
Submitted:
20 September 2023
Posted:
25 September 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
This study continued the analysis of immunization with the receptor binding domain (RBD) associated with the adjuvants Dimethyldioctadecylammonium bromide (DDA) and Saponin (Sap) (RBD+DDA/Sap) or Aluminum hydroxide (AH) and outer membrane vesicles (OMVs) of Neisseria meningitidis (RBD+AH/OMV); from adult to old age and assessed maternal-fetal transference of antibodies. Outbred Swiss mice were immunized with 2 intramuscular (IM) doses with the antigenic preparations RBD+DDA/Sap, RBD+AH/OMV or RBD alone. The humoral immune response was evaluated using ELISA, avidity-ELISA, Immunoblotting and a Omicron-surrogate-neutralization assay (BA.1), while the cellular immune response was analyzed by ELISpot. Our previous work studied the IgG response; here, we verified that IgM levels were higher right after the immunization doses, but, as IgG, it persisted until 368 days after the immunization. The avidity of IgM increased from low to intermediate-to-high after the booster dose. RBD+DDA/Sap presented neutralizing indexes against Omicron at days 47 and 176, while RBD+AH/OMV showed neutralization on days 21, 47 and 176. Cellular response, measured 465 days after immunization, revealed IFN-Y and IL-4 secretion. The offspring of RBD+DDA/Sap had detectable antibodies, which decreased around 45 days after birth, the last point we analyzed. The results suggested that DDA/Sap is a promising adjuvant mixture to enhance humoral and cellular immune response against SARS-CoV-2, improving maternal-fetal transference of antibodies; cross-reactivity with Omicron variant; and the maintenance of antibodies for a long period.
Keywords:
SARS-CoV-2
; Immune response
; Maternal-fetal IgG transference
; Antibody Functionality
; Antibody persistence
; IFN-Y and IL-4 secretion
; Receptor Binding Domain (RBD)
; Outer Membrane Vesicles (OMV)
; Saponin
; Dioctadecyldimethylammonium Bromide (DDA)
; ELISpot
1. Introduction
Brazil faced an expressive number of COVID-19 cases from 2019 to 2022. Despite the reduction in the number of cases and deaths at the present (2023), new waves could emerge at any time, considering the emergence of new variants that may not be controlled by the immune system [1,2] . Even now, several candidate vaccines are still being developed, in an attempt to meet new COVID-19 challenges: since the majority of the population was vaccinated, new vaccines will act as boosters; there is a lack of vaccine for specific populations, as newborns; and long-lasting response has not been achieved with current vaccines [2,3,4].
The development of neutralizing antibodies against COVID-19 has been considered an established correlate of protection [1,4]. Clinical and epidemiological studies support that prior exposure to SARS-CoV-2 by infection and/or vaccination is likely to attenuate the severity of subsequent infections [1]. Therefore, we must emphasize the importance of cross-reactive protection, considering the emergence of highly transmissible variants, such as Omicron [2,4].
It is relevant to study the presence of an immune response against SARS-CoV-2 during pregnancy and maternal-fetal transmission of antibodies, in order to protect newborns [5]. That is highly important considering the suceptibility of infants to COVID-19 and, although most pediatric cases are mild, children still should be protected [6]. Thus, vaccinating pregnant women reduces the risk of severe COVID-19. The literature describes that pregnant women might have an increased risk for respiratory infections and severe COVID-19. Along with its coagulopathy issues, the infection poses an crucial risk for miscarriage, pre-eclampsia and other problems [7]. Most evidence concludes that substantial transplacental and lactational antibody transfer to infants confers protection when the newborn is exposed to disease [8]. Overall, immunization of pregnant women is considered one of the best strategies to help combat infectious diseases, bringing us closer to achieving herd immunity [8].
Aluminum hydroxide (AH) was the first adjuvant to be licensed and it is widely used, supporting several vaccines so far [9]. Outer membrane vesicles (OMV) are vesicles naturally released after the blebbing of the complex membrane from Gram negative bacteria, as Neisseria meningitidis. Several Pathogen Associated Molecular Patterns (PAMPs) are carried in this process, which makes them promising immune activators [10]. Our laboratory already studied the mixture OMV/AH as an antigen for meningococcal disease immunization [11], and as adjuvant for the RBD from SARS-CoV-2 [12,13].
Dimethyldioctadecylammonium bromide (DDA) is a cationic adjuvant with a spherical lipid bilayer structure that favors interaction with APCs and slow release of antigens, being an excellent antigen nanocarrier [14], while Saponin (Sap) is an anionic molecule isolated from the bark of Quillaja saponaria, capable of stimulating the immune response [15]. The literature describes that these adjuvants are potent inducers of cellular immune responses, polarized into a Th1 profile [14,15]. Our group studied DDA and Sap before, as adjuvants for Neisseria meningitidis, and found promising results, agreeing with the literature [16,17,18].
Moreover, we observed that DDA in bilayer fragments (DDA-BF) was a promising adjuvant to enhance maternal-fetal transference of antibodies to the offspring [17], and that preparations containing OMV/AH [11], or adjuvanted by DDA-BF or Sap [16] supported long-term immune responses, from adult to old age.
In view of this, we studied an antigenic preparation using different adjuvants, aiming to induce a functional, durable and robust humoral and cellular response in mice. The first part of this study, published previously [19], showed that the same adjuvant contributed to the functionality of the humoral response and led to passive transfer of antibodies to the offspring. We used recombinant Receptor binding domain (RBD), due to its high immunogenicity and potential to induce neutralizing antibodies [4], associated with the following adjuvants mixtures: dimethyldioctadecylammonium bromide and saponin (DDA/Sap) or Outer membrane vesicles and aluminum hydroxide (OMV/AH). We already published the results concerning IgG response and its functionality by avidity and neutralization against Wuhan strain [19]. In this manuscript, we continued the study, assessing IgM levels and avidity; IgG and IgM response one year after the immunization; neutralization against Omicron strain and memory-cellular response by ELISpot. Moreover, we analyzed the kinetics of IgG transferred to the offspring for 45 days.
2. Materials and Methods
Recombinant Antigens
RBD protein was produced using the plasmid provided by Dr. Florian Krammer, from Mount Sinai Hospital (New York, NY, USA), to Dr. Carlos Prudencio, from Instituto Adolfo Lutz (São Paulo, SP, Brazil). The protein was expressed in eukaryote system (HEK 293F cells and ExpiFectamine 293 Transfection Kit, Thermo Fisher ScientificTM, Waltham, MA, USA) and purified in a fast protein liquid chromatography system (ÄKTATM Pure, Cytiva Life Sciences, Marlborough, MA, USA), as described before [20,21].
Adjuvant mixtures
Both DDA and Sap were purchased (Sigma-Aldrich), prepared in 0.01 mM sterile saline solution for a final concentration of 1 µg of DDA/1 µg of Sap per dose, and filtered using 0.45 µm membrane as described in our previous manuscript [19]. These concentrations were safe to use according to the hemolysis test that we reported before [19].
Aluminum hydroxide was also purchased (Rehydragel HPA, Reheis Chemicals). OMVs from N. meningitidis C:2a:P1.5 were isolated as described by De Gaspari and Zollinger (2001) [22], detoxified from LPS and used previously in our laboratory [11]. Final concentrations were 10 µg of OMVs and 0.1 mM of Aluminum hydroxide [19].
Antigenic preparations
As described before, the antigenic preparations were diluted in sterile 0.01 mM saline solution and consisted in the mixture of antigen and adjuvants, which were left to interact for 1 hour before the inoculation. The antigenic preparations were: 0.5 μg RBD + 1 μg Sap + 1 μg DDA (the main focus of this study); 3 μg RBD + 10 μg OMVs of N. meningitidis C:2a:P1.5 + 0.1 mM AH (an antigenic preparation used before [12,13], for comparison); 3 μg RBD; and 1 μg Sap + 1 μg DDA. An extra control group was used, consisting of naïve mice [19].
Animals, immunization and collections
Adult, female, Swiss mice (Mus musculus) were obtained from the Animal Experimentation Facility of the Instituto Adolfo Lutz and the experiments were approved by the ethics committees of the Instituto Adolfo Lutz (CEUA/IAL number 02/2022).
Mice received two intramuscular (IM) doses of the antigenic preparations 21 days apart. Blood collections were performed before (pre-immune control), 21, 47, 176 and 368 days after the immunization [19].
Females immunized with RBD+DDA/Sap and DDA/Sap were put to mating after they received the second IM dose [19]. The kinetics of IgG transference was assessed in the offspring at 15, 30 and 45 days after birth.
Detailed descriptions of antigen and adjuvant preparations and immunization calendar are in our previous manuscript [19].
ELISA
High-binding 96-well polystyrene plates (Costar) were coated with 1 μg/ml of RBD in 0.1 M carbonate bicarbonate buffer (pH 9.5), incubated overnight at 4 °C and blocked with 5% skim milk (La Serenissima) for 2 hours at 37 °C. Serum samples diluted at 1:100 were incubated overnight at 4 °C. Secondary antibodies were incubated according to the following concentrations: anti-mouse IgG-γ chain at 1:10,000; anti-mouse IgM-µ chain at 1:20,000 (both from Kirkegaard & Perry Laboratories), anti-mouse IgG2a, IgG2b or IgG3-peroxidase at 1:10,000 (Aviva Systems Biology); or anti-mouse-IgG1-biotin at 1:5,000 (Aviva Systems Biology), for 2 hours at 37ºC. Streptavidin-peroxidase (Zymed) was incubated for 1 hour at 37 °C at a 1:2,000 dilution in IgG1 plates [19]. Finally, we added 3,3,5,5-Tetramethylbenzidine (TMB)(Sigma-Aldrich) in IgG plates and isotypes and O-phenylenediamine dihydrochloride (OPD)(Sigma-Aldrich) in IgM plates. Both substrates were incubated for 20 minutes at 37 °C and we stopped the reactions using 1 N H2SO4 [19].
Avidity index (AI)
The avidity index was studied following the ELISA as described above, but adding 1.5 M potassium thiocyanate (KSCN) for 20 minutes at room temperature (RT) (20-25 °C), after serum incubation [13,23]. The IA index was measured by the following ratio: sample OD with KSCN/sample OD without KSCN. The AI was considered low if < 30%, intermediate if 30%-49%, and high if ≥ 50% [24].
Neutralization Index
We used the commercial assay cPass SARS-CoV-2 Neutralization Antibody Detection (GenScript) for Omicron variant (BA1) to measure the neutralizing index (NI) of antibodies. The methodology followed the manufacturer’s instructions. Neutralization indexes ≥ 20% were considered neutralizing [13]. The kits were kindly donated by NL Diagnostics.
Immunoblotting
To verify if the immune sera recognized the RBD in denatured form, we conducted Immunoblotting, as described before [19]. Briefly, we proceeded an electrophoresis in denaturing conditions using the RBD protein. Afterwards, the protein was transferred to 0.45 µm nitrocellulose membranes (Sigma-Aldrich), so that each strip would contain 1 µg of antigen. The strips were blocked with 5% skim milk for 2 hours at RT. Pooled serum sample, collected 368 days after the immunization, diluted at 1:100 was incubated overnight at 4 °C. Anti-IgG-γ chain (Kirkegaard & Perry Laboratories) diluted at 1:5,000 was incubated for 2 hours at RT. The reaction was developed using 4-chloro-1-naphthol (Sigma-Aldrich) and stopped with distilled water [19].
Enzyme-linked ImmunoSpot (ELISpot)
To assess immunological memory, ELISpot was performed with a pool of splenocytes from two mice per group, using Mouse INF-Y or IL-4 ELISpot PLUS (MabTech) kits. These cytokines were chosen to indicate polarization of the Th1/Th2 profile [25] and due to the importance of the Th1 response, which supports the control of viral replication, and the relevance of Th2 to support humoral response and production of neutralizing antibodies [26].
Aiming to test if the immunologic memory would last until mice were considered old, we performed ELISpot 465 days after the immunization, when mice were approaching 18 months of age (32). Unfortunately, mice from the group immunized using RBD+OMV/AH did not survive until this time and we could only analyze RBD+DDA/Sap and antigen and adjuvant controls (RBD and DDA/Sap, respectively).
The animals were anaesthetized (Ketamine 10 mg/kg and Xylazine 10 mg/kg) and euthanized by cervical dislocation. Spleens were collected and harvested to release the cells in ISCOVE’S medium (Sigma Aldrich) supplemented with 10% inactivated fetal bovine serum (Cripion, SP, Brazil), 1% antibiotics/antimycotics (penicillin G 10,000 U/mL, streptomycin sulfate 10,000 μg/mL and amphotericin B 25 μg/mL) (Sigma-Aldrich), Glutamine 20 mM (Sigma Aldrich), Hepes 9 mM (Sigma-Aldrich) and β-mercaptoethanol 5 × 10−5 M (Sigma-Aldrich). The debris were left to decant for five minutes, then, the supernatant was transferred to a new tube and centrifuged at 2,000 rpm for five minutes (Hettich Holding GmbH & Co, Germany). The cells were resuspended in culture medium and the concentration was adjusted to 1 × 108 cells, as indicated by the manufacturer.
The 96-well polyvinylidene fluoride (PVDF) plates were supplied coated with capture antibodies anti-IL-4 (clone 11B11) or anti-IFN-Y (clone AN18) (Mabtech). The plates were blocked with ISCOVES-fetal bovine serum 10% solution at RT for thirty minutes and the cell suspensions were plated and stimulated with 0.2 μg of RBD per well. Positive control wells received nonspecific stimulation of 1 μg of Concanavalin A (ConA) (Sigma-Aldrich) per well and negative control wells did not receive stimuli. The cells were incubated for 24 hours at 37oC with 5% CO2.
Afterwards, the cell suspension was removed and the plates were incubated with ethylenediaminetetraacetic acid (EDTA) 1 mM solution for 15 minutes at RT to remove cell debris from the membrane. Biotinylated anti-IL-4 or anti-IFN-Y antibodies at 1 μg/mL concentration were incubated for 2 hours, followed by streptavidin-alkaline phosphatase, incubated at 1:1,000 for 1 hour, both at RT. The plates were washed five times with PBS between each step. Finally, the BCIP/NBT substrate was incubated at RT and protected from light for 1 hour until spots were formed. The reaction was stopped by washing the plates with distilled water. After drying in the shade for 36 hours the spots were counted automatically (Aid ELISpot Reader, Germany) and the count was considered the simple average of the duplicates.
Statistical analysis
Multiple comparisons were performed using Kruskal-Wallis followed by Dunn’s post-test, in GraphPad Prism v. 8 (Graphpad Software, Inc. Software, Inc., La Jolla, CA, USA). P values ≤ 0.05 were considered significant.
3. Results
IgM antibodies and avidity
Figure 1 presents the IgM titers in serum of immunized mice, collected after different time points, while shows the avidity of such antibodies. Since the control groups (adjuvant alone and naïve) do not present specific antibodies, sera collected from these mice sera were not adequate to perform avidity assay. Because the RBD group did not present an optical density higher than those controls, it was not appropriate for the avidity test as well.
Even though we could not verify a statistical difference in IgM levels, it was higher on days 21 and 47, at the beginning of immunization and following the immunization doses, approximately 5 times higher than controls. After one dose only, the antibodies presented low avidity, but it increased to high (RBD+DDA/Sap) or intermediate (RBD+OMV/AH) after the booster. Interestingly, the AI of RBD+OMV/AH continued to increase in later time points (176 and 368 days), being considered high; while RBD+DDA/Sap kept a high avidity in day 176 and decayed for intermediate at day 368.

Table 1.
Avidity index (%) of IgM antibodies on different days after immunization.
| 21 days | 47 days | 176 days | 368 days | |
|---|---|---|---|---|
| RBD+DDA/Sap | 28.77 | 60.86 | 71.70 | 43.72 |
| RBD+OMV/AH | 20.81 | 47.05 | 77.10 | 64.72 |
Avidity is classified as low if < 30%, intermediate if between 30% and 49% and high if ≥ 50% (24). AH: aluminum hydroxide. DDA: dioctadecyldimethylammonium bromide. OMV: outer membrane vesicles. RBD: receptor binding domain. Sap: Saponin.
IgG persistence and avidity
In our previous study, we assessed the titers and avidity of IgG, as well as its subclasses (IgG1, IgG2a, IgG2b and IgG3), which showed that only adjuvanted groups (RBD+DDA/Sap and RBD+OMV/AH) produced high levels of IgG, the peak of the response being on day 47 and the same groups maintained higher levels until day 176 [19]. IgG1, IgG2a and IgG2b were the predominant subclasses of RBD+OMV/AH group and all subclasses, including IgG3, were present on serum of RBD+DDA/Sap group [19]. In this study, we followed the mice until 368 days after the immunization and we could verify the persistence of IgG antibodies (), as well as its avidity and avidity of the subclasses (). We can observe that groups immunized with adjuvants had higher IgG levels than controls. The avidity of RBD+DDA/Sap was considered high, and studying the isotypes separately, only IgG3 presented low avidity. RBD+OMV/AH presented intermediate avidity and, likewise, only IgG3 was considered low.
Table 2.
Mean ELISA OD of IgG antibodies in 368 days in a pool of serum from adult mice (n = 6 animals in each group).
Table 2.
Mean ELISA OD of IgG antibodies in 368 days in a pool of serum from adult mice (n = 6 animals in each group).
| O.D (450nm) | IgG—Day 368 | ||||
| RBD+DDA/Sap | RBD+OMV/AH | RBD | DDA/Sap | Naive | |
| 1.792 | 1.161 | 0.106 | 0.054 | 0.0795 | |
AH: aluminum hydroxide. DDA: dioctadecyldimethylammonium bromide. OMV: outer membrane vesicles of the outer membrane RBD: receptor binding domain. Sap: Saponin as controls.
Table 3.
Avidity index (%) of IgG antibodies and its isotypes on sera collected 368 days after the immunization.
Table 3.
Avidity index (%) of IgG antibodies and its isotypes on sera collected 368 days after the immunization.
| IgG | IgG1 | IgG2a | IgG2b | IgG3 | |
|---|---|---|---|---|---|
| RBD+DDA/Sap | 83.2 | 63.9 | 56.7 | 48.2 | 10.24 |
| RBD+OMV/AH | 43.72 | 62.67 | 37.5 | 44.49 | 12.12 |
Avidity is classified as low if < 30%, intermediate if between 30% and 49% and high if ≥ 50% (24). AH: aluminum hydroxide. DDA: dioctadecyldimethylammonium bromide. OMV: outer membrane vesicles of the outer membrane RBD: receptor binding domain. Sap: Saponin.
In addition, we verified that IgG antibodies from the group RBD+DDA/Sap, the main focus of this manuscript, collected 368 days after the immunization, recognized RBD antigen in denatured form, in Immunoblotting (Figure 2). The controls did not present any band in the same assay.
Neutralization against Omicron variant
According to our previous data, the immunization using RBD+DDA/Sap presented the better neutralization against Wuhan strain, especially on day 47, in the peak of the immune response [19]. In this study, we assessed the neutralization against the Omicron variant, which is presented in as neutralizing index (NI). The groups immunized with RBD alone, DDA/Sap and Naïve controls were not considered positive.
Table 4.
Neutralization index (%) of immunized groups against Omicron variant at different time points.
Table 4.
Neutralization index (%) of immunized groups against Omicron variant at different time points.
| Pre-immune | 21 days | 47 days | 176 days | |
|---|---|---|---|---|
| RBD+DDA/Sap | 14.78 | 18.62 | 21.48 | 20.37 |
| RBD+OMV/AH | 12.26 | 21.95 | 27.60 | 26.90 |
AH: aluminum hydroxide. DDA: dioctadecyldimethylammonium bromide. OMV: outer membrane vesicles of the outer membrane RBD: receptor binding domain. Sap: Saponin.
Kinetics of IgG transferred to offspring
The females immunized with RBD+DDA/Sap were put to mate and we studied the IgG transferred to the offspring. Our previous article described that transferred antibodies, collected at day 18, presented neutralizing activity despite its low avidity and were mainly IgG2a and IgG2b [19]. Here, we present the kinetics of maternal-fetal transference from day 15 to 45 (Figure 3) and its avidity (). As observed, IgG levels were higher on day 15 and started to decrease; however, we could still detect antibodies around day 30. We verified that all IgG isotypes were present at the sooner time point, day 15. However, IgG2a and IgG2b were no longer present on day 30 and, at day 45, the overall response was decreased and probably composed of only IgG1.
Table 5.
Avidity index (AI) (%) of IgG and its isotypes transferred to offspring of RBD+DDA/Sap females, in sera collected different days after birth.
Table 5.
Avidity index (AI) (%) of IgG and its isotypes transferred to offspring of RBD+DDA/Sap females, in sera collected different days after birth.
| Days after birth | |||
|---|---|---|---|
| 15 | 30 | 45 | |
| IgG | 56.06 | 39.48 | 31.57 |
| IgG1 | 66.34 | 26.54 | 26.77 |
| IgG2a | 62.90 | NR | NR |
| IgG2b | 62.69 | NR | NR |
| IgG3 | 68.69 | 41,75 | NR |
Avidity is classified as low if < 30%, intermediate between 30% and 49% and high if ≥ 50% (24). NR: non-reagent.
IL-4 and IFN-Υ secretion
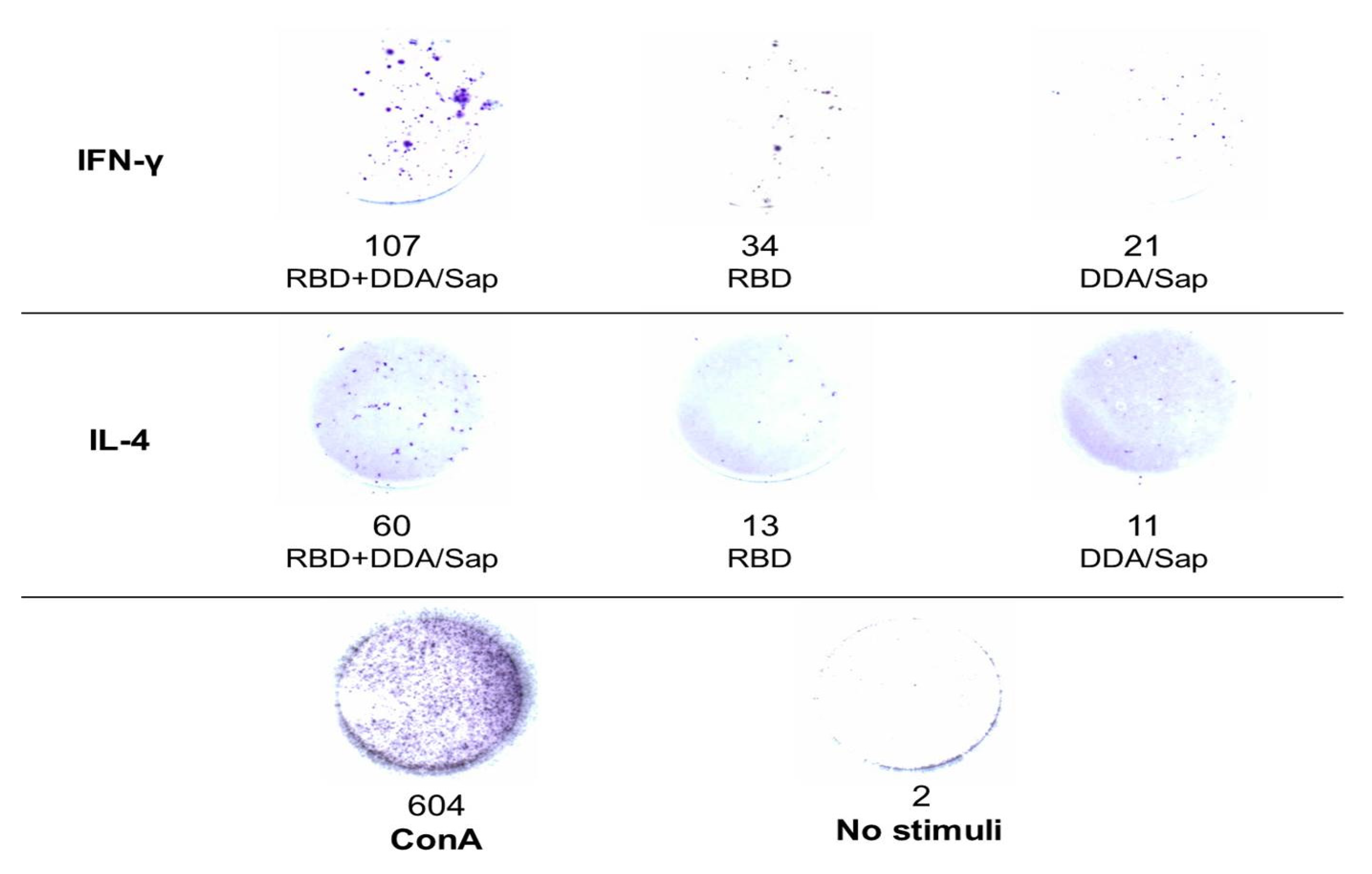
We verified IL-4 and IFN-γ secretion of splenocytes collected 465 days after the immunization by ELISpot. As mentioned in Material and Methods, to analyze immunologic memory, we decided to perform ELISpot 465 days after immunization and, unfortunately, mice from the group RBD+OMV/AH did not survive until this time point. Figure 4 shows the quantification of spots of RBD+DDA/Sap, RBD alone and DDA/Sap control groups, as well as representative pictures of the wells from each quantification. In the line below, the control wells, with cells without stimuli (negative control) or stimulated with ConA (positive control).
4. Discussion
IgM is the first class of antibody produced after antigenic stimulation, followed by the switch for other classes. As expected, the higher levels of IgM were found at day 21, after the first dose, and day 47, after the second dose, showing that the immunization activated the immune system [26]. Taken together with our previous data [19], IgG antibodies were also induced and maintained for longer periods than IgM, which is expected. Nevertheless, IgM antibodies are relevant in SARS-CoV-2 infection. Studying convalescent plasma, Kober et al. (2022) found that IgG3 and IgM strongly correlated with neutralization, even though its abundance is more limited than other antibodies [27] and, depleting plasma from immunoglobulin A, G or M, Gasser et al. (2021) found that IgM depletion caused a 5.5 fold decrease in neutralization, the major loss when the three classes were compared [28].
The persistence of antibodies is an important issue for COVID-19 prevention. It is well documented that the antibodies induced after natural infection decay within 6 to 8 months [29]. When vaccines started being administered, vaccine-antibodies were found to be more functional than infection-antibodies, but a similar scenario was observed considering their persistence, with antibodies’ levels persisting from 5 months to one year after immunization [30,31]. In the first part of our study, we assessed persistence of IgG until 176 days after the immunization [19] and here we describe that it persisted until 368 days. When mice age is considered, we see that IgG persisted from adult to middle-to-old age [32]. We attribute this result to the adjuvants used, since the literature shows persistence of antibodies when using adjuvants: Th1 adjuvants, as Saponin QS-1, and a mixture of Saponin QS-1, Cytosine-phosphate-Guanine (CpG) and Monophosphoryl Lipid A (MPL) induced a persistent humoral response against P. falciparum until day 180 [33]; in another study, an anthrax vaccine adjuvanted by CpG maintained protective antibodies titers up to one year in mice [34]. Studies of our group found similar results for Neisseria meningitidis, with a humoral response persisting from adult to old age in mice [11,16].
Avidity is an important functional parameter, specially for SARS-CoV-2 [35]. We verified that IgM had low avidity after the first dose of the immunization, which increased until intermediate-to-high avidity in the later time points. In contrast to our study, immunization of rabbits against Influenza led to IgM of high avidity early after the immunization and, in later time points, it remained stable [36]. Similarly, our results point that IgM avidity maintained intermediate-to-high levels since day 47 and until middle-to-old age. The pentameric structure of IgM is an important contribution to neutralization, since RBD-ACE-2 binding presents high affinity, thus requiring high avidity to block this reaction [35,37].
RBD, the antigen we used for immunization, is the main region of the Spike protein that undergoes mutations, especially considering the Omicron variant [4,38,39]. However, we observed some neutralizing activity in the group immunized with RBD+OMV/AH. Monoclonal IgM derived from B cells were potent cross-neutralizers [40], which might have contributed to that, although the higher neutralization index was not found exclusively at the peak of IgM response. In addition, adjuvants enhance antigen presentation and may confer a more broad epitope recognition, as observed previously for HIV [41], Dengue virus [42] and Influenza H5N1 [43], for example. Here, the main cross-neutralization occurred in the group adjuvanted by an OMV/AH mixture. We could not find similar results using OMVs for COVID-19, but OMVs provided a broader and more functional peptide recognition as adjuvant for Chlamydia [44], while Alum did the same for HIV-1 [45].
Cytokine secretion not only showed the presence of memory cells in the spleen, activated by RBD stimuli, but corroborated IgG isotypes to point to the pattern of the immune response. Based on our previous data [19], we suggested a mixed Th1/Th2 profile, due the presence of IgG2a, induced by IL-2 and IFN-γ, Th1 cytokines; and IgG1, induced by IL-4, a Th2 cytokine [25]. In ELISpot, RBD+DDA/Sap group secreted both IL-4 and IFN-γ, although the secretion of IFN-γ was higher. DDA was firstly described as a Th1-polarizing adjuvant, as confirmed in studies for different pathogens [46,47,48]; and the same Th1 profile was observed for Saponin [15]. However, some studies also found a mixed Th1/Th2 profile: Klinguer-Hamour et al. (2002) had promising results using DDA to drive the response to a Syncytial-respiratory virus away from a non-protective Th2 phenotype to a mixed Th1/Th2 pattern [49]; and a Saponin contributed to a robust anti-Influenza response, characterized by high levels of IgG1, IgG2a and IgG2b, as well as secretion of IL-2 and IFN-γ [50]. When COVID-19 vaccines are considered, a recombinant RBD associated with DDA-TDB adjuvant presented strong humoral and cellular response [51], the same was observed for a RBD/Nucleocapsid associated with Saponin [52]. Likewise, a potent humoral/cellular response was observed for Saponin as adjuvant to the Spike protein of MERS-CoV, which present similarities with SARS-CoV-2 [53].
Maternal immunity obtained passively hinders the active immunization of newborns. In addition, an immature immune system contributes to weak, Th2-polarized immunity. This state of immunity in early life sustains endemic infections in humans and continuous reinfections. Endemic infections in young people occur preferentially when the immune system is still functionally immature and when low levels of maternal antibodies are no longer protective, but still block protective immune responses. Vaccines that overcome these problems would have strong positive effects on health. A newborn’s Th2 tendency is mediated by high levels of progesterone and Th2 cytokines produced at the maternal-fetal interface. The activity of the innate system is enhanced in the mother during the prenatal period, which certainly has an effect on the offspring. Newborn mice, as young as 2 days old, can be immunized by Senday virus proteins as an example and direct the response towards Th1 and Th2 depending on the adjuvant. The same pattern has been observed in several species. In conclusion, the Th2 tendency that prevails around the parturition can be overcome by adequate treatments, allowing effective vaccination and protective immunity in the newborn [54].
COVID-19 vaccines have been administred to children showing good safety in most clinical studies [55,56]. Epidemiological evidence suggests that babies born from mothers who received two doses of the mRNA vaccine during pregnancy had a 61% lower risk of hospitalization for SARS-CoV-2 in the first six months of life. On the other hand, a study carried out in Israel found a higher risk of infection among babies aged 0 to 1 year than among older children [57]. This result is understandable because babies’ immune systems are not fully developed and are not able to produce enough immunoglobulins [58].
In addition, high levels of the SARS-CoV-2 virus in asymptomatic children can allow the infection to be transmitted to family members who are in close contact with them [56,59]. It is therefore essential to vaccinate children over 6 months of age. Studies have shown that participants aged between 6 and 23 months have a strong immunological response after three doses. This also was observed in the group of people aged between 16 and 25 who received two doses of the BNT162b2 vaccine [60,61].
In our study IgG antibodies crossed the placenta in mice, and that is likely to play a crucial role in protecting the offspring by the functionality of the transferred antibodies evaluated by the avidity of the isotypes. This transfer occurs during gestation, when the mother provides maternal IgG antibodies to the developing fetus via the placenta [54,62]. These antibodies are fundamental to the offspring’s immunity, as their immune systems are still developing. In humans, a similar process takes place, where the mother transfers IgG antibodies to the fetus via the placenta, providing temporary protection against infections [63]. This is particularly important in the first few months of life, when the baby’s immune system is less effective. However, this work represents an experimental part and further studies in humans will be necessary. The avidity property of the antibody increases over the course of the immune response, so it is suggested that avidity is directly proportional to the time of exposure to the antigen.
As avidity is a relevant functional parameter, the time of vaccination according to pregnancy stage may interfere in the results. A recent study found that avidity against pertussis antigen was independent of maternal vaccination during the second or third trimester [64].
Our data pointed to overall high avidity soon after birth, which decreased for a long time. It should be considered that this is a different pathogen and an experimental model, however, our mice did not receive any booster during pregnancy. It might be worth investigating if a booster dose during pregnancy could be interesting to improve antibody avidity.
In our previous article, blood was collected from the offspring 18 days after birth, and the antibodies passively transferred to the offspring of the RBD+DDA/Sap group were neutralized [19]. The offspring showed intermediate IgG avidity. Interestingly, the offspring showed a neutralizing index even when avidity was low [19]. Therefore, the studies were repeated in this work and a follow-up kinetics of maternal-fetal transfer was carried out, thus showing their persistence [19].
As we observed in this study, the antibodies persisted until 30 days after birth and, around 45 days, the last time point that we monitored, IgG levels decreased. Based on previous works it is already known that the antibodies can last up to 5 to 8 weeks [17,54].
It is very important to note that nanograms of the SARS-CoV-2 Spike protein administered with exosomes have recently been shown to induce potent neutralization of Delta and Omicron variants (BA.1,BA.5) [65].
Another point to emphasize is that maternal vaccination is important, as the work shows that IgG antibodies for homologous variants of SARS-CoV-2 have been transferred to neonates [66]. On the other hand, in our study we were able to observe IgG with good avidity when we used the Wuhan variant, homologous to the immunization. Even though we could not check cross-reactivity against Omicron variant in the offspring sera, their mothers presented IgG of good avidity against the homologous strain and cross-reactivity for the Omicron variant (BA.1).
5. Conclusions
Our study showed that the antigenic preparations RBD+OMV/AH and RBD+DDA/Sap improved the humoral response, inducing IgM antibodies of intermediate to high avidity; and an IgM and IgG response which lasted for one year in mice. RBD+DDA/Sap also induced cellular memory and cytokine response, mostly of IFN-γ, an important cytokine for antiviral response. Finally, IgG transferred from females immunized with RBD+DDA/Sap lasted for almost 45 days - nonetheless, until the offspring was considered young.
Studying SARS-CoV-2 vaccines is still relevant, as the scenario has changed and new vaccines must be adapted to respond to the current challenges – protection against the new variants, long-lasting immunity and maternal/fetal and pediatric immunization.
Author Contributions
Conceptualization, G.G.L. and E.D.G.; investigation, G.G.L., A.I.P and E.D.G.; resources, G.G.L., H.H.M.C., C.R.P. and E.D.G.; writing—original draft preparation, G.G.L., A.I.P. and E.D.G.; writing—review and editing, G.G.L and E.D.G.; supervision, E.D.G.; funding acquisition, E.D.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo [FAPESP] under grant number 18/04202-0; Conselho Nacional de Desenvolvimento Científico e Tecnológico [CNPq] under grant number 131308/2021-1 and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior [CAPES].
Institutional Review Board Statement
The entire process of mice manipulation complies with the recommendations of the Brazilian Society of Laboratory Animal Science (SBCAL/COBEA) and was approved by the ethics committees of the Adolfo Lutz Institute (CEUA/IAL number 02/2022).
Informed Consent Statement
Not applicable, study did not use humans.
Data Availability Statement
The manuscript is an original article and we can make the data available upon reasonable request.
Acknowledgments
We would like to thank the NL-Diagnostica for donation of the cPassTM SARS-CoV-2 Neutralization kit. We also would like to thank the receptor-binding domain (RBD) antigen that was kindly donated by Dr. Florian Krammer, from Icahn School of Medicine, Mount Sinai, Nova York, NY, EUA.
Conflicts of Interest
The authors declared no potential conflicts of interest with respect to the publication of this article.
References
- Koutsakos, M.; Ellebedy, A.H. Immunological imprinting: Understanding COVID-19. Immunity 2023, 56, 909–913. [Google Scholar] [CrossRef]
- Konyak, B.M.; Sharma, M.; Kharia, S.; Pandey, R.P.; Chang, C.-M. A Systematic Review on the Emergence of Omicron Variant and Recent Advancement in Therapies. Vaccines 2022, 10, 1468. [Google Scholar] [CrossRef]
- Chong, W.C.; Chellappan, D.K.; Shukla, S.D.; Peterson, G.M.; Patel, R.P.; Jha, N.K.; Eri, R.D.; Dua, K.; Tambuwala, M.M.; Shastri, M.D. An Appraisal of the Current Scenario in Vaccine Research for COVID-19. Viruses 2021, 13, 1397. [Google Scholar] [CrossRef]
- Krammer, F. The role of vaccines in the COVID-19 pandemic: what have we learned? Semin. Immunopathol. 2023, 1–18. [Google Scholar] [CrossRef]
- Marshall, C.L.B.; Kaplowitz, E.; Ibroci, E.; Chung, K.; Gigase, F.A.J.M.; Lieber, M.L.; Graziani, M.; Ohrn, S.B.; Lynch, J.; Castro, J.B.; et al. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Antibody Titer Levels in Pregnant Individuals After Infection, Vaccination, or Both. Obstetrics & Gynecology 2023, 141, 1199–1202. [Google Scholar] [CrossRef]
- Malcangi, G.; Inchingolo, A.D.; Inchingolo, A.M.; Piras, F.; Settanni, V.; Garofoli, G.; Palmieri, G.; Ceci, S.; Patano, A.; Mancini, A.; et al. COVID-19 Infection in Children and Infants: Current Status on Therapies and Vaccines. Children 2022, 9, 249. [Google Scholar] [CrossRef]
- Agolli, A.; Agolli, O.; Velazco, D.F.S.; Ahammed, R.; Patel, M.; Cardona-Guzman, J.; Garimella, R.; Rummaneethorn, N.; Bista, S.; Abreu, R.; et al. Fetal Complications in COVID-19 Infected Pregnant Woman: A Systematic Review and Meta-Analysis. Avicenna J. Med. 2021, 11, 200–209. [Google Scholar] [CrossRef]
- Jorgensen, S.C.J.; Burry, L.; Tabbara, N. Role of maternal COVID-19 vaccination in providing immunological protection to the newborn. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2021, 42, 58–70. [Google Scholar] [CrossRef]
- He, P.; Zou, Y.; Hu, Z. Advances in aluminum hydroxide-based adjuvant research and its mechanism. Hum. Vaccines Immunother. 2015, 11, 477–488. [Google Scholar] [CrossRef]
- Tan, K.; Li, R.; Huang, X.; Liu, Q. Outer Membrane Vesicles: Current Status and Future Direction of These Novel Vaccine Adjuvants. Front. Microbiol. 2018, 9, 783. [Google Scholar] [CrossRef]
- Portilho, A.I.; Correa, V.A.; Cirqueira, C.d.S.; De Gaspari, E. Intranasal and Intramuscular Immunization with Outer Membrane Vesicles from Serogroup C Meningococci Induced Functional Antibodies and Immunologic Memory. Immunol. Investig. 2022, 51, 2066–2085. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, E.B.; Prudencio, C.R.; De Gaspari, E. Experimental studies using OMV in a new platform of SARS-CoV-2 vaccines. Hum. Vaccines Immunother. 2021, 17, 2965–2968. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, E.B.; De Gaspari, E. Avidity assay to test functionality of anti-SARS-Cov-2 antibodies. Vaccine 2021, 39, 1473–1475. [Google Scholar] [CrossRef] [PubMed]
- Lincopan, N.; Espíndola, N.M.; Vaz, A.J.; da Costa, M.H.B.; Faquim-Mauro, E.; Carmona-Ribeiro, A.M. Novel immunoadjuvants based on cationic lipid: Preparation, characterization and activity in vivo. Vaccine 2009, 27, 5760–71. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.-X.; Xie, Y.; Ye, Y.-P. Advances in saponin-based adjuvants. Vaccine 2009, 27, 1787–1796. [Google Scholar] [CrossRef]
- Correa, V.A.; Portilho, A.I.; De Gaspari, E. Immunological Effects of Dimethyldioctadecylammonium Bromide and Saponin as Adjuvants for Outer Membrane Vesicles from Neisseria meningitidis. Diseases 2022, 10, 46. [Google Scholar] [CrossRef]
- De Almeida, A.F.; De Gaspari, E. Dioctadecyldimethylammonium bromide (DODAB-BF) as a new adjuvant for maternal-fetal immunization in mice against Neisseria meningitidis: evaluation of humoral response. Pathog. Dis. 2017, 76. [Google Scholar] [CrossRef]
- Santos, F.A.d.O.; Lincopan, N.; De Gaspari, E. Evaluation of intranasal and subcutaneous route of immunization in neonatal mice using DODAB-BF as adjuvant with outer membrane vesicles of Neisseria meningitis B. Immunobiology 2018, 223, 750–760. [Google Scholar] [CrossRef]
- Lima, G.G.; Portilho, A.I.; De Gaspari, E. Adjuvants to increase immunogenicity of SARS-CoV-2 RBD and support maternal–fetal transference of antibodies in mice. Pathog. Dis. 2022, 80. [Google Scholar] [CrossRef]
- Stadlbauer, D.; Amanat, F.; Chromikova, V.; Jiang, K.; Strohmeier, S.; Arunkumar, G.A.; Tan, J.; Bhavsar, D.; Capuano, C.; Kirkpatrick, E.; et al. SARS-CoV-2 Seroconversion in Humans: A Detailed Protocol for a Serological Assay, Antigen Production, and Test Setup. Curr. Protoc. Microbiol. 2020, 57, e100. [Google Scholar] [CrossRef]
- da Costa, H.H.M.; Orts, D.J.B.; Moura, A.D.; Duarte-Neto, A.N.; Cirqueira, C.S.; Réssio, R.A.; Kanamura, C.T.; Miguita, K.; Ferreira, J.E.; Santos, R.T.M.; et al. RBD and Spike DNA-Based Immunization in Rabbits Elicited IgG Avidity Maturation and High Neutralizing Antibody Responses against SARS-CoV-2. Viruses 2023, 15, 555. [Google Scholar] [CrossRef] [PubMed]
- De Gaspari, E.N.; Zollinger, W.D. Expression of class 5 antigens by meningococcal strains obtained from patients in Brazil and evaluation of two new monoclonal antibodies. Braz. J. Infect. Dis. 2001, 5, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Granoff, D.M.; Maslanka, S.E.; Carlone, G.M.; Plikaytis, B.D.; Santos, G.F.; Mokatrin, A.; Raff, H.V. A Modified Enzyme-Linked Immunosorbent Assay for Measurement of Antibody Responses to Meningococcal C Polysaccharide That Correlate with Bactericidal Responses. Clin. Diagn. Lab. Immunol. 1998, 5, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Chackerian, B.; Lowy, D.R.; Schiller, J.T. Conjugation of a self-antigen to papillomavirus-like particles allows for efficient induction of protective autoantibodies. J. Clin. Investig. 2001, 108, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T.R.; Coffman, R.L. TH1 and TH2 Cells: Different Patterns of Lymphokine Secretion Lead to Different Functional Properties. Annu. Rev. Immunol. 1989, 7, 145–173. [Google Scholar] [CrossRef] [PubMed]
- Júnior, D.M.; Araújo, J.A.P.; Catelan, T.T.T.; de Souza, A.W.S.; Cruvinel, W.d.M.; Andrade, L.E.C.; da Silva, N.P. Sistema imunitário - parte II: fundamentos da resposta imunológica mediada por linfócitos T e B. Rev. Bras. de Reum. 2010, 50, 552–580. [Google Scholar] [CrossRef]
- Kober, C.; Manni, S.; Wolff, S.; Barnes, T.; Mukherjee, S.; Vogel, T.; Hoenig, L.; Vogel, P.; Hahn, A.; Gerlach, M.; et al. IgG3 and IgM Identified as Key to SARS-CoV-2 Neutralization in Convalescent Plasma Pools. PLOS ONE 2022, 17, e0262162. [Google Scholar] [CrossRef]
- Gasser, R.; Cloutier, M.; Prévost, J.; Fink, C.; Ducas. ; Ding, S.; Dussault, N.; Landry, P.; Tremblay, T.; Laforce-Lavoie, A.; et al. Major role of IgM in the neutralizing activity of convalescent plasma against SARS-CoV-2. Cell Rep. 2021, 34, 108790. [Google Scholar] [CrossRef]
- Knies, A.; Ladage, D.; Braun, R.J.; Kimpel, J.; Schneider, M. Persistence of humoral response upon SARS-CoV-2 infection. Rev. Med Virol. 2021, 32, e2272. [Google Scholar] [CrossRef]
- Alharbi, N.K.; A Al-Tawfiq, J.; Alwehaibe, A.; Alenazi, M.W.; Almasoud, A.; Algaisi, A.; A Alhumaydhi, F.; Hashem, A.M.; Bosaeed, M.; A Alsagaby, S. Persistence of Anti-SARS-CoV-2 Spike IgG Antibodies Following COVID-19 Vaccines. Infect. Drug Resist. 2022, ume 15, 4127–4136. [Google Scholar] [CrossRef]
- Costa, C.; Migliore, E.; Galassi, C.; Scozzari, G.; Ciccone, G.; Coggiola, M.; Pira, E.; Scarmozzino, A.; La Valle, G.; Cassoni, P.; et al. Factors Influencing Level and Persistence of Anti SARS-CoV-2 IgG after BNT162b2 Vaccine: Evidence from a Large Cohort of Healthcare Workers. Vaccines 2022, 10, 474. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Sengupta, P. Men and mice: Relating their ages. Life Sci. 2016, 152, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Pirahmadi, S.; Zakeri, S.; Mehrizi, A.A.; Djadid, N.D.; Raz, A.-A.; Sani, J.J. Combining Monophosphoryl Lipid A (MPL), CpG Oligodeoxynucleotide (ODN), and QS-21 Adjuvants Induces Strong and Persistent Functional Antibodies and T Cell Responses against Cell-Traversal Protein for Ookinetes and Sporozoites (CelTOS) of Plasmodium falciparum in BALB/c Mice. Infect. Immun. 2019, 87. [Google Scholar] [CrossRef]
- Tross, D.; Klinman, D.M. Effect of CpG Oligonucleotides on Vaccine-Induced B Cell Memory. J. Immunol. 2008, 181, 5785–5790. [Google Scholar] [CrossRef]
- Bauer, G. High avidity of vaccine-induced immunoglobulin G against SARS-CoV-2: potential relevance for protective humoral immunity. Explor. Immunol. 2022, 2, 133–156. [Google Scholar] [CrossRef]
- Webster, R.G. The immune response to influenza virus. 3. Changes in the avidity and specificity of early IgM and IgG antibodies.. 1968, 14, 39–52. [Google Scholar]
- Khatri, I.; Staal, F.J.T.; van Dongen, J.J.M. Blocking of the High-Affinity Interaction-Synapse Between SARS-CoV-2 Spike and Human ACE2 Proteins Likely Requires Multiple High-Affinity Antibodies: An Immune Perspective. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Kudriavtsev, A.V.; Vakhrusheva, A.V.; Novoseletsky, V.N.; Bozdaganyan, M.E.; Shaitan, K.V.; Kirpichnikov, M.P.; Sokolova, O.S. Immune Escape Associated with RBD Omicron Mutations and SARS-CoV-2 Evolution Dynamics. Viruses 2022, 14, 1603. [Google Scholar] [CrossRef]
- da Costa, C.H.S.; de Freitas, C.A.B.; Alves, C.N.; Lameira, J. Assessment of mutations on RBD in the Spike protein of SARS-CoV-2 Alpha, Delta and Omicron variants. Sci. Rep. 2022, 12, 1–10. [Google Scholar] [CrossRef]
- Hale, M.; Netland, J.; Chen, Y.; Thouvenel, C.D.; Smith, K.N.; Rich, L.M.; Vanderwall, E.R.; Miranda, M.C.; Eggenberger, J.; Hao, L.; et al. IgM antibodies derived from memory B cells are potent cross-variant neutralizers of SARS-CoV-2. J. Exp. Med. 2022, 219. [Google Scholar] [CrossRef]
- Wu, J.Y.; Gardner, B.H.; I Murphy, C.; Seals, J.R.; Kensil, C.R.; Recchia, J.; A Beltz, G.; Newman, G.W.; Newman, M.J. Saponin adjuvant enhancement of antigen-specific immune responses to an experimental HIV-1 vaccine. J. Immunol. 1992, 148, 1519–1525. [Google Scholar] [CrossRef] [PubMed]
- Maeda, D.L.N.F.; Batista, M.T.; Pereira, L.R.; Cintra, M.d.J.; Amorim, J.H.; Mathias-Santos, C.; Pereira, S.A.; Boscardin, S.B.; Silva, S.d.R.; Faquim-Mauro, E.L.; et al. Adjuvant-Mediated Epitope Specificity and Enhanced Neutralizing Activity of Antibodies Targeting Dengue Virus Envelope Protein. Front. Immunol. 2017, 8, 1175. [Google Scholar] [CrossRef]
- Khurana, S.; Chearwae, W.; Castellino, F.; Manischewitz, J.; King, L.R.; Honorkiewicz, A.; Rock, M.T.; Edwards, K.M.; Del Giudice, G.; Rappuoli, R.; et al. Vaccines with MF59 Adjuvant Expand the Antibody Repertoire to Target Protective Sites of Pandemic Avian H5N1 Influenza Virus. Sci. Transl. Med. 2010, 2, 15ra5–15ra5. [Google Scholar] [CrossRef] [PubMed]
- Bartolini, E.; Ianni, E.; Frigimelica, E.; Petracca, R.; Galli, G.; Scorza, F.B.; Norais, N.; Laera, D.; Giusti, F.; Pierleoni, A.; et al. Recombinant outer membrane vesicles carrying Chlamydia muridarum HtrA induce antibodies that neutralize chlamydial infection in vitro. J. Extracell. Vesicles 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Xiao, Y.; Zhu, M.; Chen, Y.-H. HIV epitope-peptides in aluminum adjuvant induced high levels of epitope-specific antibodies. Int. Immunopharmacol. 2001, 1, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Rosenkrands, I.; Agger, E.M.; Olsen, A.W.; Korsholm, K.S.; Andersen, C.S.; Jensen, K.T.; Andersen, P. Cationic Liposomes Containing Mycobacterial Lipids: a New Powerful Th1 Adjuvant System. Infect. Immun. 2005, 73, 5817–5826. [Google Scholar] [CrossRef]
- Lindblad, E.B.; Elhay, M.J.; Silva, R.; Appelberg, R.; Andersen, P. Adjuvant modulation of immune responses to tuberculosis subunit vaccines. Infect. Immun. 1997, 65, 623–629. [Google Scholar] [CrossRef]
- Mayorga, O.; Muñoz, J.E.; Lincopan, N.; Teixeira, A.F.; Ferreira, L.C.S.; Travassos, L.R.; Taborda, C.P. The role of adjuvants in therapeutic protection against paracoccidioidomycosis after immunization with the P10 peptide. Front. Microbiol. 2012, 3, 154. [Google Scholar] [CrossRef]
- Klinguer-Hamour, C.; Libon, C.; Plotnicky-Gilquin, H.; Bussat, M.-C.; Revy, L.; Nguyen, T.; Bonnefoy, J.-Y.; Corvaı̈a, N.; Beck, A. DDA adjuvant induces a mixed Th1/Th2 immune response when associated with BBG2Na, a respiratory syncytial virus potential vaccine. Vaccine 2002, 20, 2743–2751. [Google Scholar] [CrossRef]
- Hjertner, B.; Bengtsson, T.; Morein, B.; Paulie, S.; Fossum, C. A novel adjuvant G3 induces both Th1 and Th2 related immune responses in mice after immunization with a trivalent inactivated split-virion influenza vaccine. Vaccine 2018, 36, 3340–3344. [Google Scholar] [CrossRef]
- Yu, H.; Worrall, L.J.; Berger, T.; Petric, M.; Lin, B.H.; Vuckovic, M.; Robb, C.S.; Le, Q.; Kenward, C.; Dai, C.; et al. Identification of an Optimized Receptor-Binding Domain Subunit Vaccine against SARS-CoV-2. J. Immunol. 2023, 211, 981–993. [Google Scholar] [CrossRef]
- Mekonnen, D.; Mengist, H.M.; Jin, T. SARS-CoV-2 subunit vaccine adjuvants and their signaling pathways. Expert Rev. Vaccines 2021, 21, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-C.; Algaissi, A.; Lai, C.-C.; Chang, C.-K.; Lin, J.-S.; Wang, Y.-S.; Chang, B.-H.; Chang, Y.-C.; Chen, W.-T.; Fan, Y.-Q.; et al. Subunit vaccines with a saponin-based adjuvant boost humoral and cellular immunity to MERS coronavirus. Vaccine 2023, 41, 3337–3346. [Google Scholar] [CrossRef]
- Morein, B.; Abusugra, I.; Blomqvist, G. Immunity in neonates. Veter- Immunol. Immunopathol. 2002, 87, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Song, Y.; Li, C.; Yang, W.; Ma, Q.; Jiang, Z.; Li, M.; Lian, X.; Jiao, W.; Wang, L.; et al. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine (CoronaVac) in healthy children and adolescents: a double-blind, randomised, controlled, phase 1/2 clinical trial. Lancet Infect. Dis. 2021, 21, 1645–1653. [Google Scholar] [CrossRef] [PubMed]
- Creech, C.B.; Anderson, E.; Berthaud, V.; Yildirim, I.; Atz, A.M.; Baez, I.M.; Finkelstein, D.; Pickrell, P.; Kirstein, J.; Yut, C.; et al. Evaluation of mRNA-1273 Covid-19 Vaccine in Children 6 to 11 Years of Age. New Engl. J. Med. 2022, 386, 2011–2023. [Google Scholar] [CrossRef]
- Dattner, I.; Goldberg, Y.; Katriel, G.; Yaari, R.; Gal, N.; Miron, Y.; Ziv, A.; Sheffer, R.; Hamo, Y.; Huppert, A. The role of children in the spread of COVID-19: Using household data from Bnei Brak, Israel, to estimate the relative susceptibility and infectivity of children. PLOS Comput. Biol. 2021, 17, e1008559. [Google Scholar] [CrossRef]
- Brandtzaeg, P. Mucosal immunity: integration between mother and the breast-fed infant. Vaccine 2003, 21, 3382–3388. [Google Scholar] [CrossRef]
- Halasa, N.B.; Olson, S.M.; Staat, M.A.; Newhams, M.M.; Price, A.M.; Boom, J.A.; Sahni, L.C.; Cameron, M.A.; Pannaraj, P.S.; Bline, K.E.; et al. Effectiveness of Maternal Vaccination with mRNA COVID-19 Vaccine During Pregnancy Against COVID-19–Associated Hospitalization in Infants Aged <6 Months — 17 States, July 2021–January 2022. MMWR. Morb. Mortal. Wkly. Rep. 2022, 71, 264–270. [Google Scholar] [CrossRef]
- Food D, Administration, Vaccines, Committee RBPA. FDA briefing document: EUA amendment request for Pfizer-BioNTech COVID-19 vaccine for use in children 6 months through 4 years of age. 2022. Available from: https://www.fda.gov/media/159195/download. 1591.
- Wen, J.; Du, X.; Li, A.; Zhang, S.; Shen, S.; Zhang, Z.; Yang, L.; Sun, C.; Li, J.; Zhu, S. Dilemmas and options for COVID-19 vaccination in children. Ital. J. Pediatr. 2023, 49, 1–10. [Google Scholar] [CrossRef]
- DUNCAN JR, et al. Maternal antibody transfer to neonatal mice: enhancement by glucocorticoid administratio. Clin Diagn Lab Immunol 2006, 13, 970–973.
- Simister, N.E.; Story, C.M. Human placental Fc receptors and the transmission of antibodies from mother to fetus. J. Reprod. Immunol. 1997, 37, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Sartoretti, J.; Fontannaz, P.; de Tejada, B.M.; Othenin-Girard, V.; Chilin, A.; Lemaître, B.; Blanchard-Rohner, G.; Siegrist, C.-A.; Eberhardt, C.S. Influence of Timing of Maternal Pertussis Immunization on the Avidity of Transferred Antibodies in Term and Preterm Neonates. Clin. Infect. Dis. 2023, 77, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Cacciottolo, M.; Li, Y.; Nice, J.B.; LeClaire, M.J.; Twaddle, R.; Mora, C.L.; Adachi, S.Y.; Young, M.; Angeles, J.; Elliott, K.; et al. Nanograms of SARS-CoV-2 spike protein delivered by exosomes induce potent neutralization of both delta and omicron variants. PLOS ONE 2023, 18, e0290046. [Google Scholar] [CrossRef]
- Boelig, R.C.; Chaudhury, S.; Gromowski, G.D.; Mayer, S.; King, J.; Aghai, Z.H.; Bergmann-Leitner, E. Reduced maternal immunity and vertical transfer of immunity against SARS-CoV-2 variants of concern with COVID-19 exposure or initial vaccination in pregnancy. Front. Immunol. 2023, 14, 1216410. [Google Scholar] [CrossRef]
Figure 1.
Total IgM in serum samples collected at different time points: (A) 21, (B) 47, (C) 176 and (D) 368 days after immunization. AH: aluminum hydroxide. DDA: dioctadecyldimethylammonium bromide. OMV: outer membrane vesicles. RBD: receptor binding domain. Sap: saponin. *: P<0.05. ns: non significant.
Figure 1.
Total IgM in serum samples collected at different time points: (A) 21, (B) 47, (C) 176 and (D) 368 days after immunization. AH: aluminum hydroxide. DDA: dioctadecyldimethylammonium bromide. OMV: outer membrane vesicles. RBD: receptor binding domain. Sap: saponin. *: P<0.05. ns: non significant.
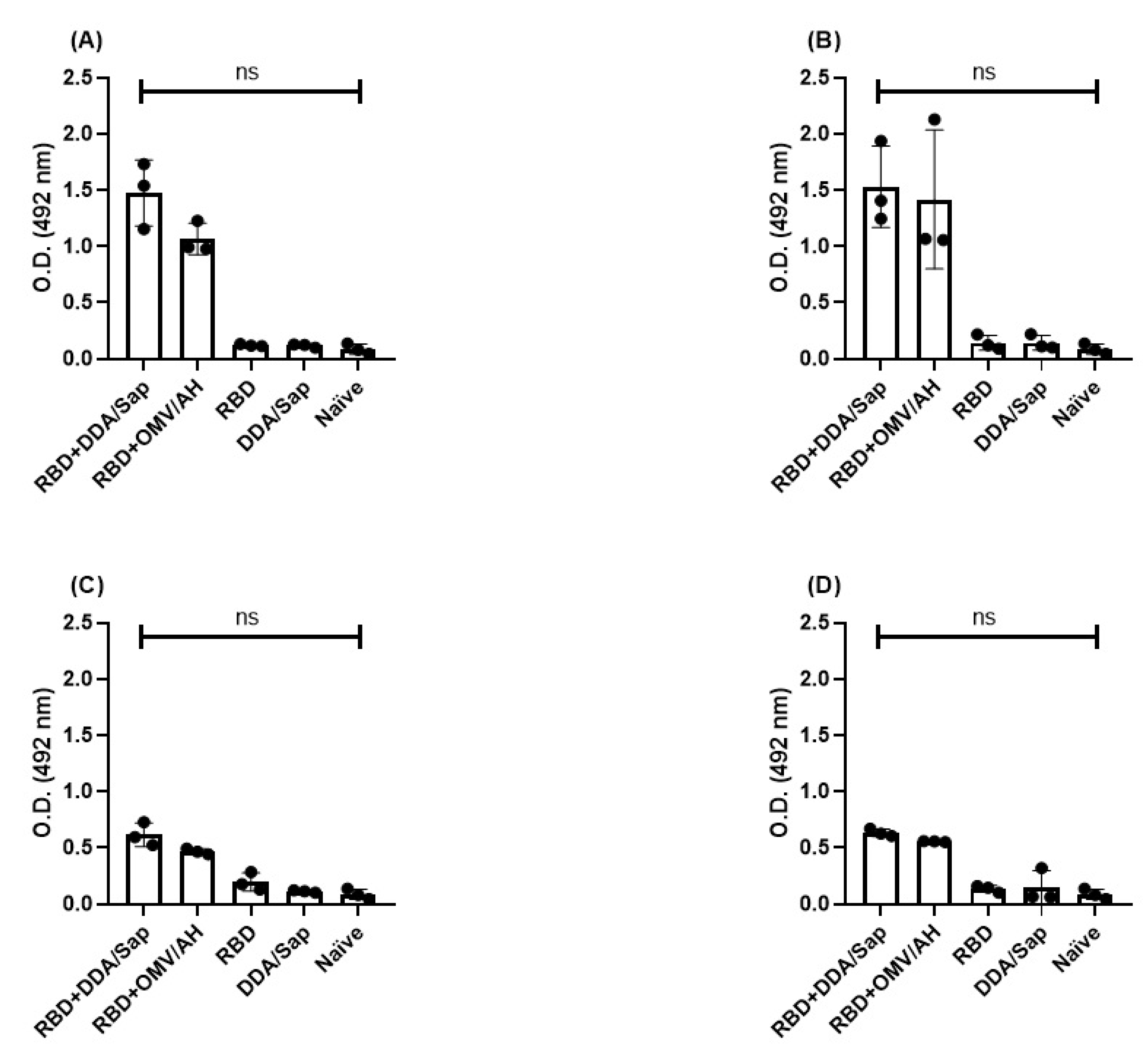
Figure 2.
(A) Immunoblotting using pooled sera of the RBD+DDA/Sap group on day 368. As observed, the antibodies recognized the denatured RBD. MW: molecular weight.
Figure 2.
(A) Immunoblotting using pooled sera of the RBD+DDA/Sap group on day 368. As observed, the antibodies recognized the denatured RBD. MW: molecular weight.

Figure 3.
Kinetics of IgG transferred to offspring in sera collected 15, 30 and 45 days after birth.
Figure 3.
Kinetics of IgG transferred to offspring in sera collected 15, 30 and 45 days after birth.
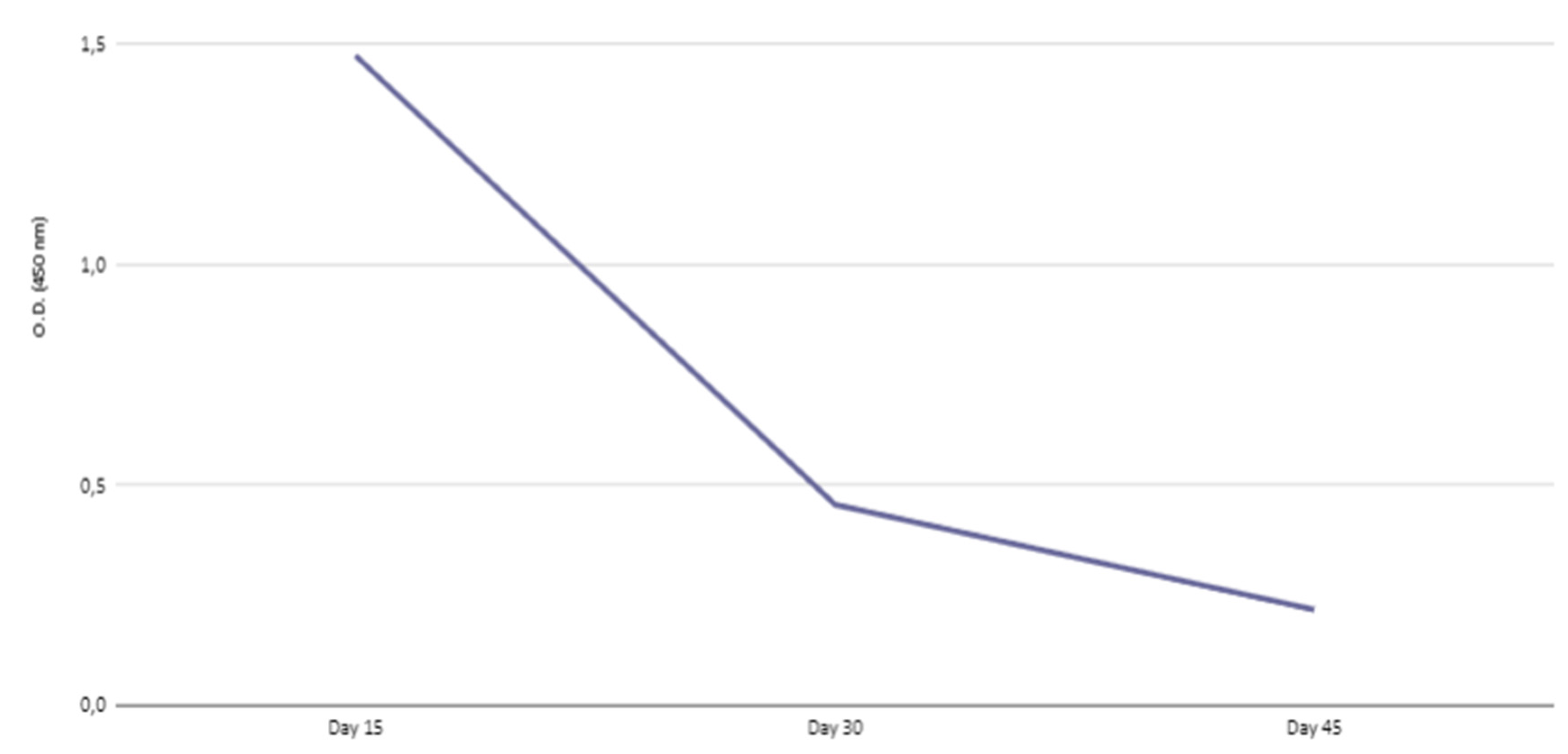
Figure 4.
Spots counted after 1 × 108 splenocytes were stimulated with RBD, considering IFN-γ and IL-4 secretion. Below, the quantification when cells were stimulated with the mitogen ConA or added without stimuli.
Figure 4.
Spots counted after 1 × 108 splenocytes were stimulated with RBD, considering IFN-γ and IL-4 secretion. Below, the quantification when cells were stimulated with the mitogen ConA or added without stimuli.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



