
Submitted:
19 September 2023
Posted:
20 September 2023
You are already at the latest version
Abstract
Information on how bacteria in plants and soil, along with extracellular enzymes, affect nutrient cycling in Encephalartos villosus growing in nutrient-poor and acidic scarp forests is lacking. Bacteria in coralloid roots, rhizosphere, and non-rhizosphere soils were isolated to determine the potential role of soil bacterial communities and their associated enzyme activities in nutrient contributions in rhizosphere and non-rhizosphere soils. The role of soil characteristics and associated bacteria on E. villosus nutrition and nitrogen source reliance was investigated. Encephalartos villosus leaves, coralloid roots, rhizosphere, and non-rhizosphere soils were collected at two scarp forests. Leaf nutrition, nitrogen source reliance, soil nutrition and extracellular enzyme activities were assayed. A phylogenetic approach was used to determine the evolutionary relationship between identified bacterial nucleotide sequences. Twenty, twelve and seven different bacterial genera were isolated from rhizosphere, non-rhizosphere, and coralloid roots, respectively. Phosphorus and nitrogen cycling enzyme activities in E. villosus rhizosphere and non-rhizosphere soils were insignificant. More than 70% of the leaf nitrogen was derived from the atmosphere. This study revealed that plant-associated bacteria with plant growth-promoting functions, soil bacteria, and associated extracellular enzymes play a role in E. villosus nutrition and nitrogen source reliance and contribute to E. villosus rhizosphere and non-rhizosphere soil nutrition.
Keywords:
Encephalartos villosus
; cycad-microbe symbiosis
; nutrient deficient soils
; biological nitrogen fixation
1. Introduction
Cycads are ancient gymnosperms that have persisted through many extinction events, survived competition with fast-growing angiosperms and environmental stresses such as drought and nutrient deficiencies [1,2,3]. The Zamiaceae is the most diverse cycad family in South Africa [4]. The Encephalartos genus belongs to the Zamiaceae family and has 37 species that are indigenous to South Africa, making South Africa an important centre for cycad diversity [5,6,7]. Encephalartos villosus is a forest understory cycad amongst the most diverse and widely distributed species in the Encephalartos genus [8,9]. According to [10], cycads are distributed in grasslands and scarp forests that are characterised as nutrient deficient [11]. The growth and development of cycads in these ecosystems may be attributed to their ability to form symbiotic associations with plant growth promoting bacteria [10] and their role in enhancing soil nutrients [12]. According to [13], cycads offer essential ecosystem services such as nutrient cycling including carbon sequestration, soil conservation, wildlife habitat provision, and medicinal resource utilization. Cycads absorb carbon dioxide from the atmosphere through photosynthesis, helping to mitigate climate change by reducing atmospheric greenhouse gas levels [14]. Their extensive root systems stabilize soil and prevent erosion, improving water quality and reducing the risk of landslides [15]. Additionally, cycads serve as a food source for numerous animal species, supporting local biodiversity and contributing to the overall health of ecosystems [16]. Finally, some cycad species have traditional medicinal uses, highlighting their potential as a renewable resource for human wellbeing [17].
[12] reported higher nitrogen (N), carbon (C), and phosphorus (P) concentrations in Cycas micronesica and Zamia integrifolia rhizosphere soils compared to surrounding soil, highlighting the soil nutrient contributions by cycads. [13] reported that soil bacterial communities and their associated enzyme activities contribute to soil nutrient inputs, enabling Encephalartos natalensis to thrive in nutrient deficient and acidic ecosystems. Thus, we can deduce that the role of cycads on soil nutrient inputs may be due to cycad-microbe symbiosis, soil bacterial communities, and associated enzyme activities [13]. The bioavailability of soil nutrients depends on soil bacteria that release extracellular enzymes to mineralise and cycle nutrients, making them available for plant assimilation [18,19,20]. Soil bacteria exude enzymes, such as phosphatase, N-acetylglucosaminidase, and β-glucosidase, that play a role in the mineralisation and cycling of P, N and C [21,22]. While studies such as [12] report that cycads improve soil N, C, and P content, but the role of soil bacterial communities and their associated enzyme activities is not accounted for.
According to [13], soil bacterial communities and their associated enzymes enhance nutrient bioavailability in rhizosphere and non-rhizosphere soils. Therefore, soil bacterial communities and their associated enzymes may influence the growth of E. villosus growing in nutrient deficient and acidic ecosystems. The growth of cycad species in nutrient deficient ecosystems is believed to be due to cycad-microbe symbiosis. Cycads are reportedly associated with plant growth-promoting (PGP) bacteria belonging to the Rhizobium, Lysinibacillus, Bradyrhizobium, and Burkholderia genera [13,23,24,25]. These microbes assist cycad growth by providing essential elements such as nitrogen (N) and phosphorus (P) [26], enabling cycads species to grow in nutrient deficient ecosystems such as scarp forests. [27] reported that symbiotic associations involving cycads and PGP bacteria produce insignificant variations in the N and P concentrations of cycad leaves, highlighting the role that cycad-microbe symbiosis plays on plant nutrition. Though a comprehensive review of the elemental composition of cycad leaves has been compiled by [28], the role of PGP bacteria on E. villosus nutrition is yet to be studied. Also, many studies reported on the N-fixing bacteria associated with cycad species [25,29,30] however, the influence of the bacterial isolates on plant nutrition and N source reliance is poorly understood. Moreover, studies that report on the composition of the 15N isotope in cycad foliage, such as [31], do not report on the N reliance and the percentage of N derived from the atmosphere in cycads.
Hence, this study isolated and identified bacteria in E. villosus coralloid roots, rhizosphere, and non-rhizosphere soils and determined the role of soil characteristics and associated bacteria on E. villosus nutrition and N source reliance. The study also investigated the potential role of soil bacterial communities and their associated enzyme activities in nutrient contributions in E. villosus rhizosphere and non-rhizosphere soils. The objectives of this study included 1) determining the soil characteristics (pH, exchange acidity, total cation, and nutrient concentrations) in E. villosus rhizosphere and non-rhizosphere soils, 2) Identifying the N-fixing, P-solubilising, and N-cycling bacteria found in E. villosus coralloid roots, rhizosphere and non-rhizosphere soils using selective media, 3) assaying the soil nutrient (P and N) cycling enzyme activities in E. villosus rhizosphere and non-rhizosphere soils, and 4) using 15N isotope to determine the N reliance of E. villosus growing in nutrient deficient and acidic scarp forest ecosystem soils. We hypothesized that the bacterial composition in coralloid roots, rhizosphere, and non-rhizosphere soils of E. villosus and associated extracellular enzyme activities would indicate the species' contribution to the bioavailability of soil nutrition and contribute to plant growth, nutrition, and reliance on N derived from the atmosphere. Conducting a study on the role of cycad-associated microbes on E. villosus growth and nutrition is essential because E. villosus has a life history similar to cycads that cannot be studied due to their conservation status [6]. Therefore, studying the role of microbes on E. villosus growth will not only shed light on how microbial interactions in E. villosus contribute to soil nutrient inputs, growth, and plant nutrition but will also give insights into other cycad species with similar life history.
2. Materials and Methods
2.1. Study sites and species
Sixteen randomly selected adult E. villosus plants of the same age, growing in two scarp forests were sampled. The scarp forests are Oceanview Farm in East London and Rhebu village in Port St Johns (eight plants were sampled from each locality), Eastern Cape, South Africa (Coordinates not included due to cycads conservation concerns). These two scarp forests, like others, occur in nutrient-poor, leached, and shallow soils [11]. The forest floor is covered with decaying leaflitter, with E. villosus as the dominant understory plant and tall trees such as Ptaeroxylon obliquum(F.Muell.) Benth. & Hook.f. ex Harv., Hyperacanthus amoenus L., Protorhus longifolia (Lam.) Small., and Vepris undulata (Aiton) Radlk., adorning the forests [11]. In addition to muthi (traditional healer) harvesting and collection of biofuels, the scarp forests in Oceanview and Rhebu village are also disturbed by cattle that graze within the forests. In the process, the cattle deposit dung and urine, enriching the forest soils. According to [32], the chemical composition of rhizosphere soils in the root and litterfall zone may be altered by cycads. Thus, rhizosphere soils were collected directly beneath the selected plants and along the dripline of the leaf canopy. The soils were collected at 0-20 cm depth at the four cardinal points of each plant (North, West, South, and East). Similarly, non-rhizosphere soils (similar depths) were collected from non-target sites defined by a radius of five meters from the base of each target plant. The direction of the non-rhizosphere samples was randomly selected using cardinal points of North, East, South, and West of the target plant. From each E. villosus target plant, soil samples (rhizosphere and non-rhizosphere) collected at different depths and cardinal points were bulked to form composite samples for soil characteristic analysis (nutrient concentrations, total cation, exchange acidity, and pH).
2.2. Soil characteristics
[33] reported that soil nutrient concentrations directly affect cycad leaf concentrations. Therefore, thirty-two 50 g compound soil samples collected from each sampled E. villosus plant were sent to the KwaZulu Natal Department of Agriculture and Rural Development Analytical services for soil nutrient concentrations, total cation concentrations, exchange acidity, and pH analysis. The soil characteristic analysis was performed per protocols explained by [34]. The total soil N concentration was measured using the Automated Dumas dry combustion method with a LECO CNS 2000 (Leco Corporation, USA). The atomic absorption method was used to measure extractable soil P and potassium (K) concentrations, where 2.5 ml soil solution was mixed with 25 ml ambic-2 solution at a pH of 8. Thereafter, the mixture was stirred at 400 rpm for 10 minutes using a multiple stirrer and filtered with Whatman no.1 paper. The same methods as above were used to determine the soil calcium and magnesium concentration, however 25 ml 1 M KCl solution was used instead of ambic-2 solution. Soil pH was determined by mixing 10 ml soil solution with 25 ml 1 M KCl solution and stirring at 400 rpm for 5 minutes using a multiple stirrer. The pH of the suspension was measured using a gel-filled combination glass electrode while stirring.
2.3. Coralloid root surface sterilisation
Coralloid roots were harvested from eight randomly selected mature E. villosus individuals in Rhebu and Oceanview, respectively. The coralloid roots were surface sterilized with 70% (v/v) ethanol for 30 seconds and soaked in 3.5% (v/v) sodium hypochlorite solution for 3 minutes. Thereafter, they were rinsed ten times with sterile distilled water and immersed in 100μl of 15% glycerol in sterile Eppendorf tubes.
Bacterial extraction and identification from coralloid roots and soils
Soil samples collected from the rhizosphere and non-rhizosphere soils in Rhebu and Oceanview were subjected to serial dilutions, and the immersed coralloid roots were crushed using sterile pipette tips for bacterial extraction [35]. Ten µL of the root suspension and 50µL of each serial dilution were cultured in sterile Petri plates containing selective media (Pikovskaya's plate containing tricalcium phosphate (TCP) for P-solubilizing bacteria, Simmons citrate agar for N-cycling bacteria, and Jensen's media agar for N-fixing bacteria) to ensure that only bacteria with P-solubilizing, N-cycling, and N-fixing traits grew on the plates [36]. Each selective media was replicated three times and incubated at 30˚C for five days. Pure bacterial colonies were obtained by repeated streaking/subculturing. A small portion of the pure bacterial colonies was amplified through polymerase chain reaction (PCR) using the 16S ribosomal RNA gene primers: 63F (5' CAGGCCTAACACATGCAAGTC 3' (21 bases) and 1387R (5' GGGCGGTGTGTACAAGGC 3' (18 bases)) [37] from Inqaba Biotechnical Industries (Pty) Ltd (South Africa). The PCR amplification was performed using an EmaraldAmp GT Master Mix with the following conditions: Initial denaturation at 94˚C for 5 minutes, followed by 30 cycles of denaturation at 94˚C for 30 seconds, annealing at 55˚C for 30 seconds and extension at 72˚C for 2 minutes, with additional extension at 72˚C for 10 minutes. The PCR products were separated by electrophoresis on 1% (w/v) agarose gel and visualized under UV light to determine the correct product size amplification. The amplicons were sent for sequencing at Inqaba Biotechnical Industries (Pty) Ltd, South Africa. The DNA sequences were edited and compared to the nucleotide sequences of known bacteria in the GenBank database of the National Centre for Biotechnology Information (NCBI) by using Basic Local Aligned Search Tool (BLAST) (https://www.ncbi.nlm.nih.gov).
Using the sequences of the identified bacteria, a phylogenetic approach was used to determine the evolutionary relationship between bacterial nucleotide sequences. The nucleotide alignment was done using the MUSCLE tool in MEGA 11 and checked manually before constructing the phylogenetic tree using the neighbour-joining likelihood tree approach. Also, a bootstrap resampling was performed with 1000 replicates in accordance with the procedure by [38].
2.4. Extracellular enzyme activities
N-acetylglucosaminidase, acid phosphatase and alkaline phosphatase activities were assayed using the fluorescence-based method described by [39] and expressed as nmol h-1g-1. Briefly, samples (10g soil/100 ml autoclaved dH2O) were homogenised at medium speed in a shaker for two hours and stored overnight at 4˚C. The supernatants were transferred into black 96-well microplates before adding their respective substrates. The sample run consisted of 200μl soil aliquot and 50μl substrate, alongside reference standards (200μl bicarbonate buffer + 50μl standard), quench standard (200μl soil aliquot + 50μl standard), sample control (200μl soil aliquot + 50μl buffer), negative controls (200μl buffer + 50μl substrate), and blanks (250μl buffer). The 96-well plate was incubated at 25˚C for 2 hours. The reaction was stopped afterward by adding 5μl of 0.5M NaOH to each well. The fluorescence was measured at 450 nm on a Glomax Multi Plus microplate reader. The buffer and standard were adjusted to pH five before determining acid phosphatase activity. Nitrate reductase activity assays were done using a modified protocol described by [40]. A volumetric flask wrapped in foil was filled with 1 ml of 25 mM KNO3, 4 ml of 0.9 mM 2,4-dinitrophenol, and 5 ml of milliQ dH2O. Thereafter, 5g of soil was added to the solution, and the flask was sealed with foil, shaken, and incubated in an oven at 30˚C for 24 hours. After incubation, 10ml of 4M KCl was added to the soil mixture, succinctly mixed, and filtered using filter paper (Whatman number 1). The enzymatic reaction was initiated by adding 2ml of the filtrate to 1.2ml of 0.19M ammonium chloride buffer (pH 8.5) and 0.8ml of a colour reagent consisting of 1% sulfanilamide, 1N HCl, and 0.02% N-(1-naphthyl) ethylenediamine dihydrochloride (NEDD). The solution was incubated at 30˚C for 30 minutes. The absorbance was measured at 520nm using an 1800 UV spectrophotometer. The nitrite (NO2-) liberated into the medium was extrapolated from a prepared standard curve with KNO3. The nitrate reductase activity was expressed as 0.1 μmol h-1 g-1.
2.5. Leaf nutrient composition
[41] reported that leaf nutrient compositions differ with age. Therefore, young leaves were sampled. Leaf N and P concentrations were considered to determine the role of N-fixing, N-cycling, and P solubilising bacteria on leaf nutrition. The number of leaves and leaf length (cm) from selected plants were measured. Leaves from E. villosus plants of the same size in Rhebu and Oceanview were collected on the leaf's basal, midpoint, and apical locations per [27]. The sampled leaves were oven dried at 80˚C till constant weight, ground into a fine powder with a tissuelyser, and sent to the Central Analytic Facilities at the University of Stellenbosch (South Africa) for P and N analysis through Inductively Coupled Mass Spectrometry (ICP-MS) and N isotope analysis at the Archeometry Department at the University of Cape Town (South Africa).
2.6. Percentage N Derived from the atmosphere (%NDFA)
The University of Cape Town Archeometry Department conducted the N isotope analysis. The isotopic ratio of N was calculated as δ = 1000 (Rsample/Rstandard), where R is the molar ratio of the heavier to the lighter isotope of the samples and standards. Between 2.10 and 2.20 mg of each milled sample were weighed into 8 mm × 5 mm tin capsules (Elemental Micro-analysis, Devon, UK) on a Sartorius microbalance (Goettingen, Germany). The samples were then combusted in a Fisons NA 1500 (Series 2) CHN analyser (Fisons Instruments SpA, Milan, Italy). The nitrogen isotope values for the N gas released were determined on a Finnigan Matt 252 mass spectrometer (Finnigan MAT GmbH, Bremen, Germany), which was connected to a CHN analyser by a Finnigan MAT Conflo control unit. Five standards were used to correct the samples for machine drift, namely, two in-house standards (Merck Gel and Nasturtium) and the IAEA (International Atomic Energy Agency) standard (NH4)2SO4.
%NDFA = 100 ((δ15N reference plant – δ15N cycad)/ (δ15 N reference plant − β)) [42]
Where NDFA is the N derived from the atmosphere, the B value represents the δ15N natural abundance of the N derived from biological N2 fixation.
2.7. Statistical analysis
R studio version 3.6.2 was used to test for differences in the micronutrients, pH, exchange acidity, soil enzyme activity, and total cation in soil samples collected in E. villosus rhizosphere and non-rhizosphere soils using independent samples T-test. The assumptions for normality and homogeneity of the variances were tested using the one-sample Kolmogorov-Smirnov normality and Levene's test, respectively (p>0.05). The Wilcoxon test, a non-parametric alternative, was used in cases where the assumptions were not met. A simple linear correlation analysis was done to determine if there is a relationship between soil enzyme activities and soil nutrition. A probability of p≤0.05 was considered significant. Relations between rhizosphere soil characteristics, extracellular enzyme activities, leaf nutrients, and N source reliance were determined using principal component analysis (PCA) on R studio version 3.6.2 using statistical package gg plot, function pr comp.
3. Results
3.1. Soil characteristics
The Magnesium (Mg) and zinc (Zn) concentrations in rhizosphere soils were significantly higher than in the non-rhizosphere soils in Oceanview (Table 1). There were no significant differences in the macronutrient (nitrogen (N), phosphorus (P), and potassium (K)) concentrations of rhizosphere and non-rhizosphere soils in both localities (Table 1). The soil samples collected from rhizosphere and non-rhizosphere soils in Rhebu and Oceanview were relatively acidic (Table 1). In Rhebu and Oceanview, rhizosphere soils had a higher pH of 4.28 and 5.43 compared to pH of 4.14 and 4.97 for the non-rhizosphere soils, respectively (Table 1).
| Study site | Oceanview | Rhebu | ||
| Parameter | Rhizosphere | Non-rhizosphere soils | Rhizosphere | Non-rhizosphere soils |
| P K |
6.1±2.9a 139.5±33.6a |
3.86±1.9a 81.9±12.5 a |
22.95±5.9 a 330.0±100.9 a |
13.23±3.7 a 179.1±54.9 a |
| N | 3093.7±741.9 a | 2638.0±438.2 a | 5189.3±297.9 a | 3473.3±1396.3 a |
| Ca | 1555.3±274.8 a | 1379.2±153.5 a | 1534.9±974.2 a | 1229.4±1087.2 a |
| Mg | 454.7±46.1a | 382.12±35.3b | 838.5±549.3 a | 697.1±548.2 a |
| Mn | 55.3±21.1 a | 44.8±15.9 a | 89.7±58.0 a | 48.1±20.2 a |
| Zn | 2.2±0.1c | 1.3±0.4d | 4.1±1.6 a | 2.8±1.2 a |
| Cu | 1.1±0.2 a | 0.6±0.1 a | 0.9±0.1 a | 0.6±0.2 a |
| Exchange acidity (cmol/L) Total cations (cmol/L) pH |
0.05±0.01 a 11.53±1.65 a 5.43±0.41e |
0.05±0.01 a 11.65±1.62 a 4.97±0.57f |
1.46±1.42 a 15.26±8.13 a 4.28±0.61 a |
1.37±1.19 a 15.10±9.48 a 4.14±0.58 a |
| In each row, different letters denote significant differences in the nutrient concentrations and relative acidity of soil samples within study sites (independent samples t-test, p≤0.05, means±SE, n=32). | ||||
3.2. Bacterial identification
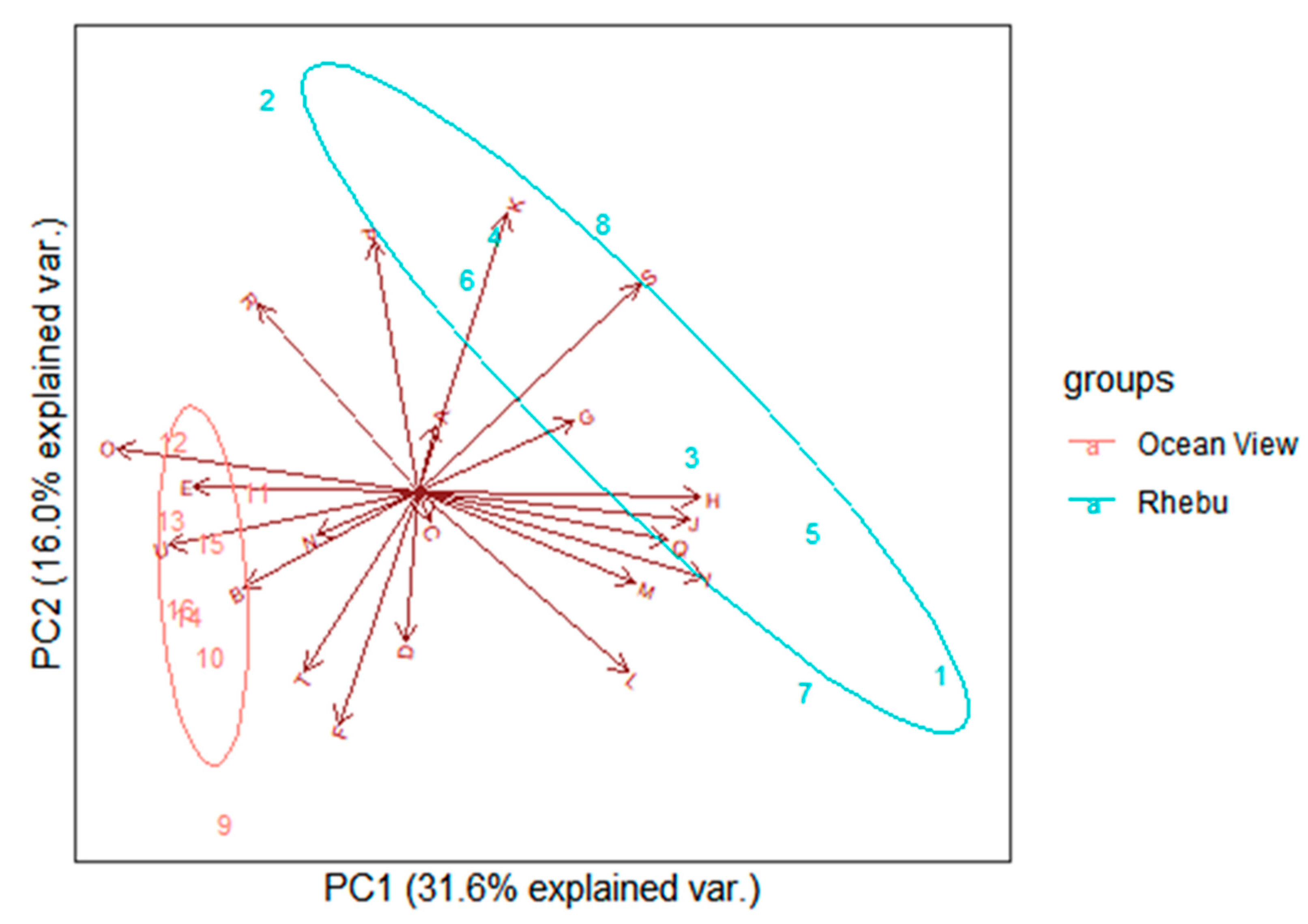
Three phylogenetic trees were generated based on bacterial isolates collected from Oceanview (Figure 1), Rhebu (Figure 2), and a combination of both sites (Figure 3). Bacteria from rhizosphere soils from both Oceanview and Rhebu were in 20 different genera, whereas those isolated from non-rhizosphere soils were in 12 genera and only 7 genera were identified from coralloid roots (Figure 1, Figure 2 and Figure 3). Notably, bacterial clusters were found to be associated with specific sampling locations, including rhizosphere soil, non-rhizosphere soil, and coralloid roots. While a few exceptions were observed, particularly in the case of Oceanview sample (Figure 1), the clustering pattern was consistently evident across all three datasets, indicating that the origin of the bacterial isolates significantly influenced their phylogenetic profiles.
3.3. Extracellular enzyme activities
There were insignificant differences in the N-cycling (nitrate reductase and N-acetylglucosaminidase) and P-cycling (acid and alkaline phosphatase) enzyme activities of rhizosphere and non-rhizosphere soils in Rhebu and Oceanview (Figure 4). Though the differences were insignificant, Rhebu and Oceanview rhizosphere soils had a higher alkaline phosphatase activity than non-rhizosphere soils (Figure 4). In Rhebu, the non-rhizosphere soils had a higher acid phosphatase activity than the rhizosphere soils (Figure 4).
3.4. Leaf nutrition and N source reliance
Encephalartos villosus leaf nutrition, number of leaves, leaf length, and N source reliance is represented in Figure 5. The mean leaf length of E. villosus plants in Rhebu and Oceanview was 186.5±49.6 cm (n = 8) and 223.6±28.5 cm, (n = 8) respectively. The sampled E. villosus plants from Rhebu and Oceanview had 11.4±5.3 (n = 8) and 10.0±4.2 (n = 8) leaves, respectively. The Leaf N concentration of E. villosus plants growing in Rhebu and Oceanview was 2.29 and 2.13 mmol g-1 respectively. The P concentration of E. villosus leaves from Rhebu and Oceanview was 8.8 and 10.9 umol g-1, respectively. The N concentration derived from the atmosphere by Rhebu plants was 0.60 and 0.70 mmol N g-1 dw for Oceanview plants. Only 0.25 and 0.21 mmol N g-1 dw was derived from the soil in Rhebu and Oceanview, respectively. Encephalartos villosus plants growing in Rhebu and Oceanview utilised 70.31% and 77.00% N derived from the atmosphere, respectively.
3.5. Correlations between rhizosphere soil characteristics, extracellular enzyme activities, leaf nutrition, and nitrogen source reliance
Correlations between rhizosphere soil characteristics, extracellular enzyme activities, leaf nutrition, and N source reliance are represented in Figure 6. The first PCA explained 31.6% of the total variation, and the second PCA explained 16.0% of the total variation. The clusters indicated that the growth of E. villosus in Rhebu and Oceanview varied. Variations in E. villosus plants growing in Rhebu were due to soil pH (represented as H) and soil P (represented as J). The PCA plot also showed relationships between soil characteristics, extracellular enzyme activities, leaf nutrition, and N source reliance. Groups closer to each other represented a high correlation. The concentration of N derived from the atmosphere (represented as A) was strongly correlated with soil N (represented as K). Percentage NDFA (represented as N) was correlated with plant N (represented as B), soil pH (represented as H) was correlated with soil P (represented as J), and soil P (represented as J) was correlated with acid phosphatase activity (represented as Q).
4. Discussion
In the present study, E. villosus coralloid roots, rhizosphere and non-rhizosphere soils were sampled to determine the role of cycad-microbe symbiosis, soil bacterial communities, and extracellular enzymes on the nutrition and growth of E. villosus in acidic and nutrient deficient scarp forest ecosystems. The first noteworthy observation in this study is the clustering pattern of the isolated bacterial strains. Initially, one might hypothesize that bacterial clusters would align with their functional attributes. However, our findings demonstrate that these clusters are primarily dictated by the ecological niches from which the bacteria originated - specifically, rhizosphere soil, non-rhizosphere soil, and coralloid roots. We propose that the underlying reasons for this pattern are related to the concept of the filter theory [43,44] and its potential interaction with the hologenome theory [45]. The filter theory suggests that environmental filters, such as soil properties and abiotic factors, play a pivotal role in shaping microbial communities. Such an effect of environment on the design of the symbionts has been previously demonstrated in several legumes [46,47] and what is more, in soils where E. villosus grows [48].
In this study, it is worth highlighting that the soil properties and environmental conditions in the scarp forests, Oceanview and Rhebu, are very similar. These shared environmental traits would typically suggest that microbial communities in both locations should exhibit a higher degree of functional similarity due to comparable selection pressures. However, our findings suggest something different, implying that factors beyond these abiotic filters are at play. The hologenome theory [49,50] hypothesizes that the collective genetic material of a host organism and its associated symbionts, including bacteria, should be considered as a single functional unit or holobiont. It suggests that the host's genotype, as well as its associated symbiotic microbial community, contribute to its ecological fitness and adaptation to specific niches. Therefore, variations in the holobiont's composition can drive the observed clustering patterns, even in environments with similar abiotic conditions. In the context of our study, it is likely that in a first step, the environment could have driven the kind of bacteria associated to E. villosus, as there are similarities in the bacterial strains identified in both Oceanview and Rhebu. Secondly, we suggest that host plants, in this case, the coralloid roots, play a pivotal role in shaping the microbial communities residing within their immediate vicinity.
The genetic makeup and specific interactions of these host plants with bacteria may exert a stronger influence on the bacterial community structure than the shared abiotic factors across the two study sites. According to [48], the availability of nutrients in the soil may shape symbiotic interactions between plants and microorganisms, thus the abundance of P solubilising bacteria in E. villosus coralloid roots, rhizosphere and non-rhizosphere soils in both localities may have been driven by P deficiency. Phosphorus solubilising bacteria belonging to the Pseudomonas, Bacillus, and Enterobacter genera have been reported to play a significant role in P solubilisation [51,52]. Phosphorus solubilising bacteria stimulate plant growth by producing phosphatases that mineralise and cycle P [53,54]. [55] studied the secretion of alkaline phosphatases and reported that Bacillus licheniformis secreted 30-40% of alkaline phosphatase in a medium containing low phosphate concentrations. Also, Variovorax paradoxus, a species isolated from Oceanview rhizosphere soils and Rhebu non-rhizosphere soils (Figure 1 and 2), reportedly produced 94.0 U/mL alkaline phosphatase in a low P liquid medium [56], elucidating that P solubilising bacteria secrete phosphatases in P deficiency.
According to the resource allocation model for extracellular enzyme activities, soil microbes exude extracellular enzymes to mineralise and cycle deficient soil nutrients [46]. Thus, the correlation between soil P and acid phosphatase activity (Figure 6) may have been caused by P deficiency in the two scarp forest soils. The two scarp forests, Rhebu and Oceanview, sampled in this study were acidic (Table 1). In acidic soils, P forms insoluble complexes with aluminium (Al) and iron (Fe), making P unavailable for plant uptake [57]. [13] reported positive correlations between soil extractable P, alkaline phosphatase, and acid phosphatase activity in the rhizosphere and non-rhizosphere soils of Encephalartos natalensis growing in P-deficient and acidic savanna woodland ecosystem soils, thus showing the link between soil nutrient deficiencies and extracellular enzyme activities. The P cycled by P cycling enzymes and the presence of N-fixing non-cyanobacterial species in E. villosus coralloid roots indicate P and N deficiency, which may trigger BNF for N nutrition [13]. Thus, the reliance of E. villosus on N derived from the atmosphere may have been attributed to N deficiency and P provision through P-solubilising bacteria, extracellular enzyme activities, and possibly arbuscular mycorrhizal fungi. In the principal component analysis, leaf N concentration was correlated with % N derived from the atmosphere, which may be attributed to the symbiotic association between cycads and N-fixing microbes. Nitrogen-fixing microbes increase the bioavailability of N for plant uptake, and the presence of N-cycling bacteria in the coralloid roots and rhizosphere soils mobilise the N [13], which may have been translocated to the leaves.
5. Conclusions
Our findings challenge the conventional expectation that environmental filters alone dictate microbial community composition in similar habitats. Instead, they suggest that host-microbe interactions, as proposed by the hologenome theory, are significant drivers of microbial community structuring. Further investigations are needed to delve deeper into the intricate interplay between environmental filters, host genetics, and microbial symbionts to fully comprehend the underlying mechanisms governing microbial community assembly in complex ecosystems like rhizosphere environments. This study shows how the wide distribution of E. villosus in nutrient deficient and acidic ecosystems may be linked to the cycad's association with bacteria with plant growth-promoting traits. Furthermore, this study revealed that E. villosus rhizosphere and non-rhizosphere soil bacterial communities and their associated enzyme activities contribute to soil nutrition in nutrient deficient and acidic scarp forests. To our knowledge, this study is the first to determine the N source reliance in cycads. These findings open a new avenue of study focused on cycad-microbe symbiosis, which will provide a result-based analysis of N-fixation in cycads. If the contributions of non-cyanobacterial cycad symbionts in N-fixation efficiency are determined, it would indicate that the N source reliance of cycad species is not only limited to cyanobacteria but also non-cyanobacterial species.
Author Contributions
All authors contributed equally to the experimental planning, field trips, analysis and manuscript writing, and all authors agreed to submit the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The research leading to these results received funding from the National Research Foundation under the grant agreement number (Grant UID 129403 and 138091).
Data Availability Statement
The data and material can be made available upon request.
Acknowledgments
We appreciate the financial support from the National Research Foundation (Grant UID 129403 and 138091). We acknowledge the support of the University of KwaZulu-Natal (School of Life Sciences) and University of Mpumalanga (School of Biology and Environmental Sciences).
Conflicts of Interest
We declare no known competing financial and non-financial interests with regards to the current research. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Brenner, E.D.; Stevenson, D.W.; Twigg, R.W. Cycads: Evolutionary innovations and the role of plant-derived neurotoxins. Trends Plant Sci. 2003, 8, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Marler, T.E.; Krishnapillai, M.V. Cycas micronesica trees alter local soil traits. Forests. 2018, 9, 565. [Google Scholar] [CrossRef]
- Erdei, B.; Coiro, M.; Miller, I.; Johnson, K.R.; Griffith, P.M.; Murphy, V. First cycad seedling foliage from the fossil record and inferences for the Cenzoic evolution cycads. Biol. Lett. 2019, 15, 1–6. [Google Scholar] [CrossRef]
- Cousins, S.R.; Williamson, V.L.; Witkowski, E.T.F. Sifting through cycads: A guide to identifying the stem fragments of six South African medicinal Encephalartos species. S. Afr. J. Bot. 2013, 84, 115–123. [Google Scholar] [CrossRef]
- Donaldson, J.S. Cycads status survey and conservation action plan; IUCN/SSC Cycad Specialist Group. IUCN Gland, Switzerland and Cambridge: United Kingdom, 2003; pp. 26–89. [Google Scholar]
- Raimondo, D.; von Staden, L.V.; Foden, W.; Victor, J.E.; Helmen, N.A.; Turner, R.C.; Kamundi, D.A.; Manyama, P.A. Red list of South African plants, Strelitzer 25; South African National Biodiversity Institute: Pretoria, 2009. [Google Scholar]
- Condamine, F.L.; Nagalingum, N.S.; Marshall, C.R.; Morlon, H. 2015. Origin and diversification of living cycads: A cautionary tale on the impact of the branching process prior in Bayesian molecular dating. BMC Evol. Biol. 2015, 15, 1–5. [Google Scholar] [CrossRef]
- Cousins, S.R.; Williamson, V.L.; Witkowski, E.T.F. Uncovering the cycad taxa (Encephalartos species) traded for traditional medicine in Johannesburg and Durban, South Africa. S. Afr. J. Bot. 2012, 78, 129–138. [Google Scholar] [CrossRef]
- Yessoufou, K.; Bamigboye, S.O.; Daru, B.H.; van der Bank, M. Evidence of constant diversification punctuated by a mass extinction in the African cycads. Ecol. Evol. 2014, 4, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Yepiz, J.C.; Cueva, A.; Dovciak, M.; Teece, M.; Enrico, A.; Yepez, E.A. 2014. Ontogenic resource-use strategies in a rare long-lived cycad along environmental gradients. Conserv. Physiol. 2014, 2, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Mucina, L.; Scott-Shaw, C.R.; Rutherford, M.C.; Camp, K.G.T.; Matthews, W.S.; Powrie, L.W.; Hoare, D.B. Indian Ocean coastal belt. Mucina, L., Rutherford, M.C., Eds.; In The vegetation of South Africa, Swaziland and Lesotho. Strelitzia 19; Pretoria: South African National Biodiversity Institute, 2006. [Google Scholar]
- Marler, T.E.; Calonje, M. Two cycad species affect the Carbon, Nitrogen, and Phosphorus content of soils. Horticulturae. 2020, 6, 24. [Google Scholar] [CrossRef]
- Ndlovu, S.; Suinyuy, T.N.; Pérez-Fernández, M.A.; Magadlela, A. Encephalartos natalensis, Their Nutrient-Cycling Microbes and Enzymes: A Story of Successful Trade-Offs. Plants. 2023, 12, 1034. [Google Scholar] [CrossRef]
- Smith, J.; Johnson, M.; Williams, D. The impact of cycads on the environment. J. Environ. Stud. 2018, 27, 123–135. [Google Scholar]
- Smith, J.; Johnson, M.; Williams, D. The impact of cycads on arid environments. J. Arid Environ. 2017, 105, 123–135. [Google Scholar]
- García, M.; Hernandez, R.; Lopez, P. The role of cycads in forest restoration efforts. Forestry Chronicle. 2019, 95, 12–23. [Google Scholar]
- Brown, T.; Davis, R.; Rodriguez, L. The economic value of cycads in developing countries. Economic Botany. 2018, 72, 34–45. [Google Scholar]
- Cenciani, K.; dos Santos Freitas, S.; Critter, S.A.M.; Airoldi, C. Microbial enzymatic activity and thermal effect in tropical soil treated with organic material. Soil Sci. Plant Nutr. 2008, 65, 1–7. [Google Scholar] [CrossRef]
- Latha, P.; Anand, T.; Prakasam, V.; Jonathan, E.I.; Paramathma, M.; Samiyappan, R. Combining Pseudomonas, Bacillus and Trichoderma stains with organic amendments and micronutrient to enhance suppression of collar and root rot disease in physic nut. Appl. Soil Ecol. 2011, 49, 215–223. [Google Scholar] [CrossRef]
- Meena, A.; Rao, K.S. Assessment of soil microbial and enzyme activity in the rhizosphere zone under different land use/cover of a semi-arid region, India Ecol. Process. 2021, 10, 1–10. [Google Scholar]
- Merino, C.; Godoy, R.; Matus, F. Soil enzymes and biological activity at different levels of organic matter stability. J Soil Sci. Plant Nutr. 2016, 16, 14–30. [Google Scholar]
- Adetunji, A.T.; Lewu, F.B.; Mulidzi, R.; Ncube, B. The biological activities of β-glucosidase, phosphatase, and urease as soil quality indicators: A review. J. Soil Sci. Plant Nutr. 2017, 17, 794–807. [Google Scholar] [CrossRef]
- Grobbelaar, N.; Hattingh, W.; Marshall, J. The occurrence of coralloid roots on the South African species of the Cycadales and their ability to fix nitrogen symbiotically. S. Afr. J. Bot. 1986, 52, 467–471. [Google Scholar] [CrossRef]
- Barea, J.; Pozo, M.J.; Azcòn-Aguilar, C. Microbial co-operation in the rhizosphere. J. Exp. Bot. 2005, 56, 1761–1778. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-García, K.; Bustos-Díaz, E.D.; Corona-Gómez, J.A.; Ramos-Aboites, H.E.; Sélem-Mojica, N.; Cruz-Morales, P.; Pérez-Farrera, M.A.; Barona-Gómez, F.; Cibrián-Jaramillo, A. Cycad coralloid roots contain bacterial communities including Cyanobacteria and Caulobacter spp. that encode niche-specific biosynthetic gene clusters. Genome Biol. Evol. 2019, 11, 319–334. [Google Scholar] [CrossRef]
- Sasse, J.; Martinoia, E.; Northern, T. Feed your friends: Do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Marler, T.E.; Lindstrӧm, A. Leaf nutrient relations of cycads in common garden. Trop. Conserv. Sci. 2021, 1, 1–10. [Google Scholar] [CrossRef]
- Deloso, B.E.; Krishnapillai, M.V.; Ferreras, U.F.; Lindström, A.J.; Calonje, M.; Marler, T.E. Chemical element concentrations of cycad leaves: Do we know enough? Horticulturae. 2020, 6, 85. [Google Scholar] [CrossRef]
- Gehringer, M.M.; Pengelly, J.J.L.; Cuddy, W.S.; Fieker, C.; Forster, P.I.; Neilan, B.A. Host selection of symbiotic cyanobacteria in 31 species of the Australian cycad genus: Macrozamia (Zamiaceae). Molecular Plant Microbe Interactions. 2010, 23, 811–82. [Google Scholar] [CrossRef]
- Suárez-Moo, P.J.; Vovides, A.P.; Griffith, M.P.; Barona-Gòmez, F.; Cibrián-Jaramillo, A. Unlocking a high bacterial diversity in the coralloid root microbiome from the cycad genus Dioon. PLoS ONE. 2019, 14, 1–20. [Google Scholar] [CrossRef]
- Kipp, M.A.; Stüeken, E.E.; Gehringer, M.M.; Sterelny, K.; Scott, J.K.; Forster, P.I.; Strӧmberg, C.A.E.; Buick, R. Exploring cycad foliage as an archive of the isotopic composition of atmospheric nitrogen. Gebiology. 2019, 18, 152–166. [Google Scholar] [CrossRef] [PubMed]
- Marler, T.E.; Lindstrӧm, A. Inserting cycads into global climate nutrient relations datasets. Plant Signal. Behav. 2018, 13, e15475781–e15475786. [Google Scholar] [CrossRef]
- Marler, T.E.; Ferreras, U.F. Disruption of leaf nutrient remobilization in coastal Cycas trees by tropical cyclone damage. Journal Geography and Natural Disasters. 2015, 5, 142. [Google Scholar]
- Manson, A.D.; Roberts, V.G. Analytical methods used by the soil fertility and analytical services section. KZN Agri-report no. N/A/2001/04, Pietermaritzburg, South Africa. 2000. [Google Scholar]
- Somasegaran, P.; Hoben, H.J. Handbook for Rhizobia: Methods in legume–Rhizobium technology; Springer-Verlag: New York, USA., 1994. [Google Scholar]
- Ndabankulu, N.; Tsvuura, Z.; Magadlela, A. Soil microbe and associated extracellular enzymes largely impact nutrient bioavailability in acidic and nutrient poor grassland ecosystem soil. Scientific Reports. 2022, 12, 12601. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, J.R.; Sato, T.; Weightman, A.J.; Martin, T.A.; Fry, J.C.; Him, S.J.; Dymock, D.; Wade, W.G. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Appl Environ Microbiol. 1998, 64, 795–9. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11, Molecular Evolutionary Genetics Analysis Version 11. Molecular Biology and Evolution. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.R.; Tyler, H.L.; Millar, J.J. Determination of microbial extracellular enzyme activity in waters, soils, and sediments using high throughput microplate assays. JoVE. 2013, 80, e5039. [Google Scholar]
- Kandeler, E. Potential Nitrification. In Schinner F et al. (eds.) Methods in Soil Biology; Spinger-Verlag Berlin Heidelberg: Berlin, 1995; pp. 146–149. [Google Scholar]
- Marler, T.E.; Krishnapillai, M.V. Incident light and leaf age influence leaflet element concentrations of Cycas micronesica trees. Horticulturae. 2019, 5, 58. [Google Scholar] [CrossRef]
- Shearer, G.; Kohl, D.H. N2-fixation in field settings: estimations based on natural 15N abundance. Funct. Plant Biol. 1986, 13, 699–756. [Google Scholar]
- Johnson, S.L.; Holt, R.D. The evolution of protein function. Science. 1989, 246, 1293–1296, Retrieved from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1228813/. [Google Scholar]
- Sparks, D.W.; et al. The evolutionary origins of biodiversity. Science. 2017, 358, 692–697. [Google Scholar]
- Kropinski, M.; et al. The hologenome: a novel concept for understanding the organization and dynamics of prokaryotic genomes. PLOS Computational Biology. 2017, 13, e1005647. [Google Scholar]
- Simonsen, A.K.; Dinnage, R.; Barrett, L.G.; Prober, S.M.; Thrall, P.H. Symbiosis limits establishment of legumes outside their native range at a global scale. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Ulkrike, M. Are legumes different? Origins and consequences of evolving nitrogen fixing symbioses. J. Plant Physiol Sep. 2022, 76, 153765. [Google Scholar] [CrossRef]
- Motsomane, N.; Suinyuy, T.N.; Pérez-Fernández, M.A.; Magadlela, A. How the right evolved partners in Cycads and Legumes drive enhanced growth in a harsh environment. Symbiosis. 2023, in press.
- Jinek, M.; Chylinski, K.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Nature. 2005, 337, 816–82. [Google Scholar] [CrossRef]
- Charpentier, E.; Valgepea, T. The holobiont: A New Paradigm for Life on Earth. Dialogues in Human Geography. 2017, 1–14. [Google Scholar] [CrossRef]
- Whitelaw, M.A. Growth promotion of plants inoculated with phosphate solubilizing fungi. Adv. Agron. 2000, 69, 99–151. [Google Scholar]
- Igual, J.M.; Valverde, A.; Cervantes, E.; Velázquez, E. Phosphate-solubilizing bacteria as inoculants for agriculture: use of updated molecular techniques in their study. Agronomie. 2001, 21, 561–568. [Google Scholar] [CrossRef]
- Jorquera, M.A.; Crowley, D.E.; Marschner, P.; Greiner, R.; Fernández, M.T.; Romero, D.; Menezes-Blackburn, D.; De La Luz Mora, M. Identification of b-propeller phytase-encoding genes in culturable Paenibacillus and Bacillus sp. from the rhizosphere of pasture plants on volcanic soils. FEMS Microbiol Ecol. 2011, 75, 163–172. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Landi, L.; Renella, G. Role of Phosphatase Enzymes in Soil. In Phosphorus in Action; Springer, 2011; pp. 215–243. [Google Scholar]
- Wouter, J.T.M.; Buijsman, P.J. Secretion of alkaline phosphatase by Bacillus licheniformis 749/C during growth in batch and chemostat cultures. FEMS Microbiol. 1980, 7, 91–95. [Google Scholar] [CrossRef]
- Magadlela, A.; Lembede, Z.; Egbewale, S.O.; Olaniran, A.O. The metabolic potential of soil microorganisms and enzymes in phosphorus deficient KwaZulu Natal grassland ecosystem soil. Appl. Soil Ecol. 2023, 181, 1–11. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Moorhead, D.L. Resource allocation to extracellular enzyme production: a model for nitrogen and phosphorus control of litter decomposition. Soil Biol. Biochem. 1994, 26, 1305–1311. [Google Scholar] [CrossRef]
- Adnan, A.; Mavinic, D.S.; Koch, F.A. Pilot-scale study of phosphorus recovery through struvite crystallization-examining to process feasibility. J. Environ. Health Sci. Eng. 2003, 2, 315–324. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic placement of the 16S rRNA gene from 32 isolates from rhizosphere soil (black), non-rhizosphere soil (brown) and coralloid roots (green) of Encephalartos villosus grown in Oceanview in the Eastern Cape Province, South Africa.
Figure 1.
Phylogenetic placement of the 16S rRNA gene from 32 isolates from rhizosphere soil (black), non-rhizosphere soil (brown) and coralloid roots (green) of Encephalartos villosus grown in Oceanview in the Eastern Cape Province, South Africa.

Figure 2.
Phylogenetic placement of the 16S rRNA gene from 36 isolates from rhizosphere soil (black), non-rhizosphere soil (brown) and coralloid roots (green) of Encephalartos villosus grown in Rhebu in the Eastern Cape Province, South Africa.
Figure 2.
Phylogenetic placement of the 16S rRNA gene from 36 isolates from rhizosphere soil (black), non-rhizosphere soil (brown) and coralloid roots (green) of Encephalartos villosus grown in Rhebu in the Eastern Cape Province, South Africa.

Figure 3.
Phylogenetic placement of the 16S rRNA gene from 68 isolates from rhizosphere soil (black), non-rhizosphere soil (brown) and coralloid roots (green) of Encephalartos villosus grown in Oceanview and Rhebu in the Eastern Cape Province, South Africa.
Figure 3.
Phylogenetic placement of the 16S rRNA gene from 68 isolates from rhizosphere soil (black), non-rhizosphere soil (brown) and coralloid roots (green) of Encephalartos villosus grown in Oceanview and Rhebu in the Eastern Cape Province, South Africa.
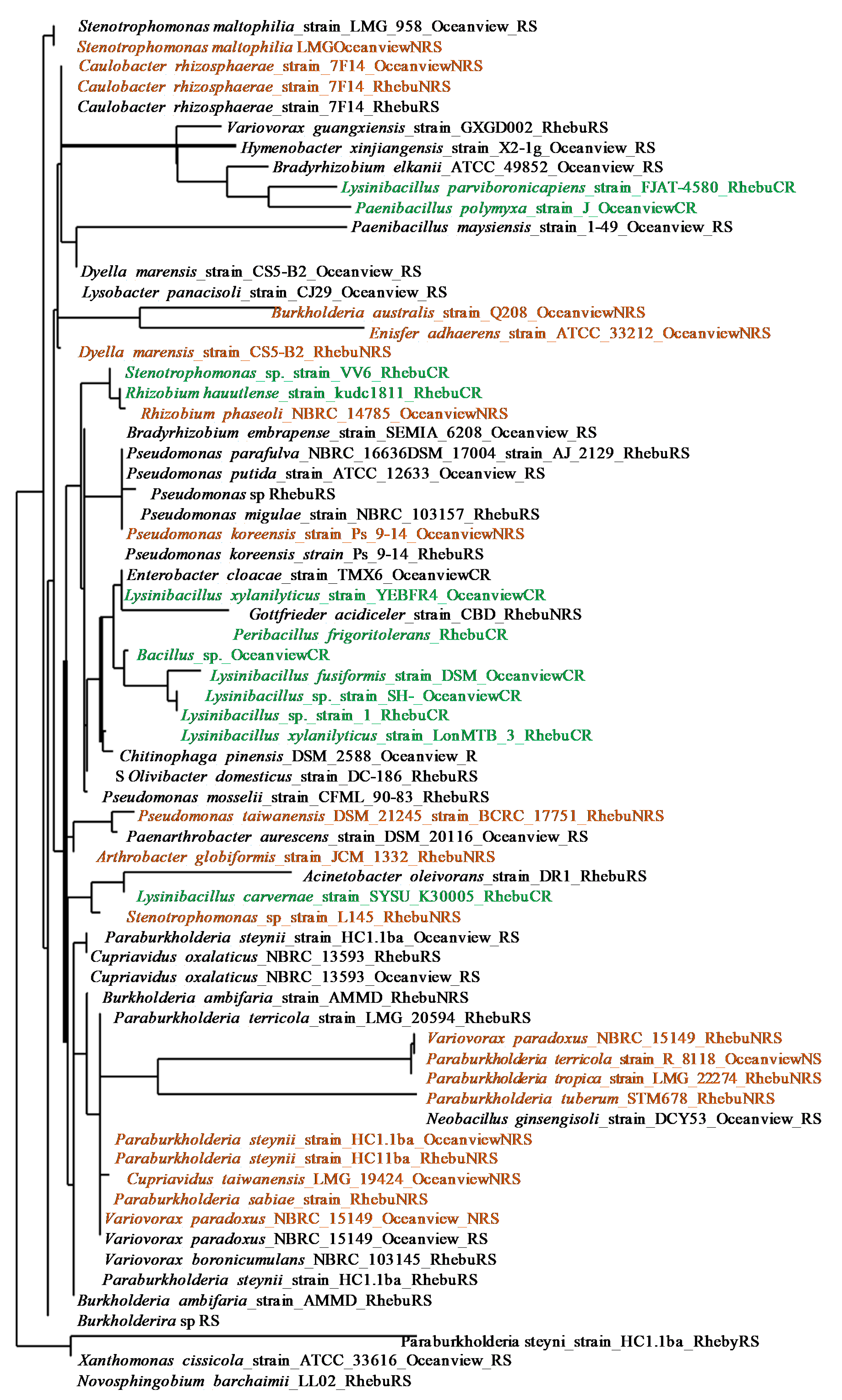
Figure 4.
Soil extracellular enzyme activities in Encephalartos villosus rhizosphere and non-rhizosphere soils collected in Rhebu and Oceanview, Eastern Cape: (a) Nitrate reductase activity; (b) Acid phosphatase activity; (c) Alkaline phosphatase activity; (d) N-acetylglucosaminidase activity. In each graph, different letters denote significant differences in the enzyme activity of soil samples within study sites (independent samples t-test, p≤0.05, means±SE, n=32).
Figure 4.
Soil extracellular enzyme activities in Encephalartos villosus rhizosphere and non-rhizosphere soils collected in Rhebu and Oceanview, Eastern Cape: (a) Nitrate reductase activity; (b) Acid phosphatase activity; (c) Alkaline phosphatase activity; (d) N-acetylglucosaminidase activity. In each graph, different letters denote significant differences in the enzyme activity of soil samples within study sites (independent samples t-test, p≤0.05, means±SE, n=32).
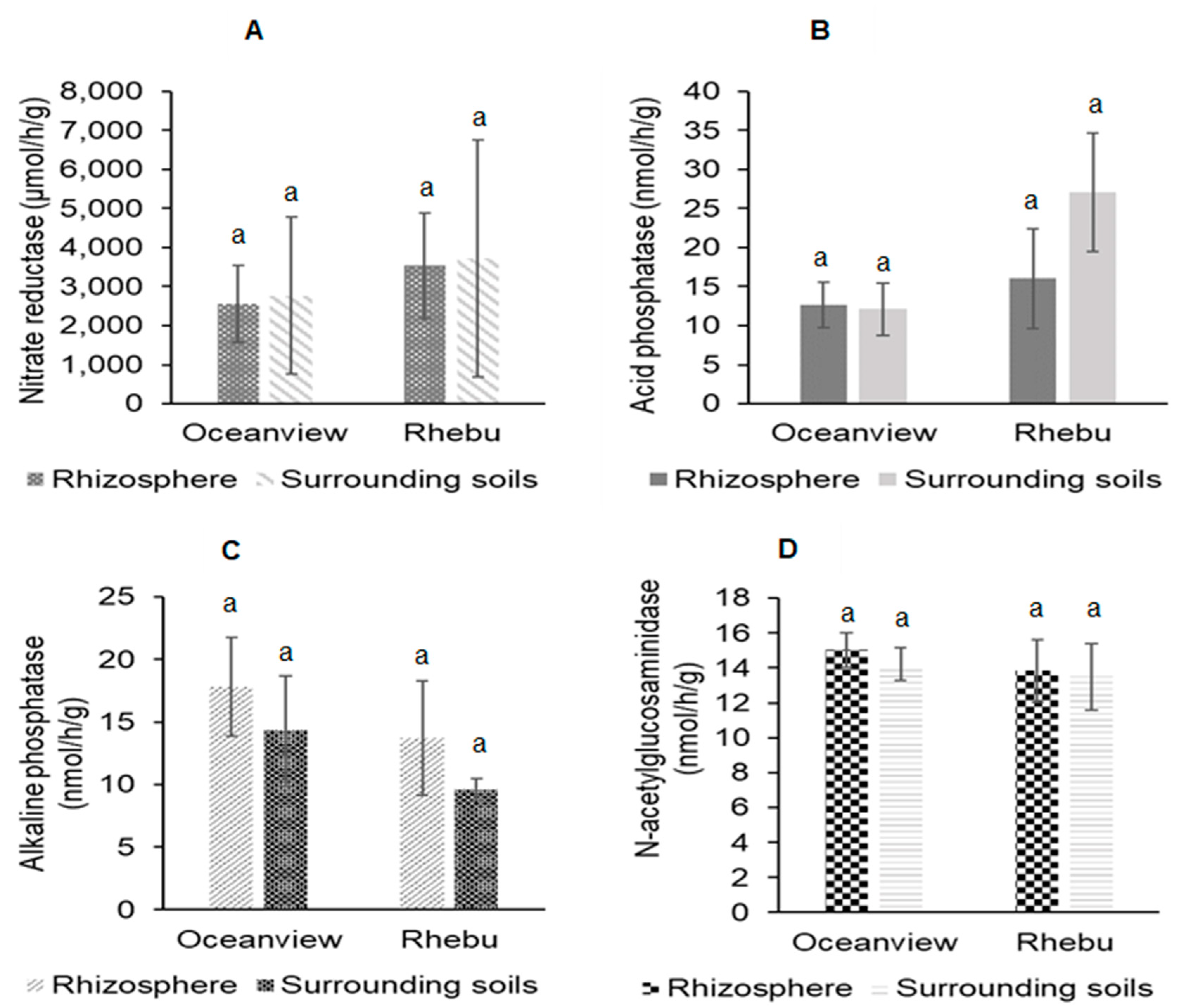
Figure 5.
Leaf length (cm), number of leaves, Leaf nitrogen (mmol g-1) and phosphorus concentration (umol g-1) and nitrogen source reliance of Encephalartos villosus plants growing in Rhebu and Oceanview, Eastern Cape. (a) Leaf length; (b) Number of leaves, (c) Leaf nitrogen; (d) Leaf phosphorus; (e) Percentage nitrogen derived from the atmosphere; (f) Nitrogen source reliance. The values represent the means ± SE, n=16.
Figure 5.
Leaf length (cm), number of leaves, Leaf nitrogen (mmol g-1) and phosphorus concentration (umol g-1) and nitrogen source reliance of Encephalartos villosus plants growing in Rhebu and Oceanview, Eastern Cape. (a) Leaf length; (b) Number of leaves, (c) Leaf nitrogen; (d) Leaf phosphorus; (e) Percentage nitrogen derived from the atmosphere; (f) Nitrogen source reliance. The values represent the means ± SE, n=16.
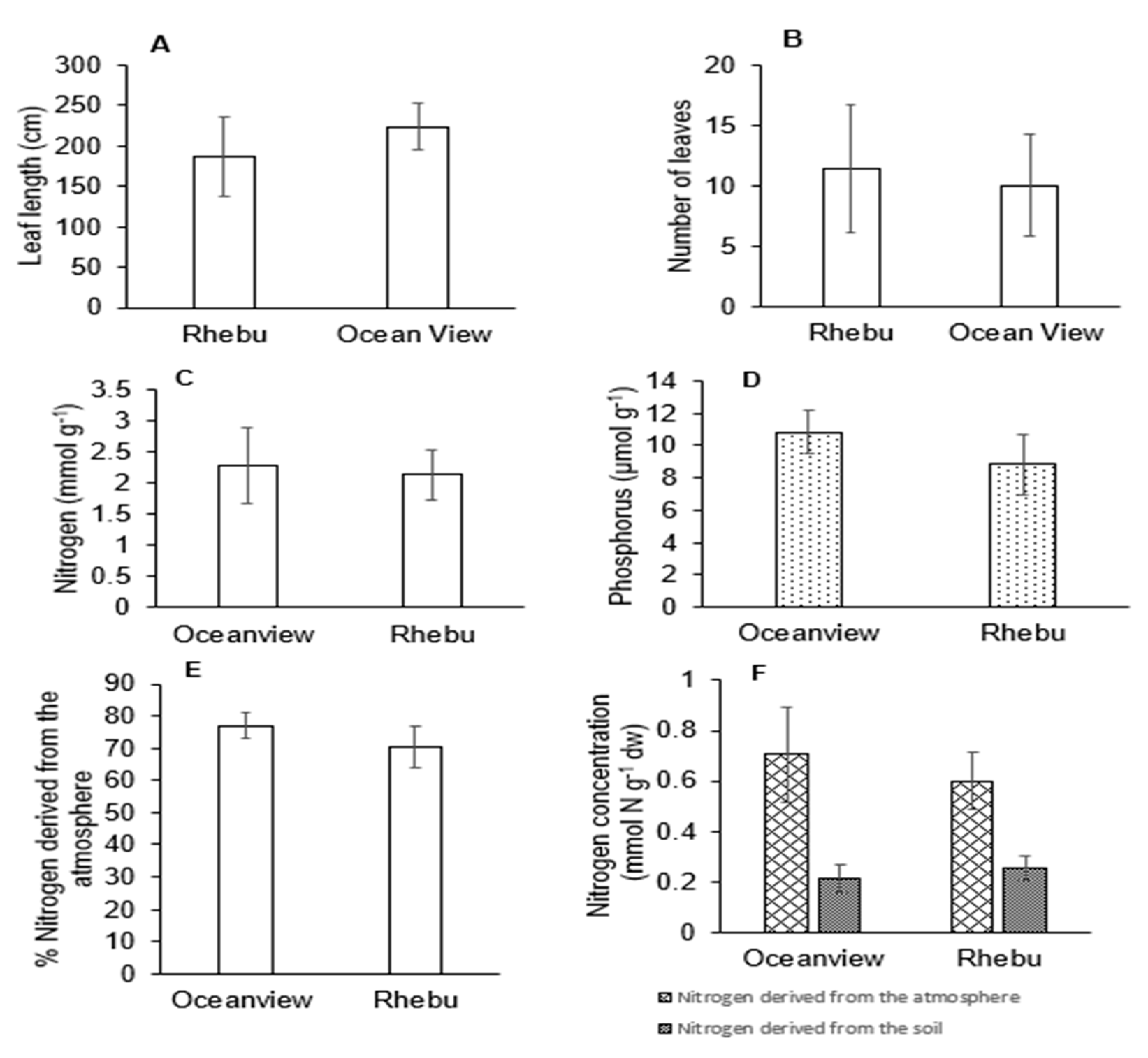
Figure 6.
Correlations between soil characteristics, leaf nutrients, extracellular enzyme activities and N source reliance of Encephalartos villosus plants growing in Rhebu and Oceanview. A: concentration of N derived from the atmosphere, B: plant N, C: soil Cu, D: alkaline phosphatase activity, E: number of leaves, F= plant P, G: concentration N derived from the soil, H: soil pH, I: N-acetylglucosaminidase activity, J: soil P, K: soil N, L: soil Mn, M: soil Mg, N: %NDFA, O: soil K, P: total cations, Q: acid phosphatase activity, R: soil Ca, S: exchange acidity, T: soil Zn, and U: leaf length (principal component analysis).
Figure 6.
Correlations between soil characteristics, leaf nutrients, extracellular enzyme activities and N source reliance of Encephalartos villosus plants growing in Rhebu and Oceanview. A: concentration of N derived from the atmosphere, B: plant N, C: soil Cu, D: alkaline phosphatase activity, E: number of leaves, F= plant P, G: concentration N derived from the soil, H: soil pH, I: N-acetylglucosaminidase activity, J: soil P, K: soil N, L: soil Mn, M: soil Mg, N: %NDFA, O: soil K, P: total cations, Q: acid phosphatase activity, R: soil Ca, S: exchange acidity, T: soil Zn, and U: leaf length (principal component analysis).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



