
Submitted:
19 September 2023
Posted:
20 September 2023
You are already at the latest version
Abstract
A new malachite beetle, Baltamauroania mirabilicornis gen. et sp. nov., belonging to the tribe Am-auroniodini (Coleoptera: Dasytidae) embedded in Eocene Baltic amber is described and illustrated. The new genus differs from the congeners of the tribe Amauroniodini in possessing a black, elongate and subparallel body and a pronotum with scalloped lateral sides; long and 11-segmented antennae, with three apical antennomeres enlarged and forming a ‘club’, with cylindrical an-tennomere 1; tarsi shortened and compressed, 5-segmented, with tarsomeres 1 and 2 equal in length, and tarsomere 5 the longest in all legs; pubescence of the dorsal surface consisting of short strong brown semi-erect and fine adpressed hairs; punctation of dorsal surface irregular, elytra lacking grooves; pronotum almost equilateral, slightly elongate, with acute anterior and obtuse posterior angles, with wide margination of basal side and with scalloped and finely margined lateral sides; lateral edges of pronotum weakly rounded, with obtuse protuberance before middle. This is the second extinct genus and third record of the subfamily Danaceinae found in Eocene amber. The fossil records of the family Dasytidae are discussed. Three recent records of different Amauroniodini taxa from Eocene East European amber (Baltic and Rovno) shows there to have been a high Danaceinae biodiversity in both the southern coast of the Subparatethys (documented by Rovno amber) and in its northern coast (documented by Baltic amber). The phenomenon of Danaceinae diversity in the Eocene Epoch is discussed.
Keywords:
Cleroidea
; Amauroniodini
; Baltamauroania
; new taxa
; fossil resin
; paleobiodiversity
; Cenozoic
1. Introduction
Members of the Malachite beetle family (Coleoptera, Dasytidae) are distributed worldwide in diverse landscapes, particularly in the Temperate Zone [1,2]. They are well known in amber inclusions as small to moderate sized (1 to 5 mm) beetles with a typically subparallel body, convex pronotum, epignathous head and dense double pubescence with long erect and short semierect hairs. In comparison with the closely related family Malachiidae, Dasytines can be recognized by well-sclerotized body, the thorax lacks evaginating vesicles, apical maxillary palpomere mostly conical, elytra mostly completely covering the entire abdomen, legs and antennae rarely modified, tarsomere 1 as long or longer than tarsomere 2 in all legs, and abdominal ventrites 1 and 2 free [1,3]. Representatives of these families in larval stage are known to be predators of small invertebrates occurring in soil litter [4]. The imagos are usually residents of flowers [4,5] in meadows and in shrubs in forested areas, but some genera such as Dasytes and Aplocnemus occur in trees [6,7,8].
The oldest record of the family Dasytidae, Protodasytes cretaceus Tihelka, Huang, Perrichot & Cai, 2021, relates to the Late Cretaceous Epoch and was described from Charentese amber originating from the sediments in the Charente-Maritime area of France [3]; according to geological dating, this amber is considered to be early Cenomanian within the Late Cretaceous Epoch, ca. 96 to 100.5 Ma [9,10]. A recent study [11] of poorly preserved remains of a Mesozoic Albian species Acanthocnemoides sukatshevae Zherikhin, 1977, described from Siberian Taimyr amber dated ca. 78 to 115 Ma, and previously attributed to the family Acanthocnemidae [12,13], resulted in its placement in the tribe Danaceini. This decision remains questionable and demands further study due to poorly visible main characters necessary to define family attribution of the beetle. Probably this species is a member of Acanthocnemidae or an ancestral form of this family which possesses special thermosensitive pores located in characteristic pit of prothorax hypomeron adjacent to sternopleural suture [14]. Traces of such a structure are visible in the hypomeron of Acanthocnemoides sukatshevae, but it is necessary to clarify if this is a natural structure or an artefact which appeared as result of chemical treatment of the amber and its inclusion. In this case, it is better to consider only Protodasytes cretaceus as a true Dasytidae reported from Cretaceous fossils.
The findings of Dasytidae in Charentese amber [3], Prionoceridae in Burmese amber [15] and Malachiidae in Burmese amber [16] provide evidence if an old history for these three families which have existed independently over at least 100 million years since the Cretaceous. Such a long history of independent evolution of these groups provides a new argument for systematic positioning of them into three separate families [17].
Representatives of Dasytidae are not rare in Eocene amber inclusions and also known from Florissant Fossil beds, Colorado, USA, where several new taxa have been described: three extinct genera, Eudasytites Wickham, 1912, Aploceble Majer, 1998 and Protomauroania Tshernyshev, 2021, one subgenus Aploceble (Chalcoaploceble) Tshernyshev, 2012, and 12 fossil species [18,19,20,21,22,23,24,25,26].
The majority of Eocene Dasytidae belong to extant genera, represented by the species with elongate, subparallel and slightly convex bodies which are densely covered with double pubescence. Usually, dasytid beetles registered in inclusions are with filiform (Dasytes) or serrate (Aplocnemus, Aploceble) antennae, but recently described Protomauroania Tshernyshev differs in its clubbed antennae [21]. This genus was attributed to the subfamily Danaceinae on the basis of clubbed antennae, scalloped lateral side of the pronotum and long, strong, reddish and semi-erect setae dorsally. Furthermore, the scalloped lateral edges of the pronotum places this genus in the tribe Amauroniodini. This was the first representative of Danaceinae from Baltic amber.
Recently, a small dasytid beetle possessing clubbed antennae was found in Baltic amber; its body is elongate and subparallel, whereas the pronotum has the scalloped lateral edges; the antennae are long, three apical antennomeres enlarged and forming a ‘club’; the tarsi are shortened and compressed, and tarsomere 1 is as long as tarsomere 2; the pubescence of the dorsal surface consists of short strong brown semi-erect and fine adpressed hairs. These brown hairs are disimilar from the adpressed scales typical of recent Danaceinae and can be considered as an ancestral character which has been modified during evolution to form scales as in Danacea species. The complex of characters mentioned above is close to Protomauroania Tshernyshev which was also described from Baltic amber, but Protomauroania differs in its small ovoid body which narrows towards the posterior, short antennae with friable club composed of six apical antennomeres, metatarsi with strongly elongated tarsomeres 1–3, short tarsomere 4 and relatively long tarsomere 5, and dorsal body surface densely covered with stiff, semi-erect, long brown setae. Obviously, the newly registered species belongs to a different genus, and both these two genera are different, but belong to one tribe of Danaceinae. This is the third record of this subfamily in the Eocene amber following those of Protomauroania chaetophorata Tshernyshev from Baltic amber [21] and P. mikhailovi Tshernyshev & Perkovsky, 2021 from Rovno amber [22].
Based on this complex of characters, Baltamauroania mirabilicornis gen. et sp. nov. is described and illustrated in the tribe Amauroniodini of the subfamily Danaceinae.
2. Materials and Methods
The specimen studied here is currently housed in the collection of Vitalii Alekseev (Kaliningrad, Russia) [collection prefix CVIA] and will ultimately be deposited in the Paleontological Institute of Russian Academy of Sciences (Moscow, Russia) for permanent preservation.
The amber piece with the inclusion was collected from the surf zone of the Baltic Sea coast near Mechnikovo village (the south-western Sambian peninsula, Kaliningrad region, Russia) on 20 June 2016. The amber piece was polished manually with emery papers of different grit sizes, allowing improved views of the included specimen. The beetle inclusion AWI-144 is preserved in a small, polished piece of transparent amber and subsequently embedded in a block of artificial resin by Hans Werner Hoffeins according to the technique described by Hoffeins [27].
Measurements of the holotype were made using an ocular micrometer in a stereoscopic microscope. Reconstruction was made based on free-hand drawings during an examination of the original specimen. The microphotographs were taken using a Canon EOS 4000D camera mounted on a Zeiss microscope. Extended depth of field at high magnifications was achieved by combining multiple images from a range of focal planes using Helicon Focus software. The resulting images were edited using Adobe Photoshop 7.0.
Nomenclatural acts introduced in the present work are registered in ZooBank (www.zoobank.org) under LSID urn:lsid:zoobank.org:pub:8BFC404A-E24D-46C3-8824-8B1AF65B5070.
3. Results
Systematics
Superfamily Cleroidea Latreille, 1802
Family Dasytidae Laporte de Castelnau, 1840
Subfamily Danaceinae C. G. Thompson, 1859
Tribe Amauroniodini Majer, 1987
Genus Baltamauroaniagen. nov.
LSID: urn:lsid:zoobank.org:act:72BB8249-538E-48C4-994A-BE4852BF0C9D
Type species: Baltamauroania mirabilicornis sp. nov., by present designation.
Diagnosis. Body black, elongate and subparallel, pronotum with scalloped lateral sides. Antennae 11-segmented, long, three apical antennomeres enlarged and forming a ‘club’, antennomere 1 cylindrical. Tarsi shortened and compressed, all tarsi 5-segmented, tarsomeres 1 and 2 of similar length, and tarsomere 5 the longest in all legs. Pubescence of the dorsal surface consists of short strong brown semierect and fine adpressed hairs. Punctation of dorsal surface irregular, elytra lacking grooves. Pronotum almost equilateral, slightly elongate, with acute anterior and obtuse posterior angles, with wide margination of basal side and scalloped and finely margined lateral edges, which are weakly rounded and provided with an obtuse protuberance below the middle of the disc.
Etymology. The name is formed from the name of the Baltic Region where the amber inclusion comes from and the name of a closely related extant genus Mauroania Majer, 1997. Gender feminine.
Comparison. Baltamauroania gen. nov. is similar to Protomauroania Tshernyshev, described from Baltic amber, but differs by the following combination of characters: subparallel and evenly rounded posteriorly body (ovoid and narrowed posteriorly body in Protomauroania), antennae elongate, with a loose 3-segmented club (antennae short with friable club composed of six antennomeres in Protomauroania), tarsi shortened and compressed, tarsomeres 1 and 2 of similar length in all legs (metatarsi with strongly elongated tarsomeres 1–3, a short tarsomere 4, and relatively long tarsomere 5 in Protomauroania), body evenly covered with short, strong, brown, semi-erect setae and fine adpressed hairs dorsally (body densely covered with stiff, semi-erect, long brown setae in Protomauroania).
Composition. Type species only.
Taxonomic assignment. The new fossil genus is placed in the family Dasytidae within the superfamily Cleroidea based on the the following combination of characters listed by Majer [28]: thorax lacking evaginating vesicles; body integument strongly sclerotized; abdominal ventrites lack membranes for contraction; bodies of adults are less flexible than the bodies of Malachiidae which have fast-opening elytra to initiate flight; body dorsal surface with erect or adpressed hairs, rarely with double pubescence; elytra are evenly rounded towards the apex and weakly flattened; most males lack special male characters indicating sexual dimorphism; tarsomeres 1 and 2 equal in length; claws short, without basal appendages; elytra lacking longitudinal carinae, with irregular punctation.
Baltamauroania mirabilicornis gen. et sp. nov. is assigned to the tribe Amauroniodini within the subfamily Danaceinae based on the complex of characters mentioned above (see Diagnosis).
Baltamauroania mirabilicornis sp. nov.
LSID: urn:lsid:zoobank.org:act:C658927F-9C1A-4FBE-A855-24EA8EF11362
Type material. Holotype: No AWI-144 [CVIA]; adult, male (due to strongly prominent "antennal club", i.e. dilated three apical antennomeres, and a shape of ultimate ventrite, which is narrow and truncate at apex so that space between the ventrite and the pygidium remains open).
. Almost complete beetle (right antennomeres 5–11 and apical segments of left protarsus were ground off during amber processing) is included in a small, transparent amber piece and subsequently embedded in a block of artificial resin to preserve inclusion. Syninclusions: absent.
Type locality. Baltic Sea coast near Mechnikovo village [former Neuhäuser], Sambian [Samland] peninsula, Kaliningrad region, W Russia, 20 June 2016, collector: VA. The collection site corresponds to the sampling sector 1.1 “Nodems” [34].
Description. Measurements: body length (i.e. length of elytra, pronotum and head along midline) 3.68 mm, body maximum width across elytra 1.25 mm; head length 0.38 mm, head maximum width at the level of eyes 0.55 mm; pronotum length 0.8 mm, pronotum maximum width 0.75 mm; elytra length 2.5 mm, elytra maximum width 1.25 mm. Body elongate, moderately convex; integument black lacking metallic lustre; surface densely covered with strong semi-erect brown setae and fine adpressed hairs.
Head partially retracted into thorax, transverse, about 1.6× wider (including eyes) than long; finely pubescent by semierect strong hairs; shallowly punctate, distance between punctures 1.0–1.5× diameter of one puncture, punctation distinctly denser on periphery. Frons convex. Frontoclypeal suture indiscernible. Mandible with bidentate apex. Compound eyes hemispherical, convex, medium-sized, shallowly impressed near antennal insertion, with fine facets, surface bare, lacking pubescence. Terminal maxillary palpomere large, broadly oval, pointed apically. Antennae inserted close to anterior margin of eyes; antennal insertions exposed in dorsal view, widely separated. Antennae 11-segmented, with loose, 3-segmented club, comparatively long, reaching slightly beyond one-fourth of elytron; all antennomeres elongate, 2.0–2.5× longer than wide; with long semierect pubescence; antennomere 1 subcylindrical; antennomeres 2–8 conical, slightly dilated apically; antennomeres 9 and 10 widened, conical, dilated apically, antennomere 9 about 1.4× wider than antennomere 8; antennomere 11 spindle-shaped, narrowly rounded apically; antennomeres 9–11 densely covered by short and fine pubescence; relative length ratios of antennomeres 1–11 equal to 10:10:7:7:7:7:7:6:10:9:9.
Pronotum weakly elongate 1.1× longer than wide, narrowed both at base and distally, at base distinctly narrower than elytral base; with the maximal width at the anterior third. Pronotal disc convex. Posterior side strongly and anterior finely marginate, lateral sides scalloped and finely margined. Punctation on pronotum dense, slightly more widely spaced than on head, distance between punctures equal to 0.5–1.0× diameter of one puncture. Posterior angles obtuse; anterior angles acute. Anterior edge weakly rounded, without excision; posterior edge evenly arcuate; lateral edges weakly rounded, with obtuse protuberance before middle, almost straight in posterior two-thirds. Prosternum short anterior of procoxae; with convex disc; sparsely punctate. Prostenal process flat, narrow.
Scutellar shield large, semicircular, as wide as long, finely punctate and pubescent. Elytra elongate-oval, slightly widened posteriorly, moderately convex, with maximum width in middle, 2.0× longer than wide, almost 3.1× as long as pronotum length; completely covering the abdomen; lateral sides slightly rounded; humeri distinct, slightly prominent. Elytral punctation irregular, coarse and dense; punctures in basal part similar diameter or slightly larger as punctures on pronotum; distance between punctures equal to 0.5–1.5× diameter of one puncture. Epipleura narrowed posteriorly, incomplete, extending to about level of abdominal ventrite 2. Mesocoxal cavities subovate, narrowly separated, closed laterally. Meso-metaventral junction straight. Metaventrite with disc convex; covered with coarse, rather dense punctation.
Legs slender, long, relatively similar in shape, finely punctate and pubescent. Procoxae large, widely sub-oval, transverse; mesocoxae rounded; metacoxae narrowly oval, strongly transverse. Femora narrowly spindle-shaped, weakly flattened. Tibiae thin, almost straight, slightly dilated apically, nearly as long as femora. Tarsal formula 5-5-5. Tarsi long and thin; tarsomere 1 bilobed, tarsomeres 2–4 simple. In all legs tarsomere 1 depressed; meso- and metatarsomeres compressed laterally. Claws strong, curved, simple, without appendages, symmetrical.
Abdomen with five visible, freely articulated ventrites, finely punctate and pubescent; intercoxal process rather wide; ventrite 1 longest; ventrite 5 wide and rounded. Relative length ratios of ventrites 1–5 (medially) equal to 25:15:12:10:12.
Note. The character state of procoxal cavities (closed or open) is not visible in the examined specimen due to the positioning of the legs in the inclusion.
Etymology. The specific epithet mirabilicornis is a compound Latin adjective formed from “mirabilis” (strange, wonderful, marvellous) and “cornu” (horn, acute apex), meaning “with wonderful antennae” in reference to the extremely unusual elongate antennal club of the beetle.
Remark. The included beetle represents an example of stressed behaviour in resin [35], namely defecation.
4. Discussion
Representatives of the superfamily Cleroidea that include the melyrid lineage of families Melyridae, Dasytidae and Rhadalidae are well known from the Eocene, the second of three major worldwide divisions of the Paleogene. Beetles have already been described from Baltic [8,18,19,20,21,36,37,38,39,40,41,42], Rovno [22,40] and French [43] amber and from imprints in Florissant deposit [23,24,25,26,44]. Melyridae lineage of the Cleroidea is reported from Cretaceous Charentese [3] and Burmese amber [45,46,47] and from imprints found in Liaoning deposit (Upper Jurassic-Lower Cretaceous, Yixian, some 145 Ma) from Inner Mongolia [48].
As mentioned above, amongst the Melyridae-group families, species of the family Dasytidae are more usual in amber inclusions, and several new taxa have been described during the past 100 years: four extinct genera, Eudasytites Wickham, 1912, Aploceble Majer, 1998 (with the subgenus Aploceble (Chalcoaploceble) Tshernyshev, 2012), Protomauroania Tshernyshev, 2021 and Protodasytes Tihelka et al., 2021, and 13 species: Eudasytites listriformis Wickham, 1912, Eutrichopleurus miocenus Wickham, 1917, Aploceble (Aploceble) berendti Majer, 1998, A. (A.) fuscipes Majer, 1998, A. (A.) kunowi Majer, 1998, A. (A.) lateantennata Tshernyshev, 2019, A. (Chalcoaploceble) viridiaeneus Tshernyshev, 2012, Aplocnemus (A.) hoffeinsorum Tshernyshev, 2019, A. (Ischnopalpus) baltiensis Tshernyshev, 2019, Dasytes (Mesodasytes) ochraceus Tshernyshev, 2019, Protomauroania chaetophorata Tshernyshev, 2021, P. mikhailovi Tshernyshev et Perkovsky, 2021, and Protodasytes cretaceus Tihelka et al., 2021 [3,18,19,20,21,22,23,26].
Furthermore, based on external morphology, some fossil beetles were assigned to the extant genera: Microjulistus sp., Aplocnemus sp. and Psilothrix sp., Xamerpus sp., Malthacodes sp., Pelecophora sp. and Aplocnemus cf. tarsalis C.R. Sahlberg, 1822, and three species closely related to Dasytes sp. [18,37]. Majer [18] also identified three inclusions as incertae sedis and noted that they belong to a separate genus closely related to Xamerpus.
An extinct genus Eudasytites Wickham and two species E. listriformis Wickham and Eutrichopleurus miocenus (Wickham) were described from Upper Eocene–Oligocene shale deposits in Florissant Fossil beds, Colorado, USA. The impressions were poorly preserved and indistinct, without legs and antenna. Wickham [23] referred Eudasytites to Dasytidae since the beetle was imprinted from the ventral side and had no distinct male specific structures on the elytra or abdomen, and therefore could not be placed in Malachiidae. The characters given for the new genus were: “slender build and coarse sculpture, with vestiture inconspicuous or wanting” [23]. The impressions of the other species contrariwise was from dorsal side with distinct elytra and regular punctured rows. This character is close to the recent species Trichochrous seriellus, and fossil species described as T. miocenus Wickham. Later, the species was transferred to the genus Eutrichopleurus Blaisdell, 1938 [44]. All other Eocene fossil of Dasytidae were found in Baltic and Rovno amber. It should be noted that catalogues [23,37,38,39,49,50,51] provide details of effective analysis of impressions and inclusions of fossil insects.
The oldest records of the family Dasytidae relate to two species: Protodasytes cretaceus Tihelka, Huang, Perrichot & Cai, 2021, described from Charentese amber of the Late Cretaceous Epoch [3] and Acanthocnemoides sukatshevae Zherikhin, 1977 described from Siberian Taimyr amber of the Mesozoic Albian [11,12].
The genus Protodasytes Tihelka, Huang, Perrichot et Cai, 2021 with the type species Protodasytes cretaceus Tihelka, Huang, Perrichot et Cai, 2021, was described from Charentese amber discovered in sediments in the Charente-Maritime area of France. This is the oldest record of Dasytidae and the first record in the Cretaceous of Europe [3]. According to geological dating, this amber age is considered as early Cenomanian of the Late Cretaceous Epoch (c. 96 to 100.5 Ma) [9,10]. The external appearance of the beetle in amber, reconstructed using high-resolution X-ray microtomography (micro-CT), looks very similar to Acantocnemus, namely: shape of the body, black colouration, irregular dense punctation of elytra with typically widened lateral sides in a middle, antennae short with 3-segmented club, legs slender with short and compressed tarsi, also prothorax hypomeron adjacent to sternopleural suture possesses an elongate pit noticeable in the reconstructed picture [3, fig. 2B]. This species differs in having longitudinal grooves in lateral sides of the pronotum, typical for the tribe Chaetomalachiini, but was probably used as a primitive thermosensor organ, typical of Acanthocnemidae. Working with the characters untypical to any tribes, Tihelka et al [3] decided to regard this species “as a fossil link between the two tribes Dasytini and Listrini” and to treat the species “as tribe incertae sedis“.
Recently, the poorly preserved remains of the Mesozoic Albian species Acanthocnemoides sukatshevae Zherikhin, 1977 were also re-examined to clarify the attribution of the species to the family Acanthocnemidae [11]. This species was described in Siberian Taimyr amber dated ca. 78 to 115 Ma [52], as a member of the family Acanthocnemidae [12]. To make such a decision in this case was very difficult, since the poorly preserved inclusion was essentially only a black contour of the beetle which masked essential characters such as tarsomeres and elytral punctation. The only character, the slender antenna with a small club, made it possible to consider this species as belonging to Danaceinae of the family Dasytidae. Kolibač and Perkovsky [11] transferred this species from Acanthocnemidae to Dasytidae, but their decision remains questionable and demands further study of the beetle remains. Very probably, this species is really a member of Acanthocnemidae or an ancestral form of this family which possesses special thermosensitive pores located in a characteristic pit of the prothorax hypomeron adjacent to the sternopleural suture [14].
The largest number of Eocene Dasytidae were probably residents of mixed forests due to repeated co-occurence of beetles with stellate fagacean trichomes [20, and original personal observation]. Eurasian malachite beetles of the Recent mainly occur in forest and forest-steppe habitats [6] and, probably, similar to those they inhabited in Paleogene. Dasytid species can be found in coniferous and mixed forests, and, probably, fossil species also occurred in Eocene biocenoses with coniferous trees. Moving to open landscapes Dasytidae occupied soil litter whereas Malachiidae used new type of plant communities appeared to find out ecological niches in plants.
Dasytidae species known from Eocene fossils are belonging both to extinct and extant genera [8. Beetles with characteristic elongate, subparallel, slightly convex body densely covered with double pubescence differ from other similar habitus beetles in possessing filiform (Dasytes) or serrate (Aplocnemus, Aploceble) antennae. These inclusions can be easily attributed to subfamilies Rhadalinae LeConte, 1862 or Dasytinae Laporte de Castelnau, 1840. That is why malachite beetles with clubbed antennae newly discovered in amber inclusions can lead to misidentifications and thereby assigned to Cucujoidea families. After a more detailed morphological study it is clear that these beetles belong to the Dasytidae subfamily Danaceinae Thomson, 1859. Representatives of this subfamily have demonstrated a high level of evolution adaptation and morphological changes. Typically, this subfamily, represented by small and slightly elongate species densely covered with adpressed scales or scale-shaped chaetae, which prevent drying and overheating, were capable of adapting to open landscapes. The first record of Dasytinae in Baltic amber was the genus Protomauroania Tshernyshev [21]. This is small ovoid beetle with narrowed posteriorly elytra, clubbed antennae, scalloped lateral side of the pronotum and long, strong, reddish, semi-erect setae on the upper side of the body. Obviously, this is an ancestral form of recent Danaceinae demonstrating an initial development of their dense cover as strong semi-erect setae, which was probably modified afterwards to flat scales in Danacea. The scalloped lateral edges of the pronotum made it possible to place this new extinct genus in the tribe Amauroniodini close to the recent genus Amauronia Westwood, 1839, for which there was formerly no ancient history evidence of genera of this subfamily. It was the first description of a new extinct genus and species of Danaceinae from Baltic amber. Later, one more species of the genus, P. mikhailovi Tshernyshev et Perkovsky, 2021, was described from Rovno amber [22]. These records in different amber confirm the widespread and species richness of Danaceinae representatives of Eocene landscapes.
The new genus and species of Dasytinae, Baltamauroania mirabilicornis gen. et sp. nov., described herein from Baltic amber, possesses clubbed antennae and pronotum with scalloped lateral sides, but differs in long antennae with only three apical antennomeres which are enlarged and club-shaped, the shortened and compressed tarsi with tarsomeres 1 and 2 equal in length, and pubescence of the dorsal surface consisting of short strong brown erect and fine adpressed hairs. Two different genera, Protomauroania Tshernyshev and Baltamauroania gen. nov., confirms the diversity of Eocene Danaceinae at the generic level.
Characters revealed in beetles described from inclusions are considered as typical for the tribe Amauroniodini, and, probably, illustrate an ancestral form for all other representatives of Danaceinae, which at present includes three tribes and 20 genera:
1. Tribe Amauroniodini Majer, 1987 (Amauronia Westwood, 1839, Baltamauroania gen. nov., Indamauronia Majer, 1997, Mauroania Majer, 1997, Perisphinctus Schilsky 1898, Protomauroania Tshernyshev, 2021 and Pseudamauronia Pic, 1915);
2. Tribe Danacaeomimini Majer, 1987 (Cyrdobaldus Majer, 1989, Cyrdothrix Majer, 1989, Danacaeomimus Champion, 1922, Piechockia Majer, 1986, Schoenia Majer, 1989;
3. Tribe Danaceini Kiesenwetter, 1863 (Amecocerus Solier, 1849, Danacea Laporte de Castelnau, 1836, Haplamaurus Kirsch, 1865, Hylodanacaea Pic, 1926, Listrocerus Majer, 1998, Setigerodasytes Pic, 1910, Bilyella Majer, 1988 and Hoppingiana Blaisdell, 1924).
Three recent records of different Amauroniodini taxa from Eocene East European amber (Baltic and Rovno) demonstrate the phenomenon of a high Danaceinae biodiversity in both the southern coast of the Subparatethys (documented by Rovno amber) and on the more northern Fennosarmatian landmass (documented by Baltic amber).
5. Conclusions
The newly described genus and species, Baltamauroania mirabilicornis gen. et sp. nov., is the third record of the subfamily Danaceinae in Eocene amber, following those of Protomauroania chaetophorata Tshernyshev in Baltic amber and P. mikhailovi Tshernyshev et Perkovsky in Rovno amber. The discovery of two different genera and at least three distinct species in different ambers confirms the widespread and high level of diversity of Danaceinae in Eocene amberiferous forest landscapes of the Fennosarmatian landmass and in the southern coast of the Subparatethys. This phenomenon of Danaceinae diversity in Eocene fossil remains has only been recorded during the past three years.
Different strategies of adaptation in ecosystems dictate morphology and bionomy of beetles, but initially they were probably connected with trees. The fact that Dasytidae are found in Charentese amber [3], Prionoceridae in Burmese amber [15] and Malachiidae in Burmese amber [16] is clear evidence of the ancient history of these families, which has sustained their independent existence over 100 million years since the Cretaceous to the present day. Such a long existence of independent groups provides a new argument for the systematic positioning of Dasytidae and Malachiidae as separate families, as previously done by Majer [1], Bocakova et al. [17], and a counterargument for considering them as subfamilies within a single diverse family Melyridae together with Melyridae s.str., as proposed by Lawrence and Leschen [4] and Gimmel et al. [53].
Author Contributions
Conceptualization, V.I.A. and S.E.T.; writing—original draft preparation, review and editing, S.E.T., G.Yu.L., V.I.A. and A.B.; visualization, S.E.T, V.I.A. and A.B.; project administration, S.E.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The specimen is deposited in the collection of Vitalii Alekseev (Kaliningrad) [collection prefix CVIA] and will ultimately be deposited in the Paleontological Institute of Russian Academy of Sciences (Moscow, Russia) for permanent preservation.
Acknowledgments
We are grateful to Prof. Mark Seaward (Bradford University, U.K.) for his most helpful advice and linguistic revision of the text. We are sincerely grateful to Mr Hans-Werner Hoffeins (Germany, Hamburg) for the conservation of the studied inclusion. We thank anonymous reviewers for their helpful comments and efforts to improve our manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Majer, K. A review of the classification of the Melyridae and related families (Coleoptera, Cleroidea). Entomol. Basil. 1994, 17, 319–390. [Google Scholar]
- Mayor, A.J. Family Malachiidae Fleming, 1821. In Catalogue of Palaearctic Coleoptera, Volume 4. Elateroidea, Derodontoidea, Bostrichoidea, Lymexyloidea, Cleroidea, Cucujoidea; Lobl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2007; pp. 415–454. [Google Scholar]
- Tihelka, E.; Huang, D.; Perrichot, V.; Cai, Ch. A previously missing link in the evolution of dasytine soft-winged flower beetles from Cretaceous Charentese amber (Coleoptera, Melyridae). Pap. Palaeontol. 2021, 7, 1753–1764. [Google Scholar] [CrossRef]
- Lawrence, J.F. ; Leschen. R., Beutel, R., Eds; De Gruyter: Berlin/New York, 2010; pp. 273–280.1815. In Handbook of zoology – Coleoptera. Volume 2; Leschen., R., Beutel, R., Eds, *!!! REPLACE !!!*, Eds.; De Gruyter: Berlin/New York, 2010; De Gruyter: Berlin/New York, 2010; pp. 273–280. [Google Scholar]
- Mawdsley, J.R. The importance of species of Dasytinae (Coleoptera: Melyridae) as pollinators in Western North America. Coleopt. Bull. 2003, 57, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Tshernyshev, S.E. A review of oedemerid and dasytid beetle fauna (Coleoptera, Oedemeridae, Melyridae, Dasytidae) of North Asia. Euroasian Entomol. J. 2017, 16, 554–565. [Google Scholar]
- Mirutenko, V.V. The beetles of Malachiidae and Dasytidae families Coleoptera) in the collections of Zoological museum and Department of entomology and biodiversity reserve of Uzhhorod university. Sci. Bull. Uzhgorod Uni. (Ser. Biol.) 2008, 23, 209–219. [Google Scholar]
- Tshernyshev, S.E. Evolution potential of eocene entomofaunas in formation of contemporary diversity of Coleoptera of the families Malachiidae and Dasytidае sensu latum. In Conceptual and applied aspects of scientific research and education in the field of invertebrate zoology: сollection of articles V International conference; Bagirov, R.T-o., Maximova, Yu.V., Subbotina, E.Yu., Shcherbakov, M.V., Simakova, A.V., Eds.; Tomsk State University Press: Tomsk, 2020; pp. 178–182. [Google Scholar]
- Néraudeau, D.; Allain, R.; Perrichot, V.; Videt, B.; De Broin, F.d.L.; Guillocheau, F.; Philippe, M.; Rage, J.-C.; Vullo, R. Découverte d'un dépôt paralique à bois fossiles, ambre insectifère et restes d'Iguanodontidae (Dinosauria, Ornithopoda) dans le Cénomanien inférieur de Fouras (Charente-Maritime, Sud-Ouest de la France). C. R. Palevol, 2003; 2, 221–230. [Google Scholar]
- Perrichot, V.; Néraudeau, D.; Tafforeau, P. Charentese amber. In Biodiversity of fossils in amber from the major world deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, 2010; pp. 192–208. [Google Scholar]
- Kolibač, J.; Perkovsky, E.E. A reclassification of Acanthocnemoides sukatshevae Zherikhin, 1977 from the mid-Cretaceous Taimyr amber (Coleoptera). Cretac. Res. 2020, 115, 104548. [Google Scholar] [CrossRef]
- Zherikhin, V.V. Series of superfamilies of Cucujiformia. In Mezozoiskie zhestkokrylye; Arnoldi, L.V., Zherikhin, V.V., Nikritin L.M., Ponomarenko A.G., Eds.; Trudy Paleontologicheskogo Instituta 1977, 161; pp. 135–142.
- Zherikhin, V.V. Series of superfamilies of Cucujiformia. In Mesozoic Coleoptera; Arnoldi, L.V., Zherikhin, V.V., Nikritin, L.M., Ponomarenko, A.G., Eds.; Oxonian Press Pvt. Ltd.: New Delhi, 1992; pp. 182–191. [Google Scholar]
- Schmitz, H.; Schmitz, A.; Trenner, S.; Bleckmann, H. A new type of insect infrared organ of low thermal mass. Naturwissenschaften 2002, 89, 226–229. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, Z.; Yu, Y.; Shih, C.; Ślipiński, A.; Ren, D. First species of Prionoceridae from mid-Cretaceous amber of northern Myanmar (Coleoptera: Cleroidea). Cret. Res. 2021, 133, 105118. [Google Scholar] [CrossRef]
- Tshernyshev, S.E.; Legalov, A.A. First record of the family Malachiidae (Coleoptera: Cleroidea) from mid-Cretaceous Burmese amber with a description of Burmalachius acroantennatus gen. et spec. nov. // Life, 2023, in litt.
- Bocakova, M.; Constantin, R.; Bocak, L. Molecular phylogenetics of the melyrid lineage (Coleoptera: Cleroidea). Cladistics 2012, 28, 117–129. [Google Scholar] [CrossRef]
- Majer, K. Rhadalinae from the Baltic amber (Coleoptera, Dasytidae). Dtsch. Entomol. Z. (N.F.) 1998, 45, 255–264. [Google Scholar] [CrossRef]
- Tschernyshev, S.E. Aploceble (Chalcoaploceble) viridiaeneus Tshernyshev – new subgenus and species of dasytid beetles (Coleopterta, Dasytidae) in Baltic amber. Euroasian Entomol. J. 2012, 11, 213–218. [Google Scholar]
- Tshernyshev, S.E. New species of malachite beetles (Coleoptera, Dasytidae) in Baltic amber. Paleontol. J. 2019, 53, 1024–1035. [Google Scholar] [CrossRef]
- Tshernyshev, S.E.; Shcherbakov, M.V. A new genus and species of malachite beetles (Coleoptera, Dasytidae) in Baltic amber. Paleontol. J. 2021, 55, 174–178. [Google Scholar] [CrossRef]
- Tshernyshev, S.E.; Perkovsky, E.E. Protomauroania mikhailovi – a new species of malachite beetles (Coleoptera, Dasytidae) in Rovno Amber. Zootaxa 2021, 5006, 189–194. [Google Scholar] [CrossRef]
- Wickham, H.F. A report of some recent collections of fossil Coleoptera from the Miocene shales of Florissant. Bull. Lab. Natl. Hist., State Univ. Iowa 1912, 6, 1–38. [Google Scholar]
- Wickham, H.F. Twenty new Coleoptera from the Florissant Shales. Trans. Am. Entomol. Soc. 1914, 40, 257–270. [Google Scholar]
- Wickham, H.F. New fossil Coleoptera from the Florissant beds. Bull. Lab. Natl. Hist. State Univ. Iowa 1916, 7, 3–20. [Google Scholar]
- Wickham, H.F. New species of fossil beetles from Florissant, Colorado. Proc. US Nat. Mus. 1917, 52, 463–472. [Google Scholar] [CrossRef]
- Hoffeins, H.W. On the preparation and conservation of amber inclusions in artificial resin. Pol. J. Entomol. 2001, 70, 215–219. [Google Scholar]
- Majer, K. Comparative morphology and proposed major taxonomy of the Family Melyridae (Insecta, Coleoptera). Pol. J. Entomol. 1987, 56, 719–859. [Google Scholar]
- Sadowski, E.-M.; Seyfullah, L.J.; Schmidt, A.R.; Kunzmann, L. Conifers of the ‘Baltic amber forest’ and their palaeoeco-logical significance. Stapfia, 2017, 106, 1–73. [Google Scholar]
- Sadowski, E.-M.; Schmidt, A.R.; Denk, T. Staminate inflorescences with in situ pollen from Eocene Baltic amber reveal high diversity in Fagaceae (oak family). Willdenowia, 2020, 50, 405–517. [Google Scholar] [CrossRef]
- Seyfullah, L.J.; Beimforde, C.; Dal Corso, J.; Perrichot, V.; Rikkinen, J.; Schmidt, A.R. Production and preservation of resins – past and present. Biol. Rev. 2018, 93, 1684–1714. [Google Scholar] [CrossRef]
- Bukejs, A.; Alekseev, V.I.; Pollock, D.A. Waidelotinae, a new subfamily of Pyrochroidae (Coleoptera: Tenebrionoidea) from Baltic amber of the Sambian peninsula and the interpretation of Sambian amber stratigraphy, age and location. Zootaxa 2019, 4664, 261–273. [Google Scholar] [CrossRef]
- Kasiński, J.R.; Kramarska, R.; Słodkowska, B.; Sivkov, V.; Piwocki, M. Paleocene and Eocene deposits on the eastern margin of the Gulf of Gdańsk (Yantarny P-1 bore hole, Kaliningrad region, Russia). Geol. Q. 2020, 64, 29–53. [Google Scholar] [CrossRef]
- Bukejs, A.; Alekseev, V.I.; Legalov, A.A. A new Eocene genus of the subtribe Tylodina (Coleoptera: Curculionidae) and notes concerning local differences of Baltic amber in the Kaliningrad Region. Foss. Rec. 2020, 23, 75–81. [Google Scholar] [CrossRef]
- Arillo, A. Paleoethology: fossilized behaviours in amber. Geol. Acta 2007, 5, 159–166. [Google Scholar]
- Berendt, G.C. Die organischen Bernstein-Einschlüsse im Allgemeinen. In Der Bernstein und die in ihm befindlichen Pflanzenreste der Vorwelt. Bd.1; Goeppert, H.R., Berendt, G.C., Eds.; Nicolai: Berlin, 1845; pp. 41–60. [Google Scholar]
- Klebs, R. Über Bernsteineinschlüsse im allgemeinen und die Coleopteren meiner Bersteinsammlung. Schr. Phys.-Okon. Ges. Königsberg, 1910, 51, 217–242. [Google Scholar]
- Spahr, U. Bibliographie der Bernstein- und Kopal-Käfer (Coleoptera). Stutt. Beitr. Natukd. Ser. B 1981, 72, 1–22. [Google Scholar]
- Spahr, U. Systematischer Katalog der Bernstein- und Kopal-Käfer (Coleoptera). Stutt. Beitr. Natukd. Ser. B 1981, 80, 1–107. [Google Scholar]
- Tshernyshev, S.E. New Taxa of Soft-winged Flower Beetles (Coleoptera, Malachiidae) in Baltic and Rovno Amber. Paleontol. J. 2016, 50, 953–962. [Google Scholar] [CrossRef]
- Tshernyshev, S.E. New taxa of soft-winged flower beetles (Coleoptera, Malachiidae) in Baltic Amber. Paleontol. J. 2020, 54, 67–75. [Google Scholar] [CrossRef]
- Kubisz, D. Beetles in the collection of the Museum of Amber Inclusions, University of Gdansk, with description of Colotes sambicus sp. n. (Coleoptera: Melyridae). Pol. J. Entomol. 2001, 70, 259–265. [Google Scholar]
- Kirejtshuk, A.G.; Nel, A. New beetles of the suborder Polyphaga from the lowermost Eocene French amber (Insecta: Coleoptera). Ann. Soc. Entomol. Fr. 2008, 44, 419–442. [Google Scholar] [CrossRef]
- Mawdsley, J.R. Review of the extant and fossil Dasytinae (Coleoptera: Melyridae) of Colorado, U.S.A. Trans. Am. Entomol. Soc. Philadelphia 1999, 125, 251–267. [Google Scholar]
- Rasnitsyn, A.P.; Ross, A.J. A preliminary list of arthropod families present in the Burmese amber collection at The Natural History Museum, London. Bull. Nat. Hist. Mus. Lond. (Geol.) 2000, 56, 21–24. [Google Scholar]
- Ross, A.; Mellish, C.; York, P.; Craighton, B. Chapter 12: Burmese amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, 2010; pp. 116–136. [Google Scholar]
- Ross, A.J. Burmese (Myanmar) amber checklist and bibliography 2018. Palaeoentomology 2019, 2, 22–84. [Google Scholar] [CrossRef]
- Kirejtshuk, A.G.; Ponomarenko, A.G.; Prokin, A.A.; Chang, H.; Nikolajev, G.V.; Ren, D. Current knowledge on Mesozoic Coleoptera from Daohugou and Liaoning (North East China). Acta Geol. Sin. 2010, 84, 783–792. [Google Scholar] [CrossRef]
- Handlirsch, A. IV. Abschnitt. Tertiäre Insekten. In Die fossilen Insekten und die Phylogenie der rezenten Formen: Ein Handbuch für Paläontologen und Zoologen; Handlirsch, A., Ed.; Wilhelm Engelman: Leipzig, 1907; pp. 673–1120. [Google Scholar]
- Bachofen-Echt, A. Der Bernstein und seine Einschlüsse; Springer Verlag: Wien, 1949. [Google Scholar]
- Larsson, S.G. Baltic Amber - a palaeobiological Study. Entomonograph 1; Scandinavian Science Press Ltd.: Klampenborg, Denmark, 1978. [Google Scholar]
- Perkovsky, E.E.; Vasilenko, D.V. A summary of recent results in the study of Taimyr amber. Paleontol. J. 2019, 53, 984–993. [Google Scholar] [CrossRef]
- Gimmel, M.L.; Bocakova, M.; Gunter, N.L.; Leschen, R.A.B. Comprehensive phylogeny of the Cleroidea (Coleoptera: Cucujiformia). Syst. Entomol. 2019, 44, 527–558. [Google Scholar] [CrossRef]
Figure 1.
Photomicrographs of Baltamauroania mirabilicornis gen. et sp. nov., holotype, No AWI-144 [CVIA], habitus: (A) Dorso-lateral view; (B) Ventral view. Scale bars represent 1 mm.
Figure 1.
Photomicrographs of Baltamauroania mirabilicornis gen. et sp. nov., holotype, No AWI-144 [CVIA], habitus: (A) Dorso-lateral view; (B) Ventral view. Scale bars represent 1 mm.

Figure 2.
Baltamauroania mirabilicornis gen. et sp. nov., holotype, No AWI-144 [CVIA]: (A) Schematic drawing of the habitus in ventral view; (B) Schematic drawing of the lateral margin of the pronotum; (C) Reconstruction of the habitus in dorsal view.
Figure 2.
Baltamauroania mirabilicornis gen. et sp. nov., holotype, No AWI-144 [CVIA]: (A) Schematic drawing of the habitus in ventral view; (B) Schematic drawing of the lateral margin of the pronotum; (C) Reconstruction of the habitus in dorsal view.
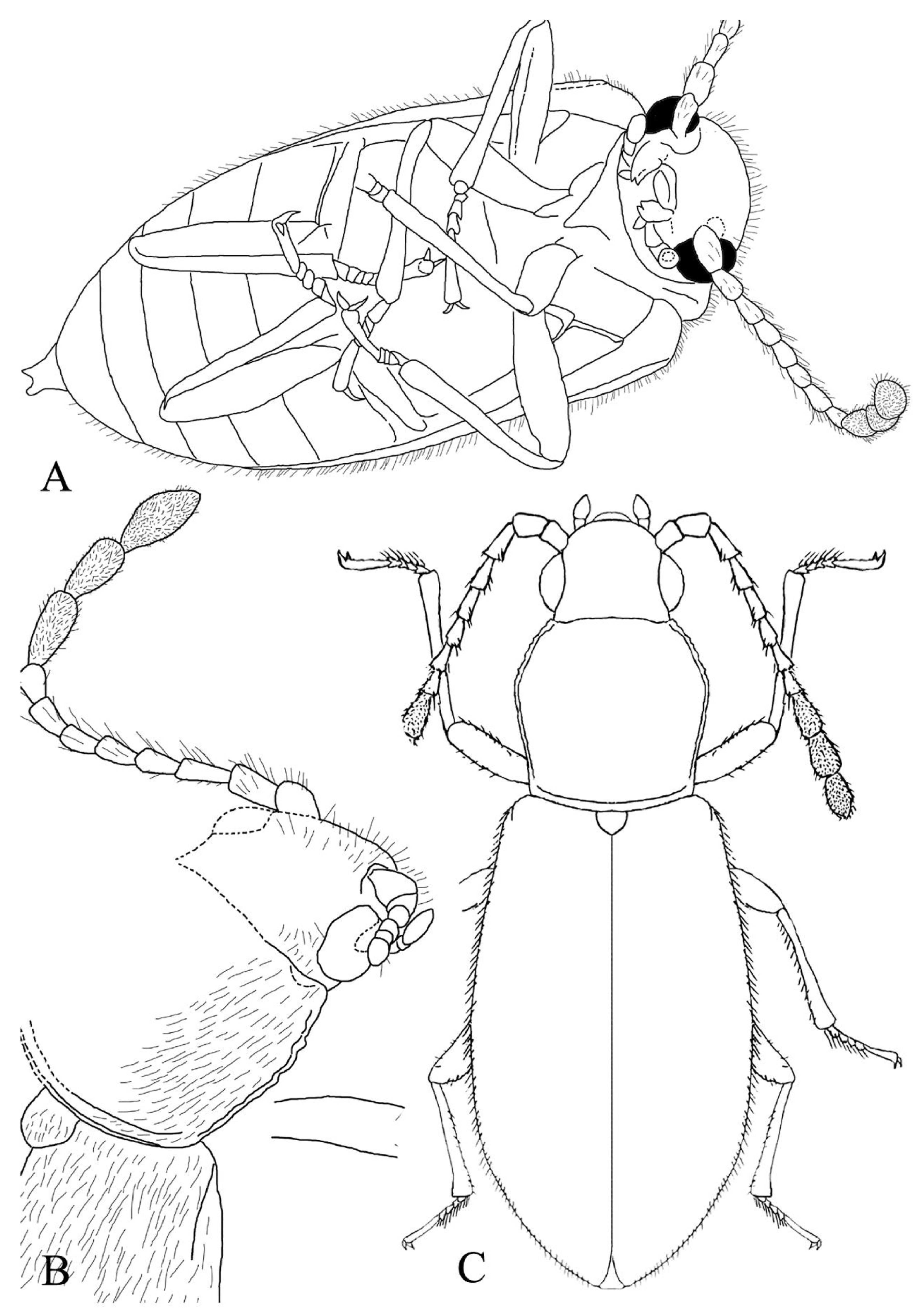
Figure 3.
Schematic drawing of Baltamauroania mirabilicornis gen. et sp. nov., holotype, No AWI-144 [CVIA]: (A) Habitus, dorso-lateral view; (B) Habitus, ventral view; (C) Right antenna; (D) Fore leg; (E) Middle leg; (F) Hind leg. Scale bars represent 1 mm.
Figure 3.
Schematic drawing of Baltamauroania mirabilicornis gen. et sp. nov., holotype, No AWI-144 [CVIA]: (A) Habitus, dorso-lateral view; (B) Habitus, ventral view; (C) Right antenna; (D) Fore leg; (E) Middle leg; (F) Hind leg. Scale bars represent 1 mm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



