘++’ mean activity demonstrated (OD570nm 0.5 – 0.1); ‘+++’ high activity demonstrated (OD570nm≥ 0.1); ‘-’ no activity; P-SOL, phosphate solubilization: ‘+’ low activity demonstrated (zone≤ 2 mm); ‘++’ mean activity demonstrated (zone 2-5 mm); ‘+++’ high activity demonstrated (zome ≥5 mm); ‘-’ no activity; SID, production of siderophores: ‘+’ low activity demonstrated (zone≤ 2 mm); ‘++’ mean activity demonstrated (zone 2-5 mm); ‘+++’ high activity demonstrated (zome ≥5 mm); ‘-’ no activity; NITRO, nitrogen fixation: ‘+’ low activity demonstrated (OD517nm≤ 0.05); ‘++’ mean activity demonstrated (OD517nm 0.05 – 0.1); ‘+++’ high activity demonstrated (OD517nm≥ 0.1); ‘-’ no activity.
Table 4.
Amount of phytohormones (ng.mL-1) produced by bacterial isolates from leaves and roots of Miscanthus giganteus.
Table 4.
Amount of phytohormones (ng.mL-1) produced by bacterial isolates from leaves and roots of Miscanthus giganteus.
| Isolates from plant tissue | Identification | Phytohormons (ng.mL−1) | ||
|---|---|---|---|---|
| iP | iPR | IAA | ||
| 50 OK 3 | Pseudomonas libanensis | 5.5 | n.d. | n.d. |
| 50 OK 5 | Variovorax paradoxus | 3.8 | n.d. | n.d. |
| 50 OK 6 | Pseudomonas marginalis | 3.7 | n.d. | n.d. |
| 5 OK 7A | Pseudomonas libanensis | 11.7 | 8.9 | 12.6 |
| 50 OL 2 | Pantoea ananatis | 1.1 | n.d. | n.d. |
50, isolation from 50% TSA; 5, isolation from 5% TSA; OK, isolates from roots; OL, isolates from leaves; iP, N6-(2-isopentenyl) adenin; iPR, N6-(2-isopentenyl) adenosin; IAA, indolyl-3-acetic acid; n.d. - no detected.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



