
Submitted:
23 September 2023
Posted:
25 September 2023
You are already at the latest version
Abstract
Dear Ladies and Gentlemen,
I thank you for suggesting to submit the manuscript with the title
“Insulin resistance develops due to an imbalance in the synthesis of cyclic AMP and the natural cyclic AMP antagonist prostaglandylinositol cyclic phosphate (cyclic PIP)”
to the Editorial Board of the MDPI journal Stresses, and I ask for consideration to publish it in the journal Stresses. It was previously submitted to the MDPI journal Int. J. Mol. Sci. (IJMS-195 6513), but it was found that 1) it would not fit within the scope of this special edition and 2) it would primarily deal with cyclic PIP and would have not enough literature citation on the topic insulin resistance.
The present version of the manuscript is improved, but the content is the same. The goal of this review is not to write again a summary on nearly all reports on insulin resistance as this was done by Dr. Petersen and Dr. Shulman, whose review has 961 literature citations. The present manuscript compiles what is known on cyclic PIP in relation to insulin resistance and the reason of writing this manuscript was that the discovered facts on cyclic PIP synthesis allow to suggest how insulin resistance most likely starts. Hitherto, reports on insulin resistance did not come close to this question. I am convinced that the manuscript makes an essential contribution to the question what causes the development of insulin resistance, and I have to say that stress overload is one of the reasons.
Unfortunately, I have to mention a topic, about which I do not like to talk. Years ago, research groups postulated with rigor that the mediator of insulin action would be a peptide and then they suggested various, extracellularly synthesized glycans as mediators. They caused confusions and a high degree of disbelief in the matter of mediators of insulin action. In my review on cyclic PIP in IUBMB Life from 2020, I wrote in the introduction that cyclic PIP should not be confused with these mediators. But I cannot write this in the introduction of every report. I am keeping fingers crossed so that the editor in charge will find fair and openminded reviewers.
Last not least I confirm that neither the manuscript nor any part of its content is currently under consideration or published in another journal.
Kind regards,
Henrich Wasner, PhD,
BioReg Biopharm, TiLab;
University of Illinois at Chicago;
2242 West Harrison St., Ste. 201,
Chicago, Il 60612; USA
Keywords:
cyclic AMP
; cyclic PIP
; insulin resistance
; prostaglandylinositol cyclic phosphate
; protein phosphorylation
; protein kinase A
; protein ser/thr phosphatase
; type 2 diabetes
1. Introduction
Type 2 diabetes and hypertension are acute health problems of today’s world. The percentage of people with type 2 diabetes is continuously increasing and worldwide there are at least 422 million people with diabetes and 1.28 billion adults with hypertension according to the WHO. Many scientists are working to find the primary causes which initiate the development of insulin resistance, a starting condition of type 2 diabetes. Using various inbred mouse strains, Nelson et al. determined the effects of diet on the development of insulin resistance primarily in muscle, liver and white adipose tissue [1] and Mejhert and Ryden commented on this report [2]. Petersen and Shulman published a comprehensive and detailed review on the “mechanism of insulin action and insulin resistance” and came to the conclusion that the “sheer complexity of biological systems means that any effort to understand insulin resistance with a unified, succinct and straightforward model may be a fool’s errand” [3]. Galicia-Garcia et al. [4] proposed that type 2 diabetes is caused by a combination of two primary factors, namely a defective insulin secretion of the pancreatic β-cells and the inability of insulin sensitive tissues to respond appropriately to insulin, and they concluded that there is still a long way to go to fully understand insulin resistance.
The mechanism of insulin action, which is a prerequisite to determine defects in the action of insulin, appears to be nearly completely solved: Insulin binds to its receptor, whose intrinsic tyrosine kinase is in this way activated; this kinase auto-phosphorylates itself and then phosphorylates primarily the insulin receptor substrates IRS1-4 and several Shc proteins [5,6]. The IRS proteins are suggested to regulate metabolism, whereas Shc proteins appear to be involved in the regulation of proliferation by insulin. The IRS proteins are considered to be docking proteins to which various enzymes bind via Src-homology-2 (SH-2) domains and send signals by phosphorylation cascades further into a cell. Additionally, the IRS proteins are inactivated by serine/threonine phosphorylation by several different protein ser/thr kinases [5].
Earl Sutherland articulated many years ago that a decrease in the strength of the insulin signal will increase the strength of the counterregulatory signal leading to dominance of the regulations triggered by cyclic AMP [7]. This view has not been adequately pursued. A characteristic of insulin resistance is that it progresses slowly and gradually for reasons not fully understood at the present time. Inhibition of single proteins/enzymes such as the IRS proteins cannot explain this slow progression, though they play a crucial role in the signal-transduction. Current thinking is that protein phosphorylation cascades are of primary importance [5,6], but fails to consider that insulin antagonizes glucagon action. Insulin rapidly reverses cyclic AMP-triggered protein ser/thr phosphorylation. The key player in this scenario is the natural cyclic AMP antagonist, prostaglandyl-inositol cyclic phosphate (cyclic PIP) [8,9]. The synthesis of cyclic PIP is stimulated by insulin and also noradrenaline. In type 2 diabetic and also hypertensive rodents a reduced cyclic PIP synthesis was found [8]. The effect of decreasing cyclic PIP synthesis and the consequently increasing cyclic AMP synthesis on the development of insulin resistance is the topic of this report.
2. Cyclic AMP
After its discovery by Earl Sutherland, cyclic AMP was for many years viewed as the unique intracellular regulator, which activates the cyclic AMP-dependent protein kinase (PKA). By protein serine/threonine phosphorylation catabolic enzymes are predominantly activated and anabolic enzymes predominantly inactivated. Further ways of action of cyclic AMP are a) its activation of guanine exchange factors, b) its binding to cyclic AMP binding proteins, and c) its regulation of cyclic nucleotide gated ion channels [10]. Cyclic AMP synthesis is stimulated for instance by glucagon and adrenaline via its β-receptors. The activation of adenylate cyclase is executed by the Gαs subunit of the stimulatory, heterotrimeric Gs protein [11]. The reversal of this activation is controlled by the GTPase activity of the Gαs protein, which converts the bound GTP to GDP inactivating adenylate cyclase. Adenylate cyclase is inhibited dose-dependently up to 100% by cyclic PIP [8]. In contrast, the Gi protein inhibits adenylate cyclase by approximately 30% at maximal hormone concentration [12]. This effect is declared to be an adrenergic α2-receptor action (see below).
3. The Natural Cyclic AMP Antagonist Cyclic PIP
The search for an antagonist to cyclic AMP started in the laboratory of the late Earl Sutherland. Cyclic PIP was primarily isolated from rat livers, which were extra-corporally perfused with buffer and stimulated with noradrenaline or insulin, homogenized within 1 to 3 minutes then put under denaturing conditions to minimize the enzymatic degradation of cyclic PIP. From 2 kg of rat livers approximately 60 g of water-soluble, low molecular compounds were obtained, which also contained cyclic PIP. A rough calculation indicated that 500.000 to 1.000.000 molecules of this extract contain 1 molecule of cyclic PIP. But more challenging was the finding that cyclic PIP is one of the most labile molecules of this extract. It decays by 80% within 30 minutes held at an ionic strength comparable to a 0.5 molar sodium chloride solution, approximately a 3-fold concentrated physiological saline solution. The labilities of cyclic PIP are connected to the tension of the 5-ring phosphodiester, which is the most labile bond of cyclic PIP, as well as the allyl-ether bond combining two secondary alcohols and the beta-keto-hydroxy structure of the C5-ring of prostaglandin E (PGE). Chemically it is O-(prostaglandyl E)-(15-4′) (myo-inositol 1′:2′-cyclic phosphate) [8]. It is biosynthesized from PGE and ‘activated inositol phosphate’ by cyclic PIP synthase [8], which is active in a tyrosine phosphorylated form [13]. The synthesis of cyclic PIP is also stimulated by adrenergic α1- and α 2-receptors [14]. This result contradicts the present view on the action of adrenergic α1- and α2-receptor action, as discussed in [14,15]. The adrenergic alpha-receptors are transmembrane, G protein-coupled receptors, thus cyclic PIP synthase should be activated also by a G protein [13]. It is not known whether there are two different variants of cyclic PIP synthase, but it is known that the biosynthesis of the substrates for cyclic PIP synthesis involves phospholipase A2 and phospholipase C, both of which are activated by both these modes of activation as summarized in [15]. The synthesis of cyclic AMP from ATP is a one-step reaction and ATP is generally always present in a high enough amount in all cells to warrant maximal synthesis of cyclic AMP. In contrast, the biosynthesis of cyclic PIP needs at least five reaction steps (Figure 1) and both substrates are obtained from membrane-bound lipids [15].
The primary action of cyclic AMP is to activate cyclic AMP-dependent protein kinase (PKA), whereas cyclic PIP inhibits PKA. The PKA is activated by increasing concentrations of cyclic AMP (10-8 to 10-6 molar) at least 10-fold. Addition of increasing concentrations of cyclic PIP leads to increasing inhibition of basal and cyclic AMP-activated PKA. For an equal percentage of inhibition of basal and cyclic AMP-activated PKA approximately 4-times more cyclic PIP is needed in case of the cyclic AMP-activated kinase [14,16]. The two regulators, cyclic AMP and cyclic PIP, perfectly regulate the activity of PKA. It was to assume that protein ser/thr phosphatase, the counter-regulatory enzyme to PKA, would also be regulated by these two compounds. The phosphatase is a rather labile enzyme. Therefore, scientists decided to isolate and characterize the catalytic subunits of this class of enzymes [17]. For this reason, the holoenzyme of protein ser/thr phosphatase was partly purified. The obtained enzyme has a slightly higher molecular weight than PKA and it is 7-fold activated by cyclic PIP and completely inhibited by cyclic AMP in the physiological action range [8]. As was the case with cyclic AMP [10], further modes of action of cyclic PIP are likely to be found in the coming years as chemical synthesis of cyclic PIP allows additional studies.
4. The Interplay of Cyclic AMP and Cyclic PIP
4.1. At the Level of Their Biosynthesis: Cyclic AMP Inhibits Cyclic PIP Synthase and Cyclic PIP Inhibits Adenylate Cyclase
Cyclic PIP synthase is activated by tyrosine phosphorylation [9,13], which correlates with the signal-transduction and the tyrosine kinase activity of the insulin receptor [5,6]. Apart from the activation and inhibition by tyrosine phosphorylation and dephosphorylation, respectively, cyclic PIP synthase is inhibited by cyclic AMP in the presence of ATP by a protein ser/thr kinase [18]. This inhibition may ensure that no cyclic PIP is synthesized at a time when cyclic AMP is required to respond to appropriate signals, as it would be counterproductive if both cyclic AMP and cyclic PIP were synthesized at the same time [9]. With respect to selectivity, it is expected that this phosphorylation is carried out by a specific kinase and not by a half dozen of enzymes as discussed for the IRS proteins [5,6]. The protein ser/thr phosphatase, which reverses the protein ser/thr phosphorylation of cyclic PIP synthase has yet to be found.
Cyclic PIP was found by its action to inhibit adenylate cyclase [8] indicating that cyclic PIP antagonizes the action of cyclic AMP comparably to the inhibition of cyclic PIP synthase by cyclic AMP. This inhibition is possibly the result of a phosphorylation reaction [19]. However, this kinase has not been characterized further and it is not known if it phosphorylates tyrosine- or serine/threonine-residues of adenylate cyclase or other regulatory structures involved in the activation of adenylate cyclase. The group of Pradipta Ghosh reported recently that receptor tyrosine kinases are activated by growth factors as EGF, PDGF and insulin. These kinases phosphorylate tyrosine-residues of the Gαi protein, which then efficiently inhibits adenylate cyclase. This inhibition was demonstrated in intact cells, but difficult to show in ‘in vitro’ assays. They explained that in the ‘in vitro’ assay a part of this complex assay mixture may be missing or not be present in the needed amount [20]. This missing component could be cyclic PIP since under the experimental conditions used the synthesis of cyclic PIP is stimulated very well in intact cells but not in disrupted cells [21]. One may question if these three different inhibitory effects on adenylate cyclase, namely the partial inhibition by Gi protein [12], the complete inhibition by cyclic PIP [8] and also by the tyrosine-phosphorylated Gαi protein [20], could be parts of one complex reaction sequence leading to inhibition of adenylate cyclase, since it would be bewildering when adenylate cyclase would be inhibited in various ways, which are triggered by the same hormones. That means, insulin stimulates the synthesis of cyclic PIP and also the tyrosine-phosphorylation of Gαi protein, and noradrenaline stimulates the synthesis of cyclic PIP and triggers the inhibition via the Gi protein, which was determined in cell homogenates by Sabol and Nirenberg. They most likely could determine only a partial inhibition of adenylate cyclase [12], since cyclic PIP is not synthesized in cell homogenates [21].
The point of this chapter is that cyclic AMP inhibits cyclic PIP synthase [18] and cyclic PIP inhibits adenylate cyclase [8]. The following two experiments visualize the effect of cyclic PIP on the hormone-stimulated adenylate cyclase of hepatocytes of untreated and diabetic rats. A) Glucagon stimulates cyclic AMP synthesis about 30-fold in isolated rat hepatocytes, whereas adrenaline triggers only a 2- to 3-fold increase. However, when hepatocytes were first stimulated with glucagon and 40 min later with adrenaline, then adrenaline stimulates cyclic AMP synthesis 6 to 9-fold [8,22]. The explanation is that on the first stimulation with adrenaline cyclic PIP synthesis is also increased, which inhibits adenylate cyclase, limiting the increase of cyclic AMP. Glucagon stimulates a much greater synthesis of cyclic AMP, which will inhibit cyclic PIP synthase. Hence, after a first stimulation with glucagon, on a second stimulation with adrenaline the synthesis of cyclic PIP is decreased, leading to an increased cyclic AMP synthesis. For a perfect intracellular control of the activity of adenylate cyclase and also cyclic PIP synthase, one would expect that both enzymes are not only inhibited by their antagonistic regulator, but that there must be mechanisms by which either cyclic PIP synthase or adenylate cyclase is specifically reactivated. B) The time course of cyclic AMP synthesis was determined in rat hepatocytes of untreated and diabetic rats, obtained by low dose streptozotocin-treatment. Both cell suspensions were stimulated with adrenaline at 0 and at 30 minutes. Adrenaline stimulated cyclic AMP and also cyclic PIP synthesis. The basal cyclic AMP level in the hepatocytes of the diabetic rats was approximately 3-fold higher than in cells of non-diabetic control rats. Additionally, adrenaline increased cyclic AMP synthesis 1.6-fold in the hepatocytes from diabetic rats and 1-fold in the controls. On the second stimulation with adrenaline at 30 minutes, in the controls the synthesis of cyclic AMP was decreased by approximately 90%, because of the stimulation of cyclic PIP synthesis by adrenaline at 0 minutes. However, in the hepatocytes of the diabetic rats, the synthesis of cyclic AMP, following a second stimulation with adrenaline was nearly as great as it was in the first stimulation. The reduced cyclic PIP synthesis, which was observed in livers of diabetic rodents is the obvious explanation [8,18]. This result is in accordance with Sutherland’s observation that cyclic AMP synthesis increases in cells with a reduced insulin response [7]. This finding is not unique to rats treated with streptozotocin. Cyclic PIP synthase activity in livers of Ksj db/db mice is 36% lower than in Ksj control mice, and cyclic PIP synthase activity in livers of spontaneously hypertensive rats (SHR) at three month of age is already decreased by 27%, when compared with the activity of Wistar-Kyoto control rats [18].
In summary, decreasing stimulation of cyclic PIP synthesis, leading to increasing cyclic AMP synthesis, appears to play a central role in the development of type 2 diabetes. Essentially, elevated cyclic AMP levels will further inhibit the stimulation of cyclic PIP synthesis and in this way continuously increase cyclic AMP synthesis and action with time. The resulting downward spiral of the stimulation of cyclic PIP synthesis can explain the nearly absent hormonal stimulation of cyclic PIP synthesis (Table 1), which will lead to a nearly unopposed dominance of cyclic AMP. In other words, the mutual inhibition of the synthesis of these two intracellular regulators, cyclic AMP and cyclic PIP is certainly necessary because it enables that the regulator, whose synthesis is stimulated by a hormone at a time, can do his task undiminished. In the case, however, that the synthesis of one of these two regulators becomes weaker, in the case of insulin resistance it is cyclic PIP, then this positive effect of mutual inhibition enforces the imbalance in the synthesis of these two regulators and consequently in the regulation of the metabolism.
4.2. At the Level of the Regulation of Metabolism: Increasing Cyclic AMP and Decreasing Cyclic PIP Synthesis Leads to Dominance of Catabolism over Anabolism
The activity of PKA and of protein ser/thr phosphatase is regulated by cyclic AMP and cyclic PIP [8,9], creating an equilibrium between de-phospho- and phospho-form of interconvertible enzymes. For instance, insulin stimulates the synthesis of cyclic PIP, which shuts off cyclic AMP synthesis. Additionally, it activates protein phosphatase and inhibits PKA [8,9] and shifts the equilibrium between de-phospho- and phospho-form of interconvertible enzymes to the left side, the dephosphorylated form (Figure 2). However, a decreasing stimulation of cyclic PIP synthesis will allow an increasing dominance of cyclic AMP [18]. This situation, in turn, will further and further decrease the stimulation of cyclic PIP synthesis. The result is that the equilibrium between de-phospho- and phospho-form of interconvertible enzymes will be more and more shifted to the phospho-form and the metabolism will be more and more stuck in the catabolic state.
5. Illnesses Connected with Decreased Synthesis of Cyclic PIP
The stimulation of cyclic PIP synthesis decreases in low dose streptozotocin diabetic rats [18,23] and additionally, streptozotocin-induced type 2 diabetes in rats can be improved by feeding arachidonic acid [24], the substrate for prostaglandin and cyclic PIP synthesis. Furthermore, inhibition of cyclic PIP synthesis by non-steroidal anti-inflammatory drugs (NSAIDs) leads to a metabolic state of the treated rats, which is comparable to type 2 diabetes at the onset of the illness [21]. Glucose tolerance tests showed that in these treated rats the blood insulin levels increased at least threefold higher as compared to the untreated control rats. Cyclic PIP inhibits the release of insulin in accordance with insulin’s action. A reduced synthesis of cyclic PIP leads to a decreasing inhibition of insulin secretion from the pancreatic β-cells and thus to increased blood insulin levels [21]. Additionally, these animals had 50% higher blood glucose levels, which declined slower in comparison with the values from untreated control rats. Accordingly, liver and muscle of these treated rats had highly reduced glycogen contents. These results correlate with the stimulation of glucose uptake by cyclic PIP in adipocytes [21].
Why and how does decreased stimulation of cyclic PIP synthesis cause insulin resistance? In the introduction, some present views on insulin resistance were mentioned. These reports describe metabolic conditions, which show the extent of derailment of metabolism in the course of the development of type 2 diabetes. However, the events that initiated the derailment have not been identified and are mostly not discussed. In particular, the action of cyclic AMP and the existence and action of cyclic PIP are rarely considered. But it has been recognized that many different pathways are derailed in the case of the development of insulin resistance [1,3,4,25]. However, since insulin triggers a multitude of regulations in all the different organs of a body, one could realize that a single, essential derailment of insulin’s signal transduction, such as the gradual loss of stimulation of cyclic PIP synthase will result in a gradual decrease of all these many regulations stimulated by insulin. The simplified model-calculation of Table 2 may visualize how the regulation of the metabolism can derail as result of a decreasing stimulation of cyclic PIP synthesis. Interconvertible enzymes are at the start 100% in the de-phospho-form and cyclic AMP-activated PKA converts 20% of these enzymes to the phospho-form. On a successive insulin stimulation, because of the lower activation of cyclic PIP synthase, the protein phosphatase is less active and only 19% of the phosphorylated enzymes are dephosphorylated, and 1% remain in the phospho-form. On a second ‘phosphorylation cycle’ the kinase is 1% more active and the protein phosphatase accordingly 1% less active because of the mutual inhibition of the two regulators and the increasing dominance of cyclic AMP (as discussed above). The result is that 4% of the enzymes remain after this second ‘phosphorylation cycle’ in the phospho-form and after the 6th ‘phosphorylation cycle’ 36% of the enzymes remain in the phospho-form. This calculation illustrates that the regulation of the metabolism will gradually and consistently get more and more out of balance and continuously increasing amounts of interconvertible enzymes will stay in the phosphorylated from. Thus, catabolic regulations will dominate more and more and anabolic regulations will decrease comparably. Certainly, in reality this will not be a straight forward, linear development, because parameters such as time and concentrations of key stimulators will vary. Finally, the magnitude of the activity-changes will be much smaller than in this brief model of Table 2, where an arbitrary 1% change was applied, and the progress of the derailment will accordingly be slower. Regardless of these details, this development will progress comparably to a temperature curve, but slowly and continuously over prolonged times, until metabolism is increasingly stuck in the catabolic state.
As mentioned in the introduction, Galicia-Garcia et al. recently concluded that type 2 diabetes is caused by a defective insulin secretion and an inability of insulin responsive tissues to respond to insulin [4]. It could have been recognized that these defects are related to the decreasing action of cyclic PIP. Firstly, cyclic PIP inhibits the release of insulin from pancreatic β-cells [8] and in case of a vanishing stimulation of cyclic PIP synthesis the shut off of insulin’s secretion seizes and the resulting continuous excretion of insulin will gradually lead to an exhaustion of the pancreatic β-cells [26]. Secondly, all organs/cells of a body are responsive to insulin and not just skeletal muscles, liver and white adipose cells, and, so far as cyclic PIP is the intracellular executor of many actions of insulin [8], these functions will vanish in case of a decreasing cyclic PIP synthesis.
What are reasons which cause the decreasing stimulation of cyclic PIP synthesis? 1) One possibility is that needed substrates can be limiting. This problem should play a minor role in the synthesis of ‘activated inositol phosphate’, since generally there should be no deficit of myo-inositol intake by food. However, an increasing number of reports show that polycystic ovarian syndrome, gestational diabetes mellitus and metabolic syndrome can be improved by increased intake of inositol [27]. Since myo-inositol is a structural component of cyclic PIP, one may ask if this could correlate with an improved synthesis of cyclic PIP, however, presently it is not known which defect in the path from myo-inositol via phosphatidylinositol to activated inositol phosphate can be improved by increased intake of myo-inositol. 2) Substrate deficiency will more likely play a role in case of the synthesis of prostaglandins. The supply of lipids containing polyunsaturated fatty acids as omega-6 fatty acids can be limiting, especially since humans cannot synthesize these acids themselves and are dependent on their sources of food. Along these lines, pain relievers of the NSAID-family inhibit the biosynthesis of prostaglandins which impair the synthesis of cyclic PIP [21], and more recently was reported that streptozotocin-induced type 2 diabetes can be improved by support with arachidonic acid [24]. 3) Too many humans have high stress levels that result from problems of daily life such as too long exposures to too much noise and to challenges at work [28], causing elevated cyclic AMP levels [29]. 4) Another consideration is that regulatory mechanisms decline with aging. Though not finally proven, key hormones such as dehydroepiandrosterone, growth hormone, insulin-like growth factor-1 and testosterone appear to be involved in aging and in insulin action [30]. Presently, the mechanisms of action of these hormones are still under investigation. Pataky, Young and Nair recommend in their review on aging that caution be taken in treatment with these hormones, since too many negative side effects of these hormonal replacements could outweigh the positive effects. They suggest that, presently, it is better to maximally apply physical exercise and calorie restrictions, since this way of lifestyle has positive effects on aging and on insulin sensitivity and has no negative side effects, though the mechanisms of action of lifestyle-changes are still largely unknown. These hormones start to decrease at around the age of 30 by around 1 to 2% annually [30]. This could be a further factor affecting the slow development of type 2 diabetes over many years. Unfortunately, not enough is known about the biochemical mechanism of action of these hormones and how their decrease might lead to type 2 diabetes and hypertension and which one of these hormones has an effect on the stimulatory mechanism of cyclic PIP synthesis.
In summary, the development of an imbalance in the mutual inhibition of the synthesis of cyclic PIP and cyclic AMP, as outlined above (Figure 2), will lead to a derailment in the regulation of metabolism. There appear to be compensatory mechanisms, which may slow down this derailment of the metabolism. Matching with cyclic PIP action, insulin inhibits skeletal muscle PKA as shown, for instance in healthy Rhesus monkeys [31]. Diabetes, apart from the disrupted insulin signaling, moreover leads to a defective signaling of adrenergic β-receptors. In the focus stays the not adequately responsive PKA [32,33,34]. The decreasing cyclic PIP synthesis leads initially to increasing cyclic AMP levels, whose actions appear to be subsequently opposed by an unresponsive PKA in a, presently, not finally discovered way. If such a down-regulation can improve diabetic conditions or if an unresponsive PKA makes the treatment of diabetes more complicated has to be found out.
There are reports that type-2 diabetes is curable through lifestyle changes in diet and exercise. Certainly, there are environmental conditions which can be altered to improve the metabolic situation. However, other factors such as the aging process need to be better understood. Of particular note here are the four hormones discussed to have effects on aging and insulin action. Such studies would lead to substantial progress in understanding insulin resistance. This report discusses only the effects of decreasing stimulation of cyclic PIP synthesis with respect to the regulation of the equilibrium between the de-phospho- and phospho-form of interconvertible enzymes. When in the coming years further regulatory properties of cyclic PIP are better characterized, then further aspects of the development of insulin resistance may be found.
Funding
This work received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The author sincerely thanks Kathy Rundell for critical suggestions and for improving the language of the manuscript and Samantha Meister for skillful help to get the figures and tables in shape.
Conflicts of Interest
The author declares no conflict of interest.
References
- Nelson, M.E.; Madsen, S.; Cooke, K.C.; Fritzen, A.M.; Thorius, I.H.; Masson, S.W.C.; Carroll, L.; Weiss, F.C.; Seldin, M.M.; Potter, M.; Hocking, S.I.; Fazakerley, D.J.; Brandon, A.E.; Thillainadesan, S.; Senior, A.M.; Cooney, G.J.; Stoeckli, J.; James, D.E. (2022). Systems-level analysis of insulin action in mouse strains provides insight into tissue- and pathway-specific interaction that drive insulin resistance. Cell Metab. 34, 227-239. [CrossRef]
- Mejhert, N. and Ryden, M. (2022). Understanding the complexity of insulin resistance. Nat. Rev. Endocrinol. 18, 269-270. [CrossRef]
- Petersen, M.C. and Shulman, G.I. (2018). Mechanisms of insulin action and insulin resistance. Physiol. Rev. 88, 2133-2233. [CrossRef]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H. and Martin, C. (2020). Pathophysiology of type 2 diabetes mellitus. Int. J. Mol. Sci. 21, 6275. [CrossRef]
- White, M.F.; Kahn, C.R. (2021). Insulin action at a molecular level – 100 years of progress. Mol. Metab. 52, 101304. [CrossRef]
- Batista, T.M.; Haider, N.; Kahn, C.R. (2021). Defining the underlying defect in insulin action in type 2 diabetes. Diabetologia 64, 994-1006. [CrossRef]
- Robison, G.A.; Butcher, R.W.; Sutherland, E.W. Cyclic AMP; Academic Press: New York, NY, USA, 1971.
- Wasner, H.K. (2020). Prostaglandylinositol cyclic phosphate, the natural antagonist of cyclic AMP. IUBMB Life 72, 2282–2289. [CrossRef]
- Wasner, H.K. (2022). Metformin’s mechanism of action is stimulation of the biosynthesis of the natural cyclic AMP antagonist prostaglandylinositol cyclic phosphate (cyclic PIP). Int. J. Mol. Sci. 23, 2200. [CrossRef]
- Yarwood, S.J. (2020). Special issue on “new advances in cyclic AMP signaling” – an editorial overview. Cells 9, 2274. [CrossRef]
- von Keulen, S.C.; Rothlisberger, U. (2017). Exploring the inhibition mechanism of adenylyl cyclase type 5 by n-terminal myristoylated Gai1. PLoS. Comput. Biol. 13(9). [CrossRef]
- Sabol, S.L. and Nirenberg, M. (1979). Regulation of adenylate cyclase of neuroblastoma x glioma hybrid cells by a-adrenergic receptors. J. Biol. Chem. 254, 1913-1920.
- Wasner, H.K.; Gebel, M.; Hucken, S.; Schaefer, M.; Kincses, M. (2000). Two different mechanisms for activation of cyclic PIP synthase: By a G protein or by protein tyrosine phosphorylation. Biol. Chem. 382, 145–153. [CrossRef]
- Wasner, H.K.; Salge, U.; Gebel, M. (1993). The endogenous cyclic AMP antagonist, cyclic PIP: Its ubiquity, hormone-stimulated synthesis and identification as prostaglandylinositol cyclic phosphate. Acta Diabetol. 30, 220–232. [CrossRef]
- Wasner, H.K.; Lessmann, M.; Conrad, M.; Amini, H.; Psarakis, E.; Mir-Mohammad-Sadegh, A. (1996). Biosynthesis of the endogenous cyclic adenosine monophosphate (AMP) antagonist, prostaglandylinositol cyclic phosphate (cyclic PIP), from prostaglandin E and activated inositol polyphosphate in rat liver plasma membranes. Acta Diabetol. 33, 126-138.
- Wasner, H.K.; Salge, U. (1987). Prostaglandylinositol cyclic phosphate, a second messenger for insulin. Excerpta Medica Int. Congr. Ser. 726, 226-231.
- Brautigan, D.L. (2013). Protein ser/thr phosphatases—The ugly ducklings of cell signaling. FEBS J. 280, 324-345. [CrossRef]
- Wasner, H.K.; Salge, U.; Psarakis, E.; Niktopoulos, A. (1997). Insulin resistance, a result of reduced synthesis of prostaglandylinositol cyclic phosphate, a mediator of insulin action? Regulation of cyclic PIP synthase activity by oral antidiabetic and antihypertensive drugs. Acta Diabetol. 34, 257–264.
- Weber, S.; Lemoine, H. and Wasner, H.K. (2000). Prostaglandin deficiency promotes sensitization of adenylyl cyclase. Biol. Chem. 381, 525-529. [CrossRef]
- Kalogriopoulos, N.A.; Lopez-Sanchez, I.; Lin, C.; Ngo, T.; Midde, K.K.; Roy, S.; Aznar, N.; Murray, F.; Garcia-Marcos, M.; Kufareva, I.; Ghassemian, M. and Ghosh, P. (2020). Receptor tyrosine kinases activate heterotrimeric G proteins via phosphorylation within the interdomain cleft of Gai. Proc. Natl. Acad. Sci. US 117, 28763-28774.
- Wasner, H.K.; Weber, S.; Partke, H.J.; Amini-Hadi-Kiashar, H. (1994). Indomethacin treatment causes loss of insulin action in rats: Involvement of prostaglandins in the mechanism of insulin action. Acta Diabetol. 31, 175–182. [CrossRef]
- Wasner, H.; Lemoine, H.; Junger, E.; Lessmann, M.; Kaufmann, R. (1991). Prostaglandyl-inositol cyclic phosphate, a new second messenger. In Prostaglandins, Leukotrienes, Lipoxins and PAF; Bailey, J.M., Ed; Plenum Press: New York, NY, USA, 1991, pp. 153-168.
- Salge, U. Untersuchungen zur biologischen Synthese des cyclo AMP-Antagonisten (PhD-thesis). Heinrich-Heine-University Duesseldorf, Duesseldorf, Germany, 1987.
- Gundula, N.K.V.; Naidu, V.G.M.; Das, U.N. (2018). Amelioration of streptozotocin-induced type 2 diabetes mellitus in Wistar rats by arachidonic acid. Biochem. Biophys. Res. Commun. 496, 105-113. [CrossRef]
- James, D.E.; Stöckli, J. and Birnbaum M.J. (2018). The aetiology and molecular landscape of insulin resistance. Nat. Rev. Mol. Cell Biol. 22, 751-771. [CrossRef]
- Suiso, A.; Glaser, B. and Dor, Y. (2017). Metabolic stress and compromised identity of pancreatic beta cells. Front. Genet. 8. [CrossRef]
- Michell, R.B. (2018). Do inositol supplements enhance phosphatidylinositol supply and thus support endoplasmic reticulum function? Br. J. Nutr. 120, 301-316. [CrossRef]
- Wong, H.; Singh, J.; Go, R.M.; Ahluwalia, N.; Guerrero-Go, M.A. (2019). The effect of mental stress on non-insulin-dependent diabetes: Determining the relationship between catecholamine and adrenergic signals from stress, anxiety, and depression on the physiological changes in the pancreatic hormone secretion. Cureus 11(8), e5474. [CrossRef]
- Pataky, M.W.; Young, W.F. and Nair, K.S. (2021). Hormonal and metabolic changes of aging and the influence of lifestyle modifications. Mayo Clin. Proc. 96(3), 788-814. [CrossRef]
- Reich, K.M.; Gay, C.V.; Frangos, J.A. (1990). Fluid shear stress as a mediator of osteoblast cyclic adenosine monophosphate production. J. Cell. Physiol. 143, 100-104. [CrossRef]
- Ortmeyer, H.K. (1997). Insulin decreases skeletal muscle cAMP-dependent protein kinase (PKA) activity in normal monkeys and increases PKA activity in insulin-resistant Rhesus monkeys. J. Basic Clin. Physiol. Pharmacol. 8, 223-236.
- Eyster, C.A.; Matsuzaki, S.; Newhardt, M.F.; Giorgione, J.R.; Humphries, K.M. (2020). Diabetes induced decreases in PKA signaling in cardiomyocytes: The role of insulin. PLos ONE 15(8): e0231806. [CrossRef]
- Bockus, L.B. and Humphries, K.M. (2015). cAMP-dependent protein kinase (PKA) signaling is impaired in diabetic heart. J. Biol. Chem. 290, 29250-29258. Doi.10.1074/jbcM115.681767.
- London, E.; Bloyd, M. and Stratakis, C.A. (2020). PKA functions in metabolism and resistance to obesity: lessons from mouse and human studies. J. Endocrinol. 246, R51-R64. [CrossRef]
Figure 1.
Cyclic PIP biosynthesis. After hormonal stimulation phospholipase A2 liberates from membrane-bound phospholipids unsaturated fatty acids, which are converted to prostaglandins; phospholipase C liberates various inositol phosphates from which one is converted to ‘activated inositol phosphate’ [8]. Cyclic PIP synthase combines the inositol 1:2-cyclic phosphate part of activated inositol phosphate and PGE to cyclic PIP. The ATP is solely needed to activate cyclic PIP synthase by tyrosine phosphorylation [13]. Apart from the insulin receptor tyrosine kinase, other protein kinases and protein phosphatases discussed in the text are not shown.
Figure 1.
Cyclic PIP biosynthesis. After hormonal stimulation phospholipase A2 liberates from membrane-bound phospholipids unsaturated fatty acids, which are converted to prostaglandins; phospholipase C liberates various inositol phosphates from which one is converted to ‘activated inositol phosphate’ [8]. Cyclic PIP synthase combines the inositol 1:2-cyclic phosphate part of activated inositol phosphate and PGE to cyclic PIP. The ATP is solely needed to activate cyclic PIP synthase by tyrosine phosphorylation [13]. Apart from the insulin receptor tyrosine kinase, other protein kinases and protein phosphatases discussed in the text are not shown.
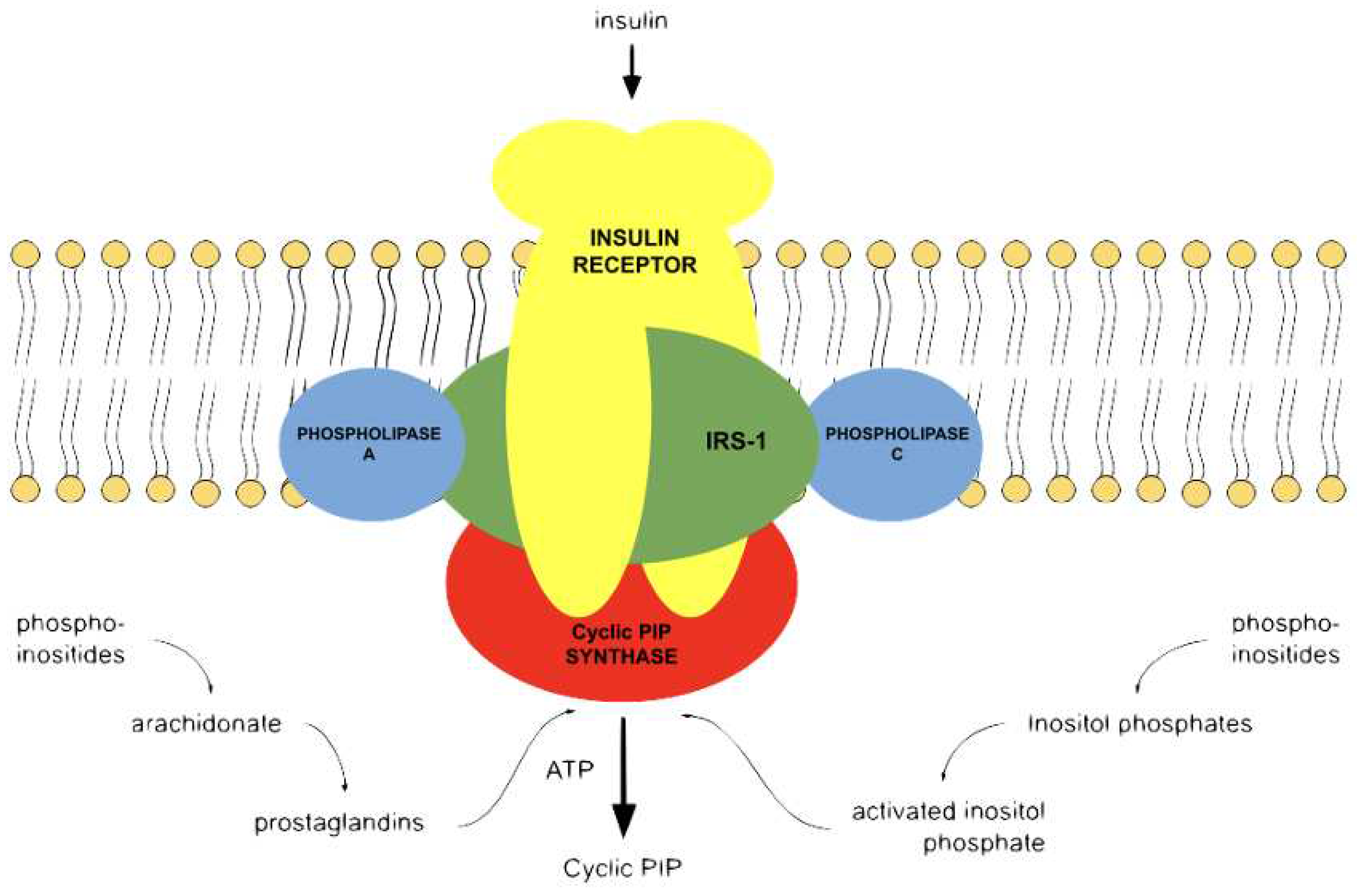
Figure 2.
Scheme illustrating the biosynthesis of cyclic AMP and cyclic PIP, their inhibition of their counterpart and one prominent way of their regulatory properties that is the regulation of the equilibrium between de-phospho- and phospho-form of interconvertible enzymes. (The blue arrows indicate inhibition, the yellow ones activation and the black arrows indicate the synthesis of either cyclic AMP from ATP or of cyclic PIP from PGE and activated inositol phosphate (act. InsP).).
Figure 2.
Scheme illustrating the biosynthesis of cyclic AMP and cyclic PIP, their inhibition of their counterpart and one prominent way of their regulatory properties that is the regulation of the equilibrium between de-phospho- and phospho-form of interconvertible enzymes. (The blue arrows indicate inhibition, the yellow ones activation and the black arrows indicate the synthesis of either cyclic AMP from ATP or of cyclic PIP from PGE and activated inositol phosphate (act. InsP).).
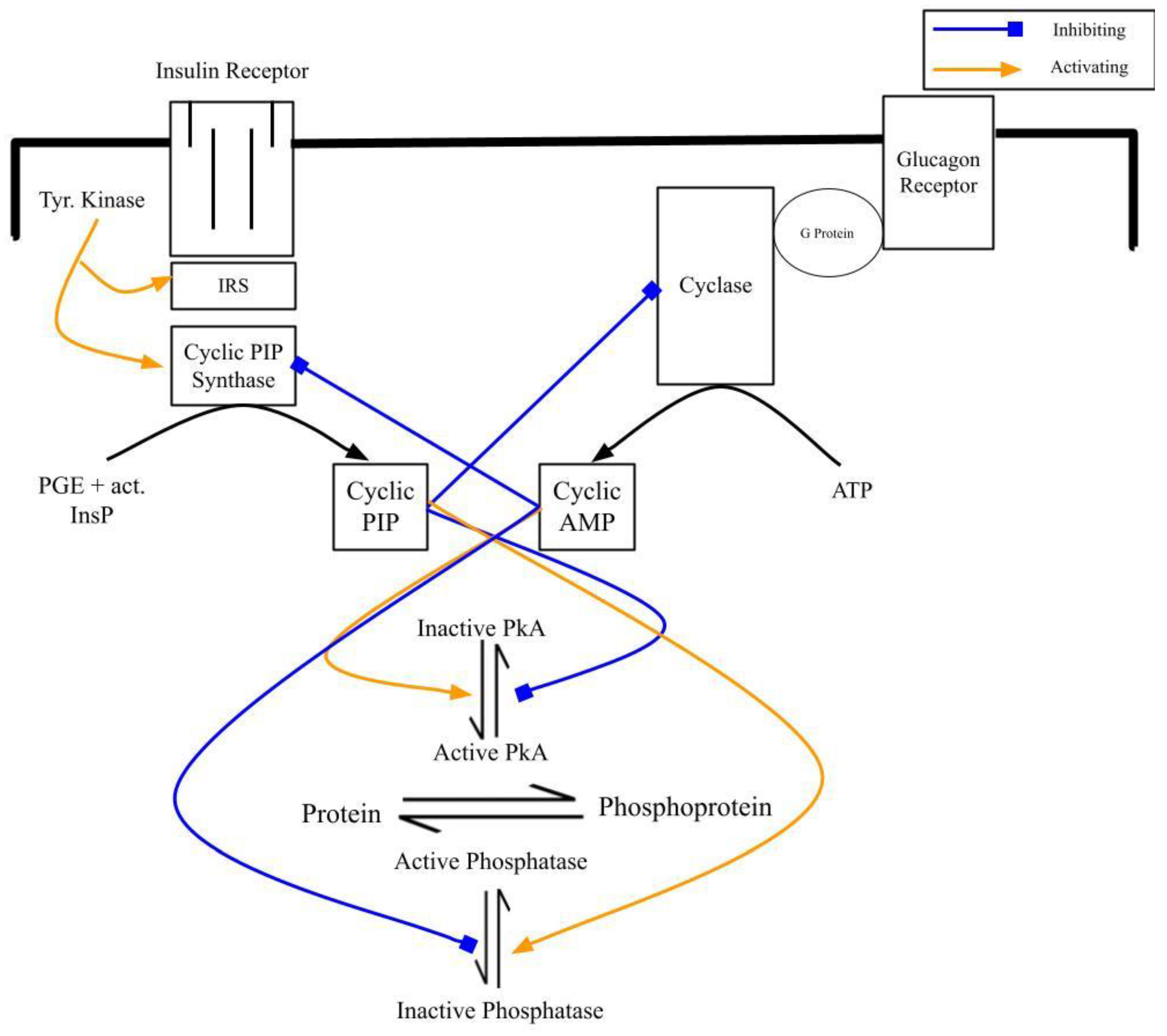

Table 1.
Insulin-stimulated cyclic PIP synthesis in organs of control rats and in organs of type 2 diabetic rats.
Table 1.
Insulin-stimulated cyclic PIP synthesis in organs of control rats and in organs of type 2 diabetic rats.
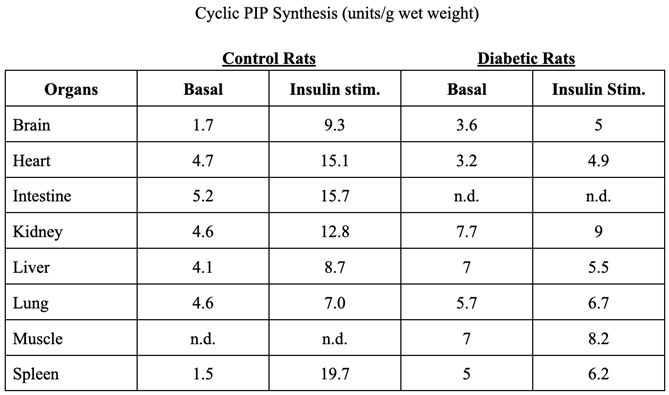
Table 2.
Theoretical calculation which visualizes that an imbalance in the synthesis of cyclic AMP and cyclic PIP leads to an increasing phosphorylation of interconvertible enzymes.
Table 2.
Theoretical calculation which visualizes that an imbalance in the synthesis of cyclic AMP and cyclic PIP leads to an increasing phosphorylation of interconvertible enzymes.
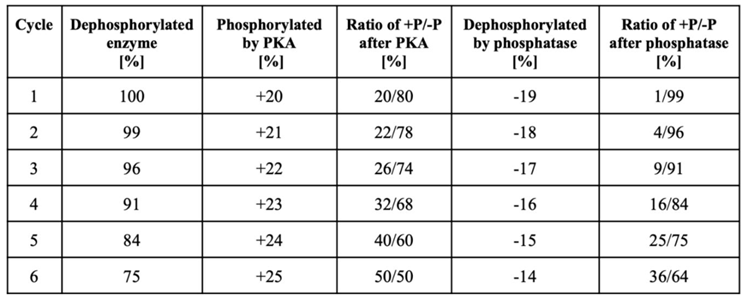
‘Phosphorylation cycle’ means successive stimulation by glucagon and insulin. Abbreviations: ratio +P/-P means the ratio of the phosphorylated and dephosphorylated enzyme.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



