
Submitted:
21 September 2023
Posted:
25 September 2023
You are already at the latest version
Abstract
Wastewater surveillance, initially conceived in the early 20th century during typhoid fever outbreaks, has evolved into a powerful tool for monitoring public health-relevant analytes. Recent applications in tracking SARS-CoV-2 infection highlight its potential. Beyond humans, it can be extended to livestock populations due to the increasing demand for livestock products. Livestock intensification poses risks of zoonotic disease emergence. Wastewater surveillance offers non-invasive, cost-effective means to detect potential outbreaks. This approach aligns with the "One Health" paradigm, emphasizing the interconnectedness of human, animal, and ecosystem health. By monitoring viruses in livestock wastewater, early detection, prevention, and control strategies can be employed, safeguarding both animal and human health, economic stability, and international trade. This integrated One Health approach enhances collaboration and a comprehensive understanding of disease dynamics, supporting proactive measures in the Anthropocene era where animal and human diseases are on the rise.
Keywords:
wastewater surveillance
; livestock
; zoonotic pathogens
; one health
; animal epidemiology
Wastewater surveillance is a burgeoning research field that has recently gained significant traction following the demonstration of its utility to follow population-level trends in SARS-CoV-2 infection (Kitajima et al., 2020). The origins of wastewater surveillance for public health can be traced back to the early 20th century when scientists recognized the presence of Salmonella typhi in wastewater during typhoid fever outbreaks (Wilson, 1933). However, it wasn't until the late 21st century, during the global campaign to eradicate poliovirus, that the concept of using wastewater as a tool for monitoring the presence of public health-relevant analytes in populations took shape (WHO, 1988). In 2005, the field expanded to include chemicals as researchers in Israel demonstrated the ability to detect illicit drug use by analyzing wastewater samples (Zuccato et al., 2005). These breakthroughs demonstrated the utility of wastewater as a non-invasive and cost-effective approach to assess community health. Wastewater surveillance has since evolved and expanded to include the tracking of pharmaceuticals, illicit drugs, pathogens, and various biomarkers in community wastewater (Diamond et al., 2022).
In the wake of the COVID-19 pandemic, wastewater surveillance is now widely recognized as a powerful tool for monitoring the incidence of infectious diseases and for assessing the overall health and well-being of communities (Pang et al., 2022). Its ability to provide unique near real-time data and early warnings of disease spread, relative to traditional disease surveillance, has made it an invaluable resource for public health officials and policymakers in more than 70 countries (Bivins et al., 2020; Tiwari et al., 2023). Most recently, wastewater surveillance has also been applied to detect monkeypox virus, respiratory syncytial virus, influenza viruses, HIV, and other viruses in wastewater samples from treatment plants, dormitories, prisons, hospitals, and transportation hubs (Toribio-Avedillo et al., 2023; Kilaru et al., 2023; Hassard et al., 2022). While current applications of wastewater surveillance focus primarily on humans, there is growing recognition that expanding the scope beyond humans could provide valuable insights into disease prevalence in animal populations. The health of humans, ecosystems, and animals are inextricably linked as has been conceptualized through the “One Health” and “Planetary Health” paradigms (Gibbs et al., 2014; Lebov et al., 2017).
The livestock sector is a pillar of the global food system and contributes approximately 40% and 20% of the global value of agricultural output in developed countries and developing countries, respectively, supporting the livelihoods of at least 1.3 billion people, and providing around 34% of global food protein (Food and Agriculture Organization, 2023). The global demand for livestock products is projected to grow substantially owing to the increasing human population, rapid urbanization, and shifts in consumption patterns to livestock-based diets (Herrero et al., 2013; Alexandratos et al., 2012). A challenge that accompanies livestock intensification is land use change that impacts wildlife habitats, increasing the frequency of livestock-wildlife interactions and the risk of emerging diseases in wildlife, livestock, and humans (Gilbert et al., 2021; Jori et al., 2021). Range shifts associated with climate change will also further increase cross-species transmission with an increased risk of first-time meetings between species creating new opportunities for pathogen transmission (Carlson et al., 2022). These include various zoonotic pathogens, driven by changing proximity interactions within the humans-livestock-wildlife nexus, particularly in species-rich ecosystems (Jones et al., 2013; Perry et al., 2013). Factors driving zoonotic spill over, many of which are accelerating, include characteristics of pathogens, increased human encroachment into natural habitats, host factors, wildlife trade and migration, climate change/climate patterns (i.e., La Niña and El Niño), and agricultural practices (Morens et al., 2004; Jones et al., 2013; Allen et al., 2017; Ellwanger et al., 2021). Therefore, understanding the mechanisms and risks associated with zoonotic virus spill-over is crucial to gain a more comprehensive understanding of environmental transmission, zoonotic diseases, and the overall health of different ecosystems.
More than 60% of human pathogens are zoonotic in origin, transmitted between animals and humans (Rahman et al., 2020). This linkage is implicated in some of the most devastating outbreaks and pandemics in history, such as Ebola, SARS, and most recently, COVID-19 (Karesh et al., 2012; Rahman et al., 2020). Mortality rates attributable to 31 recorded viral zoonoses since 1918 ranged from 0.01 to 27,322 deaths per million, with the baseline expected annual mortality at 3.3 million lives at the current world population level (Bernstein et al., 2022). Additionally, the baseline expected economic cost of zoonotic disease outbreaks is estimated to be around $212 billions of world gross national income per year, if no preventative actions are taken (Bernstein et al., 2022; Fan et al., 2018). The estimates provided are conservative since they do not account for the additional uncertainties surrounding personal and social insecurity in the post-pandemic era. The erosion of such social systems might surpass the direct economic repercussions of pandemics (World Bank, 2010).
Farmed livestock are potential hosts for some zoonotic diseases and increased intensification of production enhances the risk of disease emergence (Gilbert et al., 2021). Current disease surveillance approaches for livestock include the systematic or opportunistic collection and analysis of samples to provide data on disease occurrence in animals. Veterinarians and animal health authorities monitor livestock populations for evidence of disease and investigate unusual patterns or outbreaks using antibody-based assays, fecal swabs, or disease presentation. This epidemiological disease surveillance helps identify potential zoonotic viruses and assess their risks. However, the collection of samples from individual animals to support epidemiological surveillance may be labour and resource intensive, costly and in some cases either aversive to the animals or involves culling.
Diagnostic laboratories play a crucial role in zoonotic virus monitoring. Techniques such as polymerase chain reaction (PCR), genome sequencing, serological tests, and viral isolation are used to detect and characterize zoonotic viruses (Lee et al., 2022; Chauhan et al., 2023). However, like traditional diseases surveillance systems for human populations, such surveillance of a subset of animals from much larger populations has several limitations. Animal diseases may go unnoticed or unreported due to various factors, such as disease being pre- or sub-clinical, lack of awareness, limited resources, inadequate surveillance systems, or reluctance to report for fear of commercial or other economic implications (Gubernot et al., 2008). Laboratory testing may be expensive, time-consuming or produce inconclusive results, and certain regions may be remote or inaccessible, restricting sample collection from animals and presenting challenges to maintaining sample integrity during transport to the laboratory (Hobbs et al., 2021). These factors can lead to limited sensitivity for detecting emerging threats and underestimation of the true incidence and prevalence of existing or emerging diseases.
For livestock where aggregated waste streams are available for sampling, such as drainage systems in animal housing, wastewater pits or lagoons, wastewater surveillance of animal populations may provide advantages in early, passive detection of potential disease outbreaks, as has been observed among human populations (Ahmed et al., 2021). For example, in the case of Japanese encephalitis virus (JEV), a zoonotic pathogen for which pigs are an important amplifying host, as shown in Figure 1, wastewater surveillance offers several advantages over traditional surveillance approaches. Current laboratory diagnosis of JEV in pigs involves testing of specimens from aborted foetuses using reverse transcriptase quantitative PCR (RT-qPCR) assays which usually takes <24 h (Williams et al., 2019). This may be accompanied by serological testing of older pigs on affected farms, such as grower pigs and sows, which may take weeks to complete (in the case of cell culture-based assays) and interpretation can be clouded by cross-reaction with related viruses. As an alternative to accompany disease investigations or as part of arbovirus surveillance programs, wastewater from swine farms could be tested for the presence of JEV nucleic acid to indicate active infections and provide valuable information to support strategic animal and public health management strategies. This approach could provide an early warning system informing prompt and targeted responses, such as implementing mosquito surveillance and control measures, vaccination programs, heightened surveillance for evidence of clinical disease or other control strategies. Moreover, an advanced alert system coupled with timely intervention has the potential to limit outbreaks, resulting in significant cost savings to the industry.
Wastewater surveillance may also be helpful to identify high-risk livestock agriculture and aquaculture areas or populations using non-invasive and aggregated signals in fecal waste streams or culture waters. Besides zoonotic pathogens of concern for human populations, disease outbreaks in livestock and aquaculture can have severe health, welfare, and economic consequences, including livestock mortality and morbidity, reduced productivity, trade restrictions, market disruptions, control costs and impacts on farming communities (e.g., livelihoods, support industries, psychological etc.). As summarized in Table 1, many pathogenic viruses have the potential to cause significant animal and public health impacts. Monitoring for pathogenic viruses in livestock populations via wastewater could provide insights into their prevalence, distribution, and evolution, which could then be used to target early preventive measures and management strategies to reduce the risk of transmission to humans or to animals at other locations. Wastewater surveillance of zoonotic viruses could strategically support the identification and management of threats before they escalate. By preventing or mitigating disease spread, passive monitoring using wastewater would contribute to the economic stability of the livestock sectors, ensuring the sustainability of food production and safeguarding industries, livelihoods, and international trade.
By monitoring viruses in livestock populations through wastewater, the health status of and risks to both animals and humans can be assessed simultaneously. Such an integrated One Health approach will facilitate collaboration and information sharing among various sectors, such as agriculture, veterinary medicine, animal, and public health. This would allow for a more comprehensive understanding of both zoonotic and livestock diseases and support coordinated efforts to prevent and control them. Furthermore, the approach may be extended to provide insights into longer-range environmental transmission and into pathogens with complex ecologies. For example, multiple vectors may host JEV, with mosquitoes serving as the primary vector, along with other recognised hosts, including birds, bats, and wild pigs (Hall-Medelin et al., 2012; Lord et al., 2015; Auerswald et al., 2020; Hameed et al., 2021). The potential for monitoring of broader environmental samples (environmental waters, faeces) for a more holistic One Health view of risks also offers opportunities to investigate such environments or wildlife habitats with respect to their role in zoonotic disease spread.
The analytical approach for livestock wastewater or environmental water surveillance depends on its purpose, sensitivity requirements, available resources, and allowable cost. Various virus concentration methods have been used for SARS-CoV-2 wastewater surveillance, including membrane filtration, ultrafiltration, Concentrating Pipette, PEG precipitation, and magnetic nanoparticles (Ahmed et al., 2020; Karthikeyan et al., 2021). These methods can also be adapted and applied to a range of wastewater and environmental water surveillance systems as needed. To overcome logistical challenges in remote sampling areas, simplified virus detection methods for wastewater samples may be feasible. However, the analytical method selected must achieve the necessary sensitivity and specificity. Two categories of analysis are available: pathogen-specific methods (e.g., quantitative PCR (qPCR), RT-qPCR, digital PCR (dPCR)) and non-specific methods such as metagenomics. While pathogen-specific methods are sensitive and specific, they are not suitable for detecting unknown pathogens. PCR-based techniques, such as TaqMan array cards or microarrays could also be valuable for high-throughput screening of livestock wastewater for known viruses. Other techniques worth exploring include loop-mediated isothermal amplification (LAMP), helicase-dependent amplification (HDA), mass spectrometry, enzyme-linked immunosorbent assays (ELISA), and CRISPR-based diagnostic assays. Some of these techniques may work well in low resource settings. Next-generation sequencing (NGS) and metagenomics offer semi-targeted or untargeted analytical capabilities, enabling broad genomic surveillance of livestock wastewater or other environmental matrices.
As the emergence and re-emergence of zoonotic diseases are expected to accelerate in the Anthropocene (McMahon et al., 2018), the importance of livestock surveillance will be increasingly essential for early disease detection and control. Efforts are continually being made to address these challenges through improved surveillance systems that offer better diagnostics, are less resource intensive, less impactful on animals, that build capacity, and that enhance data sharing, and collaborative research efforts. Animal epidemiology in combination with wastewater surveillance of livestock has the potential to play a vital role in early detection, disease prevention, public health protection, One Health initiatives, and economic stability. Its successful implementation will depend on the willingness, understanding and support of farm owners, industry, and public and animal health authorities.
References
- Ahmed, W.; Bertsch, P.M.; Bivins, A.; Bibby, K.; Farkas, K.; Gathercole, A.; Haramoto, E.; Gyawali, P.; Korajkic, A.; McMinn, B.R.; et al. Comparison of virus concentration methods for the RT-qPCR-based recovery of murine hepatitis virus, a surrogate for SARS-CoV-2 from untreated wastewater. Sci. Total Environ. 2020, 739, 139960. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Tscharke, B.; Bertsch, P.M.; Bibby, K.; Bivins, A.; Choi, P.; Clarke, L.; Dwyer, J.; Edson, J.; Nguyen, T.M.H.; et al. SARS-CoV-2 RNA monitoring in wastewater as a potential early warning system for COVID-19 transmission in the community: A temporal case study. Sci. Total Environ. 2021, 761, 144216. [Google Scholar] [CrossRef] [PubMed]
- Alexandratos, N.; Bruinsma, J. World Agriculture Towards 2030/2050: The 2012 Revision; Food and Agriculture Organization of the United Nations: Rome, 2012. [Google Scholar]
- Allen, T.; Murray, K.A.; Zambrana-Torrelio, C.; Morse, S.S.; Rondinini, C.; Di Marco, M.; Breit, N.; Olival, K.J.; Daszak, P. Global hotspots and correlates of emerging zoonotic diseases. Nat. Commun. 2017, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Auerswald, H.; Ruget, A.-S.; Ladreyt, H.; In, S.; Mao, S.; Sorn, S.; Tum, S.; Duong, V.; Dussart, P.; Cappelle, J.; et al. Serological Evidence for Japanese Encephalitis and West Nile Virus Infections in Domestic Birds in Cambodia. Front. Veter- Sci. 2020, 7, 15. [Google Scholar] [CrossRef]
- Bernstein, A.S.; Ando, A.W.; Loch-Temzelides, T.; Vale, M.M.; Li, B.V.; Li, H.; Busch, J.; Chapman, C.A.; Kinnaird, M.; Nowak, K.; et al. The costs and benefits of primary prevention of zoonotic pandemics. Sci. Adv. 2022, 8, eabl4183. [Google Scholar] [CrossRef]
- Bivins, A.; North, D.; Ahmad, A.; Ahmed, W.; Alm, E.; Been, F.; Bhattacharya, P.; Bijlsma, L.; Boehm, A.B.; Brown, J.; et al. Wastewater-Based Epidemiology: Global Collaborative to Maximize Contributions in the Fight Against COVID-19. Environ. Sci. Technol. 2020, 54, 7754–7757. [Google Scholar] [CrossRef]
- Carlson, C.J.; Albery, G.F.; Merow, C.; Trisos, C.H.; Zipfel, C.M.; Eskew, E.A.; Olival, K.J.; Ross, N.; Bansal, S. Climate change increases cross-species viral transmission risk. Nature 2022, 607, 555–562. [Google Scholar] [CrossRef]
- Chauhan, R.P.; Fogel, R.; Limson, J. Overview of Diagnostic Methods, Disease Prevalence and Transmission of Mpox (Formerly Monkeypox) in Humans and Animal Reservoirs. Microorganisms 2023, 11, 1186. [Google Scholar] [CrossRef]
- Diamond, M.B.; Keshaviah, A.; Bento, A.I.; Conroy-Ben, O.; Driver, E.M.; Ensor, K.B.; Halden, R.U.; Hopkins, L.P.; Kuhn, K.G.; Moe, C.L.; et al. Wastewater surveillance of pathogens can inform public health responses. Nat. Med. 2022, 28, 1992–1995. [Google Scholar] [CrossRef]
- Ellwanger, J.H.; Chies, J.A.B. Zoonotic spillover: Understanding basic aspects for better prevention. Genet. Mol. Biol. 2021, 44, e20200355. [Google Scholar] [CrossRef]
- Fan, V.Y.; Jamison, D.T.; Summers, L.H. Pandemic risk: How large are the expected losses? Bull. World Health Organ. 2018, 96, 129–134. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. 2023. Animal production. Available online: https://www.fao.org/animal-production/en/.
- Gibbs, E.P.J. The evolution of One Health: a decade of progress and challenges for the future. Veter- Rec. 2014, 174, 85–91. [Google Scholar] [CrossRef]
- Gilbert, W.; Thomas, L.; Coyne, L.; Rushton, J. Review: Mitigating the risks posed by intensification in livestock production: the examples of antimicrobial resistance and zoonoses. Animal 2021, 15, 100123. [Google Scholar] [CrossRef]
- Gubernot, D.M.; Boyer, B.L.; Moses, M.S. Animals as Early Detectors of Bioevents: Veterinary Tools and a Framework for Animal-Human Integrated Zoonotic Disease Surveillance. Public Heal. Rep. 2008, 123, 300–315. [Google Scholar] [CrossRef] [PubMed]
- Hall-Mendelin, S.; Jansen, C.C.; Cheah, W.Y.; Montgomery, B.L.; Hall, R.A.; Ritchie, S.A.; Hurk, A.F.V.D. Culex annulirostris (Diptera: Culicidae) host feeding patterns and Japanese encephalitis virus ecology in northern Australia. J. Med Èntomol. 2012, 49, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Hameed, M.; Wahaab, A.; Nawaz, M.; Khan, S.; Nazir, J.; Liu, K.; Wei, J.; Ma, Z. Potential Role of Birds in Japanese Encephalitis Virus Zoonotic Transmission and Genotype Shift. Viruses 2021, 13, 357. [Google Scholar] [CrossRef] [PubMed]
- Hassard, F.; Smith, T.R.; Boehm, A.B.; Nolan, S.; O'Mara, O.; Di Cesare, M.; Graham, D. Wastewater surveillance for rapid identification of infectious diseases in prisons. Lancet Microbe 2022, 3, e556–e557. [Google Scholar] [CrossRef]
- Herrero, M.; Thornton, P.K. Livestock and global change: Emerging issues for sustainable food systems. Proc. Natl. Acad. Sci. 2013, 110, 20878–20881. [Google Scholar] [CrossRef]
- Hobbs, E.C.; Colling, A.; Gurung, R.B.; Allen, J. The potential of diagnostic point-of-care tests (POCTs) for infectious and zoonotic animal diseases in developing countries: Technical, regulatory and sociocultural considerations. Transbound. Emerg. Dis. 2021, 68, 1835–1849. [Google Scholar] [CrossRef]
- Jones, B.A.; Grace, D.; Kock, R.; Alonso, S.; Rushton, J.; Said, M.Y.; McKeever, D.; Mutua, F.; Young, J.; McDermott, J.; et al. Zoonosis emergence linked to agricultural intensification and environmental change. Proc. Natl. Acad. Sci. 2013, 110, 8399–8404. [Google Scholar] [CrossRef]
- Jori, F.; Hernandez-Jover, M.; Magouras, I.; Dürr, S.; Brookes, V.J. Wildlife–livestock interactions in animal production systems: what are the biosecurity and health implications? Anim. Front. 2021, 11, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Karesh, W.B.; Dobson, A.; Lloyd-Smith, J.O.; Lubroth, J.; Dixon, M.A.; Bennett, M.; Aldrich, S.; Harrington, T.; Formenty, P.; Loh, E.H.; et al. Ecology of zoonoses: Natural and unnatural histories. Lancet 2012, 380, 1936–1945. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, S.; Ronquillo, N.; Belda-Ferre, P.; Alvarado, D.; Javidi, T.; Longhurst, C.A.; Knight, R. High-throughput wastewater SARS-CoV-2 detection enables forecasting of community infection dynamics in San Diego County. mSystems 2021, 6, e00045–21. [Google Scholar] [CrossRef] [PubMed]
- Kilaru, P.; Hill, D.; Anderson, K.; Collins, M.B.; Green, H.; Kmush, B.L.; A Larsen, D. Wastewater Surveillance for Infectious Disease: A Systematic Review. Am. J. Epidemiology 2022, 192, 305–322. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, M.; Ahmed, W.; Bibby, K.; Carducci, A.; Gerba, C.P.; Hamilton, K.A.; Haramoto, E.; Rose, J.B. SARS-CoV-2 in wastewater: State of the knowledge and research needs. Sci. Total Environ. 2020, 739, 139076. [Google Scholar] [CrossRef]
- Lord, J.S.; Gurley, E.S.; Pulliam, J.R.C. Rethinking Japanese Encephalitis Virus Transmission: A Framework for Implicating Host and Vector Species. PLoS Negl. Trop. Dis. 2015, 9, e0004074. [Google Scholar] [CrossRef]
- Lebov, J.; Grieger, K.; Womack, D.; Zaccaro, D.; Whitehead, N.; Kowalcyk, B.; MacDonald, P. A framework for One Health research. One Heal. 2017, 3, 44–50. [Google Scholar] [CrossRef]
- Lee, W.L.; Gu, X.; Armas, F.; Leifels, M.; Wu, F.; Chandra, F.; Chua, F.J.D.; Syenina, A.; Chen, H.; Cheng, D.; et al. Monitoring human arboviral diseases through wastewater surveillance: Challenges, progress and future opportunities. Water Res. 2022, 223, 118904. [Google Scholar] [CrossRef]
- Lee, W.L.; Gu, X.; Armas, F.; Leifels, M.; Wu, F.; Chandra, F.; Chua, F.J.D.; Syenina, A.; Chen, H.; Cheng, D.; et al. Monitoring human arboviral diseases through wastewater surveillance: Challenges, progress and future opportunities. Water Res. 2022, 223, 118904. [Google Scholar] [CrossRef]
- McMahon, B.J.; Morand, S.; Gray, J.S. Ecosystem change and zoonoses in the Anthropocene. Zoonoses Public Heal. 2018, 65, 755–765. [Google Scholar] [CrossRef]
- Morens, D.M.; Folkers, G.K.; Fauci, A.S. The challenge of emerging and re-emerging infectious diseases. Nature 2004, 430, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Gao, T.; Ellehoj, E.; Li, Q.; Qiu, Y.; Maal-Bared, R.; Sikora, C.; Tipples, G.; Diggle, M.; Hinshaw, D.; et al. Wastewater-Based Surveillance Is an Effective Tool for Trending COVID-19 Prevalence in Communities: A Study of 10 Major Communities for 17 Months in Alberta. ACS ES&T Water 2022, 2, 2243–2254. [Google Scholar] [CrossRef]
- Perry, B.D.; Grace, D.; Sones, K. Current drivers and future directions of global livestock disease dynamics. Proc. Natl. Acad. Sci. 2011, 110, 20871–20877. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.T.; Sobur, M.A.; Islam, M.S.; levy, S.; Hossain, M.J.; El Zowalaty, M.E.; Rahman, A.T.; Ashour, H.M. Zoonotic diseases: Etiology, impact, and control. Microorganisms 2020, 8, 1405. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, A.; Adhikari, S.; Zhang, S.; Solomon, T.B.; Lipponen, A.; Islam, M.A.; Thakali, O.; Sangkham, S.; Shaheen, M.N.F.; Jiang, G.; Haramoto, E.; Mazumder, P.; Malla, B.; Kumar, M.; Pitkanen, T.; Sherchan, S.P. . Tracing COVID-19 trails in wastewater: A systematic review of SARS-CoV-2 Surveillance with viral variants. Water 2023, 15, 1018. [Google Scholar] [CrossRef]
- Toribio-Avedillo, D.; Gómez-Gómez, C.; Sala-Comorera, L.; Rodríguez-Rubio, L.; Carcereny, A.; García-Pedemonte, D.; Pintó, R.M.; Guix, S.; Galofré, B.; Bosch, A.; et al. Monitoring influenza and respiratory syncytial virus in wastewater. Beyond COVID-19. Sci. Total. Environ. 2023, 892, 164495–164495. [Google Scholar] [CrossRef]
- Toribio-Avedillo, D.; Gómez-Gómez, C.; Sala-Comorera, L.; Rodríguez-Rubio, L.; Carcereny, A.; García-Pedemonte, D.; Pintó, R.M.; Guix, S.; Galofré, B.; Bosch, A.; et al. Monitoring influenza and respiratory syncytial virus in wastewater. Beyond COVID-19. Sci. Total. Environ. 2023, 892, 164495–164495. [Google Scholar] [CrossRef]
- WHO. Expanded programme on immunization global eradication of poliomyelitis by the year 2000. Wkly. Epidemiol. Rec. 1988, 63, 161–162. [Google Scholar]
- Williams, D.T.; MacKenzie, J.S.; Bingham, J. ; 2019. Flaviviruses. In Diseases of Swine (eds J.J.; Zimmerman, L.A.; Karriker, A.; Ramirez, K.J.; Schwartz, G.W.; Stevenson and J.; Zhang).
- Wilson, W.J. Isolation of enteric Bacilli from sewage and water and its bearing on epidemiology. BMJ 1933, 2, 560–562. [Google Scholar] [CrossRef]
- World Bank. (2010). People, Pathogens and Our Planet, Vol 1: Towards a One Health Approach for Controlling Zoonotic Diseases. Report 50833-GLB.
- Zuccato, E.; Chiabrando, C.; Castiglioni, S.; Calamari, D.; Bagnati, R.; Schiarea, S.; Fanelli, R. Cocaine in surface waters: a new evidence-based tool to monitor community drug abuse. Environ. Health 2005, 4, 14. [Google Scholar] [CrossRef]
Figure 1.
Proof-of-concept of wastewater surveillance of Japanese Encephalitis Virus (JEV) at swine farms.
Figure 1.
Proof-of-concept of wastewater surveillance of Japanese Encephalitis Virus (JEV) at swine farms.
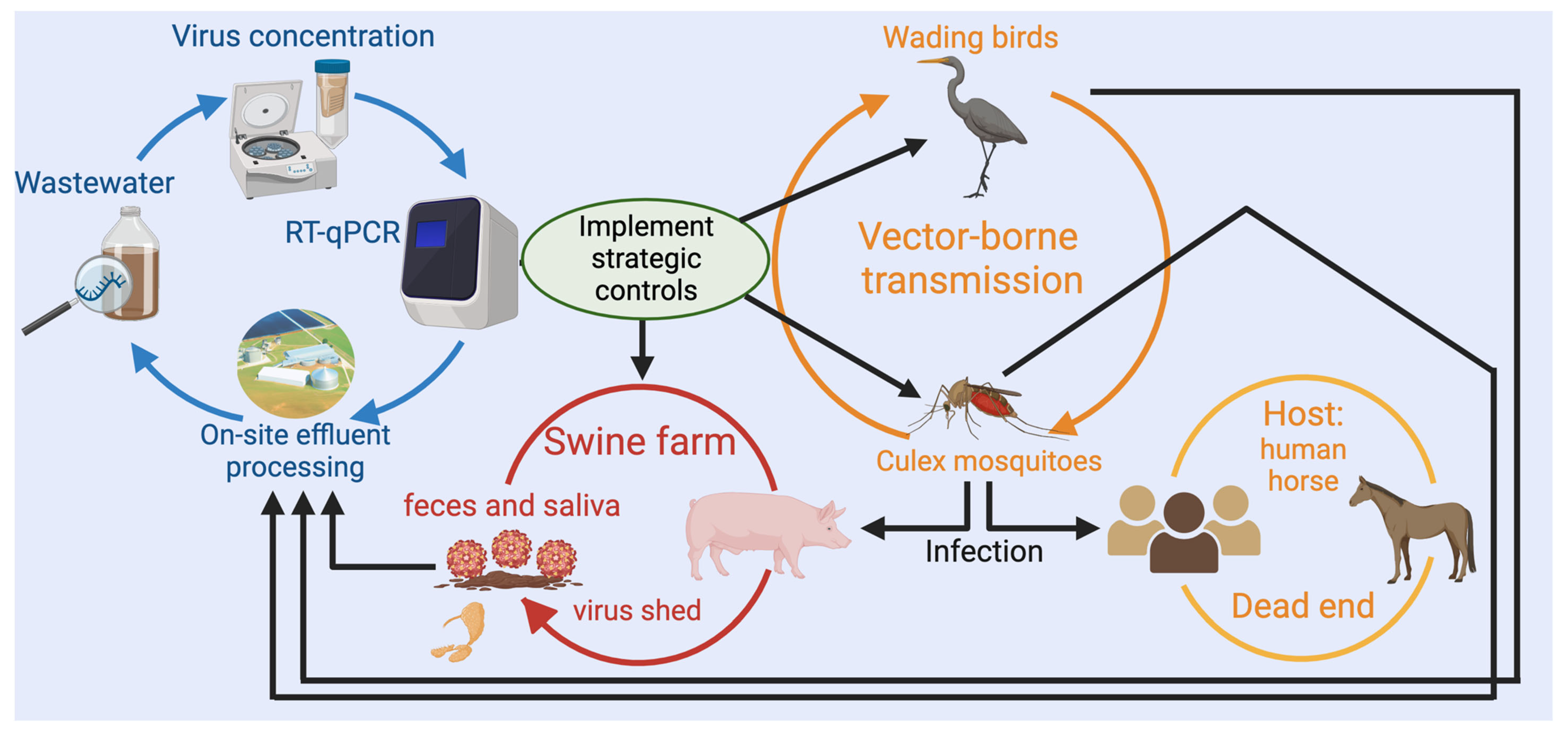

Table 1.
Epidemiology and transmission routes of livestock-associated pathogenic viruses.
| Viruses | Family | Genus | Genome type | Epidemiology | Source | Transmission | Mortality and morbidity | Hosts |
|---|---|---|---|---|---|---|---|---|
| African swine fever virus (ASFV) Non-zoonotic |
Asfarviridae | Asfivirus. | Enveloped DNA | Endemic in Africa, Europe, Asia and the Carribean | Blood, tissues, secretions and excretions of sick and dead animals | Direct and indirect contact between infected and susceptible animals; indirect transmission via contaminated fomites including trucks, feed, people, equipment and possibly some flies; swill feeding; Vector-borne transmission: soft ticks of the genus Ornithodoros (Africa) | Mortality rates up to 100% in domestic pigs | Domestic and wild Sus scrofa; warthogs (Phacochoerus spp.), bush pigs (Potamochoerus spp.) and giant forest hogs (Hylochoerus meinertzhageni); Ticks of belonging to the Ornithodoros moubata complex |
| Bluetongue virus (BTV) Non-zoonotic |
Reoviridae | Orbivirus | Non-enveloped dsRNA | Occurs on all continents except Antarctica. Endemic in many parts of the world, including Europe, Asia, Australia and North America | Infected Culicoides spp, blood, semen, placenta; | Vector-borne transmission: midges of the genus Culicoides | Mortality rates of 30-70% in susceptible sheep, with morbidity rates close to 100%; mortality rates up to 90% in wild deer and antelopes; cattle have often higher infection rates than sheep | Domestic and wild ruminants including sheep, goats, cattle, buffaloes, deer, most species of African antelope and other Artiodactyla such as camels |
| Dengue virus (DENV) Zoonotic |
Flaviviridae | Flavivirus | Enveloped ssRNA | Endemic in many parts of the world, especially in tropical parts of America, Africa, South-East Asia. | Infected mosquito vectors; humans and primates | Mosquito-borne transmission: mosquito of the genus Aedes | Many DENV infections are asymptomatic or produce only mild illness. In severe cases, dengue can be fatal | Humans are the main host. Infection also occurred in pigs, marsupials, bats, birds, horses, bovid, rodent and dogs |
| Foot-and-mouth disease virus (FMDV) |
Picornaviridae | Aphthovirus | Non-enveloped ssRNA | Endemic in parts of Asia, Africa, the Middle East and South America (sporadic outbreaks in free areas) | All secretions and excretions from acutely infected animals including expired air, saliva, milk, urine, feces, semen, fluid from FMD-associated vesicles, and in amniotic fluid and aborted fetuses in sheep | Direct and indirect contact between infected and susceptible animals; indirect transmission via contaminated fomites; airborne spread | Mortality rates of 1-5% in adult animals, and more than 20% in young animals. Morbidity can reach 100% in susceptible populations | Cloven-footed livestock species such as cattle, sheep and pigs. |
| Hepatitis E virus (HEV) Zoonotic |
Hepeviridae | Orthohepevirus | Non-enveloped ssRNA | Worldwide but most common in east and south Asia | Feces | Fecal-oral route. | Case fatality rates around 1-3% in human, and 10-30% among pregnant women; mortality rates around 1% in chicken | Pigs, wild boars, cows, deer, rabbits, chicken, camels, contaminated shellfish and humans |
| Influenza type A viruses (e.g., H1N1, H5N1, H7N9, H3N2) Zoonotic |
Orthomyxoviridae | Alphainfluenzavirus | Enveloped ssRNA | Endemic in all parts of the world | Saliva, nasal secretions and faeces of infected animals | Direct and indirect contact between infected animals; indirect transmission via contaminated fomites including feed, equipment and other materials; Airborne; droplet | Case fatality rates around 56% in human; mortality up to 90-100% in chicken for highly pathogenic subtypes | Wild aquatic birds, mammals. |
| Japanese encephalitis virus (JEV) Zoonotic |
Flaviviridae | Flavivirus | Enveloped ssRNA | Endemic in eastern, south-eastern, and southern Asian countries, and western India, western Pacific region including the eastern Indonesian archipelago, Papua New Guinea and Northern Australia | Infected mosquito vectors, water birds including the family Ardeidae (herons and egrets), pigs are an amplifying host | Vector-borne transmission cycle among mosquitoes (Culex spp.), birds and swine. Humans, horses and other species considered dead end hosts | Case morbidity in horses 1-1.4% but fatality rates in these cases from 5-15% to 30-40% in severe epizootics; human case morbidity 1% but case fatality rates can be up to 30%; morbidity and mortality rate near 0% in adult swine but can reach 100% in non-immune piglets infected in utero | Wild water birds of the family Ardeidae, pigs, human and horse (dead-end host) |
| Lumpy skin disease virus (LSDV) Non-zoonotic |
Poxviridae | Capripoxvirus | Enveloped DNA | Endemic in most African partscountries, parts of the Middle East, south-east Europe, the Balkans, Caucasus, Russian Federation and large parts of Asia | Skin nodules, scabs, crusts (contain relatively high amount of LDSV); blood, saliva, ocular, nasal discharge and semen of infected animals | Arthropod vector-borne transmission (mechanical): mosquitoes (e.g. Culex mirificens and Aedes natrionus), biting flies (e.g. Stomoxys calcitrans and Biomyia fasciata) and male ticks (Riphicephalus appendiculatus and Amblyomma hebraeum) |
Mortality rates of 1-5% and morbidity rates of 10-20% in cattle | Cattle (Bos indicus and B. taurus), water buffalo (Bubalus bubalis) |
| Porcine epidemic diarrhea virus (PEDV) Non-zoonotic |
Coronaviridae | Alphacoronavirus | Enveloped ssRNA | Endemic in Europe, large parts of Asia and the Americas | Faeces of infected animals | Direct transmission via faecal-oral route; indirect transmission via contaminated fomites including trucks, feed, equipment and people | Mortality rates less than 5 % in adult and fattening pigs, less than 10 % in piglets older than 10 days, up to 100 % in sucking piglets, morbidity rates up to 100% | Pigs |
| Porcine reproductive and respiratory syndrome virus (PRRSV) Non-zoonotic |
Arteriviridae |
Arterivirus | Enveloped ssRNA | Endemic in pig-raising areas of Europe, North America, parts of Asia | Body fluids and secretions of infected animals | Direct and indirect contact between infected and susceptible animals; indirect transmission via contaminated fomites including equipment and insects; transplacental; potential wind-borne |
Mortality rates of 10% in pregnant sows, 20% in finishers, 70% in nursery pigs and 100% in suckling pigs | Domestic pigs |
| Rift Valley fever virus (RVFV) Zoonotic |
Phenuiviridae | Phlebovirus | Enveloped ssRNA | Endemic mainly in many sub-Saharan African countries, Egypt, the Arabian Peninsula (South-west) and some Indian Ocean Islands. Usually presents in epizootic form following heavy rains/flooding | Eggs of infected mosquitoes, blood, secretions and tissues of infected animals | Arthropod-borne transmission, direct contact with the blood, secretions and tissue of infected animals; Airborne | Mortality rates of 70-100 % in young lambs and goats; 20-70% in sheep and calves; less than 10% in adult goats, cattle, buffalo, inapparent in pigs, dogs, cats, Asian monkeys, and humans; influenza-like syndrome in humans | Domestic and wild animals including cattle, sheep, goats, monkeys, wild ruminants, antelope, domestic carnivores. Humans are highly susceptible but dead end hosts |
| West Nile virus (WNV) Zoonotic |
Flaviviridae | Flavivirus | Enveloped ssRNA | Endemic in many parts of the world, particularly in Africa, Middle East, West Asia, Australia, and parts of Europe | Infected mosquito vectors; bird reservoirs | Vector-borne transmission cycle between mosquitoes (genus Culex) and birds. Mammals including horses are dead-end hosts | Case fatality rates of 23-57% in equids, close to 100% in mammals other than horses; mortality rates around 20-60% in geese; typically mild symptoms in human and horse, but can cause neurological disease. | Birds, reptiles, amphibians, mammals and marsupials, mosquitoes, and ticks |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



