
Submitted:
25 September 2023
Posted:
26 September 2023
You are already at the latest version
Abstract
The clinical relationship between diabetes and inflammation is well-established. Various branches of science independently have confirmed that disrupting oxidant-antioxidant equilibrium and elevated lipid peroxidation could be a potential mechanism for chronic kidney disease associated with Type 2 diabetes mellitus (T2DM). Under diabetic condition, hyperglycemia, especially inflammation, and increased reactive oxygen species generation are bidirectionally associated. Inflammation, oxidative stress and tissue damage are believed to have a role in the development of diabetes. Although the exact mechanism underlying the oxidative stress and its impact on diabetes progression remains uncertain, the hyperglycemia-inflammation-oxidative stress interaction clearly plays a significant role in onset and progression of vascular disease, kidney disease, hepatic injury, pancreas damage and, therefore holds promise as a therapeutic target. Evidence strongly indicates that the use of multiple antidiabetic medications therapy fails to achieve the normal range for glycated hemoglobin target, signifying treatment resistant diabetes. Antioxidants with polyphenols are considered useful as adjuvant therapy for their potential anti-inflammatory effect and antioxidant activity. We aimed to analyze the current major points reported in preclinical in vivo and clinical studies of antioxidants in the prevention or treatment of inflammation in T2DM. Then, we will share our speculative vision of the future diabetes clinical trials.
Keywords:
Oxidative stress
; Reactive oxygen and nitrogen species
; Redox state
; Antioxidant response
; diabetes
; Metabolic syndrome
; Inflammation
1. Introduction
The Type 2 diabetes mellitus (T2DM) is a common hormonal disorder in the adult population presenting in 11.3% of the US population and undiagnosed approximately 23% of adults according to the Center for Disease Control (CDC) National Diabetes Statistics Report for 2020 cases of diabetes [47]. T2DM is caused by insulin deficiency or insulin resistance. While the incident of T2DM in youth is increasing [48], it is rare in the pediatric population. Although most of the identified prediabetes are incidental findings without the need for intervention, some may present as clinically significant because they have a higher than normal blood sugar level or cause symptoms from medication or co-morbidities including hypertension, hyperlipidemia, obesity, or nephropathy.
The etiology of T2DM is diverse, and appears to be due to genetic [49,50] cause and environmental factors [51]. In certain cases, however, germline including insulin-association with cancer [52,53] or somatic genetic defects [54] are associated with the development of T2DM. Risk alleles at numerous loci such as MTNR1B, CGKAL1, and IGF2BP2 have been investigated as being potentially involved in the pathogenesis, presentation of T2DM and beta-cell function [55]. T2DM appear to be linked to dyslipidemia, atherosclerotic heart disease and cancer [56]. Additional environmental or stochastic events in T2DM, like DNA-methylation, maternal environment [57], the impact of intrauterine [58]. early infancy, and the composition and metabolic function of the gut microbiota in obese individuals [59] have also been investigated. Thus, the pathogenesis that underlies T2DM is complex and multifactorial. One of the most important components is the role of glucose autoactivation following persistent elevated circulating glucose, resulting in prooxidant production.
Oxidative stress is caused by a perturbation of prooxidant [reactive oxygen species (ROS)] and antioxidant microecosystem that favors excess production of prooxidant relative to antioxidant defense [60]. The ROS are formed by multiple overlapping and interacting mechanisms that highlights its biological complexity and its effects on individual’s genetic background. These enzymatic and non-enzymatic pathways include mainly oxidative phosphorylation, the plasma membrane proteins such as NADPH oxidases, lipid metabolism within the peroxisomes, and cyclooxygenases. The abnormality of these two pathways leads to oxidative stress, which accelerates the development of diabetes complications, both microvascular and cardiovascular (Figure 1). Readers are invited to recent reviews on this issue [61,62].
2. Diabetes
Diabetes is the most common chronic disease of childhood [63,64,65] and adulthood [66] and the most common type of diabetes is T2DM. While its exact cause is unknown, T2DM patients have elevated blood glucose levels, dyslipidemia, and impaired insulin receptor function [67]. There is a positive correlation between the progressive nature of T2DM and increased risk of the major cardiovascular diseases, including coronary artery disease, ischemic heart disease, stroke, and peripheral artery disease. Diabetic patients with silent cardiac left ventricular hypertrophy or renal disease are at a higher risk of developing both microvascular and macrovascular diseases [68], leading to organ and tissue damage. Diabetes-induced vascular aberration include anatomic, structural, and functional alteration resulting in endothelial dysfunction, reduced vascular compliance and atherosclerosis [69].
As mentioned earlier, the cause of T2DM is unknown. But the diagnosis is made on the basis of the spectrum of clinical manifestations such as impaired regulation of hepatic glucose production, incremental decline in β-cell function, destruction of pancreatic β-cells in a small subset of type 2 diabetic patients, insulin resistance (also known as impaired insulin sensitivity), genetic mutations, dyslipidemia, or hormonal diseases. Blood glucose levels change due to diet, medications, and diabetes, a chronic heterogenous metabolic disorder with complex pathogenesis. The above leading clinical features are initially characterized by monitoring blood glucose to identify glucose fluctuation patterns in response to diet. A fasting plasma glucose of 126 mg/dL or more on more than one occasion is found to be associated with increased risk of microvascular complications [70]. Elevated glucose undergoes autoactivation, which promotes accumulation of advanced glycation end (AGE) products (Figure 2). Formed AGE products generate free radicals including peroxides, superoxide, hydroxyl radical, and singlet oxygen as described in the succeeding sections. Notably, both diabetic microvascular and macrovascular complications apparently have similar etiologic characteristics.
Insulin regulates the blood glucose levels at several key peripheral organs, highlighting a complex process. Insulin plays a major role in energy metabolism by promoting glucose uptake via glucose transporter 4 (GLUT4) translocation in skeletal muscle and adipose tissue, increasing hepatic glycogen storage, and enhancing lipogenesis. Evidence suggests that elevated oxidative stress blocks not only insulin secretion but also GLUT4 translocation, which leads to glucose toxicity. Advanced glycation end product-induced free radical production and elevated oxidative stress-induced reduction in insulin secretion and GLUT4 translocation could cause a futile cycle because it involves continuous accumulation of glucose in the blood called hyperglycemia (Figure 3).
Circulating levels of free fatty acids are increased in individuals exhibiting obesity, non-alcoholic fatty liver disease, insulin resistance, or T2DM. While both growth hormone and the sympathetic nervous system promote circulating levels of free acid [71], insulin reduces free fatty acid levels through suppressing lipolysis and enhancing free fatty acid clearance [72]. Chronic elevated plasma free fatty acid levels may trigger the initiation of diabetic vascular complications via increased production of oxygen-derived free radicals and exacerbation of dyslipidemia.
Oxidative stress plays an important role in the development of diabetes. Under physiological condition, oxidants are neutralized by enzymatic and nonenzymatic antioxidants. First line defense enzymatic antioxidants superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx) counteract against free radicals to protect the integrity of biomolecules and tissues [73]. This highlights the importance of these enzymes’ expression in neutralizing excessive oxygen-derived free radicals and maintaining ROS homeostasis. While elevated levels of fatty acids cause an increase in oxidative stress through NADPH oxidative activation, the expression of the antioxidant enzymes is reduced. In addition, excessive free fatty acid liberation alters endothelium-dependent vasodilation by affecting the synthesis or degradation of nitric oxide in diabetes [74]. Thus, diabetic dyslipidemia contributes to reduced antioxidant enzyme expression and reduced nitric oxide bioavailability.
In summary, diabetes is basically a metabolic disorder characterized by hyperglycemia, elevated free fatty acids, and insulin resistance. Each of these major hallmarks of T2DM provokes molecular mechanisms that contribute to microvascular or macrovascular dysfunction. These mechanisms include decreased bioavailability of nitric oxide, increased oxidative stress, decreased antioxidants, and activation of receptors for AGEs. The prevailing model for T2DM states that diabetes is a major risk factor for cardiovascular diseases (CVD) and is responsible for widespread morbidity and mortality. A large number of physiologic and genetic studies support this model and the central role of vascular endothelial cell metabolism [75,76,77,78], which affects the metabolism of other cell types contributing to the pathogenesis of diabetes.
ROS are formed as natural byproducts of cellular metabolism [79] and are tightly controlled through well-coordinated cellular mechanisms. Increased ROS formation can range from low to severely high levels. At low levels, ROS act as regulators of certain signaling pathways [80] in both cell biology and physiology. At high levels, ROS cause an array of diseases, including cardiovascular, neurodegenerative, metabolic disorders, cancer, inflammation, and aging (Figure 1). ROS are involved in the degradation of pathogens and serve as intracellular signaling molecules in steady-state, indicating their protective roles in infection and normal biological processes. However, their prolonged effects result in abnormal cellular physiology, which exacerbates ROS production leading to progressive oxidative damage and subsequent initiation of cell death. Moreover, oxidative stress-induced cell death appears to be the most proximal event of diabetes, leading to diabetic micro- and macro- angiopathic complications. Diverse components of the oxidative stress microenvironment affect cell metabolism and cell-cell homeostasis [62]. Furthermore, damage from these stressors can include mutations in mitochondrial DNA (mDNA) in patients with diabetes mellitus [81], somatic point mutations in age-related diseases [82], and immune-related somatic point mutation genes in diabetes [83].
The literature overflows with studies involving antioxidants. The majority of scientists in the field of experimental therapeutics for diabetes have observed many preclinical antioxidants that are claimed to be potent and selective, is often not the case due to off-target effects in general. In rare cases, the effects of antioxidants on cell function is obscure, indicating a false-positive association. In some cases, the antioxidant activity of crude extract on a mechanism involved in the disease process that is unelucidated. Since a close link exists among T2DM, CVD, and oxidant-induced injury, this review focuses on the recent research done on the effects of antioxidants on the classical biomarkers of the diabetes and the components of metabolic syndrome, such as insulin resistance, atherogenic dyslipidemia, hypertension, and proinflammatory and prothrombotic state, as well as on the current biomarkers of oxidative stress, which may be useful for the treatment of diabetes.
2.1. Chronic inflammation as a cause of diabetes
The role of AGE/RAGE signaling in diabetes and their influence on proinflammatory signaling pathways in hyperglycemic condition have been elaborated in details elsewhere [84,85] (Figure 3). C-reactive protein (CRP) and IL-6 are significantly elevated in patients with diabetes, hypertension, and the metabolic syndrome. Those with diabetes have increased systemic levels of CRP, a protein in the blood. IL-6, a protein, is produced by numerous cells and is elevated with the severity of diabetes. Although tumor necrosis factor alone induces the induction of CRP expression [86], the release of IL-6 in response to tissue damage is the main activator of CRP expression, which occurs in the hepatocytes. Notably, IL-1 synergistically enhances the effects of IL-6-induced CRP expression [86]. While IL-6 promotes differentiation of B-cells, it is capable of promoting cell growth in certain cells and inhibiting growth in others. This indicates that cytokines, particularly IL-6 have multiple roles to play within the body under both physiological and pathophysiological conditions (Figure 3). However, it is still unclear whether IL-6 is causing or contributing to diabetes. Both IL-6 and CRP independently predict the risk of T2DM. However, a combined elevation levels of IL-6 and IL-1β, a cytokine, in the blood predict increased risk to develop T2DM [87]. Although the precise mechanism by which the proinflammatory state emerges in patients with diabetes remains to be determined, persistent hyperglycemia activates NF-κB that triggers expression of various cytokines [88]. NF-kB not only has prooxidant role by inducing the expression of gene such as NADPH oxidase NOX2 gp91phox [89], but it also has numerous anti-oxidant targets, such as manganese superoxide dismutase [90] and NF-kB – induced ferritin heavy chain upregulation via preventing iron-mediated generation of high reactive hydroxyl radicals from H2O2 [91,92]. The activation of NF-κB is mediated via ROS production [88]. Cytokines also activate NF-κB and promote the recruitment of monocytes producing M1 and M2 that cause β-cell destruction and insulin resistance [84]. The role of inflammation in diabetes pathophysiology has been reviewed in detailed [93].
Regardless of the triggers, low-grade inflammation and vascular diseases are believed to be principal causes of disability and premature deaths in diabetes mellitus. Dendritic cells are critical regulators of both immune and inflammatory responses. Dendritic cells sense cellular debris and modified metabolites, which are found in diabetes [94,95]. Once again, the double-edged effect of ROS raises significant concerns. Dendritic NOX2 produces low levels of ROS, which are important in promoting dendritic cell maturation and cross-presentation [96]. Damaged tissues containing dysfunctional mitochondria produce non-physiological levels of ROS. Patients with T2DM exhibit increased ROS production [77], that results in microvascular and macrovascular complications. Evidence indicates that elevated ROS inhibit cross-presentation after entering dendritic cells [97] (Figure 3). Inadequate function of dendritic cells in diabetes may feedback ROS foster generation of cytokines which can contribute to enhanced generation of ROS. Like other cells, dendritic cells are vulnerable under increased ROS level.
Diabetes and its clinical manifestations are characterized by chronically elevated levels of glucose and lipid, which leads to glucolipotoxic conditions. Of note, glucolipotoxicity alters mitochondrial function, and in particular electron transport chain activity. Impaired electron transport chain activity leads to reduced and inefficient fatty acid β-oxidation [98], and promotes an increase in ROS production triggering inflammation in highly metabolic organs including kidneys, liver, heart, and insulin secreting β-cells (Figure 3). The major regulators of fatty acid oxidation in these tissues with high oxidative rates are peroxisome proliferator-activated receptor a (PPARa) [99], a regulator of intra- and extracellular lipid metabolism, and AMP-activated kinase (AMPK) , a sensor of cellular energy change [100]. Since both PPAR and AMPK are linked to metabolic disorders, modulation of these two molecules provide therapeutic targets for diseases such as diabetes, dyslipidemia, and inflammation.
The current therapeutic approaches for the treatment of metabolic disease and the potential of immune-modulatory approaches has been reviewed in detailed elsewhere [101]. Briefly, the oxidation of LDL particles in the endothelial wall of arteries instigates monocytes infiltration [102,103]. Monocytes then are differentiated into macrophages, which accumulate oxidized lipids to form foam cells. Remarkably, oxidative stress augments intimal LDL retention and accumulation [103]. Formed foam cells stimulate macrophage proliferation and promote the recruitment of T-lymphocytes, which induce smooth muscle proliferation in the arterial walls (Figure 3). The sustained inflammatory response continues to recruit mononuclear leukocytes, both monocytes and lymphocytes. The overactive immune system and inflammatory processes [104] are driving force in a wide variety of vascular physiological mechanisms such as endothelial activation, the loss of the endothelial monolayer due to thrombus formation and neointimal thickening[105], increased oxidative stress [103], reduced endothelial trophic effect, and disrupted blood vessel function [103,106]. It has become apparent that oxidative stress and inflammation are closely linked events, and persistent oxidative stress and chronic inflammation are now known to play predominant roles in diabetes mellitus and atherosclerosis.
2.2. Understanding the relationship between hypertension and type 2 diabetes mellitus
Statistics from the Centers for Disease Control and Prevention (CDC) and National Health and Nutritional Examination Survey (NHANES) database show that majority of individuals with diabetes have hypertension [107]. Dysregulation of renin-angiotensin-aldosterone system not only has been implicated in the pathophysiology of hypertension, but it also favors T2DM [108] (Figure 3). The inhibitors of the renin-angiotensin-aldosterone system prevent diabetes in patients with hypertension. Diabetes-induced dyslipidemia and hypertension play a major role in the initiation and progression of macrovascular disease. There is substantial overlap between diabetes and hypertension in disease mechanisms, such as upregulation of renin-angiotensin-aldosterone system and inflammation [109,110]. Should we consider diabetes and hypertension as chronic inflammatory diseases?
The dogma suggests that inflammation is a symptom of many chronic diseases, such as diabetes, hypertension, arthritis, or Alzheimer’s disease. However, systemic reviews and meta-analysis indicate unwaveringly that chronic inflammation increases the risk of various age-related diseases, such as diabetes mellitus, hypertension, kidney disease, rheumatoid arthritis [111,112,113,114].
Although it is not well clear what causes age-associated chronic inflammation, it appears that cellular senescence is a major endogenous risk factor causing chronic inflammation. Senescent cells are characterized by an arrest of cell proliferation associated with changes in cell morphology, physiology, chromatin organization, which leads to the development of an unique phenotype, known as senescence-associated secretory phenotype [115]. An endogenous contributor of this phenotype is oxidative stress, which will be reviewed in the following section. Senescent cells are capable of secreting pro-inflammatory cytokines, chemokines, and other proinflammatory molecules [116]. This production leads to enhanced infiltration of immune cells and activation of other inflammatory systems, such as nitric oxide and prostaglandin. In diabetes, hyperglycemic spikes affect cytokine concentrations [117].
An increasing body of evidence suggests that diabetes-induced ROS production or excessive generation of ROS [118] independent of diabetes has a significant role in the pathogenesis of hypertension (Figure 1) [110]. Sustained elevated ROS levels contribute to both hypertension and diabetes. Hypertensive patients exhibit a substantial increase in plasma H2O2 levels [119]. Studies revealed presence of decreased endogenous antioxidant enzyme activity in peripheral mononuclear cells in hypertensive patients [120] as well as in animal models of hypertension including angiotensin II-induced hypertension [121] and nephron-vascular hypertension [122].
Nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOXs) and mitochondria produce ROS under physiological and pathological conditions, as described in the following section. Angiotensin II, a vasoactive peptide, not only control blood pressure, but it also is implicated in vascular ROS production. It stimulates NADH/NADPH oxidase, and activates kinases including Rho/Rho kinase [123], protein kinase C (PKC) [124], and mitogen-activated protein kinase (MAPK) [125]. Thus, ROS-induced vasoconstriction contributes to the pathogenesis of hypertension. The interplay between ROS imbalance and hypertension has been described and reviewed in detail elsewhere [126].
Nitric oxide (NO), a major vasodilator, is catalyzed by nitric oxide synthase. Angiotensin II and NO have opposite effects to one another [127]. The bioavailability of NO is highly dependent on the redox status [128]. The two reactive nitogen species (RNS), namely peroxynitrite and S-nitrosoglutathione, are derived from the •NO radical. Like ROS, RNS is prone to uncontrolled overproduction under stressful conditions, leading to cellular nitro-oxidative damage. Generation of ROS takes place in different compartments than that of NO generation, suggesting compartmentalization in which these molecules are being tightly regulated.
In addition, other health conditions that have key role in the development of hypertension are kidney disease, glomerulonephritis and atherosclerosis. Remarkably, both diabetes mellitus and hypertension are responsible for chronic kidney disease [129] and adversely alter the lumen of small arteries and arterioles. Diabetic patients manifest accelerated premature vascular aging characterized by enhanced vascular smooth muscle contraction and increased vascular stiffness [130]. These structural and functional vascular changes contribute to the development of hypertension. These multiple interactive mechanisms perpetuate diabetes over time, may even cause a vicious cycle. Thus, effective management of diabetes should include a multifaceted approach to ensure adequate control of blood pressure, lipid with appropriate glycemic control [131], ROS production, RNS production, and inflammation.
3. The Landscape of ROS
ROS are constantly produced and inactivated by antioxidant enzymes such as catalase, superoxide dismutase, glutathione peroxidase in biological system. Oxidative stress has been defined as an imbalance between oxidants and antioxidants and more recently as a disruption of redox signaling and control [132]. It is generally accepted that oxidative stress has a fundamental role in vascular dysfunction [132]. In diabetic retinopathy, oxidative stress - induced decreased nitric oxide bioavailability contributes to endothelial cell dysfunction, basement membrane thickening, pericyte apoptosis, resulting in altered microvasculature [133]. Lipid peroxidation is one of the major mechanisms of oxidative damage [134]. There is considerable evidence that hyperglycemia represents the main cause of complications of DM, and oxidative stress resulting from increased generation of reactive oxygen species plays a crucial role in their pathogenesis [134]. The disruption of antioxidant equilibrium and elevated lipid peroxidation mechanism for vascular damage associated with T2DM has been studied extensively.
3.1. Sources of free radicals in the cells
Both reactive oxygen species (ROS) and reactive nitrogen species (RNS) are the free radicals [135] derived from both exogenous and cellular sources. The history, chemistry, and biochemistry of both ROS and RNS are reviewed in detail elsewhere [136]. ROS are reactive molecules that are derived from oxygen and, typically involved in a biological process as free radicals. These free radicals include superoxide (O2•-), hydroxyl radical (OH•), or non-radicals (H2O2). Like ROS, RNS are products of normal cellular metabolism. However, RNS are derived from nitrogen and can be free radicals (peroxynitrite (ONOO-) or nitric oxide (NO•)). While hydroxyl radical is the most reactive species that is responsible for the induction of cytotoxicity, both nitric oxide and hydrogen peroxide are less reactive. Overproduction of hydroxyl radical and peroxynitrite can lead to impaired normal physiological redox-regulated functions, and a subsequent unregulated cell signaling, followed by apoptosis via lipid peroxidation, thus disrupting the integrity of cell membrane [137] and altering lipoprotein structures.
In diabetes, although free radicals are mainly produced in mitochondria, they are produced by other cellular organs such as peroxisome and endoplasmic reticulum [138]. Deciphering the molecular mechanisms through which ROS directly interact with pivotal signaling molecules and the physiology of cellular ROS by mitochondria and their signaling molecules in cell proliferation and survival are of utmost importance to understand how elevated levels of ROS may result in irreparable cell damage. The ROS-modulating approaches have been used to lower excessive production of mitochondrial ROS, which is considered to be a central mechanism for the development of diabetes complications. Readers are invited to recent reviews on this issue [139,140].
3.1.1. Mitochondria
The biological significance of mitochondria with special emphasis on the ROS has been summarized [141]. Compared to other cellular organelles, mitochondrial functioning is determined by mitochondrial DNA and the genome in the nucleus. Mitochondria have now emerged as multifaceted centers responsible for the site of energy production, the formation of ROS, and notably as an intracellular sink for ROS [142]. Needless to say, the mechanism by which the mitochondria perform a sink function remains unresolved. Growing evidence indicates mitochondria-related mechanisms in pathologic process. Mitochondrial defects are also found in pathological studies of chronic diseases and major neurodegenerative diseases. The homeostasis of mitochondria is disrupted in several diseases, including atherosclerosis, renal diseases, autoimmune disorders, pancreatitis, diabetes, and chronic inflammation. In these diseases, dysfunction of mitochondrial complex I could potentially favors overproduction of ROS [143] beyond mitochondrial antioxidant capacity. The mitochondrial defects found in diabetes include fragmentation [144], fusion and fission [145], morphological changes [146], elevated mutation rates in mitochondrial DNA [147], changes in permeability of mitochondrial membranes [148], fluctuations in redox potential [149], buildup of mutant proteins [150], and impaired oxidative phosphorylation [151]. Mitochondrial dysfunction and oxidative stress has been implicated in pathogenesis of diabetes [144]. While autophagy is important for maintaining balanced cell life, dysregulated or reduced autophagy [152,153,154] has been linked to diabetes-induced abnormal fuel metabolism. Based on these findings, a series of therapeutic methods targeting mitochondrial proteins and processes have been developed from bench to bedside. Mitochondria perform a pivotal role in ROS metabolism. However, mitochondrial dysfunction is not considered as a hallmark of T2DM, diabetic nephropathy or diabetic neuropathy.
3.1.2. Peroxisome
Peroxisomes serve as a sensing mechanism to sense the accumulated cellular free fatty acids and activate signaling pathway and coupling of free fatty acid degradation with that of ROS [155]. While both mitochondria and peroxisomes are involved in hydrogen peroxide formation, hydrogen peroxide is produced by leaks of electrons from donor redox centers of the mitochondrial electron transport chain [156]. Unlike mitochondria, peroxisomes transfer electrons from various metabolites to the oxygen leading to the production of H2O2 within its respiratory pathway. Since this pathway is not coupled to oxidative phosphorylation to produce ATP, its free energy is released in the form of heat. In addition, peroxisomes are important source of O2•−, •OH, and NO• via various ROS-generating enzymes [157], which play a key role in signaling pathways responsible for regulating essential processes in the cells. These free radicals are produced in a variety of metabolic pathways such as fatty acid oxidation, nucleic acid and polyamine catabolism. Of note, the discovery of ROS-metabolizing enzymes in peroxisomes has been reported [157], highlighting the importance of peroxisomes in scavenging ROS in the cells.
As mentioned earlier, diabetes causes lipid accumulation. An array of isoforms of the peroxisome proliferator-activated receptors (PPAR-α, PPAR-γ, and PPAR-σ) play essential role in the regulation of lipid and glucose metabolism. PPAR-γ isoform expression is tissue [158]. This isoform is the molecular target of a class of insulin-sensitizing drugs known as thiazolidinediones [159]. The PPAR-γ isotypes are capable of reducing inflammatory responses, which are intimately connected to ROS generation in addition to insulin resistance and atherosclerosis. Like mitochondria, peroxisomal ROS/RNS is not considered as a hallmark of T2DM. However, it appears that there is a positive correlation between dysfunctional peroxisomes and the elevation of cellular ROS levels [160] and dysregulated lipid metabolism [161]. Since removal of damaged peroxisomes is not currently possible, antioxidants have been used to maintain a correct redox balance in the cell. The structure and function of peroxisomes and peroxisomal enzymes involved in the production of different ROS are reviewed elsewhere [157].
3.1.3. Endoplasmic reticulum
Hyperglycemia causes endoplasmic reticulum (ER) stress, which has a key role in a number of pathophysiological processes, including endothelial dysfunction, apoptosis in renal cells [162], and pancreatic β-cell failure [163]. The communication networks between ER stress and oxidative stress is by no means unidirectional as previously described [164].
ER not only regulates the folding and post-translational maturation of a third of all proteins [165] and most secreted proteins, but it also plays a critical roles in lipid biosynthesis, detoxification, and reduction-oxidation balance. Numerous endoplasmic reticulum-localized enzymes including cytochrome p-450 and b5 enzymes and diamine oxidase contribute to the formation of ROS [166], suggesting the ER has a major influence to maintaining cellular integrity and viability. Another important thiol oxidase enzyme of oxidative protein-folding cycle, endoplasmic reticulum oxireductin 1 (ERO1) catalyzes the transfer of electrons from dithiols to molecular oxygen results in the formation of H2O2 [167]. During the protein folding process, activation of NADPH oxidase 4, NADPH-P450 reductase, and glutathione appears to have a causal role in the formation of ROS [168]. The enzymes of ER are responsible to improve protein-folding and coping with ongoing oxidative stress. However, the accumulation of unfolded or misfolded proteins within the lumen of the ER causes the induction of ER stress, which impairs normal cellular function via excessive buildup of oxidant H2O2. Antidiabetic drugs are used as first line of defense to improve glucose metabolism. Antioxidants, as the second line of defense, that scavenge the active radicals have been considered as an additional molecular mechanism for clinical T2DM treatment.
4. Visiting the most recent antioxidant of medicinal plants and their therapeutic potential and mechanisms in diabetes
Many plants that possess phenolic compounds are found to have important protective role in scavenging free radicals during oxidative stress [169]. These studies assess the antioxidant properties of polyphenolic compounds in both in-vitro and in-vivo models for studying diabetes as summarized in (Figure 4). Here, we provide a brief review on the most frequently used antioxidants to assess their beneficial pharmacological effect on animal body and the human.
4.1. Flavonoids
Polyphenols are diverse group of natural compounds that fall into four distinct classes: flavonoids, phenolic acids, stilbenes, and lignans. The effects of the most common polyphenols on the biomarkers of oxidative stress are summarized in (Table 1). Flavonoids are most abundant polyphenols which is found in an optimal human diet. This class of compound is further grouped into flavones, flavonols, flavan-3ols, isoflavones, anthocyanidins, and flavanones [170]. Phenolic acids are divided into two sub-groups called hydroxyl benzoic and hydroxyl cinnamic acids. The anti-inflammatory [171] and antioxidant properties of flavonoids [172] have been reviewed (Table 1 and Table 2). Though the mechanisms underlying benefits of flavonoids are complex and remain incompletely understood, flavonoids have been shown to reduce glycemia and related complications [173,174]. In contrast, others think the bioavailability and effectiveness of flavonoids are relatively low [170]. So, it was suggested that nanoparticle systems should be used to prolong circulation and flavonoid efficacy, and potentially reduce their non-specific bindings.
Seaweed and tropical papaya have been utilized as traditional remedies, and passed down through generations. Based on animal research, both papaya leaves and seaweed exhibit beneficial effects in diabetes treatment. They protect against diabetes-induced β cell damage, reduce fasting plasma glucose levels, decrease A1C, increase the expression of antioxidative enzyme, and reduce ROS production [175]. Both seaweed and papaya also contain package of antioxidants (vitamins A, C, and E complexes), other substances such as polysaccharides, phenolic compounds, crucial fatty acids, saponins, fucoidan, and phlorotannin found naturally in other fruits and vegetables. It is unlikely that flavonoids alone can accomplish all diabetes outcomes.
Linarin, a natural flavonoid compound, is capable of counteracting oxidative stress and exerting anti-inflammatory effect in diabetic mice [176]. Inhibition of aldo-keto reductase (AKR)1B, an NADP(H)-oxidoreductase, by linarin appears to serve as a mechanism for reducing oxidative stress and inflammation in a high-glucose and high-palmitic acid-induced hepatocyte injury model and a type 2 diabetic rat model [177].
Isorhamnetin, a methylated derivative of quercetin, belongs to the flavonoid group of phenolic compounds [178]. Isorhamnetin appears to hold great promise against diabetes via aldose reductase inhibition [179]. The important role the isorhamnetin plays in lowering glucose concentrations, improving oxidative status, reducing inflammation, and adjusting lipid metabolism in both in-vitro and in-vivo models is reviewed [180] suggesting that isorhamnetin may be a useful compound for treatment of diabetes.
Amomum tsao-ko Crevost et Lemarie (A. tsao-ko), a very common dietary spice, is rich in flavonoids [181]. A. tsao-ko methanol extracts appear to exert remarkable antioxidant and antidiabetic effects in both in-vitro and in-vivo studies. Nevertheless, further investigations are needed to confirm the detailed mechanisms underlying A. tsao-ko-mediated control of diabetes and oxidative stress.
With the importance of apigenin in numerous physiological functions, there has been great interest in its strong antioxidant and anti-inflammatory actions [182]. It’s action is mediated through neutralizing superoxide, singlet oxygen, and hydroxyl radicals, enhancing the function of the proliferator-activated receptor gamma (PPARγ) signaling, and suppressing CD38 [183,184,185].
Licochalcone A, a flavonoid derived from licorice, has been shown to have potential preventative and therapeutic effects on diabetic nephropathy in both in-vitro and in-vivo studies [186,187]. The antidiabetic effects of the most recent common polyphenols on diabetes are summarized in Table 3.
Myricitrin, a flavone isolated from the bark of Myrica esculenta, has demonstrated a substantial reduction in blood glucose levels in type 2 diabetic mouse and rat models [188]. It facilitates glucose absorption by skeletal muscles through the activation of IRS-1/PI3K/Akt/GLUT4 signaling, as evidenced by both in-vitro and in-vivo studies. Additionally, myricitrin is shown to mitigate oxidative stress by scavenging and neutralizing oxidative radicals and enhancing the body's natural oxidative defense through Nrf-2 activation, in both laboratory and live organism studies [189,190].
Biochanin A is a well-known isoflavone for its anti-inflammatory, antihyperlipidemic [191], antioxidant [192], and anti-cancer health benefits [193]. Biochanin A appears to have anti-diabetic effect beyond its function as antioxidant. In animal model of T2DM, it causes increased insulin sensitivity [193], reduced glucose tolerance [193], and a reduction in glycohemoglobin A1C formation. Biochanin A maintains steady blood glucose concentrations within a normal range in a T2DM rat model [194]. A meta-analysis shows that increased transforming growth factor-β (TGF-β) level was associated with a high risk of nephropathy [195]. All 4 subtypes of protease-activated receptors (PAR1-4) are mainly expressed in the renal epithelial, endothelial and podocyte cells [196]. PAR-2 inhibition improves autophagy and prevents fibrosis and inflammation [196]. Surprisingly, biochanin A reduces diabetic nephropathy via suppression of TGF-β1 and PAR-2 genes expression [191,192].
Formononetin is an isoflavone from the group of phytoestrogens that induces cell apoptosis via the intrinsic apoptosis pathway, which causes the permeabilization of the mitochondrial outer membrane [197,198]. Formononetin has a wide range of biological activities, including eliciting antioxidant [199] (Table 1) and antidiabetic properties in-vitro and in-vivo (Table 3). Sirtuin 1 (SIRT1), a nicotinamide adenine dinucleotide (NAD)-dependent histone deacetylase, protects cells from ROS. In addition, SIRT1 regulates hepatic lipid metabolism by increasing AMP-activated protein kinase leading to the inhibition of hepatic lipogenic pathways in favor of fatty acid oxidation [200]. Formononetin reduces hyperglycemia by increasing SIRT1 expression in pancreatic cells [201]. Sirtuin-induced fatty acid oxidation is important, because it lowers cytoplasmic lipid accumulation. However, elevated fatty acid oxidation could interfere with glucose metabolism in the muscle [202]. In addition, formononetin is considered as a strong apoptotic inducer [198]. Several other mechanisms of action of formononetin have been described [199,203,204]. It is suggested to be used as adjunct therapy for diabetic neuropathy and nephropathy. Clearly, further research investigation is needed in order to gain a better understanding of how precisely formononetin improves multiple aspects of metabolic syndrome, including diabetes.
Hesperetin, a derivative of hesperidin, is a bioflavonoid compound found in citrus fruits [205]. Like formononetin, hesperetin has been shown to improve diabetes (Table 3) by regulating SIRT1, alleviating inflammation [205], combating oxidative stress [206], and reducing insulin resistance [206].
Naringenin, a flavonoid compound found in propolis, exhibits potent anti-hyperglycemic and anti-hyperlipidemic properties in diabetic rat models [207]. The actions of naringenin include improving hyperglycemia, insulinemia, insulin sensitivity, pancreatic cell performance, and lipid profile [184,207].
Like other flavonoids, kaempferol exerts both antioxidant (Table 1) and anti-inflammatory effects (Table 2). Kaempferol produces anticancer effects through inhibition of epidermal growth factor receptor (EGFR) -dependent Src proto-oncogene, nonreceptor trosin kinase (SRC), ERK1/2, and AKT serine/threonine kinase (AKT) pathways [208], and cleavage of poly(ADP-ribose) polymerase (PARP) [209], an endogenous substrate of caspase [210]. Thus, kaempferol is an antiproliferative, anti-metastatic, and apoptotic drug. Kaempferol exerts antidiabetic effect (Table 3) through targeting multiple pathways, including improving glycolysis, glucose uptake, glycogen synthesis, AMPK activity, and Glut4 expression [211]. Additionally several in-vivo studies reported antidiabetic effect of kaempferol through decreasing plasma glucose level [212], increasing plasma insulin level [212], decreasing glucose synthesis [213], and increasing glucagon-like peptide 1 (GLP-1) and insulin release [214]. Readers are invited to a recent review on all pharmacological mechanisms of kaempferol in diabetes [215].
Galangin, a flavonoid, exerts an antioxidant (Table 1) and anti-inflammatory activity [216,217], whereas it elicits a potent antitumor activity in diverse cancers [218]. It is a potent inhibitor of dipeptidyl peptidase-4 (DPP-4) [219], an integral membrane protein expressed on cells. While DPP-4 cleaves a large number of bioactive molecules, its major physiological substrate is the incretin hormone, glucagon-like peptide-1 (GLP-1), which is responsible for the maintenance of normal glucose homeostasis [220]. Thus, galangin is an anti-hyperglycemic agent. Although the mechanism of action is not yet completely understood, a recent evidence suggests that galangin improves lipid profile and plasma insulin level [221].
Fisetin, a bioactive flavonol molecule, has antiproliferative [222], apoptotic [223], and antioxidant [224] activities (Table 1). Fisetin shows antidiabetic effect, which is mediated by two different mechanisms [225]. Firstly, fisetin inhibits gluconeogenesis by inhibiting the transport of pyruvate into the mitochondria and reducing the cytosolic NADH/NAD(+) potential redox [225]. Secondly, fisetin inhibits glycogen breakdown leading to a reduction in blood glucose level [226,227]. Fisetin represents a promising therapeutic strategy that may synergize with other antidiabetic therapies.
Myricetin, a flavonoid, is widely distributed in different types of fruits, herbs, and tea [228]. Recent studies have reported myricetin’s mechanism of action in diabetes, such as inhibiting DPP4 [229], an inactivator of GLP-1, or serving as a GLP-1 receptor agonist [230]. Surprisingly, myricetin appears to normalize intestinal flora of type 2 diabetic mice [231].
Anthocyanins, polyphenolic compounds of the flavonoid group, regulate digestive enzymes (α-amylase and α-glucosidase), GLUT-4, GLP-1, G6Pase, PEPCK, or PPARγ. These compounds also manage blood glucose levels by normalizing insulin secretion and insulin resistance [232]. Additional significant mechanisms involve the protection of pancreatic β cells, through their anti-inflammatory and antioxidant properties [233]. It is unlikely that anthocyanins alone can regulate various classes of enzymes. Further investigations are required to confirm the clinical utility of the anthocyanins for the treatment diabetes, and studies should be standardized and quantified to draw universal conclusions regarding their true use as suggested by the authors [232].
The extract of Delonix regia shows hypoglycemic, antioxidant, and hypolipidemic properties [234]. Of note, the antidiabetic effect of this extract was found to be similar to that of glibenclamide, a well-known antidiabetic agent, which lowers blood glucose levels by stimulating the amount of insulin produced by the pancreas. However, the constituents in Delonix regia extracts that exert antioxidant, hypoglycemic, and adverse effects remain uncharacterized.
Mulberry (Morus alba L.) leaves, used extensively as an effective traditional Chinese medicine for blood glucose management, have been reported to have a beneficial protection on skeletal muscle function [235]. It appears that the flavonoids present in mulberry leaves markedly ameliorate skeletal muscle insulin resistance and enhance mitochondrial function in diabetic mice through the AMPK-PGC-1α signaling pathway [235,236,237]. Flavonoids of mulberry leaves show hypoglycemic effects via inhibiting the TGF-β1 pathway [238] and increasing antioxidase activity. Although very interesting, more randomized controlled trials should be required to warrant the findings of this study.
Vaccarin (VAC), a potent flavonoid glycoside extracted from Vaccariae Semen, has been discovered to improve blood glucose levels and insulin resistance, reduces oxidative stress, and enhances endothelium-dependent vasodilation in T2DM mouse models [239,240], (Table 1). Thus, Vaccariae Semen may serve as an inhibitor of ROS and RNS, which are instigators of several illnesses as mentioned earlier (Table 2). It was further shown that vaccarin improves glucose metabolism and vascular endothelial function through the inhibition of the ROS/AMPK/miRNA-34a/eNOS signaling cascade [239]. The protective effects of vaccarin against ROS and RNS as a dietary supplement could be used as adjuvant therapy for T2DM.
A recent research has shown that the flavonoid extract of fenugreek restores antioxidant enzyme activities of both SOD and catalase, and causes a decrease in malondialdehyde content [241]. This extract also alleviates hyperglycemia in streptozotocin (STZ)-induced T2DM mouse model.
Taxifolin, a recognized flavonoid, is found to be most active towards inhibition of α-amylase. Thus, taxifolin has the ability to manage post-meal hyperglycemia [242]. It also prevents diabetic cardiomyopathy via inhibition of oxidative stress [243]. This function, in conjunction with its anti-inflammatory and antioxidant properties, can contribute to the treatment of Diabetes Mellitus.
A flavonoid-rich fraction derived from Trichilia emetica exhibits a prominent radical scavenging and antidiabetic activities [244]. Binding of free ferrous to oxygen yields ferric iron and superoxide, thus generating hydrogen peroxide. Formed hydrogen peroxide reacts with ferrous iron and gives rise to hydroxyl radical formation. Trichilia emetica flavonoid-rich fractions are found to be capable of chelating ferrous ions. Although this plant could potentially be a valuable agent exhibiting strong antioxidant activity for controlling high blood sugar levels, the constituents in Trichilia emetica extracts remain uncharacterized.
2,3-dihydroxybenzoic acid (DHBA), a phenol, is a weak iron chelator and radical scavenger [245] (Table 2). This colonic-derived flavonoid metabolite is found to regulate glucose uptake and production in renal tubular NRK-52E cells [246]. Additional studies are needed to confirm whether or not this phenol offers a treatment benefit in a reliable large animal diabetes model.
4.2. Catechins
Catechin, a flavonoid, is found in a variety of foods and herbs including tea. As direct antioxidants, catechins are ROS scavengers and metal ion chelators (Table 1) [247]. As indirect antioxidants, they induce the activation of antioxidant enzymes [248], although catechins become pro-oxidant at micromolar levels in cells supplemented with iron and linoleic acid [247]. The antioxidant enzyme pathway is a key player to maintain the delicate balance between ROS and RNS generation and antioxidants in both animal cells and plant cells. Although antioxidant enzyme levels do not appear to be significantly changed in various tissues of aging animals [249,250,251], stimulating this pathway to activate the expression [252] and the delivery of catalytic mimetic of antioxidant enzyme pathway [253] has been considered to be important for preventing diseases caused by oxidative stress and various exogenous substances [254,255,256]. Various pumpkin species contain large amounts of catechin and kaempferol [257]. The pumpkin pulp shows antioxidant effects based on DPPH radicals and ferric reducing antioxidant power (FRAP) radicals. However, the antioxidant potential of these polyphenols cannot be estimated based on the content of individual bioactive compounds as described in detail in the paper.
Epigallocatechin Gallate (EGCG), a sub-class of flavonoids, is an antioxidant (Table 1). EGCG is identified not only as a potential autophagy regulator, but it also is capable of diminishing the expressions of inflammation-associated genes in peripheral leukocytes and adipose tissue of non-obese type 2 diabetic Goto-Kakizaki rats [244,258,259]. Evidence indicates that EGCG has pro-oxidant activity due to its instability and autoxidation [260].
Procyanidins are oligomeric compounds composed of catechin and epicatechin molecules. Research suggests that plants rich in procyanidins could have preventive effects on hyperglycemia and type 2 diabetes [261]. The readers can find additional information, including the structure, classification, and the underlying mechanisms of catechins in regulating diabetes by (Wen et al., 2022) [262].
Although catechins have been considered beneficial in protecting against various diseases caused by oxidative stress and ROS, such as CVD, diabetes [262], and in hematology [263], their effects on CVD biomarkers such as the levels of C-reactive protein, B-type natriuretic peptide, N-terminal pro-atrial natriuretic peptide, aldosterone, renin, fibrinogen, D-dimer, plasminogen-activator inhibitor type 1, and homocysteine; and the urinary albumin-to-creatinine ratio are not fully assessed.
4.3. Phytoestrogens
Lignan, a phytoestrogen, and polyphenols isolated from Linum usitatissimum show antidiabetic properties in streptozotocin (STZ)-induced diabetic rats [264]. Daily treatment with flaxseed extract improves the HbA1c levels and blood glucose levels. It also reduces the total cholesterol, HDL, LDL, and triglyceride levels significantly in diabetic rats.
5. Clinical trial of antioxidant therapy in patients with diabetes
Inflammation is intimately tied to higher glycated hemoglobin (HbA1c) levels, a good indicator of chronic hyperglycemia. Setting the treatment target for HbA1c below 6% in high-risk diabetic patients leads to reduced 5-year nonfatal myocardial infarctions in the diabetes (ACCORD) Study Group. A new model of treating patients with antioxidant with inflammatory actions is emerging as a promising therapeutic option for patients with T2DM. The evidence provided by these various studies supports that ginger, resveratrol, and rutin flavonoid could reduce the HbA1c level, blood glucose level, and homeostatic model evaluation of insulin resistance (HOMA)-IR, an indicator of long-term glycemic control, while allium sativum and olea europaea oil and astaxanthin cause a decrease in low-density lipoprotein (LDL) and serum total cholesterol (Table 4).
Mudan granules possess neuroprotective effect in diabetic patients. Apparently, there is new opportunity for these antioxidants and anti-inflammatory like resveratrol and rutin flavonoid to demonstrate their activity in combination with mudan granules. Notably, while ginger causes an increase in NO production, resveratrol and rutin flavonoid reduce C-reactive protein and interleukin 6 (IL-6) respectively. Thus, antioxidants may be effective for the treatment of cytokine storm in diabetes. The fact that these compounds were able to pass preclinical research and enter human studies indicate their potential effectiveness. The above data show the clinical utility of antioxidants with anti-inflammatory effects in T2DM. However, the application of these antioxidants requires further research investigations to evaluate their efficacy against T2DM-related complications, including hyperlipidemia, pro-inflammatory mediators, and insulin insensitivity. There is also need for selecting diabetic patients in various stages of the disease to guarantee that the anti-inflammatory and antidiabetic effects of the compound is made evident.
6. Conclusions and future implications
ROS mediate many important signaling functions in different cellular and developmental processes of human cells. The level and sources of intracellular ROS are critically important for their actions in influencing different complex signaling and biochemical pathways under both physiological and pathophysiological conditions.
Diabetes, a metabolic disorder, is associated with reduced levels of antioxidants, which can reduce the susceptibility of pancreatic islet to oxidative stress. Oxidative stress impairs insulin-mediated intracellular signaling pathways. Moreover, while insulin resistance is a complex metabolic disorder, oxidative stress can induce insulin resistance.
Diabetes triggers changes in endothelial function, which may lead to increased oxidative stress mediators and a reduction in antioxidants. Oxidative stress can be the result of mitochondrial dysfunction or inflammation, not the cause. Evidence indicates that oxidative stress accelerates the development of complications in diabetes. Identifying novel biomarkers and therapeutic strategies to prevent diabetes-induced endothelial dysfunction are of a great interest.
The United Kingdom Prospective Diabetes Study clearly verifies the importance of glycemic control in reducing the risk for microvascular complications. While the efficacy of antioxidative treatments remain still elusive, recent clinical reports indicate that antioxidant have the ability to counteract inflammation and improve hyperglycemia, as well as beneficial effects on hepatocyte lipid metabolism through different mechanisms in diabetes (Table 4). The mechanism by which the antioxidants reduce hyperglycemia are only partly known. Since diabetes is a progressive disease, the T2DM patients’ need for different treatments changes as well. It remains to be shown whether antioxidant treatments administered together with current antidiabetic or anti-inflammatory drugs can prevent the vascular complications of diabetes. Further studies are required to clarify the role of antioxidant therapy in the management of type 2 diabetes.
Author Contributions
Conceptualization, Z.SM., and F.M.; writing—original draft preparation, A.A., S.B., Z.SM.; writing—review and editing, Z.SM., A.A., S.B., F.M.; supervision, Z.SM. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- García-Cerrillo, D.; Noriega-Cisneros, R.; Peña-Montes, D.; Huerta-Cervantes, M.; Silva-Ríos, M.; Salgado-Garciglia, R.; Montoya-Pérez, R.; Saavedra-Molina, A. Antioxidant effects of Eryngium carlinae in diabetic rats. Asian Journal of Applied Sciences 2018, 6. [Google Scholar]
- Pagliari, S.; Forcella, M.; Lonati, E.; Sacco, G.; Romaniello, F.; Rovellini, P.; Fusi, P.; Palestini, P.; Campone, L.; Labra, M.; et al. Antioxidant and Anti-Inflammatory Effect of Cinnamon (Cinnamomum verum J. Presl) Bark Extract after In Vitro Digestion Simulation. Foods 2023, 12. [Google Scholar] [CrossRef]
- Stanciauskaite, M.; Marksa, M.; Babickaite, L.; Majiene, D.; Ramanauskiene, K. Comparison of Ethanolic and Aqueous Populus balsamifera L. Bud Extracts by Different Extraction Methods: Chemical Composition, Antioxidant and Antibacterial Activities. Pharmaceuticals (Basel) 2021, 14. [Google Scholar] [CrossRef]
- Stanciauskaite, M.; Marksa, M.; Rimkiene, L.; Ramanauskiene, K. Evaluation of Chemical Composition, Sun Protection Factor and Antioxidant Activity of Lithuanian Propolis and Its Plant Precursors. Plants (Basel) 2022, 11. [Google Scholar] [CrossRef]
- Oladimeji, O.H.; Owere, P.O.; Anthony, P.C. Acetylation of Cinnamic Acid and Evaluation of Antioxidant Activity of the Resultant Derivative. Biomedical Journal of Scientific & Technical Research 2021, 39, 31084–31088. [Google Scholar] [CrossRef]
- Kiliç, I.; Yeşiloğlu, Y. Spectroscopic studies on the antioxidant activity of p-coumaric acid. Spectrochim Acta A Mol Biomol Spectrosc 2013, 115, 719–724. [Google Scholar] [CrossRef]
- Gülçin, I. Antioxidant activity of caffeic acid (3,4-dihydroxycinnamic acid). Toxicology 2006, 217, 213–220. [Google Scholar] [CrossRef]
- Bian, Y.Y.; Guo, J.; Majeed, H.; Zhu, K.X.; Guo, X.N.; Peng, W.; Zhou, H.M. Ferulic acid renders protection to HEK293 cells against oxidative damage and apoptosis induced by hydrogen peroxide. In Vitro Cell Dev Biol Anim 2015, 51, 722–729. [Google Scholar] [CrossRef]
- Nenadis, N.; Tsimidou, M. Observations on the estimation of scavenging activity of phenolic compounds using rapid 1,1-diphenyl-2-picrylhydrazyl (DPPH•) tests. Journal of the American Oil Chemists' Society 2002, 79, 1191. [Google Scholar] [CrossRef]
- Chandrasekar, V.; Belur, P.D.; Regupathi, I. Effect of hydroxybenzoic acids antioxidants on the oxidative stability of sardine oil. Resource-Efficient Technologies 2016, 2, S114–S118. [Google Scholar] [CrossRef]
- Badhani, B.; Sharma, N.; Kakkar, R. Gallic acid: a versatile antioxidant with promising therapeutic and industrial applications. RSC Advances 2015, 5, 27540–27557. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, L.; Wang, X.; Wu, W.; Qin, R. Effect of Syringic acid on antioxidant biomarkers and associated inflammatory markers in mice model of asthma. Drug Dev Res 2019, 80, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Youn, H.S.; Lee, J.Y.; Saitoh, S.I.; Miyake, K.; Kang, K.W.; Choi, Y.J.; Hwang, D.H. Suppression of MyD88- and TRIF-dependent signaling pathways of toll-like receptor by (−)-epigallocatechin-3-gallate, a polyphenol component of green tea. Biochemical Pharmacology 2006, 72, 850–859. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, S.; Zhang, W.; Zhao, P.; He, B.; Wu, N.; Han, P. Epigallocatechin-3-O-gallate (EGCG) attenuates FFAs-induced peripheral insulin resistance through AMPK pathway and insulin signaling pathway in vivo. Diabetes Res Clin Pract 2011, 93, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Hamza Sherif, S.; Gebreyohannes, B. Synthesis, Characterization, and Antioxidant Activities of Genistein, Biochanin A, and Their Analogues. Journal of Chemistry 2018, 2018, 4032105. [Google Scholar] [CrossRef]
- Jain, P.G.; Nayse, P.G.; Patil, D.J.; Shinde, S.D.; Surana, S.J. The possible antioxidant capabilities of formononetin in guarding against streptozotocin-induced diabetic nephropathy in rats. Future Journal of Pharmaceutical Sciences 2020, 6, 53. [Google Scholar] [CrossRef]
- Samie, A.; Sedaghat, R.; Baluchnejadmojarad, T.; Roghani, M. Hesperetin, a citrus flavonoid, attenuates testicular damage in diabetic rats via inhibition of oxidative stress, inflammation, and apoptosis. Life Sci 2018, 210, 132–139. [Google Scholar] [CrossRef]
- Baranowska, M.; Koziara, Z.; Suliborska, K.; Chrzanowski, W.; Wormstone, M.; Namieśnik, J.; Bartoszek, A. Interactions between polyphenolic antioxidants quercetin and naringenin dictate the distinctive redox-related chemical and biological behaviour of their mixtures. Scientific Reports 2021, 11, 12282. [Google Scholar] [CrossRef]
- Rashmi, R.; Bojan Magesh, S.; Mohanram Ramkumar, K.; Suryanarayanan, S.; Venkata SubbaRao, M. Antioxidant Potential of Naringenin Helps to Protect Liver Tissue from Streptozotocin-Induced Damage. Rep Biochem Mol Biol 2018, 7, 76–84. [Google Scholar]
- Tran, M.H.; Nguyen, H.D.; Kim, J.C.; Choi, J.S.; Lee, H.K.; Min, B.S. Phenolic glycosides from Alangium salviifolium leaves with inhibitory activity on LPS-induced NO, PGE(2), and TNF-alpha production. Bioorg Med Chem Lett 2009, 19, 4389–4393. [Google Scholar] [CrossRef]
- Aloud, A.A.; Veeramani, C.; Govindasamy, C.; Alsaif, M.A.; El Newehy, A.S.; Al-Numair, K.S. Galangin, a dietary flavonoid, improves antioxidant status and reduces hyperglycemia-mediated oxidative stress in streptozotocin-induced diabetic rats. Redox Rep 2017, 22, 290–300. [Google Scholar] [CrossRef]
- Maher, P. Modulation of the Neuroprotective and Anti-inflammatory Activities of the Flavonol Fisetin by the Transition Metals Iron and Copper. Antioxidants 2020, 9, 1113. [Google Scholar] [PubMed]
- Awad, H.M.; Boersma, M.G.; Vervoort, J.; Rietjens, I.M. Peroxidase-catalyzed formation of quercetin quinone methide–glutathione adducts. Archives of Biochemistry and Biophysics 2000, 378, 224–233. [Google Scholar]
- Park, K.-S.; Chong, Y.; Kim, M.K. Myricetin: biological activity related to human health. Applied Biological Chemistry 2016, 59, 259–269. [Google Scholar] [CrossRef]
- Topal, F.; Nar, M.; Gocer, H.; Kalin, P.; Kocyigit, U.M.; Gülçin, İ.; Alwasel, S.H. Antioxidant activity of taxifolin: an activity–structure relationship. Journal of Enzyme Inhibition and Medicinal Chemistry 2016, 31, 674–683. [Google Scholar] [CrossRef]
- Buranasudja, V.; Muangnoi, C.; Sanookpan, K.; Halim, H.; Sritularak, B.; Rojsitthisak, P. Eriodictyol Attenuates H(2)O(2)-Induced Oxidative Damage in Human Dermal Fibroblasts through Enhanced Capacity of Antioxidant Machinery. Nutrients 2022, 14. [Google Scholar] [CrossRef]
- Hu, H.-C.; Lei, Y.-H.; Zhang, W.-H.; Luo, X.-Q. Antioxidant and Anti-inflammatory Properties of Resveratrol in Diabetic Nephropathy: A Systematic Review and Meta-analysis of Animal Studies. Frontiers in Pharmacology 2022, 13. [Google Scholar] [CrossRef]
- Lu, Y.; Xue, Y.; Chen, S.; Zhu, H.; Zhang, J.; Li, X.-N.; Wang, J.; Liu, J.; Qi, C.; Du, G.; et al. Antioxidant Lignans and Neolignans from Acorus tatarinowii. Scientific Reports 2016, 6, 22909. [Google Scholar] [CrossRef]
- Alvarez Cilleros, D.; Lopez-Oliva, M.E.; Martin, M.A.; Ramos, S. (-)-Epicatechin and the colonic metabolite 2,3-dihydroxybenzoic acid protect against high glucose and lipopolysaccharide-induced inflammation in renal proximal tubular cells through NOX-4/p38 signalling. Food Funct 2020, 11, 8811–8824. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Jing, H.; Gan, L.; Li, H.; Luo, B. Resveratrol attenuated estrogen-deficient-induced cardiac dysfunction: role of AMPK, SIRT1, and mitochondrial function. Am J Transl Res 2016, 8, 2641–2649. [Google Scholar]
- Zhang, L.X.; Li, C.X.; Kakar, M.U.; Khan, M.S.; Wu, P.F.; Amir, R.M.; Dai, D.F.; Naveed, M.; Li, Q.Y.; Saeed, M.; et al. Resveratrol (RV): A pharmacological review and call for further research. Biomed Pharmacother 2021, 143, 112164. [Google Scholar] [CrossRef]
- Abu-Amero, K.K.; Kondkar, A.A.; Chalam, K.V. Resveratrol and Ophthalmic Diseases. Nutrients 2016, 8, 200. [Google Scholar]
- Michalak, B.; Filipek, A.; Chomicki, P.; Pyza, M.; Woźniak, M.; Żyżyńska-Granica, B.; Piwowarski, J.P.; Kicel, A.; Olszewska, M.A.; Kiss, A.K. Lignans From Forsythia x Intermedia Leaves and Flowers Attenuate the Pro-inflammatory Function of Leukocytes and Their Interaction With Endothelial Cells. Frontiers in Pharmacology 2018, 9. [Google Scholar] [CrossRef]
- Kuryłowicz, A. The Role of Isoflavones in Type 2 Diabetes Prevention and Treatment-A Narrative Review. Int J Mol Sci 2020, 22. [Google Scholar] [CrossRef]
- Sorrenti, V.; Raffaele, M.; Vanella, L.; Acquaviva, R.; Salerno, L.; Pittala, V.; Intagliata, S.; Di Giacomo, C. Protective Effects of Caffeic Acid Phenethyl Ester (CAPE) and Novel Cape Analogue as Inducers of Heme Oxygenase-1 in Streptozotocin-Induced Type 1 Diabetic Rats. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Kose, T.; Sharp, P.A.; Latunde-Dada, G.O. Phenolic Acids Rescue Iron-Induced Damage in Murine Pancreatic Cells and Tissues. Molecules 2023, 28. [Google Scholar] [CrossRef]
- Yang, J.; Zhu, C.; Ye, J.; Lv, Y.; Wang, L.; Chen, Z.; Jiang, Z. Protection of Porcine Intestinal-Epithelial Cells from Deoxynivalenol-Induced Damage by Resveratrol via the Nrf2 Signaling Pathway. J Agric Food Chem 2019, 67, 1726–1735. [Google Scholar] [CrossRef]
- Fukumitsu, S.; Aida, K.; Shimizu, H.; Toyoda, K. Flaxseed lignan lowers blood cholesterol and decreases liver disease risk factors in moderately hypercholesterolemic men. Nutrition Research 2010, 30, 441–446. [Google Scholar] [CrossRef]
- Memon, A.R.; Rajput, M.A.; Rizwan, F.; Akram, M.; Rizwan, M.; Iqbal, Z. Effect of Allium sativum and Olea europaea on serum lipids in patients with diabetes mellitus. J Taibah Univ Med Sci 2023, 18, 420–426. [Google Scholar] [CrossRef]
- Ciaraldi, T.P.; Boeder, S.C.; Mudaliar, S.R.; Giovannetti, E.R.; Henry, R.R.; Pettus, J.H. Astaxanthin, a natural antioxidant, lowers cholesterol and markers of cardiovascular risk in individuals with prediabetes and dyslipidaemia. Diabetes Obes Metab 2023, 25, 1985–1994. [Google Scholar] [CrossRef] [PubMed]
- Karimzadeh, L.; Sohrab, G.; Hedayati, M.; Ebrahimof, S.; Emami, G.; Razavion, T. Effects of concentrated beetroot juice consumption on glycemic control, blood pressure, and lipid profile in type 2 diabetes patients: randomized clinical trial study. Ir J Med Sci 2023, 192, 1143–1153. [Google Scholar] [CrossRef]
- Rostamkhani, H.; Veisi, P.; Niknafs, B.; Jafarabadi, M.A.; Ghoreishi, Z. The effect of zingiber officinale on prooxidant-antioxidant balance and glycemic control in diabetic patients with ESRD undergoing hemodialysis: a double-blind randomized control trial. BMC Complement Med Ther 2023, 23, 52. [Google Scholar] [CrossRef]
- Zhang, A.; Wang, Q.; Liu, M.; Tan, M.; Zhang, X.; Wu, R. Efficacy and safety of Mudan granules for painful diabetic peripheral neuropathy: A protocol for a double-blind randomized controlled trial. Medicine (Baltimore) 2022, 101, e28896. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Martinez, B.I.; Ruiz-Ramos, M.; Pedraza-Chaverri, J.; Santiago-Osorio, E.; Mendoza-Nunez, V.M. Effect of Resveratrol on Markers of Oxidative Stress and Sirtuin 1 in Elderly Adults with Type 2 Diabetes. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Fatima, S.; Khan, D.A.; Aamir, M.; Pervez, M.A.; Fatima, F. delta-Tocotrienol in Combination with Resveratrol Improves the Cardiometabolic Risk Factors and Biomarkers in Patients with Metabolic Syndrome: A Randomized Controlled Trial. Metab Syndr Relat Disord 2023, 21, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Bazyar, H.; Moradi, L.; Zaman, F.; Zare Javid, A. The effects of rutin flavonoid supplement on glycemic status, lipid profile, atherogenic index of plasma, brain-derived neurotrophic factor (BDNF), some serum inflammatory, and oxidative stress factors in patients with type 2 diabetes mellitus: A double-blind, placebo-controlled trial. Phytother Res 2023, 37, 271–284. [Google Scholar] [CrossRef]
- National Diabetes Statistics Report 2020, Estimates of Diabetes and its burden in the United States. 2020.
- Perng, W.; Conway, R.; Mayer-Davis, E.; Dabelea, D. Youth-Onset Type 2 Diabetes: The Epidemiology of an Awakening Epidemic. Diabetes Care 2023, 46, 490–499. [Google Scholar] [CrossRef]
- Willemsen, G.; Ward, K.J.; Bell, C.G.; Christensen, K.; Bowden, J.; Dalgård, C.; Harris, J.R.; Kaprio, J.; Lyle, R.; Magnusson, P.K.; et al. The Concordance and Heritability of Type 2 Diabetes in 34,166 Twin Pairs From International Twin Registers: The Discordant Twin (DISCOTWIN) Consortium. Twin Res Hum Genet 2015, 18, 762–771. [Google Scholar] [CrossRef]
- Almgren, P.; Lehtovirta, M.; Isomaa, B.; Sarelin, L.; Taskinen, M.R.; Lyssenko, V.; Tuomi, T.; Groop, L. Heritability and familiality of type 2 diabetes and related quantitative traits in the Botnia Study. Diabetologia 2011, 54, 2811–2819. [Google Scholar] [CrossRef]
- Mambiya, M.; Shang, M.; Wang, Y.; Li, Q.; Liu, S.; Yang, L.; Zhang, Q.; Zhang, K.; Liu, M.; Nie, F.; et al. The Play of Genes and Non-genetic Factors on Type 2 Diabetes. Frontiers in Public Health 2019, 7. [Google Scholar] [CrossRef]
- Udler, M.S. Type 2 Diabetes: Multiple Genes, Multiple Diseases. Curr Diab Rep 2019, 19, 55. [Google Scholar] [CrossRef]
- Caulfield, J.I.; Aizenbud, L.; Perdigoto, A.L.; Meffre, E.; Jilaveanu, L.; Michalek, D.A.; Rich, S.S.; Aizenbud, Y.; Adeniran, A.; Herold, K.C.; et al. Germline genetic variants are associated with development of insulin-dependent diabetes in cancer patients treated with immune checkpoint inhibitors. J Immunother Cancer 2023, 11. [Google Scholar] [CrossRef]
- Kamiya, J.; Aoki, Y. Associations between hyperglycaemia and somatic transversion mutations in mitochondrial DNA of people with diabetes mellitus. Diabetologia 2003, 46, 1559–1566. [Google Scholar] [CrossRef]
- Voight, B.F.; Scott, L.J.; Steinthorsdottir, V.; Morris, A.P.; Dina, C.; Welch, R.P.; Zeggini, E.; Huth, C.; Aulchenko, Y.S.; Thorleifsson, G.; et al. Twelve type 2 diabetes susceptibility loci identified through large-scale association analysis. Nat Genet 2010, 42, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Billings, L.K.; Florez, J.C. The genetics of type 2 diabetes: what have we learned from GWAS? Ann N Y Acad Sci 2010, 1212, 59–77. [Google Scholar] [CrossRef] [PubMed]
- Heijmans, B.T.; Tobi, E.W.; Stein, A.D.; Putter, H.; Blauw, G.J.; Susser, E.S.; Slagboom, P.E.; Lumey, L.H. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc Natl Acad Sci U S A 2008, 105, 17046–17049. [Google Scholar] [CrossRef] [PubMed]
- Seki, Y.; Williams, L.; Vuguin, P.M.; Charron, M.J. Minireview: Epigenetic programming of diabetes and obesity: animal models. Endocrinology 2012, 153, 1031–1038. [Google Scholar] [CrossRef]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc Natl Acad Sci U S A 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Torres, I.; Castrejón-Téllez, V.; Soto, M.E.; Rubio-Ruiz, M.E.; Manzano-Pech, L.; Guarner-Lans, V. Oxidative Stress, Plant Natural Antioxidants, and Obesity. International Journal of Molecular Sciences 2021, 22, 1786. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M.; Schmidt, A.M. Oxidative Stress and Diabetic Complications. Circulation Research 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Pitocco, D.; Tesauro, M.; Alessandro, R.; Ghirlanda, G.; Cardillo, C. Oxidative Stress in Diabetes: Implications for Vascular and Other Complications. International Journal of Molecular Sciences 2013, 14, 21525–21550. [Google Scholar]
- Kelsey, M.M.; Zaepfel, A.; Bjornstad, P.; Nadeau, K.J. Age-related consequences of childhood obesity. Gerontology 2014, 60, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Afkarian, M. Diabetic kidney disease in children and adolescents. Pediatr Nephrol 2015, 30, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Bjerregaard, L.G.; Jensen, B.W.; Angquist, L.; Osler, M.; Sorensen, T.I.A.; Baker, J.L. Change in Overweight from Childhood to Early Adulthood and Risk of Type 2 Diabetes. The New England journal of medicine 2018, 378, 1302–1312. [Google Scholar] [CrossRef] [PubMed]
- Bianco-Miotto, T.; Craig, J.M.; Gasser, Y.P.; van Dijk, S.J.; Ozanne, S.E. Epigenetics and DOHaD: from basics to birth and beyond. Journal of developmental origins of health and disease 2017, 8, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.A.; Filippi, B.M.; Kang, G.M.; Kim, M.S.; Lam, T.K. Insulin action in the hypothalamus and dorsal vagal complex. Experimental physiology 2014, 99, 1104–1109. [Google Scholar] [CrossRef]
- Cade, W.T. Diabetes-related microvascular and macrovascular diseases in the physical therapy setting. Phys Ther 2008, 88, 1322–1335. [Google Scholar] [CrossRef]
- Rahman, S.; Rahman, T.; Ismail, A.A.; Rashid, A.R. Diabetes-associated macrovasculopathy: pathophysiology and pathogenesis. Diabetes Obes Metab 2007, 9, 767–780. [Google Scholar] [CrossRef]
- Chow, L.S.; Chen, H.; Miller, M.E.; Marcovina, S.M.; Seaquist, E.R. Biomarkers related to severe hypoglycaemia and lack of good glycaemic control in ACCORD. Diabetologia 2015, 58, 1160–1166. [Google Scholar] [CrossRef]
- Fanelli, C.; Calderone, S.; Epifano, L.; De Vincenzo, A.; Modarelli, F.; Pampanelli, S.; Perriello, G.; De Feo, P.; Brunetti, P.; Gerich, J.E.; et al. Demonstration of a critical role for free fatty acids in mediating counterregulatory stimulation of gluconeogenesis and suppression of glucose utilization in humans. J Clin Invest 1993, 92, 1617–1622. [Google Scholar] [CrossRef]
- Cao, C.; Koh, H.E.; Van Vliet, S.; Patterson, B.W.; Reeds, D.N.; Laforest, R.; Gropler, R.J.; Mittendorfer, B. Increased plasma fatty acid clearance, not fatty acid concentration, is associated with muscle insulin resistance in people with obesity. Metabolism 2022, 132, 155216. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alexandria Journal of Medicine 2018, 54, 287–293. [Google Scholar] [CrossRef]
- King, G.L. The role of hyperglycaemia and hyperinsulinaemia in causing vascular dysfunction in diabetes. Ann Med 1996, 28, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J Clin Invest 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Leyane, T.S.; Jere, S.W.; Houreld, N.N. Oxidative Stress in Ageing and Chronic Degenerative Pathologies: Molecular Mechanisms Involved in Counteracting Oxidative Stress and Chronic Inflammation. International Journal of Molecular Sciences 2022, 23, 7273. [Google Scholar]
- Tangvarasittichai, S. Oxidative stress, insulin resistance, dyslipidemia and type 2 diabetes mellitus. World J Diabetes 2015, 6, 456–480. [Google Scholar] [CrossRef]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. American Journal of Physiology-Lung Cellular and Molecular Physiology 2000, 279, L1005–L1028. [Google Scholar] [CrossRef]
- Sinenko, S.A.; Starkova, T.Y.; Kuzmin, A.A.; Tomilin, A.N. Physiological Signaling Functions of Reactive Oxygen Species in Stem Cells: From Flies to Man. Frontiers in Cell and Developmental Biology 2021, 9. [Google Scholar] [CrossRef]
- Kakimoto, M.; Inoguchi, T.; Sonta, T.; Yu, H.Y.; Imamura, M.; Etoh, T.; Hashimoto, T.; Nawata, H. Accumulation of 8-hydroxy-2'-deoxyguanosine and mitochondrial DNA deletion in kidney of diabetic rats. Diabetes 2002, 51, 1588–1595. [Google Scholar] [CrossRef] [PubMed]
- Nekhaeva, E.; Bodyak, N.D.; Kraytsberg, Y.; McGrath, S.B.; Van Orsouw, N.J.; Pluzhnikov, A.; Wei, J.Y.; Vijg, J.; Khrapko, K. Clonally expanded mtDNA point mutations are abundant in individual cells of human tissues. Proc Natl Acad Sci U S A 2002, 99, 5521–5526. [Google Scholar] [CrossRef]
- Wang, K.; Zhou, W.; Meng, P.; Wang, P.; Zhou, C.; Yao, Y.; Wu, S.; Wang, Y.; Zhao, J.; Zou, D.; et al. Immune-related somatic mutation genes are enriched in PDACs with diabetes. Transl Oncol 2019, 12, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-κB, inflammation, and metabolic disease. Cell Metab 2011, 13, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Kay, A.M.; Simpson, C.L.; Stewart, J.A., Jr. The Role of AGE/RAGE Signaling in Diabetes-Mediated Vascular Calcification. J Diabetes Res 2016, 2016, 6809703. [Google Scholar] [CrossRef]
- Agrawal, A.; Cha-Molstad, H.; Samols, D.; Kushner, I. Overexpressed nuclear factor-κB can participate in endogenous C-reactive protein induction, and enhances the effects of C/EBPβ and signal transducer and activator of transcription-3. Immunology 2003, 108, 539–547. [Google Scholar] [CrossRef]
- Spranger, J.; Kroke, A.; Möhlig, M.; Hoffmann, K.; Bergmann, M.M.; Ristow, M.; Boeing, H.; Pfeiffer, A.F.H. Inflammatory Cytokines and the Risk to Develop Type 2 Diabetes: Results of the Prospective Population-Based European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam Study. Diabetes 2003, 52, 812–817. [Google Scholar] [CrossRef] [PubMed]
- Suryavanshi, S.V.; Kulkarni, Y.A. NF-κβ: A Potential Target in the Management of Vascular Complications of Diabetes. Front Pharmacol 2017, 8, 798. [Google Scholar] [CrossRef]
- Malhotra, J.D.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress: a vicious cycle or a double-edged sword? Antioxid Redox Signal 2007, 9, 2277–2293. [Google Scholar] [CrossRef] [PubMed]
- Kairisalo, M.; Korhonen, L.; Blomgren, K.; Lindholm, D. X-linked inhibitor of apoptosis protein increases mitochondrial antioxidants through NF-kappaB activation. Biochem Biophys Res Commun 2007, 364, 138–144. [Google Scholar] [CrossRef]
- Pham, C.G.; Bubici, C.; Zazzeroni, F.; Papa, S.; Jones, J.; Alvarez, K.; Jayawardena, S.; De Smaele, E.; Cong, R.; Beaumont, C.; et al. Ferritin heavy chain upregulation by NF-kappaB inhibits TNFalpha-induced apoptosis by suppressing reactive oxygen species. Cell 2004, 119, 529–542. [Google Scholar] [CrossRef]
- Lingappan, K. NF-κB in Oxidative Stress. Curr Opin Toxicol 2018, 7, 81–86. [Google Scholar] [CrossRef]
- Tsalamandris, S.; Antonopoulos, A.S.; Oikonomou, E.; Papamikroulis, G.A.; Vogiatzi, G.; Papaioannou, S.; Deftereos, S.; Tousoulis, D. The Role of Inflammation in Diabetes: Current Concepts and Future Perspectives. Eur Cardiol 2019, 14, 50–59. [Google Scholar] [CrossRef]
- Liu, C.; Dong, W.; Li, J.; Kong, Y.; Ren, X. O-GlcNAc Modification and Its Role in Diabetic Retinopathy. Metabolites 2022, 12, 725. [Google Scholar] [CrossRef]
- Wu, T.; Qiao, S.; Shi, C.; Wang, S.; Ji, G. Metabolomics window into diabetic complications. Journal of Diabetes Investigation 2018, 9, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Mantegazza, A.R.; Savina, A.; Vermeulen, M.; Pérez, L.; Geffner, J.; Hermine, O.; Rosenzweig, S.D.; Faure, F.; Amigorena, S. NADPH oxidase controls phagosomal pH and antigen cross-presentation in human dendritic cells. Blood 2008, 112, 4712–4722. [Google Scholar] [CrossRef] [PubMed]
- Chougnet, C.A.; Thacker, R.I.; Shehata, H.M.; Hennies, C.M.; Lehn, M.A.; Lages, C.S.; Janssen, E.M. Loss of Phagocytic and Antigen Cross-Presenting Capacity in Aging Dendritic Cells Is Associated with Mitochondrial Dysfunction. The Journal of Immunology 2015, 195, 2624–2632. [Google Scholar] [CrossRef]
- Gao, Z.; Chen, X. Fatty Acid β-Oxidation in Kidney Diseases: Perspectives on Pathophysiological Mechanisms and Therapeutic Opportunities. Frontiers in Pharmacology 2022, 13. [Google Scholar] [CrossRef]
- Pawlak, M.; Lefebvre, P.; Staels, B. Molecular mechanism of PPARα action and its impact on lipid metabolism, inflammation and fibrosis in non-alcoholic fatty liver disease. Journal of Hepatology 2015, 62, 720–733. [Google Scholar] [CrossRef] [PubMed]
- Ke, R.; Xu, Q.; Li, C.; Luo, L.; Huang, D. Mechanisms of AMPK in the maintenance of ATP balance during energy metabolism. Cell Biology International 2018, 42, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Rohm, T.V.; Meier, D.T.; Olefsky, J.M.; Donath, M.Y. Inflammation in obesity, diabetes, and related disorders. Immunity 2022, 55, 31–55. [Google Scholar] [CrossRef]
- Nordestgaard, B.G. Triglyceride-Rich Lipoproteins and Atherosclerotic Cardiovascular Disease: New Insights From Epidemiology, Genetics, and Biology. Circ Res 2016, 118, 547–563. [Google Scholar] [CrossRef]
- Borén, J.; Chapman, M.J.; Krauss, R.M.; Packard, C.J.; Bentzon, J.F.; Binder, C.J.; Daemen, M.J.; Demer, L.L.; Hegele, R.A.; Nicholls, S.J.; et al. Low-density lipoproteins cause atherosclerotic cardiovascular disease: pathophysiological, genetic, and therapeutic insights: a consensus statement from the European Atherosclerosis Society Consensus Panel. Eur Heart J 2020, 41, 2313–2330. [Google Scholar] [CrossRef] [PubMed]
- Mussbacher, M.; Schossleitner, K.; Kral-Pointner, J.B.; Salzmann, M.; Schrammel, A.; Schmid, J.A. More than Just a Monolayer: the Multifaceted Role of Endothelial Cells in the Pathophysiology of Atherosclerosis. Current Atherosclerosis Reports 2022, 24, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Kornowski, R.; Hong, M.K.; Tio, F.O.; Bramwell, O.; Wu, H.; Leon, M.B. In-Stent Restenosis: Contributions of Inflammatory Responses and Arterial Injury to Neointimal Hyperplasia. Journal of the American College of Cardiology 1998, 31, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Che Man, R.; Sulaiman, N.; Ishak, M.F.; Bt Hj Idrus, R.; Abdul Rahman, M.R.; Yazid, M.D. The Effects of Pro-Inflammatory and Anti-Inflammatory Agents for the Suppression of Intimal Hyperplasia: An Evidence-Based Review. Int J Environ Res Public Health 2020, 17. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. National Diabetes Statistics Report, 2020; Centers for Disease Control and Prevention, U.S. Dept of Health and Human Services: Atlanta, GA, USA, 2020. [Google Scholar]
- Favre, G.A.; Esnault, V.L.M.; Obberghen, E.V. Modulation of glucose metabolism by the renin-angiotensin-aldosterone system. American Journal of Physiology-Endocrinology and Metabolism 2015, 308, E435–E449. [Google Scholar] [CrossRef]
- Petrie, J.R.; Guzik, T.J.; Touyz, R.M. Diabetes, Hypertension, and Cardiovascular Disease: Clinical Insights and Vascular Mechanisms. Can J Cardiol 2018, 34, 575–584. [Google Scholar] [CrossRef]
- Cheung, B.M.; Li, C. Diabetes and hypertension: is there a common metabolic pathway? Curr Atheroscler Rep 2012, 14, 160–166. [Google Scholar] [CrossRef]
- Dregan, A.; Charlton, J.; Chowienczyk, P.; Gulliford, M.C. Chronic inflammatory disorders and risk of type 2 diabetes mellitus, coronary heart disease, and stroke: a population-based cohort study. Circulation 2014, 130, 837–844. [Google Scholar] [CrossRef]
- Armstrong, A.W.; Harskamp, C.T.; Armstrong, E.J. Psoriasis and the risk of diabetes mellitus: a systematic review and meta-analysis. JAMA Dermatol 2013, 149, 84–91. [Google Scholar] [CrossRef]
- Zhang, J.; Fu, L.; Shi, J.; Chen, X.; Li, Y.; Ma, B.; Zhang, Y. The risk of metabolic syndrome in patients with rheumatoid arthritis: a meta-analysis of observational studies. PLoS One 2013, 8, e78151. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic inflammation in the etiology of disease across the life span. Nature Medicine 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Sharpless, N.E. Senescence in Health and Disease. Cell 2017, 169, 1000–1011. [Google Scholar] [CrossRef] [PubMed]
- Coppé, J.P.; Desprez, P.Y.; Krtolica, A.; Campisi, J. The senescence-associated secretory phenotype: the dark side of tumor suppression. Annu Rev Pathol 2010, 5, 99–118. [Google Scholar] [CrossRef]
- Esposito, K.; Nappo, F.; Marfella, R.; Giugliano, G.; Giugliano, F.; Ciotola, M.; Quagliaro, L.; Ceriello, A.; Giugliano, D. Inflammatory Cytokine Concentrations Are Acutely Increased by Hyperglycemia in Humans. Circulation 2002, 106, 2067–2072. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, R.; González, J.; Paoletto, F. The role of oxidative stress in the pathophysiology of hypertension. Hypertension Research 2011, 34, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Lacy, F.; Kailasam, M.T.; O'Connor, D.T.; Schmid-Schönbein, G.W.; Parmer, R.J. Plasma hydrogen peroxide production in human essential hypertension: role of heredity, gender, and ethnicity. Hypertension 2000, 36, 878–884. [Google Scholar] [CrossRef]
- Redón, J.; Oliva, M.R.; Tormos, C.; Giner, V.; Chaves, J.; Iradi, A.; Sáez, G.T. Antioxidant activities and oxidative stress byproducts in human hypertension. Hypertension 2003, 41, 1096–1101. [Google Scholar] [CrossRef]
- Rajagopalan, S.; Kurz, S.; Münzel, T.; Tarpey, M.; Freeman, B.A.; Griendling, K.K.; Harrison, D.G. Angiotensin II-mediated hypertension in the rat increases vascular superoxide production via membrane NADH/NADPH oxidase activation. Contribution to alterations of vasomotor tone. J Clin Invest 1996, 97, 1916–1923. [Google Scholar] [CrossRef]
- Jung, O.; Schreiber, J.G.; Geiger, H.; Pedrazzini, T.; Busse, R.; Brandes, R.P. gp91phox-containing NADPH oxidase mediates endothelial dysfunction in renovascular hypertension. Circulation 2004, 109, 1795–1801. [Google Scholar] [CrossRef]
- Terada, Y.; Yayama, K. Angiotensin II-Induced Vasoconstriction via Rho Kinase Activation in Pressure-Overloaded Rat Thoracic Aortas. Biomolecules 2021, 11, 1076. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, A.; Kang, B.P.; Cheung, S.; Opawumi, D.; Meggs, L.G. Angiotensin II promotes glucose-induced activation of cardiac protein kinase C isozymes and phosphorylation of troponin I. Diabetes 2001, 50, 1918–1926. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Smith, R.D.; Balla, T.; Catt, K.J. Angiotensin II activates mitogen-activated protein kinase via protein kinase C and Ras/Raf-1 kinase in bovine adrenal glomerulosa cells. Endocrinology 1998, 139, 1801–1809. [Google Scholar] [CrossRef] [PubMed]
- Togliatto, G.; Lombardo, G.; Brizzi, M.F. The Future Challenge of Reactive Oxygen Species (ROS) in Hypertension: From Bench to Bed Side. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef]
- Millatt, L.J.; Abdel-Rahman, E.M.; Siragy, H.M. Angiotensin II and nitric oxide: a question of balance. Regul Pept 1999, 81, 1–10. [Google Scholar] [CrossRef]
- Chen, K.; Pittman, R.N.; Popel, A.S. Nitric oxide in the vasculature: where does it come from and where does it go? A quantitative perspective. Antioxid Redox Signal 2008, 10, 1185–1198. [Google Scholar] [CrossRef]
- Kawanami, D.; Takashi, Y.; Muta, Y.; Oda, N.; Nagata, D.; Takahashi, H.; Tanabe, M. Mineralocorticoid Receptor Antagonists in Diabetic Kidney Disease. Frontiers in Pharmacology 2021, 12. [Google Scholar] [CrossRef]
- Hill, M.A.; Yang, Y.; Zhang, L.; Sun, Z.; Jia, G.; Parrish, A.R.; Sowers, J.R. Insulin resistance, cardiovascular stiffening and cardiovascular disease. Metabolism 2021, 119, 154766. [Google Scholar] [CrossRef]
- Gaede, P.; Vedel, P.; Larsen, N.; Jensen, G.V.; Parving, H.H.; Pedersen, O. Multifactorial intervention and cardiovascular disease in patients with type 2 diabetes. N Engl J Med 2003, 348, 383–393. [Google Scholar] [CrossRef]
- Sena, C.M.; Leandro, A.; Azul, L.; Seica, R.; Perry, G. Vascular Oxidative Stress: Impact and Therapeutic Approaches. Front Physiol 2018, 9, 1668. [Google Scholar] [CrossRef]
- Tekin, K.; Tekin, M.I. Chapter 3 - Oxidative stress and diabetic retinopathy. In Pathology, Preedy, V.R., Ed.; Academic Press: 2020; pp. 29-37.
- Davi, G.; Falco, A.; Patrono, C. Lipid peroxidation in diabetes mellitus. Antioxid Redox Signal 2005, 7, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int J Biomed Sci 2008, 4, 89–96. [Google Scholar]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int J Biochem Cell Biol 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid Med Cell Longev 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Burgos-Moron, E.; Abad-Jimenez, Z.; Maranon, A.M.; Iannantuoni, F.; Escribano-Lopez, I.; Lopez-Domenech, S.; Salom, C.; Jover, A.; Mora, V.; Roldan, I.; et al. Relationship Between Oxidative Stress, ER Stress, and Inflammation in Type 2 Diabetes: The Battle Continues. J Clin Med 2019, 8. [Google Scholar] [CrossRef]
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell Signal 2012, 24, 981–990. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ Res 2010, 107, 1058–1070. [Google Scholar] [CrossRef]
- Bolisetty, S.; Jaimes, E.A. Mitochondria and reactive oxygen species: physiology and pathophysiology. Int J Mol Sci 2013, 14, 6306–6344. [Google Scholar] [CrossRef]
- Venditti, P.; Di Stefano, L.; Di Meo, S. Mitochondrial metabolism of reactive oxygen species. Mitochondrion 2013, 13, 71–82. [Google Scholar] [CrossRef]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen Res 2013, 8, 2003–2014. [Google Scholar] [CrossRef]
- Kowluru, R.A.; Mohammad, G. Mitochondrial Fragmentation in a High Homocysteine Environment in Diabetic Retinopathy. Antioxidants (Basel) 2022, 11. [Google Scholar] [CrossRef]
- Nadalutti, C.A.; Ayala-Pena, S.; Santos, J.H. Mitochondrial DNA damage as driver of cellular outcomes. Am J Physiol Cell Physiol 2022, 322, C136–C150. [Google Scholar] [CrossRef]
- Duraisamy, A.J.; Mohammad, G.; Kowluru, R.A. Mitochondrial fusion and maintenance of mitochondrial homeostasis in diabetic retinopathy. Biochim Biophys Acta Mol Basis Dis 2019, 1865, 1617–1626. [Google Scholar] [CrossRef]
- So, S.; Lee, S.; Lee, Y.; Han, J.; Kang, S.; Choi, J.; Kim, B.; Kim, D.; Yoo, H.J.; Shim, I.K.; et al. Dysfunctional pancreatic cells differentiated from induced pluripotent stem cells with mitochondrial DNA mutations. BMB Rep 2022, 55, 453–458. [Google Scholar] [CrossRef]
- Neginskaya, M.A.; Pavlov, E.V.; Sheu, S.S. Electrophysiological properties of the mitochondrial permeability transition pores: Channel diversity and disease implication. Biochim Biophys Acta Bioenerg 2021, 1862, 148357. [Google Scholar] [CrossRef]
- Scialo, F.; Fernandez-Ayala, D.J.; Sanz, A. Role of Mitochondrial Reverse Electron Transport in ROS Signaling: Potential Roles in Health and Disease. Front Physiol 2017, 8, 428. [Google Scholar] [CrossRef] [PubMed]
- Manford, A.G.; Mena, E.L.; Shih, K.Y.; Gee, C.L.; McMinimy, R.; Martinez-Gonzalez, B.; Sherriff, R.; Lew, B.; Zoltek, M.; Rodriguez-Perez, F.; et al. Structural basis and regulation of the reductive stress response. Cell 2021, 184, 5375–5390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shen, Y.; Kim, I.M.; Weintraub, N.L.; Tang, Y. The Impaired Bioenergetics of Diabetic Cardiac Microvascular Endothelial Cells. Front Endocrinol (Lausanne) 2021, 12, 642857. [Google Scholar] [CrossRef]
- Barnes, P.J. Pulmonary Diseases and Ageing. Subcell Biochem 2019, 91, 45–74. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Mehrabi Nasab, E.; Athari, S.M.; Athari, S.S. Mitochondria signaling pathways in allergic asthma. J Investig Med 2022, 70, 863–882. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.; Mathur, A.; Kakkar, P. Critical role of mitochondrial dysfunction and impaired mitophagy in diabetic nephropathy. J Cell Physiol 2019, 234, 19223–19236. [Google Scholar] [CrossRef]
- Ding, L.; Sun, W.; Balaz, M.; He, A.; Klug, M.; Wieland, S.; Caiazzo, R.; Raverdy, V.; Pattou, F.; Lefebvre, P.; et al. Peroxisomal β-oxidation acts as a sensor for intracellular fatty acids and regulates lipolysis. Nature Metabolism 2021, 3, 1648–1661. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.-S.; Dighe, P.A.; Mezera, V.; Monternier, P.-A.; Brand, M.D. Production of superoxide and hydrogen peroxide from specific mitochondrial sites under different bioenergetic conditions. Journal of Biological Chemistry 2017, 292, 16804–16809. [Google Scholar] [CrossRef] [PubMed]
- Schrader, M.; Fahimi, H.D. Peroxisomes and oxidative stress. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research 2006, 1763, 1755–1766. [Google Scholar] [CrossRef] [PubMed]
- Desvergne, B.; Wahli, W. Peroxisome proliferator-activated receptors: nuclear control of metabolism. Endocr Rev 1999, 20, 649–688. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.M.; Moore, L.B.; Smith-Oliver, T.A.; Wilkison, W.O.; Willson, T.M.; Kliewer, S.A. An antidiabetic thiazolidinedione is a high affinity ligand for peroxisome proliferator-activated receptor gamma (PPAR gamma). J Biol Chem 1995, 270, 12953–12956. [Google Scholar] [CrossRef]
- Pascual-Ahuir, A.; Manzanares-Estreder, S.; Proft, M. Pro- and Antioxidant Functions of the Peroxisome-Mitochondria Connection and Its Impact on Aging and Disease. Oxidative Medicine and Cellular Longevity 2017, 2017, 9860841. [Google Scholar] [CrossRef]
- Kim, J.; Bai, H. Peroxisomal Stress Response and Inter-Organelle Communication in Cellular Homeostasis and Aging. Antioxidants 2022, 11, 192. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, R.; Torreggiani, M.; Ting, A.; Xiong, H.; Striker, G.E.; Vlassara, H.; Zheng, F. Induction of Diabetes in Aged C57B6 Mice Results in Severe Nephropathy: An Association with Oxidative Stress, Endoplasmic Reticulum Stress, and Inflammation. The American Journal of Pathology 2010, 176, 2163–2176. [Google Scholar] [CrossRef]
- Cunard, R.; Sharma, K. The endoplasmic reticulum stress response and diabetic kidney disease. American Journal of Physiology-Renal Physiology 2011, 300, F1054–F1061. [Google Scholar] [CrossRef]
- Cao, S.S.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress in cell fate decision and human disease. Antioxid Redox Signal 2014, 21, 396–413. [Google Scholar] [CrossRef]
- Huh, W.K.; Falvo, J.V.; Gerke, L.C.; Carroll, A.S.; Howson, R.W.; Weissman, J.S.; O'Shea, E.K. Global analysis of protein localization in budding yeast. Nature 2003, 425, 686–691. [Google Scholar] [CrossRef]
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS Generation and Antioxidant Defense Systems in Normal and Malignant Cells. Oxidative Medicine and Cellular Longevity 2019, 2019, 6175804. [Google Scholar] [CrossRef]
- Zito, E. ERO1: A protein disulfide oxidase and H2O2 producer. Free Radical Biology and Medicine 2015, 83, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Zeeshan, H.M.; Lee, G.H.; Kim, H.R.; Chae, H.J. Endoplasmic Reticulum Stress and Associated ROS. Int J Mol Sci 2016, 17, 327. [Google Scholar] [CrossRef]
- Khojasteh, A.; Mirjalili, M.H.; Alcalde, M.A.; Cusido, R.M.; Eibl, R.; Palazon, J. Powerful Plant Antioxidants: A New Biosustainable Approach to the Production of Rosmarinic Acid. Antioxidants (Basel) 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Tan, B.; Murtaza, G.; Liu, G.; Rahu, N.; Saleem Kalhoro, M.; Hussain Kalhoro, D.; Adebowale, T.O.; Usman Mazhar, M.; Rehman, Z.U.; et al. Flavonoids and type 2 diabetes: Evidence of efficacy in clinical and animal studies and delivery strategies to enhance their therapeutic efficacy. Pharmacol Res 2020, 152, 104629. [Google Scholar] [CrossRef] [PubMed]
- González, R.; Ballester, I.; López-Posadas, R.; Suárez, M.D.; Zarzuelo, A.; Martínez-Augustin, O.; Sánchez de Medina, F. Effects of flavonoids and other polyphenols on inflammation. Crit Rev Food Sci Nutr 2011, 51, 331–362. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: an overview. J Nutr Sci 2016, 5, e47. [Google Scholar] [CrossRef]
- Zou, J.; Yu, X.; Qu, S.; Li, X.; Jin, Y.; Sui, D. Protective effect of total flavonoids extracted from the leaves of Murraya paniculata (L.) Jack on diabetic nephropathy in rats. Food Chem Toxicol 2014, 64, 231–237. [Google Scholar] [CrossRef]
- Jin, Y.; Arroo, R. The protective effects of flavonoids and carotenoids against diabetic complications-A review of in vivo evidence. Front Nutr 2023, 10, 1020950. [Google Scholar] [CrossRef]
- Nyakundi, B.B.; Yang, J. Uses of Papaya Leaf and Seaweed Supplementations for Controlling Glucose Homeostasis in Diabetes. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Han, X.; Wu, Y.C.; Meng, M.; Sun, Q.S.; Gao, S.M.; Sun, H. Linarin prevents LPS-induced acute lung injury by suppressing oxidative stress and inflammation via inhibition of TXNIP/NLRP3 and NF-kappaB pathways. Int J Mol Med 2018, 42, 1460–1472. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Shan, M.Y.; Tang, C.Y.; Cheng, M.Y.; Chen, B.; Yan, J.; Xu, Z.H. Linarin Ameliorates Diabetic Liver Injury by Alleviating Oxidative Stress and Inflammation through the Inhibition of AKR1B1. Comb Chem High Throughput Screen 2023. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Mitchell, A.E. Quercetin and Isorhamnetin Glycosides in Onion (Allium cepa L.): Varietal Comparison, Physical Distribution, Coproduct Evaluation, and Long-Term Storage Stability. Journal of Agricultural and Food Chemistry 2011, 59, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Abdel Motaal, A.; Salem, H.H.; Almaghaslah, D.; Alsayari, A.; Bin Muhsinah, A.; Alfaifi, M.Y.; Elbehairi, S.E.I.; Shati, A.A.; El-Askary, H. Flavonol Glycosides: In Vitro Inhibition of DPPIV, Aldose Reductase and Combating Oxidative Stress are Potential Mechanisms for Mediating the Antidiabetic Activity of Cleome droserifolia. Molecules 2020, 25. [Google Scholar] [CrossRef]
- Kalai, F.Z.; Boulaaba, M.; Ferdousi, F.; Isoda, H. Effects of Isorhamnetin on Diabetes and Its Associated Complications: A Review of In Vitro and In Vivo Studies and a Post Hoc Transcriptome Analysis of Involved Molecular Pathways. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Zhang, X.F.; Tang, Y.J.; Guan, X.X.; Lu, X.; Li, J.; Chen, X.L.; Deng, J.L.; Fan, J.M. Flavonoid constituents of Amomum tsao-ko Crevost et Lemarie and their antioxidant and antidiabetic effects in diabetic rats - in vitro and in vivo studies. Food Funct 2022, 13, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Qi, M.; Li, P.; Zhan, Y.; Shao, H. Apigenin in cancer therapy: anti-cancer effects and mechanisms of action. Cell Biosci 2017, 7, 50. [Google Scholar] [CrossRef]
- Alam, W.; Rocca, C.; Khan, H.; Hussain, Y.; Aschner, M.; De Bartolo, A.; Amodio, N.; Angelone, T.; Cheang, W.S. Current Status and Future Perspectives on Therapeutic Potential of Apigenin: Focus on Metabolic-Syndrome-Dependent Organ Dysfunction. Antioxidants (Basel) 2021, 10. [Google Scholar] [CrossRef]
- Laaroussi, H.; Bakour, M.; Ousaaid, D.; Aboulghazi, A.; Ferreira-Santos, P.; Genisheva, Z.; Teixeira, J.A.; Lyoussi, B. Effect of antioxidant-rich propolis and bee pollen extracts against D-glucose induced type 2 diabetes in rats. Food Res Int 2020, 138, 109802. [Google Scholar] [CrossRef]
- Bakour, M.; Laaroussi, H.; Ferreira-Santos, P.; Genisheva, Z.; Ousaaid, D.; Teixeira, J.A.; Lyoussi, B. Exploring the Palynological, Chemical, and Bioactive Properties of Non-Studied Bee Pollen and Honey from Morocco. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Luo, Z.; Fu, C.; Li, T.; Gao, Q.; Miao, D.; Xu, J.; Zhao, Y. Hypoglycemic Effects of Licochalcone A on the Streptozotocin-Induced Diabetic Mice and Its Mechanism Study. J Agric Food Chem 2021, 69, 2444–2456. [Google Scholar] [CrossRef]
- Luo, Z.; Li, T.; Gao, Q.; Chen, Y.; Su, G.; Zhao, Y. Impact of licochalcone A on the progression of diabetic nephropathy in type 2 diabetes mellitus of C57BL/6 mice. Food Funct 2021, 12, 10676–10689. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, Y.; Shen, Q.; Liu, G.; Ye, J.; Sun, G.; Sun, X. Myricitrin Attenuates High Glucose-Induced Apoptosis through Activating Akt-Nrf2 Signaling in H9c2 Cardiomyocytes. Molecules 2016, 21. [Google Scholar] [CrossRef]
- Zhang, B.; Shen, Q.; Chen, Y.; Pan, R.; Kuang, S.; Liu, G.; Sun, G.; Sun, X. Myricitrin Alleviates Oxidative Stress-induced Inflammation and Apoptosis and Protects Mice against Diabetic Cardiomyopathy. Sci Rep 2017, 7, 44239. [Google Scholar] [CrossRef] [PubMed]
- Ahangarpour, A.; Oroojan, A.A.; Khorsandi, L.; Kouchak, M.; Badavi, M. Solid Lipid Nanoparticles of Myricitrin Have Antioxidant and Antidiabetic Effects on Streptozotocin-Nicotinamide-Induced Diabetic Model and Myotube Cell of Male Mouse. Oxid Med Cell Longev 2018, 2018, 7496936. [Google Scholar] [CrossRef]
- Amri, J.; Alaee, M.; Babaei, R.; Salemi, Z.; Meshkani, R.; Ghazavi, A.; Akbari, A.; Salehi, M. Biochanin-A has antidiabetic, antihyperlipidemic, antioxidant, and protective effects on diabetic nephropathy via suppression of TGF-β1 and PAR-2 genes expression in kidney tissues of STZ-induced diabetic rats. Biotechnology and Applied Biochemistry 2022, 69, 2112–2121. [Google Scholar] [PubMed]
- Ram, C.; Gairola, S.; Verma, S.; Mugale, M.N.; Bonam, S.R.; Murty, U.S.; Sahu, B.D. Biochanin A Ameliorates Nephropathy in High-Fat Diet/Streptozotocin-Induced Diabetic Rats: Effects on NF-kB/NLRP3 Axis, Pyroptosis, and Fibrosis. Antioxidants 2023, 12, 1052. [Google Scholar]
- Oza, M.J.; Kulkarni, Y.A. Biochanin A improves insulin sensitivity and controls hyperglycemia in type 2 diabetes. Biomedicine & Pharmacotherapy 2018, 107, 1119–1127. [Google Scholar]
- Sadri, H.; Goodarzi, M.T.; Salemi, Z.; Seifi, M. Antioxidant effects of biochanin A in streptozotocin induced diabetic rats. Brazilian Archives of Biology and Technology 2017, 60. [Google Scholar] [CrossRef]
- Mou, X.; Zhou, D.-Y.; Zhou, D.-Y.; Ma, J.-R.; Liu, Y.-H.; Chen, H.-P.; Hu, Y.-B.; Shou, C.-M.; Chen, J.-W.; Liu, W.-H.; et al. Serum TGF-β1 as a Biomarker for Type 2 Diabetic Nephropathy: A Meta-Analysis of Randomized Controlled Trials. PLOS ONE 2016, 11, e0149513. [Google Scholar] [CrossRef] [PubMed]
- Bagang, N.; Gupta, K.; Singh, G.; Kanuri, S.H.; Mehan, S. Protease-activated receptors in kidney diseases: A comprehensive review of pathological roles, therapeutic outcomes and challenges. Chem Biol Interact 2023, 377, 110470. [Google Scholar] [CrossRef]
- Lossi, L. The concept of intrinsic versus extrinsic apoptosis. Biochem J 2022, 479, 357–384. [Google Scholar] [CrossRef]
- Tay, K.-C.; Tan, L.T.-H.; Chan, C.K.; Hong, S.L.; Chan, K.-G.; Yap, W.H.; Pusparajah, P.; Lee, L.-H.; Goh, B.-H. Formononetin: A Review of Its Anticancer Potentials and Mechanisms. Frontiers in Pharmacology 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.G.; Nayse, P.G.; Patil, D.J.; Shinde, S.D.; Surana, S.J. The possible antioxidant capabilities of formononetin in guarding against streptozotocin-induced diabetic nephropathy in rats. Future Journal of Pharmaceutical Sciences 2020, 6, 1–9. [Google Scholar] [CrossRef]
- Hou, X.; Xu, S.; Maitland-Toolan, K.A.; Sato, K.; Jiang, B.; Ido, Y.; Lan, F.; Walsh, K.; Wierzbicki, M.; Verbeuren, T.J.; et al. SIRT1 regulates hepatocyte lipid metabolism through activating AMP-activated protein kinase. J Biol Chem 2008, 283, 20015–20026. [Google Scholar] [CrossRef]
- Oza, M.J.; Kulkarni, Y.A. Formononetin treatment in type 2 diabetic rats reduces insulin resistance and hyperglycemia. Frontiers in pharmacology 2018, 9, 739. [Google Scholar] [CrossRef]
- Zhang, L.; Keung, W.; Samokhvalov, V.; Wang, W.; Lopaschuk, G.D. Role of fatty acid uptake and fatty acid beta-oxidation in mediating insulin resistance in heart and skeletal muscle. Biochim Biophys Acta 2010, 1801, 1–22. [Google Scholar] [CrossRef]
- Lv, J.; Zhuang, K.; Jiang, X.; Huang, H.; Quan, S. Renoprotective effect of formononetin by suppressing Smad3 expression in Db/Db mice. Diabetes, Metabolic Syndrome and Obesity 2020, 3313-3324.
- Zhou, Z.; Zhou, X.; Dong, Y.; Li, M.; Xu, Y. Formononetin ameliorates high glucose-induced endothelial dysfunction by inhibiting the JAK/STAT signaling pathway. Molecular Medicine Reports 2019, 20, 2893–2901. [Google Scholar] [CrossRef]
- Lee, A.; Gu, H.; Gwon, M.-H.; Yun, J.-M. Hesperetin suppresses LPS/high glucose-induced inflammatory responses via TLR/MyD88/NF-κB signaling pathways in THP-1 cells. Nutr. Res. Pract 2021, 15, 591–603. [Google Scholar]
- Tian, M.; Han, Y.-B.; Zhao, C.-C.; Liu, L.; Zhang, F.-L. Hesperidin alleviates insulin resistance by improving HG-induced oxidative stress and mitochondrial dysfunction by restoring miR-149. Diabetology & Metabolic Syndrome 2021, 13, 1–11. [Google Scholar]
- Den Hartogh, D.J.; Tsiani, E. Antidiabetic Properties of Naringenin: A Citrus Fruit Polyphenol. Biomolecules 2019, 9. [Google Scholar] [CrossRef]
- Lee, J.; Kim, J.H. Kaempferol Inhibits Pancreatic Cancer Cell Growth and Migration through the Blockade of EGFR-Related Pathway In Vitro. PLOS ONE 2016, 11, e0155264. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Y.; An, Y.; Fang, G. The mechanism of anticancer action and potential clinical use of kaempferol in the treatment of breast cancer. Biomedicine & Pharmacotherapy 2019, 117, 109086. [Google Scholar] [CrossRef]
- Timmer, J.C.; Salvesen, G.S. Caspase substrates. Cell Death & Differentiation 2007, 14, 66–72. [Google Scholar] [CrossRef]
- Alkhalidy, H.; Moore, W.; Zhang, Y.; McMillan, R.; Wang, A.; Ali, M.; Suh, K.-S.; Zhen, W.; Cheng, Z.; Jia, Z.; et al. Small Molecule Kaempferol Promotes Insulin Sensitivity and Preserved Pancreatic <b><i>β</i></b>-Cell Mass in Middle-Aged Obese Diabetic Mice. Journal of Diabetes Research 2015, 2015, 532984. [Google Scholar] [CrossRef] [PubMed]
- Chandramohan, G.; Al-Numair, K.S.; Alsaif, M.A.; Veeramani, C. Antidiabetic effect of kaempferol a flavonoid compound, on streptozotocin-induced diabetic rats with special reference to glycoprotein components. Progress in Nutrition 2015, 17, 50–57. [Google Scholar]
- Alkhalidy, H.; Moore, W.; Wang, Y.; Luo, J.; McMillan, R.P.; Zhen, W.; Zhou, K.; Liu, D. The flavonoid kaempferol ameliorates streptozotocin-induced diabetes by suppressing hepatic glucose production. Molecules 2018, 23, 2338. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Tekade, R.K.; Kalia, K. Kaempferol in ameliorating diabetes-induced fibrosis and renal damage: an in vitro and in vivo study in diabetic nephropathy mice model. Phytomedicine 2020, 76, 153235. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, Z.; Zhao, X.; Xie, H.; Du, L.; Gao, H.; Xie, C. Mechanisms of Kaempferol in the treatment of diabetes: A comprehensive and latest review. Frontiers in Endocrinology 2022, 13, 990299. [Google Scholar] [PubMed]
- Abukhalil, M.H.; Althunibat, O.Y.; Aladaileh, S.H.; Al-Amarat, W.; Obeidat, H.M.; Alayn'Al-marddyah, A.; Hussein, O.E.; Alfwuaires, M.A.; Algefare, A.I.; Alanazi, K.M. Galangin attenuates diabetic cardiomyopathy through modulating oxidative stress, inflammation and apoptosis in rats. Biomedicine & Pharmacotherapy 2021, 138, 111410. [Google Scholar]
- Al-Amarat, W.; Abukhalil, M.H.; Althunibat, O.Y.; Alfwuaires, M.A.; Alnamshan, M.M.; Alqosaibi, A.I.; Ahmeda, A.F.; Kamel, E.M.; Arab, H.H.; Mahmoud, A.M. Galangin attenuates liver injury, oxidative stress and inflammation, and upregulates Nrf2/HO-1 signaling in streptozotocin-induced diabetic rats. Processes 2021, 9, 1562. [Google Scholar]
- Kong, Y.; Feng, Z.; Chen, A.; Qi, Q.; Han, M.; Wang, S.; Zhang, Y.; Zhang, X.; Yang, N.; Wang, J.; et al. The Natural Flavonoid Galangin Elicits Apoptosis, Pyroptosis, and Autophagy in Glioblastoma. Frontiers in Oncology 2019, 9. [Google Scholar] [CrossRef]
- Kalhotra, P.; Chittepu, V.C.; Osorio-Revilla, G.; Gallardo-Velázquez, T. Discovery of galangin as a potential DPP-4 inhibitor that improves insulin-stimulated skeletal muscle glucose uptake: a combinational therapy for diabetes. International journal of molecular sciences 2019, 20, 1228. [Google Scholar]
- Deacon, C.F. Physiology and Pharmacology of DPP-4 in Glucose Homeostasis and the Treatment of Type 2 Diabetes. Frontiers in Endocrinology 2019, 10. [Google Scholar] [CrossRef]
- Aloud, A.A.; Chinnadurai, V.; Govindasamy, C.; Alsaif, M.A.; Al-Numair, K.S. Galangin, a dietary flavonoid, ameliorates hyperglycaemia and lipid abnormalities in rats with streptozotocin-induced hyperglycaemia. Pharmaceutical biology 2018, 56, 302–308. [Google Scholar] [PubMed]
- Haddad, A.Q.; Venkateswaran, V.; Viswanathan, L.; Teahan, S.J.; Fleshner, N.E.; Klotz, L.H. Novel antiproliferative flavonoids induce cell cycle arrest in human prostate cancer cell lines. Prostate Cancer Prostatic Dis 2006, 9, 68–76. [Google Scholar] [CrossRef]
- Sung, B.; Pandey, M.K.; Aggarwal, B.B. Fisetin, an inhibitor of cyclin-dependent kinase 6, down-regulates nuclear factor-kappaB-regulated cell proliferation, antiapoptotic and metastatic gene products through the suppression of TAK-1 and receptor-interacting protein-regulated IkappaBalpha kinase activation. Mol Pharmacol 2007, 71, 1703–1714. [Google Scholar] [CrossRef]
- Hou, D.X.; Fukuda, M.; Johnson, J.A.; Miyamori, K.; Ushikai, M.; Fujii, M. Fisetin induces transcription of NADPH:quinone oxidoreductase gene through an antioxidant responsive element-involved activation. Int J Oncol 2001, 18, 1175–1179. [Google Scholar] [CrossRef]
- Constantin, R.P.; Constantin, J.; Pagadigorria, C.L.S.; Ishii-Iwamoto, E.L.; Bracht, A.; Ono, M.d.K.C.; Yamamoto, N.S. The actions of fisetin on glucose metabolism in the rat liver. Cell Biochemistry and Function 2010, 28, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Vinayagam, R.; Xu, B. Antidiabetic properties of dietary flavonoids: a cellular mechanism review. Nutrition & metabolism 2015, 12, 1–20. [Google Scholar]
- Prasath, G.S.; Pillai, S.I.; Subramanian, S.P. Fisetin improves glucose homeostasis through the inhibition of gluconeogenic enzymes in hepatic tissues of streptozotocin induced diabetic rats. European journal of pharmacology 2014, 740, 248–254. [Google Scholar] [PubMed]
- Li, Y.; Ding, Y. Minireview: Therapeutic potential of myricetin in diabetes mellitus. Food Science and Human Wellness 2012, 1, 19–25. [Google Scholar]
- Lalitha, N.; Sadashivaiah, B.; Ramaprasad, T.R.; Singh, S.A. Anti-hyperglycemic activity of myricetin, through inhibition of DPP-4 and enhanced GLP-1 levels, is attenuated by co-ingestion with lectin-rich protein. PloS one 2020, 15, e0231543. [Google Scholar]
- Li, Y.; Zheng, X.; Yi, X.; Liu, C.; Kong, D.; Zhang, J.; Gong, M. Myricetin: a potent approach for the treatment of type 2 diabetes as a natural class B GPCR agonist. Faseb j 2017, 31, 2603–2611. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Chen, Y.; Li, X.; Zhu, L.; Wang, X.; Li, L.; Sun, H.; Han, X.; Li, J. Myricetin relieves the symptoms of type 2 diabetes mice and regulates intestinal microflora. Biomedicine & Pharmacotherapy 2022, 153, 113530. [Google Scholar]
- Les, F.; Casedas, G.; Gomez, C.; Moliner, C.; Valero, M.S.; Lopez, V. The role of anthocyanins as antidiabetic agents: from molecular mechanisms to in vivo and human studies. J Physiol Biochem 2021, 77, 109–131. [Google Scholar] [CrossRef]
- Casedas, G.; Les, F.; Gomez-Serranillos, M.P.; Smith, C.; Lopez, V. Anthocyanin profile, antioxidant activity and enzyme inhibiting properties of blueberry and cranberry juices: a comparative study. Food Funct 2017, 8, 4187–4193. [Google Scholar] [CrossRef]
- Khan, D.; Sharif, A.; Zafar, M.; Akhtar, B.; Akhtar, M.F.; Awan, S. Delonix regia a Folklore Remedy for Diabetes; Attenuates Oxidative Stress and Modulates Type II Diabetes Mellitus. Curr Pharm Biotechnol 2020, 21, 1059–1069. [Google Scholar] [CrossRef]
- Meng, Q.; Qi, X.; Fu, Y.; Chen, Q.; Cheng, P.; Yu, X.; Sun, X.; Wu, J.; Li, W.; Zhang, Q.; et al. Flavonoids extracted from mulberry (Morus alba L.) leaf improve skeletal muscle mitochondrial function by activating AMPK in type 2 diabetes. J Ethnopharmacol 2020, 248, 112326. [Google Scholar] [CrossRef] [PubMed]
- Thaipitakwong, T.; Numhom, S.; Aramwit, P. Mulberry leaves and their potential effects against cardiometabolic risks: a review of chemical compositions, biological properties and clinical efficacy. Pharm Biol 2018, 56, 109–118. [Google Scholar] [CrossRef]
- Thaipitakwong, T.; Supasyndh, O.; Rasmi, Y.; Aramwit, P. A randomized controlled study of dose-finding, efficacy, and safety of mulberry leaves on glycemic profiles in obese persons with borderline diabetes. Complement Ther Med 2020, 49, 102292. [Google Scholar] [CrossRef]
- Zhang, Q.; Lu, Y.; Ma, Z.; Li, Y.; Guo, J.; Meng, Q.; Bian, H. A novel formula from mulberry leaf ameliorates diabetic nephropathy in rats via inhibiting the TGF-beta1 pathway. Food Funct 2015, 6, 3307–3315. [Google Scholar] [CrossRef]
- Xu, F.; Liu, Y.; Zhu, X.; Li, S.; Shi, X.; Li, Z.; Ai, M.; Sun, J.; Hou, B.; Cai, W.; et al. Protective Effects and Mechanisms of Vaccarin on Vascular Endothelial Dysfunction in Diabetic Angiopathy. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yu, X.; Zhu, X.; Wen, Y.; Zhu, M.; Cai, W.; Hou, B.; Xu, F.; Qiu, L. Vaccarin alleviates endothelial inflammatory injury in diabetes by mediating miR-570-3p/HDAC1 pathway. Front Pharmacol 2022, 13, 956247. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Lu, S.S.; Wang, H.L.; Li, G.; He, Y.F.; Liu, X.Y.; Rong, R.; Li, J.; Lu, X.C. Effects of the fenugreek extracts on high-fat diet-fed and streptozotocin-induced type 2 diabetic mice. Animal Model Exp Med 2018, 1, 68–73. [Google Scholar] [CrossRef]
- Rehman, K.; Chohan, T.A.; Waheed, I.; Gilani, Z.; Akash, M.S.H. Taxifolin prevents postprandial hyperglycemia by regulating the activity of alpha-amylase: Evidence from an in vivo and in silico studies. J Cell Biochem 2019, 120, 425–438. [Google Scholar] [CrossRef]
- Sun, X.; Chen, R.C.; Yang, Z.H.; Sun, G.B.; Wang, M.; Ma, X.J.; Yang, L.J.; Sun, X.B. Taxifolin prevents diabetic cardiomyopathy in vivo and in vitro by inhibition of oxidative stress and cell apoptosis. Food Chem Toxicol 2014, 63, 221–232. [Google Scholar] [CrossRef]
- Konate, K.; Yomalan, K.; Sytar, O.; Zerbo, P.; Brestic, M.; Patrick, V.D.; Gagniuc, P.; Barro, N. Free Radicals Scavenging Capacity, Antidiabetic and Antihypertensive Activities of Flavonoid-Rich Fractions from Leaves of Trichilia emetica and Opilia amentacea in an Animal Model of Type 2 Diabetes Mellitus. Evid Based Complement Alternat Med 2014, 2014, 867075. [Google Scholar] [CrossRef]
- Xu, J.; Jordan, R.B. Kinetics and mechanism of the oxidation of 2,3-dihydroxybenzoic acid by iron(III). Inorganic Chemistry 1988, 27, 4563–4566. [Google Scholar] [CrossRef]
- Álvarez-Cilleros, D.; Martín, M.; Ramos, S. (-)-Epicatechin and the Colonic 2,3-Dihydroxybenzoic Acid Metabolite Regulate Glucose Uptake, Glucose Production, and Improve Insulin Signaling in Renal NRK-52E Cells. Mol Nutr Food Res 2018, 62. [Google Scholar] [CrossRef]
- Sugihara, N.; Ohnishi, M.; Imamura, M.; Furuno, K. Differences in antioxidative efficiency of catechins in various metal-induced lipid peroxidations in cultured hepatocytes. Journal of health science 2001, 47, 99–106. [Google Scholar] [CrossRef]
- Bernatoniene, J.; Kopustinskiene, D.M. The Role of Catechins in Cellular Responses to Oxidative Stress. Molecules 2018, 23. [Google Scholar] [CrossRef]
- Sohal, R.S.; Arnold, L.A.; Sohal, B.H. Age-related changes in antioxidant enzymes and prooxidant generation in tissues of the rat with special reference to parameters in two insect species. Free Radic Biol Med 1990, 9, 495–500. [Google Scholar] [CrossRef]
- Lopez-Torres, M.; Perez-Campo, R.; Barja de Quiroga, G. Aging in brown fat: antioxidant defenses and oxidative stress. Mech Ageing Dev 1991, 59, 129–137. [Google Scholar] [CrossRef]
- Lopez-Torres, M.; Perez-Campo, R.; Barja de Quiroga, G. Effect of natural ageing and antioxidant inhibition on liver antioxidant enzymes, glutathione system, peroxidation, and oxygen consumption in Rana perezi. J Comp Physiol B 1991, 160, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.-H.; Yeh, C.-T.; Yen, G.-C. Anthocyanins Induce the Activation of Phase II Enzymes through the Antioxidant Response Element Pathway against Oxidative Stress-Induced Apoptosis. Journal of Agricultural and Food Chemistry 2007, 55, 9427–9435. [Google Scholar] [CrossRef] [PubMed]
- Renjini, R.; Gayathri, N.; Nalini, A.; Srinivas Bharath, M.M. Oxidative damage in muscular dystrophy correlates with the severity of the pathology: role of glutathione metabolism. Neurochem Res 2012, 37, 885–898. [Google Scholar] [CrossRef]
- Samanta, L.; Roy, A.; Chainy, G.B. Changes in rat testicular antioxidant defence profile as a function of age and its impairment by hexachlorocyclohexane during critical stages of maturation. Andrologia 1999, 31, 83–90. [Google Scholar] [CrossRef]
- Giusti, L.; Gabriele, M.; Penno, G.; Garofolo, M.; Longo, V.; Del Prato, S.; Lucchesi, D.; Pucci, L. A Fermented Whole Grain Prevents Lipopolysaccharides-Induced Dysfunction in Human Endothelial Progenitor Cells. Oxid Med Cell Longev 2017, 2017, 1026268. [Google Scholar] [CrossRef] [PubMed]
- Rammohan, A.; Zyryanov, G.V.; Bhagath, Y.B.; Manjula, K. Antioxidants: Structure-activity of plant polyphenolics. Vitam Horm 2023, 121, 395–411. [Google Scholar] [CrossRef]
- Stryjecka, M.; Krochmal-Marczak, B.; Cebulak, T.; Kiełtyka-Dadasiewicz, A. Assessment of Phenolic Acid Content and Antioxidant Properties of the Pulp of Five Pumpkin Species Cultivated in Southeastern Poland. International Journal of Molecular Sciences 2023, 24, 8621. [Google Scholar] [PubMed]
- Uchiyama, Y.; Suzuki, T.; Mochizuki, K.; Goda, T. Dietary supplementation with a low dose of (-)-epigallocatechin-3-gallate reduces pro-inflammatory responses in peripheral leukocytes of non-obese type 2 diabetic GK rats. J Nutr Sci Vitaminol (Tokyo) 2013, 59, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, Y.; Suzuki, T.; Mochizuki, K.; Goda, T. Dietary supplementation with (-)-epigallocatechin-3-gallate reduces inflammatory response in adipose tissue of non-obese type 2 diabetic Goto-Kakizaki (GK) rats. J Agric Food Chem 2013, 61, 11410–11417. [Google Scholar] [CrossRef]
- Elbling, L.; Weiss, R.M.; Teufelhofer, O.; Uhl, M.; Knasmueller, S.; Schulte-Hermann, R.; Berger, W.; Micksche, M. Green tea extract and (-)-epigallocatechin-3-gallate, the major tea catechin, exert oxidant but lack antioxidant activities. Faseb j 2005, 19, 807–809. [Google Scholar] [CrossRef]
- Ahangarpour, A.; Afshari, G.; Mard, S.A.; Khodadadi, A.; Hashemitabar, M. Preventive effects of procyanidin A2 on glucose homeostasis, pancreatic and duodenal homebox 1, and glucose transporter 2 gene expression disturbance induced by bisphenol A in male mice. J Physiol Pharmacol 2016, 67, 243–252. [Google Scholar]
- Wen, L.; Wu, D.; Tan, X.; Zhong, M.; Xing, J.; Li, W.; Li, D.; Cao, F. The Role of Catechins in Regulating Diabetes: An Update Review. Nutrients 2022, 14. [Google Scholar] [CrossRef]
- Madkhali, H.; Tarawneh, A.; Ali, Z.; Le, H.V.; Cutler, S.J.; Khan, I.A.; Shariat-Madar, Z. Identification of Human Kinin-Forming Enzyme Inhibitors from Medicinal Herbs. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Draganescu, D.; Andritoiu, C.; Hritcu, D.; Dodi, G.; Popa, M.I. Flaxseed Lignans and Polyphenols Enhanced Activity in Streptozotocin-Induced Diabetic Rats. Biology (Basel) 2021, 10. [Google Scholar] [CrossRef]
Figure 1.
Pathological ROS in diabetes.
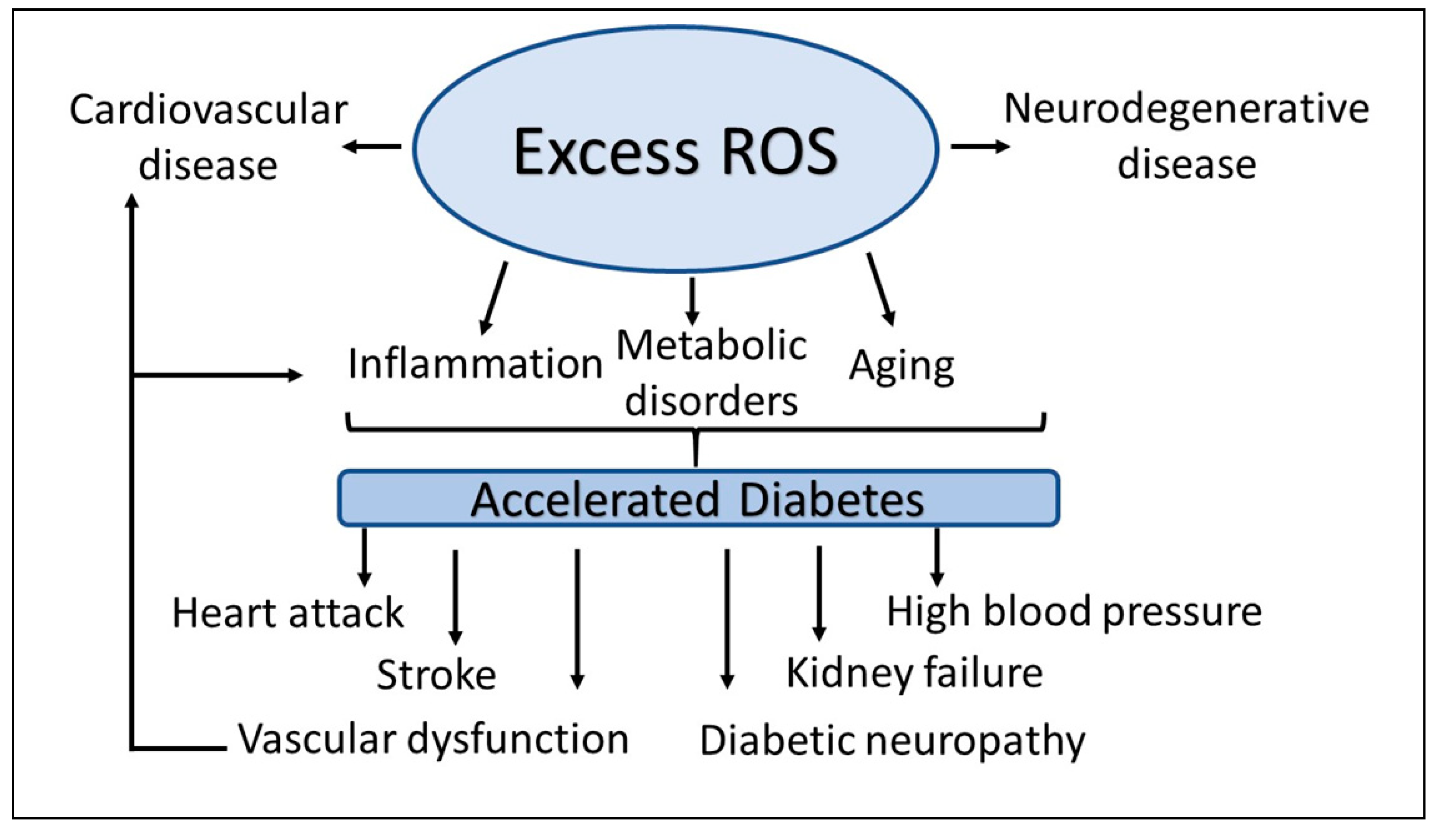
Figure 2.
Diabetes is associated with vascular dysfunction in multiple ways. Uncontrolled free radicals are the central components of cell injury. Antioxidants may reduce or prevent ROS/RNS-induced vascular complications.
Figure 2.
Diabetes is associated with vascular dysfunction in multiple ways. Uncontrolled free radicals are the central components of cell injury. Antioxidants may reduce or prevent ROS/RNS-induced vascular complications.
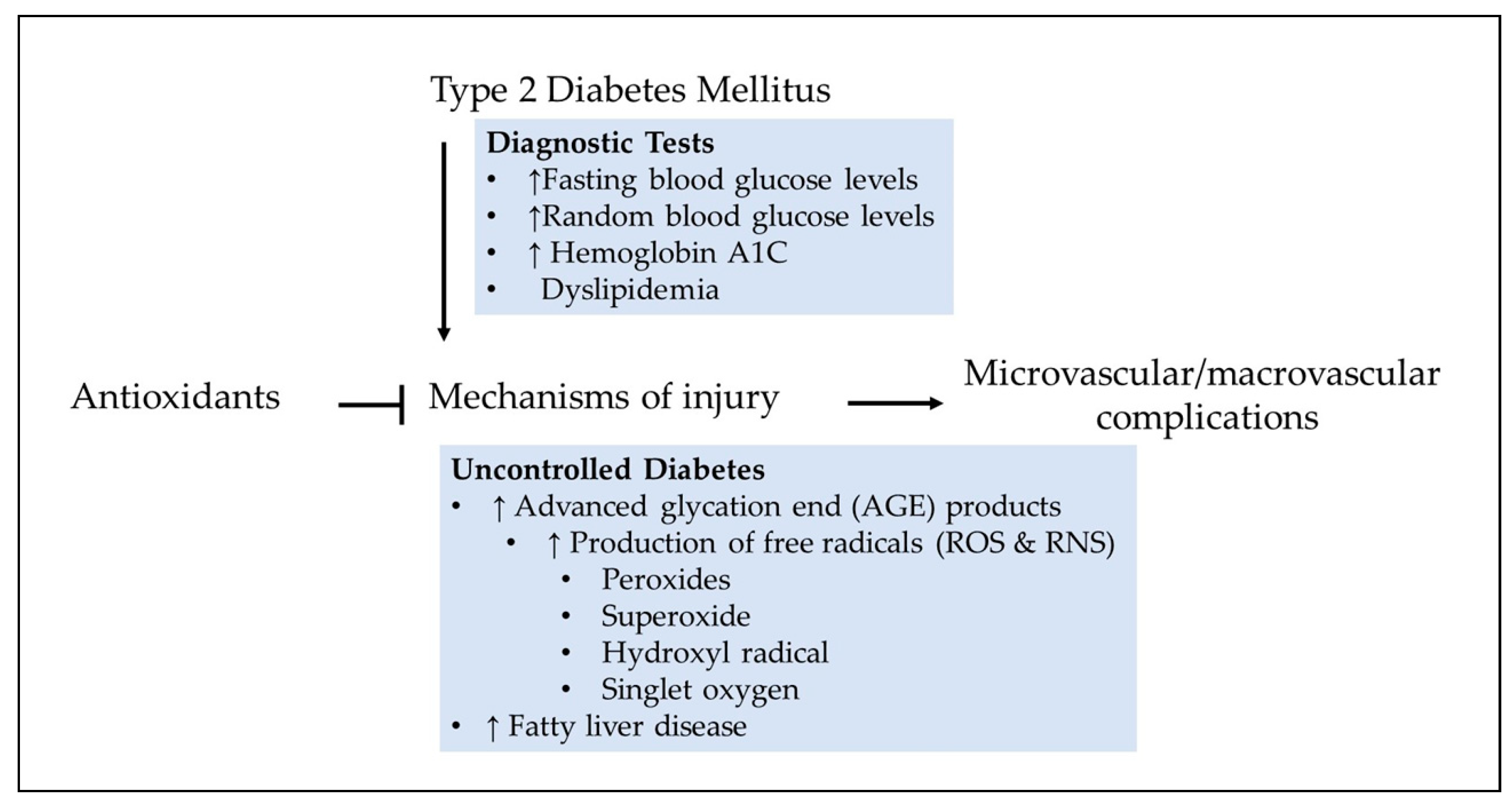
Figure 3.
Feed-forward (black arrows) and feed-back (red arrows) events where diabetes can contribute to ROS formation. Hyperglycemia, dyslipidemia, hypertension, kidney disease as well as inflammation lead to formation of ROS. As a consequence elevated levels of ROS might directly contribute to fibrosis, apoptosis, dedifferentiation, or indirectly via enhanced inflammation, reduced FFA β-oxidation, or NO bioavailability. Diabetes itself may feedback to foster generation of cytokines and vascular remodeling which also can contribute to generation of ROS. Under normal circumstances (non-diabetic response), ROS and RNS play vital roles in numerous biological processes. .
Figure 3.
Feed-forward (black arrows) and feed-back (red arrows) events where diabetes can contribute to ROS formation. Hyperglycemia, dyslipidemia, hypertension, kidney disease as well as inflammation lead to formation of ROS. As a consequence elevated levels of ROS might directly contribute to fibrosis, apoptosis, dedifferentiation, or indirectly via enhanced inflammation, reduced FFA β-oxidation, or NO bioavailability. Diabetes itself may feedback to foster generation of cytokines and vascular remodeling which also can contribute to generation of ROS. Under normal circumstances (non-diabetic response), ROS and RNS play vital roles in numerous biological processes. .
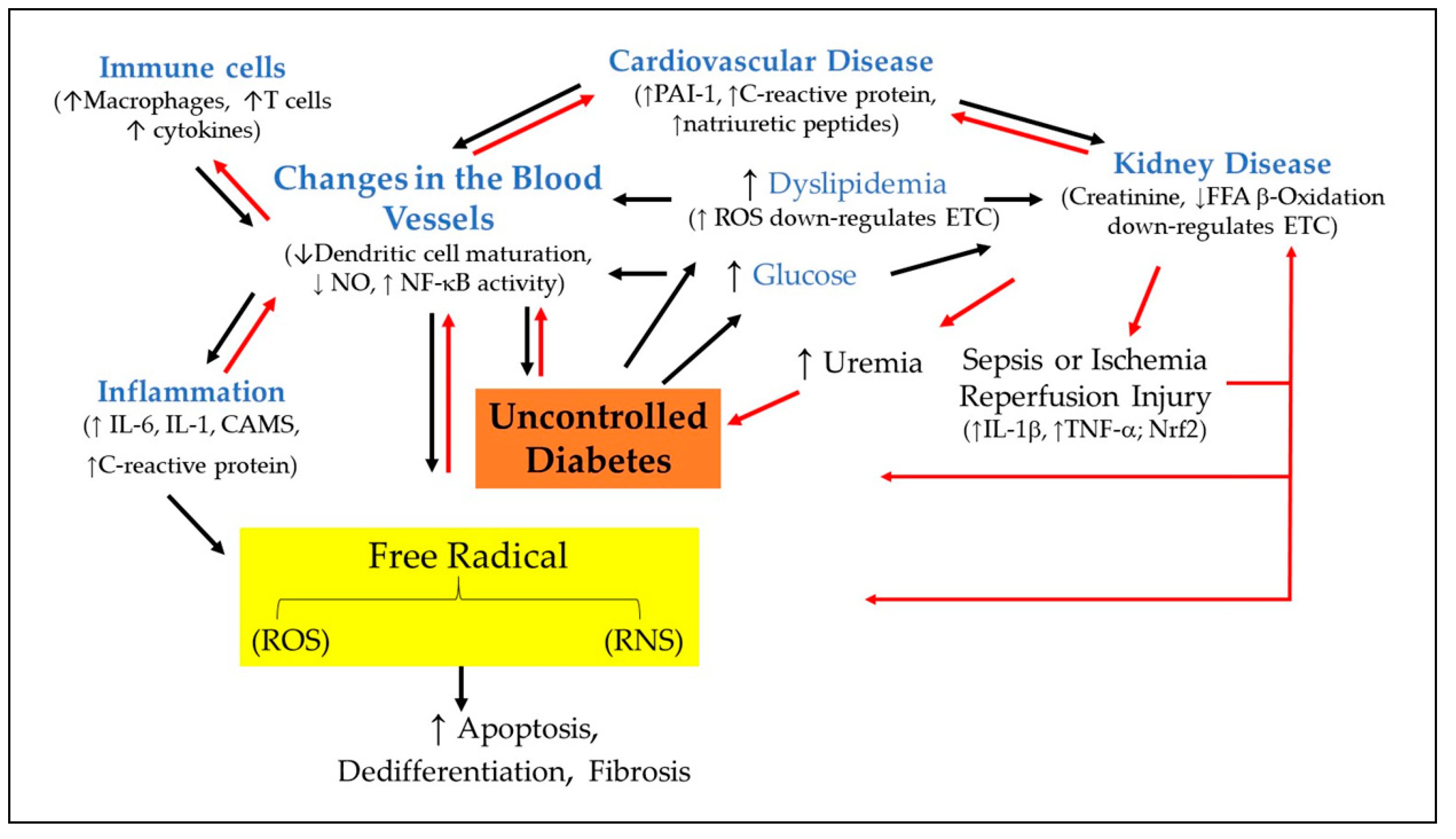
Figure 4.
An illustration depicting how the antioxidant properties of polyphenols were measured.
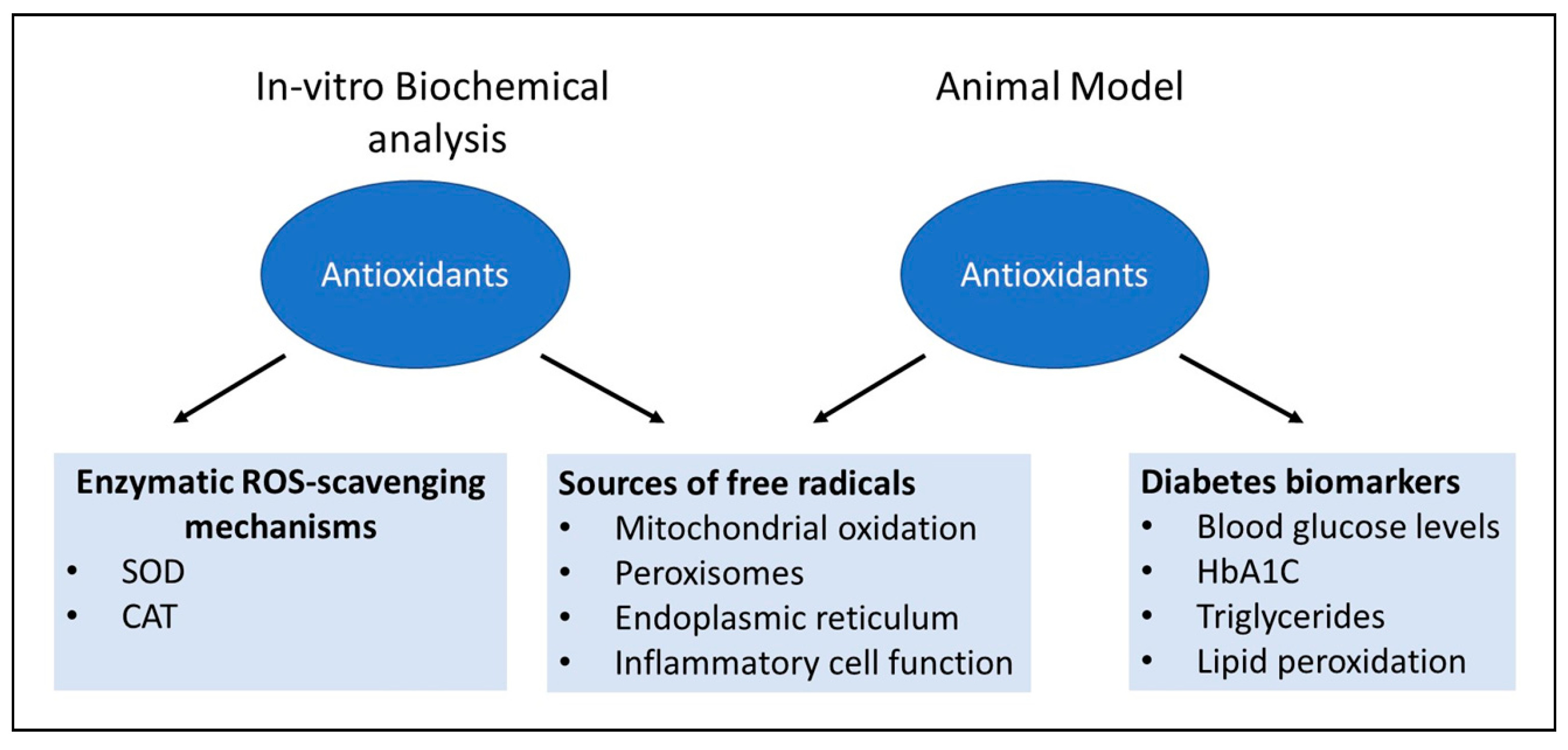

Table 1.
A summary of the effects of polyphenols in oxidative stress.
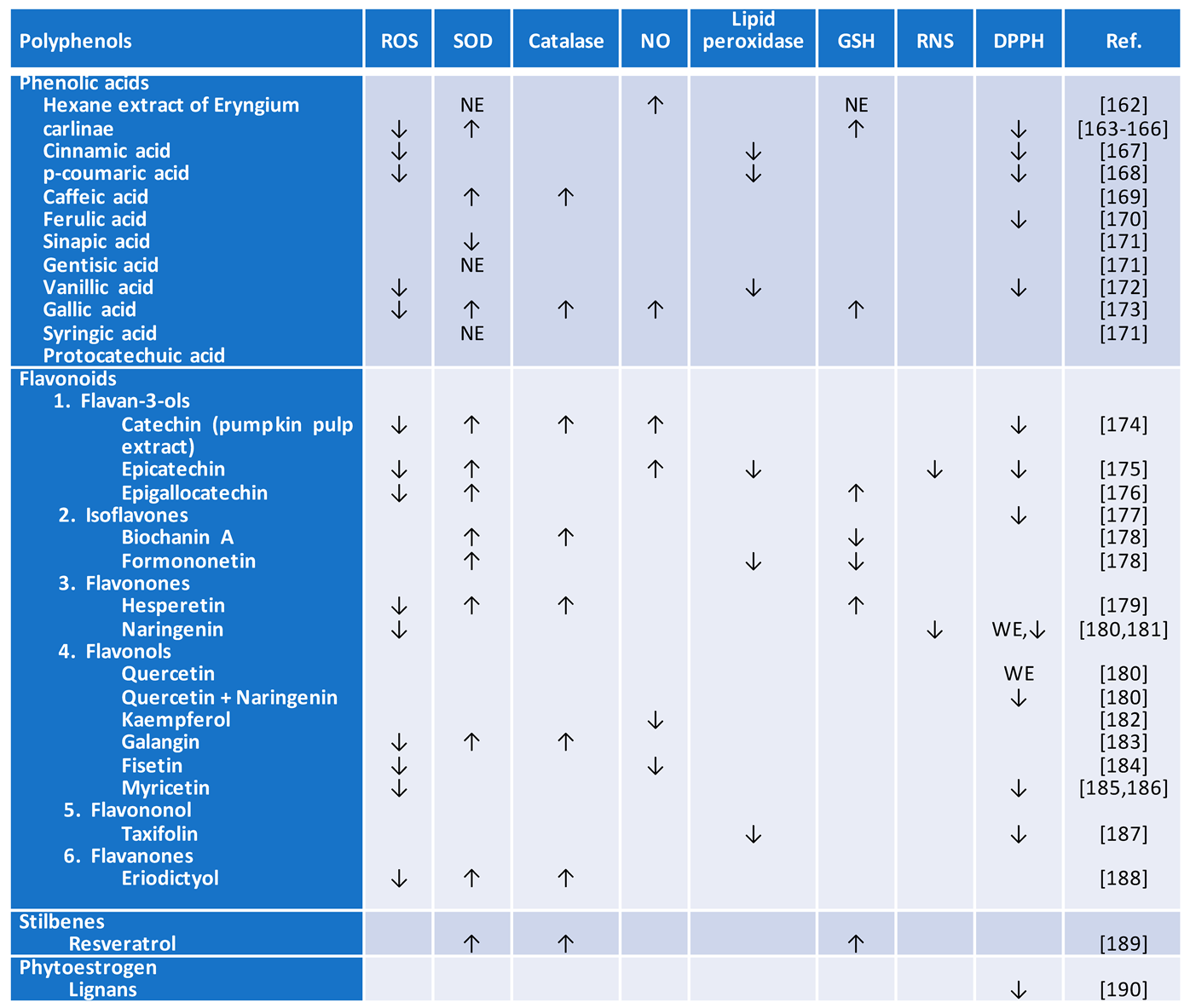
↑, represents to a significantly elevated level; ↓, refers to a significantly reduced level.
Table 2.
A summary of polyphenols-mediated protection against inflammation and associated oxidative stress.
Table 2.
A summary of polyphenols-mediated protection against inflammation and associated oxidative stress.
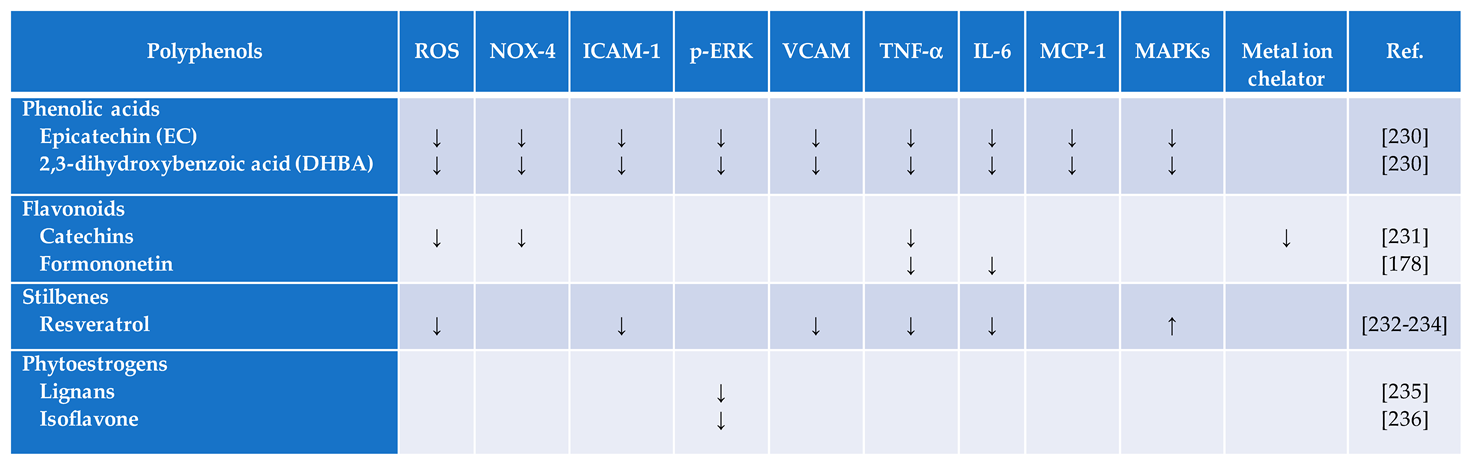
↑, represents to a significantly elevated level; ↓, refers to a significantly reduced level.
Table 3.
A summary of polyphenols on biomarkers of oxidative stress in diabetes.
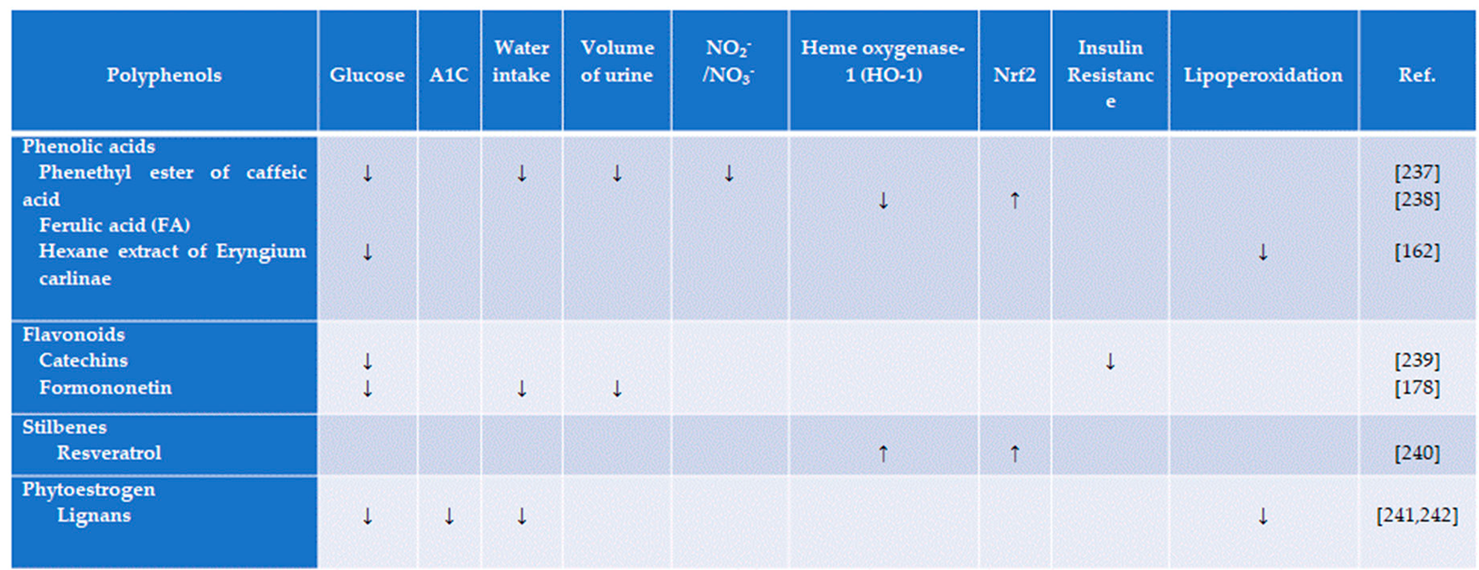
↑, represents to a significantly elevated level; ↓, refers to a significantly reduced level.
Table 4.
Summary of the acute effects of supplemental dietary antioxidant therapy for the prevention and treatment of diabetes in a double-blind placebo-controlled study.
Table 4.
Summary of the acute effects of supplemental dietary antioxidant therapy for the prevention and treatment of diabetes in a double-blind placebo-controlled study.
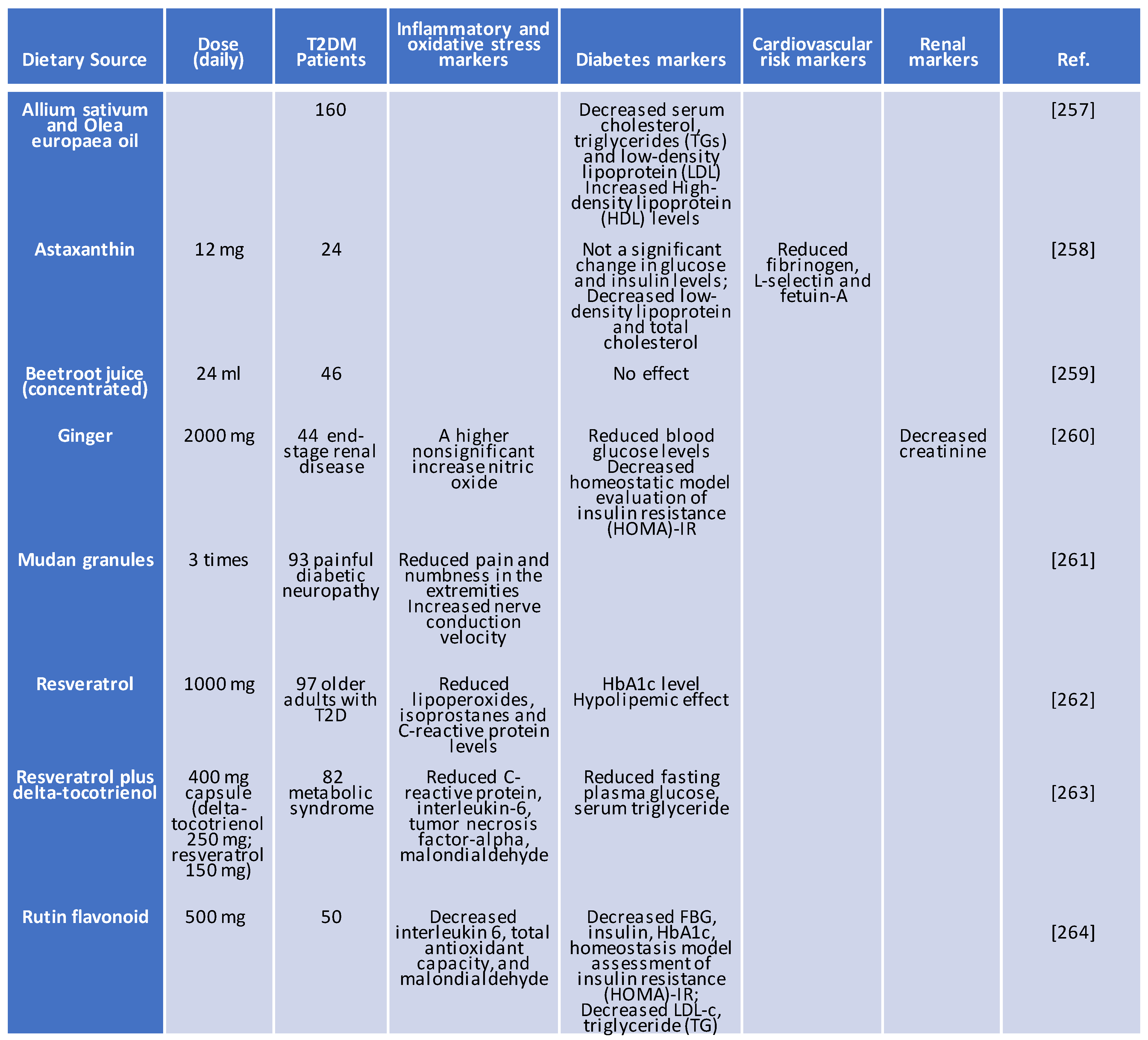
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



