
Submitted:
26 September 2023
Posted:
27 September 2023
You are already at the latest version
Abstract
Capsaicin, a lipophilic, volatile compound, is responsible for the pungent properties of chili peppers. In recent years, a significant increase in investigations into its properties has allowed the production of new formulations and the development of tools with biotechnological, diagnostic, and potential therapeutic applications. In this way, the possible clinical application of such compound is expanding every year. This opinion article aims to provide a synthesis of recent findings regarding the mechanisms by which capsaicin participates in the control of non-communicable diseases such as obesity, diabetes, and dyslipidemia. This opinion review also discusses the recent findings and clinical studies of the use of capsaicin in body weight control.
Keywords:
capsaicin
; pepper
; diabetes
; obesity
; inflammation
; gut microbiota
Capsaicinoids are a group of lipophilic and volatile compounds with different pungency intensities characterized by a vanilloid ring. They have diverse biological activities and applications, from food flavoring to therapeutic adjuncts 1. Among these, capsaicin (trans-8-methyl-N-vanylyl-6-nonenamide, C18H27NO3) is the most pungent and abundant, accounting for about 70% of all capsaicinoids, followed by dihydrocapsaicin (~22%), nordihydrocapsaicin (~7%) 2. The vanilloid ring has a high affinity for the Transient receptor potential vanilloid subtype 1 (TRPV1), a non-selective cationic channel widely distributed in different tissues. It is highly expressed in neural (peripheral and central) tissues, especially in C fibers, and, to a lesser extent, in the Aδ fibers of the nociceptive sensory pathway3. TRPV1 can also be found in other tissues, including adipose tissue 4, hepatocytes 5, immune system6, and endothelial cells 7. As a polymodal receptor, TRPV1 responds to a broad spectrum of physical and chemical stimuli such as heat, protons, and toxins. However, it exhibits affinity (in sub-μM order), sensitivity, and selectivity for capsaicin, its primary exogenous ligand 8. A better understanding of the TRPV1-capsaicin interaction at molecular levels has guided pharmaceutical efforts towards the use of capsaicin in the treatment of pain, as well as helping to explore new clinical uses 9. The biological activities of capsaicin include analgesic, anesthetic 10, anti-inflammatory 11, antioxidant 12, and thermogenic effects 13. These activities are related to capsaicin's effects in reducing adiposity, blood pressure, and blood glucose and cholesterol levels 13,14. Due to its pleiotropic action and high skin absorption, there is a growing interest in using capsaicin as a therapeutic alternative. Creams and patches with concentrations between 0.025 – 8% of capsaicin are commercially available for muscle or neuralgic pain management.
1. Metabolism and bioactivity
Since capsaicin is a food compound, the natural route of capsaicin absorption is the oral via. However, its pungency limits the quantity of capsaicin administration, either in natura (in food) or as powder and capsules. Moreover, lipophilicity directly influences the activation kinetics of its receptor TRPV1 and, consequently, its ability to generate action potentials in excitable cells 15. Thus, it becomes a challenge to overcome the capsaicin pungency maintaining its properties. In this sense, capsinoids or non-pungent synthetic analogs of capsaicin, such as olvanil and arvanil (that bind efficiently to TRPV1), could replace capsaicin in some effects 16,17.
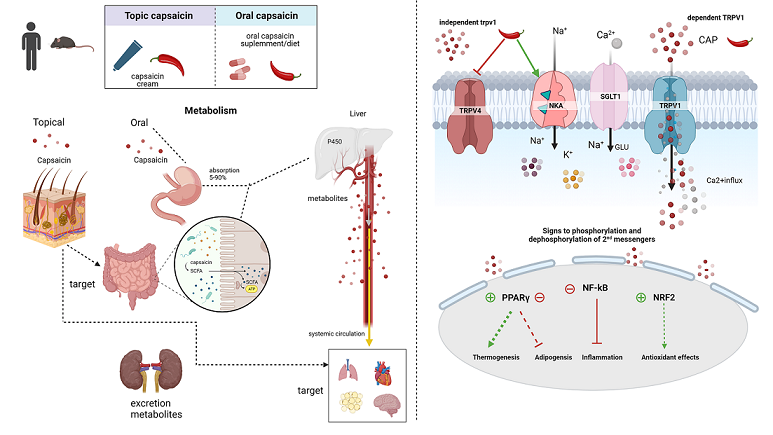
After oral ingestion, 50-90% of capsaicin is passively absorbed in the stomach, and its maximum concentration in blood is seen 1 hour later 18 (Figure 1). First-pass metabolism occurs in the liver, mediated by the cytochrome P450 system, which generates metabolites such as 16-hydroxycapsaicin, 17-hydroxycapsaicin, nordihydrocapsaicin, 16,17-dehydrocapsaicin, vanillin, vanilamine. In systemic circulation, capsaicin and its derived compounds are transported, linked to albumin, and distributed to organs and tissues 2.
When applied topically to the skin, capsaicin is rapidly absorbed and undergoes slow biotransformation, remaining mostly intact, with only a small part biotransformed into vanillin and vanillic acid. Unlike oral intake, topical applications prevent the hepatic metabolization of capsaicin, ensuring a greater bioavailability in target tissues 19. The half-life of capsaicin by topical is (1 to 24h), longer than oral intake, and its excretion is mainly by the renal via 2,19.
After topical application, the absorbed capsaicin activates its receptor TRPV1, which causes the rapid influx of sodium ions (Na +) and calcium (Ca 2+) from the extracellular environment to the cell interior. Then, a depolarization cascade is initiated and transmitted along sensory fibers from the spinal cord to the brain, causing pain sensation. A long-term refractory state follows the initial pain sensation, and the sensory neurons stop responding to an additional application of capsaicin in a process called "desensitization" 20,21 (Figure 1).
2. TRPV1-dependent mechanisms of action.
Capsaicin binds to the transmembrane segments of TRPV1 channels in a "tail up, head down" configuration and initiates calcium influx and desensitization of nerve fibers 8. In the case of chronic use of capsaicin, during the first few applications, it promotes the release of the preformed substance P, a potent local pain signal, which causes an initial state of neurogenic inflammation. The depletion of substance P and inoperability of nociceptive cells due to the constant influx of calcium occur after repeated applications, which prevents the formation of more mediators and reduces the inflammatory state 22. In summary, to minimize pain, capsaicin first causes hyperalgesia, which, by exhaustion, leads to desensitization and a state of analgesia 23.
The rapid ionic influx of Ca2+ caused by the TRPV1 activation generates activation of second messengers and modulation of several metabolic pathways. TRPV1 is also expressed in non-excitable cells, such as adipose tissue 4. In this tissue, capsaicin can exert anti-obesogenic and thermoregulatory effects via TRPV1 activation 24 by increasing thermogenic gene expression such as uncoupling protein 1 (UCP-1), Sirtuin 1 (SIRT-1) 25 and peroxisome proliferator-activated receptor γ (PPAR γ) coactivator -1α (PGC-1α). 26,27 These factors can interfere with lipid metabolism by suppressing inflammatory responses, increasing lipid oxidation, inhibiting adipogenesis, activating brown adipose tissue, and increasing satiety by interfering in the hypothalamic neuronal circuits 27,28.
The higher intracellular calcium concentration promoted by the Capsaicin-TRPV1 binding may increase the expression and activity of the endothelium-specific transcription factor (KLF2), which could increase the expression of the enzyme endothelial nitric oxide synthase (eNOS) and, consequently, the availability of nitric oxide (NO) (Figure 2). Moreover, capsaicin also induces thrombomodulin expression29 while decreasing the expression of inflammatory biomarkers such as IL-6, TNF, and CCL-2, associated with NF-κB inactivation 30. Phosphorylation of Akt is also described after capsaicin treatment, which results in disruption of the NRF2/ Keap complex and release of activated transcription factor NRF2 (Figure 2). This signaling triggers the transcription of heme-oxygenase1 genes, which are essential for heme degradation and prevention of oxidative damage 31. Other antioxidant enzymes, such as superoxide dismutase (SOD), catalase, glutathione peroxidase, as well as glutathione levels, may have their activities modified after oral treatment with Capsaicin 32. Figure 2 summarizes the main TRPV1-dependent effects of capsaicin.
3. TRPV1-independent mechanisms of action.
Although TRPV1 is the central mediator of capsaicin's effects, studies suggest this receptor is not its only target (Figure 3). The activations of TRPV1-dependent or independent pathways by capsaicin are related by the dose of capsaicin and the site where capsaicin is acting 21. For example, in the gastrointestinal tract, low concentrations of capsaicin (~1 μM) are reported to induce cell death by mechanisms involving a rapid and transient increase in TRPV1-dependent intracellular Ca 2+. However, higher concentrations of capsaicin (≥ 10 μM) induce cell death through TRPV1-independent mechanisms involving mitochondrial dysfunction and plasma membrane depolarization 27.
Despite extensive research, the exact mechanisms of non-neuronal and TRPV1-independent effects of capsaicin are poorly understood. Some non-specific and receptor-independent effect, such as the induction of changes in membrane fluidity, is also dependent on membrane lipid composition 35. Capsaicin can cause structural and physiological changes in organelles, such as mitochondria, by chemically affecting the structural properties of biological membranes. This occurs because the physicochemical modifications in this nanoenvironment can alter the membrane potential, and the ionic concentration and cause oxidative imbalance. These effects could culminate in apoptosis, as reported in pancreatic cancer cell lines 32.
The direct effects of capsaicin seem to be affected by the cell type, besides the dose and route of administration. Capsaicin acts not only through increases in Ca 2+ mediated by its binding to TRPV1 but also by modifying the availability and flux of other ions such as Na +, K +, and Ca 2+ itself for other voltage-gated ion channels, ionic transporters, and possibly interacting directly with other channels and receptors 36.
It is suggested that capsaicin induces impairment of hippocampal gamma oscillations through a TRPV1-independent pathway involving Na+/K+ ATPase (NKA). Thus, using capsaicin to reduce these oscillations to levels of healthy controls could become a promising strategy in the face of psychiatric and neurological disorders that present this dysfunction as a characteristic 21.
Another possible TRPV1-independent effect of capsaicin is its oncoprotective action. In glioblastoma and colon cancer cells, capsaicin was able to induce apoptosis by increasing PPARγ expression independently of its vanilloid receptor 37. Some of the TRPV1-independent effects of capsaicin may be related to its ability to activate or deactivate PPARs, particularly PPARγ 38. PPARγ is predominantly expressed in adipose tissue and, to a lesser extent, in other tissues such as skeletal muscle and liver. Capsaicin is described as a PPARγ activator or inhibitor. This effect in modulating PPARγ seems to be dependent on several factors, such as the balance between the PPAR's (which also include PPARα and PPARβ/δ) in the different metabolic states, the presence of TRPV1, the dose of capsaicin, and even the intestinal enterotype 39,40. Therefore, the possibility of directing the actions of capsaicin in PPARγ for therapeutic purposes and, at the same time, minimizing side effects represent major challenges 41.
Dietary capsaicin can attenuate Cl- secretion and stimulate Na+ absorption by blocking TRPV4 channels while stimulating NKA activity in intestinal epithelial cells (IEC). The intracellular Ca2+ stimulates Cl- secretion through the Ca2+-dependent Cl- secretion (CaCC) channel and apical cystic fibrosis transmembrane conductance regulator (CFTR). The blockade of TRPV4 caused by capsaicin decreases Ca2+ entry through this channel and reduces the stimulus for Cl- secretion. In contrast, stimulation of basolateral NKA activity would establish an ionic gradient and driving force used to promote apical Na+ absorption via Na+ -glucose transporter 1 (SGLT1) and the epithelial apical Na+ channel (ENaC) 42.
It is suggested that the protective effects of capsaicin against chemical carcinogens are mainly related to halogens metabolized by the cytochrome P450 (CYP450) enzyme system. Several mechanisms make capsaicin a promising agent against several types of cancer, as highlighted in a recent review 33. Capsaicin acts by inhibiting the activity of these enzymes, which prevents the metabolization of halogens into highly reactive species. Thus, the chemoprotective role of capsaicin has been mainly associated with the ability to modulate CYP enzymes 34.
Recent approaches are centered on capsaicin's ability to modulate the intestinal microbiota. Capsaicin can influence intestinal microbiota by a mechanism not totally understood. It is suggested that regular treatment with capsaicin increases diversity in the gut microbiota and abundance of short-chain fatty acid (SCFA) producing bacteria 43.
Moreover, capsaicin treatment prevents dysbiosis, gut barrier dysfunction, and low-grade chronic systemic inflammation 44 caused by dysbiosis. The ability of capsaicin to modulate the intestinal microbiota has been discussed not only as one of its TRPV1-independent effects but also as the basis for other beneficial systemic effects 45 on metabolic diseases 46 and cancer 47. Nonetheless, although this is a promising field of research, the role of capsaicin on intestinal microbiota is still controversial 48. Such studies were carried out primarily in experimental models; despite their promising effects, caution is needed when translating such effects into humans.
4. Anti-inflammatory and gastrointestinal effects of capsaicin
Capsaicin has been tested for preventing and treating intestinal pain and inflammation due to its functions in the gastrointestinal tract 49–51. Its actions on gastrointestinal sensory neurons may be dependent or independent of TRPV1 in mammals. Capsaicin triggers a painful and burning sensation in a TRPV1-dependent manner. On the other hand, as cited before, capsaicin 42appears to induce changes in anion secretions and induction of apoptosis of cancer cells by mechanisms independent of TRPV1 28,52. However, little is known about the actions of capsaicin in the gastrointestinal tract independently of TRPV1 42. Recent evidence has shown an important role for TRPV4 channels in the pathogenesis of experimental ulcerative colitis because these channels regulate ion transport in the intestinal epithelium 42. Capsaicin was able to inhibit Cl - secretion and promote Na + absorption by blocking TRPV4 channels. Moreover, the inactivation of TRPV4 channels by capsaicin in experimental colitis suppressed the overactivation of these channels that occurs in colitis 42.
Regarding anti-inflammatory effects, one of the main actions of capsaicin is the inhibition of the NFκ-B pathway that reduces proinflammatory cytokines (IL-6, TNF, and IL-1β) production and mRNA expression of the NLRP3 inflammasome, associated with a lower NFκB phosphorylation 53. In addition, the reduction in the junctional protein e-cadherin, typical in H. pylori infection, was attenuated after capsaicin treatment. It occurred as a consequence of reduced expression of the gastric cancer biomarkers miR21 and miR223, which downregulate the e-cadherin expression 53.
However, the effects of capsaicin on inflammation can be modulated by the dose administered. Xiang et al. 54 investigated the effects of oral administration of 40, 60, and 80 mg/kg weight of capsaicin for 7 days on gastrointestinal health and intestinal microbiota, using specific pathogen-free (SPF) mice. They showed that intakes above 60mg/kg seem harmful to the intestine. The administration of 80mg/kg caused inflammatory cell infiltration and loss of mucus-producing goblet cells in the colon. Such inflammation was characterized by elevated levels of inflammatory cytokines, especially TNF and IL-β1 and lower levels of the anti-inflammatory cytokine IL-10, suggesting damage to the jejunum and colon at such a dose. In addition, there was an increase in serum levels of neuropeptides (SP and CGRP) related to visceral pain, causing additional damage to the gastrointestinal system in a TRPV1-dependent manner 54. On the other hand, changes in the intestinal microbiota profile were observed when 40mg/kg was assessed. The abundance of Lactobacillus in the jejunum and ileum associated with capsaicin treatment was related to higher levels of IL-10, suggesting that the microbiota modulation by capsaicin may induce changes in the intestinal inflammatory environment.
In an experimental model of colorectal cancer, Cheng et al. 48 administered 300 mg/kg of capsaicin for 12 weeks to mice before the intraperitoneal injection of EGFP-labelled CT26 cells. Mice presented a proinflammatory microenvironment with increased cytokines IL-12, TNF, and IFN in the liver and IL-6 in the blood. In addition, there was an increase in neutrophils, macrophages, and monocyte infiltration associated with a reduction in the thickness of the intestinal mucus barrier, bacterial translocation, and destruction of the vascular epithelial barrier. The authors suggested that intake of high-dose CAP over the long term could increase the risk of metastasis in colorectal cancer 48. However, the results contradict other studies indicating capsaicin could suppress tumor cell growth 54–56. As discussed above, some effects of capsaicin are dose-dependent and may differ according to its concentration and time of use. For instance, there is no reason to recommend or avoid using capsaicin-rich foods in those with intestinal diseases 27,48,55–57.
5. Capsaicin and non-communicable chronic diseases
Capsaicin has been suggested as a potential alternative treatment for diabetes due to its influence on glucose metabolism 32. The in vitro study of Bort et al. 38 using C1C12-derived myotubes cultured with 100 and 200 µM of capsaicin found an increase in intracellular calcium induced by CAP-TRPV1 binding. It could stimulate intracellular signaling pathways through calcium-dependent protein kinase 2 or calmodulin (CAMKK2), which, in turn, phosphorylates AMP-activated protein kinase (AMPK), a regulatory kinase involved in glucose and lipid metabolisms. This action would improve insulin sensitivity due to increased fatty acid oxidation, insulin secretion, glucose uptake, and reduction of inflammatory factors 38.
Capsaicin was also related to increased glucose uptake in a TRPV1-independent, TRPV4-dependent manner, stimulating the co-transport of Na-glucose. In TRPV4 knockout mice, Na+ uptake was increased compared to the wild type, in which CAP was also shown to be able to potentiate the action of blocking TRPV4 and Na absorption via the Na-glucose cotransporter (SGLT1) 42,58.
In diabetic rats, oral administration of 0.5g/kg of capsaicin for 8 weeks reduced cholesterolemia, triglyceridemia, and cardiac fibrosis 59. Analysis of the heart showed increased expression of TRPV1 and eNOS associated with increased levels of nitric oxide and reduced reactive oxygen species (ROS). Similar results were obtained when vascular endothelial cells from these animals were treated with different concentrations of glucose and Capsaicin59.
Individuals with chronic diseases such as diabetes are more prone to vascular senescence, manifested by accelerated vascular stiffness and atherosclerosis. Senescent endothelial cells show loss of proliferation potential, decreased vasodilation, and proinflammatory and pro-arteriosclerotic phenotype 60. In this sense, the beneficial effects of capsaicin at different concentrations (0.3µM; 1.0µM, and 3.0µM) on the senescence of HUVEC cells exposed to 5 and 33 mM of glucose were studied. The findings showed a capsaicin-induced increase in sirtuin 1 (SIRT1), a protein that prevents endothelial senescence induced by intermittent hyperglycemia. In addition to increasing SIRT1 levels, capsaicin was able to suppress p21, a protein associated with ROS production and senescence. Thus, it was suggested that capsaicin could be a potential adjuvant in treating vascular aging in diabetes60.
Reports of the action of capsaicin in atherosclerosis development are numerous, mainly due to the improvement in lipid profile, inflammation, and endothelial dysfunction 60–64. Dai et al.61 studied the role of the microbiota in the anti-atherosclerotic effect of oral capsaicin (0.01% w/w) in ApoE knockout mice fed a high-fat diet (HFD) and demonstrated that the reduction of LDL-c and an increase in HDL-c was associated to reductions in atherogenesis shown by the reduction in the lesion area on aortic sinus and greater stability of atherosclerotic plaques. In the context of inflammation, capsaicin improved the HFD-induced inflammatory response in the intestinal mucosa, as seen by the reduction in inflammatory biomarkers IL-6 and LPS. In addition, capsaicin administration also altered the composition of the intestinal microbiota and metabolomic profiles, which could contribute to improving atherosclerosis. The microbiota's relevant role in capsaicin's action was confirmed when these effects disappeared in antibiotic-treated animals 61.
Regarding experimental studies addressed to body weight control, Song et al. 65 showed that diets with low (0.01%) or high (0.02%) capsaicin supplementation were not able to prevent body weight gain in Ob/Ob mice but inhibited increased fasting blood glucose and improved insulin sensitivity. Despite the absence of effects on body weight, capsaicin, in both concentrations, increased fecal levels of butyrate (short-chain fatty acid produced by microbiota fermentation) and blood GLP-1 while decreasing blood levels of ghrelin and inflammatory cytokines 65. Although both levels of supplementation failed to alter the α and β diversity of the intestinal microbiota, there was an increase in the Firmicutes/Bacteroidetes fila ratio. At the genus level, it was found an increase in the abundance of Roseburia, related to lower blood glucose levels, and a decrease in the abundance of Bacteroides and Parabacteroides associated with higher blood glucose 65.
Together, these data corroborate previous studies showing the influence of the microbiota-modulating action in the hypoglycemic and anti-inflammatory effects of Capsaicin 27,48,49,66. However, these preclinical studies need confirmation in long-lasting controlled trials.
6. Effect of Capsaicin on metabolic syndrome and obesity
Many studies with capsaicin have been done in rodents, and the result transposition to humans is not always possible, including dose matching for humans. In this way, Szallasi et al. 67 estimated the correspondence of dietary capsaicin intake from rats to humans. They concluded that a dose of 50 mg/kg of supplemented capsaicin for rats (about 250g of body weight) would be equivalent to 12.5 mg/day for humans, an intake higher than the average consumption in Korea, about 2.17 mg of Capsaicin 68. Furthermore, the 0.01% capsaicin supplementation in diet, used in several experimental models, is also above the average intake in most countries that consume spicy meals. Thus, studies on realistic amounts of capsaicin supplementation or foods rich in capsaicinoids should be conducted. Moreover, these studies should consider individual differences, such as the presence or absence of obesity and its metabolic consequences.
The described beneficial properties of capsaicin occur due to a wide variety of properties, which goes beyond the metabolism and expands its field of use from packaging conservation to effects on human health 69,70.
The metabolic actions of capsaicin, mainly related to antioxidant, anti-inflammatory, and thermogenic properties, make capsaicin a promising adjuvant in treating and preventing metabolic syndrome, obesity, and associated comorbidities such as dyslipidemia and diabetes mellitus 70–73.
Although several reviews have been published in recent years about the effect of capsaicin, capsinoids, and spicy foods on obesity 24,33,67,74,75, we are far from the confirmatory answer if this action occurs in the general as well as in the affected population, what the magnitude of this action, if it is safe, and what is the optimal therapeutic dose and treatment duration.
A previous scope review analyzed publications about capsaicin receptors signaling for its antiobesity properties 76 The analysis showed that, besides TRPV1, capsaicin can also act as an agonist for PPARs, especially PPARγ on fat metabolism 38,77. Given the role of PPARγ in the transcriptional regulation of adipogenesis, this would be one of the potential pathways for the antiobesity action of Caps. Activation of PPARγ by capsaicin has also been associated with its beneficial action on obesity, reducing inflammatory markers and inducing the expression and synthesis of adiponectin in mouse adipocytes 76.
Thus, capsaicin signaling in obesity and fat metabolism is mainly described as TRPV1 dependent 25,78–81, although some studies presented TRPV1-independent actions, including activation of PPARγ 4,79,82,83. In fact, independently of capsaicin signaling, several mechanisms mediate the action of capsaicin in weight loss, mostly linked to its ability to increase thermogenesis, induce satiety, and modify the obesogenic microbiota.
Capsaicin, through its binding to albumin, reaches the adrenal gland, which induces the release of catecholamines. It has been shown that capsaicin can increase lipolysis in adipocytes and stimulate fat oxidation by activating the sympathetic nervous system 84. This situation may affect tissues such as brown and white adipose tissue involved in body weight control. In white adipose tissue, capsaicin can exert several actions, increasing lipolysis and reducing lipogenesis 85,86 and, thus, reducing adipocyte volume and, consequently, adiposity 87. In brown adipose tissue, capsaicin activates browning by increasing UCP1 expression, mitochondrial biogenesis, energy expenditure, and glycerol recycling 86,88,89.
Studies in animal models have suggested that capsaicin may increase satietogenic hormones such as glucagon-like protein 1 (GLP-1) and the gastric inhibitory peptide (GIP) 90. The intense concentration of TRPV1 in the nervous system, mainly in the hypothalamus, supports the possible relationship between capsaicin and changes in hunger/satiety control. However, the role of capsaicin in increasing these incretins in clinical studies is still inconsistent. In this way, some reviews assessed the effect of capsaicin and spicy food on cognition, food preferences, and satiety induction 24,74,91–93. Although an increase in GLP-1 was seen after a capsaicin-supplemented meal 94, a more recent study using intraduodenal infusion of capsaicin in volunteers without obesity did not observe an increase in plasma concentrations of GLP-1 and PYY 95.
Rigamonti et al. 96 assessed the acute effect of capsaicin on appetite-regulating gastrointestinal peptides on the energy balance of young subjects with obesity. They showed that capsaicin (2mg), provided during an "ad libitum" dinner, did not change circulating levels of ghrelin, GLP-1, peptide YY, or satiety after food. Nonetheless, the pre-post meal difference in rest metabolism was higher in those receiving capsaicin. The authors suggest capsaicin could act as a metabolic activator rather than a hypophagic inducer96. In another study, supplementation for 8 weeks with 12 mg/day of capsiate, a non-pungent analog of capsaicin, increased body weight by 1 kg. However, this increase was due to increased upper body strength compared to placebo 97.
It is interesting to note that the effect of capsaicin may change between individuals, depending on body weight. It was the result of a meta-analysis that found that after ingestion of capsaicin or capsinoids, there is a modest increase in energy expenditure of around 58kcal/day and a reduction in the respiratory quotient, suggesting increased lipid oxidation in individuals with BMI >25kg/m2 98. In those with BMI<25kg/m2, capsaicin or capsinoids did not affect energy expenditure or respiratory quotient.
However, these effects in favoring healthy gut microbiota have been demonstrated in experimental animals 99–102 and in vitro studies 43,103, with the literature lacking in clinical studies. In this regard, clinical studies should explore the role of the microbiota on the capsaicin effect and its impact on metabolic conditions.
One of the factors involved in chronic low-grade inflammation linked to obesity is metabolic endotoxemia, resulting in intestinal dysbiosis. Kang et al. (Kang et al. 2017a) demonstrated that mice fed a high-fat diet supplemented with capsaicin exhibited lower levels of metabolic endotoxemia and chronic inflammation associated with lower body weight gain. Other findings in the gastrointestinal tract were increased abundance of butyrate-producing Ruminococcaceae and Lachnospiraceae, and low levels of S24-7 lipopolysaccharide (LPS)-producing bacteria and inhibition of type 1 cannabinoid receptors (CB1). Interestingly, when the microbiota from the capsaicin-treated animals was transferred to germ-free mice fed a high-fat diet, the recipient mice maintained the protective effects. Furthermore, when microbiota depletion was induced by an antibiotic cocktail, there was a loss of capsaicin protection against obesity 104.
However, Manca et al.105, in an exploratory clinical study with women presenting overweight and obesity, showed that the oral administration of capsicum extract capsules for 12 weeks produced a modest increase in the relative abundance of the genus Flavonifractor. The rise in Flavonifractor genus may be related to its ability to metabolize flavonoids such as curcumin, with which capsaicin has structural homology. Species belonging to this genus, such as F. plautii, have been involved in reducing inflammation 106.
Xia et al. 59 cultivated human fecal microbiota from healthy voluntaries with red pepper (2%) for 24h and found an increase in acetate and propionate concentrations compared with control cultures. Subdoligranulum spp.-, Blautia spp.-, Faecalibacterium prausnitzii-, P. vulgatus-, and Prevotella copri-like bacteria were defined as red pepper-responsive indigenous gut bacteria. The authors concluded that red pepper increases the short-chain fatty acid-producing bacteria and other beneficial bacteria in human fecal cultures. A similar result was seen in the study of Mahalak et al. 43, showing that capsaicin increases the diversity of human microbiota and SCFA abundance. These studies could explain the beneficial health effects of capsaicin in obesity and other non-communicable chronic diseases. The influence of capsaicin on glucose homeostasis and obesity by modulation of the gut microbiota is summarized in Figure 4.
Gupta et al. 72, in a review of the benefits of capsiate, concluded that it shares several properties with capsaicin, including increased satiety and reduced body weight. Likewise, Zsiborás et al.98, in their meta-analysis, concluded that the metabolic effects not only of capsaicin but also of capsiate are significant in individuals with high BMI. However, in the same year, Szallasi et al. 67, analyzing clinical studies carried out with capsiate, did not observe a significant influence of non-pungent capsinoids on body weight and obesity.
Some reviews and clinical studies carried out with red 72,85,107 or black pepper 108,109 have observed beneficial effects on human health. Since capsaicin and chili peppers are the main components that give spicy characteristics to foods, it could be expected to associate the increase in spicy food consumption with the increase in capsaicin's beneficial effects. In this way, Yang et al. 110 examined the association between the frequency of spicy food intake and abdominal obesity in a Chinese population in 40,877 individuals. Interestingly, the results showed that the higher consumption of spicy foods was associated with increased abdominal adiposity. However, other factors besides capsaicin content, such as total caloric and nutrient intakes, physical activity, and comorbidities, among others, could have contributed to this result.
Capsiate, dihydrocapsiate, and nordihydrocapsiate belong to the family capsinoids that possess molecular structure similar to capsaicin. Like capsaicin, capsinoids bind TRPV1 in the intestinal epithelium. However, they do not activate TRPV1 in the oral cavity. Hereafter, they can induce their biological effects without producing pungency. In this way, capsinoids such as capsiate could be, as occurs with capsaicin, a promising agent for treating obesity.
Similar results were found in a meta-analysis to evaluate the association between consumption of spicy foods and overweight/obesity, high blood pressure, and blood lipid profile 68. The results showed that greater consumption of spicy foods was associated with an increased risk of overweight/obesity, an increased LDL-c, and a modest reduction in HDL-c. However, consumption of spicy foods was negatively correlated with hypertension. Another study 111 analyzed the profile of spicy food consumption in a Chinese population. The results showed that the more frequent consumption and intense pungency of spicy foods were positively correlated with a preference for deep-fried food, salty snakes, alcohol and tea drinking, and tobacco smoking. These results highlight the importance of distinguishing between isolated capsaicin and pepper-containing foods' effects.
In addition to capsaicinoids and capsinoids, peppers are sources of nutrients and other bioactive components such as flavonoids, phenolic compounds, carotenoids, and ascorbic acid. These compounds aggregate and enhance several properties in peppers beyond capsaicin effects 93,112.
Other issues, such as the cultural consumption of pepper and the genetic characteristics of the studied populations, can influence the results. In this sense, clinical studies in populations from countries with the highest levels of per capita pepper consumption such as Bulgaria (21 mg/day), Singapore (14 mg/day) and Vietnam (0.4 mg/day) as well as studies that consider different genetic characteristics, such as the presence of polymorphisms or different proteoforms of vanilloid receptors, can provide important findings and a better understanding of the role of capsaicin in health and diseases.
7. Adverse effects
Capsaicin is not free from adverse effects, especially when given topically. The most common effects, considering all routes of administration, are burning sensation, pruritus, edema at the administration site, and pain 113. Muscle aches, chills, nasopharyngitis, sinusitis, bronchitis, cough and dyspnoea, fever, tachycardia, dizziness and headache, hypertension, nausea, and vomiting are also described, although less common. Rarer complications include dysgeusia, hypoesthesia, peripheral edema, peripheral sensory neuropathy, and throat irritation 114. Nonetheless, most effects may disappear during treatment.
8. Conclusions
Studies using capsaicinoids and capsinoids as adjuvants in obesity, diabetes, dyslipidemia, or cancer therapy are increasing in the literature. Although capsaicin is already approved as a topical treatment for neuropathic pain, its use in non-communicable diseases has not been properly and extensively studied to permit us an incontestable conclusion. Most of these studies show beneficial properties, improving antioxidant and anti-inflammatory status, inducing thermogenesis, and reducing white adipose tissue. Other mechanisms are also described, including reducing food intake and reduction of intestinal dysbiosis. Although most of these studies lead us to the benefits of capsaicin supplementation, few studies were done over 12 weeks in a population affected by these diseases. Other important issues that need to be solved before capsaicin use include the optimal dose and the best capsaicin administration schedule that brings efficiency and low adverse effects.
Furthermore, non-pungent capsinoids or synthetic analogs can conveniently substitute capsaicin. However, meta-analyses, although mostly showing beneficial effects in obesity, diabetes, dyslipidemia, and cancer, still lack confirmation. Therefore, we believe we are closer but not ready to include capsaicin as an adjuvant in treating non-communicable chronic diseases.
Funding
This research received no external funding.
Acknowledgments
The authors thank the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for research fellowship of JIAL and the "Fundação Coordenação de Aperfeiçoamento de Pessoal de Nível Superior" for the Ph.D scholarship of JLS.
Conflicts of Interest
"The authors declare no conflict of interest.
References
- Rogers, J.; Urbina, S.L.; Taylor, L.W.; Wilborn, C.D.; Purpura, M.; Jäger, R.; Juturu, V. Capsaicinoids supplementation decreases percent body fat and fat mass: adjustment using covariates in a post hoc analysis. BMC Obes. 2018, 5, 22. [Google Scholar] [CrossRef]
- Chanda, S.; Bashir, M.; Babbar, S.; Koganti, A.; Bley, K. In Vitro Hepatic and Skin Metabolism of Capsaicin. Drug Metab. Dispos. 2008, 36, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Stienstra, R.; Saudale, F.; Duval, C.; Keshtkar, S.; Groener, J.E.M.; van Rooijen, N.; Staels, B.; Kersten, S.; Müller, M. Kupffer cells promote hepatic steatosis via interleukin-1β-dependent suppression of peroxisome proliferator-activated receptor α activity. Hepatology 2009, 51, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Kida, R.; Yoshida, H.; Murakami, M.; Shirai, M.; Hashimoto, O.; Kawada, T.; Matsui, T.; Funaba, M. Direct action of capsaicin in brown adipogenesis and activation of brown adipocytes. Cell Biochem. Funct. 2016, 34, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Yang, G.; Sun, T.; Tao, J.; Zhu, C.; Yu, H.; Cheng, Y.; Yang, Z.; Xu, M.; Jiang, Y.; et al. Capsaicin receptor TRPV1 maintains quiescence of hepatic stellate cells in the liver via recruitment of SARM1. J. Hepatol. 2023, 78, 805–819. [Google Scholar] [CrossRef]
- Bertin, S.; Aoki-Nonaka, Y.; de Jong, P.R.; Nohara, L.L.; Xu, H.; Stanwood, S.R.; Srikanth, S.; Lee, J.; To, K.; Abramson, L.; et al. The ion channel TRPV1 regulates the activation and proinflammatory properties of CD4+ T cells. Nat. Immunol. 2014, 15, 1055–1063. [Google Scholar] [CrossRef]
- Negri, S.; Faris, P.; Rosti, V.; Antognazza, M.R.; Lodola, F.; Moccia, F. Endothelial TRPV1 as an emerging molecular target to promote therapeutic angiogenesis. Cells 2020, 9, 1341. [Google Scholar] [CrossRef]
- Yang, F.; Xiao, X.; Cheng, W.; Yang, W.; Yu, P.; Song, Z.; Yarov-Yarovoy, V.; Zheng, J. Structural mechanism underlying capsaicin binding and activation of the TRPV1 ion channel. Nat. Chem. Biol. 2015, 11, 518–524. [Google Scholar] [CrossRef]
- Forstenpointner, J.; Naleschinski, D.; Wasner, G.; Hüllemann, P.; Binder, A.; Baron, R. Sensitized vasoactive C-nociceptors: key fibers in peripheral neuropathic pain. PAIN Rep. 2019, 4, e709. [Google Scholar] [CrossRef]
- Irving, G.A.; Backonja, M.; Rauck, R.; Webster, L.R.; Tobias, J.K.; Vanhove, G.F. NGX-4010, a Capsaicin 8% Dermal Patch, Administered Alone or in Combination With Systemic Neuropathic Pain Medications, Reduces Pain in Patients With Postherpetic Neuralgia. Clin. J. Pain 2012, 28, 101–107. [Google Scholar] [CrossRef]
- Huang, C.-J.; Pu, C.-M.; Su, S.-Y.; Lo, S.-L.; Lee, C.H.; Yen, Y.-H. Improvement of wound healing by capsaicin through suppression of the inflammatory response and amelioration of the repair process. Mol. Med. Rep. 2023, 28, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G. E. S. et al. Biological Properties, Bioactive Constituents, and Pharmacokinetics of Some Capsicum spp. and Capsaicinoids. Int. J. Mol. Sci. 21, 1–35 (2020).
- Thongin, S.; Den-Udom, T.; Uppakara, K.; Sriwantana, T.; Sibmooh, N.; Laolob, T.; Boonthip, C.; Wichai, U.; Muta, K.; Ketsawatsomkron, P. Beneficial effects of capsaicin and dihydrocapsaicin on endothelial inflammation, nitric oxide production and antioxidant activity. Biomed. Pharmacother. 2022, 154, 113521. [Google Scholar] [CrossRef] [PubMed]
- Nawaka, N.; Wanmasae, S.; Makarasen, A.; Dechtrirat, D.; Techasakul, S.; Jeenduang, N. Allicin and Capsaicin Ameliorated Hypercholesterolemia by Upregulating LDLR and Downregulating PCSK9 Expression in HepG2 Cells. Int. J. Mol. Sci. 2022, 23, 14299. [Google Scholar] [CrossRef] [PubMed]
- Ursu, D.; Knopp, K.; Beattie, R.E.; Liu, B.; Sher, E. Pungency of TRPV1 agonists is directly correlated with kinetics of receptor activation and lipophilicity. Eur. J. Pharmacol. 2010, 641, 114–122. [Google Scholar] [CrossRef]
- Lim, S.G.; Seo, S.E.; Jo, S.; Kim, K.H.; Kim, L.; Kwon, O.S. Highly Efficient Real-Time TRPV1 Screening Methodology for Effective Drug Candidates. ACS Omega 2022, 7, 36441–36447. [Google Scholar] [CrossRef]
- Marzęda, P.; Wróblewska-Łuczka, P.; Florek-Łuszczki, M.; Drozd, M.; Góralczyk, A.; Łuszczki, J.J. Comparison of the Anticancer Effects of Arvanil and Olvanil When Combined with Cisplatin and Mitoxantrone in Various Melanoma Cell Lines—An Isobolographic Analysis. Int. J. Mol. Sci. 2022, 23, 14192. [Google Scholar] [CrossRef]
- Suresh, D.; Srinivasan, K. Tissue distribution & elimination of capsaicin, piperine & curcumin following oral intake in rats. . 2010, 131. [Google Scholar]
- Christensen, J.D.; Vecchio, S.L.; Andersen, H.H.; Elberling, J.; Arendt-Nielsen, L. Effect of Topical Analgesia on Desensitization Following 8% Topical Capsaicin Application. J. Pain 2021, 22, 778–788. [Google Scholar] [CrossRef]
- van Neerven, S. G. A. & Mouraux, A. Capsaicin-Induced Skin Desensitization Differentially Affects A-Delta and C-Fiber-Mediated Heat Sensitivity. Front. Pharmacol. 11, (2020).
- Balleza-Tapia, H. , Dolz-Gaiton, P., Andrade-Talavera, Y. & Fisahn, A. Capsaicin-Induced Impairment of Functional Network Dynamics in Mouse Hippocampus via a TrpV1 Receptor-Independent Pathway: Putative Involvement of Na+/K+-ATPase. Mol. Neurobiol. 57, 1170–1185 (2020).
- Lawrence, G. W. , Zurawski, T. H., Dong, X. & Oliver Dolly, J. Population Coding of Capsaicin Concentration by Sensory Neurons Revealed Using Ca2+ Imaging of Dorsal Root Ganglia Explants from Adult pirt-GCaMP3 Mouse. Cell. Physiol. Biochem. 55, 428–448 (2021).
- Arora, V.; Campbell, J.N.; Chung, M.-K. Fight fire with fire: Neurobiology of capsaicin-induced analgesia for chronic pain. Pharmacol. Ther. 2020, 220, 107743–107743. [Google Scholar] [CrossRef]
- Irandoost, P.; Yagin, N.L.; Namazi, N.; Keshtkar, A.; Farsi, F.; Alamdari, N.M.; Vafa, M. The effect of Capsaicinoids or Capsinoids in red pepper on thermogenesis in healthy adults: A systematic review and meta-analysis. Phytotherapy Res. 2020, 35, 1358–1377. [Google Scholar] [CrossRef]
- Baskaran, P.; Krishnan, V.; Ren, J.; Thyagarajan, B. Capsaicin induces browning of white adipose tissue and counters obesity by activating TRPV1 channel-dependent mechanisms. Br. J. Pharmacol. 2016, 173, 2369–2389. [Google Scholar] [CrossRef]
- Ghorbanpour, A.; Salari, S.; Baluchnejadmojarad, T.; Roghani, M. Capsaicin protects against septic acute liver injury by attenuation of apoptosis and mitochondrial dysfunction. Heliyon 2023, 9, e14205. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Xu, X.; Zhang, T.; Wu, X.; Fan, D.; Hu, Y.; Ding, J.; Yang, X.; Lou, J.; Du, Q.; et al. Beneficial effects of dietary capsaicin in gastrointestinal health and disease. Exp. Cell Res. 2022, 417, 113227. [Google Scholar] [CrossRef] [PubMed]
- Bouyer, P.G.; Tang, X.; Weber, C.R.; Shen, L.; Turner, J.R.; Matthews, J.B.; Obi, S.; Nakajima, T.; Hasegawa, T.; Kikuchi, H.; et al. Capsaicin induces NKCC1 internalization and inhibits chloride secretion in colonic epithelial cells independently of TRPV1. Am. J. Physiol. Liver Physiol. 2013, 304, G142–G156. [Google Scholar] [CrossRef] [PubMed]
- Ching, L.-C.; Kou, Y.R.; Shyue, S.-K.; Su, K.-H.; Wei, J.; Cheng, L.-C.; Yu, Y.-B.; Pan, C.-C.; Lee, T.-S. Molecular mechanisms of activation of endothelial nitric oxide synthase mediated by transient receptor potential vanilloid type 1. Cardiovasc. Res. 2011, 91, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.-H.; Kim, C.-S.; Han, I.-S.; Kawada, T.; Yu, R. Capsaicin, a spicy component of hot peppers, modulates adipokine gene expression and protein release from obese-mouse adipose tissues and isolated adipocytes, and suppresses the inflammatory responses of adipose tissue macrophages. FEBS Lett. 2007, 581, 4389–4396. [Google Scholar] [CrossRef]
- Joung, E.-J.; Li, M.-H.; Lee, H.G.; Somparn, N.; Jung, Y.S.; Na, H.-K.; Kim, S.-H.; Cha, Y.-N.; Surh, Y.-J.; Jang, H.J.; et al. Capsaicin Induces Heme Oxygenase-1 Expression in HepG2 CellsViaActivation of PI3K-Nrf2 Signaling: NAD(P)H:Quinone Oxidoreductase as a Potential Target. Antioxidants Redox Signal. 2007, 9, 2087–2098. [Google Scholar] [CrossRef]
- Pramanik, K.C.; Boreddy, S.R.; Srivastava, S.K. Role of Mitochondrial Electron Transport Chain Complexes in Capsaicin Mediated Oxidative Stress Leading to Apoptosis in Pancreatic Cancer Cells. PLOS ONE 2011, 6, e20151. [Google Scholar] [CrossRef]
- Adetunji, T.L.; Olawale, F.; Olisah, C.; Adetunji, A.E.; Aremu, A.O. Capsaicin: A Two-Decade Systematic Review of Global Research Output and Recent Advances Against Human Cancer. Front. Oncol. 2022, 12, 908487. [Google Scholar] [CrossRef]
- Babbar, S. , Chanda, S. & Bley, K. Inhibition and induction of human cytochrome P450 enzymes in vitro by capsaicin. Xenobiotica. 40, 807–816 (2010).
- Sharma, N.; Phan, H.T.T.; Yoda, T.; Shimokawa, N.; Vestergaard, M.C.; Takagi, M. Effects of Capsaicin on Biomimetic Membranes. Biomimetics 2019, 4, 17. [Google Scholar] [CrossRef]
- Isaev, D.; Yang, K.-H.S.; Shabbir, W.; Howarth, F.C.; Oz, M. Capsaicin Inhibits Multiple Voltage-Gated Ion Channels in Rabbit Ventricular Cardiomyocytes in TRPV1-Independent Manner. Pharmaceuticals 2022, 15, 1187. [Google Scholar] [CrossRef] [PubMed]
- Szoka, L.; Palka, J. Capsaicin up-regulates pro-apoptotic activity of thiazolidinediones in glioblastoma cell line. Biomed. Pharmacother. 2020, 132, 110741. [Google Scholar] [CrossRef] [PubMed]
- Bort, A.; Sánchez, B.G.; Mateos-Gómez, P.A.; Díaz-Laviada, I.; Rodríguez-Henche, N. Capsaicin Targets Lipogenesis in HepG2 Cells Through AMPK Activation, AKT Inhibition and PPARs Regulation. Int. J. Mol. Sci. 2019, 20, 1660. [Google Scholar] [CrossRef] [PubMed]
- Wagner, N. & Wagner, K. D. The Role of PPARs in Disease. Cells 2020, Vol. 9, Page 2367 9, 2367 (2020).
- Kang, C.; Zhang, Y.; Zhu, X.; Liu, K.; Wang, X.; Chen, M.; Wang, J.; Chen, H.; Hui, S.; Huang, L.; et al. Healthy Subjects Differentially Respond to Dietary Capsaicin Correlating with Specific Gut Enterotypes. J. Clin. Endocrinol. Metab. 2016, 101, 4681–4689. [Google Scholar] [CrossRef]
- Zeng, H.; Shi, N.; Peng, W.; Yang, Q.; Ren, J.; Yang, H.; Chen, L.; Chen, Y.; Guo, J. Effects of Capsaicin on Glucose Uptake and Consumption in Hepatocytes. Molecules 2023, 28, 5258. [Google Scholar] [CrossRef]
- Wan, H.; Chen, X.Y.; Zhang, F.; Chen, J.; Chu, F.; Sellers, Z.M.; Xu, F.; Dong, H. Capsaicin inhibits intestinal Cl- secretion and promotes Na+ absorption by blocking TRPV4 channels in healthy and colitic mice. J. Biol. Chem. 2022, 298, 101847. [Google Scholar] [CrossRef]
- Mahalak, K.K.; Bobokalonov, J.; Firrman, J.; Williams, R.; Evans, B.; Fanelli, B.; Soares, J.W.; Kobori, M.; Liu, L. Analysis of the Ability of Capsaicin to Modulate the Human Gut Microbiota In Vitro. Nutrients 2022, 14, 1283. [Google Scholar] [CrossRef]
- Kang, Y.; Cai, Y. Gut microbiota and obesity: implications for fecal microbiota transplantation therapy. Hormones 2017, 16, 223–234. [Google Scholar] [CrossRef]
- Wang, F.; Huang, X.; Chen, Y.; Zhang, D.; Chen, D.; Chen, L.; Lin, J. Study on the Effect of Capsaicin on the Intestinal Flora through High-Throughput Sequencing. ACS Omega 2020, 5, 1246–1253. [Google Scholar] [CrossRef]
- Dai, Z. et al. Capsaicin Ameliorates High-Fat Diet-Induced Atherosclerosis in ApoE-/- Mice via Remodeling Gut Microbiota. Nutrients 14, (2022).
- Garufi, A.; Pistritto, G.; Cirone, M.; D’orazi, G. Reactivation of mutant p53 by capsaicin, the major constituent of peppers. J. Exp. Clin. Cancer Res. 2016, 35, 1–9. [Google Scholar] [CrossRef]
- Cheng, P.; Wu, J.; Zong, G.; Wang, F.; Deng, R.; Tao, R.; Qian, C.; Shan, Y.; Wang, A.; Zhao, Y.; et al. Capsaicin shapes gut microbiota and pre-metastatic niche to facilitate cancer metastasis to liver. Pharmacol. Res. 2023, 188, 106643. [Google Scholar] [CrossRef] [PubMed]
- Rosca, A.E.; Iesanu, M.I.; Zahiu, C.D.M.; Voiculescu, S.E.; Paslaru, A.C.; Zagrean, A.-M. Capsaicin and Gut Microbiota in Health and Disease. Molecules 2020, 25, 5681. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.G.B.; Faria, J.V.; dos Santos, J.P.S.; Faria, R.X. Capsaicin: TRPV1-independent mechanisms and novel therapeutic possibilities. Eur. J. Pharmacol. 2020, 887, 173356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L. et al. Beneficial effect of capsaicin via TRPV4/EDH signals on mesenteric arterioles of normal and colitis mice. J. Adv. Res. 39, 291–303 (2022).
- Holzer, P. Vanilloid receptor TRPV1: hot on the tongue and inflaming the colon. Neurogastroenterol. Motil. 2004, 16, 697–699. [Google Scholar] [CrossRef] [PubMed]
- Saha, K. et al. Capsaicin Inhibits Inflammation and Gastric Damage during H. pylori Infection by Targeting NF-kB–miRNA Axis. Pathogens 11, 641 (2022).
- Xiang, Q.; Tang, X.; Cui, S.; Zhang, Q.; Liu, X.; Zhao, J.; Zhang, H.; Mao, B.; Chen, W. Capsaicin, the Spicy Ingredient of Chili Peppers: Effects on Gastrointestinal Tract and Composition of Gut Microbiota at Various Dosages. Foods 2022, 11, 686. [Google Scholar] [CrossRef]
- Liu, Y.-P.; Dong, F.-X.; Chai, X.; Zhu, S.; Zhang, B.-L.; Gao, D.-S. Role of Autophagy in Capsaicin-Induced Apoptosis in U251 Glioma Cells. Cell. Mol. Neurobiol. 2015, 36, 737–743. [Google Scholar] [CrossRef]
- Zheng, L.; Chen, J.; Ma, Z.; Liu, W.; Yang, F.; Yang, Z.; Wang, K.; Wang, X.; He, D.; Li, L.; et al. Capsaicin enhances anti-proliferation efficacy of pirarubicin via activating TRPV1 and inhibiting PCNA nuclear translocation in 5637 cells. Mol. Med. Rep. 2016, 13, 881–887. [Google Scholar] [CrossRef]
- Liang, W.; Lan, Y.; Chen, C.; Song, M.; Xiao, J.; Huang, Q.; Cao, Y.; Ho, C.-T.; Lu, M. Modulating effects of capsaicin on glucose homeostasis and the underlying mechanism. Crit. Rev. Food Sci. Nutr. 2021, 63, 3634–3652. [Google Scholar] [CrossRef]
- Ferdowsi, P.V.; Ahuja, K.D.K.; Beckett, J.M.; Myers, S. TRPV1 Activation by Capsaicin Mediates Glucose Oxidation and ATP Production Independent of Insulin Signalling in Mouse Skeletal Muscle Cells. Cells 2021, 10, 1560. [Google Scholar] [CrossRef]
- Xia, Y.; Lee, G.; Yamamoto, M.; Takahashi, H.; Kuda, T. Detection of indigenous gut bacteria related to red chilli pepper (Capsicum annuum) in murine caecum and human faecal cultures. Mol. Biol. Rep. 2022, 49, 10239–10250. [Google Scholar] [CrossRef]
- Zhu, S. L. et al. Capsaicin ameliorates intermittent high glucose-mediated endothelial senescence via the TRPV1/SIRT1 pathway. Phytomedicine 100, 154081 (2022).
- Dai, Z. et al. Capsaicin Ameliorates High-Fat Diet-Induced Atherosclerosis in ApoE−/− Mice via Remodeling Gut Microbiota. Nutrients 14, 4334 (2022).
- Zhou, Y.; Wang, X.; Guo, L.; Chen, L.; Zhang, M.; Chen, X.; Li, J.; Zhang, L. TRPV1 activation inhibits phenotypic switching and oxidative stress in vascular smooth muscle cells by upregulating PPARα. Biochem. Biophys. Res. Commun. 2021, 545, 157–163. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, D.; Huang, J.; Hu, Y.; Xu, Y. Application of capsaicin as a potential new therapeutic drug in human cancers. J. Clin. Pharm. Ther. 2019, 45, 16–28. [Google Scholar] [CrossRef]
- McCarty, M.F.; DiNicolantonio, J.J.; O'Keefe, J.H. Capsaicin may have important potential for promoting vascular and metabolic health: Table 1. Open Hear. 2015, 2, e000262. [Google Scholar] [CrossRef]
- Song, J.-X.; Ren, H.; Gao, Y.-F.; Lee, C.-Y.; Li, S.-F.; Zhang, F.; Li, L.; Chen, H. Dietary Capsaicin Improves Glucose Homeostasis and Alters the Gut Microbiota in Obese Diabetic ob/ob Mice. Front. Physiol. 2017, 8, 602. [Google Scholar] [CrossRef]
- Xue, M.; Ji, X.; Xue, C.; Liang, H.; Ge, Y.; He, X.; Zhang, L.; Bian, K.; Zhang, L. Caspase-dependent and caspase-independent induction of apoptosis in breast cancer by fucoidan via the PI3K/AKT/GSK3β pathway in vivo and in vitro. BioMedicine 2017, 94, 898–908. [Google Scholar] [CrossRef] [PubMed]
- Szallasi, A. Capsaicin for Weight Control: "Exercise in a Pill" (or Just Another Fad)? 15 851 ( 2022. [CrossRef] [PubMed]
- Wang, M. , Huang, W. & Xu, Y. Effects of spicy food consumption on overweight/obesity, hypertension and blood lipids in China: a meta-analysis of cross-sectional studies. Nutr. J. 22, 29 (2023).
- Baenas, N.; Belović, M.; Ilic, N.; Moreno, D.; García-Viguera, C. Industrial use of pepper (Capsicum annum L.) derived products: Technological benefits and biological advantages. Food Chem. 2018, 274, 872–885. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Xue, Y.; Fu, L.; Wang, Y.; He, M.; Zhao, L.; Liao, X. Extraction, purification, bioactivity and pharmacological effects of capsaicin: a review. Crit. Rev. Food Sci. Nutr. 2021, 62, 5322–5348. [Google Scholar] [CrossRef]
- Uarrota, V.G.; Maraschin, M.; Bairros. d.F.M.d.; Pedreschi, R. Factors affecting the capsaicinoid profile of hot peppers and biological activity of their non-pungent analogs (Capsinoids) present in sweet peppers. Crit. Rev. Food Sci. Nutr. 2020, 61, 649–665. [Google Scholar] [CrossRef]
- Gupta, R.; Kapoor, B.; Gulati, M.; Kumar, B.; Gupta, M.; Singh, S.K.; Awasthi, A. Sweet pepper and its principle constituent capsiate: functional properties and health benefits. Crit. Rev. Food Sci. Nutr. 2021, 62, 7370–7394. [Google Scholar] [CrossRef]
- Sanati, S.; Razavi, B.M.; Hosseinzadeh, H. A review of the effects of Capsicum annuum L. and its constituent, capsaicin, in metabolic syndrome. 2018, 21, 439–448. [Google Scholar] [CrossRef]
- Thornton, T.; Mills, D.; Bliss, E. Capsaicin: A Potential Treatment to Improve Cerebrovascular Function and Cognition in Obesity and Ageing. Nutrients 2023, 15, 1537. [Google Scholar] [CrossRef] [PubMed]
- Oz, M.; Lorke, D.E.; Howarth, F.C. Transient receptor potential vanilloid 1 (TRPV1)-independent actions of capsaicin on cellular excitability and ion transport. Med. Res. Rev. 2023, 43, 1038–1067. [Google Scholar] [CrossRef] [PubMed]
- vila, D. L. et al. Signaling Targets Related to Antiobesity Effects of Capsaicin: A Scoping Review. Adv. Nutr. 12, 2232–2243 (2021).
- Sanjay, S.; Sharma, A.; Lee, H.-J. Role of Phytoconstituents as PPAR Agonists: Implications for Neurodegenerative Disorders. Biomedicines 2021, 9, 1914. [Google Scholar] [CrossRef]
- Krishnan, V.; Baskaran, P.; Thyagarajan, B. Troglitazone activates TRPV1 and causes deacetylation of PPARγ in 3T3-L1 cells. Biochim. et Biophys. Acta (BBA) - Mol. Basis Dis. 2018, 1865, 445–453. [Google Scholar] [CrossRef]
- Baboota, R.K.; Singh, D.P.; Sarma, S.M.; Kaur, J.; Sandhir, R.; Boparai, R.K.; Kondepudi, K.K.; Bishnoi, M. Capsaicin Induces “Brite” Phenotype in Differentiating 3T3-L1 Preadipocytes. PLOS ONE 2014, 9, e103093. [Google Scholar] [CrossRef]
- Zhang, L.L.; Liu, D.Y.; Ma, L.Q.; Luo, Z.D.; Cao, T.B.; Zhong, J.; Yan, Z.C.; Wang, L.J.; Zhao, Z.G.; Zhu, S.J.; et al. Activation of Transient Receptor Potential Vanilloid Type-1 Channel Prevents Adipogenesis and Obesity. Circ. Res. 2007, 100, 1063–1070. [Google Scholar] [CrossRef]
- Chen, J. et al. Activation of TRPV1 channel by dietary capsaicin improves visceral fat remodeling through connexin43-mediated Ca2+ Influx. Cardiovasc. Diabetol. 14, (2015).
- Kida, R.; Noguchi, T.; Murakami, M.; Hashimoto, O.; Kawada, T.; Matsui, T.; Funaba, M. Supra-pharmacological concentration of capsaicin stimulates brown adipogenesis through induction of endoplasmic reticulum stress. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Fan, L.; Xu, H.; Yang, R.; Zang, Y.; Chen, J.; Qin, H. Combination of Capsaicin and Capsiate Induces Browning in 3T3-L1 White Adipocytes via Activation of the Peroxisome Proliferator-Activated Receptor γ/β3-Adrenergic Receptor Signaling Pathways. J. Agric. Food Chem. 2019, 67, 6232–6240. [Google Scholar] [CrossRef]
- Ludy, M.-J.; Moore, G.E.; Mattes, R.D. The Effects of Capsaicin and Capsiate on Energy Balance: Critical Review and Meta-analyses of Studies in Humans. Chem. Senses 2011, 37, 103–121. [Google Scholar] [CrossRef]
- Oh, M.-J.; Lee, H.-B.; Yoo, G.; Park, M.R.; Lee, C.-H.; Choi, I.; Park, H.-Y. Anti-obesity effects of red pepper (Capsicum annuum L.) leaf extract on 3T3-L1 preadipocytes and high fat diet-fed mice. Food Funct. 2022, 14, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Cao, Y.; Xiao, J.; Song, M.; Ho, C.-T. Molecular mechanisms of the anti-obesity effect of bioactive ingredients in common spices: a review. Food Funct. 2018, 9, 4569–4581. [Google Scholar] [CrossRef] [PubMed]
- Mosqueda-Solís, A.; Sánchez, J.; Portillo, M.P.; Palou, A.; Picó, C. Combination of Capsaicin and Hesperidin Reduces the Effectiveness of Each Compound To Decrease the Adipocyte Size and To Induce Browning Features in Adipose Tissue of Western Diet Fed Rats. J. Agric. Food Chem. 2018, 66, 9679–9689. [Google Scholar] [CrossRef] [PubMed]
- Silvester, A.J.; Aseer, K.R.; Yun, J.W. Dietary polyphenols and their roles in fat browning. J. Nutr. Biochem. 2019, 64, 1–12. [Google Scholar] [CrossRef]
- Takeda, Y.; Dai, P. Capsaicin directly promotes adipocyte browning in the chemical compound-induced brown adipocytes converted from human dermal fibroblasts. Sci. Rep. 2022, 12, 1–14. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, C.; Tang, Y.; Yin, H.; Liu, X. Capsaicin has an anti-obesity effect through alterations in gut microbiota populations and short-chain fatty acid concentrations. Food Nutr. Res. 2020, 64. [Google Scholar] [CrossRef]
- Siebert, E.; Lee, S.-Y.; Prescott, M.P. Chili pepper preference development and its impact on dietary intake: A narrative review. Front. Nutr. 2022, 9, 1039207. [Google Scholar] [CrossRef]
- Sirotkin, A.V. Peppers and their constituents against obesity. Biol. Futur. 2023, 1–6. [Google Scholar] [CrossRef]
- Azlan, A.; Sultana, S.; Huei, C.S.; Razman, M.R. Antioxidant, Anti-Obesity, Nutritional and Other Beneficial Effects of Different Chili Pepper: A Review. Molecules 2022, 27, 898. [Google Scholar] [CrossRef]
- Smeets, A.J.; Westerterp-Plantenga, M.S. The acute effects of a lunch containing capsaicin on energy and substrate utilisation, hormones, and satiety. Eur. J. Nutr. 2009, 48, 229–234. [Google Scholar] [CrossRef]
- van Avesaat, M.; Troost, F.J.; Westerterp-Plantenga, M.S.; Helyes, Z.; Le Roux, C.W.; Dekker, J.; Masclee, A.A.; Keszthelyi, D. Capsaicin-induced satiety is associated with gastrointestinal distress but not with the release of satiety hormones. Am. J. Clin. Nutr. 2016, 103, 305–313. [Google Scholar] [CrossRef]
- Rigamonti, A.E.; Casnici, C.; Marelli, O.; De Col, A.; Tamini, S.; Lucchetti, E.; Tringali, G.; De Micheli, R.; Abbruzzese, L.; Bortolotti, M.; et al. Acute administration of capsaicin increases resting energy expenditure in young obese subjects without affecting energy intake, appetite, and circulating levels of orexigenic/anorexigenic peptides. Nutr. Res. 2018, 52, 71–79. [Google Scholar] [CrossRef]
- Silva, V.E.L.d.M.e.; Cholewa, J.M.; Jäger, R.; Zanchi, N.E.; de Freitas, M.C.; de Moura, R.C.; Barros, E.M.L.; Antunes, B.M.; Caperuto, E.C.; Ribeiro, S.L.G.; et al. Chronic capsiate supplementation increases fat-free mass and upper body strength but not the inflammatory response to resistance exercise in young untrained men: a randomized, placebo-controlled and double-blind study. J. Int. Soc. Sports Nutr. 2021, 18, 1–9. [Google Scholar] [CrossRef]
- Zsiborás, C.; Mátics, R.; Hegyi, P.; Balaskó, M.; Pétervári, E.; Szabó, I.; Sarlós, P.; Mikó, A.; Tenk, J.; Rostás, I.; et al. Capsaicin and capsiate could be appropriate agents for treatment of obesity: A meta-analysis of human studies. Crit. Rev. Food Sci. Nutr. 2017, 58, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Guo, S.; He, H.; Gong, L.; Cui, H. Gut Microbiome and Serum Metabolome Analyses Identify Unsaturated Fatty Acids and Butanoate Metabolism Induced by Gut Microbiota in Patients With Chronic Spontaneous Urticaria. Front. Cell. Infect. Microbiol. 2020, 10, 24. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V. et al. Mucin secretory action of capsaicin prevents high fat diet-induced gut barrier dysfunction in C57BL/6 mice colon. Biomed. Pharmacother. 145, 112452 (2022).
- Xia, J.; Gu, L.; Guo, Y.; Feng, H.; Chen, S.; Jurat, J.; Fu, W.; Zhang, D. Gut Microbiota Mediates the Preventive Effects of Dietary Capsaicin Against Depression-Like Behavior Induced by Lipopolysaccharide in Mice. Front. Cell. Infect. Microbiol. 2021, 1. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Kuda, T.; Yamamoto, M.; Yano, T.; Nakamura, A.; Takahashi, H. The effect of Sichuan pepper on gut microbiota in mice fed a high-sucrose and low-dietary fibre diet. Appl. Microbiol. Biotechnol. 2023, 107, 2627–2638. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Pan, S.; Li, F.; Xu, X.; Xing, H. Plant-Derived Bioactive Compounds and Potential Health Benefits: Involvement of the Gut Microbiota and Its Metabolic Activity. Biomolecules 2022, 12, 1871. [Google Scholar] [CrossRef]
- Kang, C.; Wang, B.; Kaliannan, K.; Wang, X.; Lang, H.; Hui, S.; Huang, L.; Zhang, Y.; Zhou, M.; Chen, M.; et al. Gut Microbiota Mediates the Protective Effects of Dietary Capsaicin against Chronic Low-Grade Inflammation and Associated Obesity Induced by High-Fat Diet. Mbio 2017, 8. [Google Scholar] [CrossRef]
- Manca, C.; Lacroix, S.; Pérusse, F.; Flamand, N.; Chagnon, Y.; Drapeau, V.; Tremblay, A.; Di Marzo, V.; Silvestri, C. Oral Capsaicinoid Administration Alters the Plasma Endocannabinoidome and Fecal Microbiota of Reproductive-Aged Women Living with Overweight and Obesity. Biomedicines 2021, 9, 1246. [Google Scholar] [CrossRef]
- Mikami, A.; Ogita, T.; Namai, F.; Shigemori, S.; Sato, T.; Shimosato, T. Oral administration of Flavonifractor plautii attenuates inflammatory responses in obese adipose tissue. Mol. Biol. Rep. 2020, 47, 6717–6725. [Google Scholar] [CrossRef] [PubMed]
- Ao, Z. , Huang, Z. & Liu, H. Spicy Food and Chili Peppers and Multiple Health Outcomes: Umbrella Review. Molecular Nutrition and Food Research vol. 66 (2022).
- Zanzer, Y.C.; Plaza, M.; Dougkas, A.; Turner, C.; Östman, E. Black pepper-based beverage induced appetite-suppressing effects without altering postprandial glycaemia, gut and thyroid hormones or gastrointestinal well-being: a randomized crossover study in healthy subjects. Food Funct. 2018, 9, 2774–2786. [Google Scholar] [CrossRef] [PubMed]
- Ashokkumar, K.; Murugan, M.; Dhanya, M.K.; Pandian, A.; Warkentin, T.D. Phytochemistry and therapeutic potential of black pepper [Piper nigrum (L.)] essential oil and piperine: a review. Clin. Phytoscience 2021, 7, 1–11. [Google Scholar] [CrossRef]
- Yang, X.; Tang, W.; Mao, D.; Liu, X.; Qian, W.; Dai, Y.; Chen, L.; Ding, X. Spicy food consumption is associated with abdominal obesity among Chinese Han population aged 30–79 years in the Sichuan Basin: a population-based cross-sectional study. BMC Public Heal. 2022, 22, 1–15. [Google Scholar] [CrossRef]
- Wen, Q.; Wei, Y.; Du, H.; Lv, J.; Guo, Y.; Bian, Z.; Yang, L.; Chen, Y.; Chen, Y.; Shi, L.; et al. Characteristics of spicy food consumption and its relation to lifestyle behaviours: results from 0.5 million adults. Int. J. Food Sci. Nutr. 2020, 72, 569–576. [Google Scholar] [CrossRef]
- Duranova, H.; Valkova, V.; Gabriny, L. Chili peppers (Capsicum spp.): the spice not only for cuisine purposes: an update on current knowledge. Phytochem. Rev. 2021, 21, 1379–1413. [Google Scholar] [CrossRef]
- Names, B. Capsaicin ( Topical Route ). 1–6 (2012).
- Chang, A. , Rosani, A. & Quick, J. Capsaicin - StatPearls - NCBI Bookshelf. https://www.ncbi.nlm.nih.gov/books/NBK459168/?report=reader (2022).
Figure 1.
General metabolism of capsaicin orally or topically administered. Dietary capsaicin absorbed in the stomach reaches the hepatocytes via the portal system, where it is metabolized by the cytochrome P450 (CYP450) enzyme system. When given through topical administration, capsaicin is rapidly absorbed and undergoes slow biotransformation, mainly remaining intact, without hepatic biotransformation. After reaching systemic circulation, capsaicin and its metabolites are distributed to peripherical organs and trigger systemic effects. Finally, they are eliminated mainly through the kidneys.
Figure 1.
General metabolism of capsaicin orally or topically administered. Dietary capsaicin absorbed in the stomach reaches the hepatocytes via the portal system, where it is metabolized by the cytochrome P450 (CYP450) enzyme system. When given through topical administration, capsaicin is rapidly absorbed and undergoes slow biotransformation, mainly remaining intact, without hepatic biotransformation. After reaching systemic circulation, capsaicin and its metabolites are distributed to peripherical organs and trigger systemic effects. Finally, they are eliminated mainly through the kidneys.
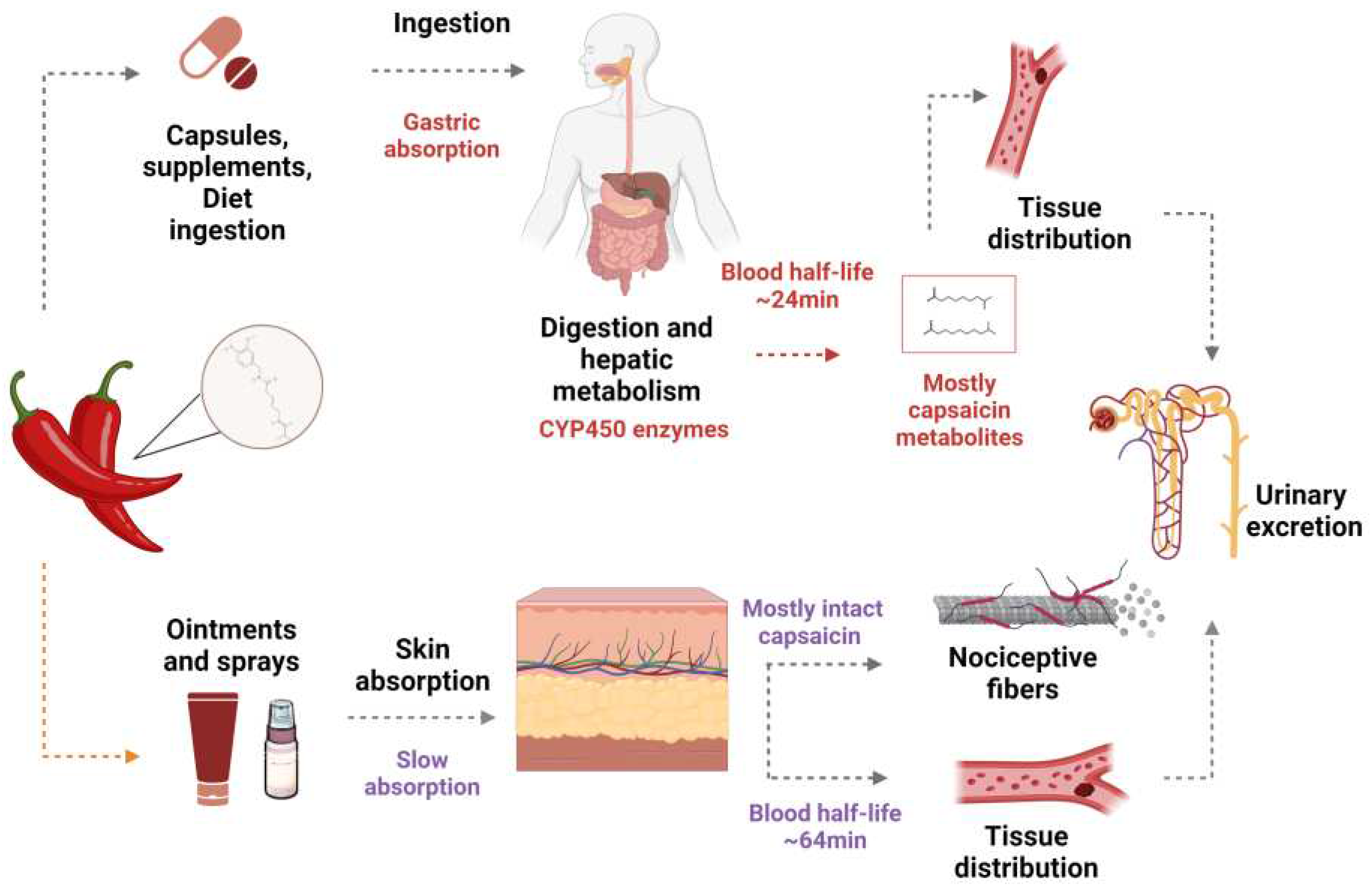
Figure 2.
General scheme of the main effects described for the TRPV1-dependent action of capsaicin. The rapid influx of calcium caused by the binding of the capsaicin molecule to its receptor triggers a cascade of signals that include phosphorylation and dephosphorylation of second messengers and enzymes, regulating mainly the pathways of nociception (in neural tissue), thermogenesis, oxidative stress, inflammation and adipogenesis in adipose tissue.
Figure 2.
General scheme of the main effects described for the TRPV1-dependent action of capsaicin. The rapid influx of calcium caused by the binding of the capsaicin molecule to its receptor triggers a cascade of signals that include phosphorylation and dephosphorylation of second messengers and enzymes, regulating mainly the pathways of nociception (in neural tissue), thermogenesis, oxidative stress, inflammation and adipogenesis in adipose tissue.
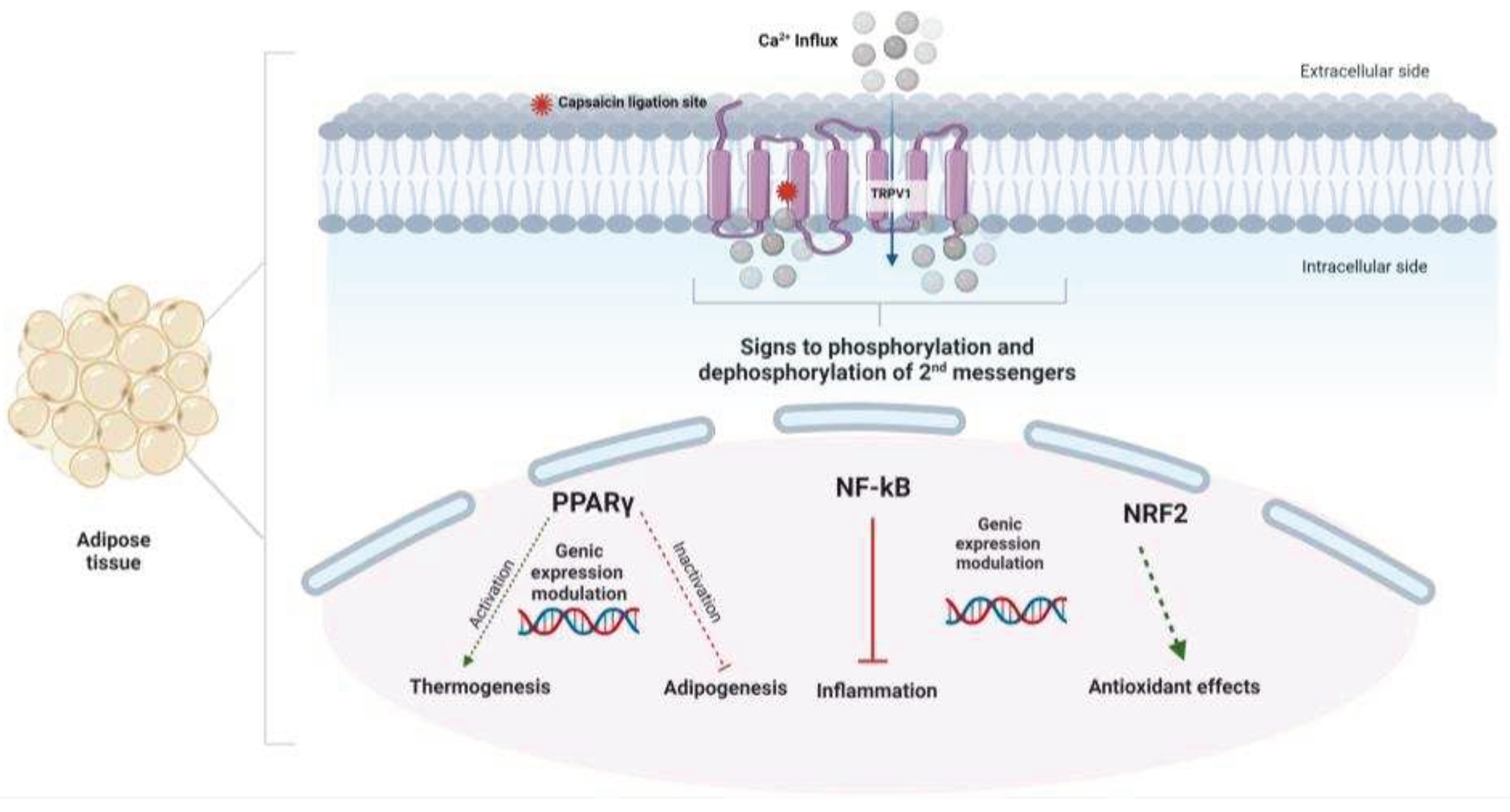
Figure 3.
Main actions carried out by capsaicin independently of TRPV1. A) Regardless of the presence of receptors, capsaicin could interact with biological membranes and modify membrane fluidity or potential, triggering signals conducted by the involved second messengers, which causes structural changes in cells and organelles, including mitochondria. These structural changes culminate in physiological responses in non-neuronal tissues that may be related to capsaicin's beneficial or adverse actions. Furthermore, it is suggested that capsaicin could also directly bind to receptors or channels other than TRPV1 and generate changes in the metabolic pathways related to these receptors. B) In intestinal epithelial cells (IECs), capsaicin could induce changes in other ion channels, such as TRPV4, blocking their actions. This blockade attenuates Cl- secretion and stimulates Na+ absorption. NKA (Na/K ATPase); (ENaC) epithelial Na+ channels; (CFTR) cystic fibrosis transmembrane conductance regulator; (SGLT1) Na+-glucose cotransporter 1.
Figure 3.
Main actions carried out by capsaicin independently of TRPV1. A) Regardless of the presence of receptors, capsaicin could interact with biological membranes and modify membrane fluidity or potential, triggering signals conducted by the involved second messengers, which causes structural changes in cells and organelles, including mitochondria. These structural changes culminate in physiological responses in non-neuronal tissues that may be related to capsaicin's beneficial or adverse actions. Furthermore, it is suggested that capsaicin could also directly bind to receptors or channels other than TRPV1 and generate changes in the metabolic pathways related to these receptors. B) In intestinal epithelial cells (IECs), capsaicin could induce changes in other ion channels, such as TRPV4, blocking their actions. This blockade attenuates Cl- secretion and stimulates Na+ absorption. NKA (Na/K ATPase); (ENaC) epithelial Na+ channels; (CFTR) cystic fibrosis transmembrane conductance regulator; (SGLT1) Na+-glucose cotransporter 1.
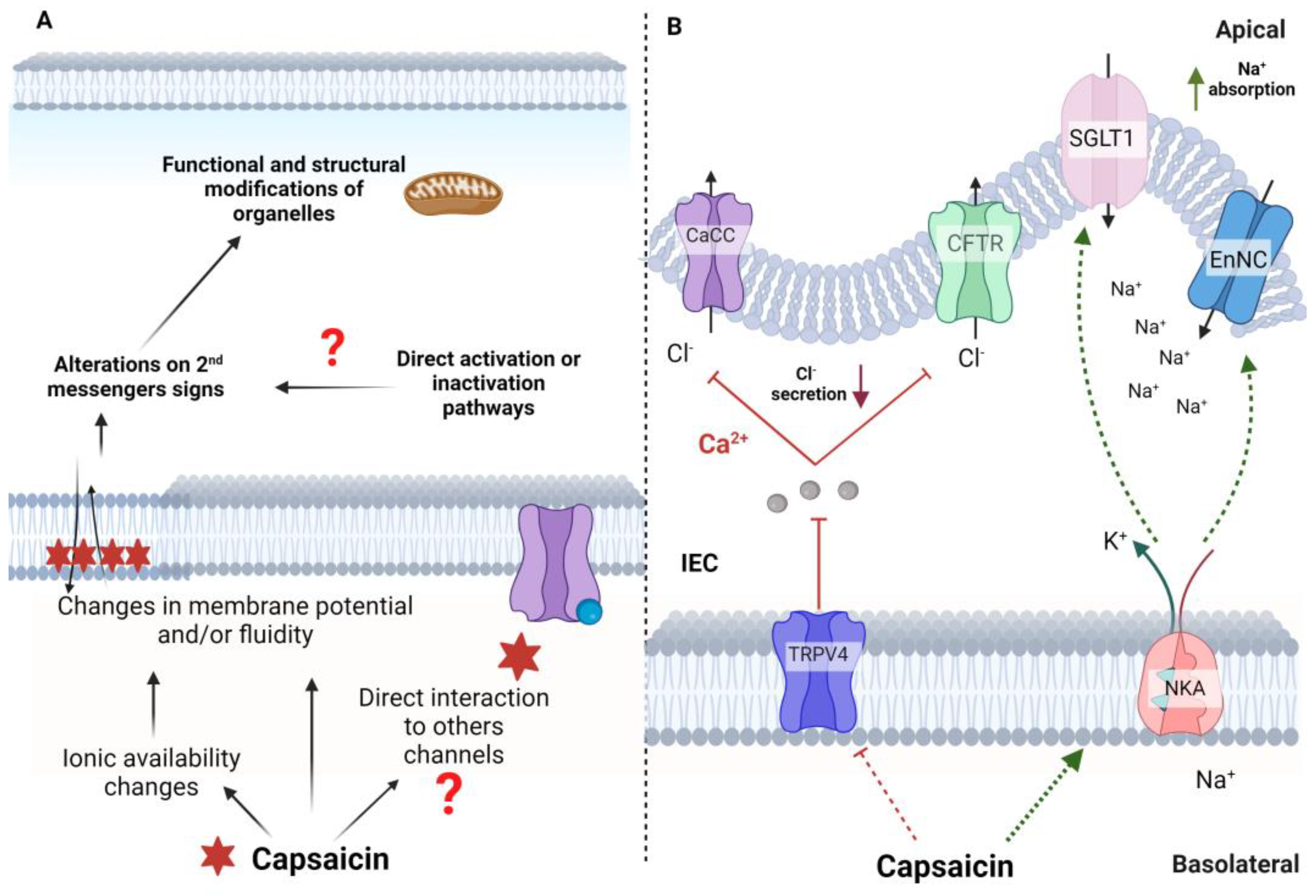
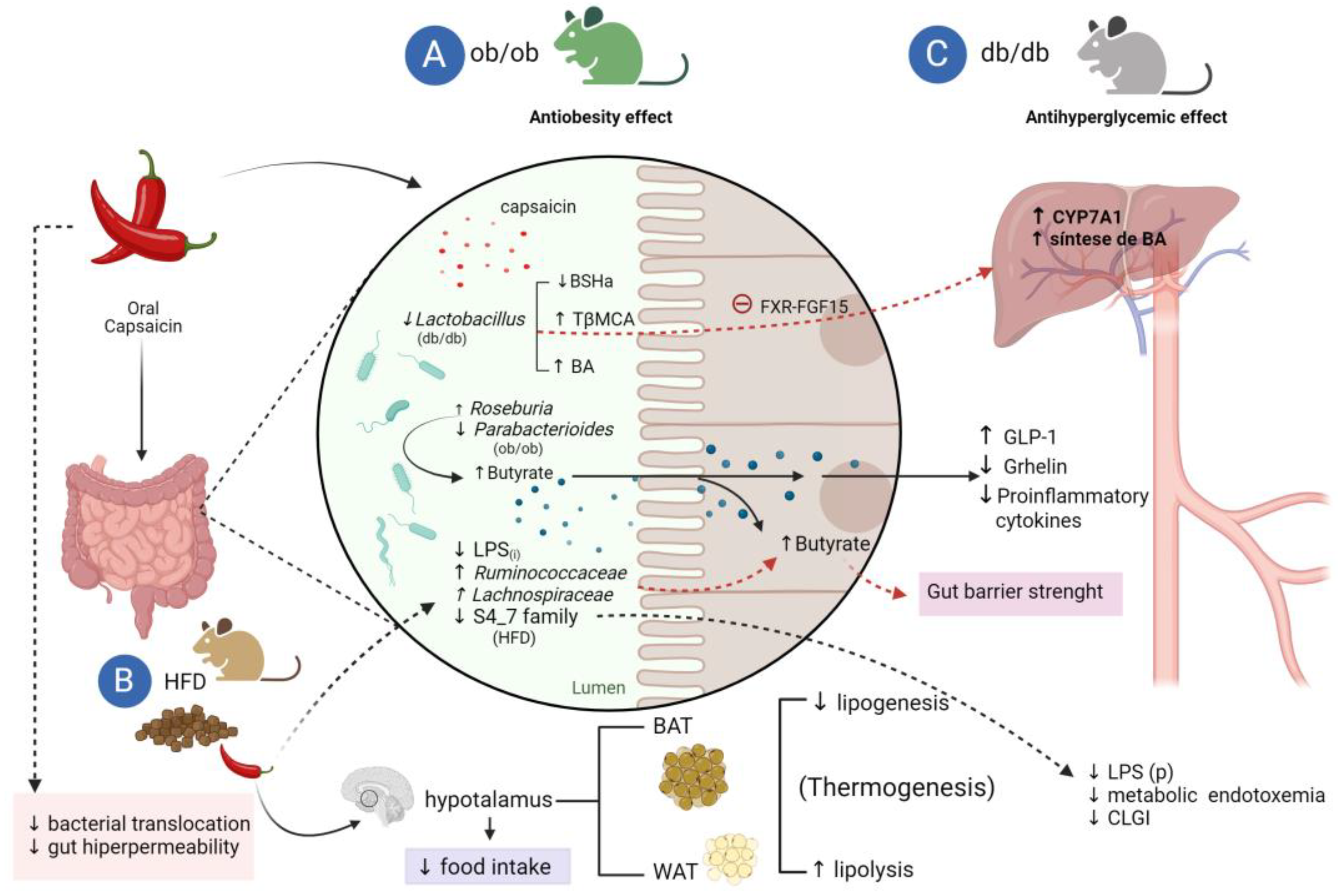
Figure 4.
Influence of dietary capsaicin on glucose homeostasis and obesity by modulation of the gut microbiota. A. capsaicin increases the abundance of Roseburia and decreases the abundance of Bacteroides and Parabacterioides in obese diabetic mice (ob/ob), leading to an increase in butyrate and GLP-1 and a decrease in total plasma ghrelin and also and proinflammatory cytokines. B. The antiobesity effects of capsaicin in mice fed a high-fat diet (HFD) modulate the gut-brain (hypothalamus) axis by decreasing food intake. In addition, capsaicin induces browning, favoring the formation of brown adipose tissue (BAT) and stimulating lipolysis of white adipose tissue (WAT). Capsaicin reduces the abundance of gram-negative bacteria that secrete intestinal lipopolysaccharide (LPS(i)) and increases the abundance of butirogenic bacteria (Ruminococcacea and Lachnospiraceae) and, consequently, butyrate. This effect attenuates the increased intestinal permeability and bacterial translocation caused by HFD and suppresses cannabinoid intestinal receptor type 1 (CB1(i)) expression. Thus, dietary capsaicin strengthens the intestinal barrier due to the increase of butyrate production, decrease of LPS(i) and plasma LPS (LPS(p)) with reduction of metabolic endotoxemia and chronic low-grade inflammation (CLGI) C. The antihyperglycemic effect of capsaicin is due to a decrease in the abundance of Lactobacillus in diabetic mice (db/db), causing a reduction in the activity of bile salt hydrolase (BSHa) and an increase in the levels of conjugated bile acids (BA) in the intestine and of β-muricholic tauro acid (TβMCA), a farsenoid receptor antagonist (FXR), leading to the suppression of the FXR-FGF15 axis and a positive regulation of the expression of cholesterol 7 hydroxylase (CYP7A 1) stimulating the hepatic synthesis of BA.
Figure 4.
Influence of dietary capsaicin on glucose homeostasis and obesity by modulation of the gut microbiota. A. capsaicin increases the abundance of Roseburia and decreases the abundance of Bacteroides and Parabacterioides in obese diabetic mice (ob/ob), leading to an increase in butyrate and GLP-1 and a decrease in total plasma ghrelin and also and proinflammatory cytokines. B. The antiobesity effects of capsaicin in mice fed a high-fat diet (HFD) modulate the gut-brain (hypothalamus) axis by decreasing food intake. In addition, capsaicin induces browning, favoring the formation of brown adipose tissue (BAT) and stimulating lipolysis of white adipose tissue (WAT). Capsaicin reduces the abundance of gram-negative bacteria that secrete intestinal lipopolysaccharide (LPS(i)) and increases the abundance of butirogenic bacteria (Ruminococcacea and Lachnospiraceae) and, consequently, butyrate. This effect attenuates the increased intestinal permeability and bacterial translocation caused by HFD and suppresses cannabinoid intestinal receptor type 1 (CB1(i)) expression. Thus, dietary capsaicin strengthens the intestinal barrier due to the increase of butyrate production, decrease of LPS(i) and plasma LPS (LPS(p)) with reduction of metabolic endotoxemia and chronic low-grade inflammation (CLGI) C. The antihyperglycemic effect of capsaicin is due to a decrease in the abundance of Lactobacillus in diabetic mice (db/db), causing a reduction in the activity of bile salt hydrolase (BSHa) and an increase in the levels of conjugated bile acids (BA) in the intestine and of β-muricholic tauro acid (TβMCA), a farsenoid receptor antagonist (FXR), leading to the suppression of the FXR-FGF15 axis and a positive regulation of the expression of cholesterol 7 hydroxylase (CYP7A 1) stimulating the hepatic synthesis of BA.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



