
Submitted:
21 September 2023
Posted:
22 September 2023
You are already at the latest version
Abstract
Aging is a complex process associated with tissue degeneration and an increased risk of age-related diseases. This study aimed to evaluate the impact of Terasen®, a nutraceutical containing standardized extracts of Euterpe oleracea, Myrciaria dubia, and purified oil of Bixa orellana and Astrocaryum aculeatum on the lifespan of Caenorhabditis elegans, a widely used model organism for aging research. The findings demonstrated that Terasen® exhibited significant antioxidant activity and influenced the feeding behavior of C. elegans, leading to a reduced pharyngeal pumping rate and a decreased number of offspring produced by treated individuals. Notably, Terasen® also displayed a remarkable ability to extend the lifespan of C. elegans. These findings suggest that Terasen® may possess promising anti-aging effects in vivo, warranting further investigation.
Keywords:
longevity
; healthspan
; lifespan
; nutraceuticals
; C. elegans
1. Introduction
Aging is a natural process during an individual’s lifetime that leads to a decline in natural body functions [1,2]. The functional changes associated with aging have been identified as risk factors for various health problems, including hypertension, osteoporosis, diabetes, cataracts, heart failure, and neurodegenerative diseases [3].
According to the World Health Organization (WHO), global life expectancy is increasing, and it is projected that the elderly population will reach 1.4 billion people by 2030 and exceed 2.1 billion by 2050 [4]. With this demographic shift, it is imperative to seek solutions that address the detrimental effects of aging, ensuring that individuals can maintain their independence and experience healthy aging [2,5].
Pharmacological interventions aimed at prolonging human longevity and preventing age-related diseases are under investigation. The nematode Caenorhabditis elegans has emerged as a promising model for studying the determinants of longevity. C. elegans has a short lifespan, compact size, and conserved genetic pathways that regulate aging, such as insulin signaling, oxidative stress response, and longevity-related genes [6].
In this study, we employed the C. elegans model to investigate the effects of Terasen®, nutraceutical formulation composed of standardized extracts of Euterpe oleracea, Myrciaria dubia, and purified oil of Bixa orellana and Astrocaryum aculeatum. The phytochemical markers of Terasen® include anthocyanins, quercetin, ellagic acid, gallic acid, carotenoids, geranylgeraniol, and tocotrienols, which have been extensively studied for their pharmacological properties and their potential impact on longevity.
2. Materials and Methods
2.1. Product
The nutraceutical product in the form of encapsulated granules, Terasen®, was provided by Ages Bioactive Compounds (São Paulo, SP, Brazil), batch URU201101.
2.2. Evaluation of Antioxidant Activity
The Terasen® sample was extracted using DMSO and vortex agitation for the antioxidant activity analysis. At the end of the procedure, all samples were prepared at a final concentration of 1 mg/ml. Gallic acid and DMSO were used as the standard and negative control, respectively. The results were expressed as the percentage of radical scavenging using the following equation: Inhibition (%) = [(Ac - As) / Ac] × 100, where Ac is the absorbance of the negative control (DMSO) and as is the absorbance of the sample. All analyses were performed in triplicates.
2.2.1. DPPH Radical Scavenging Activity
The method described by [7], with some modifications, was used to determine antioxidant activity using DPPH. The DPPH solution (2,2-diphenyl-1-picrylhydrazyl) was prepared by dissolving 1 mg of DPPH in 12 ml of absolute ethanol. Then, 270 µL of this solution and 30 µL of ethanol were added to the microplate, adjusting the solution with ethanol to achieve an absorbance of 1.00 ± 0.1 nm. For the test, 30 µL of the sample and 270 µL of the DPPH solution were added to the plate, and it was incubated in the dark for 30 minutes. After this time, the absorbance was measured at 517 nm using an ELISA reader (Kasuaki, Dr-200Bn-Bi).
2.2.2. ABTS Radical Scavenging Assay
The antioxidant activity was evaluated using the ABTS method described by [8] with some modifications. The ABTS solution (7 mM) (2,2’-azinobis(3-ethylbenzothiazoline-6-sulfonic acid)) was mixed with a potassium persulfate solution (2.45 mM) and incubated at room temperature in the dark for 16 hours. Then, 270 µL of the ABTS solution was mixed with 30 µL of the sample. The plate was incubated in the dark for 30 minutes, and the absorbance was measured at 630 nm using an ELISA reader (Kasuaki, Dr-200Bn-Bi).
2.3. Caenorhabditis Elegans Strains
The N2 Bristol (wild-type) strains of Caenorhabditis elegans used in this study were obtained from the Department of Parasitology/ICB/USP (ICB, University of São Paulo, SP). All nematodes were incubated at 20 °C and grown on NGM (nematode growth medium) seeded with Escherichia coli OP50 as a food source.
2.3.1. Reproduction Assay
The reproduction assay was conducted following the method described by [9]. Synchronized L4 nematodes (n=5) were daily transferred until the end of the reproductive period to NGM plates with only E. coli OP50 in LB medium (negative control) or E. coli in LB medium plus different concentrations of Terasen® solubilized in the medium (250, 500, or 1000 µg/mL), and the eggs were counted. This assay was performed in triplicate, and the results were presented as the mean number of offspring.
2.3.2. Pharyngeal Pumping Rate
The pharyngeal pumping rate was evaluated using the methodology proposed by [10]. Synchronized L4 nematodes were raised on NGM plates and treated with only E. coli OP50 in LB medium (negative control) or E. coli in LB medium plus different concentrations of Terasen® solubilized in the medium (250, 500, or 1000 µg/mL). On the 3rd, 6th, and 9th day of adulthood, 10 worms were randomly selected, and the number of pharyngeal contractions during a 60-second interval was quantified. The experiment was performed in triplicate.
2.3.3. Growth Alteration Assay
The growth alteration assay was performed following the method described by [11]. The animals were raised from the L1 stage on NGM plates receiving only E. coli OP50 in LB medium (negative control) or E. coli in LB medium plus different concentrations of Terasen® solubilized in the medium (250, 500, or 1000 µg/mL). On the 4th and 8th day of adulthood, the animals were photographed using a stereomicroscope and camera (Luxeo 4D, Labomed, CA, USA), and their body length was measured from head to tail using ImageJ software (v1.53u, Massachusetts, USA). The experiment was performed in triplicate.
2.3.4. Locomotion Analysis Assay
The locomotion analysis assay followed the protocol described by [12]. Initially, synchronized N2 nematodes were transferred to NGM plates and treated only E. coli OP50 in LB medium (negative control) or E. coli in LB medium plus different concentrations of Terasen® solubilized in the medium (250, 500, or 1000 µg/mL). After 4 and 8 days of treatment, each nematode was placed on a glass dish with 100 µL of M9 buffer. After 1 minute of recovery, the total number of body bends was counted over 20 seconds using a stereomicroscope.
2.3.5. Lifespan Assessment
The lifespan of nematodes was assessed following the method of [10]. Synchronized L4 stage nematodes (n=20) were transferred to NGM plates and treated with only E. coli OP50 in LB medium (negative control) or E. coli in LB medium plus different concentrations of Terasen® solubilized in the medium (250, 500, or 1000 µg/mL). The nematodes were transferred daily to fresh NGM plates with their respective treatments for the first six days, followed by transfers every two days afterward. Daily worm counts were conducted until the death of all nematodes, with worms unresponsive to a gentle touch with a platinum wire being marked as dead and excluded from the plates. The results were reported as the percentage of survival, mean lifespan, and median lifespan. The lifespan tests were conducted in triplicate at a constant temperature of 20°C.
2.4. Statistical Analysis
Statistical analysis was conducted using GraphPad Prism software (version 5.03). The results were presented as mean ± SEM. Two-Way ANOVA was used to compare data (factors: treatment; days), followed by multiple Tukey’s posthoc tests in case of statistical difference. One-Way ANOVA followed by Tukey’s test was employed to compare AUCs. The log-rank Mantel-Cox test was used to compare survival curves. Statistical significance was defined as p < 0.05.
3. Results
3.1. Radical Scavenging Assessment
In the DPPH and ABTS radical inhibition assays, the values obtained with Terasen® were 20.85 ± 2.99% and 69.95 ± 5.39% inhibition relative to DMSO, respectively, compared to 75.35 ± 4.42% and 95.82 ± 0.39% inhibition relative to DMSO in the gallic acid control. It is observed that the inhibition percentage was significantly higher in the ABTS assay, and this result is directly related to the phenolic compound content present in Terasen®. Phenolic compounds have a higher affinity for scavenging the ABTS radical due to differences in their chemical structures and the properties of free radicals [13,14].
3.2. Assessment of Pharyngeal Pumping Rate
In order to investigate whether exposure to Terasen® could affect the pharyngeal pumping rate, a crucial measure for evaluating feeding behavior in C. elegans [11], an investigation was conducted in C. elegans exposed to different concentrations of Terasen® (250, 500, or 1000 µg/mL), which revealed statistically significant results. Compared to the control group, there was a statistically significant reduction in pharyngeal pumping rate on days 3, 6, and 9 of adult life in C. elegans exposed to Terasen® (p < 0.05; Figure 1). These findings suggest that exposure to Terasen® may reduce the feeding behavior of these organisms.
3.3. Reproduction Assessment
To investigate the impact of Terasen® on C. elegans reproduction, egg-laying was evaluated in animals treated with different concentrations of the formulation (250, 500, or 1000 µg/mL). The results revealed a significant reduction in egg production in animals exposed to Terasen® compared to the control group (p < 0.001; Figure 2). However, it is observed that this drastic decrease tends to mitigate from the second day on and could be explained by possible initial stress due to an exogenous compound in the medium. Still, mainly due to the initial decrease, there was a statistical difference in overall values given by the AUC. Among the treated groups, there was a concentration-dependent increase in the progeny.
3.4. Locomotion Assessment
The movement capacity of C. elegans was evaluated in control animals and animals treated with different concentrations of Terasen® (250, 500, or 1000 µg/mL). The results demonstrate that Terasen® did not promote decline or improvement in nematode motility in the young adult phase (day 4). During the aging phase (day 8), nematode motility was reduced in animals treated with different concentrations of Terasen® (Figure 3). However, considering the data from all days, the difference was insignificant considering treatment (p = 0.0952; Two-Way ANOVA).
3.5. Size Evaluation
The results demonstrated a reduction in body length with increasing Terasen® concentration compared to the control group on day 4 (Figure 4). However, on the 8th day of adulthood, with increased exposure time, the length of the animals treated with Terasen® and the control group did not show a noticeable variation when compared to each other. Considering data from all days, however, it is observed a significant effect of the treatment (p < 0.05; Two-Way ANOVA) that was dependent on the day (Interaction: p < 0.05).
3.6. Lifespan Evaluation
The potential of Terasen® on longevity was assessed. The results showed that, compared to the control group, the average lifespan of C. elegans was significantly (p < 0.05) prolonged after exposure to Terasen® treatment (Figure 5). The median survival (days) was 6 for the control, 6 for Terasen® 250 µg/mL, 7 for 500 µg/mL, and 9 for 1000 µg/mL. The maximum lifespan of wild-type C. elegans increased by 2 days (22.22%) after treatment with 500 µg/mL of Terasen® (p<0.005) and 5 days (55.56%) after treatment with 1000 µg/mL compared to the control group. Based on the results, the use of Terasen® showed a concentration-dependent boost in the lifespan of nematodes.
4. Discussion
With aging, there is an imbalance of reactive species. Hence, we assessed the radical scavenging of Terasen® through DPPH and ABTS methods. Although the radical scavenging in the DPPH assay was low compared to the positive control gallic acid, it was high in the ABTS assay. This could be explained by the presence of the phenolic compounds from the formulation with a higher affinity for radical scavenging of ABST [13,14].
Then, the potential of Terasen® on longevity was evaluated in vivo using C. elegans as a model organism. This tiny nematode, which measures only 1 mm in length as an adult, is cultured in Petri dishes using solid and liquid culture media, and its primary food source is Escherichia coli. With its two types of sex, hermaphrodite (XX) and male (XO), the hermaphrodite can generate 300 to 1400 offspring, facilitating the creation of a uniform genetic lineage [15].
Despite its simple anatomy with few tissues and organs, the nervous system, gastrointestinal tract, gonads, and muscles of C. elegans are comparable to those of more complex animals [16]. Although it lacks adipose tissue or a liver, its intestine can perform similar functions, such as lipid synthesis and lipoprotein secretion [17]. The genetic sequence of C. elegans is well known, and genomic research has revealed that its genome has 60 to 80% of genes similar to those of human diseases and metabolic pathways [18].
C. elegans has proven to be a valuable organism for studying aging, as it undergoes noticeable changes over time, such as alterations in body movement, pharyngeal pumping, egg-laying posture, and body size, among others [19,20].
The pharynx of C. elegans has rhythmic contractions that occur at high frequencies in young adults, reaching 200-300 contractions per minute. However, with aging, there is a progressive decrease in the frequency of pharyngeal contractions [21]. This study observed that the pharyngeal pumping rate of Terasen®-treated groups was decreased compared to the control group, indicating that Terasen® likely decreased the nematode’s feeding behavior.
Caloric restriction has been widely studied and is associated with increased longevity in various organisms, including C. elegans, because reducing calorie intake promotes metabolic changes that stimulate stress resistance and cellular protection [22]. This was observed in the study conducted by (Lakowski and Hekimi, 1998), where the authors reported that caloric restriction extended the lifespan of C. elegans.
During this study, the production of eggs in C. elegans was monitored daily, revealing a decrease in the number of progenies in animals treated with Terasen® compared to the control group. Monitoring reproductive capacity is essential to age research, as several interventions that prolong adult lifespan impact progeny production. Dietary restrictions also have been associated with reductions in progeny production during the early life phase of animals and a reduction in the total number of progenies produced by self-fertile hermaphrodites [22].
The results showed that after 4 days of Terasen® treatment, there were significant changes in the body length of the nematodes compared to the control, although this difference mitigates on the eighth day. As for the body movements, there were no statistically significant changes in the animals treated (p > 0.05; Two-Way ANOVA), although a trend was observed of decreased movement. Usually, young adult hermaphrodites exhibit fluid and synchronized body movement. However, their motor coordination deteriorates as they age, becoming increasingly uncoordinated and unpredictable until they stop moving [19].
Finally, we observed that the treatment increased the average and maximum lifespan of the nematodes. As observed, other effects were seen, including decreased pharyngeal contractions, reduced body length, and decreased laid eggs. Notably, all these effects are also observed during caloric restriction [24]. Hence, it appears that Terasen® may activate the same pathways triggered by caloric restriction, even if food is normally provided.
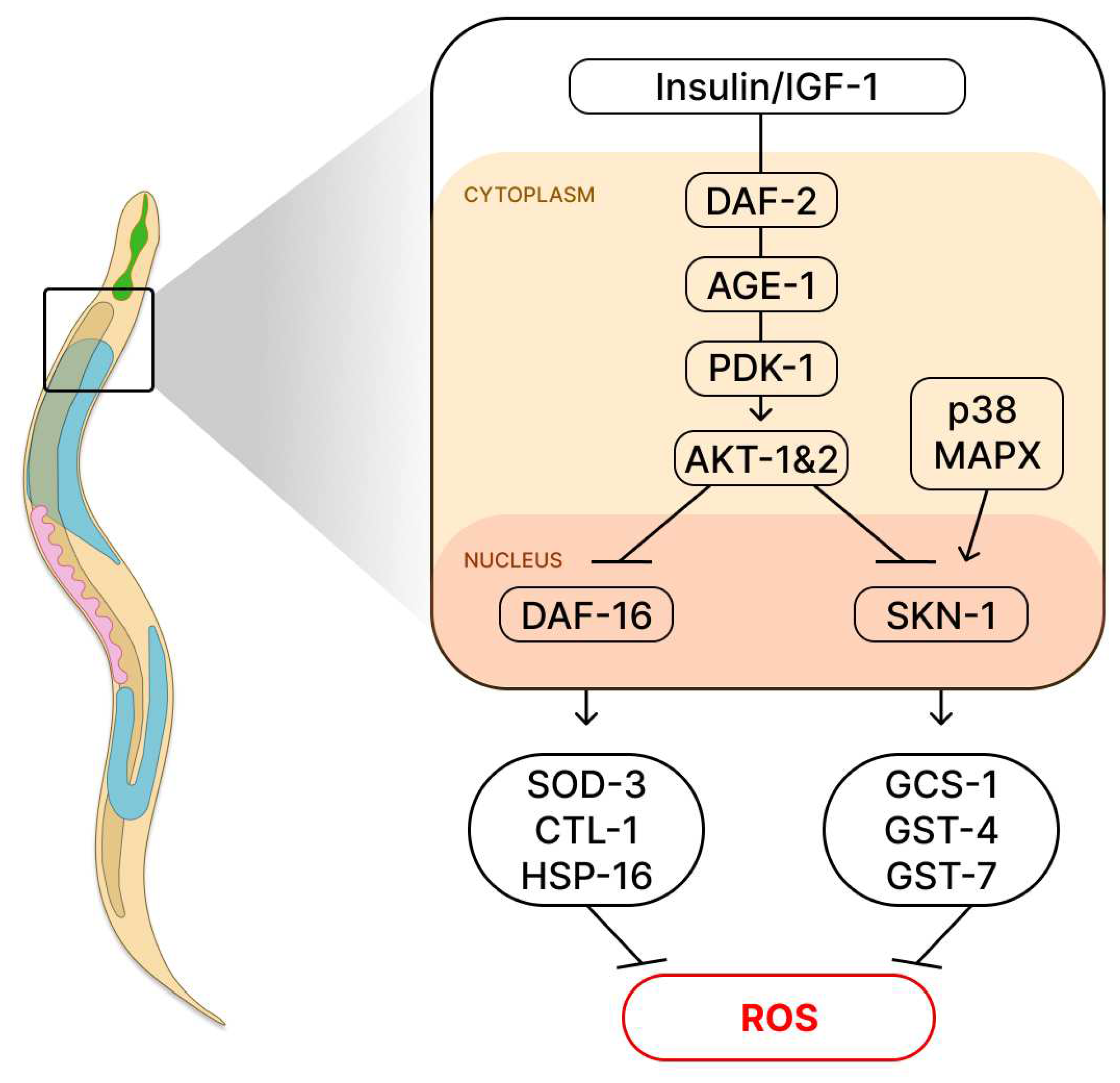
Caloric restriction is a non-genetic intervention that can increase the lifespan and decrease the incidence of age-related diseases. Hence, a body of research has been aiming to test the mechanisms beneath this process to achieve healthy aging. Some of the core components involved in the beneficial effects of caloric restriction are the mTOR [25,26] and the insulin/insulin-like growth factor-1 signaling (IIS) pathway in humans [27,28]. Accordingly, in C. elegans, the IGF-1 receptor is codified by the gene daf-2 in worms (igf1 in humans); reduction of this pathway will cause activation of DAF-16 (orthologue of the human FOXO), a transcription factor that will synthesize products involved in longevity and stress resilience [9,29]. The gene daf-16 is related to increased life expectancy in C. elegans [9,30]. Another important player involved in life expectancy in C. elegans is SKN-1 (orthologue of the human Nrf2), which is involved in the resistance to oxidative stress (Figure 6) [31].
Finally, we observed that the treatment increased the average and maximum lifespan of the nematodes. As observed, other effects were seen, including decreased pharyngeal contractions, reduced body length, and decreased laid eggs. Notably, all these effects are also observed during caloric restriction [24]. Hence, it appears that Terasen® may activate the same pathways triggered by caloric restriction, even if food is normally provided.
Caloric restriction is a non-genetic intervention that can increase the lifespan and decrease the incidence of age-related diseases. Hence, a body of research has been aiming to test the mechanisms beneath this process to achieve healthy aging. Some of the core components involved in the beneficial effects of caloric restriction are the mTOR [25,26] and the insulin/insulin-like growth factor-1 signaling (IGF-1) pathway in humans [27,28,32]. Accordingly, in C. elegans, the IGF-1 receptor is codified by the gene daf-2 in worms (IGF-1 in humans); reduction of this pathway will cause activation of DAF-16 (orthologue of the human FOXO), a transcription factor that will synthesize products involved in longevity and stress resilience [9,29]. The gene daf-16 is related to increased life expectancy in C. elegans [9,30]. Another important player involved in life expectancy in C. elegans is SKN-1 (orthologue of the human Nrf2), which is involved in the resistance to oxidative stress (Figure 6) [31,33].
Studies in the literature report that some plant secondary metabolites can aid in longevity by acting in these pathways. In line with the results observed here, some phytochemical compounds present in Terasen® have been reported to increase C. elegan’s lifespan, including anthocyanins, which are found in E. oleracea, quercetin, which is a marker from M. dubia, and tocotrienols, abundantly found in B. orellana.
[29] reported that the extract of black rice, which is a rich source of anthocyanins (representing 43% of the extract), extended the lifespan, enhanced stress resistance, increased antioxidant enzymes activity, and reduced the accumulation of lipofuscin, ROS, and MDA. The treatment downregulated the expression of age-1 and daf-2 mRNA while upregulated daf-16 mRNA and upregulated protein expression of SOD-3, CTL-1, and GST. The major anthocyanins found were cyanidin-3-O-glucoside (76.15 %) and peonidin-3-O-glucoside (22.11 %).
The increased lifespan and stress resistance in C. elegans induced by cyanidin-3-O-glucoside (the most abundant anthocyanin from E. oleracea) was also reported by [34]. Other anthocyanin-rich sources that were reported to increase C. elegans’ lifespan were Prunus cerasus [35], Paeonia suffruticosa [36], Eugenia uniflora [37], and Euterpe precatoria [38]. E. oleracea also increased longevity in Drosophila melanogaster [39].
M. dubia has quercetin as marker. [40] reported that quercetin increased the mean lifespan of C. elegans by 15%, and increased the translocation of DAF-16 to the nucleus, a mechanism correlated with stress response and longevity. This was also reported by [41], who also showed that quercetin is safer than its glycosylated forms.
The phytochemical markers of A. aculeatum include gallic acid, ellagic acid, and carotenoids. Cocoa beans, derived from Theobroma cacao, are commonly used to produce chocolate and cocoa powder. A single serving of cocoa powder (about 2 tablespoons) contains high levels of gallic acid [42]. In a study conducted by [43], the effects of cocoa powder enriched with polyphenols were evaluated on resistance to oxidative stress in biological models such as the yeast Saccharomyces cerevisiae and the nematode C. elegans. The results showed that cocoa enriched with polyphenols increased resistance to oxidative stress in both models, as well as life expectancy in the nematode. These effects were attributed to the presence of polyphenols in cocoa and the sirtuins Hst3 and SIR-2.1, in addition to the insulin/IGF-1 signaling pathway in the nematode.
A recent study [44] delved into the impact of extracts from Glochidion zeylanicum leaves on C. elegans, specifically examining their anti-aging and oxidative stress resistance properties. The study employed HPLC analysis to identify oxyresveratrol and quercetin, as well as high levels of gallic acid and catechin within the extracts. Findings revealed that these leaf extracts from G. zeylanicum provided protection against oxidative stress, regulated stress response genes like SOD-3 and GST-4, and involved transcription factors DAF-16/FoxO and SKN-1/Nrf-2 in the oxidative stress resistance properties. Additionally, the extracts improved pharyngeal pumping function and increased the life expectancy of worms, suggesting anti-aging benefits.
The impact of raspberry extracts (RE), which consist of various phytochemical compounds such as ellagic acid, salicylic acid, chlorogenic acid, p-coumaric acid, quercitrin, catechin, and luteolin, on the lifespan of C. elegans and the underlying mechanisms were studied [33]. The findings demonstrate that RE enhances resistance to oxidative stress, increases the activity of antioxidant enzymes, and decreases the excessive production of reactive oxygen species. Nevertheless, in certain mutants, the administration of RE had disparate effects, indicating that the SKN-1/Nrf2 pathway is vital in regulating RE-induced longevity and response to oxidative stress.
A study by [45] evaluated the effects of ß-carotene and fucoxanthin on the lifespans of D. melanogaster and C. elegans. The findings indicated that both carotenoids extended the lifespan of fruit flies, while only fucoxanthin exhibited beneficial effects on nematodes. The activation of the FOXO transcription factor by carotenoids initiated a series of events that triggered the activation of stress response genes, culminating in heightened resistance to stressors and an increase in lifespan.
According to research conducted by [46], mamey carotenoids were analyzed for their antioxidant properties on nematodes. The study found that both pure β-carotene and mamey pulp extract (MPCE) enhanced resistance to oxidative stress and increased the survival rate of the nematodes. However, it was observed that mamey skin extract (MSCE) had adverse effects due to the degradation of β-carotene. Even though high concentrations of MSCE were toxic, they still managed to improve survival, likely due to antioxidant mechanisms and the phenomenon of hormesis.
Finally, Terasen® has the oil of B. orellana in its composition, a rich source of tocotrienols. Tocotrienols are another group of compounds that potentially increase lifespan. [47] showed that C. elegans subjected to oxidative stress with hydrogen peroxide had decreased lifespan and increased lipofuscin accumulation. On the other hand, animals treated with a tocotrienols-rich fraction had restored lifespan and reduced lipofuscin accumulation. [48] reported that the lipophilic extract of B. orellana increased the median and maximum lifespan by 35% and 27%, increasing oxidative and thermal stress without affecting the fertility (which is different from what was observed here). According to the authors, the effect depended on the insulin/insulin growth factor-1 pathway.
5. Conclusions
Based on this research evaluating the potential of Terasen® in longevity, using C. elegans as a study model, it can be concluded that Terasen® was effective in significantly extending the median and maximum lifespan of the worms compared to untreated animals. Terasen® also affected the feeding behavior of the nematode, resulting in a reduction in the pharyngeal pumping rate. The effects observed here are similar to those of caloric restriction, including decreased progeny and size, even if food is typically provided. These results are consistent with other research showing that secondary metabolites found in Terasen® can also prolong life in C. elegans.
Author Contributions
Conceptualization, J.C.T.C. and E.L.M.; methodology, E.L.M.; validation, E.L.M.; formal analysis, H.O.C.; investigation, I.S.S., A.L.N., B.A.M.T. and N.N.D.C.; resources, J.C.T.C. and C.E.W.; writing—original draft preparation, E.L.M.; writing—review and editing, E.L.M. and A.C.M.P.; visualization, L.F.M. and A.N.C.; supervision, J.C.T.C.; project administration, E.L.M. and J.C.T.C.; funding acquisition, J.C.T.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data will be made available upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Briga, M.; Verhulst, S. What Can Long-Lived Mutants Tell Us about Mechanisms Causing Aging and Lifespan Variation in Natural Environments? Exp Gerontol 2015, 71, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Padwad, Y. Perspectives of the Potential Implications of Polyphenols in Influencing the Interrelationship between Oxi-Inflammatory Stress, Cellular Senescence and Immunosenescence during Aging. Trends Food Sci Technol 2020, 98, 41–52. [Google Scholar] [CrossRef]
- de Freitas Rodrigues, C.; Ramos Boldori, J.; Valandro Soares, M.; Somacal, S.; Emanuelli, T.; Izaguirry, A.; Weber Santos Cibin, F.; Rossini Augusti, P.; Casagrande Denardin, C. Goji Berry (Lycium Barbarum L.) Juice Reduces Lifespan and Premature Aging of Caenorhabditis Elegans: Is It Safe to Consume It? Food Research International 2021, 144, 110297. [Google Scholar] [CrossRef]
- WHO, W.H.O. Ageing.
- Trendelenburg, A.U.; Scheuren, A.C.; Potter, P.; Müller, R.; Bellantuono, I. Geroprotectors: A Role in the Treatment of Frailty. Mech Ageing Dev 2019, 180, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.E.; Lee, S.-J. v. Recent Progresses on Anti-Aging Compounds and Their Targets in Caenorhabditis Elegans. Transl Med Aging 2019, 3, 121–124. [Google Scholar] [CrossRef]
- Molyneux, P. The Use of the Stable Free Radical Diphenylpicrylhydrazyl (DPPH) for Estimating Antioxidant. Songklanakarin Journal of Science and Technology (SJST) 2004, 26, 211–219. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic Biol Med 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Li, H.; Roxo, M.; Cheng, X.; Zhang, S.; Cheng, H.; Wink, M. Pro-Oxidant and Lifespan Extension Effects of Caffeine and Related Methylxanthines in Caenorhabditis Elegans. Food Chem X 2019, 1, 100005. [Google Scholar] [CrossRef]
- Lu, L.; Zhao, X.; Zhang, J.; Li, M.; Qi, Y.; Zhou, L. Calycosin Promotes Lifespan in Caenorhabditis Elegans through Insulin Signaling Pathway via Daf-16, Age-1 and Daf-2. J Biosci Bioeng 2017, 124, 1–7. [Google Scholar] [CrossRef]
- Liao, V.H.-C.; Yu, C.-W.; Chu, Y.-J.; Li, W.-H.; Hsieh, Y.-C.; Wang, T.-T. Curcumin-Mediated Lifespan Extension in Caenorhabditis Elegans. Mech Ageing Dev 2011, 132, 480–487. [Google Scholar] [CrossRef]
- Huang, C.-W.; Liao, W.-R.; How, C.M.; Yen, P.-L.; Wei, C.-C. Chronic Exposure of Zearalenone Inhibits Antioxidant Defense and Results in Aging-Related Defects Associated with DAF-16/FOXO in Caenorhabditis Elegans. Environmental Pollution 2021, 285, 117233. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Wu, X. Anthocyanins: Structural Characteristics That Result in Unique Metabolic Patterns and Biological Activities. Free Radic Res 2006, 40, 1014–1028. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Cano, A.; Acosta, M. The Hydrophilic and Lipophilic Contribution to Total Antioxidant Activity. Food Chem 2001, 73, 239–244. [Google Scholar] [CrossRef]
- Yue, Y.; Li, S.; Shen, P.; Park, Y. Caenorhabditis Elegans as a Model for Obesity Research. Curr Res Food Sci 2021, 4, 692–697. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Guo, P.; Wang, P.; Zheng, S.; Qu, Z.; Liu, N. The Review of Anti-Aging Mechanism of Polyphenols on Caenorhabditis Elegans. Front Bioeng Biotechnol 2021, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lemieux, G.A.; Ashrafi, K. Investigating Connections between Metabolism, Longevity, and Behavior in Caenorhabditis Elegans. Trends in Endocrinology and Metabolism 2016, 27, 586–596. [Google Scholar] [CrossRef]
- Lee, S.A.; Lim, W.H.; Van Le, V.; Ko, S.R.; Kim, B.; Oh, H.M.; Ahn, C.Y. Lifespan Extension and Anti-Oxidant Effects of Carotenoid Pigments in Caenorhabditis Elegans. Bioresour Technol Rep 2022, 17. [Google Scholar] [CrossRef]
- Huang, C.; Xiong, C.; Kornfeld, K. Measurements of Age-Related Changes of Physiological Processes That Predict Lifespan of Caenorhabditis Elegans. Proc Natl Acad Sci U S A 2004, 101, 8084–8089. [Google Scholar] [CrossRef]
- Wang, Q.; Li, H.; Zhang, G.; Chen, X.; Wang, X. Itaconate Prolongs the Healthy Lifespan by Activating UPRmt in Caenorhabditis Elegans. Eur J Pharmacol 2022, 923, 174951. [Google Scholar] [CrossRef]
- Chow, D.K.; Glenn, C.F.; Johnston, J.L.; Goldberg, I.G.; Wolkow, C.A. Sarcopenia in the Caenorhabditis Elegans Pharynx Correlates with Muscle Contraction Rate over Lifespan. Exp Gerontol 2006, 41, 252–260. [Google Scholar] [CrossRef]
- Evason, K.; Collins, J.J.; Huang, C.; Hughes, S.; Kornfeld, K. Valproic Acid Extends Caenorhabditis Elegans Lifespan. Aging Cell 2008, 7, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Lakowski, B.; Hekimi, S. The Genetics of Caloric Restriction in Caenorhabditis Elegans. Proc Natl Acad Sci U S A 1998, 95, 13091–13096. [Google Scholar] [CrossRef] [PubMed]
- Liao, V.H.C.; Yu, C.W.; Chu, Y.J.; Li, W.H.; Hsieh, Y.C.; Wang, T.T. Curcumin-Mediated Lifespan Extension in Caenorhabditis Elegans. Mech Ageing Dev 2011, 132, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Lapierre, L.R.; Hansen, M. Lessons from C. Elegans: Signaling Pathways for Longevity. Trends in Endocrinology & Metabolism 2012, 23, 637–644. [Google Scholar] [CrossRef]
- Vellai, T.; Takacs-Vellai, K.; Zhang, Y.; Kovacs, A.L.; Orosz, L.; Müller, F. Influence of TOR Kinase on Lifespan in C. Elegans. Nature 2003, 426, 620–620. [Google Scholar] [CrossRef]
- Cao, L.; Lee, S.G.; Park, S.-H.; Kim, H.-R. Sargahydroquinoic Acid (SHQA) Suppresses Cellular Senescence through Akt/MTOR Signaling Pathway. Exp Gerontol 2021, 151, 111406. [Google Scholar] [CrossRef]
- Wilkinson, D.S.; Taylor, R.C.; Dillin, A. Analysis of Aging in Caenorhabditis Elegans. In; 2012; pp. 353–381.
- Li, X.; Wang, X.; Wang, K.; Yang, X.; Liu, X.; Chen, J.; Li, J.; Wang, J.; Guo, Q.; Wang, H. Black Rice Anthocyanin Extract Enhances the Antioxidant Capacity in PC12 Cells and Improves the Lifespan by Activating IIS Pathway in Caenorhabditis Elegans. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2023, 265, 109533. [Google Scholar] [CrossRef]
- Zhu, J.; Jia, Y.; Wang, C.; Zhou, W.; Shu, Y.; Zhang, K.; Zeng, X.; Guo, R. Lonicera Japonica Polysaccharides Improve Longevity and Fitness of Caenorhabditis Elegans by Activating DAF-16. Int J Biol Macromol 2023, 229, 81–91. [Google Scholar] [CrossRef]
- Cui, X.; Zhang, B.; Li, Z.; Li, C.; Li, J. Zhuyeqing Liquor Promotes Longevity through Enhancing Stress Resistance via Regulation of SKN-1 and HSF-1 Transcription Factors in Caenorhabditis Elegans. Exp Gerontol 2023, 174, 112131. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, L.; Zhou, L. Oleanolic Acid Activates Daf-16 to Increase Lifespan in Caenorhabditis Elegans. Biochem Biophys Res Commun 2015, 468, 843–849. [Google Scholar] [CrossRef]
- Song, B.; Zheng, B.; Li, T.; Liu, R.H. Raspberry Extract Ameliorates Oxidative Stress in Caenorhabditis Elegans via the SKN-1/Nrf2 Pathway. J Funct Foods 2020, 70, 103977. [Google Scholar] [CrossRef]
- Chen, W.; Chen, Z.; Shan, S.; Wu, A.; Zhao, C.; Ye, X.; Zheng, X.; Zhu, R. Cyanidin-3-O-Glucoside Promotes Stress Tolerance and Lifespan Extension of Caenorhabditis Elegans Exposed to Polystyrene via DAF-16 Pathway. Mech Ageing Dev 2022, 207, 111723. [Google Scholar] [CrossRef] [PubMed]
- van de Klashorst, D.; van den Elzen, A.; Weeteling, J.; Roberts, M.; Desai, T.; Bottoms, L.; Hughes, S. Montmorency Tart Cherry (Prunus Cerasus L.) Acts as a Calorie Restriction Mimetic That Increases Intestinal Fat and Lifespan in Caenorhabditis Elegans. J Funct Foods 2020, 68, 103890. [Google Scholar] [CrossRef]
- Wang, S.; Xue, J.; Zhang, S.; Zheng, S.; Xue, Y.; Xu, D.; Zhang, X. Composition of Peony Petal Fatty Acids and Flavonoids and Their Effect on Caenorhabditis Elegans Lifespan. Plant Physiology and Biochemistry 2020, 155, 1–12. [Google Scholar] [CrossRef]
- Tambara, A.L.; de Los Santos Moraes, L.; Dal Forno, A.H.; Boldori, J.R.; Gonçalves Soares, A.T.; de Freitas Rodrigues, C.; Mariutti, L.R.B.; Mercadante, A.Z.; de Ávila, D.S.; Denardin, C.C. Purple Pitanga Fruit (Eugenia Uniflora L.) Protects against Oxidative Stress and Increase the Lifespan in Caenorhabditis Elegans via the DAF-16/FOXO Pathway. Food and Chemical Toxicology 2018, 120, 639–650. [Google Scholar] [CrossRef]
- Peixoto, H.; Roxo, M.; Krstin, S.; Röhrig, T.; Richling, E.; Wink, M. An Anthocyanin-Rich Extract of Acai (Euterpe Precatoria Mart.) Increases Stress Resistance and Retards Aging-Related Markers in Caenorhabditis Elegans. J Agric Food Chem 2016, 64, 1283–1290. [Google Scholar] [CrossRef]
- Sun, X.; Seeberger, J.; Alberico, T.; Wang, C.; Wheeler, C.T.; Schauss, A.G.; Zou, S. Açai Palm Fruit (Euterpe Oleracea Mart.) Pulp Improves Survival of Flies on a High Fat Diet. Exp Gerontol 2010, 45, 243–251. [Google Scholar] [CrossRef]
- Kampkötter, A.; Timpel, C.; Zurawski, R.F.; Ruhl, S.; Chovolou, Y.; Proksch, P.; Wätjen, W. Increase of Stress Resistance and Lifespan of Caenorhabditis Elegans by Quercetin. Comp Biochem Physiol B Biochem Mol Biol 2008, 149, 314–323. [Google Scholar] [CrossRef]
- Dueñas, M.; Surco-Laos, F.; González-Manzano, S.; González-Paramás, A.M.; Gómez-Orte, E.; Cabello, J.; Santos-Buelga, C. Deglycosylation Is a Key Step in Biotransformation and Lifespan Effects of Quercetin-3-O-Glucoside in Caenorhabditis Elegans. Pharmacol Res 2013, 76, 41–48. [Google Scholar] [CrossRef]
- Chattopadhyay, D.; Thirumurugan, K. Longevity Promoting Efficacies of Different Plant Extracts in Lower Model Organisms. Mech Ageing Dev 2018, 171, 47–57. [Google Scholar] [CrossRef]
- Martorell, P.; Forment, J.V.; de Llanos, R.; Montón, F.; Llopis, S.; González, N.; Genovés, S.; Cienfuegos, E.; Monzó, H.; Ramón, D. Use of Saccharomyces Cerevisiae and Caenorhabditis Elegans as Model Organisms To Study the Effect of Cocoa Polyphenols in the Resistance to Oxidative Stress. J Agric Food Chem 2011, 59, 2077–2085. [Google Scholar] [CrossRef] [PubMed]
- van de Klashorst, D.; van den Elzen, A.; Weeteling, J.; Roberts, M.; Desai, T.; Bottoms, L.; Hughes, S. Montmorency Tart Cherry (Prunus Cerasus L.) Acts as a Calorie Restriction Mimetic That Increases Intestinal Fat and Lifespan in Caenorhabditis Elegans. J Funct Foods 2020, 68, 103890. [Google Scholar] [CrossRef]
- Lashmanova, E.; Proshkina, E.; Zhikrivetskaya, S.; Shevchenko, O.; Marusich, E.; Leonov, S.; Melerzanov, A.; Zhavoronkov, A.; Moskalev, A. Fucoxanthin Increases Lifespan of Drosophila Melanogaster and Caenorhabditis Elegans. Pharmacol Res 2015, 100, 228–241. [Google Scholar] [CrossRef] [PubMed]
- González-Peña, M.A.; Lozada-Ramírez, J.D.; Ortega-Regules, A.E. Carotenoids from Mamey (Pouteria Sapota) and Carrot (Daucus Carota) Increase the Oxidative Stress Resistance of Caenorhabditis Elegans. Biochem Biophys Rep 2021, 26, 100989. [Google Scholar] [CrossRef] [PubMed]
- Aan, G.; Zainudin, M.; Karim, N.; Ngah, W. Effect of the Tocotrienol-Rich Fraction on the Lifespan and Oxidative Biomarkers in Caenorhabditis Elegans under Oxidative Stress. Clinics 2013, 68, 599–604. [Google Scholar] [CrossRef]
- Gómez-Linton, D.R.; Alavez, S.; Navarro-Ocaña, A.; Román-Guerrero, A.; Pinzón-López, L.; Pérez-Flores, L.J. Achiote (Bixa Orellana) Lipophilic Extract, Bixin, and δ-Tocotrienol Effects on Lifespan and Stress Resistance in Caenorhabditis Elegans. Planta Med 2021, 87, 368–374. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of treatment with Terasen® 250 µg/mL (G 0.25), 500 µg/mL (G 0.5), or 1000 µg/mL (G1) and Escherichia coli on Pharyngeal Pumping Rate of C. elegans in the treated groups. Results are expressed as mean ± SEM, where * indicates a significant difference vs. the control group on the same day and # indicates a significant difference vs. the same group on day 3. Treatment factor: p < 0.0001; Day factor: p < 0.0001; Interaction: ns (p < 0.05; Two-Way ANOVA followed by the posthoc Tukey’s test).
Figure 1.
Effect of treatment with Terasen® 250 µg/mL (G 0.25), 500 µg/mL (G 0.5), or 1000 µg/mL (G1) and Escherichia coli on Pharyngeal Pumping Rate of C. elegans in the treated groups. Results are expressed as mean ± SEM, where * indicates a significant difference vs. the control group on the same day and # indicates a significant difference vs. the same group on day 3. Treatment factor: p < 0.0001; Day factor: p < 0.0001; Interaction: ns (p < 0.05; Two-Way ANOVA followed by the posthoc Tukey’s test).
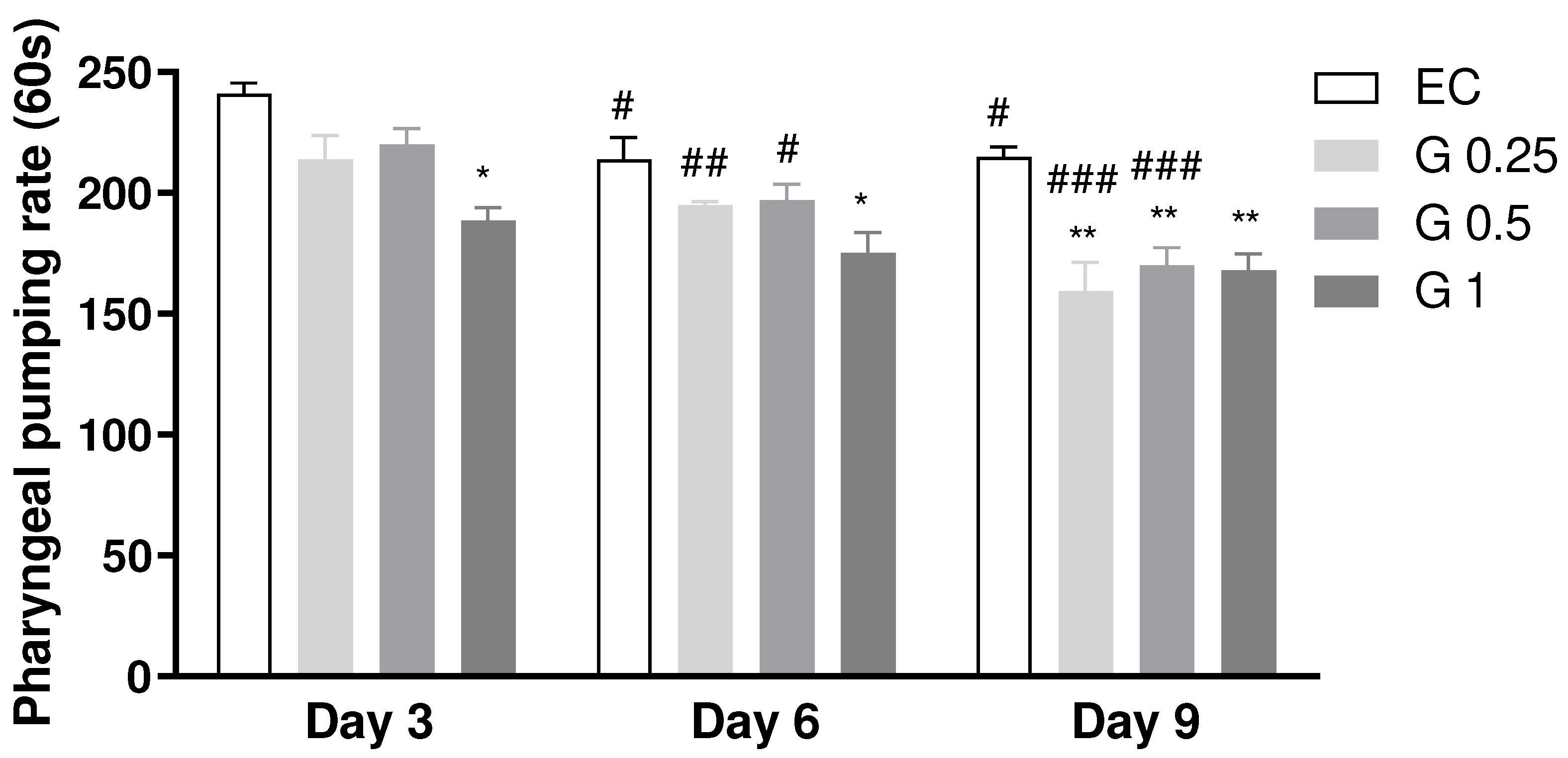
Figure 2.
Effect of treatment with Terasen® 250 µg/mL (G 0.25), 500 µg/mL (G 0.5), or 1000 µg/mL (G1) and Escherichia coli on C. elegans reproduction. (A) Represents the results with mean and SEM of the groups according to the days evaluated; (B) Expresses the area under the curve, ***statistically significant results (p < 0.001) vs. EC group (One-Way ANOVA followed by Tukey posthoc test).
Figure 2.
Effect of treatment with Terasen® 250 µg/mL (G 0.25), 500 µg/mL (G 0.5), or 1000 µg/mL (G1) and Escherichia coli on C. elegans reproduction. (A) Represents the results with mean and SEM of the groups according to the days evaluated; (B) Expresses the area under the curve, ***statistically significant results (p < 0.001) vs. EC group (One-Way ANOVA followed by Tukey posthoc test).
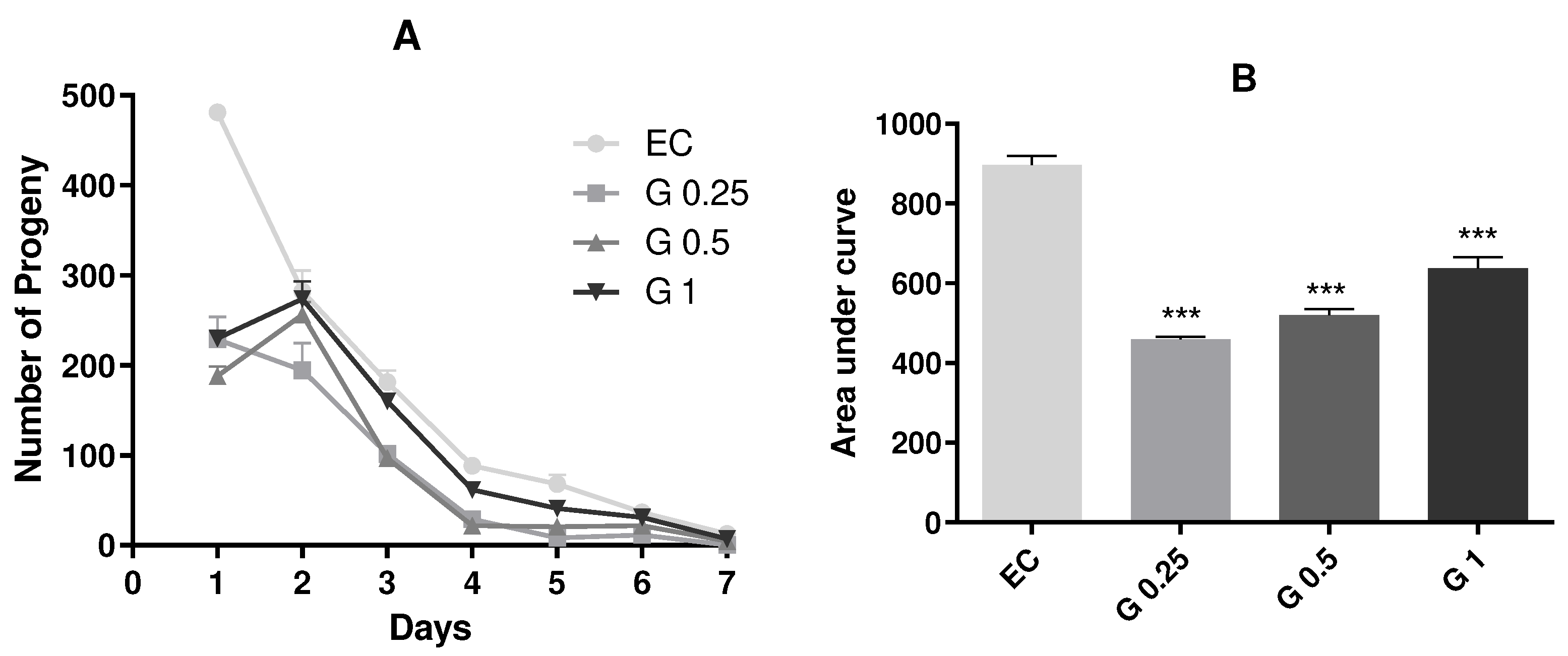
Figure 3.
Effect of treatment with Terasen® at 250 µg/mL (G 0.25), 500 µg/mL (G 0.5), or 1000 µg/mL (G1) and Escherichia coli on the locomotion of C. elegans. Results are expressed as mean and SEM, where # indicates a significant difference vs. the same group on day 4. Treatment factor: ns; Day factor: p < 0.0001; Interaction: ns (p < 0.05; Two-Way ANOVA followed by the posthoc Tukey’s test).
Figure 3.
Effect of treatment with Terasen® at 250 µg/mL (G 0.25), 500 µg/mL (G 0.5), or 1000 µg/mL (G1) and Escherichia coli on the locomotion of C. elegans. Results are expressed as mean and SEM, where # indicates a significant difference vs. the same group on day 4. Treatment factor: ns; Day factor: p < 0.0001; Interaction: ns (p < 0.05; Two-Way ANOVA followed by the posthoc Tukey’s test).
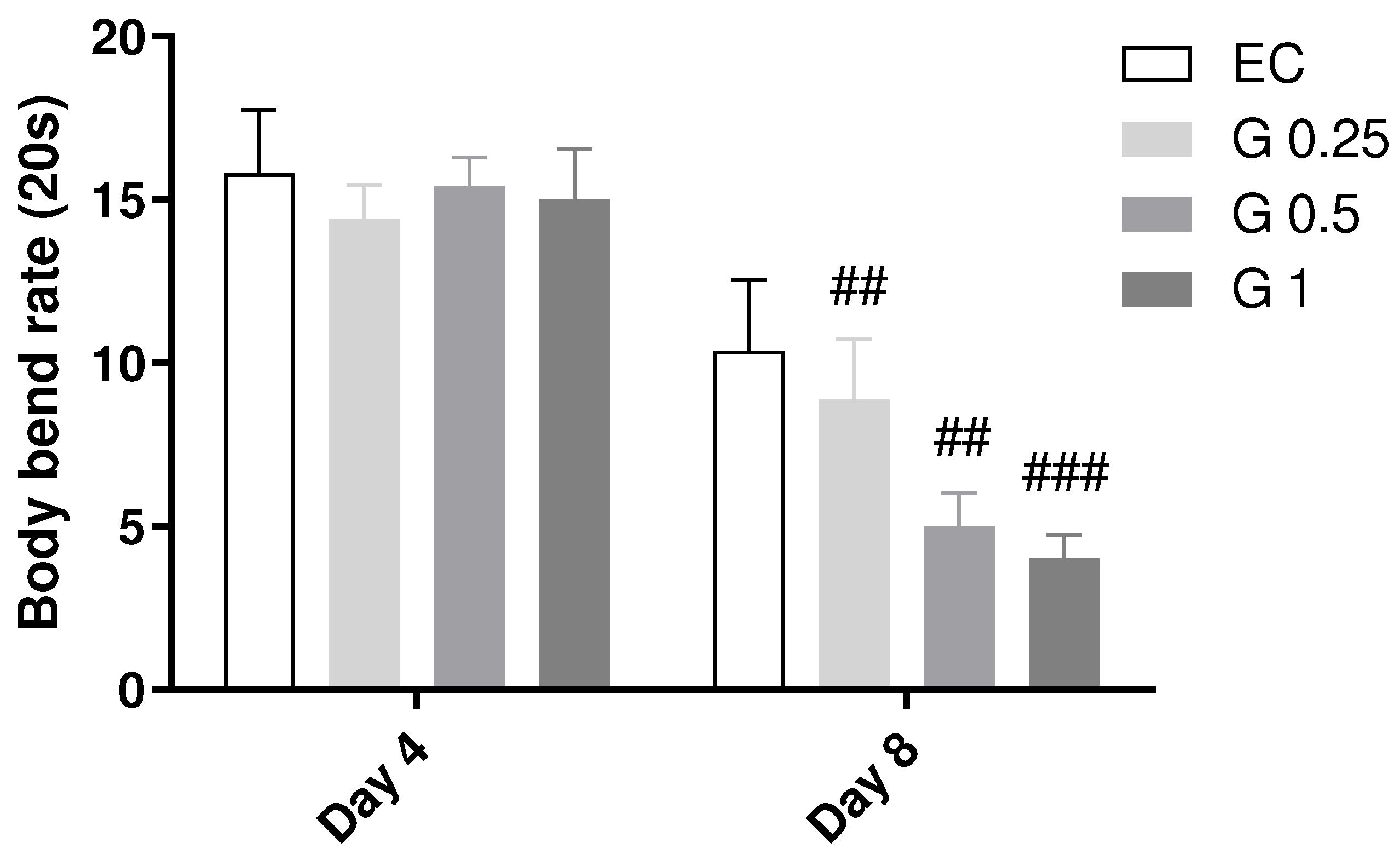
Figure 4.
Effect of treatment with Terasen® at 250 µg/mL (G 0.25), 500 µg/mL (G 0.5), or 1000 µg/mL (G1) and Escherichia coli on the size of C. elegans. Results are expressed as mean and SEM, where * indicates a significant difference vs. the control group on the same day and # indicates a significant difference vs. the same group on day 4. Treatment factor: p < 0.05; Day factor: p < 0.0001; Interaction: p < 0.05 (p < 0.05; Two-Way ANOVA followed by the posthoc Tukey’s test).
Figure 4.
Effect of treatment with Terasen® at 250 µg/mL (G 0.25), 500 µg/mL (G 0.5), or 1000 µg/mL (G1) and Escherichia coli on the size of C. elegans. Results are expressed as mean and SEM, where * indicates a significant difference vs. the control group on the same day and # indicates a significant difference vs. the same group on day 4. Treatment factor: p < 0.05; Day factor: p < 0.0001; Interaction: p < 0.05 (p < 0.05; Two-Way ANOVA followed by the posthoc Tukey’s test).
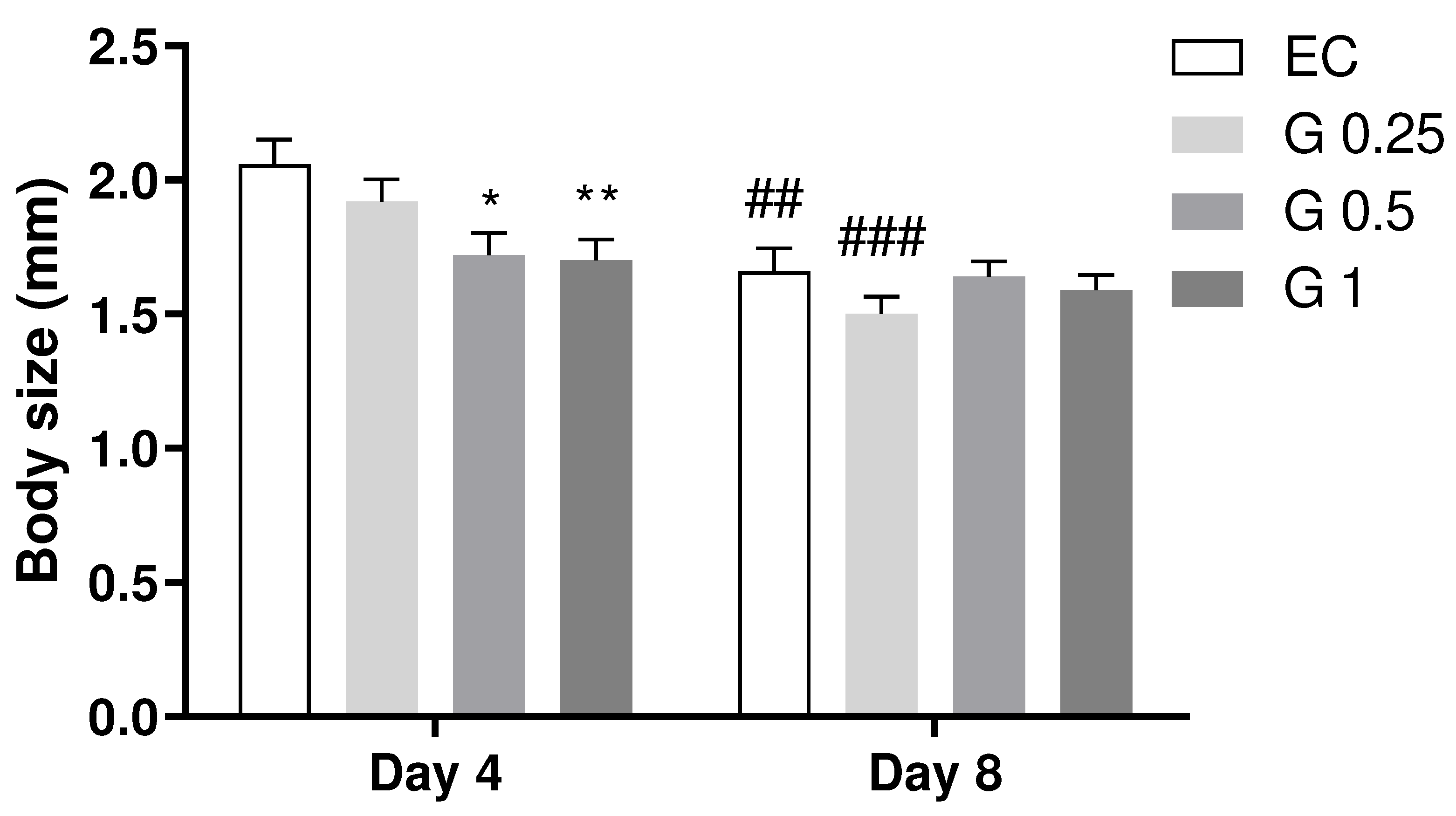
Figure 5.
Effect of treatment with Terasen® at 250 µg/mL (G 0.25), 500 µg/mL (G 0.5), or 1000 µg/mL (G1) and Escherichia coli on the lifespan of C. elegans. The log-rank test using Mantel-Cox survival analysis demonstrated the statistical significance (p<0.05) of the difference between the curves.
Figure 5.
Effect of treatment with Terasen® at 250 µg/mL (G 0.25), 500 µg/mL (G 0.5), or 1000 µg/mL (G1) and Escherichia coli on the lifespan of C. elegans. The log-rank test using Mantel-Cox survival analysis demonstrated the statistical significance (p<0.05) of the difference between the curves.
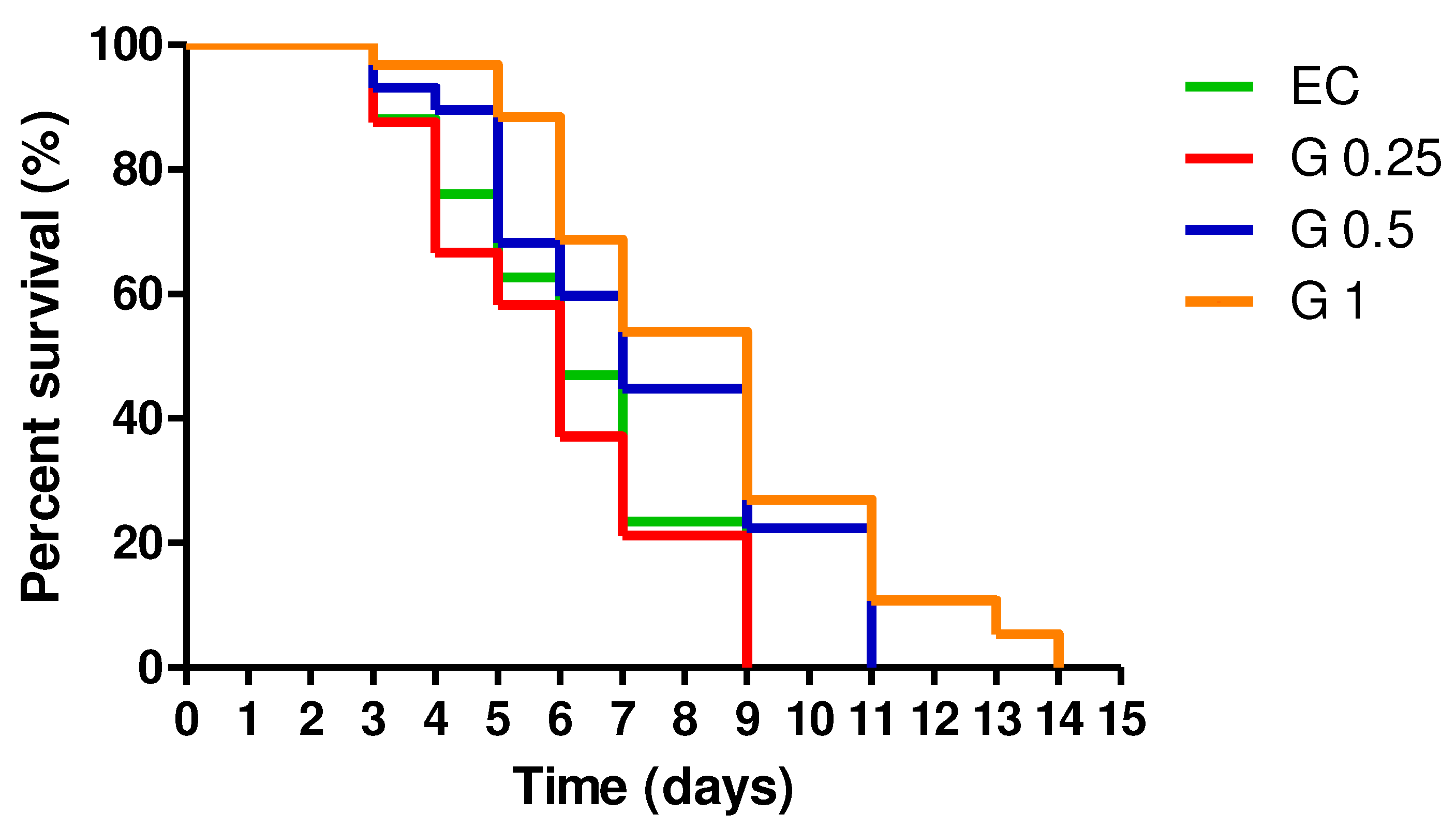
Figure 6.
The insulin/IGF-1 (insulin/insulin-like growth factor-1) signaling pathway and its correlation with the SKN-1 transcription factor is an area of study that warrants a formal overview. The IGF receptor in C. elegans, which is encoded by the DAF-2 gene, initiates a cascade of phosphorylation events involving AGE-1/PI3K, PDK-1, AKT-1/2. By inhibiting the activity of DAF-2, the transcription factor DAF-16 is activated, which in turn activates genes such as superoxide dismutase-3 (sod-3), catalase-1 (ctl-1), and small heat shock protein-16.2 (hsp-16.2), thus providing protection against oxidative stress. The regulation of life expectancy, which involves the transcription factor SKN-1, acts through the activation of p38 mitogens (MAPK). SKN-1 plays a crucial role in regulating antioxidant genes, cellular detoxification, and longevity. The activation of SKN-1 regulates target genes in the nucleus, including GCS-1, GST-4, and GST-7.
Figure 6.
The insulin/IGF-1 (insulin/insulin-like growth factor-1) signaling pathway and its correlation with the SKN-1 transcription factor is an area of study that warrants a formal overview. The IGF receptor in C. elegans, which is encoded by the DAF-2 gene, initiates a cascade of phosphorylation events involving AGE-1/PI3K, PDK-1, AKT-1/2. By inhibiting the activity of DAF-2, the transcription factor DAF-16 is activated, which in turn activates genes such as superoxide dismutase-3 (sod-3), catalase-1 (ctl-1), and small heat shock protein-16.2 (hsp-16.2), thus providing protection against oxidative stress. The regulation of life expectancy, which involves the transcription factor SKN-1, acts through the activation of p38 mitogens (MAPK). SKN-1 plays a crucial role in regulating antioxidant genes, cellular detoxification, and longevity. The activation of SKN-1 regulates target genes in the nucleus, including GCS-1, GST-4, and GST-7.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



