
Submitted:
21 September 2023
Posted:
22 September 2023
You are already at the latest version
Abstract
Fungal pathogens pose a major threat to food production worldwide. Traditionally, chemical fungicides have been the primary means of controlling these pathogens, but many of these fungicides have recently come under increased scrutiny due to their negative effects on the health of humans, animals, and the environment. However, the use of fungicides with less of an environmental impact tends to result in the development of resistance in populations of phytopathogenic fungi. Therefore, new environmentally friendly alternatives that provide adequate levels of disease control are needed to replace chemical fungicides—if not completely, at least partially. A number of alternatives to conventional chemical fungicides have been developed, including plant defence elicitors (PDEs), biological control agents (fungi, bacteria and mycoviruses), biofungicides, RNA interference (RNAi) methods, and resistance breeding. This article reviews the conventional and alternative methods available to manage fungal pathogens, discusses their strengths and weaknesses, and identifies potential areas for future research.
Keywords:
alternative methods
; fungal disease management
; plants
Introduction
Plant pathogens pose a significant threat to the agricultural industry and are one of the most important factors in agricultural yield losses and food insecurity across the globe. Fungal pathogens alone may account for up to 20% of worldwide yield losses [1]. Conventional breeding techniques cannot provide adequate protection against fungal pathogens for many crops because fungal pathogens are able to overcome introduced genetic resistance [2]. These pathogens pose a particularly serious problem in perennial crops such as apples and other tree fruits. Orchards are expected to last 20–30 years, making it unrealistic to replace vulnerable cultivars with resistant ones, especially since resistance can be overcome long before the end of an orchard’s productive life. Considering that genetic resistance is generally not sustainable and the development of resistant cultivars takes many years, disease control has relied for decades on the application of chemical fungicides [2]. Although very effective, these fungicides are notoriousfor their hazardous effects on human and animal health as well as for their environmental toxicity [3,4]. This has led to increased restrictions on their use, ranging from reductions in the number of applications per crop season to the outright removal of specific products from the market [3]. In addition, plant pathogenic fungi may develop resistance to chemical fungicides, particularly single-site fungicides are more likely to lead to the development of resistance in fungal pathogen populations. In the past few decades, numerous disease management strategies have been developed as an alternative to traditional chemical fungicides and breeding methods, including the use of plant defence elicitors (PDEs), biocontrols or biofungicides, and RNA interference (RNAi). We have reached a crossroads, in which these alternatives to chemical fungicides will be called on to play an increasingly important role in disease management. The present article summarizes the current state of fungal disease management strategies, discusses the strengths and weaknesses of their mode of action, and draws conclusions on the future of fungal disease management.
Chemical Fungicides
Fungicides can be broadly defined as chemical substances used to control fungal diseases by inhibiting the growth of pathogenic fungi or by eradicating them completely. Fungicides can be classified as inorganic or organic based on their chemical composition. Inorganic fungicides do not contain carbon in their structure and are typically based on sulphur or metal ions. This group of fungicides has been in use since the discovery of Bordeaux mixture (copper sulphate pentahydrate and lime) by Pierre-Marie Alexis Millardet in the in the late 19th century [5]. Centuries after this discovery, copper- and sulphur-based fungicides are still used extensively in conventional and organic agriculture [6,7,8,9]. Examples of modern inorganic fungicides include copper sulphate, copper oxychloride, and copper hydroxide [10,11]. In contrast, organic fungicides contain carbon atoms in their structure [6]. These organic synthetic compounds have become more popular, although inorganic fungicides continue to be used in modern agriculture. All synthetic inorganic or organic fungicides, regardless of their composition, can be divided into two distinct classes based on their mobility in the plant: contact (protective) fungicides, which remain on the surface of the plant, and systemic (mobile/curative) fungicides, which are absorbed into the plant.
Contact fungicides typically have a wide range of action against different fungal pathogens and are effective in preventing the infection of plant tissues. They usually achieve this by killing fungal spores or by inhibiting their germination before they penetrate and colonize the host tissues [6,12]. Contact fungicides are not effective in a curative strategy and to be effective, must be applied before the pathogen infects the plant [13]. Most contact fungicides are not absorbed by the plant and remain on the plant tissue surface. However, the frequency of application must be carefully monitored, since contact fungicides can become phytotoxic in the rare cases when they are absorbed [14]. Because contact fungicides usually remain on the plant surface, protection is temporary and can be quickly lost due to rainfall or other weather conditions [15,16]. Contact fungicides can also be eliminated from the plant surface by wind or degraded by UV radiation, and therefore their protective action does not exceed 10–12 days [6]. It is also important to bear in mind that contact fungicides are only effective on the leaf surface present at the time of application and thus are less efficacious during heavy leaf growth. Therefore, the effectiveness of contact fungicides is particularly reliant on the proper timing of application, which generally must be done prior to the known or predicted infection periods of the targeted pathogens. Furthermore, since these fungicides are gradually removed from the plant surface, repeated applications during the growing season are necessary for sustained protection. In some circumstances, this characteristic is actually beneficial to growers, since contact fungicides, unlike systemic fungicides, have the advantage of being easily removed from treated produce before it reaches the consumer [6].
Systemic fungicides are a more recent development in disease control and are considered to be more promising than contact fungicides [17]. While providing a protective effect by suppressing spore germination, these fungicides can also be absorbed into plant tissues, either locally or more broadly, and are therefore able to kill fungal pathogens after they have penetrated and infected host tissues [18]. The degree of systemic activity—which ranges from simple translaminar activity in leaf tissues to local spread from the absorption site and mobility within the xylem of the plant—is generally determined by the chemistry of the compound and can play an important role in determining the efficacy of a fungicide against specific fungal pathogens [18,19,20]. Because of the ability of systemic fungicides to be absorbed in plant tissues, using them to treat plant materials has become routine practice, and the seeds of most agricultural crops are treated with systemic fungicides to protect against both seed- and soil-borne pathogens [21]. Although systemic fungicides are highly effective, most of the compounds involved operate through a single mode of action (i.e. they generally target a single essential fungal enzyme or metabolic pathway) and therefore are extremely vulnerable to the development of resistance by target fungal pathogens [22].
Disadvantages of Chemical Fungicides: Environmental Toxicity and Resistance Development
Despite their high efficacy, both contact and systemic fungicides have numerous drawbacks associated with their use. One substantial shortcoming is that, due to their lack of specificity, chemical fungicides can disrupt both beneficial and pathogenic micro-organisms. For instance, the application of fungicides to mango leaves has been demonstrated to eliminate many endophytes, creating a window of opportunity for pathogens to colonize the tissues formerly that the endophytes formerly inhabited [23]. While this phenomenon has been most readily observed in foliar spray treatments, fungicidal seed treatments have also been associated with similar reductions in beneficial endophytes, and the negative consequences of fungicides on soil microbial communities are well documented [24,25,26,27,28]. Indeed, mounting evidence suggests that seeds protected with systemic fungicides may negatively impact plant health and vigour by targeting beneficial endophytes in the absence of pathogen pressure, the practice may actually be counterproductive [21]. Therefore, the application of chemical fungicides can have negative consequences for plant health and yield by eliminating beneficial microbes that promote growth, development, and resistance to biotic and abiotic stresses.
Fungicides can severely impact the aquatic environment, as they are able to enter aquatic ecosystems through different ways including wastewater, runoff, and subsurface drainage [29{Bereswill, 2012 #260]], and can be toxic to a wide range of aquatic organisms, including algae, fish, and invertebrates [30,31]. Furthermore, fungicides could harm important pollinators like bees through mechanisms such as the impairment of larval and physiological development, the promotion of increased sensitivity to other pesticides, and increased mortality [32]. Exposure to fungicides can lead to acute and chronic neurotoxicity in humans, and thus significantly impact human health [33].
In addition to fungicides’ environmental toxicity, concerns have been raised over the durability of fungicide efficacy. Fungi have tremendous evolutionary potential to rapidly develop resistance against fungicides, due to the intense selective pressure exerted by repeated fungicide applications [22]. Mutations in DNA sequences can arise from errors in DNA replication, damage from UV radiation, or exposure to mutagens or viral infections. Environmental stress may play a significant role in determining the rate of mutation. For example, stress from increasing temperatures accelerated the rate observed in the fungal pathogen Zymoseptoria tritici [34]. Although mutations are inherently random, those that result in enhanced resistance to fungicides will be positively selected for by the eradication of strains without resistance. Over time, this inevitably results in fungicide-resistant strains of the targeted pathogen [22].
Fungicide resistance is a stable and heritable change in an individual fungus that results in a reduction in its susceptibility to fungicides. Fungicide resistance is well documented to develop more often against single-site fungicides than those with a multi-site mode of action, making modern synthetic fungicides especially vulnerable to resistance development [22]. The threat of fungicide resistance is a major concern to growers worldwide, and numerous strategies are employed in order to prevent its emergence. The Fungicide Resistance Action Committee broadly divides fungicides by their mode of action in order to identify those with potential resistance concerns. Its recommendations include applying multiple fungicides with varying modes of action over the course of the growing season, restricting the use of the fungicides most likely to induce resistance with repeated use, and prioritizing multi-site fungicides, to which fungi are less likely to develop resistance [35]. To date, over 43 different modes of action have been identified, although the mechanisms for some of these are not yet known [35].
In conclusion, while chemical fungicides are an extremely effective tool—at least in the short term—for reducing disease incidence in the crops, they have harmful effects on beneficial plant microbiota, the health of humans and other animals, and on the environment. These factors, in addition to the rising threat of fungicide resistance, have led to increasing restrictions on the use of chemical fungicides. Given these challenges, conventional chemical fungicides must be complemented with cost-effective, eco-friendly alternatives in order to maintain appropriate levels of disease control with the absence or reduced usage of these vital compounds.
Alternative Management of Fungal Diseases
Improving Plants’ Genetic Resistance Through the Use of R and S Genes
Plants have evolved numerous genetic defence mechanisms to protect themselves from pathogens. Growers have long relied on the manipulation of these mechanisms, traditionally by breeding for resistance as a way to reduce crops’ susceptibility to fungal pathogens. Host plants can recognize non-specialized fungal pathogens by toll-like receptors that detect pathogen-associated molecular patterns, in turn activating the host’s downstream defense mechanisms. Specialized pathogens are able to overcome these basal defence mechanisms by secreting effector molecules into host plants [36]. However, the co-evolution of plants and fungal pathogens over millennia has provided plants with a means of defence against effector molecules: resistance (R) genes. The R-gene family is incredibly diverse and well conserved in plant species. R-genes encode for nucleotide-binding leucine-rich receptors (NLRs), which collectively recognize a broad spectrum of plant pathogens and pests, inducing an array of resistance mechanisms in response to infection or predation [37,38]. NLRs are activated by the binding and recognition of pathogen effectors but, in some cases, may detect a pathogen indirectly, generally by recognizing pathogen modified host proteins.. An example of this form of recognition occurs in Arabidopsis in response to Pseudomonas syringae infection, where the effector (in this case, a protease) cleaves the Arabidopsis PBS1 kinase, triggering its recognition by the NLR RPS5 [39]. The successful recognition of effector molecules or effector-modified host proteins in plants by the associated NLR typically results in effector-triggered immunity, a localized response characterized by a hypersensitive reaction (HR) in which the plant cells near the infection undergo apoptosis [40,41]. In addition, recognition of a pathogen by plant pattern-recognition receptors (PAMPs) or an effector protein by R proteins triggers the production of salicylic acid (SA) and the downstream induction of broad, systemic defence mechanisms against subsequent infections, triggered independently of the HR response [42,43].
The direct or indirect recognition of effector proteins by R-gene-encoded receptors involves a gene-for-gene relationship in which the R-gene receptor identifies a single effector protein (encoded by a matching avirulence [Avr] gene); therefore, a host with a given R-gene will be resistant to a pathogen with the matching Avr gene [44]. In most plant-pathogen systems, the host and pathogen species may collectively have numerous R- or Avr genes. For example, twenty R-genes have been identified in apples (Malus x domestica) that match the corresponding Avr genes identified in the pathogen Venturia inaequalis, which causes apple scab. However, it should be noted that no single cultivar or individual line will contain all these resistance genes—for example, Honeycrisp apples have Rvi19 and Rvi20 in their genomes, while Golden Delicious cultivars contain Rvi1. Therefore, while many apple cultivars have some resistance to Venturia inaequalis, these cultivars are still vulnerable to some Venturia inaequalis strains that do not have corresponding Avr genes [45].
A typical mechanism in pathogens for overcoming host resistance is Avr gene mutations, to prevent the product (or activity, in the case of indirect mechanisms) from being recognized by R-gene-encoded receptors. If the effector is recognized, pathogens can also overcome resistance by interfering with the host response [46]. The presence of an R-gene in a host plant population will naturally select for pathogens in which the corresponding Avr gene has been lost or modified so that it is no longer recognized by the R-gene-encoded receptor. In turn, successful mutations in the Avr gene will induce selection pressure on host plants for R-genes which impart resistance to the mutated effector. Thus, host plants and their pathogens are continuously engaged in an evolutionary arms race and, in wild populations, the frequencies of Avr and R-genes will cycle over time [47,48]. In modern agricultural settings, the uniformity of resistance genes in a population may accelerate the selection process, leading to rapid loss of resistance in these settings [49].
The identification of R-genes and their incorporation in economically important crops is a vital pillar in the development of resistant plants. Along with the use of conventional fungicides, resistance breeding techniques have served as the most effective method of disease control for decades, particularly in annual crops [50]. Although resistance breeding is also practiced in perennial crops, its effectiveness is often limited by the lifetime of the crop. Modern tree fruit crops, for instance, are expected to have lifespans of 20–30 years, giving ample time for selective pressure from resistant cultivars to result in pathogens overcoming the associated R-genes [49]. This is particularly problematic since introducing cultivars with new resistance genes is difficult due to orchards’ long lifespans. Furthermore, plant breeding is very time consuming and, in recent years, plant breeders have relied on transgenic tools or gene transformation to expedite resistance-breeding efforts, since they allow the faster introduction of R-genes from otherwise incompatible species as well as from compatible species [51].
Numerous strategies have been developed to reduce the ability of fungal pathogens to overcome R-genes, such as rotating R-genes in a field (most suitable for annual crops), mixing cultivars with distinct R-genes in a field or between fields, and pyramiding multiple R-genes in a single cultivar to confer more durable resistance [49]. Somewhat like multi-site fungicides, pyramiding R-genes in a single cultivar makes it more difficult for pathogens to overcome resistance despite their evolutionary potential [52]. However, pathogens are still capable of overcoming multiple R-genes in the same host plant. For example, the oomycete pathogen Phytopthora infestans can escape multiple resistance genes in potato [53]. The breakdown of resistance to rust fungi in cereal crops under different strategies was recently modelled, and it was demonstrated that, although pyramiding could provide the most effective pathogen resistance, this resistance is less durable when mutation rates in the pathogen population are high [49]. Under such conditions, mixing or rotating crops was more successful at delaying the breakdown of resistance to different R-genes. For example, in mixed populations, the breakdown of resistance to one major R-gene was correlated with increased durability of the other R-genes in the population. Rotations were particularly successful, since they were modelled so that pathogens were consistently challenged with new R-genes. Consequently, the authors concluded that rotating different pyramids of R-genes was the most promising method of ensuring durable R-gene resistance [49]. Of course, although this strategy is feasible in annual crops, it is unlikely to be useable or effective in perennial crops, such as apples, pears, and cherries, which have longer lifespans and a juvenile period.
Beyond the introduction of R-genes in susceptible genotypes, advances in genome editing have allowed researchers to identify other mechanisms for reducing disease severity or improving resistance, such as targeting susceptibility (S) genes [54]. S-genes are genes in the host plant required for pathogen infection. Interaction of a pathogen’s effector/toxin molecules with S-genes can assist the pathogen in a variety of ways, such as the recognition and penetration of the host, sustained compatibility between pathogen and host, and the inhibition of immune signalling [55]. Therefore, the genetic silencing or knocking out of S-genes can improve the host plant’s resistance to the pathogen, and is one of the newest frontiers in conferring durable pathogen resistance [54]. Recently, CRISPR/Cas9-mediated knockouts of three S-genes in potato, StDND1, StCHL1 and StDMR6, increased resistance to potato late blight, caused by Phytophthora infestans [56]. Likewise, in apple, the expression of the MdCNGC2 gene, which encodes a cyclic nucleotide-gated ion channel, was observed to be strongly induced by Botryosphaeria dothidea infection in susceptible cultivars [57]. Improved resistance to the pathogen was observed with both virus-induced gene silencing and CRISPR/Cas9-mediated mutagenesis of MdCNGC2 [57]. To date, targeting S-genes has proven to be a successful strategy for inducing disease resistance in a number of crop systems, including cucumbers, rice and tomato [58].
Directly introducing resistance in crops is an effective disease management strategy. However, while both R-genes and S-genes can be modified or integrated in the host genome to improve disease resistance, the process is costly, laborious, and time consuming. In addition, the rapid breakdown of resistance in the field makes resistant cultivars less effective in long-lived crops. Therefore, complementary tools are needed to help delay the breakdown of resistance in crops that cannot be rotated annually.
Use of Plant Defence Elicitors
While pathogen-triggered immunity coordinates the host’s defence against specific pathogens, systemic acquired resistance (SAR) or induced systemic resistance (ISR) mediates prolonged, broad-range resistance to plant pathogens [59,60]. The induction of SAR is characterized by a local increase in levels of the phytohormone SA, which in turn results in the accumulation of reactive oxygen species (ROS) and promotes the expression of β1,3-glucanase, chitinase, and classical pathogenesis-related (PR) genes (Pr1, Pr2, Pr5). The increased expression of these genes promotes plant defence through various downstream targets [61]. Pathogen infection is associated with a rapid increase in SA levels in apple leaf tissue, and this activity is necessary to induce SAR [62,63]. ISR, however, does not require SA accumulation and instead is associated with the accumulation of jasmonic acid (JA) and ethylene (ET). JA and ET accumulation leads to the activation of downstream targets, including chitinase, β1,3-glucanase and an alternative set of PR genes (PDF1.2, PR3, and PR4), as well as the accumulation of ROS (like in SAR) [64]. Significant overlap and crosstalk occur between the SAR and ISR signalling pathways, and both are associated with protection against different pathogens. For example, SAR provides greater protection against biotrophic or hemi-biotrophic pathogens, but may leave the plant more vulnerable to necrotrophic pathogens. Conversely, ISR provides protection against necrotrophic pathogens and chewing insects at the expense of protection against biotrophs and hemi-biotrophs [65,66]. This is a direct result of the crosstalk between these pathways, since the upregulation of SA is typically associated with a decrease in JA and vice versa, although synergism occurs between these two defence mechanisms in some cases [66].
Inducing plant defence by applying exogenous elicitors is a promising alternative to conventional fungicide applications [67]. The application of exogenous PDEs has been associated with enhanced resistance against a wide range of pathogens in different crops, including cereals, tomato, rubber tree, and apple [68,69,70,71]. Pre-treatment with SA of the leaves of the susceptible apple cultivar Gala was associated with a marked (albeit temporary) increase in the expression of the plant pathogenesis-related genes PR1, PR5 and PR8, as well as of chitinase and β1,3-glucanase, with the treated leaves demonstrating increased resistance to Glomerella leaf spot following in-vitro inoculation with Glomerella cingulate, indicating the induction of SAR [71]. Similarly, exogenous applications of phenylacetic acid have been observed to induce ISR in tobacco, imparting significant resistance to the bacterial soft rot pathogen Pectobacterium carotovum [72]. Elicitor treatment may protect fruit from plant pathogens for a significantly longer period than the temporary uptick in defence-gene-related expression suggests. In this regard, field treatments of Ya Li pear with a 2.5-mM SA spray were demonstrated to produce a remarkable decline in disease incidence and lesion diameter compared to the untreated control. Finally, activities of defence-related enzymes such as peroxidase, phenylalanine ammonia-lyase, chitinase, and β-1,3 glucanase not only increased significantly in the four days following SA application, but these increased activities were still observed after harvest [73].
Since the discovery of SA as an exogenous inducer of SAR, several synthetic PDEs that are analogues of the phytohormones SA and JA have been developed. The ones most commonly used in the past few decades are benzo(1,2,3)thiadiazole-7-carbothioic acid S-methyl ester (BTH) and 2,6-dichloro-isonicotonic acid (INA), which are preferred for their increased efficiency and reduced phytotoxicity compared to the original SA [61].
Phytohormones and their synthetic analogues are not the only methods for inducing plant defence mechanisms. Peptides, polysaccharides, and lipids isolated from plants, fungi, and bacteria can also serve as PDEs. For example, chitosan, which can be readily obtained from fungal cell walls, is well known for its ability to induce host defenses, although this compound also demonstrates direct fungicidal activity [74]. The application of ZhiNenCong (ZNC), an extract of Paecilomyces variotii, stimulated immunity in Aribidopsis thaliana to bacterial infection and in potato to Phytopthora infestans infection via SA-dependent signalling pathways. Cell wall extract from the mushroom Pleurotus ostreatus enhanced defence against Septoria and mildew in wheat and grapevine, respectively [75,76]. Similarly, extracts of giant knotweed (Reynoutria sachalinensis) improved the resistance of courgette to Podosphaera xanthii in an SA-dependent manner [77]. Extracts from the seaweeds Ascophyllum nodosum, Cystoseira myriophylloides, Laminaria digitata, and Fucus spiralis represent a significant portion of plant-based elicitors [78,79].
The application of exogenous plant defence elicitors that can stimulate SAR or ISR responses before pathogen infection is widely considered to be an eco-friendlier alternative for disease control in plants [80]. However, PDEs have been shown to be significantly less effective than conventional chemical fungicides in many instances [73], which could be partially attributed to the inherent crosstalk between the SAR and ISR signalling pathways, since the induction of one pathway is often associated with the inhibition of the other. Therefore, the activation of SAR may improve a host’s defence against biotrophic and hemi-biotrophic fungal pathogens, but leave the host more vulnerable to necrotrophs [66]. Furthermore, the induction of plant defences requires the significant allocation of the host plant’s resources and thus may be associated with a decrease in overall plant fitness [81,82]. For this reason, many PDEs are very rarely used, and therefore do not fully replace conventional chemical fungicides. Conversely, the use of elicitor products improved retention and yield, particularly in the case of seaweed extracts, which may also act as biostimulants [83,84]. These products can improve disease protection when applied in conjunction with fungicides. For example, the use of Actigard® (an SA analogue) increased the efficacy of dimethomorph, mancozeb, and azoxystrobin in preventing tobacco blue mould caused by Peronospora tabacina [85]. Consequently, despite their limitations, PDEs remain an attractive disease control tool, with their usefulness determined by the context of their use. Although PDEs are inefficient when used on their own, when incorporated in integrated disease management programs, they could potentially reduce the use of chemical fungicides, thus lessening the environmental exposure to these pesticides and potentially slowing the development of fungicide resistance in pathogens. The increased use of PDEs combined with biological control agents or biofungicides could provide an economically acceptable level of control.
Biological Control and Biofungicides
Plant tissue is colonized by a wide range of microbiota, which may be endophytic (colonizing the inner surface of the plant) or epiphytic (residing on the outer surface of the plant). Endophytic and epiphytic microbes do not cause disease symptoms, but their interaction with the host plant is not strictly mutualistic. Many plant-associated microbes play vital roles in promoting plant health, including influencing their hosts’ disease resistance [86]. This resident microbiota can antagonize plant pathogens, and thus are a focus of current research on plant disease management [87]. A common mechanism for this antagonism is direct competition for resources, which reduces the availability of both physical space and nutrients in host tissues and, in turn, the opportunity for pathogen infection [88,89]. Plant-associated microbiota can also significantly impact host resistance to pathogen infection through colonization, which triggers the host’s localized defence mechanisms through ISR, resulting in a more rapid induction of the defence response when a plant pathogen subsequently colonizes the host [90]. Furthermore, plant-associated microbes can target and antagonize plant pathogens either directly, by secreting antifungal or antibacterial secondary metabolites that reduce pathogens’ growth, or indirectly, by secreting metabolites that enhance the host’s production of antifungal or antibacterial metabolites [91,92,93]. Some endophytes, such as members of the genus Trichoderma, can antagonize plant pathogens directly through mycoparasitism [94,95].
Biocontrol agents (also known as bioagents) are generally plant-associated microbiota that have been screened and selected for use in crop systems in order to improve plant fitness, induce plant defence mechanisms, and antagonize pathogens [96,97,98]. Microbes can be beneficial to plant health and disease control through a multitude of mechanisms, and many different biocontrols have been developed to protect against pathogens, particularly at the post-harvest stage. The current literature on biocontrol focuses heavily on a small number of beneficial genera. For example, species of the soil-borne genus Trichoderma are extremely effective in controlling soil-borne and foliar pathogens, and have been demonstrated to perform well against a number of pathogenic species, including Rhizoctonia solani, Fusarium oxysporum, and Bipolaris sorokinia. The antifungal properties of these soil-borne species typically arise from a combination of direct competition, mycoparasitism, and the secretion of antifungal compounds [95,98,99,100,101,102]. Members of the genus Aureobasidium, most notably Aureobasidium pullulans, show promise in controlling post-harvest pathogens, such as Botrytis cinerea, Penicillum expansum, and Diplodia seriata [103,104,105,106,107,108]. Like Trichoderma, A. pullulans antagonizes pathogens through direct competition for space and resources as well as by the secretion of antimicrobial compounds [109]. Additionally, bacterial genera such as Pseudomonas, Bacillus, and Agrobacterium have demonstrated an excellent ability to suppress economically significant plant pathogens [110,111,112].
Extensive laboratory research has resulted in commercial biocontrol agents being available in varying degrees around the world. Trichoderma spp. is the best established biocontrol agent, and several products containing various Trichoderma species have been commercialized since the 1970s [113]. Products incorporating fungi such as Aspergillus flavus and A. pullulans, yeasts such as Candida oleophila, and bacteria such as Pseudomonas, Bacillus, and Agrobacterium, have been approved in Europe and the United States [113]. Biocontrol agents are often considered to be less vulnerable to resistance development in pathogen populations, because, unlike synthetic fungicides, they often have multiple modes of action, although this is not always the case and more research is needed to determine their modes of action [113,114]. Indeed, very few studies have been conducted on the probability of resistance development in fungal pathogens to biological control agents [115]. Despite the potential advantages of biocontrol products, they are not universally preferred to chemical fungicides because their efficacy can vary significantly. Conditions in the field are variable and difficult to predict, and microbial biocontrol agents can behave unreliably in uncontrolled conditions [116].
Mycoviruses also demonstrate remarkable promise as candidates for biocontrol, since mycoviral infections in plant pathogens are generally associated with a reduction in virulence as a result of the RNA silencing (RNAi) of pathogen and host genes [117]. A major advantage of this form of biocontrol is its ability to spread within a pathogen population following introduction. This spread occurs primarily through fungal hyphal anastomosis (hyphal fusion), a key process required in these pathogens for homeostasis and genetic exchange [118,119]. Upon a mycovirus’ successful infection of a pathogen, the mycovirus will be passed down to the pathogen’s progeny, thereby reducing pathogenicity across multiple generations [120]. To date, numerous mycoviruses capable of inducing hypovirulence in pathogen populations have been identified, including AsHV1, which reduces the virulence and growth of Alternaria alternata, and BcPV2, which induces hypovirulence and an absence of conidia in Botrytis cinerea [121,122].
In addition to mycoviruses, bacteriophages (viruses that target bacteria) have also been used in the biocontrol of bacterial plant pathogens. Bacteriophages have a number of advantages, because they are usually genus-specific and able to replicate within their hosts, but do not accumulate in the environment in their hosts’ absence [123]. In some cases, phages can degrade extracellular polysaccharides involved in virulence, as in the case of Erwinia amylovora [124]. Bacteriophage-based biocontrol products typically use a combination of different phages to increase the product’s modes of action as well as improve the range of target pathogen genotypes, and to reduce the chance of acquired pathogen resistance [125].
In contrast to biocontrol agents, biochemical fungicides (or biofungicides) are naturally occurring compounds that can be used to control fungal diseases. Biofungicides can increase a plant’s disease resistance by inducing its defence responses or by directly inhibiting the pathogen. Biofungicides may include compounds screened and selected based on their antimicrobial activity, many derived from biocontrol agents [126] or, alternatively, take the form of whole-cell extracts. Biofungicides have been proven effective against many fungal pathogens. For example, thethe free cell extracts of Pseudomonas have been shown to inhibit the mycelial growth of Alternaria alternata and Fusarium solani in vitro, while SH2, an antifungal compound derived from Streptomyces hydrogenas, was found to control Alternaria brassicicola on radish seeds in vivo [127,128]. Plant extracts are another important source of biofungicides, and can inhibit diverse phytopathogenic fungi such as Botrytis cinerea, Fusarium oxysporum, and Leptosphaeria sacchari (syn. Phoma sorghina) [129,130]. Similarly, natural oils can have potent antifungal activity, reducing the severity of diseases caused by numerous pathogens, such as Fusarium oxysporum, Fusarium solani, Fusarium monliforme, Thilaviopsis paradoxa, Botryodiplodia theobromae, and Rhizoctonia solani in date palm seedlings; Phytophthora parasitica var. nicotianae in tobacco; and Venturia inaequalis in apple [131,132,133].
Currently, both biocontrol agents and biofungicides are viewed as eco-friendly means of disease control. These methods are typically less phytotoxic than synthetic fungicides and are considered to be a more environmentally friendly method for pathogen inhibition or eradication as they do not leave toxic residues [126,134,135]. The continued identification of biocontrol agents and antimicrobial compounds and extracts will be of vital importance in the future development of disease management strategies.
RNAi-Based Disease Management
Small RNA-Based Fungicides
RNA silencing (or RNAi) is a highly conserved mechanism in eukaryotes that allows gene silencing at the transcriptional and post-transcriptional levels by small RNAs (sRNAs). At the transcriptional level, small, double-stranded RNA (dsRNA) is cleaved by the ribonuclease Dicer, and loaded into the RNA-induced initiation of transcriptional gene silencing complex (RITSC), which, utilizing the sRNA as a guide, binds to homologous DNA, leading to methylation of the heterochromatin in this region and, consequently, gene silencing [136]. A similar mechanism is responsible for post-transcriptional gene regulation, in which small dsRNA, after being cleaved by Dicer, is loaded into the RNA induced-silencing complex (RISC), and guides the latter to complementary RNA targets through base pairing [137]. RISC then “slices” the homologous mRNA directly, leading to the degradation of the mRNA transcript [137,138]. The recognition of the mRNA sequence can also lead to translation repression [137,138,139], since RNAi does not require perfect complementarity. The degree of complementarity between the small RNA guide and its target is a major factor in determining the interference mechanism, with lower degrees of complementarity associated with translational repression and higher ones, with mRNA degradation, although translational repression may still occur in plants under conditions of high complementarity [140,141,142,143]. These mechanisms are originally thought to have evolved primarily as a means of protecting the host from viral infections and transposable elements; however, a body of evidence suggests that hosts and pathogens are engaged in an evolutionary arms race, with rapid co-evolution of antiviral RNAi genes in hosts and viral RNAi suppressor genes in viruses [144].
In the last decade, RNAi has generated immense scientific interest as a way to achieve effective and eco-friendly disease control in agricultural settings. This research can broadly be divided into two separate categories: that focusing on host-induced gene silencing (HIGS), which involves the genetic modification of a host plant to express dsRNA that targets a specific pathogen’s pathogenicity related genes, and that focusing on spray-induced gene silencing (SIGS), which involves the exogenous application of similar dsRNAs or sRNA [145,146].
i. Host-Induced Gene Silencing
RNAi can be triggered by the presence of dsRNA, hairpin or transgenic foreign RNA, or viral dsRNA, culminating in the use of the dsRNA as a template sequence to locate and silence matching foreign dsRNA [145]. Taking advantage of this inherent disease control mechanism, researchers have developed transgenic crops that express dsRNA targeting known pathogens and pests [147]. HIGS offers significant advantages over conventional resistance breeding techniques. For instance, HIGS does not require a pool of established resistance genes. Its only limitation is that the researcher must design the dsRNA to target the relevant pathogenicity genes. The use of HIGS to target the parasitism genes of root-knot nematodes is just one example of the degree of improvement provided by HIGS in fine-tuning disease resistance. For years, efforts to combat root-knot nematode parasitism in many crops were hindered largely by the lack of effective, broad-range resistance genes [148]. However, this problem was overcome by creating transgenic Arabidopsis plants that express dsRNA for the parasitism gene 16D10, which confers resistance against a wide range of root-knot nematode species [149]. Thus, HIGS was able to provide a degree of resistance unattainable by conventional methods, and, in the future, other varieties of vulnerable crops will likely be transformed with 16D10 dsRNA [149].
HIGS techniques show significant promise in the control of fungal pathogens. After confirmation was obtained that dsRNA targeting fungal transcripts of Blumeria graminis (the causal agent of powdery mildew) drastically impedes the development of this disease in wheat and barley, transgenic plants were developed that express dsRNA to silence pathogenicity genes. Significant disruption of host-pathogen interactions was demonstrated in these transgenic plants, demonstrating HIG’s potential in controlling fungal pathogens [150]. Since then, HIGS techniques have been applied successfully against the fungal pathogens of a wide variety of crops, including Verticillium in tomato and Magnaporthe oryzae in rice [151,152].
Despite HIGS’ advantages over conventional breeding methods, this emerging technology has many of the same limitations as conventional techniques, namely that introducing new dsRNA in economically important crops and cultivars is a time-consuming process. In addition, significant opposition to genetic modifications may be present in some jurisdictions. Furthermore, although it may be more difficult for fungi to develop resistance to RNAi, there is significant evidence supporting an evolutionary arms race between fungal pathogens and host plants in regard to RNAi [144]. Therefore, there might be a need to “pyramid” dsRNA to target several relevant (and unrelated) pathogenicity genes [52]. Even with the “pyramiding” of dsRNA, the introduction of resistant cultivars remains a suboptimal option for perennial crops with lengthy life cycles. In these crops, HIGS will likely need to be complemented with conventional spray programs or the new SIGS techniques.
ii. Using Spray-Induced Gene Silencing as the Basis for RNA-based Fungicides
SIGS techniques involve the spray application of exogenous dsRNA or sRNA to plants. The efficacy of this method was first demonstrated in barley, where the application of exogenous dsRNA targeting three ergosterol biosynthesis genes critical to the integrity of fungal cell membranes resulted in the effective inhibition of the fungal pathogen Fusarium graminearum [146]. Since then, SIGS has proved effective against a number of fungal pathogens, including Botrytis cinerea and Sclerotinia scletiorum, in a variety of crop systems, including strawberry, tomato, and Arabidopsis [153,154].
While dsRNA was originally thought to be processed directly by the fungal RNAi machinery, recent evidence suggests that SIGS requires the uptake of sprayed dsRNA by the host plant’s stomata[155]. In their study, Biekenkopf et al. demonstrated the processing of dsRNA by host plants and visualized fluorescently labelled dsRNA travelling through the vascular system of plants, indicating its long-distance spreading to distal areas [155]. Significant (~60%) gene silencing of the targeted Shp gene was reported even in aphids that fed on unsprayed distal tissues, confirming the travel of dsRNA through the vascular system to tissues [155]. Indeed, the fungal pathogen Fusarium graemarium was also impacted by the dsRNA, even in the plant roots [155]. Previously, it had been demonstrated that the use of larger dsRNAs resulted in the decreased inhibition of target mRNA when dsRNAs were applied exogenously, suggesting that the lower efficacy of larger dsRNA in SIGS must be a result of poor uptake by fungi (while this has no relation to the efficacy of HIGS) [156]. However, since more recent evidence suggests that dsRNA uptake and processing by the host plant plays a significant role in the SIGS of target pathogens, it seems that the size of dsRNA may instead inhibit the uptake of these molecules by the plant, a process required in SIGS [155]. Unfortunately, while these studies provide some insight into the uptake and dispersal of dsRNA in host plants, more research is needed to elucidate the exact mechanism for dsRNA uptake in plant tissues and, subsequently, by fungal pathogens.
While the use of SIGS in crop systems is advantageous, it should also come with the caveat that dsRNAs are not continuously transcribed by the crop, as is the case with HIGS. Until recently, this represented a major hurdle in the practical application of SIGS biopesticides, especially since dsRNAs are known to have a short half-life. However, the development of environmentally friendly, non-toxic, degradable clay nanosheets allows the sustained release of dsRNA for up to 30 days after application, drastically increasing the duration of protection offered by a single spray application [157]. Since both the clay nanosheets and dsRNA have been demonstrated to be environmentally non-toxic, SIGS offers significant environmental advantages over traditional chemical fungicides [157]. A recent RT-qPCR method for quantifying dsRNA in agricultural soils demonstrated that exogenous dsRNA dissipated to below-detectable levels within hours of application, providing further evidence that this spray technique may be far more eco-friendly than conventional fungicides, which are known to persist in the environment over a longer time scale [3,158]. In addition, because dsRNAs are designed to target specific pathogens, SIGS has the potential to avoid many of conventional fungicides’ off-target effects, which greatly alter the microbiome of crops [159]. In this regard, the specific design of the dsRNA is vital, since RNAi has recently been noted to have off-target effects, such as targeting mRNA with as few as eleven contiguous nucleotides in common with the dsRNA-provided template, thus posing a substantial risk to both human health and the host plant [160,161]. Consequently, extensive research will be required to determine the off-target effects of specific dsRNA on human health, as well as on target crops and their microbiomes. Nevertheless, the potential advantages of SIGS over conventional fungicides are undeniable.
Future Directions
Despite recent advances in disease control, comparing a given method’s efficacy across different studies can be quite difficult, due to differences in crops, cultivars, the timing of treatment, environmental conditions, and targeted pathogens. Although numerous eco-friendly options have been developed to control fungal diseases, many of these are challenging to use effectively in the field. For example, PDEs were initially considered a viable alternative to conventional pesticides, until subsequent field trials demonstrated that they were usually less effective than conventional fungicides [162]. Direct comparisons of these control methods will be required in order to determine their relative efficacy under field conditions.
While biofungicides have been a focus of research efforts for decades and are available in commercial formulations, new bio-degradable, environmentally safe products have significant potential and there is a strong need for their development [163]. Many synthetic fungicides, such as strobilurins, have been developed from natural antimicrobial compounds, and the continued identification, isolation, and production of these compounds could provide a pipeline of highly efficient antifungal agents [128,164]. Although the isolation and use of these antifungal compounds may be appealing, the resulting individual compounds are likely to be similar to modern synthetic fungicides in their reliance on a single target site and therefore, like strobilurins, may be extremely vulnerable to the development of resistance in the targeted pathogens [35]. In contrast, biofungicides produced from the whole-cell extracts of known mutualists, as well as plant extracts and essential oils, appear to be less vulnerable to resistance development due to their potential for multi-site activity. Essential oils and plant extracts may have up to six different modes of action, such as (i) inhibition of cell wall formation; (ii) inhibition of ergosterol synthesis; (iii) inhibition of mitochondrial electron transport; (iv) interference with RNA/DNA and protein synthesis; (v) interference with efflux pumps; and (vi) inhibition of cell division. On the other hand, substantial losses in the efficacy of plant extracts and essential oils in field treatments have been observed compared to experiments under controlled conditions, most likely due to a decrease in the stability of these products during storage or transport, or under field conditions [165]. However, in recent years, significant advances have been made in biopesticide formulation, and several non-liquid preparations have been developed for insecticidal purposes [166]. Similar advances in essential-oil- and plant-extract-based biofungicides will improve their efficacy under field conditions and allow for the more widespread adoption of these important technologies, since the formulation and stability of these products remain the greatest obstacles to their effective use.
Although research on RNAi techniques is in its infancy, both SIGS and HIGS have significant potential as eco-friendly disease control methods. In addition to their high potency and reduced environmental impacts, RNAi techniques may also be less vulnerable to resistance development than modern single-site fungicides. While a single mutation can change the conformation of a target enzyme (for example, CYP51 in ergosterol synthesis) and prevent fungicide binding and activity, RNAi mechanisms are not nearly as vulnerable to evasion, since even the imperfect recognition of an mRNA transcript can still lead to translational repression, and thus deprive the pathogen of a vital protein [140]. Owing to the relatively recent introduction of the HIGS and SIGS techniques, the rate at which fungal pathogens can develop resistance to dsRNA targeting of pathogenicity related genes has yet to be assessed. Nevertheless, the dsRNA targeting of multiple unique pathogenicity genes, presenting the pathogen with a challenge that would require mutations in multiple loci to overcome, should obviously be investigated. However, the generation of hosts expressing dsRNA is a lengthy and laborious process, and would need to be repeated with several dsRNAs unique to each targeted pathogen to ensure enduring broad-spectrum resistance. Considering the strong consumer sentiment against the genetic modification of crops, SIGS will most likely become the preferred method for controlling fungal pathogens through RNAi. Nevertheless, the development of resistant lines with HIGS and, in particular, combining the pyramiding of the expression of unique dsRNA with R-genes and the modification of susceptibility genes could significantly reduce growers’ costs—particularly labour costs—by reducing the need for pesticide applications.
The timing of fungicide applications is usually informed by plant growth stage and/or the predictive modelling of pathogen risk based on weather forecasting, resulting in greater efficiency and reduced fungicide use [167,168,169,170,171]. More recently, assays to detect inoculum levels in the field have been developed, which allow pathogen pressures throughout the growing season to be assessed and the most effective timing for fungicide application to be determined, resulting in a potential reduction in pesticide use and in the rate of fungicide resistance development [172,173]. The monitoring of inoculum levels will be particularly useful in guiding the application of SIGS, since, unlike many chemical fungicides, well-designed dsRNA should be highly specific to the targeted pathogen. The incorporation of environmentally friendly disease control methods in integrated spray programs, informed by predictive modelling and the monitoring of inoculum levels in the field, could significantly reduce our reliance on environmentally toxic chemical fungicides and delay the development of fungicide resistance in economically significant pathogens. For instance, Colletotrichum acutatum, a fungal pathogen causing bitter rot of apple, has been demonstrated to overwinter in trees by infecting the outer bud scales asymptomatically; the primary inoculum identified in orchards in early spring is mainly dispersed by the buds opening in the canopy [174]. Therefore, to manage Colletotrichum acutatum, early applications of fungicides during bud burst are critical to control the levels of primary inoculum and thus prevent secondary infections later in the growing season [174]. Subsequent to bud-break treatments with systemic fungicides, biofungicides or SIGS could be used to control pathogen population levels, reducing the need for additional systemic fungicide applications later in the growing season, delaying the development of fungicide resistance, and reducing the environmental impacts of fungicide use. Weather-basedforecasting models for biocontrol and pathogen populations need to be developed to optimize the timing of applications.
Although PDEs and biocontrols are generally outperformed by other disease control methods in comparative assays, these methods are still beneficial in reducing disease under actual field conditions, particularly when used in combination with other techniques. For instance, co-application of the PDE Actigard® with conventional fungicides significantly increased protection against blue mould of tobacco caused by the oomycete Peronospora tabacina compared to the use of fungicides alone [85]. Combining biocontrol agents with chemical and biological plant defence inducers and chemical fungicides has been suggested as a way to reduce fungicide use, while still providing effective disease control [175]. In the case of biological PDEs and biocontrol agents, this approach requires compatibility with specific conventional fungicides. However, this is another major advantage of RNAi technologies, because RNA constructs, which are designed specifically to target pathogenicity related genes, are likely to be compatible with unrelated biocontrol agents, enabling their co-application. The successful co-application of biocontrol agents and SIGS would present pathogens with simultaneous multiple challenges, such as RNAi targeting key pathogenicity genes, direct competition for space and resources, the induction of host defences, and direct antagonism. Consequently, at a time when SIGS research is still in its infancy, the investigation of combined treatments with biocontrol agents or PDEs could clearly present a significant opportunity for improved disease management.
Conclusion
While chemical fungicides have been one of the most effective methods of disease control for decades, restrictions on their use are increasing due to their negative environmental impacts and consequences for animals and human health, as well as the growing threat of fungicide-resistant pathogens. However, the gap left by the elimination of these fungicides cannot be bridged by traditional breeding methods, because the introduction of new resistance genes in plant species is time and labor intensive, and pathogenic fungi are able to quickly overcome introduced resistance genes in the field. Consequently, for decades, researchers have been searching for effective and environmentally friendly methods for controlling fungal pathogens.
According to the literature, both biofungicides and RNAi-based techniques are highly effective and have significant advantages over conventional methods in terms of environmental sustainability. The rapid breakdown of RNA in the environment suggests that RNAi techniques will be an eco-friendly alternative to chemical fungicides. In addition, owing to the specificity of RNAi, it can be more easily incorporated in integrated pest management systems than conventional fungicides, leading to combined treatments with biocontrols or PDEs. More research is required in order to determine the relative efficacy of these methods under field conditions, and to develop integrated pest management systems that challenge fungal pathogens through a variety of different mechanisms. This will allow a reduction in chemical fungicide use and therefore prolong pathogens’ sensitivity to these vital tools, while reducing environmental and human exposure. In conclusion, the successful integration of SIGS, biofungicides, biocontrols, and PDEs in conventional spray programs will be beneficial not only to crop health and yield, but also to the broader environment and human health, and preserve the efficacy of fungicides by challenging pests with different selection pressures.
Funding Information
This work was supported by NSFGA and Agriculture and Agri-Food Canada through CAP project (J-002190), Agriculture and Agri-Food Canada A base projects (J-002516).
Acknowledgements
Additional support was provided by USDA ARS Project 2090-21000-003-00D to SNY. The authors would like to thank Elizabeth McFarlane, Senior Translator, Translation Bureau, Public Services and Procurement Canada for professional editing of this manuscript.
Conflicts of Interest
The authors declare no conflict of interests related to this study.
References
- Gullino, M.; Leroux, P.; Smith, C. Uses and challenges of novel compounds for plant disease control. Crop Protection 2000, 19, 1–11. [Google Scholar] [CrossRef]
- Kelman, A. Introduction: the importance of research on the control of postharvest diseases of perishable food crops. Phytopathology 1989, 79, 1374. [Google Scholar]
- Ragsdale, N.N.; Sisler, H.D. Social and political implications of managing plant diseases with decreased availability of fungicides in the United States. Annu Rev Phytopathol 1994, 32, 545–557. [Google Scholar] [CrossRef]
- Steinhauer, N.; Kulhanek, K.; Antúnez, K.; Human, H.; Chantawannakul, P.; Chauzat, M.P.; vanEngelsdorp, D. Drivers of colony losses. Curr Opin Insect Sci 2018, 26, 142–148. [Google Scholar] [CrossRef]
- Millardet, P.M.A. The Discovery of Bordeaux Mixture; The American Phytopathological Society: 2018; pp. –1.
- Baibakova, E.; Nefedjeva, E.; Suska-Malawska, M.; Wilk, M.; Sevriukova, G.; Zheltobriukhov, V. Modern Fungicides: Mechanisms of Action, Fungal Resistance and Phytotoxic Effects. Annual Research & Review in Biology 2019, 1-16. [CrossRef]
- Tamm, L.; Thuerig, B.; Apostolov, S.; Blogg, H.; Borgo, E.; Corneo, P.E.; Fittje, S.; de Palma, M.; Donko, A.; Experton, C.; et al. Use of Copper-Based Fungicides in Organic Agriculture in Twelve European Countries. Agronomy 2022, 12. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Osdaghi, E.; Behlau, F.; Köhl, J.; Jones, J.B.; Aubertot, J.-N. Thirteen decades of antimicrobial copper compounds applied in agriculture. A review. Agronomy for Sustainable Development 2018, 38, 28. [Google Scholar] [CrossRef]
- Karuppuchamy, P.; Venugopal, S. Chapter 21 - Integrated Pest Management. In Ecofriendly Pest Management for Food Security, Omkar, Ed.; Academic Press: San Diego, 2016; pp. 651–684. [Google Scholar]
- Anna La, T.; Valeria, I.; Federica, C. Copper in plant protection: current situation and prospects. Phytopathologia Mediterranea 2018, 57. [Google Scholar] [CrossRef]
- Oziengbe; Osazee. Antifungal Activity of Copper Sulphate Against Colletotrichum Gloeosporioides. Journal of Asian Scientific Research 2012, 2, 835–839. [Google Scholar]
- Oliver, R.; Hewitt, H.G. Fungicides in crop protection: Second edition. CABI Journal. CABI International 2014, 1–190. [Google Scholar]
- Dias, M. Phytotoxicity: An Overview of the Physiological Responses of Plants Exposed to Fungicides. Journal of Botany 2012, 2012. [Google Scholar] [CrossRef]
- Petit, A.N.; Fontaine, F.; Vatsa, P.; Clément, C.; Vaillant-Gaveau, N. Fungicide impacts on photosynthesis in crop plants. Photosynth Res 2012, 111, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Kromann, P.; Taipe, A.; Perez, W.; Forbes, G. Rainfall Thresholds as Support for Timing Fungicide Applications in the Control of Potato Late Blight in Ecuador and Peru. Plant Disease - PLANT DIS 2009, 93, 142–148. [Google Scholar] [CrossRef]
- Vicent, A.; Armengol, J.; García-Jiménez, J. Rain Fastness and Persistence of Fungicides for Control of Alternaria Brown Spot of Citrus. Plant Disease - PLANT DIS 2007, 91, 393–399. [Google Scholar] [CrossRef]
- Garcia, P.; Rivero, R.; Ruiz, J.; Romero, L. The Role of Fungicides in the Physiology of Higher Plants: Implications for Defense Responses. The Botanical Review 2003, 69, 162–172. [Google Scholar] [CrossRef]
- Klittich, C.J.R. Fungicide Mobility and the Influence of Physical Properties. In Retention, Uptake, and Translocation of Agrochemicals in Plants; ACS Symposium Series; American Chemical Society: 2014; Volume 1171, pp. 95–109.
- Klittich, C.J.; Ray, S.L. Effects of physical properties on the translaminar activity of fungicides. Pestic Biochem Physiol 2013, 107, 351–359. [Google Scholar] [CrossRef]
- Warneke, B.; Thiessen, L.; Mahaffee, W. Effect of Fungicide Mobility and Application Timing on the Management of Grape Powdery Mildew. Plant Disease 2019, 104. [Google Scholar] [CrossRef]
- Ayesha, M.S.; Suryanarayanan, T.S.; Nataraja, K.N.; Prasad, S.R.; Shaanker, R.U. Seed Treatment With Systemic Fungicides: Time for Review. Front Plant Sci 2021, 12, 654512. [Google Scholar] [CrossRef]
- Deising, H.; Reimann, S.; Pascholati, S. Mechanisms and significance of fungicide resistance†. Brazilian journal of microbiology : [publication of the Brazilian Society for Microbiology] 2008, 39, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Mohandoss, J.; Suryanarayanan, T. Effect of fungicide treatment on foliar fungal endophyte diversity in mango. Sydowia 2009, 61, 11–24. [Google Scholar]
- Leyronas, C.; Mériaux, B.; Raynal, G. Chemical Control of Neotyphodium spp. Endophytes in Perennial Ryegrass and Tall Fescue Seeds. Crop Science 2006, 46, 98–104. [Google Scholar] [CrossRef]
- Kalia, A.; Gosal, S.K. Effect of pesticide application on soil microorganisms. Archives of Agronomy and Soil Science 2011, 57, 569–596. [Google Scholar] [CrossRef]
- Murphy, B.R.; Doohan, F.M.; Hodkinson, T.R. A seed dressing combining fungal endophyte spores and fungicides improves seedling survival and early growth in barley and oat. Symbiosis 2017, 71, 69–76. [Google Scholar] [CrossRef]
- Lloyd, A.W.; Percival, D.; Yurgel, S.N. Effect of Fungicide Application on Lowbush Blueberries Soil Microbiome. Microorganisms 2021, 9, 1366. [Google Scholar] [CrossRef]
- Lloyd, A.W.; Percival, D.; Langille, M.G.I.; Yurgel, S.N. Changes to Soil Microbiome Resulting from Synergetic Effects of Fungistatic Compounds Pyrimethanil and Fluopyram in Lowbush Blueberry Agriculture, with Nine Fungicide Products Tested. Microorganisms 2023, 11, 410. [Google Scholar] [CrossRef]
- Kahle, M.; Buerge, I.J.; Hauser, A.; Müller, M.D.; Poiger, T. Azole Fungicides: Occurrence and Fate in Wastewater and Surface Waters. Environmental Science & Technology 2008, 42, 7193–7200. [Google Scholar] [CrossRef]
- Zubrod, J.P.; Bundschuh, M.; Arts, G.; Brühl, C.A.; Imfeld, G.; Knäbel, A.; Payraudeau, S.; Rasmussen, J.J.; Rohr, J.; Scharmüller, A.; et al. Fungicides: An Overlooked Pesticide Class? Environ Sci Technol 2019, 53, 3347–3365. [Google Scholar] [CrossRef]
- Kiaune, L.; Singhasemanon, N. Pesticidal copper (I) oxide: environmental fate and aquatic toxicity. Rev Environ Contam Toxicol 2011, 213, 1–26. [Google Scholar] [CrossRef]
- Belsky, J.; Joshi, N.K. Effects of Fungicide and Herbicide Chemical Exposure on Apis and Non-Apis Bees in Agricultural Landscape. Frontiers in Environmental Science 2020, 8. [Google Scholar] [CrossRef]
- Weisenburger, D.D. Human health effects of agrichemical use. Hum Pathol 1993, 24, 571–576. [Google Scholar] [CrossRef]
- Habig, M.; Lorrain, C.; Feurtey, A.; Komluski, J.; Stukenbrock, E.H. Epigenetic modifications affect the rate of spontaneous mutations in a pathogenic fungus. Nature Communications 2021, 12, 5869. [Google Scholar] [CrossRef]
- Hermann, D.; Stenzel, K. FRAC Mode-of-action Classification and Resistance Risk of Fungicides. In Modern Crop Protection Compounds; 2019; pp. 589–608.
- Bent, A.F.; Mackey, D. Elicitors, effectors, and R genes: the new paradigm and a lifetime supply of questions. Annu Rev Phytopathol 2007, 45, 399–436. [Google Scholar] [CrossRef]
- Jones, J.D.; Vance, R.E.; Dangl, J.L. Intracellular innate immune surveillance devices in plants and animals. Science 2016, 354. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.-Q.; Xue, J.-Y.; Wu, P.; Zhang, Y.-M.; Wu, Y.; Hang, Y.-Y.; Wang, B.; Chen, J.-Q. Large-Scale Analyses of Angiosperm Nucleotide-Binding Site-Leucine-Rich Repeat Genes Reveal Three Anciently Diverged Classes with Distinct Evolutionary Patterns. Plant physiology 2016, 170, 2095–2109. [Google Scholar] [CrossRef] [PubMed]
- Shao, F.; Golstein, C.; Ade, J.; Stoutemyer, M.; Dixon, J.E.; Innes, R.W. Cleavage of Arabidopsis PBS1 by a bacterial type III effector. Science 2003, 301, 1230–1233. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-Microbe Interactions: Shaping the Evolution of the Plant Immune Response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef]
- Mur, L.A.J.; Kenton, P.; Lloyd, A.J.; Ougham, H.; Prats, E. The hypersensitive response; the centenary is upon us but how much do we know? Journal of Experimental Botany 2008, 59, 501–520. [Google Scholar] [CrossRef]
- Liu, P.-P.; Yang, Y.; Pichersky, E.; Klessig, D.F. Altering Expression of Benzoic Acid/Salicylic Acid Carboxyl Methyltransferase 1 Compromises Systemic Acquired Resistance and PAMP-Triggered Immunity in Arabidopsis. Molecular Plant-Microbe Interactions® 2010, 23, 82–90. [Google Scholar] [CrossRef]
- Tsuda, K.; Sato, M.; Stoddard, T.; Glazebrook, J.; Katagiri, F. Network properties of robust immunity in plants. PLoS Genet 2009, 5, e1000772. [Google Scholar] [CrossRef]
- Kaur, B.; Bhatia, D.; Mavi, G.S. Eighty years of gene-for-gene relationship and its applications in identification and utilization of R genes. J Genet 2021, 100. [Google Scholar] [CrossRef]
- Khajuria, Y.P.; Kaul, S.; Wani, A.A.; Dhar, M.K. Genetics of resistance in apple against Venturia inaequalis (Wint.) Cke. Tree Genetics & Genomes 2018, 14, 16. [Google Scholar] [CrossRef]
- Kou, Y.; Wang, S. Broad-spectrum and durability: understanding of quantitative disease resistance. Current Opinion in Plant Biology 2010, 13, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Ashfield, T.; Ong, L.E.; Nobuta, K.; Schneider, C.M.; Innes, R.W. Convergent evolution of disease resistance gene specificity in two flowering plant families. Plant Cell 2004, 16, 309–318. [Google Scholar] [CrossRef]
- Stahl, E.A.; Dwyer, G.; Mauricio, R.; Kreitman, M.; Bergelson, J. Dynamics of disease resistance polymorphism at the Rpm1 locus of Arabidopsis. Nature 1999, 400, 667–671. [Google Scholar] [CrossRef]
- Rimbaud, L.; Papaïx, J.; Barrett, L.G.; Burdon, J.J.; Thrall, P.H. Mosaics, mixtures, rotations or pyramiding: What is the optimal strategy to deploy major gene resistance? Evolutionary Applications 2018, 11, 1791–1810. [Google Scholar] [CrossRef]
- Mundt, C.C. Durable resistance: A key to sustainable management of pathogens and pests. Infection, Genetics and Evolution 2014, 27, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Pandolfi, V.; Neto, J.; da Silva, M.D.; Amorim, L.L.B.; Wanderley-Nogueira, A.C.; de Oliveira Silva, R.L.; Kido, E.A.; Crovella, S.; Iseppon, A.M.B. Resistance (R) Genes: Applications and Prospects for Plant Biotechnology and Breeding. Curr Protein Pept Sci 2017, 18, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Mundt, C. Pyramiding for Resistance Durability: Theory and Practice. Phytopathology 2018, 108. [Google Scholar] [CrossRef]
- Perez, W.; Salas, A.; Raymundo, R.; Huamán, Z.; Nelson, R.; Bonierbale, M. Evaluation of Wild Potato Species for Resistance to Late Blight. 2001.
- Zaidi, S.S.; Mukhtar, M.S.; Mansoor, S. Genome Editing: Targeting Susceptibility Genes for Plant Disease Resistance. Trends Biotechnol 2018, 36, 898–906. [Google Scholar] [CrossRef]
- van Schie, C.C.; Takken, F.L. Susceptibility genes 101: how to be a good host. Annu Rev Phytopathol 2014, 52, 551–581. [Google Scholar] [CrossRef]
- Kieu, N.P.; Lenman, M.; Wang, E.S.; Petersen, B.L.; Andreasson, E. Mutations introduced in susceptibility genes through CRISPR/Cas9 genome editing confer increased late blight resistance in potatoes. Sci Rep 2021, 11, 4487. [Google Scholar] [CrossRef]
- Zhou, H.; Bai, S.; Wang, N.; Sun, X.; Zhang, Y.; Zhu, J.; Dong, C. CRISPR/Cas9-Mediated Mutagenesis of MdCNGC2 in Apple Callus and VIGS-Mediated Silencing of MdCNGC2 in Fruits Improve Resistance to Botryosphaeria dothidea. Front Plant Sci 2020, 11, 575477. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Qiu, J.L. Genome editing for plant disease resistance: applications and perspectives. Philosophical Transactions of the Royal Society B 2019, 374. [Google Scholar] [CrossRef] [PubMed]
- Sticher, L.; Mauch-Mani, B.; Métraux, J.P. Systemic acquired resistance. Annu Rev Phytopathol 1997, 35, 235–270. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Dong, X. Systemic acquired resistance: turning local infection into global defense. Annu Rev Plant Biol 2013, 64, 839–863. [Google Scholar] [CrossRef]
- Bektas, Y.; Eulgem, T. Synthetic plant defense elicitors. Frontiers in plant science 2015, 5, 804–804. [Google Scholar] [CrossRef]
- Malamy, J.; Carr, J.P.; Klessig, D.F.; Raskin, I. Salicylic Acid: a likely endogenous signal in the resistance response of tobacco to viral infection. Science 1990, 250, 1002–1004. [Google Scholar] [CrossRef]
- Gaffney, T.; Friedrich, L.; Vernooij, B.; Negrotto, D.; Nye, G.; Uknes, S.; Ward, E.; Kessmann, H.; Ryals, J. Requirement of salicylic Acid for the induction of systemic acquired resistance. Science 1993, 261, 754–756. [Google Scholar] [CrossRef]
- Yu, Y.; Gui, Y.; Li, Z.; Jiang, C.; Guo, J.; Niu, D. Induced Systemic Resistance for Improving Plant Immunity by Beneficial Microbes. Plants (Basel) 2022, 11. [Google Scholar] [CrossRef]
- Thomma, B.P.; Eggermont, K.; Penninckx, I.A.; Mauch-Mani, B.; Vogelsang, R.; Cammue, B.P.; Broekaert, W.F. Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. Proc Natl Acad Sci U S A 1998, 95, 15107–15111. [Google Scholar] [CrossRef]
- Koornneef, A.; Leon-Reyes, A.; Ritsema, T.; Verhage, A.; Den Otter, F.C.; Van Loon, L.C.; Pieterse, C.M.J. Kinetics of salicylate-mediated suppression of jasmonate signaling reveal a role for redox modulation. Plant physiology 2008, 147, 1358–1368. [Google Scholar] [CrossRef]
- Höfte, M. Induced Resistance for Plant Defence. A Sustainable Approach to Crop Protection - Edited by Dale Walters, Adrian Newton and Gary Lyon. Plant Pathology 2007, 56, 1036–1037. [Google Scholar] [CrossRef]
- Wise, M.L. Plant Defense Activators: Application and Prospects in Cereal Crops. In 50 Years of Phytochemistry Research: Volume 43, Gang, D.R., Ed.; Springer International Publishing: Cham, 2013; pp. 55–70. [Google Scholar]
- Li, T.; Huang, Y.; Xu, Z.-S.; Wang, F.; Xiong, A.-S. Salicylic acid-induced differential resistance to the Tomato yellow leaf curl virus among resistant and susceptible tomato cultivars. BMC Plant Biology 2019, 19, 173. [Google Scholar] [CrossRef]
- Deenamo, N.; Kuyyogsuy, A.; Khompatara, K.; Chanwun, T.; Ekchaweng, K.; Churngchow, N. Salicylic Acid Induces Resistance in Rubber Tree against Phytophthora palmivora. Int J Mol Sci 2018, 19, 1883. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, X.; Li, B.; Zhang, Q.; Liang, W.; Wang, C. Salicylic acid confers enhanced resistance to Glomerella leaf spot in apple. Plant Physiol Biochem 2016, 106, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Sumayo, M.S.; Son, J.S.; Ghim, S.Y. Exogenous application of phenylacetic acid promotes root hair growth and induces the systemic resistance of tobacco against bacterial soft-rot pathogen Pectobacterium carotovorum subsp. carotovorum. Funct Plant Biol 2018, 45, 1119–1127. [Google Scholar] [CrossRef]
- Cao, J.; Zeng, K.; Jiang, W. Enhancement of Postharvest Disease Resistance in Ya Li Pear ( Pyrus bretschneideri ) Fruit by Salicylic Acid Sprays on the Trees during Fruit Growth. European Journal of Plant Pathology 2006, 114, 363–370. [Google Scholar] [CrossRef]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in Plant Protection. Marine Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Liu, H.; Jiang, D.; Wang, L.; Jiang, Y.; Tang, S.; Hou, X.; Han, X.; Liu, Z.; Zhang, M.; et al. Paecilomyces variotii extracts (ZNC) enhance plant immunity and promote plant growth. Plant and Soil 2019, 441, 383–397. [Google Scholar] [CrossRef]
- Faugeron-Girard, C.; Gloaguen, V.; Koçi, R.; Célérier, J.; Raynaud, A.; Moine, C. Use of a Pleurotus ostreatus Complex Cell Wall Extract as Elicitor of Plant Defenses: From Greenhouse to Field Trial. Molecules 2020, 25, 1094. [Google Scholar] [CrossRef] [PubMed]
- Margaritopoulou, T.; Toufexi, E.; Kizis, D.; Balayiannis, G.; Anagnostopoulos, C.; Theocharis, A.; Rempelos, L.; Troyanos, Y.; Leifert, C.; Markellou, E. Reynoutria sachalinensis extract elicits SA-dependent defense responses in courgette genotypes against powdery mildew caused by Podosphaera xanthii. Sci Rep 2020, 10, 3354. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, J.; Wan, A.; Rahman, M.; Punja, Z. Application of seaweed extract reduces foliar fungal diseases on carrot. Canadian Journal of Plant Pathology 2009, 31, 137–138. [Google Scholar]
- Esserti, S.; Smaili, A.; Rifai, L.A.; Koussa, T.; Makroum, K.; Belfaiza, M.; Kabil, E.M.; Faize, L.; Burgos, L.; Alburquerque, N.; et al. Protective effect of three brown seaweed extracts against fungal and bacterial diseases of tomato. Journal of Applied Phycology 2016, 29, 1081–1093. [Google Scholar] [CrossRef]
- Wang, Z.; Jia, C.; Li, J.; Huang, S.; Xu, B.; Jin, Z. Activation of salicylic acid metabolism and signal transduction can enhance resistance to Fusarium wilt in banana (Musa acuminata L. AAA group, cv. Cavendish). Funct Integr Genomics 2015, 15, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Heidel, A.J.; Clarke, J.D.; Antonovics, J.; Dong, X. Fitness costs of mutations affecting the systemic acquired resistance pathway in Arabidopsis thaliana. Genetics 2004, 168, 2197–2206. [Google Scholar] [CrossRef] [PubMed]
- Heil, M. The Ecological Concept of Costs of Induced Systemic Resistance (ISR). European Journal of Plant Pathology 2001, 107, 137–146. [Google Scholar] [CrossRef]
- Ngullie, C.R.; Tank, R.; Bhanderi, D. Effect of salicylic acid and humic acid on flowering, fruiting, yield and quality of mango (Mangifera indica L.) cv. KESAR. ADVANCE RESEARCH JOURNAL OF CROP IMPROVEMENT 2014, 5, 136–139. [Google Scholar] [CrossRef]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulant Properties of Seaweed Extracts in Plants: Implications towards Sustainable Crop Production. Plants 2021, 10, 531. [Google Scholar] [CrossRef]
- LaMondia, J.A. Actigard Increases Fungicide Efficacy Against Tobacco Blue Mold. Plant Disease 2008, 92, 1463–1467. [Google Scholar] [CrossRef]
- Bosamia, T.C.; Barbadikar, K.M.; Modi, A. 9 - Genomic insights of plant endophyte interaction: prospective and impact on plant fitness. In Microbial Endophytes, Kumar, A., E.K, R., Eds.; Woodhead Publishing: 2020; pp. 227–249.
- Busby, P.E.; Ridout, M.; Newcombe, G. Fungal endophytes: modifiers of plant disease. Plant Mol Biol 2016, 90, 645–655. [Google Scholar] [CrossRef]
- Backman, P.; Sikora, R. Endophytes: An emerging tool for biological control. Biological Control - BIOL CONTROL 2008, 46, 1–3. [Google Scholar] [CrossRef]
- Rodriguez, R.J.; White Jr, J.F.; Arnold, A.E.; Redman, R.S. Fungal endophytes: diversity and functional roles. New Phytologist 2009, 182, 314–330. [Google Scholar] [CrossRef]
- Redman, R.S.; Freeman, S.; Clifton, D.R.; Morrel, J.; Brown, G.; Rodriguez, R.J. Biochemical Analysis of Plant Protection Afforded by a Nonpathogenic Endophytic Mutant of Colletotrichum magna1. Plant Physiology 1999, 119, 795–804. [Google Scholar] [CrossRef]
- Gunatilaka, A.A. Natural products from plant-associated microorganisms: distribution, structural diversity, bioactivity, and implications of their occurrence. J Nat Prod 2006, 69, 509–526. [Google Scholar] [CrossRef] [PubMed]
- Kusari, S.; Hertweck, C.; Spiteller, M. Chemical Ecology of Endophytic Fungi: Origins of Secondary Metabolites. Chemistry & Biology 2012, 19, 792–798. [Google Scholar] [CrossRef]
- Ludwig-Müller, J. Plants and endophytes: equal partners in secondary metabolite production? Biotechnology letters 2015, 37, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-L.; Sun, S.-Z.; Miao, C.-P.; Wu, K.; Chen, Y.-W.; Xu, L.-H.; Guan, H.-L.; Zhao, L.-X. Endophytic Trichoderma gamsii YIM PH30019: a promising biocontrol agent with hyperosmolar, mycoparasitism, and antagonistic activities of induced volatile organic compounds on root-rot pathogenic fungi of Panax notoginseng. Journal of Ginseng Research 2016, 40, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Qualhato, T.F.; Lopes, F.A.; Steindorff, A.S.; Brandão, R.S.; Jesuino, R.S.; Ulhoa, C.J. Mycoparasitism studies of Trichoderma species against three phytopathogenic fungi: evaluation of antagonism and hydrolytic enzyme production. Biotechnol Lett 2013, 35, 1461–1468. [Google Scholar] [CrossRef]
- Quimby, P.C.; King, L.R.; Grey, W.E. Biological control as a means of enhancing the sustainability of crop/land management systems. Agriculture, Ecosystems & Environment 2002, 88, 147–152. [Google Scholar] [CrossRef]
- Griffin, M.R. Biocontrol and bioremediation: two areas of endophytic research which hold great promise. Advances in Endophytic Research 2014, 257–282. [Google Scholar]
- Acosta-González, U.; Silva-Rojas, H.V.; Fuentes-Aragón, D.; Hernández-Castrejón, J.; Romero-Bautista, A.; Rebollar-Alviter, A. Comparative Performance of Fungicides and Biocontrol Products in the Management of Fusarium Wilt of Blackberry. Plant Disease 2022, 106, 1419–1427. [Google Scholar] [CrossRef]
- Anees, M.; Tronsmo, A.; Edel-Hermann, V.; Hjeljord, L.G.; Héraud, C.; Steinberg, C. Characterization of field isolates of Trichoderma antagonistic against Rhizoctonia solani. Fungal Biol 2010, 114, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Ahamad, L.; Zaidi, R.K. Seed Priming Through Fungicides, Biocontrol Agents and Botanicals as Effective Method for Controlling Spot Blotch Pathogen, Bipolaris sorokiniana in Barley. Gesunde Pflanzen 2022, 1–10. [Google Scholar] [CrossRef]
- Hermosa, R.; Cardoza, R.E.; Rubio, M.B.; Gutiérrez, S.; Monte, E. Secondary metabolism and antimicrobial metabolites of Trichoderma. In Biotechnology and biology of Trichoderma; Elsevier: 2014; pp. 125–137.
- Mukhopadhyay, R.; Kumar, D. Trichoderma: a beneficial antifungal agent and insights into its mechanism of biocontrol potential. Egyptian Journal of Biological Pest Control 2020, 30, 133. [Google Scholar] [CrossRef]
- Iqbal, M.; Jützeler, M.; França, S.C.; Wäckers, F.; Andreasson, E.; Stenberg, J.A. Bee-Vectored Aureobasidium pullulans for Biological Control of Gray Mold in Strawberry. Phytopathology® 2022, 112, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Pinto, C.; Custódio, V.; Nunes, M.; Songy, A.; Rabenoelina, F.; Courteaux, B.; Clément, C.; Gomes, A.C.; Fontaine, F. Understand the potential role of Aureobasidium pullulans, a resident microorganism from grapevine, to prevent the infection caused by Diplodia seriata. Frontiers in microbiology 2018, 9, 3047. [Google Scholar] [CrossRef]
- Bencheqroun, S.; Bajji, M.; Sebastien, M.; Bentata, F.; Labhilili, M.; el hassan, A.; el Jaafari, S.; Jijakli, M. Biocontrol of blue mold on apple fruits by Aureobasidium pullulans (strain Ach 1-1): in vitro and in situ evidence for the possible involvement of competition for nutrients. Communications in agricultural and applied biological sciences 2006, 71, 1151–1157. [Google Scholar]
- Schena, L.; Nigro, F.; Pentimone, I.; Ligorio, A.; Ippolito, A. Control of postharvest rots of sweet cherries and table grapes with endophytic isolates of Aureobasidium pullulans. Postharvest Biology and Technology 2003, 30, 209–220. [Google Scholar] [CrossRef]
- Agirman, B.; Erten, H. Biocontrol ability and action mechanisms of Aureobasidium pullulans GE17 and Meyerozyma guilliermondii KL3 against Penicillium digitatum DSM2750 and Penicillium expansum DSM62841 causing postharvest diseases. Yeast 2020, 37, 437–448. [Google Scholar] [CrossRef]
- Di francesco, A.; Ugolini, L.; Lazzeri, L.; Mari, M. Production of volatile organic compounds by Aureobasidium pullulans as a potential mechanism of action against postharvest fruit pathogens. Biological Control 2015, 81. [Google Scholar] [CrossRef]
- Bozoudi, D.; Tsaltas, D. The Multiple and Versatile Roles of Aureobasidium pullulans in the Vitivinicultural Sector. Fermentation 2018, 4, 85. [Google Scholar] [CrossRef]
- Pujol, M.; Badosa, E.; Cabrefiga, J.; Montesinos, E. Development of a strain-specific quantitative method for monitoring Pseudomonas fluorescens EPS62e, a novel biocontrol agent of fire blight. FEMS Microbiology Letters 2005, 249, 343–352. [Google Scholar] [CrossRef]
- Santoyo, G.; Orozco-Mosqueda, M.d.C.; Govindappa, M. Mechanisms of biocontrol and plant growth-promoting activity in soil bacterial species of Bacillus and Pseudomonas: a review. Biocontrol Science and Technology 2012, 22, 855–872. [Google Scholar] [CrossRef]
- Vicedo, B.; Peñalver, R.; Asins, M.J.; López, M.M. Biological Control of Agrobacterium tumefaciens, Colonization, and pAgK84 Transfer with Agrobacterium radiobacter K84 and the Tra Mutant Strain K1026. Appl Environ Microbiol 1993, 59, 309–315. [Google Scholar] [CrossRef]
- Collinge, D.B.; Jensen, D.F.; Rabiey, M.; Sarrocco, S.; Shaw, M.W.; Shaw, R.H. Biological control of plant diseases – What has been achieved and what is the direction? Plant Pathology 2022, 71, 1024–1047. [Google Scholar] [CrossRef]
- Bardin, M.; Ajouz, S.; Comby, M.; Lopez-Ferber, M.; Graillot, B.; Siegwart, M.; Nicot, P.C. Is the efficacy of biological control against plant diseases likely to be more durable than that of chemical pesticides? Front Plant Sci 2015, 6, 566. [Google Scholar] [CrossRef]
- Bardin, M.; Ajouz, S.; Comby, M.; Lopez-Ferber, M.; Graillot, B.; Siegwart, M.; Nicot, P.C. Is the efficacy of biological control against plant diseases likely to be more durable than that of chemical pesticides? Frontiers in Plant Science 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Guetsky, R.; Shtienberg, D.; Elad, Y.; Dinoor, A. Combining biocontrol agents to reduce the variability of biological control. Phytopathology 2001, 91, 621–627. [Google Scholar] [CrossRef]
- García-Pedrajas, M.D.; Cañizares, M.C.; Sarmiento-Villamil, J.L.; Jacquat, A.G.; Dambolena, J.S. Mycoviruses in Biological Control: From Basic Research to Field Implementation. Phytopathology® 2019, 109, 1828–1839. [Google Scholar] [CrossRef] [PubMed]
- Chagnon, P.-L. Ecological and evolutionary implications of hyphal anastomosis in arbuscular mycorrhizal fungi. FEMS microbiology ecology 2014, 88, 437–444. [Google Scholar] [CrossRef]
- Xie, J.; Jiang, D. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annual Review of Phytopathology 2014, 52, 45–68. [Google Scholar] [CrossRef]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Molecular Plant Pathology 2009, 10, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Bian, R.; Liu, Q.; Yang, L.; Pang, T.; Salaipeth, L.; Andika, I.B.; Kondo, H.; Sun, L. Identification of a Novel Hypovirulence-Inducing Hypovirus From Alternaria alternata. Frontiers in microbiology 2019, 10, 1076–1076. [Google Scholar] [CrossRef]
- Kamaruzzaman, M.; He, G.; Wu, M.; Zhang, J.; Yang, L.; Chen, W.; Li, G. A Novel Partitivirus in the Hypovirulent Isolate QT5-19 of the Plant Pathogenic Fungus Botrytis cinerea. Viruses 2019, 11, 24. [Google Scholar] [CrossRef]
- Buttimer, C.; McAuliffe, O.; Ross, R.P.; Hill, C.; O'Mahony, J.; Coffey, A. Bacteriophages and Bacterial Plant Diseases. Front Microbiol 2017, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Born, Y.; Fieseler, L.; Klumpp, J.; Eugster, M.R.; Zurfluh, K.; Duffy, B.; Loessner, M.J. The tail-associated depolymerase of Erwinia amylovora phage L1 mediates host cell adsorption and enzymatic capsule removal, which can enhance infection by other phage. Environ Microbiol 2014, 16, 2168–2180. [Google Scholar] [CrossRef]
- Das, M.; Bhowmick, T.S.; Ahern, S.J.; Young, R.; Gonzalez, C.F. Control of Pierce's Disease by Phage. PLOS ONE 2015, 10, e0128902. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Gomes, A.C.; da Silva, R.R.; Moreira, S.I.; Vicentini, S.N.C.; Ceresini, P.C. Biofungicides: An Eco-Friendly Approach for Plant Disease Management. In Encyclopedia of Mycology, Zaragoza, Ó., Casadevall, A., Eds.; Elsevier: Oxford, 2021; pp. 641–649. [Google Scholar]
- Trejo-Raya, A.B.; Rodríguez-Romero, V.M.; Bautista-Baños, S.; Quiroz-Figueroa, F.R.; Villanueva-Arce, R.; Durán-Páramo, E. Effective in vitro control of two phytopathogens of agricultural interest using cell-free extracts of pseudomonas fluorescens and chitosan. Molecules 2021, 26, 6359. [Google Scholar] [CrossRef]
- Kaur, T.; Kaur, A.; Sharma, V.; Manhas, R.K. Purification and Characterization of a New Antifungal Compound 10-(2, 2-dimethyl-cyclohexyl)-6, 9-dihydroxy-4, 9-dimethyl-dec-2-enoic Acid Methyl Ester from Streptomyces hydrogenans Strain DH16. Frontiers in microbiology 2016, 7, 1004. [Google Scholar] [CrossRef] [PubMed]
- Polo, K.J.J.; Campos, H.L.M.; Olivera, C.C.; Nakayo, J.L.J.; Flores, J.W.V. Biofungicide for the Control of Botrytis Cinerea and Fusarium Oxysporum: a Laboratory Study. Chemical Engineering Transactions 2021, 87, 517–522. [Google Scholar]
- Wulff, E.; Zida, E.; Torp, J.; Lund, O. Yucca schidigera extract: a potential biofungicide against seedborne pathogens of sorghum. Plant Pathology 2012, 61, 331–338. [Google Scholar] [CrossRef]
- Baraka, M.; Fatma; Shaban, M. ; H, A. Efficiency of Some Plant Extracts, Natural Oils, Biofungicides and Fungicides Against Root Rot of Date Palm. J. Biol. Chem. Environ. Sci. 2011, 6, 405–429. [Google Scholar]
- Lu, M.; Han, Z.; Yao, L. In vitro and in vivo antimicrobial efficacy of essential oils and individual compounds against Phytophthora parasitica var. nicotianae. Journal of Applied Microbiology 2013, 115, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Géza, N.; Hochbaum, T.; Sarosi, S.; Ladanyi, M. In vitro and in planta activity of some essential oils against Venturia inaequalis (Cooke) G. Winter. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 2014, 42, 109–114. [Google Scholar]
- Sharma, M.; Tarafdar, A.; Ghosh, R.; Gopalakrishanan, S. Biological Control as a Tool for Eco-friendly Management of Plant Pathogens. 2017; pp. 153–188.
- Al-Samarrai, G.; Singh, H.; Syarhabil, M. Evaluating eco-friendly botanicals (natural plant extracts) as alternatives to synthetic fungicides. Annals of Agricultural and Environmental Medicine 2012, 19. [Google Scholar]
- Verdel, A.; Jia, S.; Gerber, S.; Sugiyama, T.; Gygi, S.; Grewal, S.I.; Moazed, D. RNAi-mediated targeting of heterochromatin by the RITS complex. Science 2004, 303, 672–676. [Google Scholar] [CrossRef]
- Pratt, A.J.; MacRae, I.J. The RNA-induced silencing complex: a versatile gene-silencing machine. J Biol Chem 2009, 284, 17897–17901. [Google Scholar] [CrossRef]
- Tang, Y.; Yan, X.; Gu, C.; Yuan, X. Biogenesis, Trafficking, and Function of Small RNAs in Plants. Front Plant Sci 2022, 13, 825477. [Google Scholar] [CrossRef]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Yi, R.; Cullen, B.R. MicroRNAs and small interfering RNAs can inhibit mRNA expression by similar mechanisms. Proc Natl Acad Sci U S A 2003, 100, 9779–9784. [Google Scholar] [CrossRef]
- López-Dolz, L.; Spada, M.; Daròs, J.A.; Carbonell, A. Fine-tune control of targeted RNAi efficacy by plant artificial small RNAs. Nucleic Acids Res 2020, 48, 6234–6250. [Google Scholar] [CrossRef]
- Dugas, D.V.; Bartel, B. Sucrose induction of Arabidopsis miR398 represses two Cu/Zn superoxide dismutases. Plant Mol Biol 2008, 67, 403–417. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wang, F.; Axtell, M.J. Analysis of complementarity requirements for plant microRNA targeting using a Nicotiana benthamiana quantitative transient assay. Plant Cell 2014, 26, 741–753. [Google Scholar] [CrossRef] [PubMed]
- Obbard, D.J.; Gordon, K.H.; Buck, A.H.; Jiggins, F.M. The evolution of RNAi as a defence against viruses and transposable elements. Philos Trans R Soc Lond B Biol Sci 2009, 364, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Harvey, J.J.W.; Lewsey, M.G.; Patel, K.; Westwood, J.; Heimstädt, S.; Carr, J.P.; Baulcombe, D.C. An Antiviral Defense Role of AGO2 in Plants. PLOS ONE 2011, 6, e14639. [Google Scholar] [CrossRef]
- Koch, A.; Biedenkopf, D.; Furch, A.; Weber, L.; Rossbach, O.; Abdellatef, E.; Linicus, L.; Johannsmeier, J.; Jelonek, L.; Goesmann, A.; et al. An RNAi-Based Control of Fusarium graminearum Infections Through Spraying of Long dsRNAs Involves a Plant Passage and Is Controlled by the Fungal Silencing Machinery. PLoS Pathog 2016, 12, e1005901. [Google Scholar] [CrossRef]
- Nunes, C.C.; Dean, R.A. Host-induced gene silencing: a tool for understanding fungal host interaction and for developing novel disease control strategies. Mol Plant Pathol 2012, 13, 519–529. [Google Scholar] [CrossRef]
- Williamson, V.M.; Kumar, A. Nematode resistance in plants: the battle underground. Trends Genet 2006, 22, 396–403. [Google Scholar] [CrossRef]
- Huang, G.; Allen, R.; Davis, E.L.; Baum, T.J.; Hussey, R.S. Engineering broad root-knot resistance in transgenic plants by RNAi silencing of a conserved and essential root-knot nematode parasitism gene. Proceedings of the National Academy of Sciences 2006, 103, 14302–14306. [Google Scholar] [CrossRef]
- Nowara, D.; Gay, A.; Lacomme, C.; Shaw, J.; Ridout, C.; Douchkov, D.; Hensel, G.; Kumlehn, J.; Schweizer, P. HIGS: Host-Induced Gene Silencing in the Obligate Biotrophic Fungal Pathogen Blumeria graminis Plant Cell 2010, 22, 3130–3141. [CrossRef]
- Song, Y.; Thomma, B. Host-induced gene silencing compromises Verticillium wilt in tomato and Arabidopsis. Mol Plant Pathol 2018, 19, 77–89. [Google Scholar] [CrossRef]
- Guo, X.Y.; Li, Y.; Fan, J.; Xiong, H.; Xu, F.X.; Shi, J.; Shi, Y.; Zhao, J.Q.; Wang, Y.F.; Cao, X.L.; et al. Host-Induced Gene Silencing of MoAP1 Confers Broad-Spectrum Resistance to Magnaporthe oryzae. Front Plant Sci 2019, 10, 433. [Google Scholar] [CrossRef]
- Wang, M.; Weiberg, A.; Lin, F.M.; Thomma, B.P.; Huang, H.D.; Jin, H. Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat Plants 2016, 2, 16151. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, A.G.; Wytinck, N.; Walker, P.L.; Girard, I.J.; Rashid, K.Y.; de Kievit, T.; Fernando, W.G.D.; Whyard, S.; Belmonte, M.F. Identification and application of exogenous dsRNA confers plant protection against Sclerotinia sclerotiorum and Botrytis cinerea. Sci Rep 2018, 8, 7320. [Google Scholar] [CrossRef] [PubMed]
- Biedenkopf, D.; Will, T.; Knauer, T.; Jelonek, L.; Furch, A.C.U.; Busche, T.; Koch, A. Systemic spreading of exogenous applied RNA biopesticides in the crop plant Hordeum vulgare. ExRNA 2020, 2, 12. [Google Scholar] [CrossRef]
- Höfle, L.; Biedenkopf, D.; Werner, B.T.; Shrestha, A.; Jelonek, L.; Koch, A. Study on the efficiency of dsRNAs with increasing length in RNA-based silencing of the Fusarium CYP51 genes. RNA Biol 2020, 17, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Li, P.; Jain, R.G.; Taochy, C.; Fletcher, S.J.; Carroll, B.J.; Lu, G.Q.; Xu, Z.P. Clay nanosheets for topical delivery of RNAi for sustained protection against plant viruses. Nat Plants 2017, 3, 16207. [Google Scholar] [CrossRef]
- Zhang, K.; Wei, J.; Huff Hartz, K.E.; Lydy, M.J.; Moon, T.S.; Sander, M.; Parker, K.M. Analysis of RNA Interference (RNAi) Biopesticides: Double-Stranded RNA (dsRNA) Extraction from Agricultural Soils and Quantification by RT-qPCR. Environ Sci Technol 2020, 54, 4893–4902. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Wisniewski, M.; Droby, S.; Schena, L. Spatial and compositional variation in the fungal communities of organic and conventionally grown apple fruit at the consumer point-of-purchase. Hortic Res 2016, 3, 16047–16047. [Google Scholar] [CrossRef]
- Jackson, A.L.; Bartz, S.R.; Schelter, J.; Kobayashi, S.V.; Burchard, J.; Mao, M.; Li, B.; Cavet, G.; Linsley, P.S. Expression profiling reveals off-target gene regulation by RNAi. Nat Biotechnol 2003, 21, 635–637. [Google Scholar] [CrossRef]
- Neumeier, J.; Meister, G. siRNA Specificity: RNAi Mechanisms and Strategies to Reduce Off-Target Effects. Front Plant Sci 2020, 11, 526455. [Google Scholar] [CrossRef]
- Percival, G.; Noviss, K.; Haynes, I. Field evaluation of systemic inducing resistance chemicals at different growth stages for the control of apple (Venturia inaequalis) and pear (Venturia pirina) scab. Crop Protection 2009, 28, 629–633. [Google Scholar] [CrossRef]
- Zaker, M. Natural Plant Products as Eco-friendly Fungicides for Plant Diseases Control- A Review. The Agriculturists 2016, 14, 134. [Google Scholar] [CrossRef]
- Balba, H. Review of strobilurin fungicide chemicals. Journal of environmental science and health. Part. B, Pesticides, food contaminants, and agricultural wastes 2007, 42, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential Oils as Potential Alternative Biocontrol Products against Plant Pathogens and Weeds: A Review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [PubMed]
- López, M.D.; Cantó-Tejero, M.; Pascual-Villalobos, M.J. New Insights into biopesticides: Solid and liquid formulations of essential oils and derivatives. Frontiers in Agronomy 2021, 3, 763530. [Google Scholar] [CrossRef]
- Llorente, I.; Vilardell, P.; Bugiani, R.; Gherardi, I.; Montesinos, E. Evaluation of BSPcast Disease Warning System in Reduced Fungicide Use Programs for Management of Brown Spot of Pear. Plant Disease 2000, 84, 631–637. [Google Scholar] [CrossRef]
- Holb, I.J. Timing of first and final sprays against apple scab combined with leaf removal and pruning in organic apple production. Crop Protection 2008, 27, 814–822. [Google Scholar] [CrossRef]
- de Kraker, J.; van den Ende, J.E.; Rossing, W.A.H. Control strategies with reduced fungicide input for Botrytis leaf blight in lily—a simulation analysis. Crop Protection 2005, 24, 157–165. [Google Scholar] [CrossRef]
- Mertely, J.C.; MacKenzie, S.J.; Legard, D.E. Timing of Fungicide Applications for Botrytis cinerea Based on Development Stage of Strawberry Flowers and Fruit. Plant Disease 2002, 86, 1019–1024. [Google Scholar] [CrossRef]
- Madden, L.; Pennypacker, S.; MacNab, A. FAST, a forecast system for Alternaria solani on tomato. Phytopathology 1978, 68, 1354–1358. [Google Scholar] [CrossRef]
- Dhar, N.; Mamo, B.E.; Subbarao, K.V.; Koike, S.T.; Fox, A.; Anchieta, A.; Klosterman, S.J. Measurements of Aerial Spore Load by qPCR Facilitates Lettuce Downy Mildew Risk Advisement. Plant Disease 2020, 104, 82–93. [Google Scholar] [CrossRef]
- Thiessen, L.D.; Keune, J.A.; Neill, T.M.; Turechek, W.W.; Grove, G.G.; Mahaffee, W.F. Development of a grower-conducted inoculum detection assay for management of grape powdery mildew. Plant Pathology 2016, 65, 238–249. [Google Scholar] [CrossRef]
- Everett, K.R.; Pushparajah, I.P.S.; Timudo, O.E.; Ah Chee, A.; Scheper, R.W.A.; Shaw, P.W.; Spiers, T.M.; Taylor, J.T.; Wallis, D.R.; Wood, P.N. Infection criteria, inoculum sources and splash dispersal pattern of Colletotrichum acutatum causing bitter rot of apple in New Zealand. European Journal of Plant Pathology 2018, 152, 367–383. [Google Scholar] [CrossRef]
- Ons, L.; Bylemans, D.; Thevissen, K.; Cammue, B.P.A. Combining Biocontrol Agents with Chemical Fungicides for Integrated Plant Fungal Disease Control. Microorganisms 2020, 8, 1930. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Methods for controlling fungal plant pathogens. The potential advantages (green) and disadvantages (red) inherent in conventional and alternative methods of disease control are also shown.
Figure 1.
Methods for controlling fungal plant pathogens. The potential advantages (green) and disadvantages (red) inherent in conventional and alternative methods of disease control are also shown.
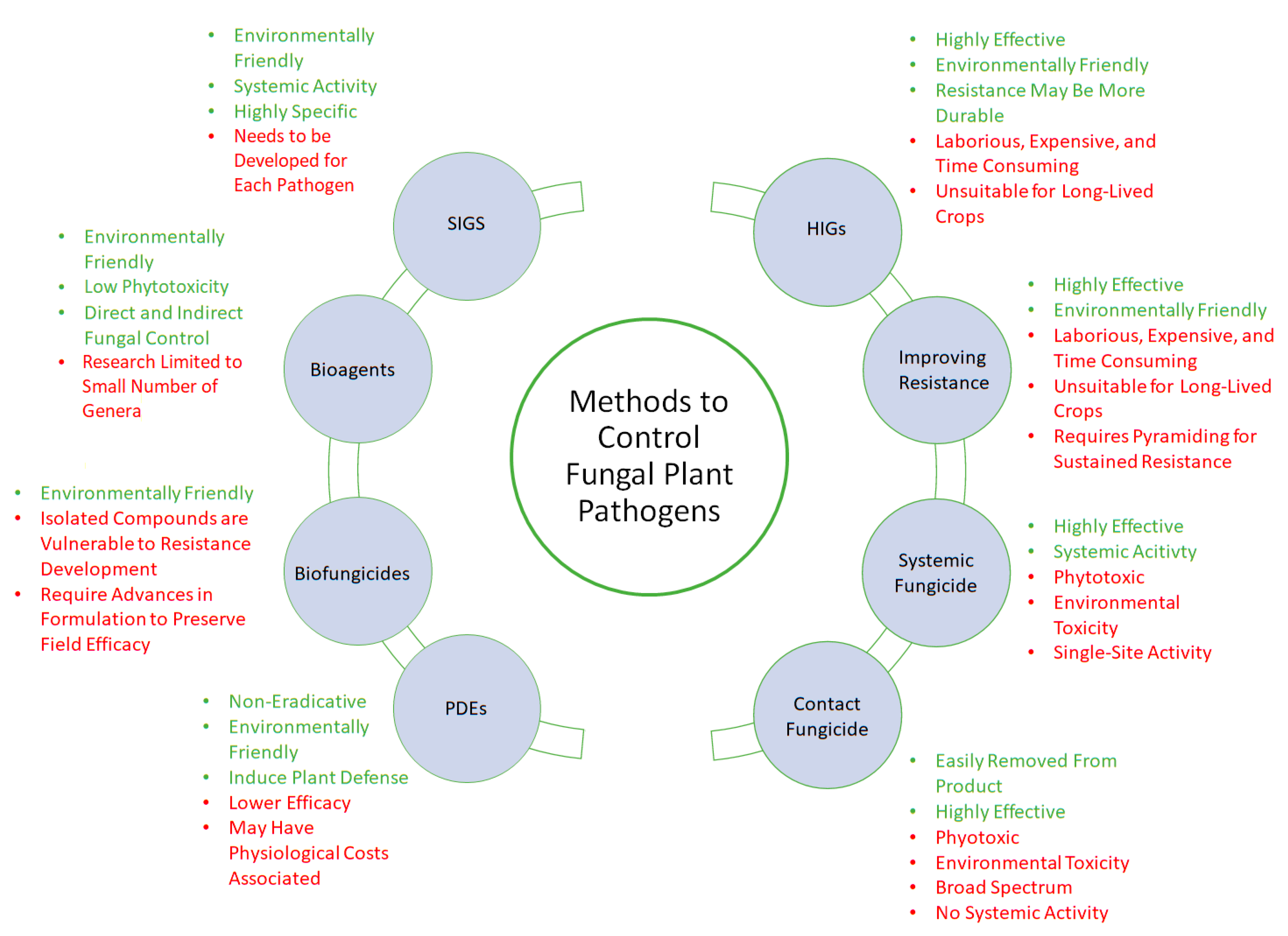
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



