Submitted:
19 September 2023
Posted:
20 September 2023
You are already at the latest version
Abstract
Bifenthrin is a synthetic pyrethroid insecticide. It is used worldwide against different agricultural pests. Nervous system (NS) is the primary target organ for the pyrethroid group of insecticide which causes hyper excitation. The aim of this study to determine the effects of bifenthrin on the brain of rabbits. The animals were treated for 14 days with three different doses (1/5th, 1/10th and 1/15th of LD50) of bifenthrin. Neurotransmitters and Histopathological alterations were observed at the end of the experiments. Neurochemical parameters were checked 24 hours after the last dose. Significant changes in the level of biogenic amines were observed. The levels of epinephrine (EPN), dopamine (DA), 3, 4 dihydroxyphenylacetic acid (DOPAC) were decreased as compared to controls group with the increase in insecticide. However, norepinephrine (NE) and homovanillic acid (HVA) levels were found to be significantly higher than those of the controls. A significant reduction in the activity of the acetyl cholinesterase enzyme was found in the frontal cortex’s erythrocyte, plasma and brain of rabbit with the increase in administered bifenthrin. The oxidative stress showed significantly decrease in the activity of glutathione peroxidase (GPx), superoxide dismutase (SOD), catalase (CAT) and glutathione S- transferase (GST) with the increase in dose of bifenthrin. Histopathological study of brain tissues showed normal GC (glial cells) and NC (neuronal cells) in the frontal cortex of brain tissue of control groups while the treated groups showed evacuation (odema), elongation of pyramidal nucleus, glial cell damage and brain haemorrhage.
Keywords:
Rabbit
; Bifenthrin
; Neurotransmitter parameters
; oxidative stress
; histopathology
1. Introduction
Pakistan is an agricultural country and more than 70% of its population is related to agriculture directly or indirectly. Farmers are trying to boost their yield of crops to feed Pakistan’s overgrown population. For this purpose, they are using different insecticides indiscriminately with over and under dose. They also use these chemicals in their houses to protect themselves and their families from different insect pests like mosquitoes, house flies, cockroaches, ants etc [1]. This causes environmental pollution, human health hazards, insect resistance and damages the non-target organisms too [2]. These insecticides are also harmful to aquatic life like fish. The issue of pesticide toxicity should be addressed scientifically within cooperative action plans at the national, regional, and worldwide levels [2].
Different groups of insecticides (organchlorines, organophsophates, synthetic pyrethroides, carbamates and IGR’s) are being used in the world. Among these insecticides, synthetic pyrethroids (SPs) are the common insecticides used more than 25% in the global insecticide market for home and agricultural pest management. Owing to their high efficacy, they have replaced the classical insecticides like organochlorine and organophosphate. Synthetic pyrethroids entered the environment due to their wide spread use and they cause many hazardous effects on targeted and non-targeted organisms. The hazardous effects include headache, reduced energy, muscle twitching and loss of consciousness [3]. Pyrethroids cause neurotoxicity as they target the central nervous system in both targeted and non-targeted organisms and lead to quick paralysis and ultimately cause their death.
Bifenthrin is a member of third-generation synthetic pyrethroids and is widely used on account of its high stability and great insecticidal activity. It causes headache, body shaking, a severe and protracted tremorigenic reaction, overheating, vocalization, prostration, and at high doses it causes convulsions and even death of organisms [4]. It has an environmental half-life that resulted in the existence of substantial concentrations of it in the sediment and irrigation water run-off [5]. Acute poisoning with bifenthrin in mammals produces aggressive sensitivity to stimuli [6]. As bifenthrin is used in agro-industry for pest control, there is a high risk of chronic exposure to humans through food products [7].
When harmful substances enter the body, liver is the first organ that is affected [8]. It carries out crucial tasks like metabolism, detoxification, transportation, and biosynthesis, all of which can be negatively impacted by harmful chemicals when they rise above a certain threshold level [9]. When exposed to bifenthrin, individuals with impaired liver function may experience cell death and liver disease as a result of oxidative stress, DNA damage, mitochondrial malfunction, and lipid peroxidation [10]. The number of synthetic substances that are known to harm the liver grows early [11]. There are more and more reports of endocrine disturbance caused by pyrethyroid pesticides [12].
In the membrane of postsynaptic cells, acetyl cholinesterase enzyme plays a vital role for the hydrolysis of acetylcholine [13]. In the central nervous system (CNS), parasympathetic nervous system (PNS) and somatic nervous system are major neurotransmitters [14]. Upon the inhibition of acetylcholinesterase, there is accumulation of acetylcholine in the synapse which over-stimulates the receptors, leading to a disruption of the nervous system [15]. Organophosphates also inhibit the acetylcholinesterase best when the phosphorus atom is double-bonded some time with oxygen atom or sulphur atom [13]. Synthetic pyrethroids are the newest major class of insecticides, having been introduced in the 1980s [16]. They are derived from the natural pyrethrin of Chrysanthemum cinerariaefolium, which has been used in China since the first century A.D. [17]. Despite their high cost, they are excellent insecticides with low mammalian toxicity, high field stability, and low soil persistence [17]. These reasons resulted in its extensive use domestically and in agriculture as well. Bifenthrin is highly toxic not only for mammals including human beings but also has killing effects for aquatic life too. So, this study is carried out to investigate the neurotoxic effects of bifenthrin in the brain tissues of rabbits by exposing them to its different doses.
2. Materials and methods
2.1. Experimental materials
The bifenthrin (Bifenthrin 10% EC, Jaffer Group of Companies, Pakistan) was bought from market and its different concentrations (1/15th, 1/10th and 1/5th of its LD50 mg/kg body weight) were formulated in normal saline (0.9% NaCl).
Healthy rabbits weighing (870 ± 30 grams) were purchased from a local market (Jhang bazar, Faisalabad, Pakistan) and were kept in Animal House, Department of Zoology, Government College University, Faisalabad, Pakistan. The animals were maintained under standard housing laboratory conditions (25℃ ± 2℃ temperature and 60 ± 5% relative humidity), 12 h light/dark cycles, and fed on alfalfa.
2.2. Ethical statement
Animal handling and subsequent sacrifice were carried out according to the guidelines provided by the ‘Ethical Committee of the Faculty of Biological Sciences’, Government College University, Faisalabad, Pakistan.
2.3. Determination of LD50 of bifenthrin
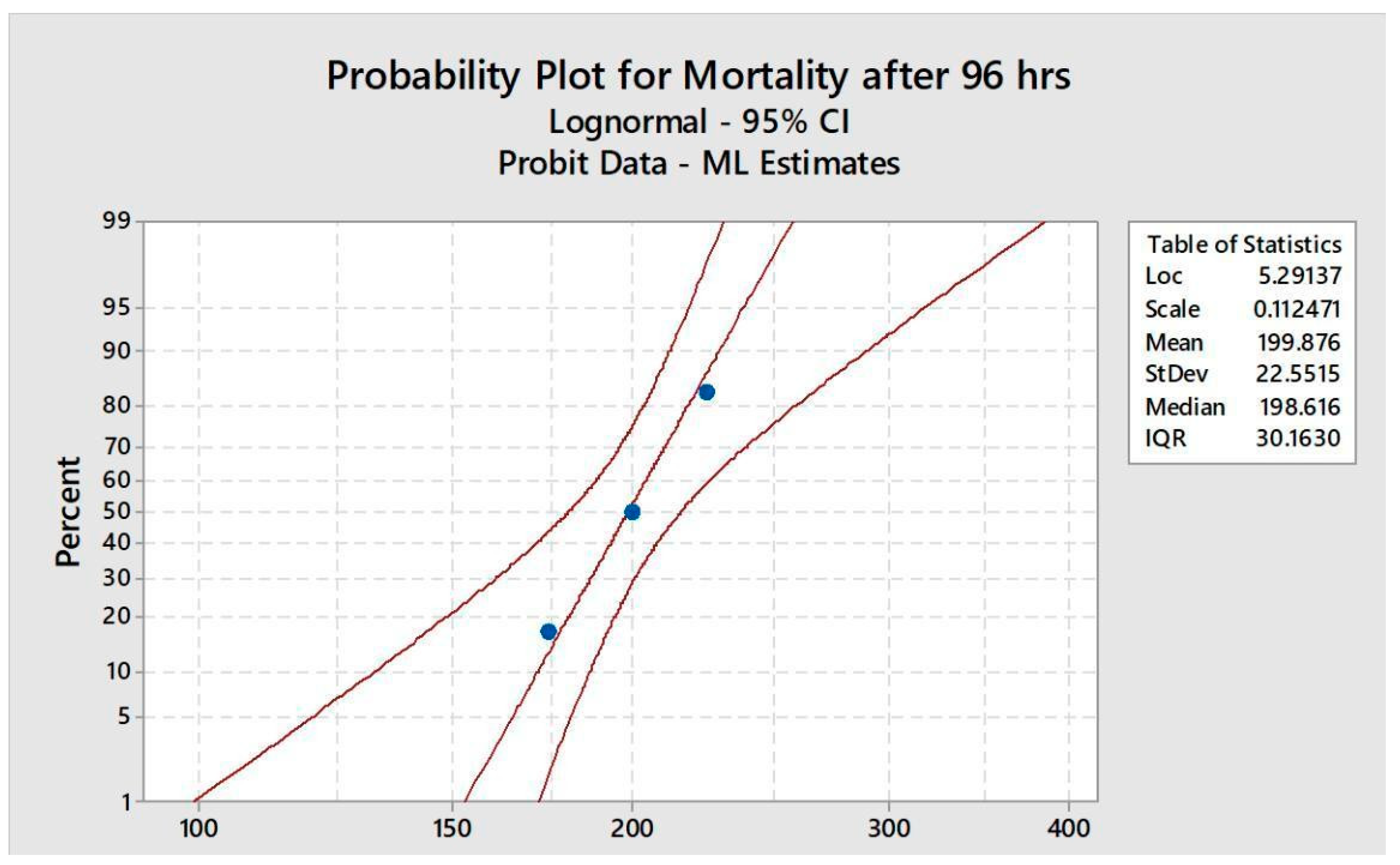
Five groups with (n=6) were made and five different doses of bifenthrin @ 150, 175, 200, 225 and 250mg/kg BW were intraperitoneally administered for 96 hours. The movement and the behaviour along with the mortality of rabbits were continuously monitored throughout the study. Rabbits without any movement for a long period were considered dead and were removed immediately to prevent contamination. Mortality rate was observed after every 24, 48, 72 and 96 hours. During the LD50 assessment, rabbits were not fed and tests were conducted in accordance with the standard methods [18].
2.4. Experimental design
The rabbits were placed in polypropylene-metal cages with 5 animals per cage and were allowed to acclimatize for one week prior to treatment. All the experimental protocols conducted on rabbits were performed in accordance with the internationally accepted principles for laboratory animal use and care and Institutional animal care and use committee. The toxic effects of bifenthrin was investigated through biochemical and histopathological parameters. For this purpose, five groups were made: G1, G2, G3, G4 and G5 with five animals in each group. G1 is a control group which did not treat any dose of pesticide while G2 was treated with 0.9% saline and G3, G4 and G5 were treated with different doses of pesticide (1/15th, 1/10th and 1/5th of LD50) respectively for 14 days [19].
2.5. Collection of samples and processing
The animal’s rabbits were anaesthetised by using diethyl ether. After that 2 ml of heparinized blood sample per rabbit was collected from the ear of rabbits. For the harvesting of plasma, blood samples were centrifuged at 3000 rpm for fifteen minutes. After the harvesting of plasma, erythrocytes were left behind. After the removal of the buffy coat, it was subjected to three times washing with equal volume of saline, and was used to obtain 10% erythrocyte suspension in phosphate buffer saline (pH 7.4). By virits tissue homogenizer, 1 gram of whole brain tissue was homogenised in 10 ml ice and 0.5 mol /L buffer of potassium phosphate having pH 7.4. For the biochemical analysis the homogenate was centrifuged at 4000 rpm for fifteen minutes for the harvesting of superananent. Homogenate was kept in ice packs throughout the procedure.
2.6. Acetyl cholinesterase activity in the brain region:
Acetyl cholinesterase activity was measured by using the method of [20] in which 0.1mM acetylthiocholine iodide as a substrate and DTNB (5, 5- DITHIOBIS-2 nitro benzoic acid) used as a colouring agent. Acetylthiocholine iodide degradation occurs at 412 nm and the result was measured as u mol ACTI hydrolysed per minute per mg protein.
2.7. Histological examination of tissues:
For the histology of brain tissues were fixed in a solution containing formalin, ethanol, and acetic acid was stored in 70 percent alcohol. 5 1mm section of tissues were embedded in paraffin and for the staining of slides haematoxylin and eosin stain was being used. Light microscope was used for the examining of tissues [21] as a reference, and photographed using a digital camera.
2.8. Neurochemical studies:
In the brain, biogenic amines and metabolites were determined. NE (norepinephrine) EPN non epinephrine DA dopamine, 5-HT (serotonin), HVA homovanillic acid and DOPAC 3,4- dihydroxyphenyl acetic acid in the frontal cortex of the brain were measured by reversed phase high performance liquid chromatography (HPLC) with the help of electrochemical detector by using the method of [22] with slight changes in his method. The high performance liquid chromatography (Waters, Melford, USA) consists of isocratic pumps. A sample injector valve, electrochemical detector, reverse phase column. The tissue of the frontal region of the brain was homogenised in 0.1 M perchloric acid having 3, 4-dihydroxybenzylamine, an internal standard the final concentration of 25ng ml was centrifuged at 36000 rpm for 10 minutes. After that the superanent was isolated and filtered through 0.25mm nylon filters and biogenic amines were determined from the filtered liquid. Twenty µl sample was injected into the injector port of HPLC and an electrochemical detector was activated at a potential of +0.800 V and maintaining the sensitivity of n A at ambient temperature. The obtained data was analysed by using the empower 2 software and result was shown as ng per gram tissue weight.
2.9. Oxidative stress activity in brain regions:
The activity of catalase in the brain tissue was checked spectrophotometrically by following the method of [23] hydrogen peroxide used as substrate and the value was measured in µmole per min per mg protein. By using the method of [24] the assay for superoxide dismutase was performed. And the obtaining results are expressed as units per mg protein. The method of [25] was used for glutathione estimation. The obtained results were expressed as nmol GSH oxidised per mg protein.
2.10. Statistical analysis:
All the parameters which are included in this research were carried out in triplicate for each group. The different observation was measured by using a student unpaired t-test. p < 0.05 and p < 0.01 significance level was used between the control and treated groups [26]. The observed results of treatment groups were also expressed as a percentage of control values (100%).
3. Results and Discussion
3.1. LD50 of bifenthrin
Figure 1.
The percentage mortality of O. cuniculus at different concentrations of bifenthrin during 96 hrs acute toxicity test.
Figure 1.
The percentage mortality of O. cuniculus at different concentrations of bifenthrin during 96 hrs acute toxicity test.
3.2. Body Weight and Somatic Index
The data presented in Table 1 showed significant decrease in the weight with respect to dose and the initial weight for G1, G2, G3, G4 and G5 (881, 880, 877, 875 and 872 gms) while weight after first week was recorded as (945, 942, 832, 818 and 802 gms) respectively. After 2nd week, weight of control and saline was increased while treated ones lost their weight as shown in Table 1.
The levels of biogenic amines in the cerebral cortex of rabbit’s brain were significantly affected after 14 days of exposure to bifenthrin. Rabbits' frontal brain was studied with three different doses at 3.5, 7 and 14 mg kg-1, significant reductions in the levels of EPN (252.66, 220.66 and 190.55) DA (404.6, 370.8 and 347.44), DOPAC (38.06, 26.07 and 17.09) and 5-HT (465.67, 395.73 and 3245.88±54) were observed as compared to the case of controls (control and saline). However, NE (327, 336 and 349) and HVA (33.37, 45.55 and 61.29) levels were found to be significantly higher than those of the controls. The levels of all biogenic amines were, however, comparable to those of controls 14 days after cessation of exposure shown in Table 2.
3.3. Acetyl cholinesterase activity
Compared to the case of controls (G1 and G2), significant reductions in the activity of the acetyl cholinesterase enzyme was found in the frontal cortex erythrocyte (3.6, 3.2 and 2.9), Plasma (0.55, 0.50 and 0.48), Brain (6.2, 5.7 and 5.0) of rabbits administered bifenthrin (3.5, 7 and 14 mg kg-1) as indicated in Table 3.
The data presented in (Table 4) showed significantly decrease in the activity of GPx (1.31, 1.06 and 0.57), SOD (1.10, 0.89 and 0.43), CAT (13.05, 11.83 and 5.71) and GST (48.74, 40.49 and 20.24) with the increase in dose of bifenthrin from 1/15th to 1/5th of LD50 in groups G3, G4 and G5 respectively as compared with the control groups G1 and G2.
3.4. Histopathological study
In histopathological study, rabbits treated with different doses of bifenthrin showed alteration in the brain tissue. Control groups (G1 and G2) showed normal GC (glial cells) and NC (neuronal cells) in the frontal cortex of brain tissue while the treated groups with three different doses (3.5 and 7 mg and 14 mg per kg body weight) of bifenthrin for two weeks showed evacuation (oedema), elongation of pyramidal nucleus, glial cell damage and brain haemorrhage as shown in Figure 2.
4. Discussion
The use of organophosphate and organochlorine was restricted and pyrethroid is commonly used due to limited persistence and low mammalian toxicity. Synthetic pyrethroid is currently used in the domestic and agriculture sector to control different pests [27]. This insecticide has the ability to generate ROS (Reactive oxygen species), oxidative and hydrolytic enzymes. Due to this mechanism, pyrethroids have the ability to metabolise in the mammalian metabolic system [28] and oxidative stress is induced due to the anti-oxidative enzymes like SOD, CAT and GPx [29].
The activities of SOD, GPx, and CAT were significantly decreased when treated with different doses of bifenthrin in comparison with those of the control. Our results showed a significant decrease in the activity of GPx, SOD, CAT and GST with the increase in dose of bifenthrin and these findings are matched with the observation of Syed et al. [22] and Gargouri et al. [30] who also observed similar activity of oxidative enzymes in their experiments with deltamethrin and lambda-cyhalothrin against rats due to production of superoxide and hydrogen peroxide free radicals in the in the tissues of brain.
In our study, the level of neurotransmitters and biogenic amines in frontal cortex of brain was reduced due to different doses of bifenthrin exposure. The level of epinephrine (EPN), dopamine (DA), 3, 4 dihydroxyphenylacetic acid (DOPAC) and serotonin (5-HT) was reduced in treated groups as compared with the control groups while the level neurotransmitter norepinephrine (Ne) and homovanillic acid (HVA) was significantly increased in treated groups as compared to the control groups. These results are at par with the results of Leutner et al. [31] who also observed the similar findings (the level of DA and metabolites DOPAC were significantly decreased and the level of other DA metabolites HVA were increased with respect to the dose of bifenthrin as compared to the control group). Syed et al. [22] observed in his experiments that the dopaminergic modulation was caused due to the induced drug which can be observed by locomotion and rearing frequency in open fields. He concluded that the insecticide bifenthrin damages the motor activity in rats and he correlated the locomotion and rearing frequency with the dopaminergic neurodegeneration.
Our study showed that the level of AChE in the brain, blood and erythrocytes was decreased due to the dose dependent pesticide treatment. These findings are in accordance with the findings of Shah & Gupta, [32] who also observed in their study that the permethrin disturbed the AChE enzyme. Rao & Rao, [33] also showed in their study that bifenthrin play a key role to interact aromatic, hydrophobic surface of AChE and it also inhibit the activity of AChE. Oxidative stress is also correlated to the activity of AChE [34]. For the better understanding of the effect of oxidant on AChE enzyme, free radicals are main cause of alteration in AChE enzyme.
Noeman et al. [35] and Dar et al. [36] also observed in their respective studies that the MDA cross linking and oxidative damage can also cause the inhibition of AChE enzyme due to the different pollutants like insecticides. The level of AChE in the brain, blood and erythrocytes was decreased with the increasing dose of bifenthrin, it indicates that bifenthrin has a higher tendency to inhibit AChE. Our results also match with the results of Khan et al. [37] and Yang and Li [38] who also observed the inhibition of voltage gated sodium channels and neuro excitatory effects caused due to the inhibition of AChE in rats when they were treated with insecticide bifenthrin.
The histopathological study of the brain in our results showed evacuation (oedema), elongation of pyramidal nucleus, glial cell damage and brain haemorrhage. Our findings were related with the study of [39] who observed in his study that insecticide bifenthrin causes brain lesion, congestion of blood vessels, evacuation and proliferation in neuron cells in the brain of rats when they were treated with insecticide bifenthrin. Britt et al. [40] observed in his study that when rhesus macaques were exposed with pesticide, organophosphorus showed histopathological alteration necrosis, gliosis, astrocytes, spongiosis and vascularization in neurons.
5. Conclusion:
It is concluded that the insecticide bifenthrin has the potential to cause neurotoxicity. It causes alteration in neurotransmitters, oxidative stress and histopathological changes. So it may be advised that proper care should be taken to avoid bifenthrin.
Conflicts of Interest
The Authors declare that there is no conflict of interest.
References
- Younas, A.; Naqvi, S. A.; Khan, M. R.; Shabbir, M. A.; Jatoi, M. A.; Anwar, F.; Aadil, R. M. Functional food and nutra-pharmaceutical perspectives of date (Phoenix dactylifera) fruit. Journal of Food Biochemistry. 2020, 44, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Ismail, W. I. W.; Radzi, M. N. F. M. Evaluation on the benefits of date palm (Phoenix dactylifera) to the brain. Alternative and Integrative Medicine. 2013, 5, 1–3. [Google Scholar]
- Weston, K. S.; Wisløff, U.; Coombes, J. S. High-intensity interval training in patients with lifestyle-induced cardiometabolic disease: a systematic review and meta-analysis. British Journal of Sports Medicine. 2014, 48, 1227–1234. [Google Scholar] [CrossRef]
- Wolansky, M. J.; McDaniel, K. L.; Moser, V. C.; Crofton, K. M. Influence of dosing volume on the neurotoxicity of bifenthrin. Neurotoxicology and Teratology. 2007, 29, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wu, N.; Wang, C. Toxicity of the pyrethroid bifenthrin insecticide. Environmental Chemistry Letters, 2020, 16, 1377–1391. [Google Scholar] [CrossRef]
- Cao, Z.; Shafer, T. J.; Crofton, K. M.; Gennings, C.; Murray, T. F. Additivity of pyrethroid actions on sodium influx in cerebrocortical neurons in primary culture. Environmental Health Perspectives. 2011, 119, 1239–1246. [Google Scholar] [CrossRef]
- Wolansky, M. J.; Gennings, C.; Crofton, K. M. Relative potencies for acute effects of pyrethroids on motor function in rats. Toxicological Sciences. 2006, 89, 271–277. [Google Scholar] [CrossRef]
- Mohan, G. K.; Pallavi, E.; Kumar, R.; Ramesh, M.; Venkatesh, S. Hepatoprotective activity of F. carica Linn leaf extract against carbon tetrachloride-induced hepatotoxicity in rats. Journal of Pharmaceutical Sciences, 2007, 15, 162–166. [Google Scholar]
- Dar, M. A.; Khan, A. M.; Raina, R.; Verma, P. K.; Wani, N. M. Effect of bifenthrin on oxidative stress parameters in the liver, kidneys, and lungs of rats. Environmental Science and Pollution Research, 2019, 26, 9365–9370. [Google Scholar] [CrossRef]
- Holt, M. P.; Ju, C. Mechanisms of drug-induced liver injury. The AAPS Journal. 2006, 8, 48–54. [Google Scholar] [CrossRef]
- Jaeschke, H.; Gores, G. J.; Cederbaum, A. I.; Hinson, J. A.; Pessayre, D.; Lemasters, J. J. Mechanisms of hepatotoxicity. Toxicological Sciences. 2002, 65, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Brander, S. M.; He, G.; Smalling, K. L.; Denison, M. S.; Cherr, G. N. The in vivo estrogenic and in vitro anti-estrogenic activity of permethrin and bifenthrin. Environmental Toxicology and Chemistry. 2012, 31, 2848–2855. [Google Scholar] [CrossRef] [PubMed]
- Colovic, M. B.; Krstic, D. Z.; Lazarevic-Pasti, T. D.; Bondzic, A. M.; Vasic, V. M. Acetylcholinesterase inhibitors: pharmacology and toxicology. Current Neuropharmacology. 2013, 11, 315–335. [Google Scholar] [CrossRef]
- Malomouzh, A.; Ilyin, V.; Nikolsky, E. Components of the GABAergic signaling in the peripheral cholinergic synapses of vertebrates: a review. Amino Acids. 2019, 51, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Zandona, A.; Katalinić, M. Nicotinic acetylcholine receptors: Diversity and physiological importance for neurodegenerative disorders and development of organophosphate antidotes. Periodicum Biologorum. 2020, 121, 115–128. [Google Scholar] [CrossRef]
- Osteen, C. D.; Fernandez-Cornejo, J. Economic and policy issues of US agricultural pesticide use trends. Pest Management Science. 2013, 69, 1001–1025. [Google Scholar] [CrossRef]
- Mofunanya, A. O. The effect of a common household pesticide, bifenthrin on neuronal cell survival and differentiation. Adelphi University. 2003.
- APHA. Standard methods for the examination of water and waste waters, 20th ed. American Public Health Association, Washington DC. 1998.
- Cao, Z.; Shafer, T. J.; Murray, T. F. Mechanisms of pyrethroid insecticide- induced stimulation of calcium influx in neocortical neurons. Journal of Pharmacology and Experimental Therapeutics. 2011, 336, 197–205. [Google Scholar] [CrossRef]
- Ellman, G. L.; Courtney, K. D.; Andres Jr, V.; Featherstone, R. M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochemical Pharmacology. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Takashima, F.; Hibiya, T. An atlas of fish histology: normal and pathological features. 1995.
- Syed, F.; Awasthi, K. K.; Chandravanshi, L. P.; Verma, R.; Rajawat, N. K.; Khanna, V. K.; Soni, I. Bifenthrin-induced neurotoxicity in rats: involvement of oxidative stress. Toxicology Research. 2018, 7, 48–58. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods in Enzymology. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. European Journal of Biochemistry. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Paglia, D. E.; Valentine, W. N. N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. Journal of Laboratory and Clinical Medicine. 1967, 70, 158–169. [Google Scholar] [PubMed]
- Ross, R. G.; Selvasubramanian, S.; Jayasundar, S. Immunomodulatory activity of Punica granatum in rabbits a preliminary study. Journal of Ethnopharmacology, 2001, 78, 85–87. [Google Scholar] [CrossRef] [PubMed]
- Silva, A. P. B.; Santos, J. M. M.; Martins, A.J. Mutations in the voltage-gated sodium channel gene of anophelines and their association with resistance to pyrethroids–a review. Parasite and Vectors. 2014, 7, 1–14. [Google Scholar] [CrossRef]
- Agrawal, A.; Sharma, B. Pesticides induced oxidative stress in mammalian systems. International Journal of Current Biological and Medical Science. 2010, 1, 90–104. [Google Scholar]
- Mekircha, F.; Chebab, S.; Gabbianelli, R. The possible ameliorative effect of Olea europaea L. oil against deltamethrin-induced oxidative stress and alterations of serum concentrations of thyroid and reproductive hormones in adult female rats. Ecotoxicology and Environmental Safety. 2018, 161, 374–382. [Google Scholar] [CrossRef]
- Gargouri, B.; Boukholda, K.; Kumar, A. Bifenthrin insecticide promotes oxidative stress and increases inflammatory mediators in human neuroblastoma cells through NF-kappa B pathway. Toxicology in Vitro. 2020, 65, 104792. [Google Scholar] [CrossRef]
- Leutner, S.; Eckert, A.; Müller, W. E. ROS generation, lipid peroxidation and antioxidant enzyme activities in the aging brain. Journal of Neural Transmission. 2001, 108, 955–967. [Google Scholar] [CrossRef]
- Shah, D.; Gupta, P. Fast updating algorithms for TCAM. IEEE Micro. 2001, 21, 36–47. [Google Scholar] [CrossRef]
- Rao, G. V.; Rao, J. K. S. Modulation in acetylcholinesterase of rat brain by pyrethroids in vivo and an in vitro kinetic study. Journal of Neurochemistry. 1995, 65, 2259–2266. [Google Scholar] [CrossRef] [PubMed]
- Bond, C. E.; Greenfield, S. A. Multiple Cascade Effects of Oxidative Stress on Astroglia. Glia. 2007, 55, 1348–1361. [Google Scholar] [CrossRef] [PubMed]
- Noeman, S. A.; Hamooda, H. E.; Baalash, A. A. Biochemical study of oxidative stress markers in the liver, kidney and heart of high fat diet induced obesity in rats. Diabetology and Metabolic Syndrome, 2011, 3, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Dar, M. A.; Khan, A. M.; Raina, R. Effect of repeated oral administration of bifenthrin antioxidant status and acetylcholinesterase activity in brain of rats. Environmental Toxicology and Chemistry, 2015, 97, 961–967. [Google Scholar] [CrossRef]
- Khan, A. M.; Rampal, S. Effects of repeated oral administration of pazufloxacin mesylate and meloxicam on the antioxidant status in rabbits. Journal of the American Association for Laboratory Animal Science, 2014, 53, 399–403. [Google Scholar]
- Yang, L.; Li, L. Actions of the pyrethroid insecticide bifenthrin on sodium channels expressed in rat cerebral cortical neurons. Toxicology Mechanisms and Methods. 2015, 25, 63–69. [Google Scholar] [CrossRef]
- Mahmoud, S.M.; Abdel Moneim, A.E.; Qayed, M.M.; El-Yamany, N.A. Potential role of N-acetylcysteine on chlorpyrifos-induced neurotoxicity in rats. Environmental Science and Pollution Research, 2019, 26, 20731–20741. [Google Scholar] [CrossRef]
- Britt, J. O.; Martin, J. L.; Okerberg, C. V.; Dick, E. J. Histopathologic changes in the brain, heart, and skeletal muscle of rhesus macaques, ten days after exposure to soman (an organophosphorus nerve agent). Comparative Medicine, 2002, 50, 133–139. [Google Scholar]
Figure 2.
Photomicrograph of brain tissue, A showed normal GC (glial cells) and NC (neuronal cells), B showed evacuation (odema), C showed elongation of pyramidal nucleus and glial cell damage, D showed the brain haemorrhage.
Figure 2.
Photomicrograph of brain tissue, A showed normal GC (glial cells) and NC (neuronal cells), B showed evacuation (odema), C showed elongation of pyramidal nucleus and glial cell damage, D showed the brain haemorrhage.
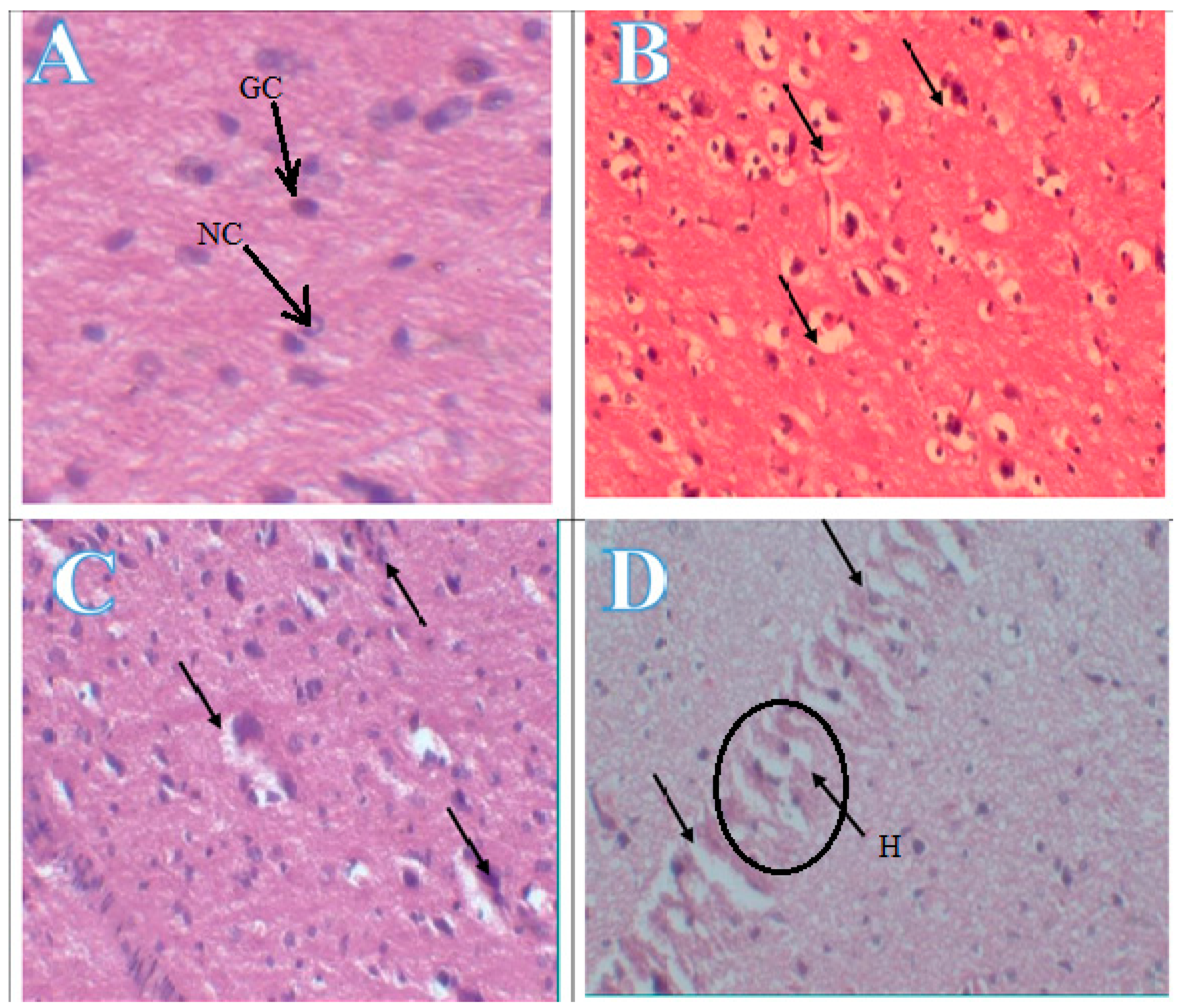

Table 1.
Weekly body weight (g) changes in rabbits exposed to bifenthrin.
| Weight in grams | Experimental Group | ||||
|---|---|---|---|---|---|
| G1 (Control) | G2 (Saline) | G3 | G4 | G5 | |
| Initial weight | 881±9.5 | 880±9.3 | 877±9.3 | 875±9.8 | 872±8.8 |
| 1stweeks | 945±9.7 | 942±9.2 | 832±9.2 | 818±8.1 | 802±9.3 |
| 2nd week | 1011±9.5 | 1005±9.1 | 782±9.1 | 758±8.2 | 705±8.5 |
Values are represented as an overall mean ± SE; n=5 for each treatment group at P<0.05.
Table 2.
Levels of biogenic amines and metabolites in the hippocampus of rabbits following treatment with bifenthrin.
Table 2.
Levels of biogenic amines and metabolites in the hippocampus of rabbits following treatment with bifenthrin.
| Brain region of hippocampus | Treatment for 14 days | P value | ||||
|---|---|---|---|---|---|---|
| G1 (Control) |
G2 (Saline) |
G3 (1/15th of LD50) | G4 (1/10th of LD50) | G5 (1/5th of LD50) | ||
| Norepinephrine | 317±0.66 | 315±0.65 | 327±0.73 | 336±0.44 | 349±0.44 | 0.00 |
| Epinephrine | 276.73±0.5 | 271.72±0.6 | 252.66±0.30 | 220.66±0.55 | 190.55±0.50 | 0.00 |
| Dopamine | 500.54±0.11 | 501.52±0.1 | 404.6±0.08 | 370.8±0.12 | 347.44±0.66 | 0.00 |
| 3,4 Dihydroxyphe nylacetic acid |
40.01±0.24 | 39.41±0.22 | 38.06±0.22 | 26.07±0.20 | 17.09±0.17 | 0.00 |
| Homovanillic acid | 30.66±0.44 | 31.86±0.43 | 33.37±0.5 | 45.55±0.55 | 61.29±0.66 | 0.00 |
| Serotonin | 505.66±0.77 | 503.96±0.6 | 465.67±0.7 | 395.73±0.6 | 324.88±0.57 | 0.00 |
Values are represented as an overall mean ± SE; n=5 for each treatment group. The significant difference is P<0.05.
Table 3.
Effect of repeated oral administration of bifenthrin on the activity of acetylcholinesterase in rabbits.
Table 3.
Effect of repeated oral administration of bifenthrin on the activity of acetylcholinesterase in rabbits.
| Parameters | Experimental Groups | P values | ||||
| G1 (Control) | G2 (Saline) | G3 | G4 | G5 | ||
| Erythrocyte | 4.0±0.01 | 3.86±0.02 | 3.6±0.05 | 3.2±0.04 | 2.9±0.03 | 0.00 |
| Plasma | 0.60±0.05 | 0.58±0.04 | 0.55±0.06 | 0.50±0.03 | 0.48±0.05 | 0.00 |
| Brain | 6.8±0.04 | 6.9±0.03 | 6.2±0.05 | 5.7±0.06 | 5.0±0.05 | 0.00 |
Values are represented as an overall mean ± SE; n=5 for each treatment group. The significant difference is P<0.05.
Table 4.
Antioxidant status of rabbits as affected by three different concentrations of bifenthrin.
| Parameters | Experimental Groups | P value | ||||
|---|---|---|---|---|---|---|
| G1 (Control) | G2 (Saline) |
G3 | G4 | G5 | ||
| GPx (U/L) | 2.04±0.05 | 2.00±0.04 | 1.31±0.06 | 1.06± 0.04 | 0.57±0.03 | 0.00 |
| SOD (U/ml) | 1.75±0.06 | 1.71±0.05 | 1.10±0.05 | 0.89± 0.04 | 0.43±0.03 | 0.00 |
| GST (U/L) | 74.99±0.07 | 72.89±0.05 | 48.74±0.07 | 40.49±0.06 | 20.24±0.04 | 0.00 |
| CAT (U/ml) | 20.40±0.04 | 19.72±0.02 | 13.05±0.03 | 11.83±0.04 | 5.71±0.03 | 0.00 |
Values are represented as an overall mean ± SE; n=5 for each treatment group. The significant difference is P<0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



