Submitted:
24 September 2023
Posted:
25 September 2023
You are already at the latest version
Abstract
Macrophages serve as vital defenders, protecting the body by exhibiting remarkable cellular adaptability in response to invading pathogens and various stimuli. These cells express nicotinic acetylcholine receptors, with the α7-nAChR being extensively studied due to its involvement in activating the cholinergic anti-inflammatory pathway. Activation of this pathway plays a crucial role in suppressing macrophages' production of proinflammatory cytokines, thus mitigating excessive inflammation and maintaining host homeostasis. Macrophage polarization, which occurs in response to specific pathogens or insults, is a process that has received limited attention concerning the activation of the cholinergic anti-inflammatory pathway and the contributions of the α7-nAChR in this context. This review aims to present evidence highlighting how the cholinergic machinery in macrophages, led by the α7-nAChR, facilitates the polarization of macrophages towards anti-inflammatory phenotypes. Additionally, we explore the influence of viral infections on macrophage inflammatory phenotypes, taking into account cholinergic mechanisms. We also review the current understanding of macrophage polarization in response to these infections. Finally, we provide insights into the relatively unexplored partial duplication of the α7-nAChR, known as dup α7, which is emerging as a significant factor in macrophage polarization and inflammation scenarios.
Keywords:
macrophage
; inflammation
; acetylcholine receptor
; cholinergic receptor
; polarization
; cholinergic agonist
; alpha 7 nicotinic acetylcholine receptor
; CHRFAM7A
; cholinergic machinery
; cholinergic anti-inflammatory pathway
1. Introduction
Macrophages are a vital component of the immune system responsible for defending the body against pathogens and foreign substances. Found throughout the body, particularly in tissues and organs, macrophages play a crucial role in innate immunity. Their primary function is to engulf and eliminate harmful microorganisms, debris, and dead cells through phagocytic processes involving cytokines [1,2,3,4,5].
Macrophages exhibit remarkable plasticity, capable of adopting different phenotypes and functions in response to signals from their microenvironment. In the presence of pathogens, insults, or tissue alterations, macrophages fulfill their critical role by polarizing and differentiating into distinct subtypes with specific roles and functions. Currently, four main subgroups (M1-M4) have been identified, working in conjunction with other immune cells to defend the body and maintain organ and systemic homeostasis. Of note, macrophages are equipped with cholinergic machinery that enables them to respond in an anti-inflammatory manner through a neuroimmune circuit known as the cholinergic anti-inflammatory pathway (CAP) [6,7]. In macrophages, this pathway is dependent on the expression of the alpha7 nicotinic acetylcholine receptor (α7-nAChR) [6], an ion channel with anti-inflammatory properties upon stimulation.
Although significant progress has been made in understanding the role and function of the CAP, there are still many aspects that require further clarification and investigation. For instance, the role of macrophage polarization in limiting inflammation through the cholinergic anti-inflammatory response (CAR) is still not fully understood. Moreover, the response of polarized macrophages (M1-M4) to the CAR during disease and infection remains understudied. In this review, we aim to discuss the available evidence on how the cholinergic machinery in macrophages contributes to the polarization of these cells towards stimulus-dependent phenotypes. Furthermore, we explore scenarios where viruses disrupt macrophage polarization and alter α7-nAChR expression, thereby contributing to inflammation. Last, we delve into published research that establishes the connection between the CAP and macrophage polarization, with a particular emphasis on the role of the α7-nAChR and its partial duplication, dup α7.
2. Activation and Polarization of Human Macrophages
Human macrophages are white immune cells that destroy pathogens; participate in tissue remodeling, wound healing, and angiogenesis; they remove dead immune cells, participate in metabolism; and secrete cytokines and chemokines to stimulate other immune system cells to participate in the immune response, among other. Macrophages become activated and polarized as part of the natural response process that maximizes their antimicrobial properties and maintains host homeostasis. Their cellular plasticity allows them to polarize into two distinct populations: classically activated inflammatory (M1) and alternatively activated anti-inflammatory (M2) macrophages (Figure 1). The latter can be divided into four subsets (M2a, b, c, and d) based on the type of stimulus they receive (Figure 2). The specialization and tropism of macrophages toward organs is an example of cellular plasticity in both health and disease. This is the case with the relatively recent discovery of five types of macrophages (M4, Mhem, M(Hb), HA-mac, and Mox) associated with the development of atherosclerosis and atheroinflammation [8,9,10,11,12,13] (Figure 3). There are also M3 macrophages known as commuting macrophages. The first study discussing the emergence of this previously unknown lineage showed that their appearance depended on the switching response between M1 and M2 macrophage pathways [14]. It has recently been demonstrated that these macrophages have antitumor activity, limiting tumor proliferation [15], and that they may be a pharmacological target for inducing antitumor immunity [16]. Finally, M4 macrophages are a subset of macrophages that are polarized by platelet factor 4. This population is found in human lesions and is distinguished by high levels of matrix metalloprotease 7 and S100A8 expression. M4 macrophages are considered atherogenic because they produce proinflammatory cytokines (IL-6 and TNF-α) and have poor phagocytic properties [17,18]. It has been reported that these cells are reproducibly found in coronary artery plaques [19]. In HIV infection, these inflammatory M4-macrophages form a major tissue reservoir of replication-competent HIV-1, which reactivates viral production upon autocrine/paracrine S100A8-mediated glycolytic stimulation [20]. Furthermore, it has been discovered that the level of cholesterol oxidation influences the profile of these M4 macrophages, with LDL being the molecule that causes the most significant changes. This results in the polarization of M2 macrophages and Kupffer cells towards the M4 phenotype. These cells also demonstrate increased neutrophil recruitment and more effective induction of neutrophil extracellular traps [21].
2.1. Polarization of Macrophages during Viral Infection
One of the hallmark events of macrophages during viral infections is their polarization switch during the immune response. This polarization could be triggered by TLR4 or IL-1R ligand activation, IFN-γ binding to its receptor (IFN-γR), interaction of Notch proteins with Delta-like and Jagged ligands, and IL-4 or IL-13 binding to its corresponding receptor [22]. Depending on the virus, the stage of infection, and even the infected person's gender, macrophages adopt different inflammatory phenotypes, either M1, M2, or a biphasic identity beginning with M1 during the acute phase of infection and then changing to M2 during the chronic phase. Depending on the virus, macrophage polarization plays a variety of roles during viral infection. For example, HIV-1-induced polarization has been shown to influence macrophage susceptibility to infection [23]. Also, studies on the Epstein Barr virus have shown that M1 macrophage polarization persists even in asymptomatic patients, despite the presence of anti-inflammatory cytokines like IL-10 and TGF-β [24]. Moreover, Japanese encephalitis virus and dengue virus can cause microglia (macrophages from CNS) and infiltrated macrophages to undergo M1 polarization and related proinflammatory activation in mice, and it has been proposed that targeting the occurrence of type 1 immunity may alleviate the pathologically lethal effect of viral encephalitis [25]. Furthermore, during COVID-19, both classically polarized macrophages (M1) and alternatively polarized macrophages (M2) inhibit SARS-CoV-2 infection. However, upon viral infection, M1 and non-activated (M0) macrophages, but not M2 macrophages, significantly up-regulate inflammatory factors [26]. This up-regulation of inflammatory factors is undoubtedly a significant contributor to the inflammation observed in COVID-19 [27]. Additionally, recent studies on the street rabies virus (RABV) have demonstrated its capability to affect macrophage polarization, shifting the macrophages toward an M2-c phenotype. Remarkably, the authors also discovered that a RABV glycoprotein can activate the α7-nAChR in monocyte-derived macrophages (MDMs), thus triggering the cholinergic anti-inflammatory response (CAR) [28]. These collective findings suggest that the RABV can induce an anti-inflammatory phenotype in human macrophages, potentially impacting the functioning of T cells [28]. On the other hand, the finding that viral proteins can bind to and activate α7-nAChR is highly significant as it reveals the complexity of viral infections in relation to inflammation. It also sheds light on a relatively unexplored area where only a few authors have delved into studying the potential interactions between viral proteins and the anti-inflammatory α7-nAChR. Indeed, viruses such as SARS-CoV-2 [29,30], and RABV[28] have been among the few viruses investigated in this context. It appears that the activation of α7-nAChR is not universally beneficial, as its activation in HIV-1 infected CD4+ T cells promotes HIV-1 transcription [31]. This observation highlights the complex and context-dependent nature of α7-nAChR signaling in different cellular environments. For comprehensive information on macrophage polarization during viral infections, particularly in noncholinergic settings, we highly recommend consulting the insightful reviews authored by Yu et al. [22] and Atmeh et al. [32].
It is not always the virus itself that polarizes macrophages, but rather viral proteins or cytokines released during infection. This is the case with HIV, where, in addition to the polarizing activity of granulocyte-macrophage colony-stimulating factor (M1) and macrophage colony-stimulating factor (M2) [33,34], the viral protein nef polarizes macrophages to an M1-like phenotype [35]. Similarly, hepatitis C virus core protein engagement with Toll-like receptor 2 of macrophages inhibits M2a, M2b, and M2c macrophage polarization [36]. Furthermore, in the case of a respiratory syncytial virus (RSV) infection, the polarization of alveolar macrophages occurs as a result of cytokines (IFN and GM-CSF), intercellular communication via the Notch-Jagged pathway, and RSV's direct activation signal [37]. Interestingly, the soluble spike protein of SARS-CoV-2 has recently gained prominence as a potential culprit in the deregulation of macrophage polarization via the α7-nAChR in COVID-19 [38]. The latter is crucial because the activation of this cholinergic ion channel in macrophages plays a significant role in reducing the production of proinflammatory cytokines in these cells [6]. The well-known CAR is responsible for achieving this effect, emphasizing the significance of this nAChR in suppressing inflammation during disease states.
3. The Expression of nAChRs and the Potential Role of α7-nAChR in Shaping Macrophage Polarization through the Cholinergic Anti-Inflammatory Response
3.1. Expression of nAChRs in Macrophages
Although the presence of neuronal nAChRs in non-neuronal cells, such as macrophages, has long been recognized, we are only now beginning to gain a better understanding of their precise roles and functions. Indeed, we know very little about how these receptors function in macrophages in both health and disease. Macrophages express α1, α2, α3, α4, α5, α6, α7, α9, α10, β1, β2, β3, and β4 subunits [39], resulting in the assembly of various stoichiometries. The expression of nAChRs in macrophages has been extensively studied in humans using primary cultures and representative cell lines from various body tissues. Likewise, macrophages have been extensively examined in a wide range of tissues and cell lines from rats and mice. In recent years, there has been increasing understanding of the roles and functions of nAChRs in macrophages [40,41]. The α7-nAChR is by far the most studied nAChR in human macrophages, as it is known that α7-nAChR-expressing macrophages are effectors of the cholinergic anti-inflammatory pathway, with activation resulting in CAR activation and decreased cytokine production [6], ultimately mitigating inflammation.
In human MDMs, the ion-conducting action of α7-nAChR appears to be regulated by intracellular proteins that restrict its ion-translocation activity [42]. This suggests that the physiological and natural activation of α7-nAChR depends on intracellular signaling pathways, rather than solely relying on ion conduction through its function as an ion channel. In line with this, recent studies suggest that the α7-nAChR can engage in G protein-coupled receptor-like signaling [43,44,45,46], independent of ion transport through it. In the context of macrophages, this area remains relatively unexplored. For instance, it merits deeper investigation of whether the α7-nAChR is capable of ion transport at the physiological concentrations of endogenous agonists (such as ACh and choline) naturally present in our body or during disease state. Also, it is worth investigating whether exogenous α7-nAChR agonists, such as nicotine, can activate this receptor at concentration levels commonly observed in cigarette smokers. This investigation would provide valuable insights into the potential modulation of α7-nAChR activity by endogenous and external agents and their impact on macrophage polarization and inflammatory response. Surprisingly, smoking is protective in ulcerative colitis, possibly owing to nicotine’s activating α7-nAChRs, causing polarization into M2 macrophages, upregulation of IL-10, and downregulation of IL-6 and TNF-α [47]. Remarkably, only two studies have successfully recorded electrophysiological currents from α7-nAChR in human macrophages [42,48]. This emphasizes the complexity associated with the intracellular regulation of α7-nAChR activity in macrophages and its response to agonists.
Macrophages are known for their significant cytokine production capacity, and they also possess an activatable cholinergic machinery capable of mitigating or inhibiting cytokine production through the CAR. This innate immune response relies on the expression of the α7-nAChR in macrophages. This receptor is physiologically activated by ACh and choline, which are endogenous agonists synthesized by other immune cells [49,50] or by macrophages themselves [39,51]. The efferent vagus nerve can also release ACh to activate the α7-nAChR. The net effect of α7-nAChR activation is that it prevents macrophages from overproducing cytokines [6], allowing inflammation to be controlled through the CAR. However, bacterial [52] and viral infections, such as HIV [53,54], seem to disrupt this delicate innate balance. Therefore, it is not surprising that inflammation becomes a prominent feature in the central axis of bacterial and viral infections.
3.2. Cholinergic Machinery of Macrophages
ACh has a number of effects on immune cells, with the macrophage playing a key role in dampening inflammation via the CAR. The most common components of the cholinergic system present in immune cells are choline acetyl-transferase (ChAT), vesicular ACh transporters (VAChT), acetylcholinesterase (AChE), and nAChRs. ChAT and VAChT expression has been demonstrated in human alveolar macrophages, lung macrophages, monocytes, and monocyte-derived macrophages (MDMs) [55,56,57], but there is still work to be done to establish the presence of ChAT in macrophages that reside in other tissues and their ability to synthesize/secrete ACh. However, the coexpression of ChAT and VAChT strongly suggests that macrophages have the potential to synthesize and secrete ACh in the lungs and bloodstream. Still in question is whether ACh is stored in macrophages or is produced and immediately released without any storage. In terms of mode of action, experiments in macrophage cell lines have shown that ACh can act in nAChRs in a paracrine and autocrine manner to reduce cytokines production [39]. AChE enters the picture when it comes time to hydrolyze ACh and end its activity. AChE has been found in human immune cells such as mononuclear lymphocytes and T cells [58], and in human macrophages [59]. Interestingly, α7-nAChR has been shown to interact with AChE in mouse macrophages, which appears to contribute to the reduction of the inflammatory response in macrophages, implicating this enzyme in active anti-inflammatory processes [60]. The cholinergic activation initiated by ACh or choline has the ability to influence the macrophage phenotype. Specifically, it can transform an inflammatory macrophage (M1) into an anti-inflammatory one (M2) by activating the CAR. As a result, it is plausible to consider that this phenotypic conversion, aimed at counteracting excessive inflammation, is also associated with macrophage polarization and subsequent differentiation into the various subpopulations (M3, M4, Mhem, M(Hb), HA-mac, and Mox) that have been identified thus far (see Figure 2 and Figure 3).
3.3. The Cholinergic Anti-Inflammatory Pathway (CAP) Is Closely Associated with the Polarization of Macrophages
In recent years, there has been a growing understanding of the changes in macrophage phenotype following CAR activation. In vivo studies conducted in rats have demonstrated that vagus nerve stimulation, resulting in the release of ACh, leads to a conversion of lung macrophages from the M1 to the M2 phenotype. This conversion is dependent on CAR activation and involves the participation of STAT3 [61]. In murine bone marrow-derived macrophages, it has been shown that the stimulation of α7nAChR activates the STAT3 pro-survival pathway, which serves as a protective mechanism against endoplasmic reticulum stress-induced apoptosis [62]. Additionally, studies conducted on rats with myocardial infarction have demonstrated that the administration of a specific α7-nAChR agonist (PNU 282,987) reduces the infiltration of inflammatory M1 macrophages into the infarcted area and increases the recruitment of M2-type anti-inflammatory peripheral macrophages to the infarcted tissue [63]. Moreover, in vitro studies using rodent cells focused on atherosclerosis have shown that CAP agonists (ACh and GTS-21) suppress M1 macrophage polarization while promoting M2 macrophage polarization through the upregulation of TNFAIP3 and phosphorylation of STAT3 [64]. This response strongly supports the presence of anti-inflammatory macrophages (M2) to counteract atheroinflammation. Importantly, this study provides evidence that CAP plays a role in the development of atherosclerosis by regulating macrophage function and promoting polarization toward anti-inflammatory (M2) phenotypes. Furthermore, another study on atherosclerosis using mouse peritoneal macrophages has reported that macrophage polarization is associated with the expression of nAChRs, specifically with the suppression of α4β2-nAChR expression in M1 macrophages [65]. One of the most compelling pieces of evidence supporting the medullary role of the α7-nAChR is the study on preeclampsia conducted by Han et al. [66]. This study provides strong evidence that women with preeclampsia exhibit downregulated levels of α7-nAChR in their decidual macrophages, which is accompanied by a decrease in the number of M2 phenotype macrophages and an increase in the number of M1 phenotype macrophages. The findings of this study demonstrate that the transition from the anti-inflammatory (M2) phenotype to the pro-inflammatory (M1) phenotype in macrophages can occur simply by reducing α7-nAChR levels. The polarization of macrophages appears to have broad implications, as they have also been implicated in the anti-inflammatory effects of acupuncture [67] and electroacupuncture [68].
The precise immunological identity (M1-4) of the macrophages involved in the CAR remains under study. This raises the question of whether M2 macrophages are more avid to activation by ACh or choline as compared to M1 macrophages. Given the vast diversity of macrophage subpopulations upon polarization, it is highly improbable that they all exhibit the same response to endogenous ACh and choline. In fact, both published and unpublished findings from our group agree as to the heterogeneity of their responses. For example, electrophysiological studies have demonstrated that only a fraction of MDMs exhibit electrophysiological currents indicative of the α7-nAChR. It is worth noting that the prevalence of these currents varies across different electrophysiological configurations, with approximately 15% observed in outside-out recordings, around 30% in inside-out recordings, and approximately 40% in single-channel current measurements [42], and an even smaller percentage (≈10%) of these cells display calcium mobilization in response to ACh (unpublished results). Recent findings from Victor Tsetlin's group demonstrated similar results in human macrophages (THP-1). Patch-clamp experiments revealed the presence of currents in approximately 22% of the tested cells [48]. Another crucial consideration is the distinct levels of α7-nAChR expression among different macrophage populations. Could it be that macrophages expressing higher levels of α7-nAChR demonstrate a stronger anti-inflammatory response upon ACh/choline activation? Currently, we lack an answer to this question. However, what we do know is that elevated levels of α7-nAChR induced by the HIV protein gp120 do not potentiate the anti-inflammatory response (CAR), suggesting a disruption of the CAR [53]. From early stages, these cells already exhibit glimpses of their cellular and functional complexity. We are referring to monocytes, the precursors of macrophages. Studies have shown that primary monocytes can be categorized into two distinct populations based on their nAChR expression levels [53]. The phenotype of these cells upon differentiation into macrophages remains unknown. Additionally, we have yet to determine whether there are differences in the expression and functionality of the cholinergic machinery, particularly the anti-inflammatory α7-nAChR, among macrophages. In summary, the precise cellular basis of CAR remains under study. Gaining a better understanding of these processes would offer valuable insights into the pharmacological modulation of inflammation through the α7-nAChR expressed by macrophages.
3.4. Experimental Challenges in Testing the Cholinergic Anti-Inflammatory Response in Macrophages: The Choline Issue
There is a significant challenge regarding choline when studying human macrophages in vitro, whether using primary cultures, macrophages or monocytic cancer cell lines. Choline is a precursor and metabolite of ACh. Because endogenous choline serves as an agonist of the α7-nAChR, it is important to know the concentrations present in the culture medium where these cells are grown and to monitor the accumulation of choline during cell culture to avoid desensitization of the α7-nAChR [69]. While macrophages are being cultured, choline can accumulate in the culture medium for various reasons. The first may be due to the release from cellular components. During cell growth and turnover, cellular components containing choline, such as phospholipids, may be broken down and release choline into the cell culture media. This process could contribute to the increased choline levels observed over time. Another possible reason for choline accumulation in the culture medium may be due to choline metabolism by macrophages. Macrophages have enzymatic pathways that metabolize choline, and these pathways may become more active as the cells respond to environmental cues or specific stimuli. The increased metabolic activity could result in higher choline levels as a byproduct or as part of intracellular signaling processes. It is worth mentioning that another important caution is that primary cells such as MDMs and cancer cell lines vary greatly in terms of choline metabolism whereas cancer cells exhibit altered choline metabolism [70]. Therefore, the extrapolation of the results between both types of cells when studying inflammation or cell polarization must be done with care. Of note, it was recently discovered that the uptake and metabolism of choline are important for macrophage inflammation because the polarization of primary bone marrow macrophages with lipopolysaccharide (LPS) resulted in an increased rate of choline uptake and higher levels of phosphatidylcholine synthesis. Furthermore, choline uptake and metabolism modulate macrophage IL-1β and IL-18 production [71]. Therefore, could macrophages incorporate choline into themselves for anti-inflammatory purposes? This newly incorporated choline can easily be used by the macrophage to synthesize acetylcholine and release it autocrinely and paracrinely [39].
In the body, the systemic and local concentration of choline varies depending on the organ where it is measured and the physical status of the person. In healthy adults, the concentration of choline in plasma ranges from 7 to 20 micromol/L [72] whereas that of sick people is usually high [73,74,75] or very low [76]. The desensitizing concentration of choline reported for neuronal α7-nAChR in rats is at concentrations above 10 µM [69,77]. Therefore, if it is desired to study the CAR in MDMs in vitro, care should be taken to keep the choline concentration away from 10 uM to activate the α7-nAChR effectively. It turns out that the accumulation of choline during the cultures of MDMs occurs naturally, even when they are not exposed to any treatment (Figure 4). For example, our studies testing the cholinergic anti-inflammatory response in MDMs (primary culture) exposed to the HIV-1 viral protein, gp120IIB, demonstrate a time-dependent accumulation of choline. Said accumulation occurs irrespective of the application of gp120IIIB, pyridostigmine, or the co-application of both (Figure 4). Importantly, in these experiments, the culture medium already included choline. After evaluating four different preparations using four different lots of the base (RPMI-1640), fetal bovine serum, and human serum, we identified that the average initial concentration of choline is 113.70 µM ± 0.2. Thus, the concentration of choline exceeds the desensitizing concentration for the α7-nAChR, 10 uM.
Therefore, when it is desired to stimulate the α7-nAChR to activate CAR in macrophages, the previous medium should be removed and the remainder washed and replaced with fresh low-choline medium to ensure that the α7-nAChR can be stimulated with the agonist of interest. To activate α7-nAChR, ACh, nicotine, or more elaborate agonists such as GTS-21 and PNU 282,987 are typically used. The application of ACh must be preceded by treatment with pyridostigmine to prevent acetylcholinesterase from hydrolyzing ACh and converting it into choline and acetate, thus contributing to increasing choline concentration at the expense of ACh.
3.5. Function and Role of CHRFAM7A in Macrophage Polarization and Cholinergic Antiinflammatory Pathway Operation
CHRFAM7A is a gene that encodes a protein (called duplicated or dup α7) found in humans, specifically in the brain and immune cells such as macrophages. This fusion gene results from a gene duplication event [78] involving the ULK4 (Unc51-like kinase-4) and the FAM7A genes [79]. CHRFAM7A has been of interest due to its potential involvement in various physiological processes and diseases. While its exact function is not yet fully understood, studies suggest that CHRFAM7A may play a role in modulating cholinergic signaling pathways, including those related to inflammation and cognitive function. According to the available information, CHRFAM7A functions as a negative regulator of the expression and activity of the α7-nAChR [80], thereby diminishing the anti-inflammatory activity in macrophages. Theoretically, this suggests that elevated levels of CHRFAM7A lead to an inflammatory phenotype (M1) in the impacted cell, as it inhibits the activity of the α7-nAChR. In line with this idea, a recent report by Li et al. indicated that the expression of CHRFAM7A was identified as a promoter of inflammation [81]. In a more recent study, transgenic CHRFAM7A mice were employed to examine the impact of this human-specific gene on the development of knee osteoarthritis. The findings revealed that CHRFAM7A serves as an exacerbating factor in inflammation and tissue damage associated with osteoarthritis [82]. Interestingly, studies conducted with primary macrophages have revealed that co-expression of CHRNA7 (gene coding for α7-nAChR) and CHRFAM7A leads to a greater anti-inflammatory response upon stimulation with lipopolysaccharide (LPS), as compared to macrophages expressing only one of these genes [83]. This indicates a potential modulation or regulation between the two proteins. Similar evidence has also emerged from studies carried out in humanized mouse models of radiation-induced lacrimal gland injury, involving cell types other than macrophages [84]. These studies demonstrate that elevating CHRFAM7A levels yields beneficial outcomes in these contexts. These findings emphasize the relevance of CHRFAM7A in modulating the inflammatory response mediated by α7-nAChR. This also highlights its significance as a factor to be taken into account during the development and evaluation of drugs targeting α7-nAChR to dampen inflammation.
The role of CHRFAM7A in macrophages extends beyond inflammation and encompasses macrophage polarization as well. Studies have demonstrated that CHRFAM7A facilitates M2 macrophage polarization through the Notch pathway, ameliorating hypertrophic scar formation in humans [81]. Furthermore, in a mouse model of renal fibrosis, the overexpression of CHRFAM7A was found to suppress M1 macrophage activation following unilateral ureteral obstruction (UUO) when compared to wild-type mice. Conversely, the expression of M2 macrophage markers such as CD206 and FIZZ1 was elevated in mice overexpressing CHRFAM7A after UUO, indicating an increased transition of macrophages from an M1 to an M2 phenotype [85]. Nevertheless, in contrast to the findings in the hypertrophic scar mouse model [81], overexpression of CHRFAM7A in the central nervous system (CNS) has been shown to mitigate cerebral ischemia-reperfusion injury. This effect is achieved through the inhibition of microglia pyroptosis, which is mediated by the NLRP3/Caspase-1 pathway [86]. Consequently, the upregulation of CHRFAM7A expression in macrophages proved to be anti-inflammatory.
Collectively, the existing findings indicate that the impact of CHRFAM7A on inflammation is cell-dependent. However, comprehensive studies, particularly in human subjects, are limited. Further investigations are required to clarify/elucidate its cellular function and the extent of its involvement in inflammation before considering pharmacological modulation of its activity or expression.
4. Conclusions and Future Directions
The information presented here clearly highlights the fundamental role of macrophages in regulating inflammatory processes, with their polarization being associated with the functioning of the CAR, particularly on M1 and M2 phenotypes. However, the role of CAR in the differentiation of M0 macrophages to more sophisticated phenotypes like M3, M4, Mhem, M(Hb), HA-mac, and Mox remains unknown, and the exact order of polarization is yet to be determined. In response to a stimulating signal or insult, the CAR may polarize macrophages, or macrophages may autopolarize by releasing autocrine polarizing substances, such as ACh [39], which are capable of activating the CAR. The available literature suggests that macrophage polarization depends on the specific insult activating it (virus, bacteria, parasite, fungus, etc.), the macrophage's location at the time of activation, and the surrounding biochemical microenvironment. Within this chemical environment, endogenous agonists (ACh and choline) released by the body activate α7-nAChR. Studies on influenza [87] and bacterial infections, like Mycobacterium tuberculosis [88], have shown increased ACh levels as infections progress, indicating a more complex scenario where ACh release by both host and pathogen may lead to desensitization of α7-nAChR in macrophages and CAR activation. Interestingly, ACh release has also been reported to have bactericidal properties, further influencing host defenses against pathogens [89]. Moreover, recent discoveries, such as ACh functioning as a chemotactic agent for certain bacteria, have opened new scientific directions and added complexity to the intricate mechanisms influencing CAR activity in macrophages [90]. These advancements pave the way for innovative approaches to treat inflammatory problems.
From a therapeutic standpoint, α7-nAChR emerges as a promising option for modulating the CAR in macrophages to counteract local and systemic inflammation. However, optimizing its activation remains a subject of investigation, and understanding the role of CHRFAM7A in the pharmacological activation of α7-AChR is critical. It is possible that α7-nAChR may or may not require its inherent ion translocation ability and instead function primarily as a GPCR to exhibit an anti-inflammatory phenotype. Additionally, it is worth considering that the anti-inflammatory profile of macrophages (M2) may not solely rely on the activation of α7-nAChR, as other nAChRs, such as α4β2, also appear to play a contributory role. Therefore, when formulating therapeutic strategies, both α7-nAChR and α4β2 should be taken into account for a comprehensive approach. In conclusion, the future shows significant promise for pharmacologically harnessing the cholinergic machinery expressed by macrophages, and ongoing efforts in the field are moving in that direction.
Funding
The authors thank the National Institute of General Medical Sciences (NIGMS) of the National Institutes of Health (NIH) under Award Number P20GM103642, and the COBRE NIEF Grant: 5P20GM103642-100. Also, we recognize the important funding provided by the NIGMS RISE Program (2R25GM061151-5A1) to MD-V and CMC-V.
Acknowledgments
We extend our genuine gratitude to Mr. Bismark Madera for his invaluable assistance in conducting the calcium mobilization experiments mentioned in this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chung, E.Y.; Kim, S.J.; Ma, X.J. Regulation of Cytokine Production during Phagocytosis of Apoptotic Cells. Cell Res 2006, 16, 154–161. [CrossRef]
- Fagundes, D.L.G.; França, E.L.; Morceli, G.; Rudge, M.V.C.; Calderon, I. de M.P.; Honorio-França, A.C. The Role of Cytokines in the Functional Activity of Phagocytes in Blood and Colostrum of Diabetic Mothers. Journal of Immunology Research 2013, 2013, e590190. [CrossRef]
- Fadok, V.A.; McDonald, P.P.; Bratton, D.L.; Henson, P.M. Regulation of Macrophage Cytokine Production by Phagocytosis of Apoptotic and Post-Apoptotic Cells. Biochemical Society Transactions 1998, 26, 653–656. [CrossRef]
- Geiger, S.S.; Curtis, A.M.; O’Neill, L.A.J.; Siegel, R.M. Daily Variation in Macrophage Phagocytosis Is Clock-Independent and Dispensable for Cytokine Production. Immunology 2019, 157, 122–136. [CrossRef]
- Fu, Y.L.; Harrison, R.E. Microbial Phagocytic Receptors and Their Potential Involvement in Cytokine Induction in Macrophages. Frontiers in Immunology 2021, 12.
- Wang, H.; Yu, M.; Ochani, M.; Amella, C.A.; Tanovic, M.; Susarla, S.; Li, J.H.; Wang, H.; Yang, H.; Ulloa, L.; et al. Nicotinic Acetylcholine Receptor Alpha7 Subunit Is an Essential Regulator of Inflammation. Nature 2003, 421, 384–388. [CrossRef]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H.; Botchkina, G.I.; Watkins, L.R.; Wang, H.; Abumrad, N.; Eaton, J.W.; Tracey, K.J. Vagus Nerve Stimulation Attenuates the Systemic Inflammatory Response to Endotoxin. Nature 2000, 405, 458–462. [CrossRef]
- Colin, S.; Chinetti-Gbaguidi, G.; Staels, B. Macrophage Phenotypes in Atherosclerosis. Immunol Rev 2014, 262, 153–166. [CrossRef]
- De Paoli, F.; Staels, B.; Chinetti-Gbaguidi, G. Macrophage Phenotypes and Their Modulation in Atherosclerosis. Circ J 2014, 78, 1775–1781. [CrossRef]
- Mushenkova, N.V.; Nikiforov, N.G.; Melnichenko, A.A.; Kalmykov, V.; Shakhpazyan, N.K.; Orekhova, V.A.; Orekhov, A.N. Functional Phenotypes of Intraplaque Macrophages and Their Distinct Roles in Atherosclerosis Development and Atheroinflammation. Biomedicines 2022, 10, 452. [CrossRef]
- Kadl, A.; Meher, A.K.; Sharma, P.R.; Lee, M.Y.; Doran, A.C.; Johnstone, S.R.; Elliott, M.R.; Gruber, F.; Han, J.; Chen, W.; et al. Identification of a Novel Macrophage Phenotype That Develops in Response to Atherogenic Phospholipids via Nrf2. Circ Res 2010, 107, 737–746. [CrossRef]
- Erbel, C.; Tyka, M.; Helmes, C.M.; Akhavanpoor, M.; Rupp, G.; Domschke, G.; Linden, F.; Wolf, A.; Doesch, A.; Lasitschka, F.; et al. CXCL4-Induced Plaque Macrophages Can Be Specifically Identified by Co-Expression of MMP7+S100A8+in Vitro and in Vivo. Innate Immun 2015, 21, 255–265. [CrossRef]
- Chinetti-Gbaguidi, G.; Colin, S.; Staels, B. Macrophage Subsets in Atherosclerosis. Nat Rev Cardiol 2015, 12, 10–17. [CrossRef]
- Malyshev, I.; Malyshev, Y. Current Concept and Update of the Macrophage Plasticity Concept: Intracellular Mechanisms of Reprogramming and M3 Macrophage “Switch” Phenotype. Biomed Res Int 2015, 2015, 341308. [CrossRef]
- Kalish, S.; Lyamina, S.; Manukhina, E.; Malyshev, Y.; Raetskaya, A.; Malyshev, I. M3 Macrophages Stop Division of Tumor Cells In Vitro and Extend Survival of Mice with Ehrlich Ascites Carcinoma. Med Sci Monit Basic Res 2017, 23, 8–19. [CrossRef]
- Jackaman, C.; Yeoh, T.L.; Acuil, M.L.; Gardner, J.K.; Nelson, D.J. Murine Mesothelioma Induces Locally-Proliferating IL-10(+)TNF-α(+)CD206(-)CX3CR1(+) M3 Macrophages That Can Be Selectively Depleted by Chemotherapy or Immunotherapy. Oncoimmunology 2016, 5, e1173299. [CrossRef]
- Gleissner, C.A.; Shaked, I.; Erbel, C.; Böckler, D.; Katus, H.A.; Ley, K. CXCL4 Downregulates the Atheroprotective Hemoglobin Receptor CD163 in Human Macrophages. Circulation Research 2010, 106, 203–211. [CrossRef]
- Gleissner, C.A.; Shaked, I.; Little, K.M.; Ley, K. CXC Chemokine Ligand 4 Induces a Unique Transcriptome in Monocyte-Derived Macrophages. The Journal of Immunology 2010, 184, 4810–4818. [CrossRef]
- Erbel, C.; Wolf, A.; Lasitschka, F.; Linden, F.; Domschke, G.; Akhavanpoor, M.; Doesch, A.O.; Katus, H.A.; Gleissner, C.A. Prevalence of M4 Macrophages within Human Coronary Atherosclerotic Plaques Is Associated with Features of Plaque Instability. Int J Cardiol 2015, 186, 219–225. [CrossRef]
- Real, F.; Zhu, A.; Huang, B.; Belmellat, A.; Sennepin, A.; Vogl, T.; Ransy, C.; Revol, M.; Arrigucci, R.; Lombès, A.; et al. S100A8-Mediated Metabolic Adaptation Controls HIV-1 Persistence in Macrophages in Vivo. Nat Commun 2022, 13, 5956. [CrossRef]
- Maretti-Mira, A.C.; Golden-Mason, L.; Salomon, M.P.; Kaplan, M.J.; Rosen, H.R. Cholesterol-Induced M4-Like Macrophages Recruit Neutrophils and Induce NETosis. Front Immunol 2021, 12, 671073. [CrossRef]
- Yu, S.; Ge, H.; Li, S.; Qiu, H.-J. Modulation of Macrophage Polarization by Viruses: Turning Off/On Host Antiviral Responses. Front Microbiol 2022, 13, 839585. [CrossRef]
- Cassol, E.; Cassetta, L.; Rizzi, C.; Alfano, M.; Poli, G. M1 and M2a Polarization of Human Monocyte-Derived Macrophages Inhibits HIV-1 Replication by Distinct Mechanisms. J Immunol 2009, 182, 6237–6246. [CrossRef]
- Moyano, A.; Ferressini Gerpe, N.M.; De Matteo, E.; Preciado, M.V.; Chabay, P. M1 Macrophage Polarization Prevails in Epstein-Barr Virus-Infected Children in an Immunoregulatory Environment. Journal of Virology 2022, 96, e01434-21. [CrossRef]
- Jhan, M.-K.; Chen, C.-L.; Shen, T.-J.; Tseng, P.-C.; Wang, Y.-T.; Satria, R.D.; Yu, C.-Y.; Lin, C.-F. Polarization of Type 1 Macrophages Is Associated with the Severity of Viral Encephalitis Caused by Japanese Encephalitis Virus and Dengue Virus. Cells 2021, 10, 3181. [CrossRef]
- Lian, Q.; Zhang, K.; Zhang, Z.; Duan, F.; Guo, L.; Luo, W.; Mok, B.W.-Y.; Thakur, A.; Ke, X.; Motallebnejad, P.; et al. Differential Effects of Macrophage Subtypes on SARS-CoV-2 Infection in a Human Pluripotent Stem Cell-Derived Model. Nat Commun 2022, 13, 2028. [CrossRef]
- Tan, L.Y.; Komarasamy, T.V.; RMT Balasubramaniam, V. Hyperinflammatory Immune Response and COVID-19: A Double Edged Sword. Frontiers in Immunology 2021, 12.
- Embregts, C.W.E.; Begeman, L.; Voesenek, C.J.; Martina, B.E.E.; Koopmans, M.P.G.; Kuiken, T.; GeurtsvanKessel, C.H. Street RABV Induces the Cholinergic Anti-Inflammatory Pathway in Human Monocyte-Derived Macrophages by Binding to NAChr A7. Front Immunol 2021, 12, 622516. [CrossRef]
- Chrestia, J.F.; Oliveira, A.S.; Mulholland, A.J.; Gallagher, T.; Bermúdez, I.; Bouzat, C. A Functional Interaction Between Y674-R685 Region of the SARS-CoV-2 Spike Protein and the Human A7 Nicotinic Receptor. Mol Neurobiol 2022, 59, 6076–6090. [CrossRef]
- O’Brien, B.C.V.; Weber, L.; Hueffer, K.; Weltzin, M.M. SARS-CoV-2 Spike Ectodomain Targets A7 Nicotinic Acetylcholine Receptors. Journal of Biological Chemistry 2023, 104707. [CrossRef]
- Wen, J.; Zhao, C.; Chen, J.; Song, S.; Lin, Z.; Xie, S.; Qi, H.; Wang, J.; Su, X. Activation of A7 Nicotinic Acetylcholine Receptor Promotes HIV-1 Transcription. Cell Insight 2022, 1, 100028. [CrossRef]
- Atmeh, P.A.; Mezouar, S.; Mège, J.-L.; Atmeh, P.A.; Mezouar, S.; Mège, J.-L. Macrophage Polarization in Viral Infectious Diseases: Confrontation with the Reality. In Macrophages - Celebrating 140 Years of Discovery; IntechOpen, 2022 ISBN 978-1-80355-625-3.
- Cassetta, L.; Cassol, E.; Poli, G. Macrophage Polarization in Health and Disease. ScientificWorldJournal 2011, 11, 2391–2402. [CrossRef]
- Herbein, G.; Varin, A. The Macrophage in HIV-1 Infection: From Activation to Deactivation? Retrovirology 2010, 7, 33. [CrossRef]
- Chihara, T.; Hashimoto, M.; Osman, A.; Hiyoshi-Yoshidomi, Y.; Suzu, I.; Chutiwitoonchai, N.; Hiyoshi, M.; Okada, S.; Suzu, S. HIV-1 Proteins Preferentially Activate Anti-Inflammatory M2-Type Macrophages. J Immunol 2012, 188, 3620–3627. [CrossRef]
- Zhao, S.; Si, M.; Deng, X.; Wang, D.; Kong, L.; Zhang, Q. HCV Inhibits M2a, M2b and M2c Macrophage Polarization via HCV Core Protein Engagement with Toll-like Receptor 2. Exp Ther Med 2022, 24, 522. [CrossRef]
- Wang, Y.; Zheng, J.; Wang, X.; Yang, P.; Zhao, D. Alveolar Macrophages and Airway Hyperresponsiveness Associated with Respiratory Syncytial Virus Infection. Frontiers in Immunology 2022, 13.
- Tanmay, S.; Labrou, D.; Farsalinos, K.; Poulas, K. Is SARS-CoV-2 Spike Glycoprotein Impairing Macrophage Function via A7-Nicotinic Acetylcholine Receptors? Food Chem Toxicol 2021, 152, 112184. [CrossRef]
- Chernyavsky, A.I.; Arredondo, J.; Skok, M.; Grando, S.A. Auto/Paracrine Control of Inflammatory Cytokines by Acetylcholine in Macrophage-like U937 Cells through Nicotinic Receptors. Int. Immunopharmacol. 2010, 10, 308–315. [CrossRef]
- Mashimo, M.; Moriwaki, Y.; Misawa, H.; Kawashima, K.; Fujii, T. Regulation of Immune Functions by Non-Neuronal Acetylcholine (ACh) via Muscarinic and Nicotinic ACh Receptors. IJMS 2021, 22, 6818. [CrossRef]
- Richter, K.; Grau, V. Signaling of Nicotinic Acetylcholine Receptors in Mononuclear Phagocytes. Pharmacological Research 2023, 191, 106727. [CrossRef]
- Báez-Pagán, C.A.; Delgado-Vélez, M.; Lasalde-Dominicci, J.A. Activation of the Macrophage A7 Nicotinic Acetylcholine Receptor and Control of Inflammation. J Neuroimmune Pharmacol 2015. [CrossRef]
- Kabbani, N.; Nichols, R.A. Beyond the Channel: Metabotropic Signaling by Nicotinic Receptors. Trends Pharmacol Sci 2018, 39, 354–366. [CrossRef]
- Kabbani, N.; Nordman, J.C.; Corgiat, B.A.; Veltri, D.P.; Shehu, A.; Seymour, V.A.; Adams, D.J. Are Nicotinic Acetylcholine Receptors Coupled to G Proteins? Bioessays 2013, 35, 1025–1034. [CrossRef]
- King, J.R.; Kabbani, N. Alpha 7 Nicotinic Receptor Coupling to Heterotrimeric G Proteins Modulates RhoA Activation, Cytoskeletal Motility, and Structural Growth. J. Neurochem. 2016, 138, 532–545. [CrossRef]
- King, J.R.; Gillevet, T.C.; Kabbani, N. A G Protein-Coupled A7 Nicotinic Receptor Regulates Signaling and TNF-α Release in Microglia. FEBS Open Bio 2017, 7, 1350–1361. [CrossRef]
- AlQasrawi, D.; Qasem, A.; Naser, S.A. Divergent Effect of Cigarette Smoke on Innate Immunity in Inflammatory Bowel Disease: A Nicotine-Infection Interaction. Int J Mol Sci 2020, 21, 5801. [CrossRef]
- Siniavin, A.E.; Streltsova, M.A.; Kudryavtsev, D.S.; Shelukhina, I.V.; Utkin, Y.N.; Tsetlin, V.I. Activation of A7 Nicotinic Acetylcholine Receptor Upregulates HLA-DR and Macrophage Receptors: Potential Role in Adaptive Immunity and in Preventing Immunosuppression. Biomolecules 2020, 10. [CrossRef]
- Rosas-Ballina, M.; Olofsson, P.S.; Ochani, M.; Valdés-Ferrer, S.I.; Levine, Y.A.; Reardon, C.; Tusche, M.W.; Pavlov, V.A.; Andersson, U.; Chavan, S.; et al. Acetylcholine-Synthesizing T Cells Relay Neural Signals in a Vagus Nerve Circuit. Science 2011, 334, 98–101. [CrossRef]
- Reardon, C.; Duncan, G.S.; Brüstle, A.; Brenner, D.; Tusche, M.W.; Olofsson, P.; Rosas-Ballina, M.; Tracey, K.J.; Mak, T.W. Lymphocyte-Derived ACh Regulates Local Innate but Not Adaptive Immunity. Proc. Natl. Acad. Sci. U.S.A. 2013. [CrossRef]
- Knights, A.J.; Liu, S.; Ma, Y.; Nudell, V.S.; Perkey, E.; Sorensen, M.J.; Kennedy, R.T.; Maillard, I.; Ye, L.; Jun, H.; et al. Acetylcholine-Synthesizing Macrophages in Subcutaneous Fat Are Regulated by Β2 -Adrenergic Signaling. EMBO J 2021, 40, e106061. [CrossRef]
- Richter, K.; Koch, C.; Perniss, A.; Wolf, P.M.; Schweda, E.K.H.; Wichmann, S.; Wilker, S.; Magel, I.; Sander, M.; McIntosh, J.M.; et al. Phosphocholine-Modified Lipooligosaccharides of Haemophilus Influenzae Inhibit ATP-Induced IL-1β Release by Pulmonary Epithelial Cells. Molecules 2018, 23, 1979. [CrossRef]
- Delgado-Vélez, M.; Báez-Pagán, C.A.; Gerena, Y.; Quesada, O.; Santiago-Pérez, L.I.; Capó-Vélez, C.M.; Wojna, V.; Meléndez, L.; León-Rivera, R.; Silva, W.; et al. The A7-Nicotinic Receptor Is Upregulated in Immune Cells from HIV-Seropositive Women: Consequences to the Cholinergic Anti-Inflammatory Response. Clin Transl Immunology 2015, 4, e53. [CrossRef]
- Ríos, S.C.; Colón Sáez, J.O.; Quesada, O.; Figueroa, K.Q.; Lasalde Dominicci, J.A. Disruption of the Cholinergic Anti-Inflammatory Response by R5-Tropic HIV-1 Protein Gp120JRFL. J Biol Chem 2021, 100618. [CrossRef]
- Fujii, T.; Mashimo, M.; Moriwaki, Y.; Misawa, H.; Ono, S.; Horiguchi, K.; Kawashima, K. Expression and Function of the Cholinergic System in Immune Cells. Front Immunol 2017, 8, 1085. [CrossRef]
- Wessler, I.K.; Kirkpatrick, C.J. The Non-Neuronal Cholinergic System: An Emerging Drug Target in the Airways. Pulm Pharmacol Ther 2001, 14, 423–434. [CrossRef]
- Koarai, A.; Traves, S.L.; Fenwick, P.S.; Brown, S.M.; Chana, K.K.; Russell, R.E.K.; Nicholson, A.G.; Barnes, P.J.; Donnelly, L.E. Expression of Muscarinic Receptors by Human Macrophages. Eur Respir J 2012, 39, 698–704. [CrossRef]
- Fujii, T.; Mashimo, M.; Moriwaki, Y.; Misawa, H.; Ono, S.; Horiguchi, K.; Kawashima, K. Physiological Functions of the Cholinergic System in Immune Cells. J Pharmacol Sci 2017, 134, 1–21. [CrossRef]
- de Oliveira, P.; Gomes, A.Q.; Pacheco, T.R.; Vitorino de Almeida, V.; Saldanha, C.; Calado, A. Cell-Specific Regulation of Acetylcholinesterase Expression under Inflammatory Conditions. Clin Hemorheol Microcirc 2012, 51, 129–137. [CrossRef]
- Liu, E.Y.L.; Xia, Y.; Kong, X.; Guo, M.S.S.; Yu, A.X.D.; Zheng, B.Z.Y.; Mak, S.; Xu, M.L.; Tsim, K.W.K. Interacting with A7 NAChR Is a New Mechanism for AChE to Enhance the Inflammatory Response in Macrophages. Acta Pharm Sin B 2020, 10, 1926–1942. [CrossRef]
- Li, S.; Qi, D.; Li, J.-N.; Deng, X.-Y.; Wang, D.-X. Vagus Nerve Stimulation Enhances the Cholinergic Anti-Inflammatory Pathway to Reduce Lung Injury in Acute Respiratory Distress Syndrome via STAT3. Cell Death Discov 2021, 7, 63. [CrossRef]
- Lee, R.H.; Vazquez, G. Evidence for a Prosurvival Role of Alpha-7 Nicotinic Acetylcholine Receptor in Alternatively (M2)-activated Macrophages. Physiol Rep 2013, 1. [CrossRef]
- Niu, X.-H.; Liu, R.-H.; Lv, X.; He, R.-L.; Lv, F.-Z.; Wu, S.-J.; Li, X.-Q.; Li, L.; Lin, J.-F. Activating A7nAChR Helps Post-Myocardial Infarction Healing by Regulating Macrophage Polarization via the STAT3 Signaling Pathway. Inflamm Res 2023, 72, 879–892. [CrossRef]
- Qian, Z.; Yang, H.; Li, H.; Liu, C.; Yang, L.; Qu, Z.; Li, X. The Cholinergic Anti-Inflammatory Pathway Attenuates the Development of Atherosclerosis in Apoe-/- Mice through Modulating Macrophage Functions. Biomedicines 2021, 9, 1150. [CrossRef]
- Suzuki, M.; Katayama, T.; Suzuki, C.; Nakajima, K.; Magata, Y.; Ogawa, M. Uptake of Nicotinic Acetylcholine Receptor Imaging Agent Is Reduced in the Pro-Inflammatory Macrophage. Nucl Med Biol 2021, 102–103, 45–55. [CrossRef]
- Han, X.; Li, W.; Li, P.; Zheng, Z.; Lin, B.; Zhou, B.; Guo, K.; He, P.; Yang, J. Stimulation of A7 Nicotinic Acetylcholine Receptor by Nicotine Suppresses Decidual M1 Macrophage Polarization Against Inflammation in Lipopolysaccharide-Induced Preeclampsia-Like Mouse Model. Frontiers in Immunology 2021, 12.
- Wang, J.; Lu, S.; Yang, F.; Guo, Y.; Chen, Z.; Yu, N.; Yao, L.; Huang, J.; Fan, W.; Xu, Z.; et al. The Role of Macrophage Polarization and Associated Mechanisms in Regulating the Anti-Inflammatory Action of Acupuncture: A Literature Review and Perspectives. Chinese Medicine 2021, 16, 56. [CrossRef]
- Torres-Rosas, R.; Yehia, G.; Peña, G.; Mishra, P.; del Rocio Thompson-Bonilla, M.; Moreno-Eutimio, M.A.; Arriaga-Pizano, L.A.; Isibasi, A.; Ulloa, L. Dopamine Mediates Vagal Modulation of the Immune System by Electroacupuncture. Nat Med 2014, 20, 291–295. [CrossRef]
- Alkondon, M.; Pereira, E.F.; Cortes, W.S.; Maelicke, A.; Albuquerque, E.X. Choline Is a Selective Agonist of Alpha7 Nicotinic Acetylcholine Receptors in the Rat Brain Neurons. Eur. J. Neurosci. 1997, 9, 2734–2742.
- Roci, I.; Watrous, J.D.; Lagerborg, K.A.; Jain, M.; Nilsson, R. Mapping Choline Metabolites in Normal and Transformed Cells. Metabolomics 2020, 16, 125. [CrossRef]
- Sanchez-Lopez, E.; Zhong, Z.; Stubelius, A.; Sweeney, S.R.; Booshehri, L.M.; Antonucci, L.; Liu-Bryan, R.; Lodi, A.; Terkeltaub, R.; Lacal, J.C.; et al. Choline Uptake and Metabolism Modulate Macrophage IL-1β and IL-18 Production. Cell Metab 2019, 29, 1350-1362.e7. [CrossRef]
- Institute of Medicine (US) Standing Committee on the Scientific Evaluation of Dietary Reference Intakes and its Panel on Folate, Other B Vitamins, and Choline Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Choline; The National Academies Collection: Reports funded by National Institutes of Health; National Academies Press (US): Washington (DC), 1998; ISBN 978-0-309-06411-8.
- Song, M.; Xu, B.P.; Liang, Q.; Wei, Y.; Song, Y.; Chen, P.; Zhou, Z.; Zhang, N.; He, Q.; Liu, L.; et al. Association of Serum Choline Levels and All-Cause Mortality Risk in Adults with Hypertension: A Nested Case–Control Study. Nutrition & Metabolism 2021, 18, 108. [CrossRef]
- Elble, R.; Giacobini, E.; Higgins, C. Choline Levels Are Increased in Cerebrospinal Fluid of Alzheimer Patients. Neurobiology of Aging 1989, 10, 45–50. [CrossRef]
- Yang, Y.S.; Smucny, J.; Zhang, H.; Maddock, R.J. Meta-Analytic Evidence of Elevated Choline, Reduced N-Acetylaspartate, and Normal Creatine in Schizophrenia and Their Moderation by Measurement Quality, Echo Time, and Medication Status. Neuroimage Clin 2023, 39, 103461. [CrossRef]
- Judd, J.M.; Jasbi, P.; Winslow, W.; Serrano, G.E.; Beach, T.G.; Klein-Seetharaman, J.; Velazquez, R. Low Circulating Choline, a Modifiable Dietary Factor, Is Associated with the Pathological Progression and Metabolome Dysfunction in Alzheimer’s Disease. bioRxiv 2023, 2023.05.06.539713. [CrossRef]
- Mike, A.; Castro, N.G.; Albuquerque, E.X. Choline and Acetylcholine Have Similar Kinetic Properties of Activation and Desensitization on the Alpha7 Nicotinic Receptors in Rat Hippocampal Neurons. Brain Res 2000, 882, 155–168. [CrossRef]
- Gault, J.; Robinson, M.; Berger, R.; Drebing, C.; Logel, J.; Hopkins, J.; Moore, T.; Jacobs, S.; Meriwether, J.; Choi, M.J.; et al. Genomic Organization and Partial Duplication of the Human Alpha7 Neuronal Nicotinic Acetylcholine Receptor Gene (CHRNA7). Genomics 1998, 52, 173–185. [CrossRef]
- Riley, B.; Williamson, M.; Collier, D.; Wilkie, H.; Makoff, A. A 3-Mb Map of a Large Segmental Duplication Overlapping the Alpha7-Nicotinic Acetylcholine Receptor Gene (CHRNA7) at Human 15q13-Q14. Genomics 2002, 79, 197–209. [CrossRef]
- de Lucas-Cerrillo, A.M.; Maldifassi, M.C.; Arnalich, F.; Renart, J.; Atienza, G.; Serantes, R.; Cruces, J.; Sánchez-Pacheco, A.; Andrés-Mateos, E.; Montiel, C. Function of Partially Duplicated Human A77 Nicotinic Receptor Subunit CHRFAM7A Gene: Potential Implications for the Cholinergic Anti-Inflammatory Response. J. Biol. Chem. 2011, 286, 594–606. [CrossRef]
- Li, T.; Chen, W.; Zhang, Q.; Deng, C. Human-Specific Gene CHRFAM7A Mediates M2 Macrophage Polarization via the Notch Pathway to Ameliorate Hypertrophic Scar Formation. Biomedicine & Pharmacotherapy 2020, 131, 110611. [CrossRef]
- Courties, A.; Olmer, M.; Myers, K.; Ordoukhanian, P.; Head, S.R.; Natarajan, P.; Berenbaum, F.; Sellam, J.; Lotz, M.K. Human-Specific Duplicate CHRFAM7A Gene Is Associated with More Severe Osteoarthritis and Amplifies Pain Behaviours. Ann Rheum Dis 2023, 82, 710–718. [CrossRef]
- Maroli, A.; Di Lascio, S.; Drufuca, L.; Cardani, S.; Setten, E.; Locati, M.; Fornasari, D.; Benfante, R. Effect of Donepezil on the Expression and Responsiveness to LPS of CHRNA7 and CHRFAM7A in Macrophages: A Possible Link to the Cholinergic Anti-Inflammatory Pathway. J Neuroimmunol 2019, 332, 155–166. [CrossRef]
- Zhang, Y.; Qian, J.; Ren, H.; Meng, F.; Ma, R.; Xu, B. Human-Specific CHRFAM7A Protects against Radiotherapy-Induced Lacrimal Gland Injury by Inhibiting the P38/JNK Signalling Pathway and Oxidative Stress. Int J Clin Exp Pathol 2017, 10, 9001–9011.
- Zhou, B.; Zhang, Y.; Dang, X.; Li, B.; Wang, H.; Gong, S.; Li, S.; Meng, F.; Xing, J.; Li, T.; et al. Up-Regulation of the Human-Specific CHRFAM7A Gene Protects against Renal Fibrosis in Mice with Obstructive Nephropathy. Journal of Cellular and Molecular Medicine 2023, 27, 52–65. [CrossRef]
- Cao, X.; Wang, Y.; Gao, L. CHRFAM7A Overexpression Attenuates Cerebral Ischemia-Reperfusion Injury via Inhibiting Microglia Pyroptosis Mediated by the NLRP3/Caspase-1 Pathway. Inflammation 2021, 44, 1023–1034. [CrossRef]
- Horkowitz, A.P.; Schwartz, A.V.; Alvarez, C.A.; Herrera, E.B.; Thoman, M.L.; Chatfield, D.A.; Osborn, K.G.; Feuer, R.; George, U.Z.; Phillips, J.A. Acetylcholine Regulates Pulmonary Pathology During Viral Infection and Recovery. Immunotargets Ther 2020, 9, 333–350. [CrossRef]
- Islas-Weinstein, L.; Marquina-Castillo, B.; Mata-Espinosa, D.; Paredes-González, I.S.; Chávez, J.; Balboa, L.; Marín Franco, J.L.; Guerrero-Romero, D.; Barrios-Payan, J.A.; Hernandez-Pando, R. The Cholinergic System Contributes to the Immunopathological Progression of Experimental Pulmonary Tuberculosis. Front Immunol 2020, 11, 581911. [CrossRef]
- Ramirez, V.T.; Godinez, D.R.; Brust-Mascher, I.; Nonnecke, E.B.; Castillo, P.A.; Gardner, M.B.; Tu, D.; Sladek, J.A.; Miller, E.N.; Lebrilla, C.B.; et al. T-Cell Derived Acetylcholine Aids Host Defenses during Enteric Bacterial Infection with Citrobacter Rodentium. PLOS Pathogens 2019, 15, e1007719. [CrossRef]
- Matilla, M.A.; Velando, F.; Tajuelo, A.; Martín-Mora, D.; Xu, W.; Sourjik, V.; Gavira, J.A.; Krell, T. Chemotaxis of the Human Pathogen Pseudomonas Aeruginosa to the Neurotransmitter Acetylcholine. mBio 2022, 13, e0345821. [CrossRef]
Figure 1.
M1 and M2 macrophages exhibit distinct inflammatory phenotypes by expressing and secreting pro-inflammatory and anti-inflammatory cytokines, respectively. M1 macrophages, classically activated, play a vital role in pro-inflammatory responses, expressing receptors like MHC-II, iNOS, TLR-2, TLR-4, CD86, and CD80. They secrete pro-inflammatory cytokines (TNF-α, IL-1α, IL-1β, IL-6, IL-12, IL-23, CXCL9 (MIG), CXCL10, (IP-10), CXCL11 (I-TAC), CXCL16, and CCL5 (RANTES) produce ROS and NO, and promote Th1 responses. In contrast, M2 macrophages, alternatively activated, exhibit anti-inflammatory and tissue repair functions, expressing receptors such as CD163, CD206, CD209, FIZZ1, and Ym1/2. They secrete anti-inflammatory cytokines (IL-10, TGF-β, CCL1 (I-309), CCL17 (TARC), CCL18 (MIP-4), CCL22 (MDC), CCL24 (Eotaxin-2), CXCL13 (BCA-1), and VEGF), enhance phagocytosis, and support Th2 responses. The balance between M1 and M2 states is crucial for immune homeostasis and effective immune responses.
Figure 1.
M1 and M2 macrophages exhibit distinct inflammatory phenotypes by expressing and secreting pro-inflammatory and anti-inflammatory cytokines, respectively. M1 macrophages, classically activated, play a vital role in pro-inflammatory responses, expressing receptors like MHC-II, iNOS, TLR-2, TLR-4, CD86, and CD80. They secrete pro-inflammatory cytokines (TNF-α, IL-1α, IL-1β, IL-6, IL-12, IL-23, CXCL9 (MIG), CXCL10, (IP-10), CXCL11 (I-TAC), CXCL16, and CCL5 (RANTES) produce ROS and NO, and promote Th1 responses. In contrast, M2 macrophages, alternatively activated, exhibit anti-inflammatory and tissue repair functions, expressing receptors such as CD163, CD206, CD209, FIZZ1, and Ym1/2. They secrete anti-inflammatory cytokines (IL-10, TGF-β, CCL1 (I-309), CCL17 (TARC), CCL18 (MIP-4), CCL22 (MDC), CCL24 (Eotaxin-2), CXCL13 (BCA-1), and VEGF), enhance phagocytosis, and support Th2 responses. The balance between M1 and M2 states is crucial for immune homeostasis and effective immune responses.
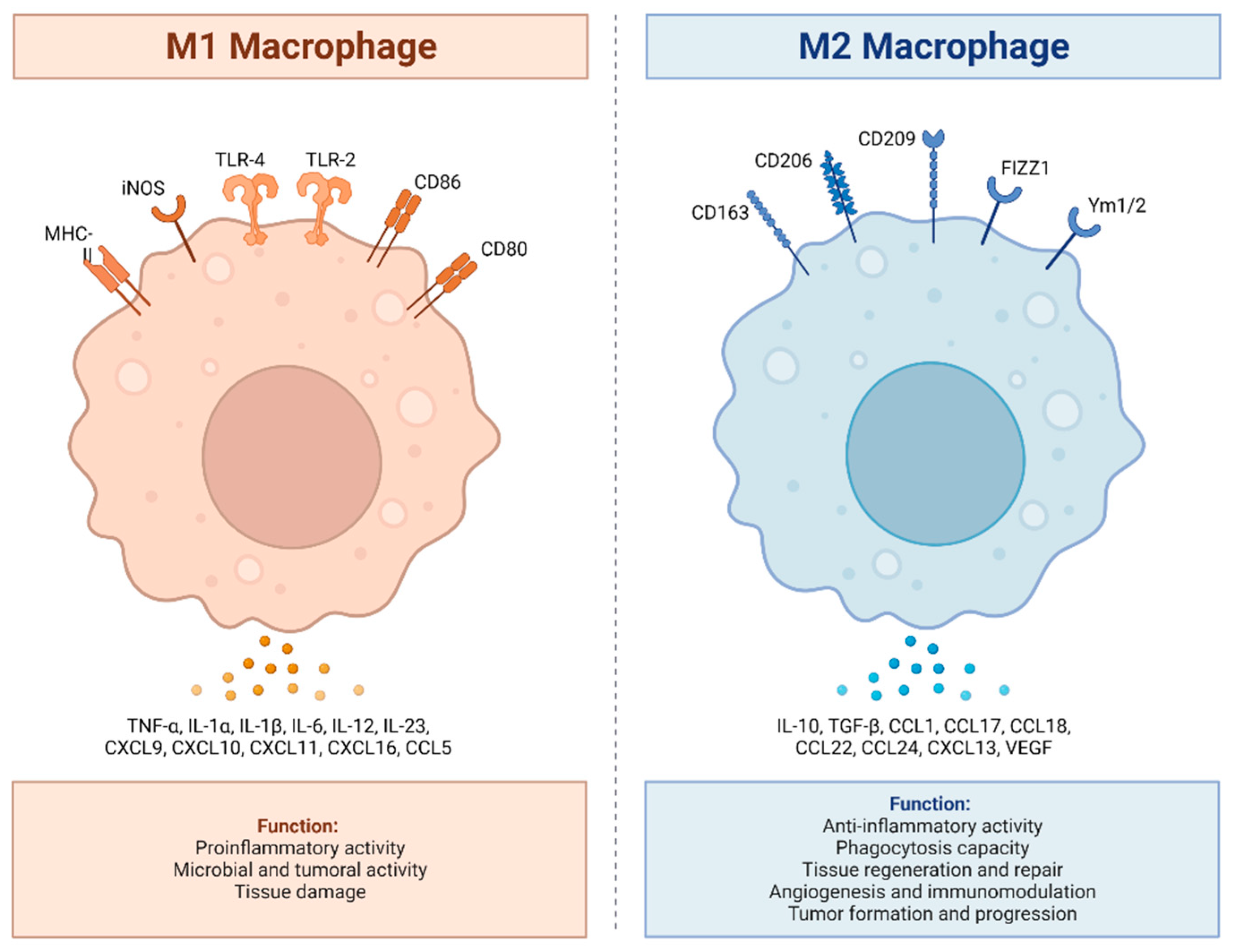
Figure 2.
M2 macrophages are divided into M2a, M2b, M2c, and M2d subtypes. These macrophages differ in their cell surface markers, secreted cytokines, and biological functions. Once stimulated, each subtype exhibits unique functions and expresses characteristic receptors that influence their distinct roles in immune regulation and tissue homeostasis. M2a macrophages, alternatively activated by IL-4 and IL-13, are involved in tissue repair, high endocytic activity, and anti-inflammatory responses. They express CD206, IL-1R, Arg-1, FIZZ1, and Ym1/2 which contribute to their polarization. M2b macrophages, activated by immune complexes, IL-1β, and Toll-like receptor (TLR) signaling, exhibit regulatory functions in immune responses and Th2 differentiation. Their characteristic receptors include IL-10R, IL-12R, CD86, and IL-6R. M2c macrophages, activated by IL-10, TGF-β, and glucocorticoids, are known for their anti-inflammatory and immune-regulatory functions such as phagocytosis of dying cells. They express TLR-8, TLR-1, Arg-1, CD163, and CD206. M2d macrophages, activated by TLR ligands, are involved in immune modulation and express IL-10R and IL-12R.
Figure 2.
M2 macrophages are divided into M2a, M2b, M2c, and M2d subtypes. These macrophages differ in their cell surface markers, secreted cytokines, and biological functions. Once stimulated, each subtype exhibits unique functions and expresses characteristic receptors that influence their distinct roles in immune regulation and tissue homeostasis. M2a macrophages, alternatively activated by IL-4 and IL-13, are involved in tissue repair, high endocytic activity, and anti-inflammatory responses. They express CD206, IL-1R, Arg-1, FIZZ1, and Ym1/2 which contribute to their polarization. M2b macrophages, activated by immune complexes, IL-1β, and Toll-like receptor (TLR) signaling, exhibit regulatory functions in immune responses and Th2 differentiation. Their characteristic receptors include IL-10R, IL-12R, CD86, and IL-6R. M2c macrophages, activated by IL-10, TGF-β, and glucocorticoids, are known for their anti-inflammatory and immune-regulatory functions such as phagocytosis of dying cells. They express TLR-8, TLR-1, Arg-1, CD163, and CD206. M2d macrophages, activated by TLR ligands, are involved in immune modulation and express IL-10R and IL-12R.
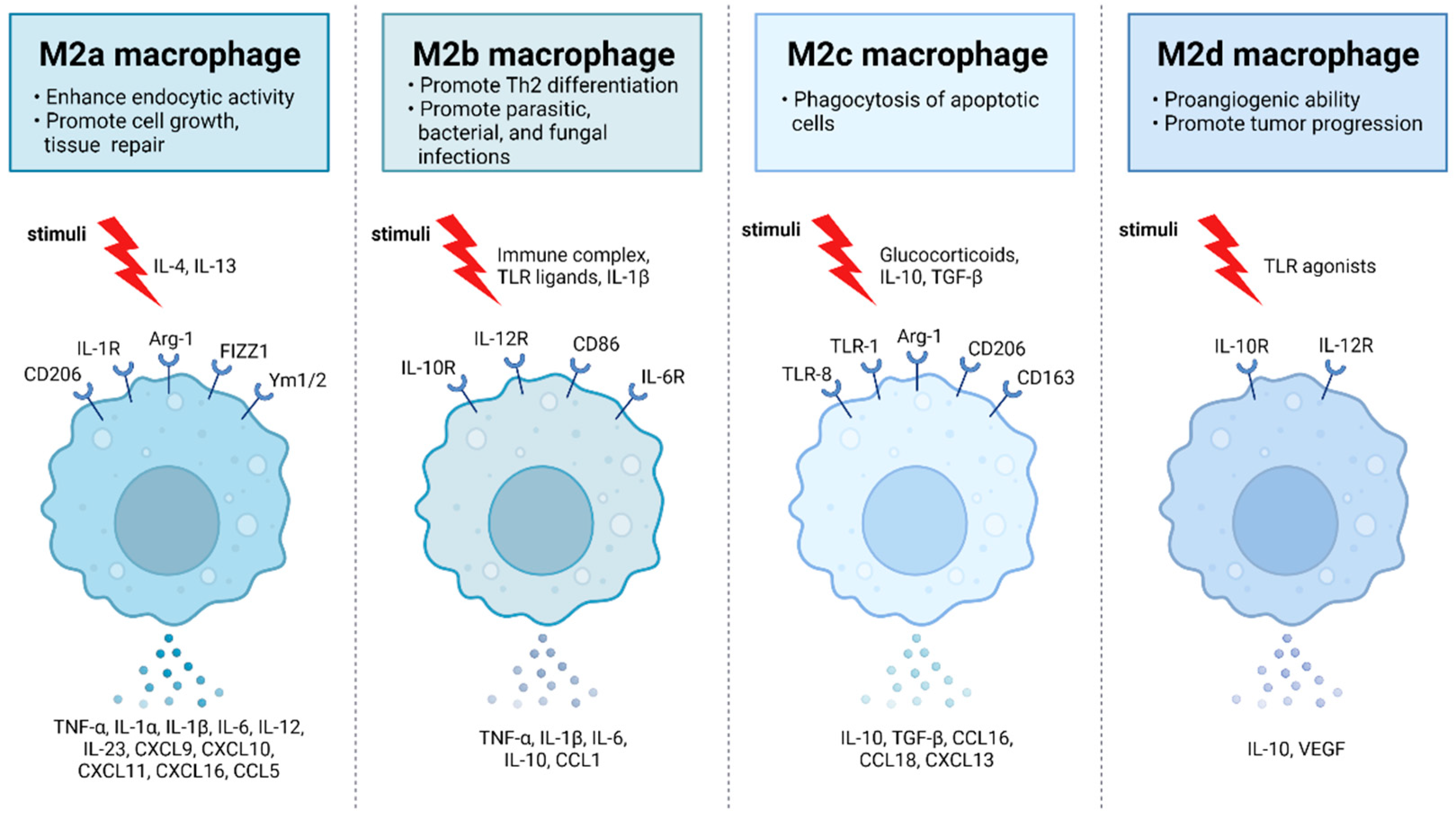
Figure 3.
M0 macrophages are the precursors that give rise to specialized subtypes found in atherosclerotic lesions. Monocytes are differentiated to M0 macrophages via monocyte colony stimulating factor (MCSF). Depending on the tissue microenvironment, the M0 macrophage is polarized to a particular subtype. The primary subtypes are pro-inflammatory M1 and anti-inflammatory M2 macrophages with immunoregulatory properties. In addition, multiple other subtypes have been identified including M(Hb), Mhem, Mox, and M4. All of these subtypes produce various chemokines and cytokines and have different functions within the atherosclerotic plaque.
Figure 3.
M0 macrophages are the precursors that give rise to specialized subtypes found in atherosclerotic lesions. Monocytes are differentiated to M0 macrophages via monocyte colony stimulating factor (MCSF). Depending on the tissue microenvironment, the M0 macrophage is polarized to a particular subtype. The primary subtypes are pro-inflammatory M1 and anti-inflammatory M2 macrophages with immunoregulatory properties. In addition, multiple other subtypes have been identified including M(Hb), Mhem, Mox, and M4. All of these subtypes produce various chemokines and cytokines and have different functions within the atherosclerotic plaque.
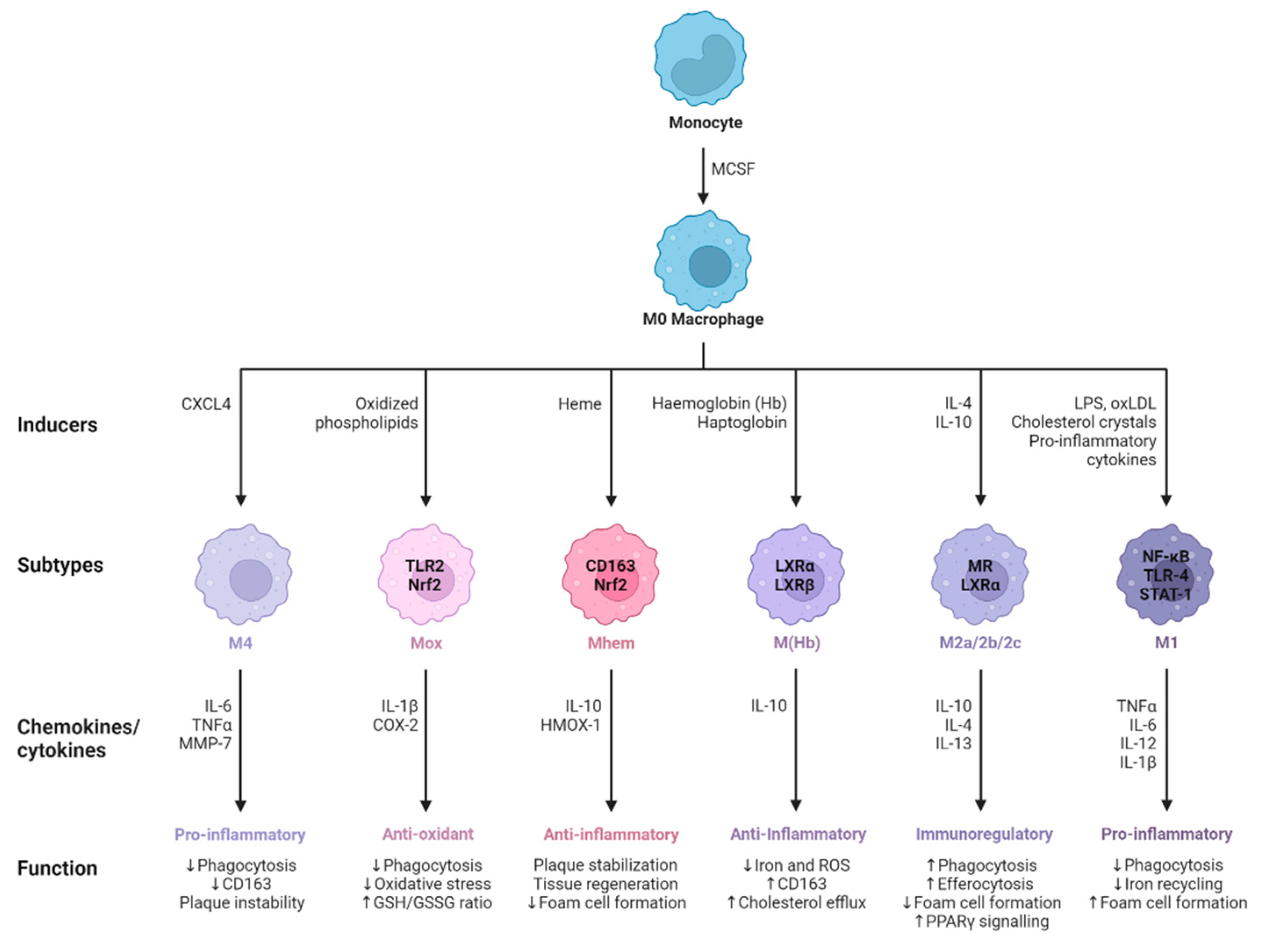
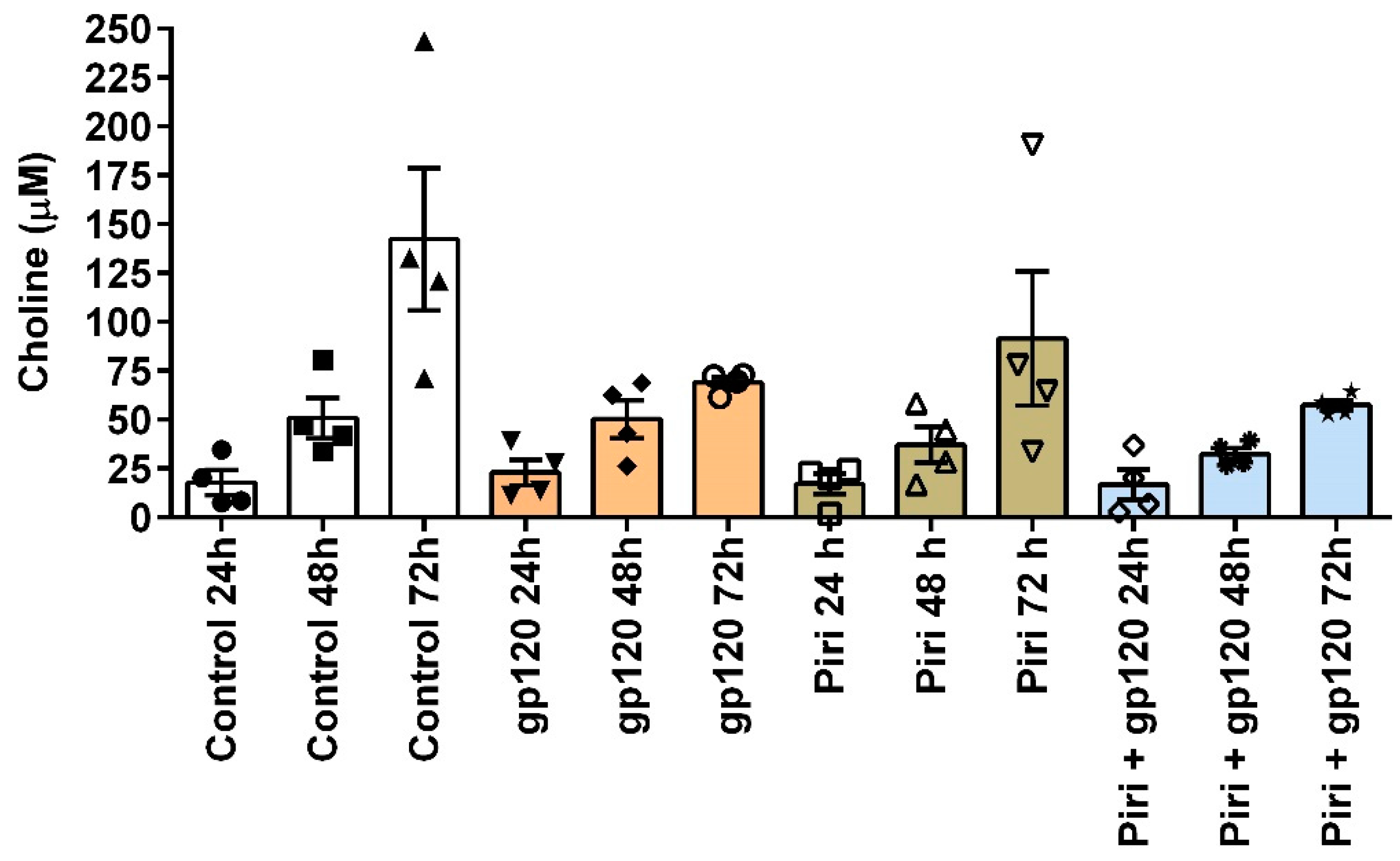
Figure 4.
Choline concentrations in primary cultures of MDMs. Choline levels were determined using Biovision's Choline/Acetylcholine Quantification Colorimetric/Fluorometric Kit (cat# K615-100) following the manufacturer's instructions. The graph was constructed by subtracting the choline values from the supplemented culture medium (initially 113.70 uM) to show the choline concentration produced exclusively by the MDMs. The data were derived from four independent experiments, and error bars represent the standard error of the mean (SEM). Over time, the choline concentration gradually increased for all tested conditions, underscoring the significance of considering choline accumulation when aiming to activate the cholinergic anti-inflammatory response through the α7-nAChR. The control condition consisted of MDMs without any treatment, incubated for 24, 48, and 72 hours. A One-Way ANOVA followed by a Sidak's multiple comparisons test revealed no significant difference when comparing all conditions against controls at 24, 48, and 72 hours. The cell culture media used was RPMI-1640 supplemented with 20% inactivated fetal bovine serum, 10% inactivated human serum, 2 μg/ml of macrophage colony-stimulating factor, and 1% Penicillin Streptomycin. The abbreviation "Piri" refers to pyridostigmine; gp120 refers to gp120IIB.
Figure 4.
Choline concentrations in primary cultures of MDMs. Choline levels were determined using Biovision's Choline/Acetylcholine Quantification Colorimetric/Fluorometric Kit (cat# K615-100) following the manufacturer's instructions. The graph was constructed by subtracting the choline values from the supplemented culture medium (initially 113.70 uM) to show the choline concentration produced exclusively by the MDMs. The data were derived from four independent experiments, and error bars represent the standard error of the mean (SEM). Over time, the choline concentration gradually increased for all tested conditions, underscoring the significance of considering choline accumulation when aiming to activate the cholinergic anti-inflammatory response through the α7-nAChR. The control condition consisted of MDMs without any treatment, incubated for 24, 48, and 72 hours. A One-Way ANOVA followed by a Sidak's multiple comparisons test revealed no significant difference when comparing all conditions against controls at 24, 48, and 72 hours. The cell culture media used was RPMI-1640 supplemented with 20% inactivated fetal bovine serum, 10% inactivated human serum, 2 μg/ml of macrophage colony-stimulating factor, and 1% Penicillin Streptomycin. The abbreviation "Piri" refers to pyridostigmine; gp120 refers to gp120IIB.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



