Submitted:
25 September 2023
Posted:
25 September 2023
You are already at the latest version
Abstract
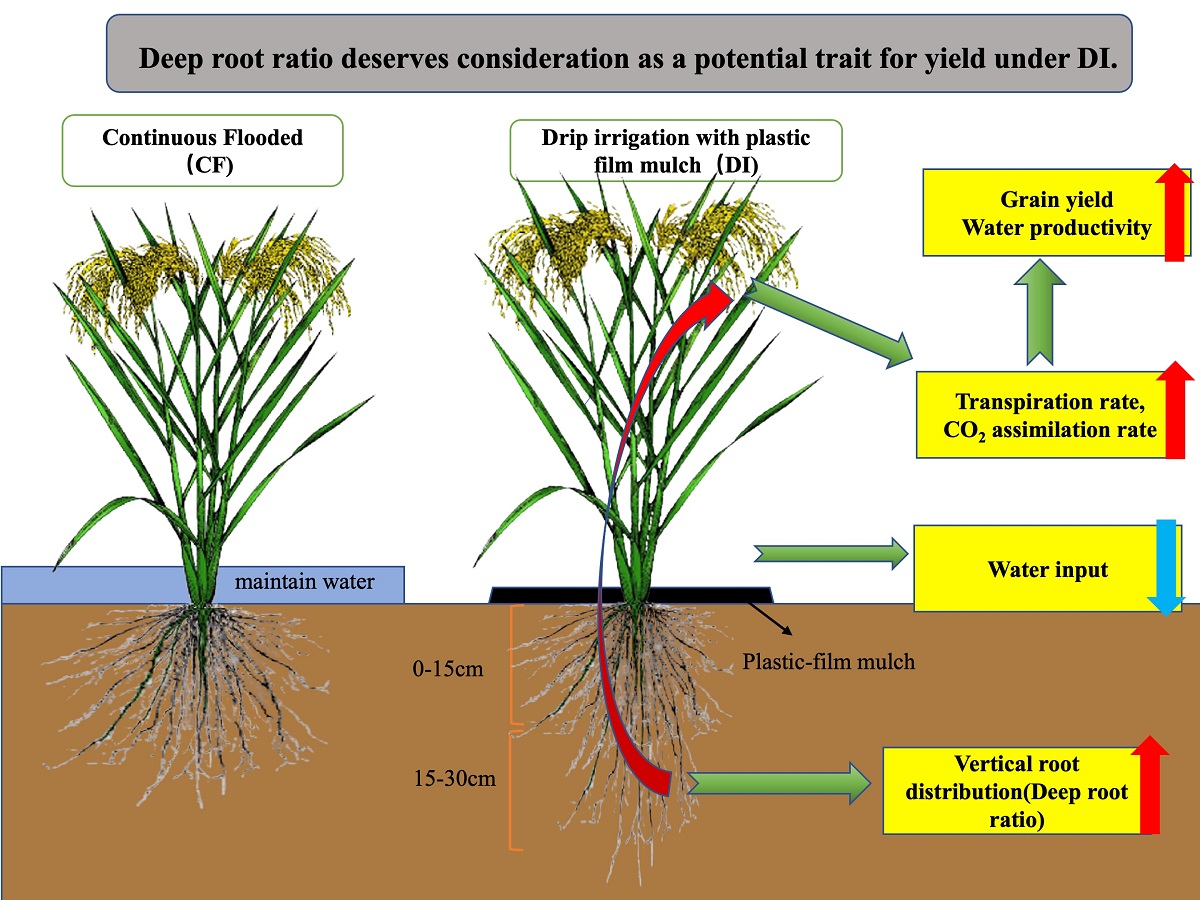
With the escalating water scarcity in agriculture, a novel water-saving technique has emerged: drip irrigation with plastic-film mulch (DI). Root function is crucial for sustaining rice production, and understanding its response to DI is essential. However, few studies have evaluated root systems in rice varietals and examined which kind of root system contributes to improving rice grain yield and water productivity in DI. To fill this knowledge gap, we conducted a two-year field experiment comparing two irrigation systems: continuous flooding (CF) and DI. We analyzed their effectiveness with four rice varieties, including upland, F1 lowland, animal feed lowland, and lowland varieties. Vertical root distribution, root bleed-ing rate, photosynthetic-associated parameters, water productivity, and yield performance were analyzed. In our study, the average grain yield of varieties in the DI system (6.4t/ha) was equivalent to those in the CF system (6.6 t/ha). Compared to CF, DI demonstrated significant water-saving potential, saving approximately 35% of the total water supplied, resulting in higher water productivity. Among the varieties, the deep-root weight of the upland variety significantly increased by 51%. The deep-root ratio was positively correlated to transpiration rate, grain yield, and water productivity, which suggested that it contributed to high transpiration, thus maintaining a high carbon assimilation rate resulting in high yield and water productivity. Therefore, deep roots deserve consideration as a trait potentially corresponding to high yield under DI.
Keywords:
drip irrigation
; root
; varietal difference
; deep root
; plastic-film mulch
1. Introduction
With the escalating water scarcity in agriculture, the need to enhance water-use efficiency in rice cultivation (Oryza sativa L.) has become paramount. Agriculture accounts for 70% of the world's freshwater consumption, a figure that has been exacerbated by factors such as population growth, rapid increases in urbanization, and industrial expansion [1,2]. The dwindling availability of water resources due to these challenges accentuates the need for innovative solutions [3]. Addressing the pressing challenge of increasing rice production to meet the demand of a growing population amidst the backdrop of diminishing water resources, a novel water-saving technique has emerged: drip irrigation with plastic-film mulch (DI) [4,5]. DI is a novel approach to rice cultivation wherein an opaque plastic covering is used to mulch the soil surface without standing water throughout the entire rice growth cycle. This water-efficient method is designed to optimize the utilization of water resources for crop production by inhibiting the transfer of water vapor between the soil and the atmosphere, ultimately minimizing the reduction in crop yields associated with conventional rice farming practices.
As an integral component of a plant’s organs, the root plays a pivotal role by providing anchorage for the plant, enhancing nutrient and water uptake, and synthesizing plant hormones, organic acids, and amino acids [6,7]. However, the ability of the root to effectively carry out its role depends on its type, structure, and modification. This significance extends to water-saving rice production systems, where root growth and structure can be instrumental to the development of the rice plant and grain yield production [8]. Therefore, it is necessary to determine the variation in the root responses of different rice varieties under DI. Enhancements in root morpho-physiological traits are closely intertwined with heightened shoot biomass accumulation, improved efficiency in the use of nitrogen, increased water productivity, and augmented resistance to plant lodging in various crops [9,10,11]. The alteration of root structure stands out as a strategy to foster the development of a deep root system in rice. This can be accomplished by stimulating deeper elongation, particularly in conditions of low soil moisture, and by encouraging increased root branching to optimize soil water extraction [12,13]. The ability of roots to penetrate the soil to greater depths holds paramount importance in establishing a robust root system, which holds particular significance in gramineous crops with their robust roots [14,15]. While differences among cultivars and the application of breeding practices can influence the formation of deep root systems, the potential to induce such systems through effective cultivation management practices remains a compelling avenue for exploration [16].
In recent years, up-to-date knowledge has been provided by many researchers on water productivity, plant growth, physiological characteristics, panicle structure, greenhouse gas emissions (GWP), and grain yield potential of DI [4,5,17,18,19]. However, only a few studies evaluate the varying responses of rice root systems to DI. Therefore, the objectives of this study were (1) to compare the root characteristics, water productivity, and grain yield performance of rice varieties under CF and DI, (2) to compare varietal differences in root patterns in response to DI, and (3) to examine which of the root factors contributes to the improvement of rice grain yield and water productivity in DI.
2. Materials and Methods
2.1. Information on experimental design, soil, and weather
Rice crops were cultivated at the Faculty of Environmental Horticulture research farm, Chiba University, Japan (35.78N, 139.9E), in the summer seasons of 2020 and 2021. The field experimental design of crop management has been previously reported [19]. The soil at the experimental site was a low-humic Andosol with 39.9% sand, 29.9% silt, and 30.2% clay at 0-20 cm depth. The pH of the soil was 5.2, while the amount of organic matter and total N at 0-20 cm depth were 33.3 g kg-1 and 3.6 g kg-1, respectively. The weather parameters, including the mean temperature, sunshine hours, and rainfall during rice-growing periods, are shown in Figure 1.
The field experiment was arranged in a split-plot design (main plot: irrigation system; subplot: variety) with three replicates. The two rice irrigation systems were as follows. (1) CF: continuous flooded system. In this rice production system, each plot had concrete boundaries with an area of 7.5m2. The field was flood-irrigated so that a 4-5 cm water layer above the soil surface was kept until two weeks before harvest. (2) DI: water-saving drip irrigation with plastic-film mulch. In this system, raised beds were covered with black polyethylene film. The plots were buried within the rows at a depth of 40 cm to prevent lateral percolation between neighboring plots. Irrigation was applied via the tube when volumetric soil moisture content fell below 75% of the field capacity in both years. The irrigation schedule was monitored using an FDR soil moisture meter (DIK321A, Daiki Rika Kogyo Co., Ltd). The two irrigation systems were located adjacent to each other, and both were adopted for direct seeding.
Four rice varieties were examined. (1) Norin24 (upland rice, abbreviated as Norin); (2) Mitsuhikari 2003 (F1 lowland rice, abbreviated as Mitsu); (3) Hokuriku 193 (animal feed lowland rice, abbreviated as Hoku); (4) Koshihikari (lowland rice, abbreviated as Koshi). Viable seeds were directly sown in the field on June 8 and June 1 in 2020 and 2021, respectively. The plant density was 69.6 hills m-2. Before planting, chemical fertilizers were administered at a rate of 40 kg ha-1 (N:P: K = 20:20:20) on each field, and then top-dressing fertilization was carried out by applying 10 kg ha-1 of nitrogen through foliar spraying at the panicle initiation stage in both years. Other key cultivation management tasks, such as weed control and pesticide spraying, were conducted following local recommendations.
2.2. Sampling and Measurements
Aboveground and root biomass were estimated 4 or 5 times at the panicle-initiation, heading, grain-filling, and maturity stages. During each sampling instance, we gathered samples from ten rice hills, which collectively occupied an area of 0.5 m2 within each plot. Roots were sampled from each replicate at the panicle-initiation stage (DAS50) and heading stage (DAS85), using the dimensions of the monolith soil sampler (150 mm diameter, 30 cm depth) with the aid of five soil-sampling cores. Soil samples were divided into 0-15 cm and 15-30 cm segments and washed carefully on a 0.5 mm stainless steel sieve under running water to separate the roots. The data ignored the roots below 30 cm for both CF and DI. The portion of each root sample was used for the measurement of root length. Roots were arranged on a glass tray, scanned using a scanner (Epson GT-X970, Seiko Epson Corp., Nagano, Japan), and analyzed using ImageJ as described by Tajima et al., 2013 [20]. Finally, the remaining roots were oven-dried at 80℃ for 72 h and weighed.
To determine the differences in root bleeding rate among varieties grown under CF and DI, 5 hills were marked on each plot at the panicle-initiation stage on July 20 and July 24 in 2020 and 2021. The rice stems were trimmed to a height of 10 centimeters above the soil level at 6:00 PM local time. We then placed 5 grams of cotton wool on the upper portion of each cut stem and enclosed it with a polyethylene bag. After 12 hours, we retrieved the cotton wool and determined the root bleeding rate by measuring the increase in weight of the cotton wool.
A portable photosynthetic apparatus system of Li-6400 (LI-COR, Lincoln, NE, USA) was used to measure the CO2 assimilation rate (An), stomatal conductance (gs), and transpiration rate in the flag leaf of each rice variety under the two water regimes at the early grain-filling stage in 2021. The measurements were carried out when photosynthesis was most active (09:00 to 12:00 local time). Throughout the measurement period, the photosynthetic photon flux density level, average leaf temperature, and average vapor pressure deficit were maintained at 1000 μmolm⁻²s⁻¹, 25°C, and 1.5 kPa, respectively.
Rice plants from a 1 m2 area in each plot were collected to determine grain yield at physiological maturity each year. We recorded the count of panicles for each treatment. To determine the rough grain yield, the panicles were threshed and weighed for each rice variety. The grain weights were recorded for each variety under CF and DI after the grains were dried to a moisture content of 14%. The percentage of grains that had ripened was calculated by dividing the count of filled grains by the total number of grains. Harvest index was defined as grain yield per biomass at maturity. Water productivity was calculated as the ratio of the total quantity of water supplied (irrigation and rainfall) to each variety per grain yield.
2.3. Data analysis
Analysis of variance (ANOVA) was performed using SPSS 20.0 (IBM, New York, USA). The statistical model used included sources of variation due to replication, year (Y), variety (V), irrigation regime (IR), and the interaction of Y x V, Y x IR, V x IR, and Y x V x IR. Data from each sampling date were analyzed separately. The means were separated by Duncan’s multiple range test when the effects were significant in each year.
3. Results
3.1. Soil water content under DI
The precipitation in the CF and DI areas during the growth period is presented in Figure 1 and Figure 2, respectively. The levels were generally similar over two years of field experiments except for certain months. The amounts of irrigation water supplied to the CF area were 1085 and 985 mm in 2020 and 2021, respectively. The total irrigation water supplied in 2020 under DI conditions increased by 25% in 2021. However, the seasonal precipitation differed in 2020, lower by 19.4% compared to 2021. Soil moisture content ranged from 67.4 to 87.2% in 2021 and 62.3 to 83.4% in 2020. Average soil moisture during the growth period under DI was 76.0% in 2021, higher than the 73.9% average in 2020, due to the higher total quantities of water supplied. Compared with CF, DI saved 35.1±1.5% of the total water supplied in both years.
3.2. Root growth in DI
3.2.1. Shoot, root biomass, and root-to-shoot ratio
Shoot and root biomass were not significantly affected by the irrigation system, although significant differences among the varieties were found (Figure 3). Shoot biomass in DI was equivalent to or lower than that in CF in 2021, except for the variety Norin, which had a longer growth duration under DI. Varietal differences in shoot biomass were observed from the early tillering stage (DAS65) in DI, earlier than the heading stage (DAS90) in CF. Root biomass was generally higher mid-season (from tillering to early grain filling) than early or late in the season. Root biomass in DI was similar to or lower than that in CF in 2021. The root-to-shoot ratio of all varieties decreased with growth. The root-to-shoot ratio in DI was lower than that in CF in 2021, except for the variety Hoku, which probably competed more for soil nutrients under DI in the early tillering stage. The differences in DI were most obvious in the early growth stage but were reduced at the heading stage.
3.2.2. Root distribution pattern
Root length and weight at the soil depth of 0-30 cm were significantly higher in CF than in DI at the heading stage of rice growth (Figure 4). Root length and weight of all varieties were significantly lower in DI than in CF in both the surface and subsurface layers at the heading stage, except for Norin. As to the varietal difference, Norin developed more subsurface root biomass under DI at the heading stage, followed by Hoku.
The deep-root ratio tended to be lower under DI than under CF, even though no significant differences in deep-root ratio were observed between the water regimes of either DAS50 or DAS85 (Figure 5). The deep-root ratio ranged from 0.1-0.19 under CF and ranged from 0.08-0.24 under DI, respectively. Generally, varietal differences in deep-root ratio in CF were smaller in DAS50 than in DAS85. However, the upland variety (Norin) was significantly higher in DI compared to CF in both DAS50 and DAS85. No significant differences in deep-root ratio were observed between water regimes of either DAS50 or DAS85.
3.2.3. Root bleeding rate
The average root bleeding rate of rice varieties under CF was significantly higher than that under DI at the panicle-initiation stage in both years (Figure 6). In particular, Koshi and Mitsu were significantly higher under CF than DI in 2020 and 2021, respectively. The average root bleeding rate of the 2-year data decreased by 17.5% and 15.3%, respectively, under DI compared with CF. Within the various rice varieties, Mitsu consistently displayed the highest value under CF conditions, whereas Koshi consistently exhibited the lowest value under DI in both years.
3.3. Photosynthesis-associated parameters
An, gs, and E were not significantly different between CF and DI (Table 1). However, the variety and the interaction between the water regime and variety had a significant effect on the photosynthesis-associated parameters. Mitsu had the highest value in all photosynthetic-associated parameters in CF. An, gs, and E were significantly higher in Norin under DI than in the other three varieties. However, they were significantly lower in Koshi under DI compared to CF.
3.4. Yield, yield component, harvest index, and water productivity
Table 2 shows the yield component. The panicle number, spikelet per panicle, ripening ratio, and 1000-grain weight were all significantly affected by the irrigation regime. On average over two years, DI led to an increase in the panicle number and spikelet per panicle, but a decrease in the ripening ratio and 1000-grain weight of the varieties compared to CF. Among the varieties under DI, the upland variety (Norin) significantly increased the number of spikelets per panicle, which contributed to a higher yield than occurred under CF. The F1 lowland and lowland varieties significantly decreased in ripening ratio and 1000-grain weight; this contributed to the lower yield under DI compared to CF. The ripening ratio and 1000-grain weight of rice varieties were significantly lower under DI than CF.
Across both years, the average grain yield of the different rice varieties showed no significant difference when DI and CF were compared. However, there was a significant impact on yield due to the interaction between the irrigation system and rice variety in the year 2021. Among these varieties, lowland types, such as Mitsu and Koshi, consistently yielded better results under CF conditions compared to DI, while the upland variety, Norin, demonstrated higher yields under DI. Furthermore, variations in yields were observed for the animal feed lowland variety Hoku under the different irrigation systems. Though not significant, the small deep-root ratio trend was found to be stronger under CF than DI, except for Norin, at the panicle-initiation stage and heading stage. The irrigation system did not demonstrate a significant effect on the harvest index for the examined varieties in either of the two years. Nevertheless, there were discernible differences in the harvest indexes of the varieties between CF and DI in 2021. Notably, among all the varieties and across both irrigation systems, Mitsu consistently displayed the highest harvest index. DI significantly increased the average water productivity in both years compared with CF. The average water productivity in DI was 0.56 kg grain m-3 water; 1.5-1.6 times higher than in CF. All the varieties achieved higher water productivity under DI in comparison to CF, leading to more efficient use of irrigation water.
4. Discussion
Prior to this study, little information was available describing varietal differences in rice root morphology and physiology under CF and DI systems. Our results indicated that in DI, root growth at the soil surface was comparable with that under CF during the vegetative phase (Figure 4). Araki et al., 2005 [21] and Luo et al., 2023 [22] revealed that plants first draw water from the surface layer, and subsequently, the area of water extraction gradually shifts downward through the soil profile. When the water content in the shallow layer (0-25 cm depth) was abundant, over 90% of the water uptake occurred in this shallower zone [23,24]. Maintaining paddy soils in a wet but not continuously saturated state during the vegetative phase has the effect of preserving predominantly aerobic soil conditions [25]. These particular soil management practices have significantly facilitated better root growth at the surface layer with the DI approach. The results of our study suggest that current levels of root growth at the surface in CF can be achieved under DI. Our results are consistent with those of Bouman et al., 2006 [26], who showed no significant differences in root characteristics of rice grown under CF compared with an alternate wetting and drying (AWD) irrigation system. We observed that there were large genotypic differences in root distribution patterns. Regarding the deep-root ratio, the lowland varieties produced comparable biomass to subsurface root growth in DI; however, upland varieties significantly allocated more root growth at the lower soil horizons in DI compared to CF (Figure 5). This is in line with the conclusion of Kato et al., 2010 [27], who used aerobic cultivation technology. It is known that the soil moisture content, soil nutrient status, and soil physical properties can profoundly influence plant root growth [28].
The deep root system in upland rice varieties enables them to harness water from the deeper soil layers, a resource that remains out of reach for cultivars with shallow root systems. This capability has proven effective in preserving yields, especially under reduced soil moisture conditions [29,30,31,32]. For a more precise assessment of rice's deep-root characteristics under water management, soil samples were taken and measured up to a depth of 60 cm by Kato et al., 2007 [33]. However, due to the presence of concrete boundaries under CF, our study was limited to measurements up to 30 cm below the surface. This discrepancy revealed that the rice varieties under DI had greater deep root growth potential than under CF. The root bleeding rates conducted during the panicle-initiation stage over two consecutive years identified substantial genotypic differences among the rice varieties (Figure 6). Interestingly, lowland varieties, such as Mitsu and Koshi, exhibited significantly higher root activity under CF than under DI. This observation aligns with the findings reported by Matsuo et al., 2009 [34], where a decrease in root activity for lowland variety Koshi under water-saving cultivation systems was similarly noted. Furthermore, significant differences in root activity among upland (Norin) and animal feed varieties (Hoku) under CF and DI were not observed (Figure 6). This could be attributed to the influence of root- and stem-xylem resistance mechanisms. The presence and impact of root- and stem-xylem resistance mechanisms represent essential factors that warrant consideration when assessing disparities in root activity between two irrigation systems [35,36]. These resistance mechanisms can substantially influence the transport of water and sap within the plant, potentially masking or reducing variations in root bleeding rates [37].
The results of this study showed no significant difference among the photosynthetic-associated parameters under CF and DI. However, it was observed that substantial genotypic variation existed among the tested rice varieties (Table 1). The transpiration rate of Norin showed a significant increase of 11% under DI due to greater deep-root development (Figure 7a). Deep roots are instrumental in enhancing a plant's resilience in drought conditions, ensuring that transpiration can persist even when surface soil dries out. This strategy promotes drought tolerance and improves transpiration efficiency [38,39]. The regulation of deep-root functioning and distribution, adjustments in canopy size, and the synthesis of the hormone abscisic acid (ABA) in both roots and leaves collectively play a central role in controlling leaf water potential (LWP) and enhancing leaf transpiration efficiency [40,41,42]. Under severe drought-stress conditions, abscisic acid (ABA) typically assumes a more critical and pronounced role in the regulation of plant responses to water stress. However, the significance of ABA in plant responses may not be as pronounced in less severe drought-stress conditions. Shi et al., 2015 [43] reported that both upland cultivars (Gaoshan1) and lowland cultivars (Nipponbare) demonstrated the ability to open their stomata for transpiration even under slight-to-moderate drought conditions, which allowed the effective loading of root-generated ABA into the xylem through apoplastic by-pass flow, subsequently reducing ABA exudation. Furthermore, root structural distributions, including deep-root ratio and root branching, are likely to be promoted under aerobic conditions [44]. A study by Kato et al., 2013 [45] in northeastern Thailand utilized an irrigation system for aerobic cultivation and obtained similar results, supporting the notion that aerobic conditions can enhance root structure distributions. Deep-rooted traits often exhibit better transpiration efficiency, which means they can achieve higher photosynthesis per unit of water transpired [46]. Our results indicated that the deep roots of the upland variety (Norin) could potentially contribute to a high transpiration rate, thus maintaining a high assimilation rate under DI in comparison to CF (Figure 7a, Fig 8). Additionally, while deep roots are one of the factors influencing transpiration, other environmental factors (i.e., temperature, humidity, and light intensity) also contribute to the regulation of transpiration rates [47]. In future research, it will be essential to continue investigating the complex interplay among these factors to gain a more comprehensive understanding of how plants manage water resources and optimize transpiration rates to adapt to various environmental conditions, especially in aerobic and mild water-stress conditions. On the other hand, the lowland variety (Koshi) significantly decreased gas-exchange performance (An, gs, and E), likely due to the lower plasticity index for lower root bleeding rate (Table 1, Figure 6). Morita et al., 2000; He et al., 2019; Ansari et al., 2004 and Liu et al., 2021 [48,49,50,51] reported that root bleeding is closely linked to the active absorption of water and nutrients, photosynthate transport, and redistribution efficiency in the root system. In a study by Li et al. conducted on winter maize in 2022 [52], they similarly observed a significant effect of water management on various factors, such as root oxidation activity, cytokinin concentrations in both roots and shoots, the rate of photosynthesis in leaves, and the activities of essential enzymes responsible for converting sucrose to starch in grains. The reduction in yield observed in the lowland varieties under the DI system could be attributed to alterations in root activity, function, and morphology, which may disrupt the effective transport and redistribution of photosynthates within the plants.
In general, the primary goals of water-saving technologies are to achieve higher water-use efficiency while maintaining or even increasing crop yields [53]. Our research findings indicate that the average grain yield under DI was equivalent to that under the CF system. Meanwhile, DI demonstrated significant water-saving potential, saving approximately 35% of the total water supplied compared to CF, resulting in water productivity that was on average 53% higher (Table 2). Among the varieties, DI significantly boosted the grain yield of the upland variety (Norin) and the comparable yield of animal-feed lowland variety (Hoku). The increase in yield can be primarily attributed to the higher number of spikelets per square meter (m²) in these cultivars, despite a trade-off in ripening ratio and individual grain weight. However, lowland rice varieties (Mitsu and Koshi) produced significantly higher yields under CF. This higher yield can be attributed to their superior ripening ratio and grain weight under CF conditions, offsetting potential trade-offs in other components. On the other hand, a sizeable genotypic variation was observed between root traits and grain yield. The root systems of the upland variety (Norin) under DI were characterized by well-developed root growth on the surface and produced more deep roots compared to the CF sample. Therefore, it can be inferred that those characteristics contributed to a high transpiration rate, thus maintaining a high assimilation rate resulting in the high-yield performance and water productivity of the upland variety in production. Furthermore, there was a significant positive relationship between deep-root ratio, grain yield (0.745*), and water productivity (0.861*) under DI (Figure 7b, c). The results indicated that vertical root distribution (i.e., the deep-root ratio) during the vegetative phase influenced the yield and water productivity. The fact that animal-feed lowland rice (Hoku) maintained a comparable yield under DI compared to CF is likely due to the high root-to-shoot ratio during the early growth stage (Figure 3). This high root-to-shoot ratio of Hoku during the early growth stage contributed to the prioritization of root development, which can increase a plant's capacity to take up essential nutrients, promoting healthy growth and development [54]. These adaptations in the root system facilitated a greater allocation of photosynthetic products to the shoots in particular leaves, ultimately leading to a reduced root-to-shoot ratio and an enhanced yield while conserving water [55,56]. The low yield of lowland varieties in DI was likely due to the low root vigor and water uptake. The relationship between root bleeding rate and grain yield in DI differed between the lowland varieties (Mitsu and Koshi) and the other two. There was a significant positive relationship between root bleeding rate and grain yield under CF (Figure 4). This highlights the importance of the root activity of lowland rice in influencing grain yield in CF conditions, where root activity plays a critical role in crop performance.
Development and identification of suitable genotypes and crop management options are underway worldwide to make agriculture more resource-use efficient and productive under drip irrigation with plastic-film mulch [57]. As a first for future studies, exploring varietal differences under drip irrigation is an essential avenue for future research in agriculture. To further investigate and select adaptable crop varieties, developing guidelines for selecting rice varieties optimized for drip irrigation with plastic-film mulch is crucial, considering local climate, soil conditions, plant density, and fertilizer management. Secondly, the alterations in the root structures of deep root systems result from increased root branches [14]. Further research is needed to improve the development of the rice root system in drip irrigation systems, including the study of adaptive responses, such as adventitious root emergence, lateral root branching, and water uptake. Thirdly, because of the limited extent of the research on root morphology and physiological characteristics in DI systems, additional studies are essential to elucidate the specific traits that contribute to optimal crop performance under DI conditions.
5. Conclusions
In our study, the average grain yield under DI was equivalent to that under the CF system. Meanwhile, DI demonstrated significant water-saving potential, saving approximately 35% of the total water supplied in comparison with CF, resulting in higher water productivity. Under DI, the underlying factors in the high yield of the upland variety were the well-developed shallow roots and the greater number of deep roots contributing to high transpiration, maintaining a high carbon assimilation rate that resulted in higher yield and water productivity. The comparable yield of the animal feed variety was likely due to the high root-to-shoot ratio during the early growth stage, whereas the low yield in the lowland variety under DI was attributed to a significant decrease in the root bleeding rate compared to CF. Our results indicate that deep roots deserve consideration as a trait potentially corresponding to high yield under DI. Meanwhile, the ideal rice varieties for DI should have well-developed shallow roots, more deep-root morphological traits, higher root-to-shoot ratio at the early stage, and greater root activity, contributing to photosynthetic performance; these are essential for the productivity of the DI system, resulting in higher grain yield and water productivity. Further research is needed to improve the development of the morphological or physiological traits of the rice root system in DI, including the study of adaptive responses, such as adventitious root emergence, lateral root branching, and water uptake.
Author Contributions
Conceptualization, J.W., O.F., and A.I.; methodology, J.W.; software, J.W.; validation, J.W., O.F., and A.I.; formal analysis, J.W.; investigation, J.W.; resources, J.W.; data curation, J.W. and A.I.; writing—original draft preparation, J.W.; writing—review and editing, O.F. and A.I.; visualization, J.W. and A.I.; supervision, J.W. and A.I.; project administration, J.W.; funding acquisition, J.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by JST SPRING, grant number JPMJSP2019.
Data Availability Statement
Not applicable
Acknowledgments
We would like to express our appreciation for the assistance provided by team members during the experiments. Additionally, we sincerely appreciate the work of the editor and the reviewers of the present paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Belder, P. Water Saving in Lowland Rice Production: An Experimental and Modelling Study. PhD thesis, Wageningen university, Netherlands, 15 June 2005. https://edepot.wur.nl/121681.
- Bouman, B. A. M. A Conceptual Framework for the Improvement of Crop Water Productivity at Different Spatial Scales. Agric. Syst. 2007, 93 (1–3), 43–60. [CrossRef]
- Asian Development Bank. Asian Water Development Outlook 2020: Advancing Water Security across Asia and the Pacific, 0 ed.; Asian Development Bank: Manila, Philippines, 2020. [CrossRef]
- He, H.; Ma, F.; Yang, R.; Chen, L.; Jia, B.; Cui, J.; Fan, H.; Wang, X.; Li, L. Rice Performance and Water Use Efficiency under Plastic Mulching with Drip Irrigation. PLoS ONE 2013, 8 (12), e83103. [CrossRef]
- Fawibe, O. O.; Honda, K.; Taguchi, Y.; Park, S.; Isoda, A. Greenhouse Gas Emissions from Rice Field Cultivation with Drip Irrigation and Plastic Film Mulch. Nutr. Cycl. Agroecosystems 2019, 113 (1), 51–62. [CrossRef]
- Ortíz-Castro, R.; Contreras-Cornejo, H. A.; Macías-Rodríguez, L.; López-Bucio, J. The Role of Microbial Signals in Plant Growth and Development. Plant Signal. Behav. 2009, 4 (8), 701–712. [CrossRef]
- Jung, J. K. H.; McCouch, S. Getting to the Roots of It: Genetic and Hormonal Control of Root Architecture. Front. Plant Sci. 2013, 4. [CrossRef]
- Islam, M. D. D.; Price, A. H.; Hallett, P. D. Contrasting Ability of Deep and Shallow Rooting Rice Genotypes to Grow through Plough Pans Containing Simulated Biopores and Cracks. Plant Soil. 2021, 467 (1–2), 515–530. [CrossRef]
- Kassam, A.; Stoop, W.; Uphoff, N. Review of SRI Modifications in Rice Crop and Water Management and Research Issues for Making Further Improvements in Agricultural and Water Productivity. Paddy Water Environ. 2011, 9 (1), 163–180. [CrossRef]
- Chu, G.; Chen, T.; Wang, Z.; Yang, J.; Zhang, J. Morphological and Physiological Traits of Roots and Their Relationships with Water Productivity in Water-Saving and Drought-Resistant Rice. Field Crops Res. 2014, 162, 108–119. [CrossRef]
- Zhao, X.; Hu, Y.; Liang, B.; Chen, G.; Feng, L.; Pu, T.; Sun, X.; Yong, T.; Liu, W.; Liu, J.; Du, J.; Yang, F.; Wang, X.; Yang, W. Coordination of Density and Nitrogen Fertilization Improves Stalk Lodging Resistance of Strip-Intercropped Maize with Soybeans by Affecting Stalk Quality Traits. Agriculture 2023, 13 (5), 1009. [CrossRef]
- Zhan, A.; Lynch, J. P. Reduced Frequency of Lateral Root Branching Improves N Capture from Low-N Soils in Maize. J. Exp. Bot. 2015, 66 (7), 2055–2065. [CrossRef]
- Kou, X.; Han, W.; Kang, J. Responses of Root System Architecture to Water Stress at Multiple Levels: A Meta-Analysis of Trials under Controlled Conditions. Front. Plant Sci. 2022, 13, 1085409. [CrossRef]
- Miyazaki, A.; Arita, N. Deep Rooting Development and Growth in Upland Rice NERICA Induced by Subsurface Irrigation. Plant Prod. Sci. 2020, 23 (2), 211–219. [CrossRef]
- Feddes, R. A.; Raats, P. A. C. Parameterizing the Soil – Water – Plant Root System. Wageningen UR Frontis Series. 2004, vol.6, Unsaturated-zone Modeling, Progress, Challenges and Applications, pp.95-141 Kluwer, Dordrecht. https://edepot.wur.nl/35358.
- Tatsumi, Y.; Murakami, S.; Ishibashi, Y.; Iwaya-Inoue, M. Characteristics for Deep Root System of a Drought Tolerant Cowpea Cultivar. Cryobiology and Cryotechnology, Japan, 1st September 2009. [CrossRef]
- Fawibe, O. O.; Hiramatsu, M.; Taguchi, Y.; Wang, J.; Isoda, A. Grain Yield, Water-Use Efficiency, and Physiological Characteristics of Rice Cultivars under Drip Irrigation with Plastic-Film-Mulch. J. Crop Improv. 2020, 34 (3), 414–436. [CrossRef]
- Park, S.; Nishikoji, H.; Takahashi, S.; Fawibe, O. O.; Wang, P.; Isoda, A. Rice Cultivation under Drip Irrigation with Plastic Film Mulch in the Kanto Area of Japan. Univers. J. Agric. Res. 2021, 9 (4), 101–110. [CrossRef]
- Wang, J.; Fawibe, O. O.; Fawibe, K. O.; Isoda, A. Water Productivity, Sink Production and Varietal Differences in Panicle Structure of Rice (Oryza Sativa L.) under Drip Irrigation with Plastic-Film Mulch. Field Crops Res. 2023, 291, 108790. [CrossRef]
- Tajima, R.; Kato, Y. 【Short Report】A Quick Method to Estimate Root Length in Each Diameter Class Using Freeware ImageJ. Plant Prod. Sci. 2013, 16 (1), 9–11. [CrossRef]
- Araki, R.; Yamanaka, M. D.; Murata, F.; Hashiguchi, H.; Oku, Y.; Sribimawati, T.; Kudsy, M.; Renggono, F. Seasonal and Interannual Variations of Diurnal Cycles of Wind and Cloud Activity Observed at Serpong, West Jawa, Indonesia. J. Meteorol. Soc. Jpn. Ser II 2006, 84A, 171–194. [CrossRef]
- Luo, Z.; Fan, J.; Shao, M.; Hu, W.; Yang, Q.; Zhang, S. Soil Water Dynamics and Groundwater Evolutions of Check Dams under Natural Rainfall Reduction in Semi-Arid Areas. J. Hydrol. 2023, 617, 129099. [CrossRef]
- Gabriel, C. E.; Kellman, L. Examining Moisture and Temperature Sensitivity of Soil Organic Matter Decomposition in a Temperate Coniferous Forest Soil; preprint; Biogeochemistry: Soils, 2011. [CrossRef]
- Wu, Y.; Du, T.; Li, F.; Li, S.; Ding, R.; Tong, L. Quantification of Maize Water Uptake from Different Layers and Root Zones under Alternate Furrow Irrigation Using Stable Oxygen Isotope. Agric. Water Manag. 2016, 168, 35–44. [CrossRef]
- Alam, Md. K.; Bell, R. W.; Hasanuzzaman, M.; Salahin, N.; Rashid, M. H.; Akter, N.; Akhter, S.; Islam, M. S.; Islam, S.; Naznin, S.; Anik, M. F. A.; Apu, Md. M. R. B.; Saif, H. B.; Alam, M. J.; Khatun, Mst. F. Rice (Oryza Sativa L.) Establishment Techniques and Their Implications for Soil Properties, Global Warming Potential Mitigation and Crop Yields. Agronomy 2020, 10 (6), 888. [CrossRef]
- Bouman, B. A. M.; Yang, X.; Wang, H.; Wang, Z.; Zhao, J.; Chen, B. Performance of Aerobic Rice Varieties under Irrigated Conditions in North China. Field Crops Res. 2006, 97 (1), 53–65. [CrossRef]
- Kato, Y.; Okami, M. Root Growth Dynamics and Stomatal Behaviour of Rice (Oryza Sativa L.) Grown under Aerobic and Flooded Conditions. Field Crops Res. 2010, 117 (1), 9–17. [CrossRef]
- Price, A. H.; Steele, K. A.; Gorham, J.; Bridges, J. M.; Moore, B. J.; Evans, J. L.; Richardson, P.; Jones, R. G. W. Upland Rice Grown in Soil-Filled Chambers and Exposed to Contrasting Water-Deficit Regimes I. Root Distribution, Water Use and Plant Water Status. Field Crops Res. 2002, 76 (1), 11–24. [CrossRef]
- Kato, Y.; Katsura, K. Rice Adaptation to Aerobic Soils: Physiological Considerations and Implications for Agronomy. Plant Prod. Sci. 2014, 17 (1), 1–12. [CrossRef]
- Uga, Y.; Kitomi, Y.; Ishikawa, S.; Yano, M. Genetic Improvement for Root Growth Angle to Enhance Crop Production. Breed. Sci. 2015, 65 (2), 111–119. [CrossRef]
- Kim, Y.; Chung, Y. S.; Lee, E.; Tripathi, P.; Heo, S.; Kim, K.-H. Root Response to Drought Stress in Rice (Oryza Sativa L.). Int. J. Mol. Sci. 2020, 21 (4), 1513. [CrossRef]
- Ajmera, I.; Henry, A.; Radanielson, A. M.; Klein, S. P.; Ianevski, A.; Bennett, M. J.; Band, L. R.; Lynch, J. P. Integrated Root Phenotypes for Improved Rice Performance under Low Nitrogen Availability. Plant Cell Environ. 2022, 45 (3), 805–822. [CrossRef]
- Kato, Y.; Kamoshita, A.; Yamagishi, J.; Imoto, H.; Abe, J. Growth of Rice ( Oryza Sativa L.) Cultivars Under Upland Conditions With Different Levels of Water Supply3. Root System Development, Soil Moisture Changeand Plant Water Status. Plant Prod. Sci. 2007, 10 (1), 3–13. [CrossRef]
- Matsuo, N.; Mochizuki, T. Growth and Yield of Six Rice Cultivars under Three Water-Saving Cultivations. Plant Prod. Sci. 2009, 12 (4), 514–525. [CrossRef]
- Brodersen, C. R.; McElrone, A. J. Maintenance of Xylem Network Transport Capacity: A Review of Embolism Repair in Vascular Plants. Front. Plant Sci. 2013, 4. [CrossRef]
- Lamarque, L. J.; Corso, D.; Torres-Ruiz, J. M.; Badel, E.; Brodribb, T. J.; Burlett, R.; Charrier, G.; Choat, B.; Cochard, H.; Gambetta, G. A.; Jansen, S.; King, A.; Lenoir, N.; Martin-StPaul, N.; Steppe, K.; Van Den Bulcke, J.; Zhang, Y.; Delzon, S. An Inconvenient Truth about Xylem Resistance to Embolism in the Model Species for Refilling Laurus Nobilis L. Ann. For. Sci. 2018, 75 (3), 88. [CrossRef]
- Vilagrosa, A.; Chirino, E.; Peguero-Pina, J. J.; Barigah, T. S.; Cochard, H.; Gil-Pelegrín, E. Xylem Cavitation and Embolism in Plants Living in Water-Limited Ecosystems. In Plant Responses to Drought Stress; Aroca, R., Ed.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2012; pp 63–109. [CrossRef]
- Kulkarni, M.; Soolanayakanahally, R.; Ogawa, S.; Uga, Y.; Selvaraj, M. G.; Kagale, S. Drought Response in Wheat: Key Genes and Regulatory Mechanisms Controlling Root System Architecture and Transpiration Efficiency. Front. Chem. 2017, 5, 106. [CrossRef]
- Li, H.; Ma, X.; Lu, Y.; Ren, R.; Cui, B.; Si, B. Growing Deep Roots Has Opposing Impacts on the Transpiration of Apple Trees Planted in Subhumid Loess Region. Agric. Water Manag. 2021, 258, 107207. [CrossRef]
- Siopongco, J. Dlc.; Yamauchi, A.; Salekdeh, H.; Bennett, J.; Wade, L. J. Growth and Water Use Response of Doubled-Haploid Rice Linesto Drought and Rewatering during the Vegetative Stage. Plant Prod. Sci. 2006, 9 (2), 141–151. [CrossRef]
- Zegada-Lizarazu, W.; Monti, A. Deep Root Growth, ABA Adjustments and Root Water Uptake Response to Soil Water Deficit in Giant Reed. Ann. Bot. 2019, 124 (4), 605–615. [CrossRef]
- Parkash, V.; Singh, S. A Review on Potential Plant-Based Water Stress Indicators for Vegetable Crops. Sustainability 2020, 12 (10), 3945. [CrossRef]
- Shi, L.; Guo, M.; Ye, N.; Liu, Y.; Liu, R.; Xia, Y.; Cui, S.; Zhang, J. Reduced ABA Accumulation in the Root System Is Caused by ABA Exudation in Upland Rice (Oryza Sativa L. Var. Gaoshan1) and This Enhanced Drought Adaptation. Plant Cell Physiol. 2015, 56 (5), 951–964. [CrossRef]
- Gowda, V. R. P.; Henry, A.; Yamauchi, A.; Shashidhar, H. E.; Serraj, R. Root Biology and Genetic Improvement for Drought Avoidance in Rice. Field Crops Res. 2011, 122 (1), 1–13. [CrossRef]
- Kato, Y.; Tajima, R.; Homma, K.; Toriumi, A.; Yamagishi, J.; Shiraiwa, T.; Mekwatanakarn, P.; Jongdee, B. Root Growth Response of Rainfed Lowland Rice to Aerobic Conditions in Northeastern Thailand. Plant Soil 2013, 368 (1–2), 557–567. [CrossRef]
- Vadez, V.; Kholova, J.; Medina, S.; Kakkera, A.; Anderberg, H. Transpiration Efficiency: New Insights into an Old Story. J. Exp. Bot. 2014, 65 (21), 6141–6153. [CrossRef]
- Aroca, R.; Porcel, R.; Ruiz-Lozano, J. M. Regulation of Root Water Uptake under Abiotic Stress Conditions. J. Exp. Bot. 2012, 63 (1), 43–57. [CrossRef]
- Morita, S.; Okamoto, M.; Abe, J.; Yamagishi, J. Bleeding Rate of Field-Grown Maize with Reference to Root System Development. Jpn. J. Crop Sci. 2000, 69 (1), 80–85. [CrossRef]
- He, Z.; Xu, C.; Liu, B.; Yao, B.; Wang, H.; Chen, Z. Y.; Li, D. Y.; Bai, Z. Y.; Zhang, Z. A. Relationship between Photosynthesis, Bleeding-Sap Mass, and Bleeding Components in Maize Hybrids and Corresponding Parents in Northern China. Photosynthetica 2019, 57 (2), 698–704. [CrossRef]
- Ansari, T. H.; Yamamoto, Y.; Yoshida, T.; Sakagami, K.; Miyazaki, A. Relation between Bleeding Rate during Panicle Formation Stage and Sink Size in Rice Plant. Soil Sci. Plant Nutr. 2004, 50 (1), 57–66. [CrossRef]
- Liu, C.; Wang, X.; Tu, B.; Li, Y.; Chen, H.; Zhang, Q.; Liu, X. Root K Affinity Drivers and Photosynthetic Characteristics in Response to Low Potassium Stress in K High-Efficiency Vegetable Soybean. Front. Plant Sci. 2021, 12, 732164. [CrossRef]
- Li, H.; Li, L.; Liu, N.; Chen, S.; Shao, L.; Sekiya, N.; Zhang, X. Root Efficiency and Water Use Regulation Relating to Rooting Depth of Winter Wheat. Agric. Water Manag. 2022, 269, 107710. [CrossRef]
- Evans, R. G.; Sadler, E. J. Methods and Technologies to Improve Efficiency of Water Use: INCREASING WATER USE EFFICIENCIES. Water Resour. Res. 2008, 44 (7). [CrossRef]
- Wang, S.; Li, L.; Zhou, D. Root Morphological Responses to Population Density Vary with Soil Conditions and Growth Stages: The Complexity of Density Effects. Ecol. Evol. 2021, 11 (15), 10590–10599. [CrossRef]
- Foxx, A. J.; Fort, F. Root and Shoot Competition Lead to Contrasting Competitive Outcomes under Water Stress: A Systematic Review and Meta-Analysis. PLOS ONE 2019, 14 (12), e0220674. [CrossRef]
- Nadeem, M.; Wu, J.; Ghaffari, H.; Kedir, A. J.; Saleem, S.; Mollier, A.; Singh, J.; Cheema, M. Understanding the Adaptive Mechanisms of Plants to Enhance Phosphorus Use Efficiency on Podzolic Soils in Boreal Agroecosystems. Front. Plant Sci. 2022, 13, 804058. [CrossRef]
- Zhang, W.; Dong, A.; Liu, F.; Niu, W.; Siddique, K. H. M. Effect of Film Mulching on Crop Yield and Water Use Efficiency in Drip Irrigation Systems: A Meta-Analysis. Soil Tillage Res. 2022, 221, 105392. [CrossRef]
Figure 1.
Average daily air temperature and sunshine hour during the rice growing seasons of 2020(A) and 2021(B).
Figure 1.
Average daily air temperature and sunshine hour during the rice growing seasons of 2020(A) and 2021(B).
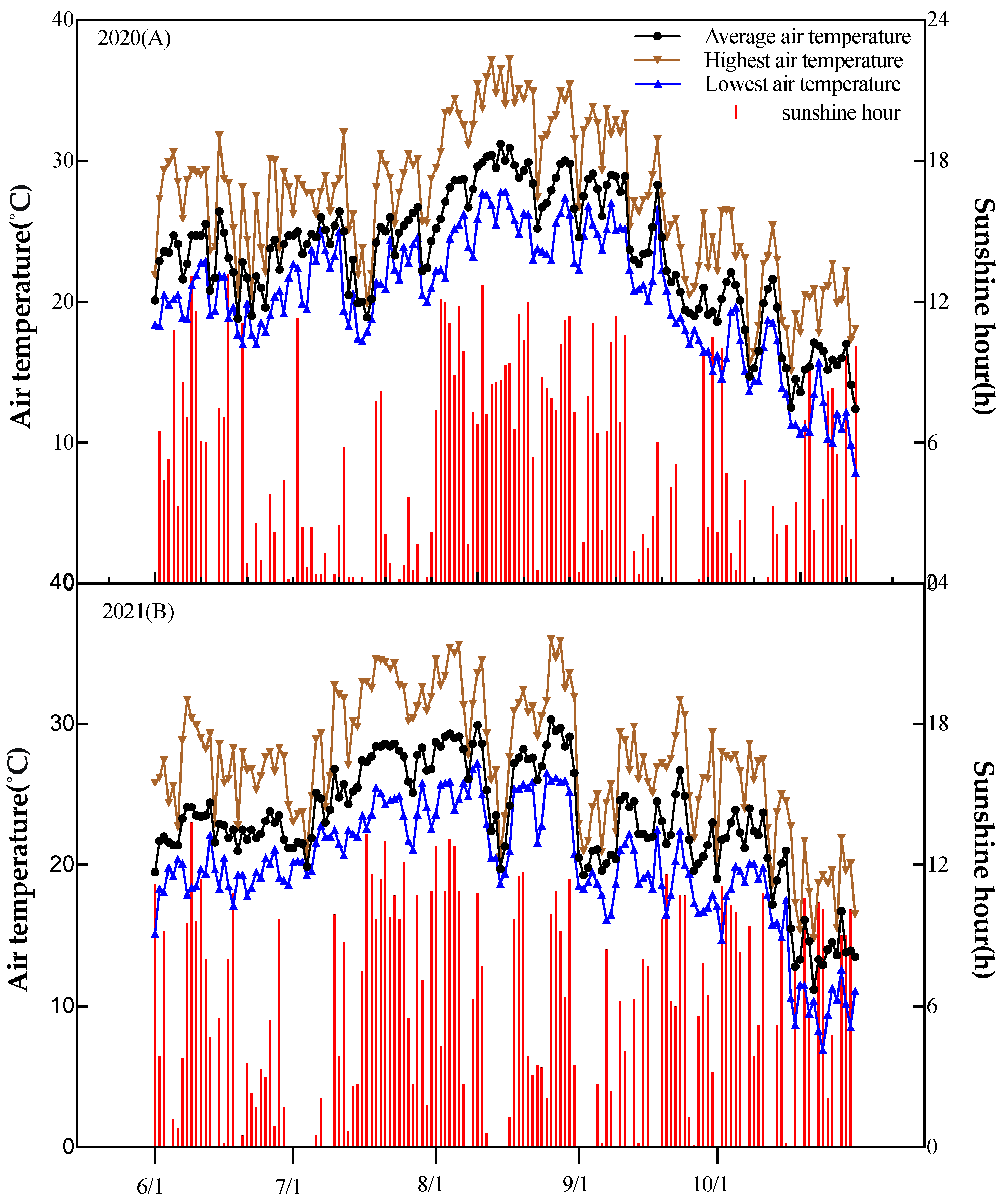
Figure 2.
Rainfall, irrigation, and volumetric soil moisture content under drip irrigation system with plastic-film mulch during the rice growing seasons of 2020(A) and 2021(B).
Figure 2.
Rainfall, irrigation, and volumetric soil moisture content under drip irrigation system with plastic-film mulch during the rice growing seasons of 2020(A) and 2021(B).
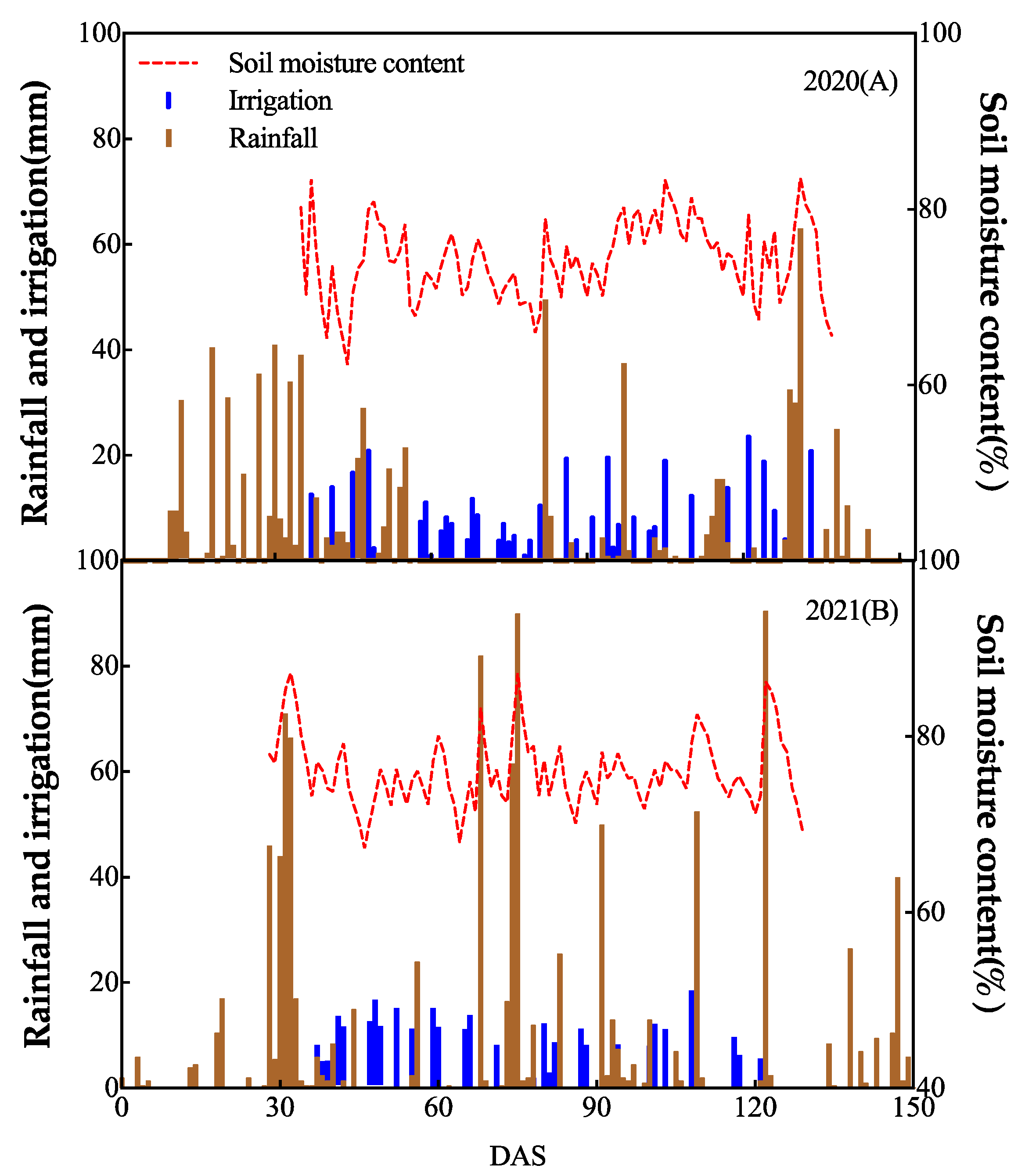
Figure 3.
Shoot dry matter (a and b), root dry matter (c and d), and root-to-shoot ratio (e and f) of four rice varieties under CF and DI in 2021. Data indicate the mean of three replications. Open symbols and solid symbols indicate CF and DI, respectively.
Figure 3.
Shoot dry matter (a and b), root dry matter (c and d), and root-to-shoot ratio (e and f) of four rice varieties under CF and DI in 2021. Data indicate the mean of three replications. Open symbols and solid symbols indicate CF and DI, respectively.
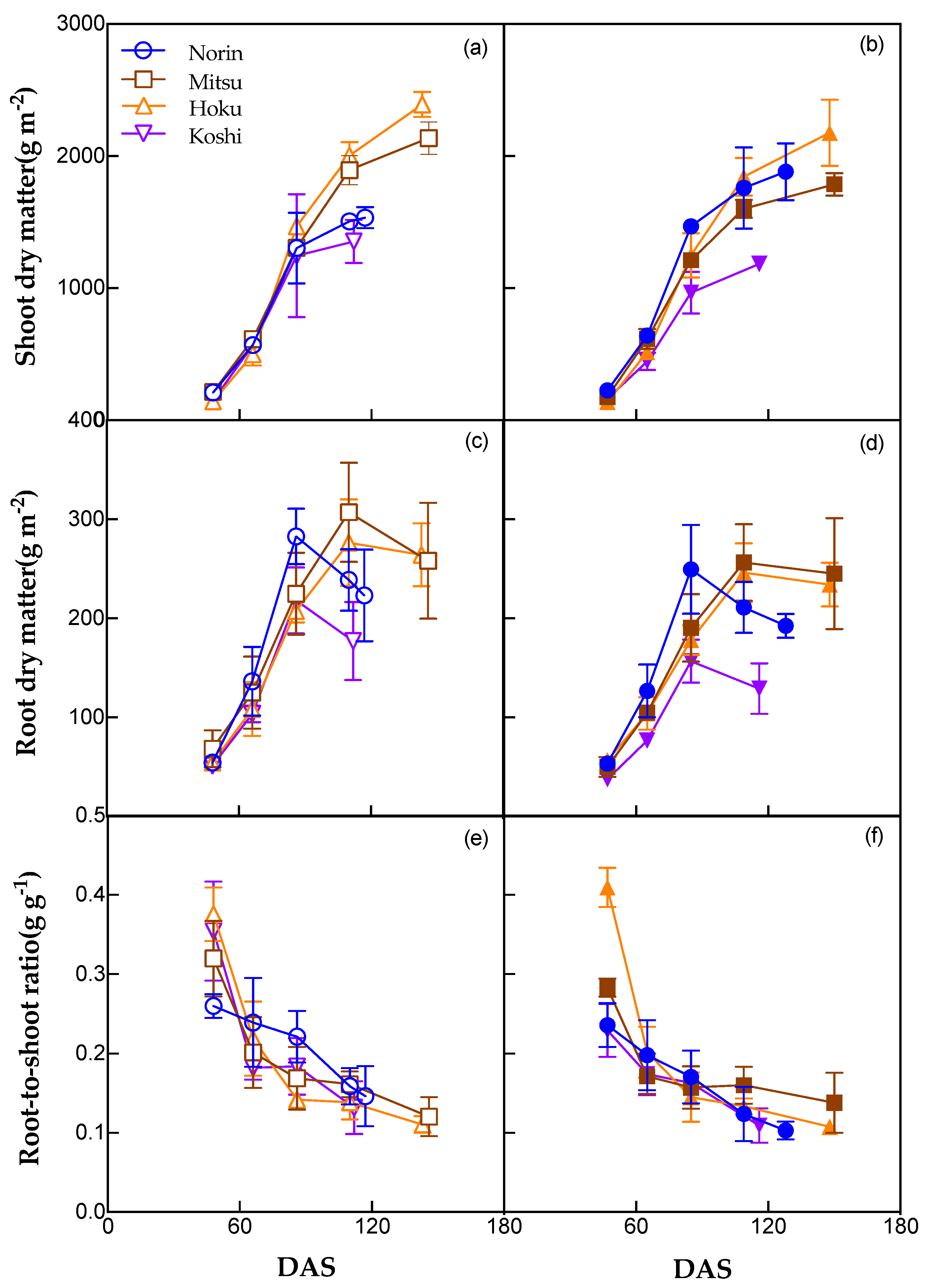
Figure 4.
Root length, and root weight at panicle-initiation stage(a)(b) and heading stage(c)(d) as affected by varieties and irrigation regimes at the soil depths of 0-15, and 15-30cm, respectively. Different capital letters beside bars show significant differences (P<0.05) between CF and DI, and different lowercase letters beside bars show significant differences (P<0.05) among varieties for each irrigation system by Duncan’s multiple range test.
Figure 4.
Root length, and root weight at panicle-initiation stage(a)(b) and heading stage(c)(d) as affected by varieties and irrigation regimes at the soil depths of 0-15, and 15-30cm, respectively. Different capital letters beside bars show significant differences (P<0.05) between CF and DI, and different lowercase letters beside bars show significant differences (P<0.05) among varieties for each irrigation system by Duncan’s multiple range test.
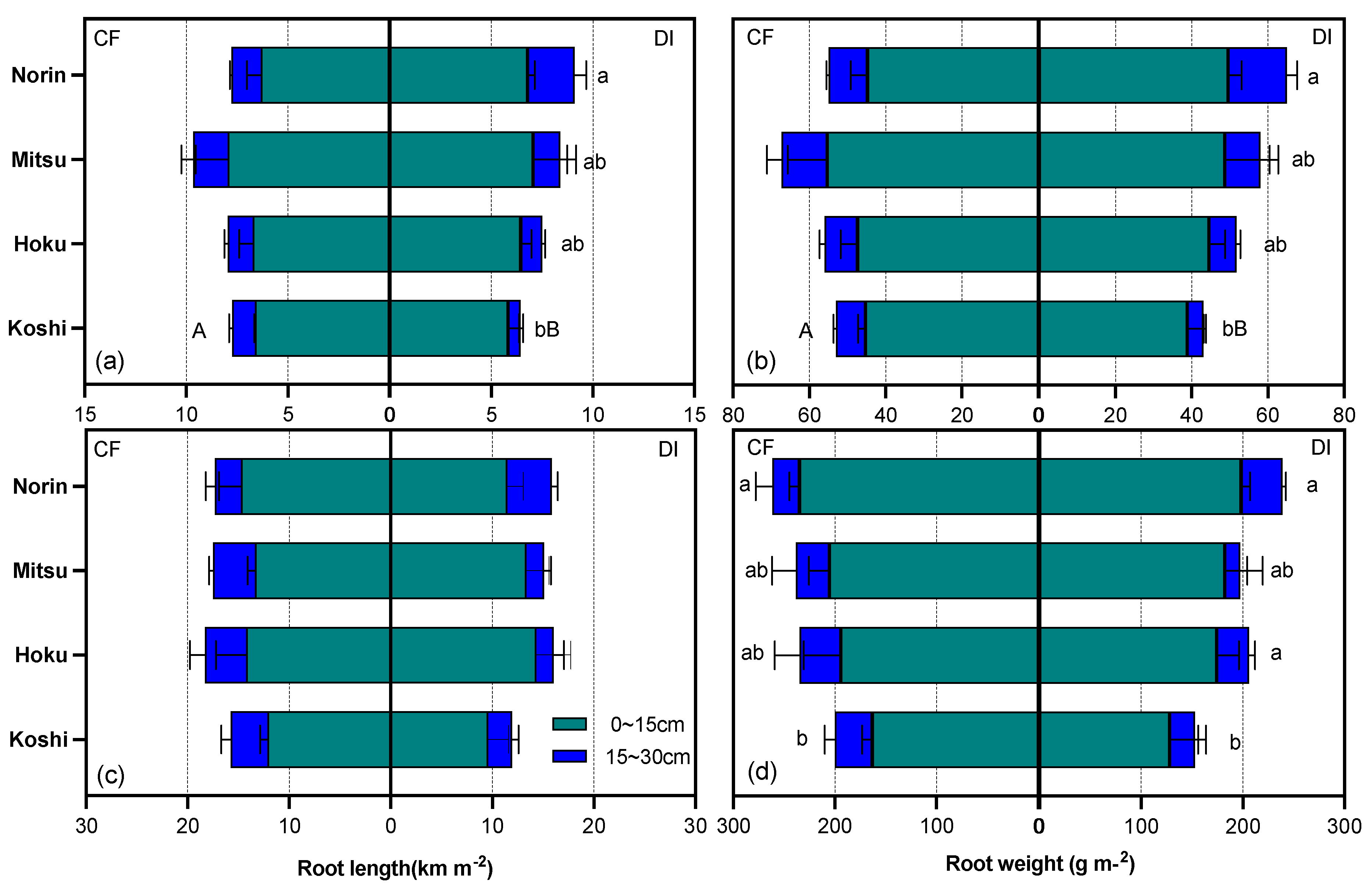
Figure 5.
Deep root ratio at panicle-initiation stage(a) and heading stage(b) as affected by irrigation system and rice variety. Different capital letters beside bars show significant differences(P<0.05) between CF and DI, and different lowercase letters beside bars show significant differences(P<0.05) among varieties for each irrigation system by Duncan’s multiple range test. Vertical bars represent the standard error of the mean.
Figure 5.
Deep root ratio at panicle-initiation stage(a) and heading stage(b) as affected by irrigation system and rice variety. Different capital letters beside bars show significant differences(P<0.05) between CF and DI, and different lowercase letters beside bars show significant differences(P<0.05) among varieties for each irrigation system by Duncan’s multiple range test. Vertical bars represent the standard error of the mean.
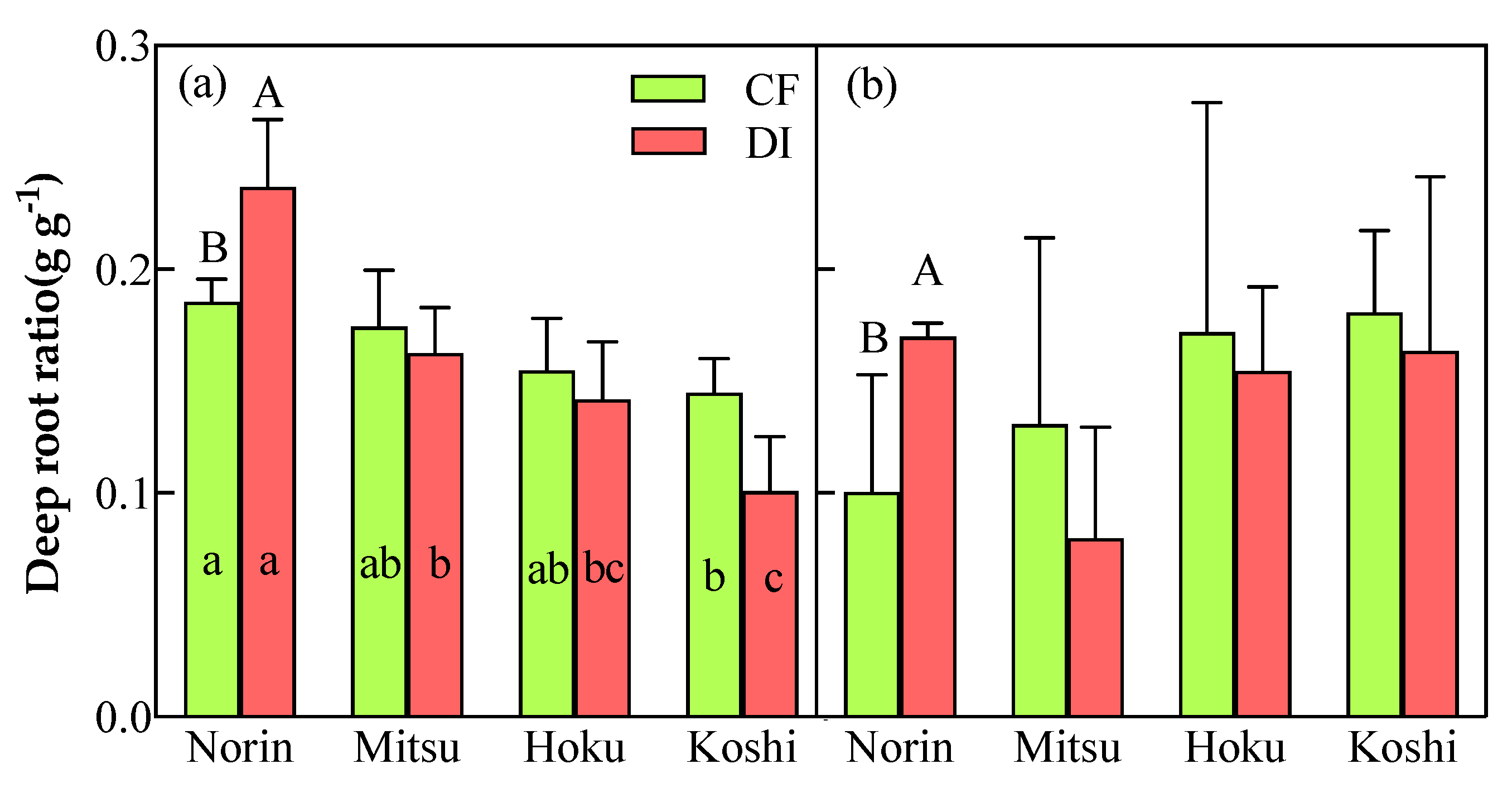
Figure 6.
Root bleeding rate at the panicle-initiation stage as affected by the irrigation system and rice variety in 2020(a) and 2021(b), respectively. Different capital letters beside bars show significant differences(P<0.05) between CF and DI, and different lowercase letters beside bars show significant differences(P<0.05) among varieties for each irrigation system by Duncan’s multiple range test. Vertical bars represent the standard error of the mean.
Figure 6.
Root bleeding rate at the panicle-initiation stage as affected by the irrigation system and rice variety in 2020(a) and 2021(b), respectively. Different capital letters beside bars show significant differences(P<0.05) between CF and DI, and different lowercase letters beside bars show significant differences(P<0.05) among varieties for each irrigation system by Duncan’s multiple range test. Vertical bars represent the standard error of the mean.
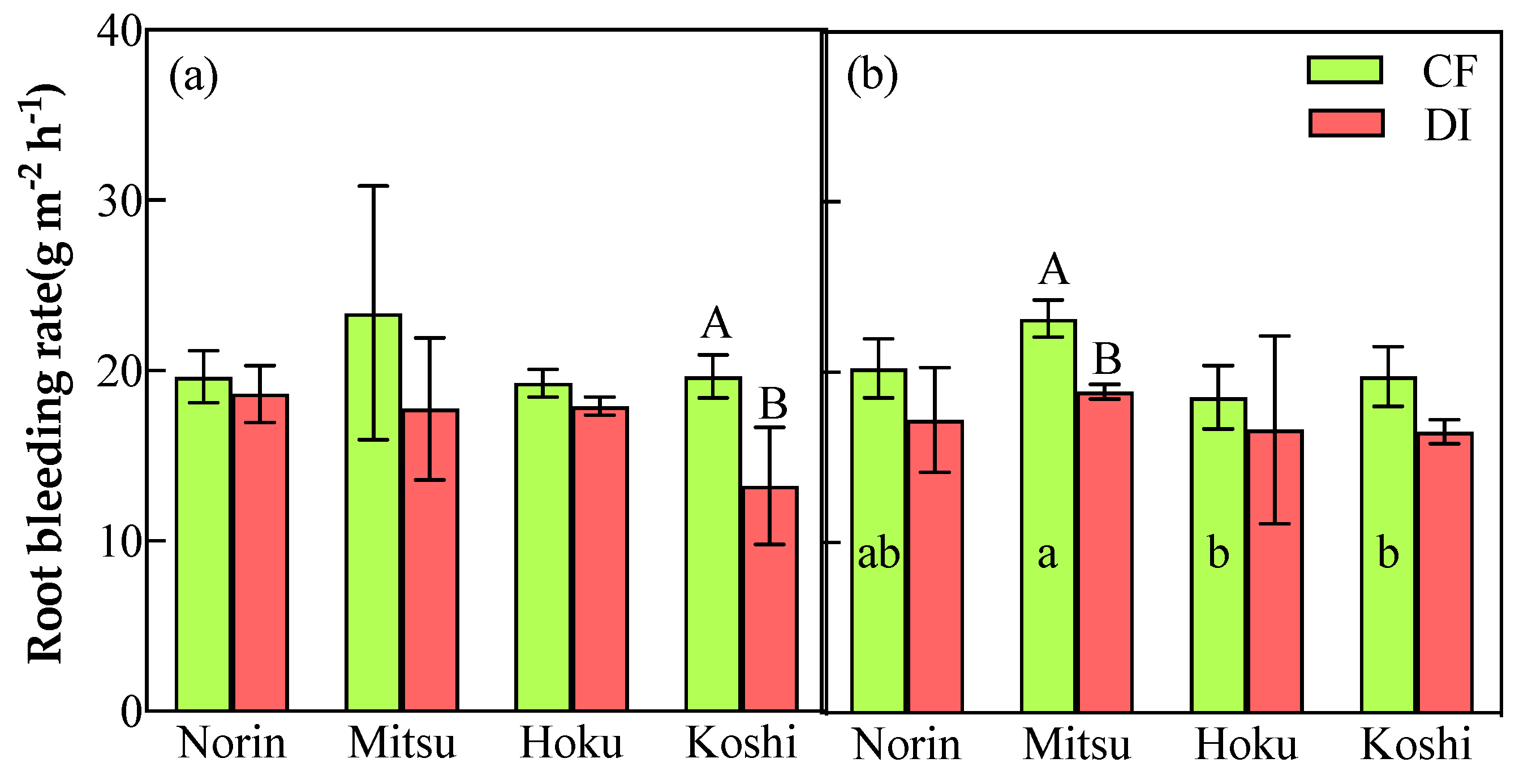
Figure 7.
Relationship between deep root ratio and transpiration rate(a), grain yield(b), and water productivity(c) in 2021. Open and solid symbols indicate CF and DI, respectively. * means significant differences at P<0.05. n=24.
Figure 7.
Relationship between deep root ratio and transpiration rate(a), grain yield(b), and water productivity(c) in 2021. Open and solid symbols indicate CF and DI, respectively. * means significant differences at P<0.05. n=24.
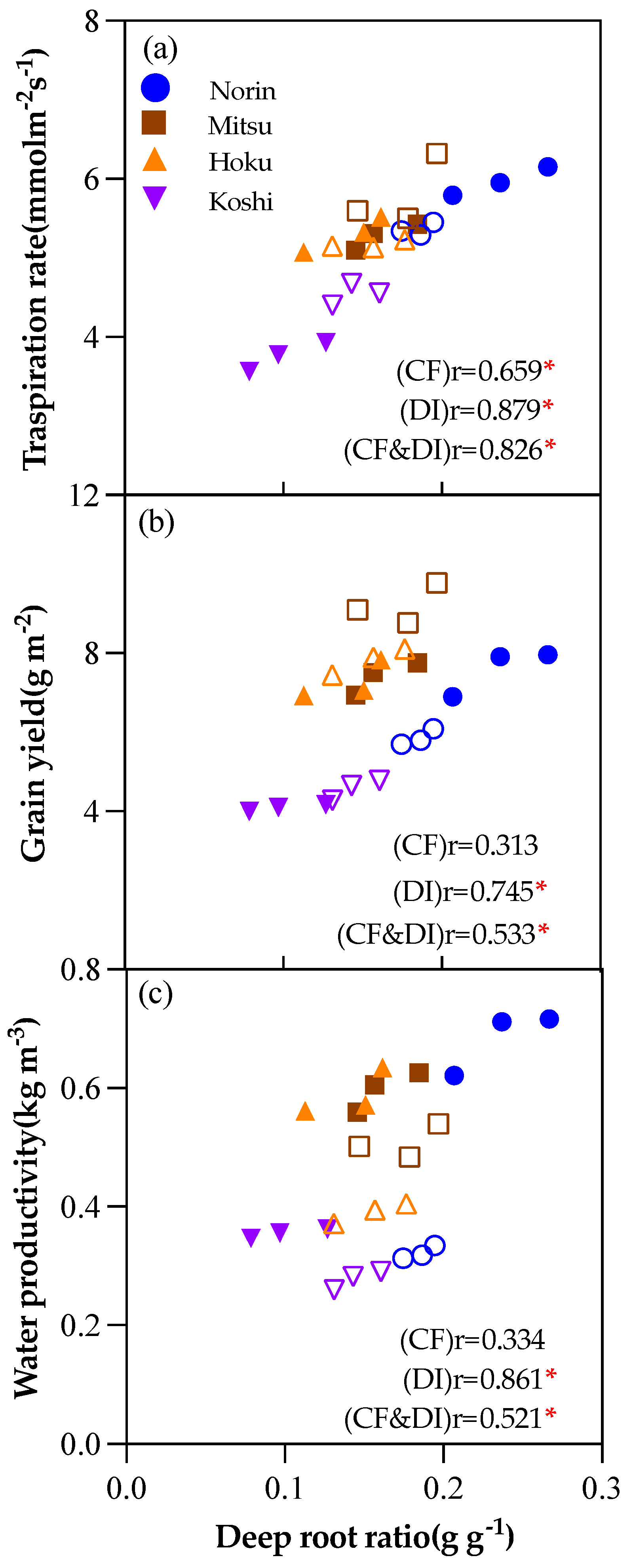
Figure 8.
Pearson’s correlation coefficient value of yield, water productivity (Wp), CO2 assimilation rate (An), stomatal conductance(gs), deep root ratio (DR), root length (RL), root weight (RW), and root bleeding rate (BR) under CF (a), DI (b) and CF&DI (c), respectively.
Figure 8.
Pearson’s correlation coefficient value of yield, water productivity (Wp), CO2 assimilation rate (An), stomatal conductance(gs), deep root ratio (DR), root length (RL), root weight (RW), and root bleeding rate (BR) under CF (a), DI (b) and CF&DI (c), respectively.
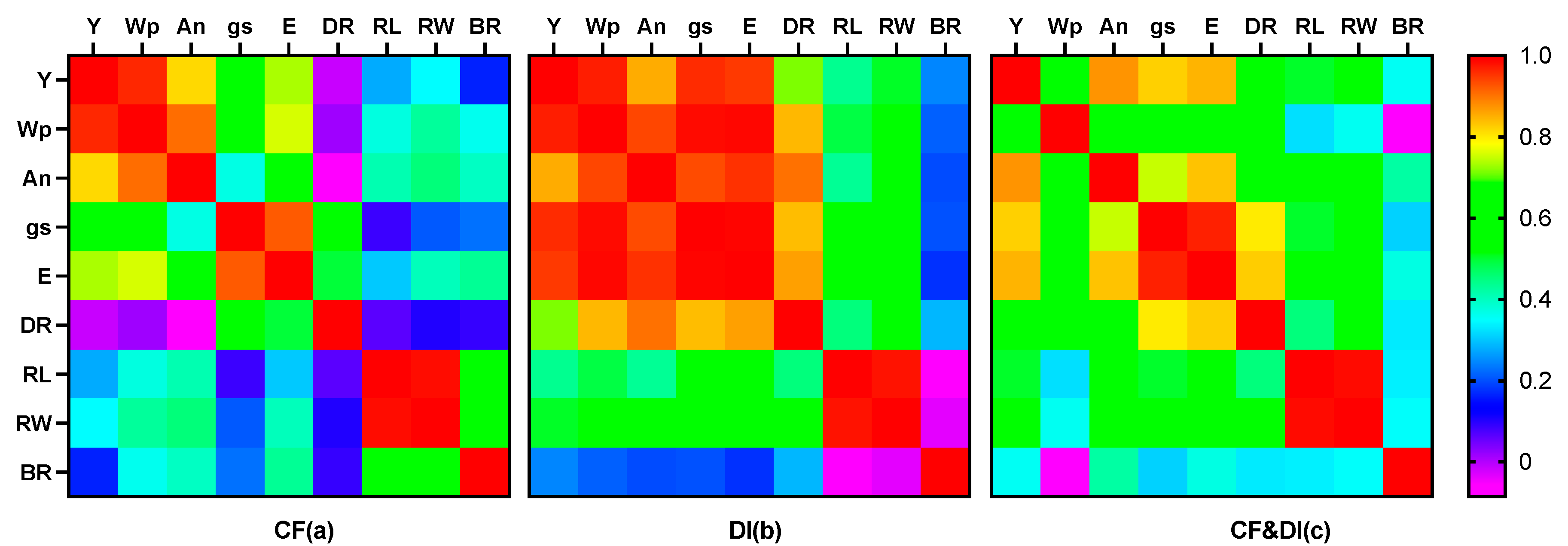

Table 1.
Photosynthetic-associated parameters of four rice cultivars at the early grain-filling stage under CF and DI in 2021.
Table 1.
Photosynthetic-associated parameters of four rice cultivars at the early grain-filling stage under CF and DI in 2021.
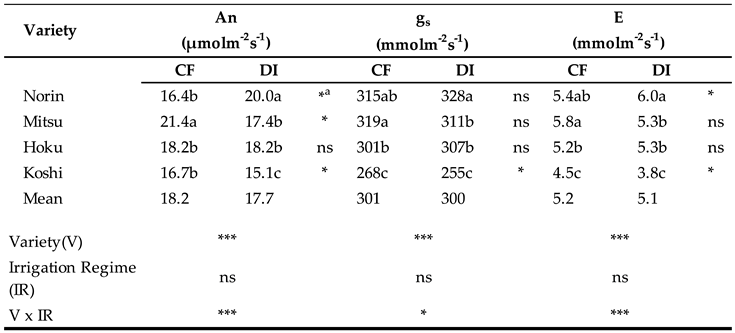
Noted: Data indicate the mean of three replications. Values within a column for each irrigation system followed by different letters are significantly different at P<0.05 by Duncan's multiple range test. a indicates the significance within some cultivars under CF and DI by T-test. *, ** and *** are significant differences at P<0.05, P<0.01 and P<0.001 respectively; n.s means non-significant by ANOVA.
Table 2.
Water productivity, harvest index, yield, and yield component of four rice varieties under CF and DI in 2020 and 2021.
Table 2.
Water productivity, harvest index, yield, and yield component of four rice varieties under CF and DI in 2020 and 2021.
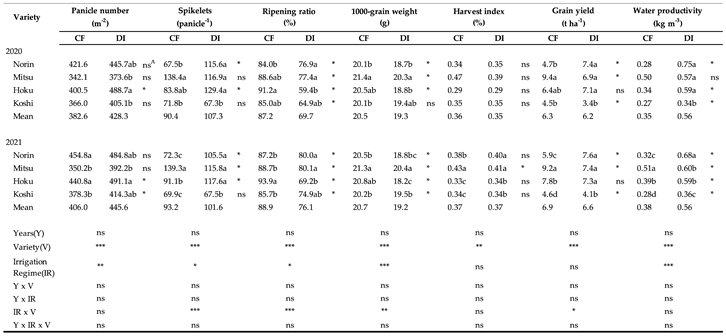
Noted: Data indicate the mean of three replications. Values within a column for each irrigation system followed by different letters are significantly different at P<0.05 by Duncan's multiple range test. A indicates the significance within some cultivars under CF and DI by T-test. *, ** and *** are significant differences at P<0.05, P<0.01 and P<0.001 respectively; n.s means non-significant by ANOVA.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



