
Submitted:
26 September 2023
Posted:
27 September 2023
Read the latest preprint version here
Abstract
The maintenance of skeletal muscle mass plays a fundamental role in health and issues associated with quality of life. Mechanical signals are one of the most potent regulators of muscle mass, with a decrease in mechanical loading leading to a decrease in muscle mass. This concept has been supported by a plethora of of human- and animal-based studies during the last 100 years and has resulted in the commonly used term of “disuse atrophy”. These same studies have also provided a great deal of insight into the structural adaptations that mediate disuse-induced atrophy. For instance, disuse results in radial atrophy of fascicles, and this is driven, at least in part, by radial atrophy of the muscle fibers. However, the ultrastructural adaptations that mediate these changes remain far from defined. Indeed, even the most basic questions, such as whether the radial atrophy of muscle fibers is driven by the radial atrophy of myofibrils and/or myofibril hypoplasia, have yet to be answered. In this review, we thoroughly summarize what is known about the macroscopic, microscopic, and ultrastructural adaptations that mediated disuse-induced atrophy and highlight some of the major gaps in knowledge that need to be filled.
Keywords:
disuse
; fascicle
; hypoplasia
; longitudinal atrophy
; muscle fibers
; myofibril
; myofilaments
; radial atrophy
; sarcomere
1. Introduction
Skeletal muscle is the largest organ in the human body and, on average, contributes to 35% of an individual’s body mass [1]. Skeletal muscles are not only the motors that drive basic movements, but they also play numerous physiological functions (e.g., regulation of body temperature and metabolism) and can function as endocrine organs via the secretion of myokines [2,3,4,5]. Based on these points, it is not surprising that the maintenance of muscle mass has been widely implicated in health and issues associated with quality of life. For instance, low muscle mass has been correlated with an increased risk of type II diabetes, heart disease, and death [6,7,8,9,10,11], and the maintenance of muscle mass becomes particularly important during aging as the age-associated loss of muscle has been strongly linked to disability, lack of independence, mortality, and an estimated $20-40 billion in annual healthcare costs in the United States alone [12,13,14,15]. Thus, the development of therapies that can maintain, restore, or even enhance muscle mass is of great clinical and fiscal significance. However, to develop such therapies, we will first need to develop a deeper understanding of the mechanisms that regulate the size of this vital organ.
It is well established that skeletal muscle mass can be regulated by a variety of different stimuli, and one of the most potent stimuli is mechanical loading [16,17]. Indeed, even in the 5th century BC, Hippocrates appreciated that “exercise strengthens and inactivity wastes” [18]. Thus, the notion that a loss in mechanical loading can lead to a loss of muscle mass appears to have been recognized for at least 2,500 years [18]. Over the last 100 years, a plethora of human- and animal-based studies have provided further support for this concept and resulted in the commonly used term “disuse atrophy”. These same studies have also provided insight into the macroscopic and microscopic changes that contribute to the loss of muscle mass (i.e., atrophy). Surprisingly, however, the ultrastructural adaptations that drive these changes remain far from defined. In this review, we will summarize what is currently known about the structural adaptations that mediate disuse-induced atrophy and highlight the critical gaps in knowledge that need to be filled.
2. Disuse atrophy and the models that can be used to study it
Before discussing the models that can be used to study disuse atrophy it is necessary to have a precise definition for the term “disuse”. In this review, disuse will be defined as any condition in which there is a maintenance or reduction in the mean passive tension along with a reduction in the mean active tension generated by the muscle. This definition is important to keep in mind because it will exclude certain skeletal muscles that are often examined in commonly used models of disuse atrophy. Furthermore, it is known that a variety of diseases such as congestive heart failure, cirrhosis, and cancer can lead to disuse [19,20,21]. However, such diseases often present with an increase in circulating atrophic factors such as cytokines and cortisol [21,22,23,24]. The structural adaptations that contribute to disuse atrophy in diseased versus otherwise healthy individuals may be different and, therefore, this review will only focus on the structural adaptations that mediate disuse atrophy in subjects that are free of any apparent disease.
2.1. Space flight
Skeletal muscle atrophy is a widely recognized consequence of prolonged spaceflight, and the disuse that results from the microgravity environment of space is considered to be the key driver of the atrophic response [25,26,27]. However, as recently summarized by Lee et al., 2022, additional factors such as radiation and the stresses associated with living in space might also play a role [27]. Nonetheless, the most prominent atrophic response occurs in antigravity muscles (e.g., the soleus) which suggests that disuse is the primary driver [27,28]. Importantly, as defined in this review, some muscles of the body would not qualify as being subjected to disuse during space flight. For instance, humans assume a plantarflexed foot drop posture during spaceflight, and in this position, the plantarflexors are maintained in a shortened position whereas the dorsiflexors are maintained in a lengthed position [29,30]. The lengthening of the dorsiflexors would result in a chronic increase in the passive tension that is generated by the dorsiflexor muscles, and thus, these muscles would not fulfill the definition of being subjected to disuse. While this may seem like a trivial point, a chronic increase in length / passive tension can profoundly influence the structural adaptions that occur when a muscle is subjected to a chronic decrease in active force generation [31,32,33,34], and it is our hope that the reader will gain a deep appreciation for this concept as they work through the remainder of the review.
2.2. Bed rest
In the 19th century, bedrest was advocated as the primary line of treatment for a variety of ailments [35]. However, shortly after World War II, classic studies in humans such as the one performed by Dietrick and Whedon (1948) began to expose the negative impact that prolonged bedrest can have on various metabolic and physiological parameters (e.g., skeletal muscle mass and strength) [36]. Since that time, it has become clear that the disuse that occurs during prolonged bed rest can lead to profound atrophy, particularly of the primary weight-bearing muscles of the human body such as the soleus and quadriceps [37,38,39,40].
2.3. Hindlimb suspension
Hindlimb suspension (also known as hindlimb unloading) is an animal model that has been extensively used in the study of disuse atrophy. The model was originally developed as a complement to the 6º head-down bed rest model of spaceflight in humans, and similar to space flight, the hindlimbs of the suspended animals assume a plantarflexed position [41,42,43,44,45]. As such, the dorsiflexors (e.g., tibialis anterior and extensor digitorum longus) experience a chronic increase in passive tension and therefore do not fulfill the definition of being subjected to disuse. Again, this may seem like a trivial point, but it should not be overlooked. For instance, previous studies have shown that the soleus is particularly prone to hindlimb suspension-induced atrophy, yet the atrophic response can be prevented if the soleus is maintained in a lengthened position [46,47]. Thus, we would like to reiterate that, as defined in this review, disuse requires a maintenance or reduction in the mean passive tension as well as a reduction in the mean active tension generated by the muscle.
2.4. Unilateral Limb Suspension
The unilateral limb suspension model is a human model that can be used to study the impact that disuse has on muscles that are maintained in a relatively relaxed/neutral position. Specifically, in the unilateral limb suspension model, participants wear a shoe with a 100-mm-thick sole on the weight-bearing leg and then use crutches when they need to move around. With this approach, the unloaded limb can swing freely around the hip and numerous studies have shown that it induces atrophy of the knee extensors and the plantar flexors [48,49].
2.5. Immobilization
Immobilization is often used in the treatment of traumatic injuries such as fractured bones and torn ligaments. It is well recognized that immobilization can lead to disuse atrophy, and a variety of human and animal models of immobilization have been developed to study the mechanisms of this phenomenon. In humans, the most commonly employed approach involves the use of a cast or brace to immobilize the ankle or knee in a relatively neutral position [50,51]. More extreme variations have been used in animals in which splints, casts, or staples have been used to immobilize the lower hindlimb in a fully plantarflexed, neutral, or fully dorsiflexed position [52,53,54]. However, as defined in this review, only muscles that are immobilized at resting/neutral or shorter than resting length would qualify as being subjected to disuse. Again, the importance of the point was highlighted many years ago when it was shown that the length at which a muscle is immobilized can determine whether it atrophies. Specifically, in 1944, Thomsen and Luco reported that when the soleus or tibialis anterior muscles of cats were immobilized at a shorter-than-normal length the muscles atrophied, whereas immobilization of the same muscles at a longer-than-normal length led to an increase in muscle mass [55]. Thus, unless otherwise stated, any discussions in this review that refer to immobilization will imply that the muscles being discussed were immobilized at resting, or shorter than resting, length.
2.6. Denervation
Denervation can be caused by a variety of different conditions (e.g., motor neuron diseases such as ALS, physical transection of the nervous input, etc.), and various forms of denervation have been used as models of disuse atrophy. It is clear that denervation can lead to a dramatic atrophic response, however, numerous studies have also shown that denervation can lead to signs of muscle fiber death and a concomitant regenerative response [56,57,58,59]. These types of events are rarely, if ever, observed in the other models of disuse described above. Thus, the atrophy that occurs in response to denervation is likely the result of more than just disuse. Accordingly, studies that have employed denervation as a model of disuse will not be further considered in this review.
3. Overview of Skeletal Muscle Structure
Before we discuss what is known about the structural adaptations that mediate disuse atrophy, we first want to ensure that the reader appreciates the basic structural design of skeletal muscle. In our opinion, the easiest way to appreciate this design is to consider skeletal muscle as a hierarchy of structural elements that is visible at the macroscopic level (viewable without magnification), followed by the microscopic level (viewable with standard microscopy), and finally the ultrastructural level (viewable with high-resolution microscopy). Below we will describe the structural elements that are found at each of these levels. For a more detailed description of these components please refer to the following reviews [60,61,62,63].
Beginning with the macroscopic level, one will find that skeletal muscles are attached to bones via tendons (Figure 1A). Moving away from the bony connection, the tendon transitions into the musculotendinous junction and then into the epimysium which is a layer of connective tissue that encases the whole muscle [60]. Underneath the epimysium are smaller organizational units known as fascicles. Each fascicle consists of a bundle of muscle fibers that is encased by a layer of connective tissue called the perimysium [64,65]. It bears noting that in the majority of skeletal muscles, fascicles are not arranged in parallel with the long axis of the muscle, but rather run along an angle known as the pennation angle which plays an important role in muscle force production and size [63].
At the microscopic level, a cross-section of skeletal muscle will reveal the presence of individual muscle fibers (Figure 1A-C). Each muscle fiber is encased by a membrane called the sarcolemma that, in turn, is surrounded by a layer of connective tissue called the endomysium. Running between the muscle fibers are a variety of different types of interstitial cells including fibro-adipogenic/mesenchymal progenitors, fibroblasts, immune cells, and pericytes [66,67,68]. There is also a unique class of cells that reside between the endomysium and the sarcolemma called satellite cells which function as muscle stem cells and are critically important for myogenesis (Figure 1A, C) [67,68,69].
At the ultrastructural level, one will find that ~80% of each muscle fiber is filled with structures called myofibrils [70,71]. Myofibrils contain the basic contractile machinery of the muscle (i.e., the myosin (thick) and actin (thin) myofilaments) and are surrounded by mitochondria and a mesh-like membrane layer called the sarcoplasmic reticulum (Figure 1A, D, and E). Within the myofibril, the thick and thin myofilaments are arranged in a repetitive series of structures called sarcomeres (Figure 1A). The length of sarcomeres is highly conserved within a given species, typically measuring 2.0-2.5 µm in length [72,73], and this length is determined by the distance between the two Z-discs of the sarcomere. Within the sarcomere, the thin myofilaments attach to the Z-disc and extend toward the middle of the sarcomere (M-line) where thick myofilaments attach. The area closest to the Z-discs that are occupied solely by the thin myofilaments is known as the I-band whereas the middle portion of the sarcomere that is occupied by the thick myofilaments is known as the A-band (Figure 1A) [60,74,75]. Within the A-band, there is a region called the H-zone which only contains thick myofilaments. Throughout the remainder of the A-band, there is overlap between the thick and thin myofilaments and this overlap endows the sarcomere with the ability to actively produce force as described by the sliding filament theory of muscle contraction [76,77].
4. Disuse-Induced Atrophy at the Macroscopic Level
4.1. Whole muscle
At the whole muscle level, disuse-induced atrophy can be driven by a decrease in the length and/or a decrease in the diameter / cross-sectional area (CSA) of the muscle. Atrophy resulting from a decrease in the length is known as longitudinal atrophy, whereas atrophy that results from a decrease in the CSA is referred to as radial atrophy. As described in the ‘overview of skeletal muscle structure’, muscles are attached to bones, and in adults, the length of bones is fixed [78,79,80]. Accordingly, the resting length of muscles is generally viewed as being invariable and therefore has seldom been measured in studies of disuse atrophy. Nonetheless, a handful of studies have consistently shown that when a muscle is immobilized in a shortened position it can undergo substantial longitudinal atrophy [81,82,83,84]. For instance, Coutinho et al., (2004) reported that the length of the soleus muscle in rats decreased by 19% after 3 weeks of being immobilized in a shortened position [84], and the magnitude of this effect increased to 27% after 6 weeks of immobilization [82].
Although a few studies have examined the length of muscles that were subjected to disuse while being maintained in a shortened position, we are only aware of one study that has reported on the length of muscles that were subjected to disuse while being maintained a neutral/resting position. In this case, Spector et al. (1982) immobilized the hindlimb of rats in a neutral position for 4 weeks and found that several muscles including the soleus, medial gastrocnemius, and tibialis anterior experienced a 41-49% decrease in mass, but none of these muscles revealed a significant decrease in length [81]. Importantly, in the same study, additional experiments were performed in which the aforementioned muscles were immobilized in a shortened position and, in this case, all of the muscles underwent a significant 12-21% decrease in length [81]. Thus, according to the limited data in the field, it appears that when disuse occurs in conjunction with a chronic shortening of the muscle it will lead to longitudinal atrophy at the whole muscle level, but longitudinal atrophy at the whole muscle level will not transpire when muscles are subjected to disuse in a chronically neutral position.
The work of Spector et al. (1982) illustrates that a very large disuse-induced decrease in muscle mass can occur in the absence of longitudinal atrophy [81]. Like all materials, the mass of a given muscle will be a function of its volume and density, and we are not aware of any studies that have reported significant disuse-induced changes in muscle density. Thus, the decreases in muscle mass reported by Spector et al., were likely mediated by a decrease in volume. In skeletal muscle, volume is a function of the whole muscle CSA and the whole muscle length. Given that Spector et al. (1982) were able to observe decreases in mass in the absence of changes in length, it can be argued that the decreases in mass were driven by a decrease in the whole muscle CSA (i.e., radial atrophy) [81]. Indeed, radial atrophy at the whole muscle level has been observed in all of the models of disuse atrophy described in section 2 including, space flight [85,86], bed rest [87,88,89,90,91,92,93,94,95,96,97], unilateral lower limb suspension [98,99,100], and immobilization [81,101,102,103,104,105,106,107]. Of note, most of these studies obtained measurements of the whole muscle CSA at the midpoint along the length of the muscle (i.e., at 50% of the whole muscle length) [85,89,99]. However, disuse-induced changes in whole-muscle CSA are not always homogenous along the length of the muscle [92,95,96,97]. For instance, Franchi et al., (2022), reported that after just 10 days of bed rest in humans, a 5.8% and 7.0% reduction in the CSA of the biceps femoris short head could be detected at 30% and 50% of the muscle length (distal to proximal), respectively. Yet, at 70% of the muscle length, no significant change in CSA was observed [97]. Likewise, the same study also reported an 8.5% decrease in the CSA of the semitendinosus muscle at 30% of the muscle length, with no significant changes being observed at 50% or 70% of the muscle length. Hence, when taken together, it appears that all models of disuse can induce radial atrophy at the whole muscle level, and the magnitude of the radial atrophy can vary along the length of the muscle.
4.2. Muscle fascicles
As a basic guideline, it is helpful to consider that disuse-induced changes that occur at one level of muscle structure will be mediated by the changes that occur at the preceding level within the hierarchy of the structural elements (e.g., myofilaments → myofibrils → fibers → fascicles → whole muscle). According to this guideline, the longitudinal and radial atrophy that occurs at the whole muscle level will be mediated by the changes that occur at the level of the fascicles, and when evaluating how changes at the level of the fascicles could influence the whole muscle structure, the architectural properties of the muscle must be kept in mind. The importance of this point cannot be understated because it would be very counterintuitive to propose that robust longitudinal atrophy at the whole muscle level could be mediated exclusively by radial atrophy of the fascicles, or that robust radial atrophy at the whole muscle level could be mediated exclusively by longitudinal atrophy of the fascicles. However, as illustrated in Figure 2, these are both very realistic possibilities.
Readers who are not familiar with the geometric principles described in Figure 2 are strongly encouraged to view the work of Maxwell et al. (1974) because further consideration of this work will reveal that a decrease in whole muscle length and/or whole muscle diameter/CSA could be mediated by radial atrophy of the fascicles and/or longitudinal atrophy of the fascicles, and the specific mechanism(s) involved will depend on the inherent architectural properties of the muscle as well as the physical constraints under which the adaptations occur [108]. For instance, the examples shown in Figure 2A vs. 2C illustrate that when whole muscle length is considered as a fixed variable, a 25% decrease in fascicle length could lead to a 34% decrease in the whole muscle diameter but no change in the whole muscle length or the pennation angle of the fascicles. Alternatively, when whole muscle length is not considered as a fixed variable, a 25% decrease in fascicle length could lead to a 14% decrease in whole muscle length, a 20% decrease in whole muscle diameter, and a 16% increase in the pennation angle of the fascicles. From a geometric perspective, there are an endless number of possibilities and thus the key point here is worth repeating: a decrease in whole muscle length and/or whole muscle diameter/CSA could be mediated by radial atrophy of the fascicles and/or longitudinal atrophy of the fascicles, and the specific mechanism(s) involved will depend on the inherent architectural properties of the muscle as well as the physical constraints under which the adaptations occur.
Having described how longitudinal atrophy of fascicles could induce changes in whole muscle length and/or whole muscle CSA, we will now address whether this type of atrophy occurs in response to disuse. Surprisingly, we could not find any animal-based studies that have examined the effects of disuse on fascicle length. However, numerous studies in humans have reported such measurements and have shown that longitudinal atrophy of the fascicles occurs in response to various forms of disuse including space flight, bed rest, and unilateral lower limb suspension [97,99,109,110,111,112,113,114,115]. Importantly, although it appears that longitudinal atrophy at the whole muscle level only transpires when disuse occurs in conjunction with a chronic shortening of the muscle, longitudinal atrophy at the level of the fascicles can be observed when muscles are subjected to disuse in a predominantly neutral position [99,113,114]. For instance, unilateral lower limb suspension maintains the knee and ankle in a natural/neutral position, and work by De Boer et al. (2007) revealed that the length of fascicles in the vastus lateralis muscle decreased by 8% after 23 days of unilateral lower limb suspension [99]. Likewise, Campbell et al. (2013), reported that 23 days of unilateral lower limb suspension led to a 6-11% decrease in fascicle length in all four heads of the quadriceps [113].
Longitudinal atrophy at the level of the fascicles has also been reported to occur with bed rest [97,109,110,112]. As a case in point, Reeves et al. (2002) reported that 90 days of bed rest led to a 10% decrease in the fascicle length of the gastrocnemius [109]. However, the length at which various muscles are maintained during bedrest has not been described. During typical sleep postures, the ankle will assume a plantarflexed position while the position of the knee can be highly variable [116,117,118,119]. If these positions are dominant during the awake portion of bedrest, then it could be assumed that the plantarflexors are maintained in a shortened position whereas the position of the knee flexors and extensors would be unpredictable. With this point in mind, one is faced with the question of whether the bedrest-induced decrease in fascicle length reported by Reeves et al. (2002) was dependent on a chronic shortening of the muscle [109]. To date, no studies have directly addressed this question, however, De Boer et al. (2008) reported a 6% decrease in the fascicle length of the vastus lateralis muscle after five weeks of bed rest [110], and Sarto et al. (2021) reported a 2.8% decrease in fascicle length of the biceps femoris after just 10 days of bed rest [112]. The vastus lateralis and biceps femoris exert opposing actions on the position of the knee, and thus, if one muscle is maintained in a shortened position then the other would have to be maintained in the lengthened position. Yet, both muscles revealed a bedrest-induced decrease in fascicle length. Thus, the available evidence indicates that disuse leads to the longitudinal atrophy of fascicles, this phenomenon does not appear to be dependent on a chronic shortening of the muscle, and according to Maxwell’s geometric model, the longitudinal atrophy of fascicles could lead to changes in whole muscle length and/or whole muscle CSA.
Just like longitudinal atrophy, radial atrophy of fascicles could also induce changes in whole muscle length and/or whole muscle CSA (Figure 2), but whether disuse leads to radial atrophy of fascicles has not been reported. Nonetheless, a multitude of studies have examined changes in the pennation angle of fascicles and, as illustrated by Maxwell et al. (1974), there is a direct relationship between fascicle diameter and the pennation angle of the fascicles [99,108,110,113,114,115]. Specifically, Maxwell’s geometric model indicates that when muscle length is a fixed variable, then a decrease in fascicle diameter will lead to a decrease in the pennation angle of the fascicles and vice versa (Figure 2E). As described in section 4.1, it is generally thought changes in whole muscle length do not occur when muscles are subjected to disuse in a neutral position. Thus, if a decrease in pennation angle is observed in muscles subjected to disuse in a neutral position, then it can be inferred that the decrease in pennation angle was due to a decrease in the diameter of the fascicles (i.e., radial atrophy of the fascicles). Fortunately, several studies have reported such an effect [99,113,114]. For example, Campbell et al. (2013), reported that, after 23 days of unilateral lower limb suspension, the pennation angle of the fascicles in all four heads of the quadriceps decreased by 7-10% [113]. Moreover, the work of De Boer et al. (2007) revealed that the pennation angle of the fascicles in the vastus lateralis muscle decreased by 7.6% after 23 days of unilateral lower limb suspension [99].
In addition to the aforementioned studies, a decrease in the pennation angle of fascicles has also been reported when the average position of the muscle during disuse was less clear (e.g., during bed rest) [109,110,111,115]. As mentioned above, the plantarflexor muscles are likely maintained in a shortened position during bed rest and therefore the length of the plantarflexor muscles would be expected to either slightly decrease or remain unchanged. According to Maxwell’s geometric model, if the length of the muscle did not change during bed rest then the decrease in the pennation angle of the fascicles would be directly mediated by a decrease in fascicle diameter. Importantly, however, if the length of the muscle decreased, then a decrease in the pennation angle of the fascicles would likely underrepresent the actual decrease in fascicle diameter that occurred [108]. Thus, although no studies have directly addressed whether disuse leads to radial atrophy of fascicles, inferences can be made and such inferences consistently indicate that disuse leads to the radial atrophy of fascicles.
5. Disuse-Induced Atrophy at the Microscopic Level
As mentioned above, disuse-induced changes that occur at one level of muscle structure reflect the changes that occur at the preceding level within the hierarchy of the structural elements. Thus, having established that disuse atrophy can lead to longitudinal and/or radial atrophy of the fascicles, we will now describe how these alterations are driven by microscopic changes that occur at the level of the muscle fibers.
5.1. Longitudinal Atrophy of Fascicles
Fascicles consist of a bundle of muscle fibers and, in general, the muscle fibers run the entire length of the fascicles [120,121]. For such fascicles, longitudinal atrophy would most likely be mediated by a decrease in the length of the individual fibers. However, in some muscles, fascicles are composed of muscle fibers that only run a fraction of the fascicle length. In such fascicles, the muscle fibers possess intrafascicular terminations which consist of tapered ends that interdigitate with the tapered ends of other fibers within the fascicle [122,123,124,125]. In theory, longitudinal atrophy of fascicles with intrafascicular terminations could be mediated by a decrease in the length of the individual muscle fibers and/or the loss of muscle fibers in series. However, we are not aware of any studies that have addressed whether disuse-induced atrophy can lead to the loss of muscle fibers in series, and therefore, this section will focus on whether disuse leads to a decrease in muscle fiber length.
Despite an exhaustive search, we could not find any human-based studies that have directly investigated the effects of disuse on muscle fiber length. Nevertheless, numerous animal-based studies have shown that when muscles are subjected to disuse in a shortened position it leads to a decrease in fiber length [81,82,84,126,127,128,129]. For instance, Widrick et al. (2008), reported an 11% decrease in the length of soleus muscle fibers following 15 days of hindlimb unloading [128]. Likewise, immobilization of the soleus and medial gastrocnemius muscles in a shortened position for four weeks led to 14% and 26% decreases in the length of the muscle fibers, respectively [81]. Similarly, Witzmann et al. (1982) found that immobilization of the soleus in a shortened position for 42 days led to a 27% decrease in fiber length [82].
Although a consistent body of literature indicates that disuse in a shortened position leads to a decrease in fiber length (i.e., longitudinal atrophy of the fibers), whether the same effect happens when muscles are subjected to disuse in a neutral position is not clear. Specifically, we are only aware of one study that has directly addressed this question, and in this study, Spector et al. (1982) concluded that the length of the fibers in the soleus, medial gastrocnemius, and tibialis anterior muscles of rats was not significantly altered after 4 weeks of immobilization in a neutral position [81]. However, it bears noting that the rats in this study were quite young (e.g., 200–240-gram males, age not specified but estimated to be 6-7 weeks) [130]. This is noteworthy because the longitudinal growth of bones in male rats continues until approximately 20 weeks of age [131]. Thus, in the work by Spector et al. (1982), the limb of the rats may have been immobilized with the muscles in a neutral position, but the muscles were likely still experiencing a chronic lengthening stimulus from the longitudinal growth of the bones. As such, the results from Spector et al. (1982) should be interpreted with caution. Continuing with this point in mind, it is important to acknowledge the studies from the previous section which indicated that longitudinal atrophy of the fascicles occurs when muscles are subjected to disuse in a neutral position [99,113,114]. Specifically, in these studies, a decrease in fascicle length was observed in several muscles including all four heads of the quadriceps and the lateral head of the gastrocnemius. Available evidence indicates that, in all of these muscles, the fibers run the entire length of the fascicles (i.e., there are no intrafascicular terminations) [64,132,133]. Thus, it would seem fair to argue that the disuse-induced decrease in fascicle length was mediated by longitudinal atrophy of the fibers. However, we recognize that this argument is based purely on speculation and the only responsible conclusion that can be drawn from this section is that additional studies are needed to determine whether disuse in a neutral position leads to longitudinal atrophy of the muscle fibers.
5.2. Radial Atrophy of Fascicles
At the microscopic level, there are two primary mechanisms via which radial atrophy of fascicles could be mediated: (i) a decrease in the CSA of the existing muscle fibers (i.e., radial atrophy of the muscle fibers, Figure 3A), and/or (ii) a decrease in the number of muscle fibers per cross-section (i.e., muscle fiber hypoplasia, Figure 3B). These mechanisms have been investigated in the context of disuse-induced atrophy, and the available literature on these topics will be summarized in the following sections. However, before doing so, it should be noted that, in fascicles with intrafascicular terminations, radial atrophy could also result from the longitudinal atrophy of muscle fibers (Figure 3C). Despite this possibility, we are not aware of any studies that have addressed it, and therefore, this potential mechanism will not be further discussed.
5.2.1. Radial Atrophy of Muscle Fibers
Radial atrophy of muscle fibers is defined as a decrease in the CSA of the muscle fibers. Having illustrated how radial atrophy of the muscle fibers can lead to radial atrophy of fascicles, we will now consider the studies that have investigated whether it occurs in response to disuse. Fortunately, this subject has been extensively studied in all models of disuse atrophy described in section 2 including, space flight [134,135,136,137,138], bed rest [139,140,141], unilateral lower limb suspension [98,113,142], hindlimb unloading [47,128,143,144,145,146], and immobilization [84,106,147,148,149], and the results of these studies consistently demonstrate that disuse leads to radial atrophy of the muscle fibers. For instance, Arentson-Lantz et al. (2016), reported a 24% decrease in the CSA of human vastus lateralis muscle fibers after 14 days of bed rest [141]. Likewise, mice flown aboard the space shuttle for 13 days showed a 39% decrease in the CSA of the gastrocnemius muscle fibers [138]. In addition, Desplanches et al. (1990), reported that five weeks of hindlimb suspension led to a 75% decrease in the CSA of the soleus muscle fibers in rats [143]. Thus, when taken together, it appears that radial atrophy of fascicles could be mediated, at least in part, by radial atrophy of the muscle fibers.
5.2.2. Changes in the number of muscle fibers per cross-section
Hypoplasia refers to the loss of muscle fibers, and as illustrated in Figure 3B, hypoplasia could promote radial atrophy of the muscle fascicles. However, there are conflicting reports with regard to whether disuse induces hypoplasia, and this conflict can be attributed to the different types of methods that have been used to address the question. Specifically, two primary methods have been employed: (i) counting the total number of muscle fibers per mid-belly cross-section of the muscle [150,151,152], or (ii) digestion of the muscle’s connective tissue followed by direct counting of the total number of muscle fibers per muscle [147,153]. The direct counting method should be the most accurate, but the use of this method requires the manual dissociation and counting of thousands of muscle fibers per muscle. Consequently, most studies simply count the number of muscle fibers per mid-belly cross-section and assume that this is reflective of the total fiber number. However, it is critical to appreciate that changes in the architectural properties of the muscle (e.g., fiber length, fiber CSA, etc.) can dramatically alter the number of muscle fibers that appear in a mid-belly cross-section. Therefore, to help the reader appreciate this, we developed to-scale replicas of the soleus and plantaris muscles of the rat (Figure 4). As detailed in the figure legend, the replicas are based on architectural properties that were reported in previous studies [84,127,128,143,154,155,156], and with these replicas, we used Maxwell’s geometric model to examine how a 16% decrease in fiber length [81,82,84,127,128] and/or a 61% decrease in fiber CSA [84,127,128,143] would affect the number of muscle fibers that appear in a mid-belly cross-section. The first thing for the reader to note in this figure is that, after zooming in on panel 4B, it should be evident that all of the fibers within the soleus muscle pass through the mid-belly of the muscle (hatched box). The reason for this is that, in the soleus, [fiber length * cosine θ] is greater than one-half of the muscle length. Importantly, in muscles that exceed this critical threshold value, changes in fiber length and/or fiber CSA will not affect the number of fibers that appear in a mid-belly cross-section (Figure 4C-D). Nonetheless, in many muscles (e.g., the plantaris), [fiber length * cosine θ] does not exceed one-half of the muscle length. When this critical threshold is not exceeded, all of the fibers in the muscle will not pass through the mid-belly and, as illustrated in Figure 4B-D, changes in fiber length and/or fiber CSA of such muscles can dramatically alter the number of fibers that appear in a mid-belly cross-section. For instance, in the rat plantaris, Maxwell’s geometric model predicts that a 16% decrease in fiber length would lead to a 41% decrease in the number of fibers in a mid-belly cross-section. Thus, without an appreciation of the architectural properties of the muscle, one might mistakenly conclude the decrease in the number of fibers per cross-section was indicative of hypoplasia when, in reality, it was exclusively mediated by longitudinal atrophy of the pre-existing fibers.
Having illustrated how the number of fibers per cross-section can be influenced by the architectural properties of the muscle, we will now consider the studies that have investigated whether disuse induces hypoplasia. To the best of our knowledge, no human studies have addressed this question and there are only a handful of animal-based studies that have investigated it [147,150,151,152,153]. For instance, using the direct counting method, Nicks et al., (1989) concluded that the total number of fibers in the long head of the triceps brachii muscle of rats was not altered after eight weeks of forelimb immobilization [147]. Likewise, Templeton et al. (1988) found that four weeks of hindlimb suspension did not alter the number of fibers in the rat soleus [153]. Consistent with this finding, Cardenas et al. (1977) subjected the soleus muscle of rats to four weeks of immobilization in a shortened position and, after counting the number of fibers per mid-belly cross-section, concluded that the disuse did not alter the number of fibers per muscle [151]. Indeed, we are only aware of one study that has reported disuse-induced hypoplasia. Specifically, in this study, Booth and Kelse (1973) subjected the soleus muscle of rats to four weeks of immobilization in a neutral position [150]. The number of muscle fibers per cross-section was then counted and, in contrast to the work of Carenas et al. (1977), it was concluded that immobilization led to a 24% reduction in the number of fibers. Importantly, in this study, the region of the muscle via which the cross-sections were obtained was not specified, and this is important because, as highlighted in Figure 4B, taking a cross-section from the soleus at a region that is proximal or distal to the mid-belly could lead one to mistakenly conclude that there was disuse-induced hypoplasia. Thus, after considering this explanation, we feel fairly confident in concluding that disuse does not induce hypoplasia.
6. Disuse-Induced Atrophy at the Ultrastructural Level
Up to this point, we have concluded that disuse can induce both longitudinal and radial atrophy of muscle fibers. According to our basic guideline, these changes will be mediated by the alterations that occur at the preceding level within the hierarchy of the structural elements. In this case, the preceding level represents the ultrastructure of the muscle, and at this level, one will find that ~80% of the muscle fiber volume is composed of myofibrils [70,71]. Given that myofibrils make up the bulk of the muscle fiber volume, it can be reasoned that longitudinal and/or radial atrophy of the muscle fibers will be largely mediated by changes that occur at the level of the myofibrils. Accordingly, in this section, we will explore how disuse induces atrophy at the level of the myofibrils.
6.1. Longitudinal Atrophy of Muscle Fibers
As summarized in the previous section, a compelling body of studies has shown that disuse in a shortened position can induce the longitudinal atrophy of the muscle fibers. Since muscle fibers are filled with myofibrils, and the myofibrils are composed of an in-series repetition of sarcomeres (Figure 1A), it follows that a decrease in the length of muscle fibers should be driven by a decrease in the length of the sarcomeres and/or the loss of in-series sarcomeres. When considering these two possibilities it is important to recognize that if the decrease in fiber length were mediated by a decrease in sarcomere length, then the shortened sarcomeres would not be able to function at their optimal length for force production [82,157]. This point, coupled with the fact that the resting/optimal length of sarcomeres (2.0-2.5 µm) is highly conserved within a given species [72,73], suggests that the muscle fibers will be faced with a strong drive to restore the resting/optimal length of its sarcomeres. To accomplish this, the muscle fibers would have to eliminate in-series sarcomeres, and, not surprisingly, this is what has been observed in numerous studies [41,82,84,126,127,129,148,158,159]. For instance, Shah et al. (2001) reported that 28 days of immobilizing the mouse soleus muscle in a shortened position led to a 26% decrease in fiber length and this was accompanied by a 26% decrease in the number of in-series sarcomeres [129]. Likewise, Tabary et al. (1972) found that 21-43 days of immobilizing the soleus muscle of cats in a shortened position led to an ~34% decrease in fiber length and that this was associated with a ~40% reduction in the number of in-series sarcomeres [126]. In addition to immobilization, similar decrements in both fiber length and the number of in-series sarcomeres have also been reported to occur in response to hindlimb suspension [41,127]. Thus, it can be strongly concluded that when muscles are subjected to disuse in a shortened position it will lead to longitudinal atrophy of the muscle fibers and this effect is largely mediated by the elimination of in-series sarcomeres. With that being said, it bears noting that all of the aforementioned studies were focused on muscles that were subjected to disuse in a shortened position, and whether the same phenomena happens when muscles are subjected to disuse in a neutral position has, to the best of our knowledge, not been addressed.
Having established that disuse can lead to the elimination of in-series sarcomeres, one is faced with the question of how this occurs. Unfortunately, to date, the mechanism(s) that drive this phenomenon remain far from defined. Nonetheless, multiple studies have suggested that the elimination of in-series sarcomeres might occur in regions of myofibrillar disruption that are generally referred to as: (i) central core-like lesions (CCLs) and (ii) segmental necrosis.
CCLs are the most widely reported type of myofibrillar disruption [42,43,47,54,146,160,161,162,163,164,165,166,167], and conventional CCLs can be described as centrally located circular disruptions of the myofibrils within a cross-section of a muscle fiber (Figure 5A). However, less widely appreciated patches of irregularly shaped CCL-like disruptions can also be found throughout the entirety muscle fibers that are undergoing disuse-induced atrophy (Figure 5A-C) [42,43,167]. In both cases, a close examination of these regions reveals that they are enriched with signs of active remodeling including a disarray of the myofilaments, loss of mitochondria, and fragmentation of Z-lines [42,43,54,160,162]. Interestingly, a large number of studies have reported that CCLs are very common in muscles that have been subjected to disuse in a shortened position [42,43,47,54,146,163,164,165,166,167]. Yet, such regions are rarely found in muscles that have been subjected to immobilization in a lengthened position [47,54,164]. This is an important point because when muscles are subjected to immobilization in a lengthened position, they will generally still undergo atrophy, but the atrophic response will not involve the in-series elimination of sarcomeres [81,126,129,158,159]. Thus, the presence of CCLs appears to occur specifically in response to atrophic conditions that lead to the in-series elimination of sarcomeres.
The aforementioned points are all consistent with the notion that the elimination of in-series sarcomeres occurs at CCLs. However, numerous studies have also reported that CCLs are largely confined to Type I fibers [43,47,54,146,163,164,165]. For example, Riley et al. (1990), subjected the soleus muscle of rats to 13 days of hindlimb suspension and found that ~92% of the fibers with CCLs were Type I fibers [43]. To the best of our knowledge, there is no evidence, or reason to think, that the loss of in-series sarcomeres would be confined to Type I fibers. Thus, although intriguing, the notion that the elimination of in-series sarcomeres occurs at CCLs remains highly questionable and additional studies will be needed to resolve this issue. Moreover, although conventional CCLs and patches of CCL-like disruptions both show signs of active remodeling, they might be involved in distinct biological processes. Indeed, Murakami et al. (2008) reported that irregularly shaped areas of ‘sarcomeric disarray’ (i.e., patches of CCL-like disruptions) were readily apparent in rat soleus muscles after 7 days of hindlimb suspension, but the presence of conventional CCLs only became apparent after 14 days of hindlimb suspension [167]. The different temporal nature of their appearance suggests that convential CCLs and patches of CCL-like disruptions could represent biologically distinct areas of remodeling, and thus, future studies should take care to classify these areas as different types of myofibrillar disruptions. Consistent with the nomenclature of Murakami et al. (2008), we propose that the centrally located circular disruptions should be classified as conventional CCLs, and the irregularly shaped CCL-like disruptions should be referred to as areas of sarcomeric disarray (SDA).
The other type of myofibrillar disruption that has been implicated in the in-series elimination of sarcomeres is called segmental necrosis [42,43,162,166,168,169]. Unlike CCLs, segmental necrosis typically presents as a destruction of an entire segment of the muscle fiber, and is associated with the infiltration of mononuclear cells and neutrophils as well as signs of elevated phagocytic activity [162,168,169]. Two examples of segmental necrosis are shown in Figure 5D-E, and it has been proposed that these regions could serve as areas in which large segments of in-series sarcomere are eliminated [166,168]. Importantly, just like CCLs, areas of segmental necrosis are generally confined to muscles that have been subjected to disuse in a shortened position, but their appearance is quite rare when compared with CCLs. For instance, Riley et al. (1990) reported that only ~3% of rat soleus muscle fibers possessed regions of segmental necrosis after 10 days of hindlimb suspension, whereas ~30% of the fibers showed signs of CCLs after 13 days [43]. Undoubtedly, there is still much to learn about the function of CCLs and areas of segmental necrosis, and the mechanisms that give rise to them. The only thing that we can confidently conclude at this point is that additional studies will be needed to determine whether these are truly the sites in which in-series sarcomeres are eliminated during the longitudinal atrophy of muscle fibers.
6.2. Radial Atrophy of Muscle Fibers
In section 5.2 we reviewed the evidence which indicates that radial atrophy of the muscle fibers is a major contributor to disuse-induced atrophy. Since the bulk of the muscle fiber volume is composed of myofibrils, it can be reasoned that radial atrophy of the muscle fibers will primarily be mediated by either: (i) a decrease in the CSA of the existing myofibrils (i.e., radial atrophy of the myofibrils), and/or (ii) a decrease in the number of myofibrils per muscle fiber (i.e., myofibril hypoplasia) (Figure 6). Importantly, a loss of the interstitial elements (e.g., sarcoplasmic reticulum, mitochondria, etc.) could also contribute to the radial atrophy of the muscle but we are not aware of any studies that have addressed this possibility. Therefore, in this section, we will review the studies that have addressed whether the disuse-induced radial atrophy of muscle fibers is mediated by radial atrophy of the myofibrils and/or myofibril hypoplasia.
6.2.1. Radial Atrophy of Myofibrils
Radial atrophy of myofibrils is defined as a decrease in the CSA of the myofibrils. Having illustrated how radial atrophy of the myofibrils can lead to radial atrophy of the muscle fiber (Figure 6), we will now consider the studies that have investigated whether disuse leads to changes in myofibril CSA. Sadly, we could not find any studies that have directly measured myofibril CSA; however, there are at least two studies that reported measurements of myofibril diameter, and in both cases, it was concluded that disuse leads to a decrease in myofibril diameter [162,170]. For instance, Riley et al. (1990), reported a 23% decrease in the diameter of the myofibrils of the adductor longus muscles of rats after 12.5 days of space flight [162]. Likewise, Giordano et al. (2014), reported a 50% reduction in the diameter of myofibrils in the plantaris muscle of rats that had been subjected to 10 days of hindlimb suspension [170]. Consistent with these results, several qualitative studies have also reported that disuse leads to a reduction in myofibril diameter [43,171,172,173,174]. Hence, the available data suggest that radial atrophy of muscle fibers is mediated, at least in part, by radial atrophy of the myofibrils.
6.2.2. Myofibril Hypoplasia
Myofibril hypoplasia refers to the loss of myofibrils, and as illustrated in Figure 6, myofibril hypoplasia could also lead to the radial atrophy of the muscle fibers. However, we are not aware of any studies that have directly addressed this topic. Nevertheless, there is one study that, after some extrapolation, suggests that the radial atrophy of muscle fibers that occurs during space flight is not mediated by myofibril hypoplasia [162]. Specifically, Riley et al. (1990) reported that 12.5 days of space flight led to a 36% decrease in the CSA of the muscle fibers and that was accompanied by a 23% decrease in myofibril diameter. If myofibrils are viewed as round cylinders, then a 23% decrease in the myofibril diameter would be equivalent to a 41% decrease in the myofibril CSA. Hence, at least in this study, the decrease in muscle fiber CSA could be fully explained by the radial atrophy of the myofibrils, and therefore suggests that disuse did not lead to myofibril hypoplasia. Despite this point, there is a major gap in our knowledge about whether disuse leads to myofibril hypoplasia and this is certainly a topic that is worthy of further investigation.
6.2.3. Mechanisms that might promote the radial atrophy of myofibrils
As described above, current evidence indicates that the radial atrophy of muscle fibers during disuse is mediated, at least in part, by radial atrophy of the myofibrils. Since myofibrils are largely composed of thick and thin myofilaments, it can be inferred that radial atrophy of the myofibrils will be primarily mediated by the loss of these myofilaments. Importantly, the number of myofilaments per myofibril depends on the size of the myofibril and the density of the myofilaments. Thus, if the size of the myofibril gets smaller and the density of the myofilaments does not increase then the number of myofilaments must decrease. Fortunately, myofilament density has been investigated in several models of disuse atrophy, and none of these studies observed an increase in the density of either the thick or thin myofilaments [43,171,172,173,174,175]. Accordingly, it can be strongly concluded that the disuse-induced radial atrophy of myofibrils is associated with the loss of myofilaments.
In the simplest terms, a disuse-induced loss of myofilaments will occur when there is a negative balance between the rate of protein synthesis and the rate of protein degradation and a plethora of human and animal studies have shown that disuse leads to a decrease in the rate of protein synthesis [53,176,177,178,179,180,181,182,183,184,185,186,187,188,189,190,191,192,193,194,195,196]. For instance, nearly 50 years ago, Goldspink reported that immobilization of the rat soleus muscle in a shortened position led to a rapid (within 6 hr) and sustained (> 7 days) decrease in the rate of protein synthesis [176], and a multitude of similar observation have been reported when muscles are subjected to disuse in a neutral position. As a case in point, Gibson et al. (1987) reported that 5 weeks of immobilizing the quadriceps of humans in a neutral position led to a 26% decrease in the rate of protein synthesis [180]. Likewise, De Boer et al. (2007) found that 10 days of unilateral lower limb suspension led to a 53% decrease in the rate of myofibrillar protein synthesis in the quadriceps [186].
Although a clear and consistent body of evidence indicates that disuse leads to a decrease in the rate of protein synthesis, whether disuse alters the rate of protein degradation is questionable. For instance, some human and animal studies have revealed that disuse is associated with an increase in the rate of protein degradation [176,178,197,198], and numerous studies have found that disuse leads to an increase in the expression of genes that regulate the ubiquitin-proteasome pathway of protein degradation [106,199,200,201,202,203,204]. However, several studies have also shown that disuse does not alter the rate of protein degradation [182,189,205,206]. Given these contradictory results, we believe that the notion that disuse alters the rate of protein degradation should be viewed as a subject of debate [207,208,209].
Based on the studies described above, it seems fair to conclude that the disuse-induced loss of myofilaments is mediated, at least in part, by a decrease in the rate of protein synthesis. Given this conclusion, one is then faced with the question of where new myofilaments are synthesized and deposited within the myofibrils, as the answer to this question would provide insight into how disuse leads to a loss in the total number of myofilaments within the myofibril. New myofilaments are synthesized by ribosomes, and previous studies have reported that ribosomes are particularly prominent in the intermyofibrillar space, as well as in the subsarcolemmal and perinuclear regions of the fiber [210,211,212]. Importantly, these same studies have also reported that ribosomes are not found within the intramyofibrillar space. Thus, according to the positions of the ribosomes, one would expect new myofilaments to be synthesized and initially deposited at the periphery of the myofibrils. Consistent with this proposition, Dr. Morkin (1970) used 3H-leucine to label newly synthesized proteins and then used electron microscope-based autoradiography to identify where the newly synthesized proteins accumulated, and the results indicated that the deposition was primarily confined to the periphery of the myofibrils [213]. As such, it would appear that the radial atrophy of myofibrils that occurs during disuse is due, at least in part, to a decrease in the rate at which new myofilaments are synthesized and deposited at the periphery of myofibrils.
In addition to the above-mentioned conclusion, it also bears noting that a handful of studies have reported that muscles subjected to disuse have myofibrils that exhibit a moth-eaten appearance [43,171,173,175,214,215]. For instance, as shown in Figure 7, Riley et al. (1987) reported that 7 days of spaceflight led to an increase in the prevalence of gaps in the myofilaments around the periphery of the myofibrils, as well as an increase in the focal loss of myofilaments within the myofibrils [173]. The same type of peripheral and intramyofibrillar moth-eaten appearance has also been reported by others [43,175]. Hence, when taken together, these observations provide support for the notion that radial atrophy of myofibrils is not simply mediated by decreased myofilament deposition at the periphery of myofibrils, but also by the focal loss of myofilaments.
Having established that radial atrophy of myofibrils is associated with the focal loss of myofilaments, one is faced with questions about the mechanism that drives this phenomenon. To the best of our knowledge, no studies have addressed this topic, but we suspect that it may be attributed to a sub-group of myofilaments that have been referred to as easily releasable myofilaments (ERMs). Specifically, ERMs are a sub-group of myofilaments that can be easily released from the myofibrillar fraction of a muscle lysate via gentle agitation with an ATP-containing solution [216,217,218,219,220]. It is thought that the presence of ATP dissociates the actin-myosin interaction and facilitates the release of myofilaments that are weakly associated with the Z disc. For instance, Belcastro et al. (1991) reported that only ~2-4% of the myofilaments are released from rabbit myofibrils when they are agitated with a solution that does not contain ATP, and this proportion increases to ~28% when ATP is added to the agitation solution [219]. It has also been reported that ERMs contain the major constituents of the myofibrils “myosin heavy chain and actin”, but very little alpha-actinin and desmin [218,219,220], suggesting that the contractile myofilaments are more susceptible to release in an ATP-containing solution than the Z-line components and intermediate myofilaments. It also bears noting that previous studies have reported that the proportion of ERMs in muscle lysates increases during atrophic conditions such as food deprivation and sepsis [221,222,223], as well as when the myofibrils are pretreated with proteases [218]. Thus, the potential for ERMs to play a role in the disuse-induced loss of myofilaments is intriguing and seems worthy of further investigation.
7. Closing Remarks
As illustrated throughout this review, disuse can lead to a profound loss of skeletal muscle mass, and in Table 1 we have summarized the structural adaptations that are thought to mediate this response. In this table, we also list our opinion about the level of evidence that supports each of these adaptations, as well as some of the major gaps in knowledge that still exist. After reading this review we hope that the reader can appreciate that our knowledge about the structural adaptations that mediate disuse atrophy is far from complete. Indeed, a number of foundationally important questions have yet to be answered and, in many instances, we believe that the answers to these questions will be essential for the advancement of the field.
Acknowledgements
The work in this manuscript was supported by the National Institute of Arthritis and Musculoskeletal and Skin Diseases of the National Institutes of Health (NIH) under Awards AR074932 and AR082816 to T.A.H.
References
- Janssen, I.; Heymsfield, S.B.; Wang, Z.M.; Ross, R. Skeletal muscle mass and distribution in 468 men and women aged 18-88 yr. J Appl Physiol (1985) 2000, 89, 81-88. [CrossRef]
- Rowland, L.A.; Bal, N.C.; Periasamy, M. The role of skeletal-muscle-based thermogenic mechanisms in vertebrate endothermy. Biol Rev Camb Philos Soc 2015, 90, 1279-1297. [CrossRef]
- Thyfault, J.P.; Bergouignan, A. Exercise and metabolic health: beyond skeletal muscle. Diabetologia 2020, 63, 1464-1474. [CrossRef]
- Egan, B.; Zierath, J.R. Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Cell Metab 2013, 17, 162-184. [CrossRef]
- Severinsen, M.C.K.; Pedersen, B.K. Muscle-Organ Crosstalk: The Emerging Roles of Myokines. Endocrine reviews 2020, 41, 594-609. [CrossRef]
- Abramowitz, M.K.; Hall, C.B.; Amodu, A.; Sharma, D.; Androga, L.; Hawkins, M. Muscle mass, BMI, and mortality among adults in the United States: A population-based cohort study. PloS one 2018, 13, e0194697. [CrossRef]
- Zhang, X.; Wang, C.; Dou, Q.; Zhang, W.; Yang, Y.; Xie, X. Sarcopenia as a predictor of all-cause mortality among older nursing home residents: a systematic review and meta-analysis. BMJ Open 2018, 8, e021252. [CrossRef]
- Srikanthan, P.; Karlamangla, A.S. Relative muscle mass is inversely associated with insulin resistance and prediabetes. Findings from the third National Health and Nutrition Examination Survey. The Journal of clinical endocrinology and metabolism 2011, 96, 2898-2903. [CrossRef]
- Sizoo, D.; Stam, S.P.; de Heide, L.J.M.; Emous, M.; van Zutphen, T.; van Dijk, P.R.; van Beek, A.P. The association of low muscle mass with prevalence and incidence of type 2 diabetes in different BMI classes. Diabetes Res Clin Pract 2023, 195, 110197. [CrossRef]
- Wolfe, R.R. The underappreciated role of muscle in health and disease. Am J Clin Nutr 2006, 84, 475-482. [CrossRef]
- Tyrovolas, S.; Panagiotakos, D.; Georgousopoulou, E.; Chrysohoou, C.; Tousoulis, D.; Haro, J.M.; Pitsavos, C. Skeletal muscle mass in relation to 10 year cardiovascular disease incidence among middle aged and older adults: the ATTICA study. J Epidemiol Community Health 2020, 74, 26-31. [CrossRef]
- Seguin, R.; Nelson, M.E. The benefits of strength training for older adults. American journal of preventive medicine 2003, 25, 141-149. [CrossRef]
- Proctor, D.N.; Balagopal, P.; Nair, K.S. Age-related sarcopenia in humans is associated with reduced synthetic rates of specific muscle proteins. J Nutr 1998, 128, 351S-355S. [CrossRef]
- Pahor, M.; Kritchevsky, S. Research hypotheses on muscle wasting, aging, loss of function and disability. J Nutr Health Aging 1998, 2, 97-100.
- Goates, S.; Du, K.; Arensberg, M.B.; Gaillard, T.; Guralnik, J.; Pereira, S.L. Economic Impact of Hospitalizations in US Adults with Sarcopenia. J Frailty Aging 2019, 8, 93-99. [CrossRef]
- Bodine, S.C. Disuse-induced muscle wasting. Int J Biochem Cell Biol 2013, 45, 2200-2208. [CrossRef]
- Adams, G.R.; Bamman, M.M. Characterization and regulation of mechanical loading-induced compensatory muscle hypertrophy. Comprehensive Physiology 2012, 2, 2829-2870. [CrossRef]
- Hippocrates. The Genuine Works of Hippocrates; Dover New York, 1868.
- Bellar, A.; Welch, N.; Dasarathy, S. Exercise and physical activity in cirrhosis: opportunities or perils. J Appl Physiol (1985) 2020, 128, 1547-1567. [CrossRef]
- Pina, I.L.; Apstein, C.S.; Balady, G.J.; Belardinelli, R.; Chaitman, B.R.; Duscha, B.D.; Fletcher, B.J.; Fleg, J.L.; Myers, J.N.; Sullivan, M.J.; et al. Exercise and heart failure: A statement from the American Heart Association Committee on exercise, rehabilitation, and prevention. Circulation 2003, 107, 1210-1225. [CrossRef]
- Rausch, V.; Sala, V.; Penna, F.; Porporato, P.E.; Ghigo, A. Understanding the common mechanisms of heart and skeletal muscle wasting in cancer cachexia. Oncogenesis 2021, 10, 1. [CrossRef]
- Dasarathy, S. Cause and management of muscle wasting in chronic liver disease. Curr Opin Gastroenterol 2016, 32, 159-165. [CrossRef]
- Lavine, K.J.; Sierra, O.L. Skeletal muscle inflammation and atrophy in heart failure. Heart Fail Rev 2017, 22, 179-189. [CrossRef]
- Braun, T.P.; Marks, D.L. The regulation of muscle mass by endogenous glucocorticoids. Front Physiol 2015, 6, 12. [CrossRef]
- Vandenburgh, H.; Chromiak, J.; Shansky, J.; Del Tatto, M.; Lemaire, J. Space travel directly induces skeletal muscle atrophy. FASEB J 1999, 13, 1031-1038. [CrossRef]
- National Research, C.; Division on, E.; Physical, S.; Aeronautics; Space Engineering, B.; Space Studies, B.; Committee for the Decadal Survey on, B.; Physical Sciences in, S. Recapturing a Future for Space Exploration : Life and Physical Sciences Research for a New Era; National Academies Press: Washington, D.C., UNITED STATES, 2012.
- Lee, P.H.U.; Chung, M.; Ren, Z.; Mair, D.B.; Kim, D.H. Factors mediating spaceflight-induced skeletal muscle atrophy. Am J Physiol Cell Physiol 2022, 322, C567-C580. [CrossRef]
- Fitts, R.H.; Riley, D.R.; Widrick, J.J. Functional and structural adaptations of skeletal muscle to microgravity. The Journal of experimental biology 2001, 204, 3201-3208. [CrossRef]
- Clement, G.; Lestienne, F. Adaptive modifications of postural attitude in conditions of weightlessness. Exp Brain Res 1988, 72, 381-389. [CrossRef]
- Thornton, W.E., Hoffler, G.W. Rummel, J.A. Anthropometric changes and fluid shifts. Proc. of the Skylab Life Sci. Symp. 1974, Vol. 2.
- Wisdom, K.M.; Delp, S.L.; Kuhl, E. Use it or lose it: multiscale skeletal muscle adaptation to mechanical stimuli. Biomech Model Mechanobiol 2015, 14, 195-215. [CrossRef]
- Jorgenson, K.W.; Phillips, S.M.; Hornberger, T.A. Identifying the Structural Adaptations that Drive the Mechanical Load-Induced Growth of Skeletal Muscle: A Scoping Review. Cells 2020, 9. [CrossRef]
- Kruse, A.; Rivares, C.; Weide, G.; Tilp, M.; Jaspers, R.T. Stimuli for Adaptations in Muscle Length and the Length Range of Active Force Exertion-A Narrative Review. Front Physiol 2021, 12, 742034. [CrossRef]
- Zollner, A.M.; Abilez, O.J.; Bol, M.; Kuhl, E. Stretching skeletal muscle: chronic muscle lengthening through sarcomerogenesis. PloS one 2012, 7, e45661. [CrossRef]
- Hilton, J. On the influence of Mechanical and Physiological Rest in the Treatment of Accidents and Surgical Diseases, and the Diagnostic Value of Pain. Br Foreign Med Chir Rev 1864, 34, 53-58.
- Deitrick, J.E.; Whedon, G.D.; Shorr, E. Effects of immobilization upon various metabolic and physiologic functions of normal men. The American journal of medicine 1948, 4, 3-36. [CrossRef]
- LeBlanc, A.D.; Schneider, V.S.; Evans, H.J.; Pientok, C.; Rowe, R.; Spector, E. Regional changes in muscle mass following 17 weeks of bed rest. J Appl Physiol (1985) 1992, 73, 2172-2178. [CrossRef]
- Mulder, E.R.; Horstman, A.M.; Stegeman, D.F.; de Haan, A.; Belavy, D.L.; Miokovic, T.; Armbrecht, G.; Felsenberg, D.; Gerrits, K.H. Influence of vibration resistance training on knee extensor and plantar flexor size, strength, and contractile speed characteristics after 60 days of bed rest. J Appl Physiol (1985) 2009, 107, 1789-1798. [CrossRef]
- Hides, J.A.; Belavy, D.L.; Stanton, W.; Wilson, S.J.; Rittweger, J.; Felsenberg, D.; Richardson, C.A. Magnetic resonance imaging assessment of trunk muscles during prolonged bed rest. Spine (Phila Pa 1976) 2007, 32, 1687-1692. [CrossRef]
- LeBlanc, A.; Gogia, P.; Schneider, V.; Krebs, J.; Schonfeld, E.; Evans, H. Calf muscle area and strength changes after five weeks of horizontal bed rest. Am J Sports Med 1988, 16, 624-629. [CrossRef]
- Kawano, F.; Ishihara, A.; Stevens, J.L.; Wang, X.D.; Ohshima, S.; Horisaka, M.; Maeda, Y.; Nonaka, I.; Ohira, Y. Tension- and afferent input-associated responses of neuromuscular system of rats to hindlimb unloading and/or tenotomy. Am J Physiol Regul Integr Comp Physiol 2004, 287, R76-86. [CrossRef]
- Anzil, A.P.; Sancesario, G.; Massa, R.; Bernardi, G. Myofibrillar disruption in the rabbit soleus muscle after one-week hindlimb suspension. Muscle Nerve 1991, 14, 358-369. [CrossRef]
- Riley, D.A.; Slocum, G.R.; Bain, J.L.; Sedlak, F.R.; Sowa, T.E.; Mellender, J.W. Rat hindlimb unloading: soleus histochemistry, ultrastructure, and electromyography. J Appl Physiol (1985) 1990, 69, 58-66. [CrossRef]
- Morey-Holton, E.R.; Globus, R.K. Hindlimb unloading rodent model: technical aspects. J Appl Physiol (1985) 2002, 92, 1367-1377. [CrossRef]
- Hargens, A.R.; Vico, L. Long-duration bed rest as an analog to microgravity. J Appl Physiol (1985) 2016, 120, 891-903. [CrossRef]
- Ohira, Y. Effects of hindlimb suspension with stretched or shortened muscle length on contractile properties of rat soleus; 2000; Vol. 16, pp. S. 80-87.
- Fujita, N.; Arakawa, T.; Matsubara, T.; Ando, H.; Miki, A. Influence of fixed muscle length and contractile properties on atrophy and subsequent recovery in the rat soleus and plantaris muscles. Archives of Histology and Cytology 2009, 72, 151-163.
- Tesch, P.A.; Lundberg, T.R.; Fernandez-Gonzalo, R. Unilateral lower limb suspension: From subject selection to "omic" responses. J Appl Physiol (1985) 2016, 120, 1207-1214. [CrossRef]
- Hackney, K.J.; Ploutz-Snyder, L.L. Unilateral lower limb suspension: integrative physiological knowledge from the past 20 years (1991-2011). Eur J Appl Physiol 2012, 112, 9-22. [CrossRef]
- Yasuda, N.; Glover, E.I.; Phillips, S.M.; Isfort, R.J.; Tarnopolsky, M.A. Sex-based differences in skeletal muscle function and morphology with short-term limb immobilization. J Appl Physiol (1985) 2005, 99, 1085-1092. [CrossRef]
- Nedergaard, A.; Jespersen, J.G.; Pingel, J.; Christensen, B.; Sroczynski, N.; Langberg, H.; Kjaer, M.; Schjerling, P. Effects of 2 weeks lower limb immobilization and two separate rehabilitation regimens on gastrocnemius muscle protein turnover signaling and normalization genes. BMC Res Notes 2012, 5, 166. [CrossRef]
- Caron, A.Z.; Drouin, G.; Desrosiers, J.; Trensz, F.; Grenier, G. A novel hindlimb immobilization procedure for studying skeletal muscle atrophy and recovery in mouse. J Appl Physiol (1985) 2009, 106, 2049-2059. [CrossRef]
- You, J.S.; Anderson, G.B.; Dooley, M.S.; Hornberger, T.A. The role of mTOR signaling in the regulation of protein synthesis and muscle mass during immobilization in mice. Disease models & mechanisms 2015, 8, 1059-1069. [CrossRef]
- Bruce-Gregorios, J.; Chou, S.M. Core myofibers and related alterations induced in rats' soleus muscle by immobilization in shortened position. J Neurol Sci 1984, 63, 267-275. [CrossRef]
- Thomsen, P.; Luco, J.V. CHANGES OF WEIGHT AND NEUROMUSCULAR TRANSMISSION IN MUSCLES OF IMMOBILIZED JOINTS. Journal of Neurophysiology 1944, 7, 245-251. [CrossRef]
- Carraro, U.; Gargiulo, P.; Kern, H. Induced Muscle Fiber Regeneration in Permanent Skeletal Muscle Denervation: Implication for Functional Electrical Stimulation of Denervated Degenerated Muscles in Spinal Cord Injury. J Transl Sci Res 2017, 1.
- Borisov, A.B.; Carlson, B.M. Cell death in denervated skeletal muscle is distinct from classical apoptosis. Anat Rec 2000, 258, 305-318. [CrossRef]
- Dedkov, E.I.; Kostrominova, T.Y.; Borisov, A.B.; Carlson, B.M. Reparative myogenesis in long-term denervated skeletal muscles of adult rats results in a reduction of the satellite cell population. Anat Rec 2001, 263, 139-154. [CrossRef]
- Yoshimura, K.; Harii, K. A regenerative change during muscle adaptation to denervation in rats. J Surg Res 1999, 81, 139-146. [CrossRef]
- Frontera, W.R.; Ochala, J. Skeletal muscle: a brief review of structure and function. Calcif Tissue Int 2015, 96, 183-195. [CrossRef]
- Henderson, C.A.; Gomez, C.G.; Novak, S.M.; Mi-Mi, L.; Gregorio, C.C. Overview of the Muscle Cytoskeleton. Compr Physiol 2017, 7, 891-944. [CrossRef]
- Lieber, R.L. Skeletal muscle adaptability. I: Review of basic properties. Dev Med Child Neurol 1986, 28, 390-397. [CrossRef]
- Narici, M.; Franchi, M.; Maganaris, C. Muscle structural assembly and functional consequences. J Exp Biol 2016, 219, 276-284. [CrossRef]
- Friederich, J.A.; Brand, R.A. Muscle fiber architecture in the human lower limb. J Biomech 1990, 23, 91-95. [CrossRef]
- Lieber, R.L.; Friden, J. Functional and clinical significance of skeletal muscle architecture. Muscle Nerve 2000, 23, 1647-1666. [CrossRef]
- Biferali, B.; Proietti, D.; Mozzetta, C.; Madaro, L. Fibro-Adipogenic Progenitors Cross-Talk in Skeletal Muscle: The Social Network. Front Physiol 2019, 10, 1074. [CrossRef]
- Murray, I.R.; Baily, J.E.; Chen, W.C.W.; Dar, A.; Gonzalez, Z.N.; Jensen, A.R.; Petrigliano, F.A.; Deb, A.; Henderson, N.C. Skeletal and cardiac muscle pericytes: Functions and therapeutic potential. Pharmacol Ther 2017, 171, 65-74. [CrossRef]
- Tedesco, F.S.; Moyle, L.A.; Perdiguero, E. Muscle Interstitial Cells: A Brief Field Guide to Non-satellite Cell Populations in Skeletal Muscle. Methods Mol Biol 2017, 1556, 129-147. [CrossRef]
- Murach, K.A.; Fry, C.S.; Kirby, T.J.; Jackson, J.R.; Lee, J.D.; White, S.H.; Dupont-Versteegden, E.E.; McCarthy, J.J.; Peterson, C.A. Starring or Supporting Role? Satellite Cells and Skeletal Muscle Fiber Size Regulation. Physiology (Bethesda) 2018, 33, 26-38. [CrossRef]
- MacDougall, J.D.; Sale, D.G.; Elder, G.C.; Sutton, J.R. Muscle ultrastructural characteristics of elite powerlifters and bodybuilders. Eur J Appl Physiol Occup Physiol 1982, 48, 117-126. [CrossRef]
- Seiden, D. Quantitative analysis of muscle cell changes in compensatory hypertrophy and work-induced hypertrophy. Am J Anat 1976, 145, 459-465. [CrossRef]
- Burkholder, T.J.; Lieber, R.L. Sarcomere length operating range of vertebrate muscles during movement. J Exp Biol 2001, 204, 1529-1536. [CrossRef]
- Gordon, A.; Huxley, A.F.; Julian, F. The variation in isometric tension with sarcomere length in vertebrate muscle fibres. The Journal of physiology 1966, 184, 170-192. [CrossRef]
- Burkholder, T.J.; Fingado, B.; Baron, S.; Lieber, R.L. Relationship between muscle fiber types and sizes and muscle architectural properties in the mouse hindlimb. Journal of morphology 1994, 221, 177-190. [CrossRef]
- Hanson, J.; Huxley, H.E. Structural basis of the cross-striations in muscle. Nature 1953, 172, 530-532. [CrossRef]
- Huxley, A.F.; Niedergerke, R. Structural changes in muscle during contraction; interference microscopy of living muscle fibres. Nature 1954, 173, 971-973. [CrossRef]
- Huxley, H.; Hanson, J. Changes in the cross-striations of muscle during contraction and stretch and their structural interpretation. Nature 1954, 173, 973-976. [CrossRef]
- Anderson, M.; Green, W.T. Lengths of the femur and the tibia; norms derived from orthoroentgenograms of children from 5 years of age until epiphysial closure. Am J Dis Child (1911) 1948, 75, 279-290. [CrossRef]
- Beaucage, K.L.; Pollmann, S.I.; Sims, S.M.; Dixon, S.J.; Holdsworth, D.W. Quantitative in vivo micro-computed tomography for assessment of age-dependent changes in murine whole-body composition. Bone Rep 2016, 5, 70-80. [CrossRef]
- Berg, B.N.; Harmison, C.R. Growth, disease, and aging in the rat. J Gerontol 1957, 12, 370-377. [CrossRef]
- Spector, S.; Simard, C.; Fournier, M.; Sternlicht, E.; Edgerton, V. Architectural alterations of rat hind-limb skeletal muscles immobilized at different lengths. Experimental neurology 1982, 76, 94-110. [CrossRef]
- Witzmann, F.A.; Kim, D.; Fitts, R.H. Hindlimb immobilization: length-tension and contractile properties of skeletal muscle. Journal of applied physiology 1982, 53, 335-345. [CrossRef]
- Gossman, M.R.; Rose, S.J.; Sahrmann, S.A.; Katholi, C.R. Length and circumference measurements in one-joint and multijoint muscles in rabbits after immobilization. Physical therapy 1986, 66, 516-520. [CrossRef]
- Coutinho, E.; Gomes, A.; França, C.; Oishi, J.; Salvini, T. Effect of passive stretching on the immobilized soleus muscle fiber morphology. Brazilian journal of medical and biological research 2004, 37, 1853-1861. [CrossRef]
- Tesch, P.A.; Berg, H.E.; Bring, D.; Evans, H.J.; LeBlanc, A.D. Effects of 17-day spaceflight on knee extensor muscle function and size. European journal of applied physiology 2005, 93, 463-468. [CrossRef]
- Sandonà, D.; Desaphy, J.-F.; Camerino, G.M.; Bianchini, E.; Ciciliot, S.; Danieli-Betto, D.; Dobrowolny, G.; Furlan, S.; Germinario, E.; Goto, K. Adaptation of mouse skeletal muscle to long-term microgravity in the MDS mission. PloS one 2012, 7, e33232. [CrossRef]
- Berry, P.; Berry, I.; Manelfe, C. Magnetic resonance imaging evaluation of lower limb muscles during bed rest--a microgravity simulation model. Aviation, space, and environmental medicine 1993, 64, 212-218.
- Ferretti, G.; Antonutto, G.; Denis, C.; Hoppeler, H.; Minetti, A.E.; Narici, M.V.; Desplanches, D. The interplay of central and peripheral factors in limiting maximal O2 consumption in man after prolonged bed rest. The Journal of physiology 1997, 501, 677. [CrossRef]
- Mulder, E.R.; Stegeman, D.F.; Gerrits, K.; Paalman, M.; Rittweger, J.; Felsenberg, D.; De Haan, A. Strength, size and activation of knee extensors followed during 8 weeks of horizontal bed rest and the influence of a countermeasure. European journal of applied physiology 2006, 97, 706-715. [CrossRef]
- Berg, H.E.; Eiken, O.; Miklavcic, L.; Mekjavic, I.B. Hip, thigh and calf muscle atrophy and bone loss after 5-week bedrest inactivity. European journal of applied physiology 2007, 99, 283-289. [CrossRef]
- Mulder, E.R.; Horstman, A.M.; Stegeman, D.F.; De Haan, A.; Belavý, D.L.; Miokovic, T.; Armbrecht, G.; Felsenberg, D.; Gerrits, K.H. Influence of vibration resistance training on knee extensor and plantar flexor size, strength, and contractile speed characteristics after 60 days of bed rest. Journal of applied physiology 2009, 107, 1789-1798. [CrossRef]
- Miokovic, T.; Armbrecht, G.; Felsenberg, D.; Belavý, D.L. Heterogeneous atrophy occurs within individual lower limb muscles during 60 days of bed rest. Journal of Applied Physiology 2012, 113, 1545-1559. [CrossRef]
- Mulder, E.; Clément, G.; Linnarsson, D.; Paloski, W.; Wuyts, F.; Zange, J.; Frings-Meuthen, P.; Johannes, B.; Shushakov, V.; Grunewald, M. Musculoskeletal effects of 5 days of bed rest with and without locomotion replacement training. European journal of applied physiology 2015, 115, 727-738. [CrossRef]
- Dirks, M.L.; Wall, B.T.; van de Valk, B.; Holloway, T.M.; Holloway, G.P.; Chabowski, A.; Goossens, G.H.; van Loon, L.J. One week of bed rest leads to substantial muscle atrophy and induces whole-body insulin resistance in the absence of skeletal muscle lipid accumulation. Diabetes 2016, 65, 2862-2875. [CrossRef]
- Liphardt, A.M.; Bolte, V.; Eckstein, F.; Wirth, W.; Brüggemann, G.P.; Niehoff, A. Response of thigh muscle cross-sectional area to 21-days of bed rest with exercise and nutrition countermeasures. Translational Sports Medicine 2020, 3, 93-106. [CrossRef]
- Smeuninx, B.; Elhassan, Y.S.; Manolopoulos, K.N.; Sapey, E.; Rushton, A.B.; Edwards, S.J.; Morgan, P.T.; Philp, A.; Brook, M.S.; Gharahdaghi, N. The effect of short-term exercise prehabilitation on skeletal muscle protein synthesis and atrophy during bed rest in older men. Journal of cachexia, sarcopenia and muscle 2021, 12, 52-69. [CrossRef]
- Franchi, M.V.; Sarto, F.; Simunič, B.; Pišot, R.; Narici, M.V. Early Changes of Hamstrings Morphology and Contractile Properties During 10 Days of Complete Inactivity. Medicine and Science in Sports and Exercise 2022.
- Hather, B.M.; Adams, G.R.; Tesch, P.A.; Dudley, G.A. Skeletal muscle responses to lower limb suspension in humans. Journal of applied physiology 1992, 72, 1493-1498. [CrossRef]
- De Boer, M.D.; Maganaris, C.N.; Seynnes, O.R.; Rennie, M.J.; Narici, M.V. Time course of muscular, neural and tendinous adaptations to 23 day unilateral lower-limb suspension in young men. The Journal of physiology 2007, 583, 1079-1091.
- Abadi, A.; Glover, E.I.; Isfort, R.J.; Raha, S.; Safdar, A.; Yasuda, N.; Kaczor, J.J.; Melov, S.; Hubbard, A.; Qu, X. Limb immobilization induces a coordinate down-regulation of mitochondrial and other metabolic pathways in men and women. PloS one 2009, 4, e6518. [CrossRef]
- Vandenborne, K.; Elliott, M.A.; Walter, G.A.; Abdus, S.; Okereke, E.; Shaffer, M.; Tahernia, D.; Esterhai, J.L. Longitudinal study of skeletal muscle adaptations during immobilization and rehabilitation. Muscle & Nerve: Official Journal of the American Association of Electrodiagnostic Medicine 1998, 21, 1006-1012.
- Hespel, P.; Op't Eijnde, B.; Leemputte, M.V.; Ursø, B.; Greenhaff, P.L.; Labarque, V.; Dymarkowski, S.; Hecke, P.V.; Richter, E.A. Oral creatine supplementation facilitates the rehabilitation of disuse atrophy and alters the expression of muscle myogenic factors in humans. The Journal of physiology 2001, 536, 625-633. [CrossRef]
- Thom, J.; Thompson, M.; Ruell, P.; Bryant, G.; Fonda, J.; Harmer, A.; De Jonge, X.J.; Hunter, S. Effect of 10-day cast immobilization on sarcoplasmic reticulum calcium regulation in humans. Acta physiologica scandinavica 2001, 172, 141-147. [CrossRef]
- Yasuda, N.; Glover, E.I.; Phillips, S.M.; Isfort, R.J.; Tarnopolsky, M.A. Sex-based differences in skeletal muscle function and morphology with short-term limb immobilization. Journal of applied physiology 2005, 99, 1085-1092. [CrossRef]
- Psatha, M.; Wu, Z.; Gammie, F.M.; Ratkevicius, A.; Wackerhage, H.; Lee, J.H.; Redpath, T.W.; Gilbert, F.J.; Ashcroft, G.P.; Meakin, J.R. A longitudinal MRI study of muscle atrophy during lower leg immobilization following ankle fracture. Journal of Magnetic Resonance Imaging 2012, 35, 686-695. [CrossRef]
- Wall, B.T.; Dirks, M.L.; Snijders, T.; Senden, J.M.; Dolmans, J.; Van Loon, L.J. Substantial skeletal muscle loss occurs during only 5 days of disuse. Acta physiologica 2014, 210, 600-611. [CrossRef]
- Kilroe, S.P.; Fulford, J.; Jackman, S.R.; Van Loon, L.J.; Wall, B.T. Temporal muscle-specific disuse atrophy during one week of leg immobilization. 2019. [CrossRef]
- Maxwell, L.C.; Faulkner, J.A.; Hyatt, G.J. Estimation of number of fibers in guinea pig skeletal muscles. J Appl Physiol 1974, 37, 259-264. [CrossRef]
- Reeves, N.D.; Maganaris, C.N.; Ferretti, G.; Narici, M.V. Influence of simulated microgravity on human skeletal muscle architecture and function. In Proceedings of Life in Space for Life on Earth; pp. 373-374.
- De Boer, M.D.; Seynnes, O.R.; Di Prampero, P.E.; Pišot, R.; Mekjavić, I.B.; Biolo, G.; Narici, M.V. Effect of 5 weeks horizontal bed rest on human muscle thickness and architecture of weight bearing and non-weight bearing muscles. European journal of applied physiology 2008, 104, 401-407.
- Koryak, Y.A. Architectural and functional specifics of the human triceps surae muscle in vivo and its adaptation to microgravity. Journal of applied physiology 2019, 126, 880-893. [CrossRef]
- Sarto, F.; Monti, E.; Šimunič, B.; Pišot, R.; Narici, M.V.; Franchi, M.V. Changes in Biceps Femoris Long Head Fascicle Length after 10-d Bed Rest Assessed with Different Ultrasound Methods. Medicine and Science in Sports and Exercise 2021, 53, 1529-1536.
- Campbell, E.-L.; Seynnes, O.R.; Bottinelli, R.; McPhee, J.S.; Atherton, P.; Jones, D.; Butler-Browne, G.; Narici, M.V. Skeletal muscle adaptations to physical inactivity and subsequent retraining in young men. Biogerontology 2013, 14, 247-259. [CrossRef]
- Seynnes, O.R.; Maganaris, C.N.; De Boer, M.D.; Di Prampero, P.E.; Narici, M.V. Early structural adaptations to unloading in the human calf muscles. Acta physiologica 2008, 193, 265-274. [CrossRef]
- Narici, M.; Cerretelli, P. Changes in human muscle architecture in disuse-atrophy evaluated by ultrasound imaging. Journal of gravitational physiology: a journal of the International Society for Gravitational Physiology 1998, 5, P73-74.
- Cary, D.; Jacques, A.; Briffa, K. Examining relationships between sleep posture, waking spinal symptoms and quality of sleep: A cross sectional study. PloS one 2021, 16, e0260582. [CrossRef]
- Tetley, M. Instinctive sleeping and resting postures: an anthropological and zoological approach to treatment of low back and joint pain. BMJ 2000, 321, 1616-1618. [CrossRef]
- Skarpsno, E.S.; Mork, P.J.; Nilsen, T.I.L.; Holtermann, A. Sleep positions and nocturnal body movements based on free-living accelerometer recordings: association with demographics, lifestyle, and insomnia symptoms. Nat Sci Sleep 2017, 9, 267-275. [CrossRef]
- Cary, D.; Collinson, R.; Sterling, M.; Briffa, K. Examining the validity and reliability of a portable sleep posture assessment protocol, using infrared cameras, under a variety of light and bed cover situations in the home environment. Work 2019, 63, 291-298. [CrossRef]
- Paul, A.C. Muscle length affects the architecture and pattern of innervation differently in leg muscles of mouse, guinea pig, and rabbit compared to those of human and monkey muscles. The Anatomical Record: An Official Publication of the American Association of Anatomists 2001, 262, 301-309.
- Paul, A.C.; Rosenthal, N. Different modes of hypertrophy in skeletal muscle fibers. The Journal of cell biology 2002, 156, 751-760. [CrossRef]
- Loeb, G.; Pratt, C.; Chanaud, C.; Richmond, F. Distribution and innervation of short, interdigitated muscle fibers in parallel-fibered muscles of the cat hindlimb. journal of Morphology 1987, 191, 1-15.
- Gans, C.; Loeb, G.E.; De Vree, F. Architecture and consequent physiological properties of the semitendinosus muscle in domestic goats. journal of Morphology 1989, 199, 287-297. [CrossRef]
- Heron, M.I.; Richmond, F.J. In-series fiber architecture in long human muscles. Journal of Morphology 1993, 216, 35-45. [CrossRef]
- Richmond, F.; MacGillis, D.; Scott, D. Muscle-fiber compartmentalization in cat splenius muscles. Journal of Neurophysiology 1985, 53, 868-885. [CrossRef]
- Tabary, J.; Tabary, C.; Tardieu, C.; Tardieu, G.; Goldspink, G. Physiological and structural changes in the cat's soleus muscle due to immobilization at different lengths by plaster casts. The Journal of physiology 1972, 224, 231-244.
- Wang, X.; Kawano, F.; Matsuoka, Y.; Fukunaga, K.; Terada, M.; Sudoh, M.; Ishihara, A.; Ohira, Y. Mechanical load-dependent regulation of satellite cell and fiber size in rat soleus muscle. American Journal of Physiology-Cell Physiology 2006, 290, C981-C989. [CrossRef]
- Widrick, J.J.; Maddalozzo, G.F.; Hu, H.; Herron, J.C.; Iwaniec, U.T.; Turner, R.T. Detrimental effects of reloading recovery on force, shortening velocity, and power of soleus muscles from hindlimb-unloaded rats. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 2008, 295, R1585-R1592. [CrossRef]
- Shah, S.B.; Peters, D.; Jordan, K.A.; Milner, D.J.; Fridén, J.; Capetanaki, Y.; Lieber, R.L. Sarcomere number regulation maintained after immobilization in desmin-null mouse skeletal muscle. Journal of Experimental Biology 2001, 204, 1703-1710. [CrossRef]
- Coelho, M.S.; Passadore, M.D.; Gasparetti, A.L.; Bibancos, T.; Prada, P.O.; Furukawa, L.L.; Furukawa, L.N.; Fukui, R.T.; Casarini, D.E.; Saad, M.J.; et al. High- or low-salt diet from weaning to adulthood: effect on body weight, food intake and energy balance in rats. Nutr Metab Cardiovasc Dis 2006, 16, 148-155. [CrossRef]
- Horton, J.A.; Bariteau, J.T.; Loomis, R.M.; Strauss, J.A.; Damron, T.A. Ontogeny of skeletal maturation in the juvenile rat. Anat Rec (Hoboken) 2008, 291, 283-292. [CrossRef]
- Wickiewicz, T.L.; Roy, R.R.; Powell, P.L.; Edgerton, V.R. Muscle architecture of the human lower limb. Clinical Orthopaedics and Related Research® 1983, 179, 275-283.
- Ward, S.R.; Eng, C.M.; Smallwood, L.H.; Lieber, R.L. Are current measurements of lower extremity muscle architecture accurate? Clinical orthopaedics and related research 2009, 467, 1074-1082. [CrossRef]
- Edgerton, V.; Zhou, M.; Ohira, Y.; Klitgaard, H.; Jiang, B.; Bell, G.; Harris, B.; Saltin, B.; Gollnick, P.; Roy, R. Human fiber size and enzymatic properties after 5 and 11 days of spaceflight. Journal of Applied Physiology 1995, 78, 1733-1739. [CrossRef]
- Widrick, J.J.; Knuth, S.T.; Norenberg, K.M.; Romatowski, J.; Bain, J.L.; Riley, D.A.; Karhanek, M.; Trappe, S.; Trappe, T.A.; Costill, D. Effect of a 17 day spaceflight on contractile properties of human soleus muscle fibres. Journal of Physiology 1999. [CrossRef]
- Belozerova, I.; Nemirovskaya, T.; Shenkman, B.; Kozlovskaya, I. Characteristic of changes in the structure and metabolism of the vastus lateralis muscles in monkeys after space flight. Neuroscience and Behavioral Physiology 2003, 33, 735-740. [CrossRef]
- Fitts, R.; Trappe, S.; Costill, D.; Gallagher, P.M.; Creer, A.C.; Colloton, P.; Peters, J.R.; Romatowski, J.; Bain, J.; Riley, D.A. Prolonged space flight-induced alterations in the structure and function of human skeletal muscle fibres. The Journal of physiology 2010, 588, 3567-3592. [CrossRef]
- Sung, M.; Li, J.; Spieker, A.; Spatz, J.; Ellman, R.; Ferguson, V.; Bateman, T.; Rosen, G.; Bouxsein, M.; Rutkove, S. Spaceflight and hind limb unloading induce similar changes in electrical impedance characteristics of mouse gastrocnemius muscle. Journal of musculoskeletal & neuronal interactions 2013, 13, 405.
- Berg, H.; Larsson, L.; Tesch, P. Lower limb skeletal muscle function after 6 wk of bed rest. Journal of applied physiology 1997.
- Yamashita-Goto, K.; Okuyama, R.; Honda, M.; Kawasaki, K.; Fujita, K.; Yamada, T.; Nonaka, I.; Ohira, Y.; Yoshioka, T. Maximal and submaximal forces of slow fibers in human soleus after bed rest. Journal of Applied Physiology 2001, 91, 417-424. [CrossRef]
- Arentson-Lantz, E.J.; English, K.L.; Paddon-Jones, D.; Fry, C.S. Fourteen days of bed rest induces a decline in satellite cell content and robust atrophy of skeletal muscle fibers in middle-aged adults. Journal of applied physiology 2016, 120, 965-975. [CrossRef]
- Brocca, L.; Longa, E.; Cannavino, J.; Seynnes, O.; de Vito, G.; McPhee, J.; Narici, M.; Pellegrino, M.A.; Bottinelli, R. Human skeletal muscle fibre contractile properties and proteomic profile: adaptations to 3 weeks of unilateral lower limb suspension and active recovery. The Journal of Physiology 2015, 593, 5361-5385. [CrossRef]
- Desplanches, D.; Kayar, S.; Sempore, B.; Flandrois, R.; Hoppeler, H. Rat soleus muscle ultrastructure after hindlimb suspension. Journal of Applied Physiology 1990, 69, 504-508. [CrossRef]
- Mortreux, M.; Rosa-Caldwell, M.E.; Stiehl, I.D.; Sung, D.M.; Thomas, N.T.; Fry, C.S.; Rutkove, S.B. Hindlimb suspension in Wistar rats: Sex-based differences in muscle response. Physiological Reports 2021, 9, e15042. [CrossRef]
- Haida, N.; Fowler Jr, W.M.; Abresch, R.T.; Larson, D.B.; Sharman, R.B.; Taylor, R.G.; Entrikin, R.K. Effect of hind-limb suspension on young and adult skeletal muscle: I. normal mice. Experimental neurology 1989, 103, 68-76.
- Murakami, T.; Hijikata, T.; Yorifuji, H. Staging of disuse atrophy of skeletal muscles on immunofluorescence microscopy. Anatomical science international 2008, 83, 68-76. [CrossRef]
- Nicks, D.K.; Beneke, W.M.; Key, R.M.; Timson, B.F. Muscle fibre size and number following immobilisation atrophy. Journal of anatomy 1989, 163, 1.
- Gomes, A.; Coutinho, E.; França, C.; Polonio, J.; Salvini, T. Effect of one stretch a week applied to the immobilized soleus muscle on rat muscle fiber morphology. Brazilian Journal of Medical and Biological Research 2004, 37, 1473-1480. [CrossRef]
- Gomes, A.R.; Cornachione, A.; Salvini, T.F.; Mattiello-Sverzut, A.C. Morphological effects of two protocols of passive stretch over the immobilized rat soleus muscle. Journal of Anatomy 2007, 210, 328-335. [CrossRef]
- Booth, F.; Kelso, J. Effect of hind-limb immobilization on contractile and histochemical properties of skeletal muscle. Pfluegers Archiv 1973, 342, 231-238. [CrossRef]
- Cardenas, D.; Stolov, W.; Hardy, R. Muscle fiber number in immobilization atrophy. Archives of physical medicine and rehabilitation 1977, 58, 423-426.
- Boyes, G.; Johnston, I. Muscle fibre composition of rat vastus intermedius following immobilisation at different muscle lengths. Pfluegers Archiv 1979, 381, 195-200.
- Templeton, G.; Sweeney, H.; Timson, B.F.; Padalino, M.; Dudenhoeffer, G. Changes in fiber composition of soleus muscle during rat hindlimb suspension. Journal of applied physiology 1988, 65, 1191-1195. [CrossRef]
- Eng, C.M.; Smallwood, L.H.; Rainiero, M.P.; Lahey, M.; Ward, S.R.; Lieber, R.L. Scaling of muscle architecture and fiber types in the rat hindlimb. Journal of Experimental Biology 2008, 211, 2336-2345. [CrossRef]
- Miller, T.A.; Lesniewski, L.A.; Muller-Delp, J.M.; Majors, A.K.; Scalise, D.; Delp, M.D. Hindlimb unloading induces a collagen isoform shift in the soleus muscle of the rat. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 2001, 281, R1710-R1717. [CrossRef]
- Al-Sarraf, H.; Mouihate, A. Muscle Hypertrophy in a Newly Developed Resistance Exercise Model for Rats. Frontiers in Physiology 2022, 962. [CrossRef]
- Winters, T.M.; Takahashi, M.; Lieber, R.L.; Ward, S.R. Whole muscle length-tension relationships are accurately modeled as scaled sarcomeres in rabbit hindlimb muscles. Journal of biomechanics 2011, 44, 109-115. [CrossRef]
- Williams, P.E.; Goldspink, G. The effect of immobilization on the longitudinal growth of striated muscle fibres. Journal of Anatomy 1973, 116, 45.
- Williams, P.E.; Goldspink, G. Changes in sarcomere length and physiological properties in immobilized muscle. Journal of anatomy 1978, 127, 459.
- Hikida, R.S.; Gollnick, P.D.; Dudley, G.A.; Convertino, V.A.; Buchanan, P. Structural and metabolic characteristics of human skeletal muscle following 30 days of simulated microgravity. Aviation, Space, and Environmental Medicine 1989, 60, 664-670.
- Sakakima, H.; Yoshida, Y.; Sakae, K.; Morimoto, N. Different frequency treadmill running in immobilization-induced muscle atrophy and ankle joint contracture of rats. Scandinavian journal of medicine & science in sports 2004, 14, 186-192. [CrossRef]
- Riley, D.; Ilyina-Kakueva, E.; Ellis, S.; Bain, J.; Slocum, G.; Sedlak, F. Skeletal muscle fiber, nerve, and blood vessel breakdown in space-flown rats. The FASEB Journal 1990, 4, 84-91.
- Nonaka, I.; Miyazawa, M.; Sukegawa, T.; Yonemoto, K.; Kato, T. Muscle fiber atrophy and degeneration induced by experimental immobility and hindlimb suspension. International journal of sports medicine 1997, 18, S292-S294. [CrossRef]
- Baewer, D.V.; Hoffman, M.; Romatowski, J.G.; Bain, J.; Fitts, R.H.; Riley, D.A. Passive stretch inhibits central corelike lesion formation in the soleus muscles of hindlimb-suspended unloaded rats. Journal of Applied Physiology 2004, 97, 930-934. [CrossRef]
- Cornachione, A.; Cação-Benedini, L.; Shimano, M.; Volpon, J.B.; Martinez, E.Z.; Mattiello-Sverzut, A.C. Morphological comparison of different protocols of skeletal muscle remobilization in rats after hindlimb suspension. Scandinavian journal of medicine & science in sports 2008, 18, 453-461.
- Baker, J.H.; Matsumoto, D.E. Adaptation of skeletal muscle to immobilization in a shortened position. Muscle & Nerve: Official Journal of the American Association of Electrodiagnostic Medicine 1988, 11, 231-244.
- Murakami, T.; Hijikata, T.; Yorifuji, H. Staging of disuse atrophy of skeletal muscles on immunofluorescence microscopy. Anat Sci Int 2008, 83, 68-76. [CrossRef]
- Baker, J.H. Segmental necrosis in tenotomized muscle fibers. Muscle & Nerve: Official Journal of the American Association of Electrodiagnostic Medicine 1983, 6, 29-39.
- Appell, H.-J. Morphology of immobilized skeletal muscle and the effects of a pre-and postimmobilization training program. International journal of sports medicine 1986, 7, 6-12. [CrossRef]
- Giordano, F.; Vizziello, E.; Tidball, J.; Falcieri, E.; Curzi, D. Plantaris muscle adaptation to atrophy generated by disuse: an ultrastructural study. Microscopie 2014, 22, 31-36.
- Widrick, J.J.; Romatowski, J.; Bain, J.L.; Trappe, S.W.; Trappe, T.A.; Thompson, J.L.; Costill, D.L.; Riley, D.A.; Fitts, R.H. Effect of 17 days of bed rest on peak isometric force and unloaded shortening velocity of human soleus fibers. American Journal of Physiology-Cell Physiology 1997, 273, C1690-C1699. [CrossRef]
- Riley, D.A.; Bain, J.L.; Thompson, J.L.; Fitts, R.H.; Widrick, J.J.; Trappe, S.W.; Trappe, T.A.; Costill, D.L. Decreased thin filament density and length in human atrophic soleus muscle fibers after spaceflight. Journal of applied physiology 2000, 88, 567-572. [CrossRef]
- Riley, D.A.; Ellis, S.; Slocom, G.R.; Satyanarayana, T.; Bain, J.L.; Sedlak, F.R. Hypogravity-induced atrophy of rat soleus and extensor digitorum longus muscles. Muscle & Nerve: Official Journal of the American Association of Electrodiagnostic Medicine 1987, 10, 560-568.
- Riley, D.A.; Bain, J.L.; Thompson, J.L.; Fitts, R.H.; Widrick, J.J.; Trappe, S.W.; Trappe, T.A.; Costill, D.L. Disproportionate loss of thin filaments in human soleus muscle after 17-day bed rest. Muscle & Nerve: Official Journal of the American Association of Electrodiagnostic Medicine 1998, 21, 1280-1289.
- Jakubiec-Puka, A.; Szczepanowska, J.; Wieczorek, U. Myosin heavy chains in striated muscle immobilized in a shortened position. Basic Appl Myol 1995, 5, 147-153.
- Goldspink, D. The influence of immobilization and stretch on protein turnover of rat skeletal muscle. The Journal of physiology 1977, 264, 267-282. [CrossRef]
- Booth, F.; Seider, M. Early change in skeletal muscle protein synthesis after limb immobilization of rats. Journal of Applied Physiology 1979, 47, 974-977. [CrossRef]
- Jaspers, S.R.; Tischler, M.E. Atrophy and growth failure of rat hindlimb muscles in tail-cast suspension. Journal of Applied Physiology 1984, 57, 1472-1479. [CrossRef]
- Loughna, P.; Goldspink, G.; Goldspink, D. Effect of inactivity and passive stretch on protein turnover in phasic and postural rat muscles. Journal of applied physiology 1986, 61, 173-179. [CrossRef]
- Gibson, J.; Halliday, D.; Morrison, W.; Stoward, P.; Hornsby, G.; Watt, P.; Murdoch, G.; Rennie, M. Decrease in human quadriceps muscle protein turnover consequent upon leg immobilization. Clinical science 1987, 72, 503-509. [CrossRef]
- Thomason, D.B.; Biggs, R.B.; Booth, F.W. Protein metabolism and beta-myosin heavy-chain mRNA in unweighted soleus muscle. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 1989, 257, R300-R305. [CrossRef]
- Ferrando, A.A.; Lane, H.W.; Stuart, C.A.; Davis-Street, J.; Wolfe, R.R. Prolonged bed rest decreases skeletal muscle and whole body protein synthesis. American Journal of Physiology-Endocrinology and Metabolism 1996, 270, E627-E633.
- Gamrin, L.; Berg, H.; Essen, P.; Tesch, P.; Hultman, E.; Garlick, P.; McNurlan, M.; Wernerman. The effect of unloading on protein synthesis in human skeletal muscle. Acta physiologica scandinavica 1998, 163, 369-377.
- Fluckey, J.; Dupont-Versteegden, E.; Montague, D.; Knox, M.; Tesch, P.; Peterson, C.; Gaddy-Kurten, D. A rat resistance exercise regimen attenuates losses of musculoskeletal mass during hindlimb suspension. Acta Physiologica Scandinavica 2002, 176, 293-300.
- Paddon-Jones, D.; Sheffield-Moore, M.; Cree, M.G.; Hewlings, S.J.; Aarsland, A.; Wolfe, R.R.; Ferrando, A.A. Atrophy and impaired muscle protein synthesis during prolonged inactivity and stress. The Journal of Clinical Endocrinology & Metabolism 2006, 91, 4836-4841.
- De Boer, M.D.; Selby, A.; Atherton, P.; Smith, K.; Seynnes, O.R.; Maganaris, C.N.; Maffulli, N.; Movin, T.; Narici, M.V.; Rennie, M.J. The temporal responses of protein synthesis, gene expression and cell signalling in human quadriceps muscle and patellar tendon to disuse. The Journal of physiology 2007, 585, 241-251.
- Kortebein, P.; Ferrando, A.; Lombeida, J.; Wolfe, R.; Evans, W.J. Effect of 10 days of bed rest on skeletal muscle in healthy older adults. Jama 2007, 297, 1769-1774.
- Glover, E.I.; Phillips, S.M.; Oates, B.R.; Tang, J.E.; Tarnopolsky, M.A.; Selby, A.; Smith, K.; Rennie, M.J. Immobilization induces anabolic resistance in human myofibrillar protein synthesis with low and high dose amino acid infusion. The Journal of physiology 2008, 586, 6049-6061.
- Symons, T.B.; Sheffield-Moore, M.; Chinkes, D.L.; Ferrando, A.A.; Paddon-Jones, D. Artificial gravity maintains skeletal muscle protein synthesis during 21 days of simulated microgravity. Journal of applied physiology 2009, 107, 34-38.
- Lang, S.M.; Kazi, A.A.; Hong-Brown, L.; Lang, C.H. Delayed recovery of skeletal muscle mass following hindlimb immobilization in mTOR heterozygous mice. PloS one 2012, 7, e38910.
- Phillips, S.M.; McGlory, C. CrossTalk proposal: The dominant mechanism causing disuse muscle atrophy is decreased protein synthesis. The Journal of physiology 2014, 592, 5341.
- Wall, B.T.; Dirks, M.L.; Snijders, T.; van Dijk, J.-W.; Fritsch, M.; Verdijk, L.B.; van Loon, L.J. Short-term muscle disuse lowers myofibrillar protein synthesis rates and induces anabolic resistance to protein ingestion. American journal of physiology-endocrinology and metabolism 2016.
- Mirzoev, T.; Tyganov, S.; Vilchinskaya, N.; Lomonosova, Y.; Shenkman, B. Key markers of mTORC1-dependent and mTORC1-independent signaling pathways regulating protein synthesis in rat soleus muscle during early stages of hindlimb unloading. Cellular Physiology and Biochemistry 2016, 39, 1011-1020.
- Tyganov, S.A.; Mochalova, E.P.; Belova, S.P.; Sharlo, K.A.; Rozhkov, S.V.; Vilchinskaya, N.A.; Paramonova, I.I.; Mirzoev, T.M.; Shenkman, B.S. Effects of plantar mechanical stimulation on anabolic and catabolic signaling in rat postural muscle under short-term simulated gravitational unloading. Frontiers in Physiology 2019, 10, 1252.
- Rozhkov, S.V.; Sharlo, K.A.; Mirzoev, T.M.; Shenkman, B.S. Temporal changes in the markers of ribosome biogenesis in rat soleus muscle under simulated microgravity. Acta Astronautica 2021, 186, 252-258.
- Lin, K.H.; Wilson, G.M.; Blanco, R.; Steinert, N.D.; Zhu, W.G.; Coon, J.J.; Hornberger, T.A. A deep analysis of the proteomic and phosphoproteomic alterations that occur in skeletal muscle after the onset of immobilization. J Physiol 2021, 599, 2887-2906. [CrossRef]
- Tesch, P.A.; von Walden, F.; Gustafsson, T.; Linnehan, R.M.; Trappe, T.A. Skeletal muscle proteolysis in response to short-term unloading in humans. Journal of applied physiology 2008, 105, 902-906.
- Steffen, J.M.; Musacchia, X. Spaceflight effects on adult rat muscle protein, nucleic acids, and amino acids. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 1986, 251, R1059-R1063.
- Bodine, S.C.; Latres, E.; Baumhueter, S.; Lai, V.K.-M.; Nunez, L.; Clarke, B.A.; Poueymirou, W.T.; Panaro, F.J.; Na, E.; Dharmarajan, K. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science 2001, 294, 1704-1708.
- Nikawa, T.; Ishidoh, K.; Hirasaka, K.; Ishihara, I.; Ikemoto, M.; Kano, M.; Kominami, E.; Nonaka, I.; Ogawa, T.; Adams, G.R. Skeletal muscle gene expression in space-flown rats. The FASEB journal 2004, 18, 522-524.
- Hussain, S.N.; Mofarrahi, M.; Sigala, I.; Kim, H.C.; Vassilakopoulos, T.; Maltais, F.; Bellenis, I.; Chaturvedi, R.; Gottfried, S.B.; Metrakos, P. Mechanical ventilation–induced diaphragm disuse in humans triggers autophagy. American journal of respiratory and critical care medicine 2010, 182, 1377-1386.
- Reich, K.A.; Chen, Y.-W.; Thompson, P.D.; Hoffman, E.P.; Clarkson, P.M. Forty-eight hours of unloading and 24 h of reloading lead to changes in global gene expression patterns related to ubiquitination and oxidative stress in humans. Journal of Applied Physiology 2010, 109, 1404-1415.
- Andrianjafiniony, T.; Dupré-Aucouturier, S.; Letexier, D.; Couchoux, H.; Desplanches, D. Oxidative stress, apoptosis, and proteolysis in skeletal muscle repair after unloading. American Journal of Physiology-Cell Physiology 2010, 299, C307-C315.
- Nandi, D.; Tahiliani, P.; Kumar, A.; Chandu, D. The ubiquitin-proteasome system. J Biosci 2006, 31, 137-155. [CrossRef]
- Stuart, C.A.; Shangraw, R.E.; Peters, E.J.; Wolfe, R.R. Effect of dietary protein on bed-rest-related changes in whole-body-protein synthesis. The American journal of clinical nutrition 1990, 52, 509-514.
- Stein, T.; Schluter, M. Human skeletal muscle protein breakdown during spaceflight. American Journal of Physiology-Endocrinology and Metabolism 1997, 272, E688-E695.
- Reid, M.B.; Judge, A.R.; Bodine, S.C. CrossTalk opposing view: The dominant mechanism causing disuse muscle atrophy is proteolysis. J Physiol 2014, 592, 5345-5347. [CrossRef]
- Phillips, S.M.; McGlory, C. CrossTalk proposal: The dominant mechanism causing disuse muscle atrophy is decreased protein synthesis. J Physiol 2014, 592, 5341-5343. [CrossRef]
- Goodman, C.A.; Hornberger, T.A.; al., e. Comments on the CrossTalk proposal and opposing view: The dominant mechanism causing disuse muscle atrophy is decreased protein synthesis/proteolysis. Journal of Physiology 2015, 1-10.
- Galavazi, G. Identification of helical polyribosomes in sections of mature skeletal muscle fibers. Zeitschrift für Zellforschung und Mikroskopische Anatomie 1971, 121, 531-547.
- Galavazi, G.; Szirmai, J.A. The influence of age and testosterone on the ribosomal population in the m. levator ani and a thigh muscle of the rat. Zeitschrift für Zellforschung und Mikroskopische Anatomie 1971, 121, 548-560.
- Horne, Z.; Hesketh, J. Immunological localization of ribosomes in striated rat muscle. Evidence for myofibrillar association and ontological changes in the subsarcolemmal: myofibrillar distribution. Biochemical Journal 1990, 268, 231-236.
- Morkin, E. Postnatal muscle fiber assembly: localization of newly synthesized myofibrillar proteins. Science 1970, 167, 1499-1501.
- Kannus, P.; Jozsa, L.; Jarvinen, T.L.; Kvist, M.; Vieno, T.; Jarvinen, T.A.; Natri, A.; Jarvinen, M. Free mobilization and low-to high-intensity exercise in immobilization-induced muscle atrophy. Journal of Applied Physiology 1998, 84, 1418-1424.
- Järvinen, T.A.; Józsa, L.; Kannus, P.; Järvinen, T.L.; Hurme, T.; Kvist, M.; Pelto-Huikko, M.; Kalimo, H.; Järvinen, M. Mechanical loading regulates the expression of tenascin-C in the myotendinous junction and tendon but does not induce de novo synthesis in the skeletal muscle. Journal of cell science 2003, 116, 857-866.
- Etlinger, J.; Zak, R.; Fischman, D.; Rabinowitz, M. Isolation of newly synthesised myosin filaments from skeletal muscle homogenates and myofibrils. Nature 1975, 255, 259-261.
- Etlinger, J.; Zak, R.; Fischman, D. Compositional studies of myofibrils from rabbit striated muscle. The Journal of cell biology 1976, 68, 123-141.
- Van der Westhuyzen, D.; Matsumoto, K.; Etlinger, J. Easily releasable myofilaments from skeletal and cardiac muscles maintained in vitro. Role in myofibrillar assembly and turnover. Journal of Biological Chemistry 1981, 256, 11791-11797.
- Belcastro, A.N.; Scrubb, J.; Gilchrist, J.S. Regulation of ATP-stimulated releasable myofilaments from cardiac and skeletal muscle myofibrils. Molecular and cellular biochemistry 1991, 103, 113-120.
- Neti, G.; Novak, S.M.; Thompson, V.F.; Goll, D.E. Properties of easily releasable myofilaments: are they the first step in myofibrillar protein turnover? American Journal of Physiology-Cell Physiology 2009, 296, C1383-C1390.
- Williams, A.B.; Decourten-Myers, G.M.; Fischer, J.E.; Luo, G.; Sun, X.; Hasselgren, P.O. Sepsis stimulates release of myofilaments in skeletal muscle by a calcium-dependent mechanism. The FASEB Journal 1999, 13, 1435-1443.
- Dahlmann, B.; Rutschmann, M.; Reinauer, H. Effect of starvation or treatment with corticosterone on the amount of easily releasable myofilaments in rat skeletal muscles. Biochemical Journal 1986, 234, 659-664.
- Smuder, A.J.; Nelson, W.B.; Hudson, M.B.; Kavazis, A.N.; Powers, S.K. Inhibition of the ubiquitin–proteasome pathway does not protect against ventilator-induced accelerated proteolysis or atrophy in the diaphragm. Anesthesiology 2014, 121, 115-126.
Figure 1.
Overview of skeletal muscle structure. (A) Illustration of major structural components of skeletal muscle. (B) Immunohistochemical staining (IHC) of a mouse plantaris muscle cross-section for laminin (white) as well as Type IIA (cyan) and Type IIB muscle fibers (magenta) was used to highlight the endomysium that surrounds individual muscle fibers. (C) IHC of a tibialis anterior muscle cross-section was used to identify a major component of the endomysium (laminin, white), a marker of satellite cells (Pax7, red), and nuclei (green). (D-E) Electron microscopy of mouse plantaris muscle cross-section was used to highlight the sarcoplasmic reticulum (SR) and mitochondria (mito) that surround individual myofibrils. (E) Higher magnification of the boxed region in D reveals the presence of the myosin (thick) and actin (thin) myofilaments. The figure is adapted from Jorgenson, et al., 2020 [32].
Figure 1.
Overview of skeletal muscle structure. (A) Illustration of major structural components of skeletal muscle. (B) Immunohistochemical staining (IHC) of a mouse plantaris muscle cross-section for laminin (white) as well as Type IIA (cyan) and Type IIB muscle fibers (magenta) was used to highlight the endomysium that surrounds individual muscle fibers. (C) IHC of a tibialis anterior muscle cross-section was used to identify a major component of the endomysium (laminin, white), a marker of satellite cells (Pax7, red), and nuclei (green). (D-E) Electron microscopy of mouse plantaris muscle cross-section was used to highlight the sarcoplasmic reticulum (SR) and mitochondria (mito) that surround individual myofibrils. (E) Higher magnification of the boxed region in D reveals the presence of the myosin (thick) and actin (thin) myofilaments. The figure is adapted from Jorgenson, et al., 2020 [32].
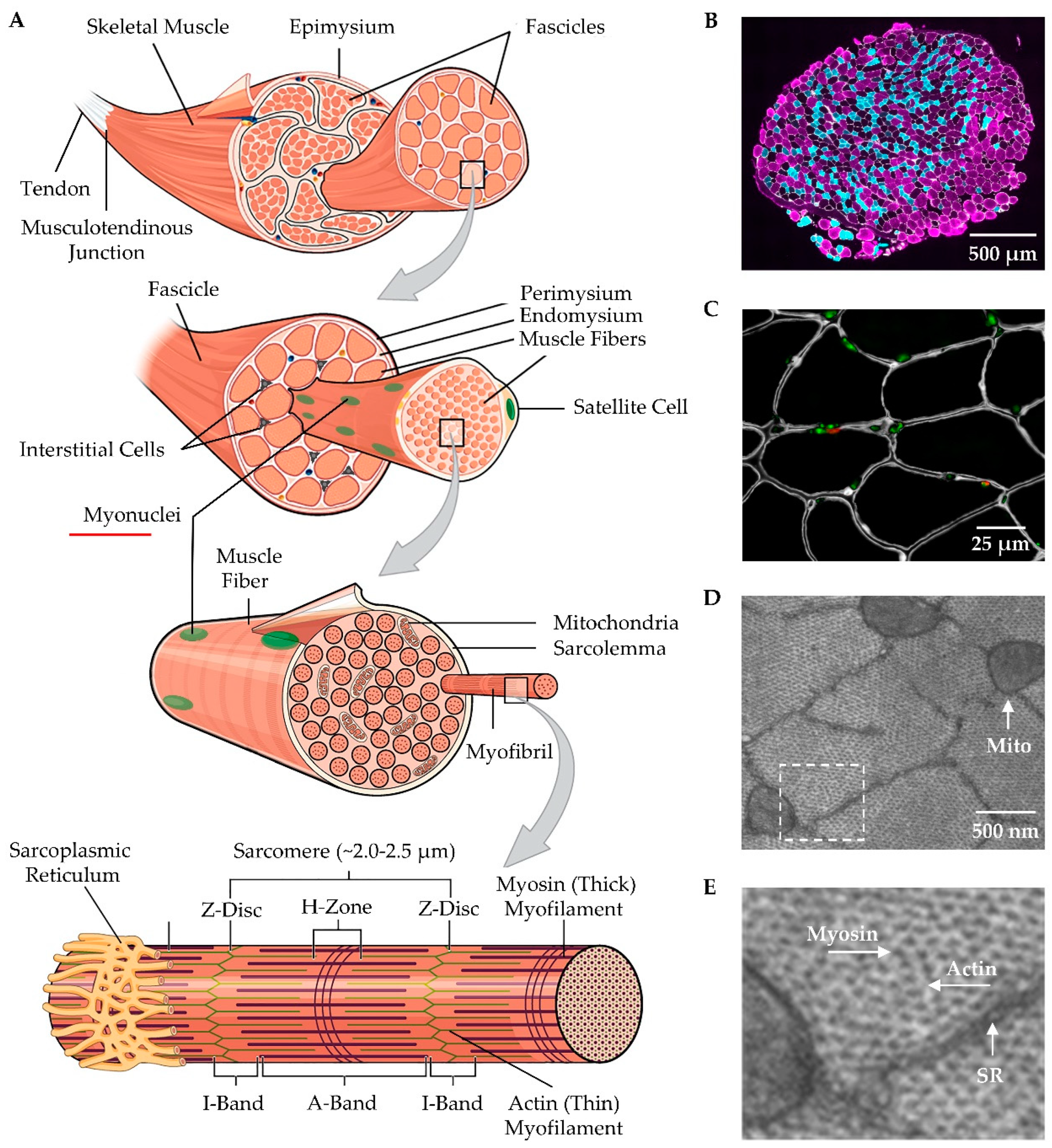
Figure 2.
Illustration of how longitudinal and radial atrophy of fascicles could influence the structural adaptations observed at the whole muscle level. (A) Description of a hypothetical muscle and its basic architectural properties. The geometric model of Maxwell et al., (1974) [108] was used to predict how only a 25% decrease in fascicle diameter (B), or only a 25% decrease in fascicle length (C), would impact measurements of whole muscle length and whole muscle diameter. (D) Prediction of how a 25% decrease in fascicle length plus a 14% decrease in whole muscle length would impact the whole muscle diameter and pennation angle of the fascicles. (E) Prediction of how a 25% decrease in fascicle diameter would impact the whole muscle diameter and pennation angle of the fascicles when whole muscle length is fixed.
Figure 2.
Illustration of how longitudinal and radial atrophy of fascicles could influence the structural adaptations observed at the whole muscle level. (A) Description of a hypothetical muscle and its basic architectural properties. The geometric model of Maxwell et al., (1974) [108] was used to predict how only a 25% decrease in fascicle diameter (B), or only a 25% decrease in fascicle length (C), would impact measurements of whole muscle length and whole muscle diameter. (D) Prediction of how a 25% decrease in fascicle length plus a 14% decrease in whole muscle length would impact the whole muscle diameter and pennation angle of the fascicles. (E) Prediction of how a 25% decrease in fascicle diameter would impact the whole muscle diameter and pennation angle of the fascicles when whole muscle length is fixed.
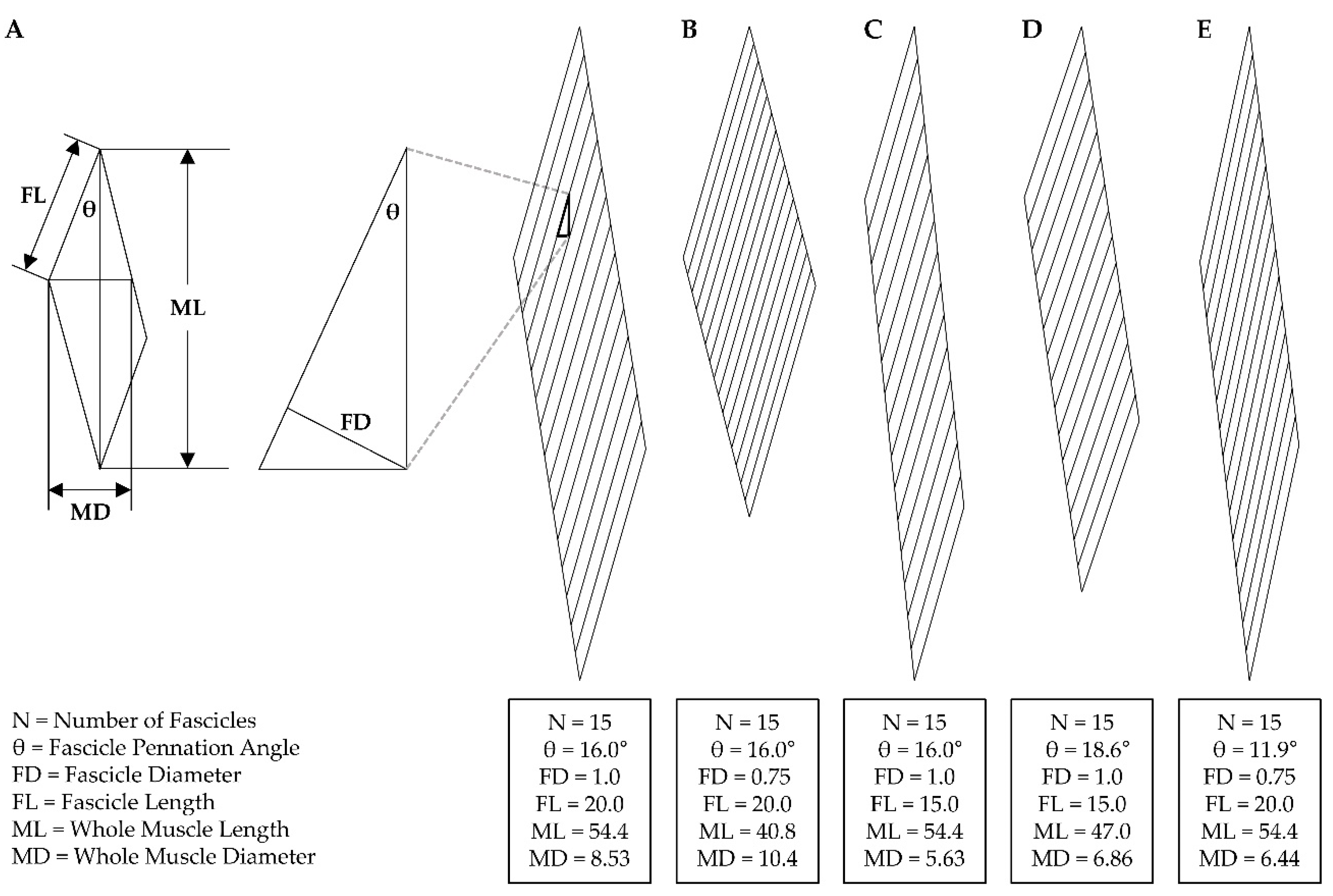
Figure 3.
Illustration of the mechanisms that could lead to radial atrophy of fascicles. (A) Radial atrophy of muscle fibers, (B) muscle fiber hypoplasia, and (C) longitudinal atrophy of muscle fibers that exhibit intrafascicular terminations such as those found in the sartorius and gracilis muscles of humans [120,124]. The figure is adapted from Jorgenson, et al., 2020 [32].
Figure 3.
Illustration of the mechanisms that could lead to radial atrophy of fascicles. (A) Radial atrophy of muscle fibers, (B) muscle fiber hypoplasia, and (C) longitudinal atrophy of muscle fibers that exhibit intrafascicular terminations such as those found in the sartorius and gracilis muscles of humans [120,124]. The figure is adapted from Jorgenson, et al., 2020 [32].
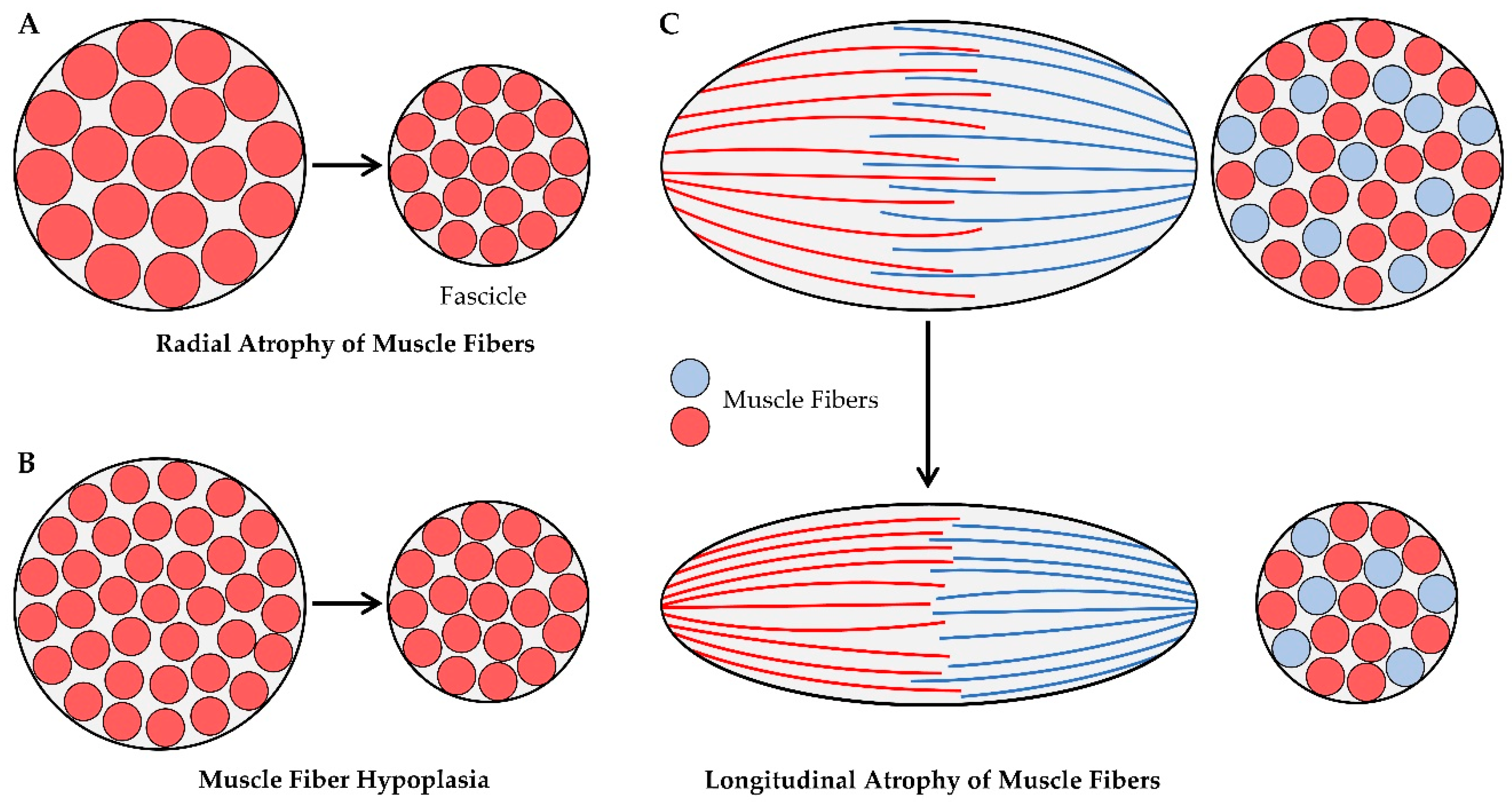
Figure 4.
Illustration of how changes in fiber length and fiber CSA can alter the number of muscle fibers that appear in a mid-belly cross-section. (A) Key formulas from the geometric model that was used to predict the architectural properties of skeletal muscle [108]. (B) Illustration of how a 16% decrease in the fiber length and/or a 61% decrease in the fiber CSA would exhibit different effects on the number of muscle fibers that appear in a mid-belly cross-section of the soleus and the plantaris muscles. Note: these are high-resolution images and details at the single fiber level should be visible when zooming in. (C) Quantitative results from the illustrations in B, where the data for the control muscles was derived from previously reported architectural properties [84,127,128,143,154-156], and the rationale for the 16% decrease in the fiber length and the 61% decrease in the fiber CSA being derived from the average decrease in these values that were reported in the following studies [81,82,84,127,128,143]. (D) Illustration of how a 16% decrease in fiber length and/or a 61% decrease in fiber CSA have no impact on the number of fibers that appear in mid-belly cross-sections (hatched boxes in B) of the soleus muscle, but the same changes in fiber length and/or fiber CSA dramatically alter the number of fibers that appear in mid-belly cross-sections of the plantaris.
Figure 4.
Illustration of how changes in fiber length and fiber CSA can alter the number of muscle fibers that appear in a mid-belly cross-section. (A) Key formulas from the geometric model that was used to predict the architectural properties of skeletal muscle [108]. (B) Illustration of how a 16% decrease in the fiber length and/or a 61% decrease in the fiber CSA would exhibit different effects on the number of muscle fibers that appear in a mid-belly cross-section of the soleus and the plantaris muscles. Note: these are high-resolution images and details at the single fiber level should be visible when zooming in. (C) Quantitative results from the illustrations in B, where the data for the control muscles was derived from previously reported architectural properties [84,127,128,143,154-156], and the rationale for the 16% decrease in the fiber length and the 61% decrease in the fiber CSA being derived from the average decrease in these values that were reported in the following studies [81,82,84,127,128,143]. (D) Illustration of how a 16% decrease in fiber length and/or a 61% decrease in fiber CSA have no impact on the number of fibers that appear in mid-belly cross-sections (hatched boxes in B) of the soleus muscle, but the same changes in fiber length and/or fiber CSA dramatically alter the number of fibers that appear in mid-belly cross-sections of the plantaris.
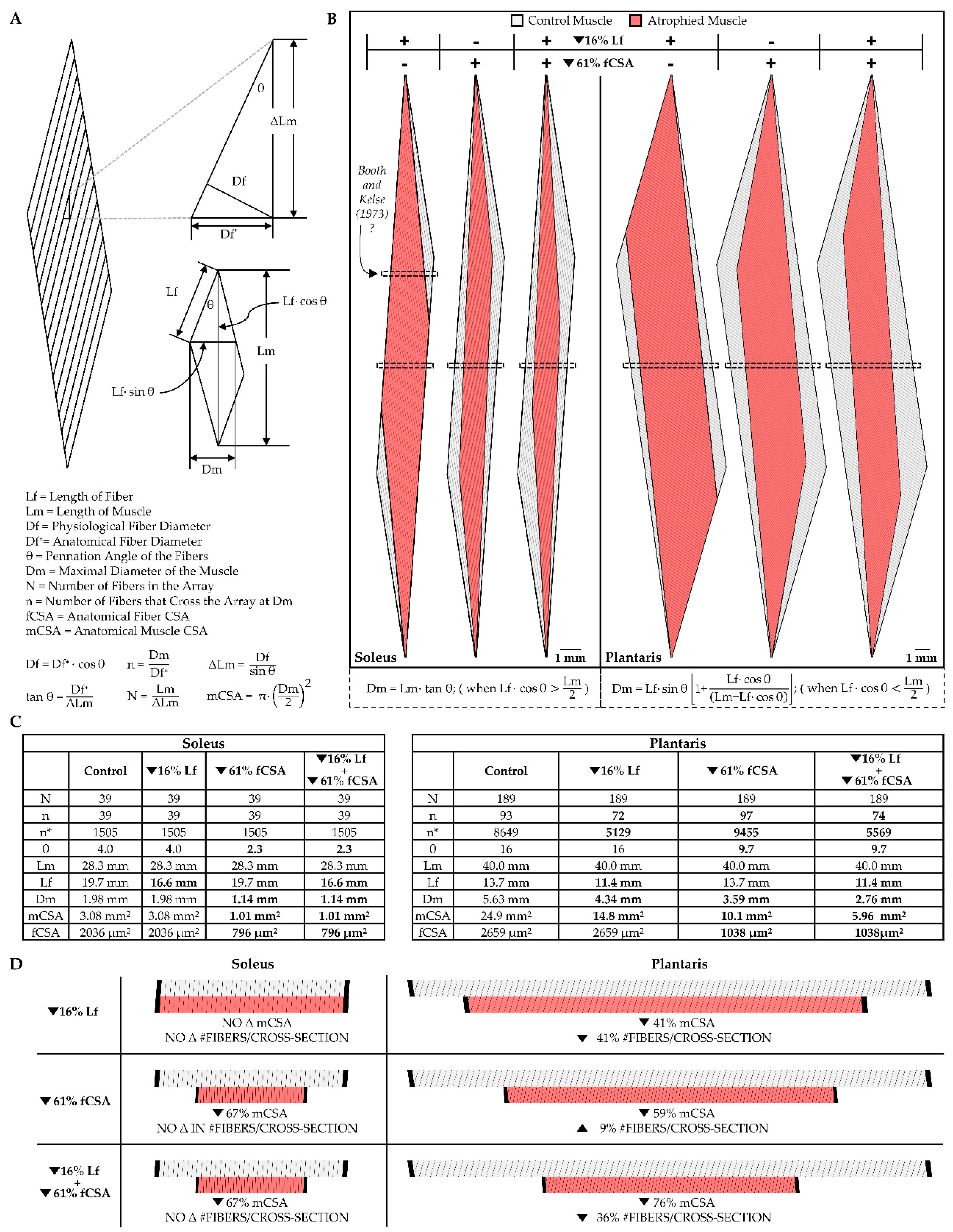
Figure 5.
Representations of CCLs and segmental necrosis. (A) Cross-section of soleus muscle fibers from rats that had been subjected to 12 days of hindlimb suspension. The black arrow points to a conventional CCL, and the white arrow points to a fiber with patches of CCL-like disruptions, copied with permission from [164]. (B and C) Cross- and longitudinal-sections of the soleus muscle of a rabbit that had been subjected to 7 days of hindlimb suspension, respectively. The white arrows point to patches of CCL-like disruptions that are found throughout the muscle fiber, copied with permission from [42]. (D) Longitudinal-section of a soleus muscle fiber from a rat 1 day after being subjected to disuse via tenotomy. The black arrow heads point to a large portion of muscle fibers that has undergone segmental necrosis, copied with permission from [168]. (E) Longitudinal-section of soleus muscle fibers from rats that had been subjected to 7 days of immobilization in a shortened position. The black arrow refers to an area of segmental necrosis near the muscle-tendon junction, copied with permission from [166]. Scale bar = 10 µm in all images.
Figure 5.
Representations of CCLs and segmental necrosis. (A) Cross-section of soleus muscle fibers from rats that had been subjected to 12 days of hindlimb suspension. The black arrow points to a conventional CCL, and the white arrow points to a fiber with patches of CCL-like disruptions, copied with permission from [164]. (B and C) Cross- and longitudinal-sections of the soleus muscle of a rabbit that had been subjected to 7 days of hindlimb suspension, respectively. The white arrows point to patches of CCL-like disruptions that are found throughout the muscle fiber, copied with permission from [42]. (D) Longitudinal-section of a soleus muscle fiber from a rat 1 day after being subjected to disuse via tenotomy. The black arrow heads point to a large portion of muscle fibers that has undergone segmental necrosis, copied with permission from [168]. (E) Longitudinal-section of soleus muscle fibers from rats that had been subjected to 7 days of immobilization in a shortened position. The black arrow refers to an area of segmental necrosis near the muscle-tendon junction, copied with permission from [166]. Scale bar = 10 µm in all images.
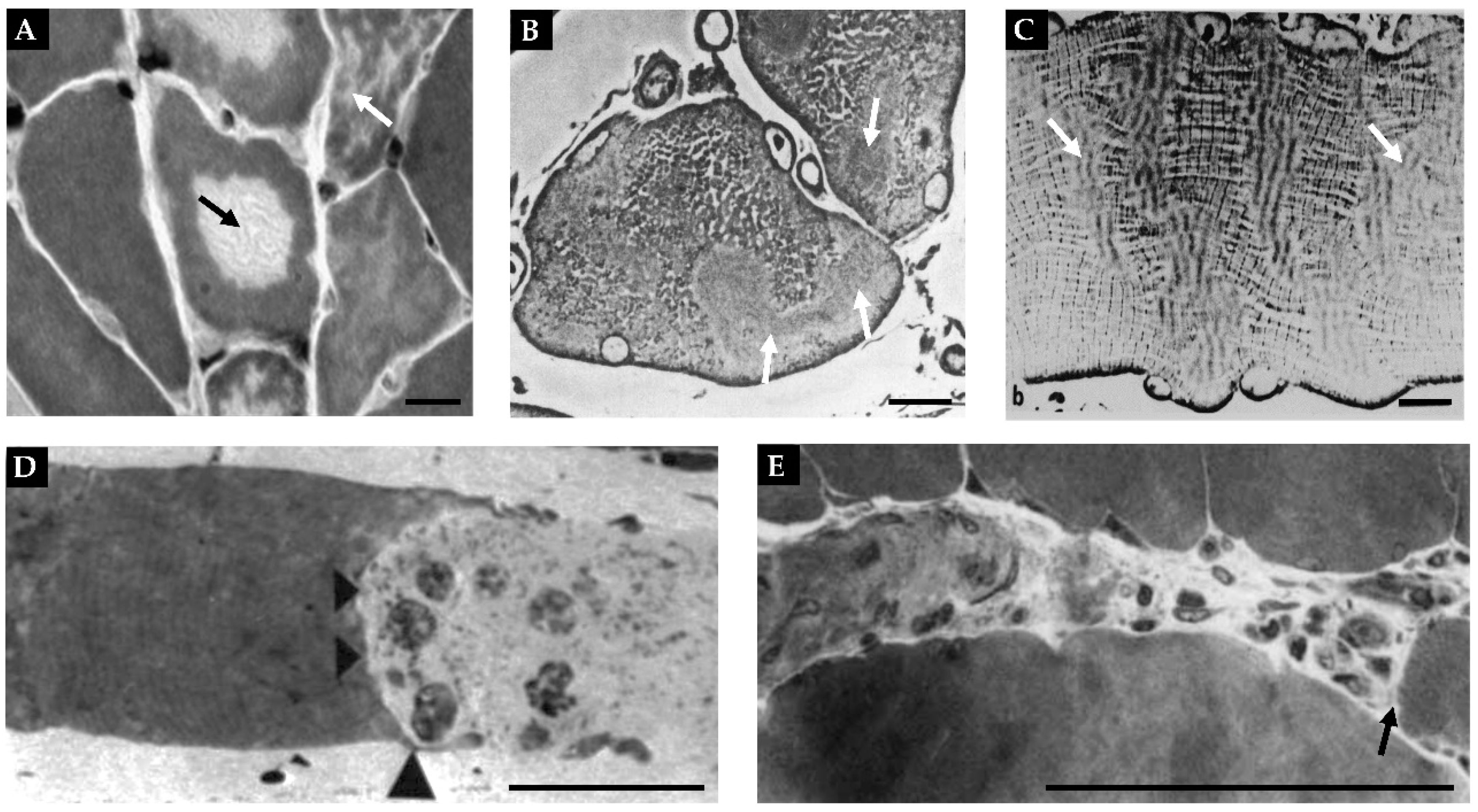
Figure 6.
Illustration of how radial atrophy of myofibrils and/or myofibril hypoplasia can contribute to the radial atrophy of the muscle fibers. The figure is adapted from Jorgenson, et al., 2020 [32].
Figure 6.
Illustration of how radial atrophy of myofibrils and/or myofibril hypoplasia can contribute to the radial atrophy of the muscle fibers. The figure is adapted from Jorgenson, et al., 2020 [32].
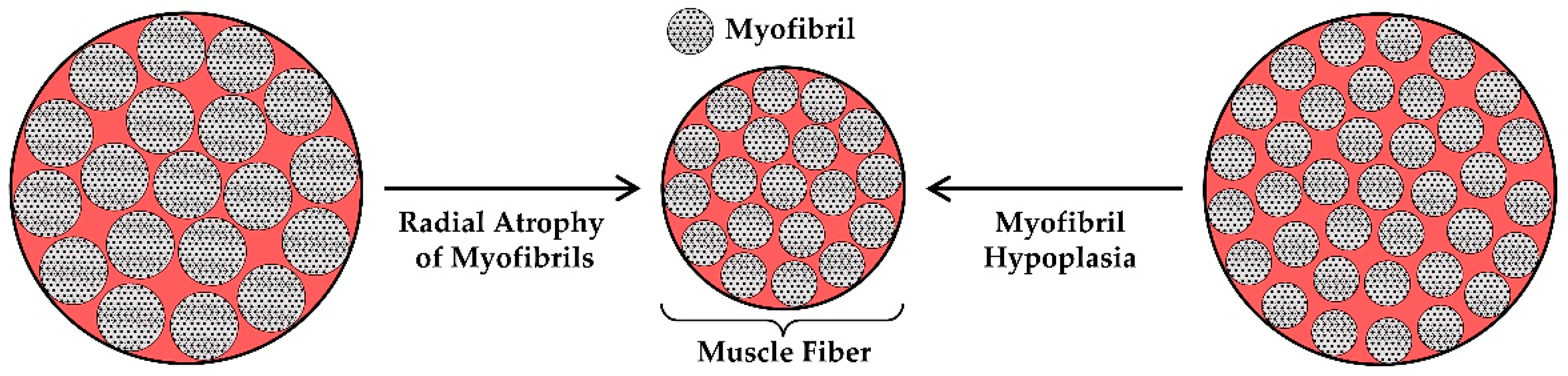
Figure 7.
Representations of the moth-eaten appearance of myofibrils that occurs during disuse. Electron micrographs from rat soleus muscles that have been subjected to 7 days of a control condition (A), or 7 days of spaceflight (B). Red arrows point to the presence of gaps in the myofilaments around the periphery of the myofibrils. Black arrows point to the focal loss of myofilaments within the myofibrils. Scale bar = 0.5 µm, copied with permission from [173].
Figure 7.
Representations of the moth-eaten appearance of myofibrils that occurs during disuse. Electron micrographs from rat soleus muscles that have been subjected to 7 days of a control condition (A), or 7 days of spaceflight (B). Red arrows point to the presence of gaps in the myofilaments around the periphery of the myofibrils. Black arrows point to the focal loss of myofilaments within the myofibrils. Scale bar = 0.5 µm, copied with permission from [173].
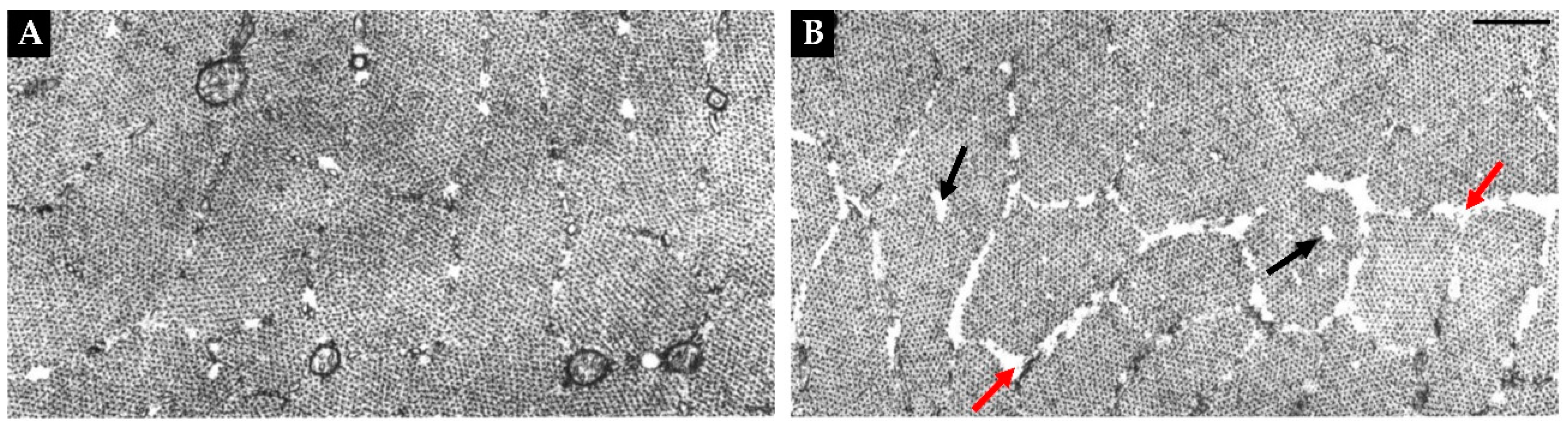

Table 1.
The Structural Adaptations that Mediate the Disuse-Induced Atrophy of Skeletal Muscle.
| Adaptation | Evidence | Major Gaps in Knowledge |
|---|---|---|
| Longitudinal Atrophy of Fascicles | High (for disuse in a shortened position) |
|
| Radial Atrophy of Fascicles | High |
|
| Longitudinal Atrophy of Muscle Fibers | High (for disuse in a shortened position) |
|
| Radial Atrophy of Muscle Fibers | Extremely High |
|
| Radial Atrophy of Myofibrils | Moderate |
|
| Myofibril Hypoplasia | Very Low |
|
* The underlined major gaps in knowledge are considered to be essential for the advancement of the field.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



