Submitted:
27 September 2023
Posted:
28 September 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Neutrophils are the principal trouper of innate immune system. Activated neutrophils undergo a noble cell death termed NETosis and release a mesh-like structure called neutrophil extracellular traps (NETs) as a part of their defensive strategy against microbial pathogen attack. This web-like architecture includes a DNA backbone embedded with antimicrobial proteins like myeloperoxidase (MPO), neutrophil elastase (NE), histones etc. and deploys in the entrapment and clearance of encountered pathogens. Thus NETs play an inevitable beneficial role in the host's protection. However, recent accumulated evidence shows that dysregulated and enhanced NET formation has various pathological aspects including promotion of sepsis, pulmonary, cardiovascular, hepatic, nephrological, thrombotic, autoimmune, pregnancy, cancer diseases etc. and the list is increasing gradually. In this review, we summarize NETs mediated pathophysiology of different diseases, focus on some updated potential therapeutic approaches against NETs and share our future perspectives.
Keywords:
neutrophil extracellular traps (NETs)
; innate immunity
; sepsis
; lung disease
; cardiovascular disease
; liver disease
; kidney disease
; diabetes
; COVID-19
; coagulopathy and thrombotic microangiopathy
; cancer
; autoimmunity
; preeclampsia
; Kawasaki disease
1. Introduction to Neutrophil Extracellular Traps (NETs)
1.1. Neutrophil and Innate Immunity
Innate immunity is the first line of defense of the body. The main function of the innate immunity is to protect our body from invading microorganisms and limit the growth and proliferation and finally kill the microorganisms. Polymorphonuclear neutrophils are the most abundant granular leukocytes produced at a rate of 5 × 1010-10 × 1010 cells per day, contributing approximately 50%–70% of all circulating white blood cells in humans having an average short half-life of 6-8 hours in circulation and key player of the innate immunity [1,2]. They are the first responder of the inflammatory cascades [3,4]. Neutrophils are derived from the bone marrow and enter into the circulation from where they quickly move into the sites of infection or inflammation in response to pathogen attack in the body and roll, adhere to endothelial layer, crawl and thus transmigrated from the vessel and kill the microbes [5,6,7]. After accomplishment of their role they undergo apoptosis and are cleared by macrophages [8]. As the arsenal of innate immunity neutrophil appear first to the infected site in the body and imply several strategies to eliminate the infection. In fact, neutrophils evolved to fulfill the key role in innate immunity through rapid deployment and effective antimicrobial action against a broad range of pathogens and noninfectious inflammation. Hence they are equipped with multiple weapons that can be deployed as a part of their antimicrobial strategies [9].
1.2. Antimicrobial actions of Neutrophils:
Neutrophils were recognized as the vanguard of the innate immunity and a vital protector against microbial infection and foreign invasion. In past decades it was thought that when neutrophil encounter invading pathogen in the body, they kill them either by phagocytosis, where they engulf and inactivate the pathogen and rapidly kill them by releasing proteolytic enzymes, antimicrobial proteins or reactive oxygen species (ROS) or by exocytosis, where they de-granulate and release antimicrobial factors in the extracellular space. However, in 2004, Brinkmann et al. described another way of neutrophil derived novel antimicrobial defense mechanism of innate immunity termed NETosis that can participate in pathogen killing extracellularly through the release of NETs [10]. Although neutrophils are classically known to die primarily by apoptosis or necrosis and highly regulated necroptois but NETosis is a cell death program that is distinct from apoptosis and necrosis [11,12].
1.3. Neutrophil Extracellular Traps (NETs)
NETs are released by activated neutrophils and the structure is composed meshwork of decondensed chromatin fibers decorated with antimicrobial granular proteins such as histones, myeloperoxidase (MPO), neutrophil elastase (NE), and cathepsin G etc. This web-like architectural design and composition allow NETs to prevent the dissemination of pathogen in the body. NETs can only be released robustly by matured neutrophils upon stimulation. Immature neutrophils are less potent compared to mature neutrophils at promoting innate immune defenses [13,14,15] and neutrophils from term and preterm infants fail to form NETs upon activation by inflammatory agonist [16]. Several infectious and sterile stimuli have been reported to trigger NET formation including bacteria [10,12], virus [17], fungi [18,19], parasites [20], pro-inflammatory cytokines like interleukin 8 (IL-8) [21], tumor necrosis factor α (TNF-α) [22], placental micro-debris [23], activated platelets [24], cholesterol crystals, monosodium uric acid [25], immune complexes [26], autoantibodies [27], complements such as C5a [28] and even cancer cells [29,30,31]. Phorbol esters such as phorbol myristate acetate (PMA), bacterial products such as lipopolysaccharide (LPS) are most widely used non physiological agonist to generate NETs ex vivo [10,32,33].
1.4. NETosis Mechanism
Two different mechanisms, NADPH Oxidase 2 (Nox 2)-dependent and Nox 2 independent NETosis have been well validated by published literatures. Agonist like PMA, LPS, bacteria such as Pseudomonas aeruginosa etc. induce Nox-dependent NETosis while agonist like calcium ionophores (A231128, ionomycin), uric acid crystals, certain microbes, UV ight etc. triggers Nox-independent NETosis through the formation of different ROS; Nox-ROS and mitochondrial ROS respectively [34,35,36]. The generated ROS activates Nox-dependent and Nox-independent specific different sets of kinases (MAPK, ERK, p38 and JNK) leading to transcriptional firing and stimulation of MPO. In Nox-independent NETosis histone citrullination is facilitated by a nuclear enzyme protein-arginine deiminase 4 (PAD4) activation [37,38]. MPO trigger the stimulation and translocation of NE from azurophilic granules to the nucleus and decorate the chromatin [39,40,41]. Ultimately nuclear membrane disintegrates and NETs are released.
1.5. NETosis Pathways
NET formation occurs via two pathways; suicidal NETosis and vital NETosis. Suicidal NETosis represents a cell death pathway that starts with nuclear delobulation, the nuclear membrane disassembly followed by continuous loss of cellular polarization, chromatin decondensation and finally finished with plasma membrane rupture and expulsion of NETs [42]. This lytic slow cell death process usually takes 2-4 hours [33]. On the other hand vital NETosis is cell death independent non-lytic process where expulsion of nuclear chromatin is accompanied by the release of granular proteins and fabrication of these components occur extracellularly leaving active anucleated phagocytic cytoplast having the potency to ingest microorganisms and chemotaxis [42]. Vital NETosis happens faster within 5-60 minutes depending on the inducer [43]. In vital NETosis the released extracellular DNA by neutrophils could be of either nuclear or mitochondrial [44,45,46].
1.6. Physiological role of NETs
Decades ago, when the NETs emerged, scientists became very interested observing its antibacterial capacity. Wide range of killing capacities of NETs made the immunologist to work with great haste. NETs conquer over infections by trapping, immobilizing and neutralizing gram-negative and gram-positive bacteria [10], viruses [17] fungi [47], parasites [48,49] and thus it prevents the dissemination of intruding microbes and protects the host. Although NETs trap microbes through charge interaction [50] but some pathogens evade NETs. Many gram positive pathogens including but not limited to pathogens of the Streptococcus and Staphylococcus genera release endonucleases, secreted endonuclease degrades the extracellular DNA scaffold, destroy and evade NETs [51,52]. The escaped bacteria with endonuclease may promote further invasion and spread from the local sites to distant organ and blood stream [53,54,55]. Again Streptococcus pneumoniae can escape from NETs in a charge dependent manner. This gram-positive bacterial strain expresses anti-phagocytic polysaccharide capsules and lipoteichoic acid which can produce a positive charge on their surface and thus prevent them to be trapped by positively charged NET fibers and histone residues [56]. Interestingly, the beneficial effect of NETosis over phagocytosis is the size of NETs. Large microbes and parasites evade phagocytosis and can prove difficult to clear. But neutrophil has a unique microbe sensing mechanism that allows them to selectively tailor their antimicrobial responses to pathogen on basis of microbial size. The large filamentous form of fungi cannot be removed by phagocytosis. NETs play a significant role to control these large filamentous fungi [57,58,59].
In addition, NETs also contribute to immunothrombosis. Immunothrombosis is a physiological process having bidirectional role with innate immune system. Although excessive activation of coagulation cascade leads to many clinical conditions including sepsis, disseminated intravascular coagulation (DIC), myocardial infarction and coronavirus disease 2019 (COVID-19) etc. [60] in contrast it also plays an essential protective role in maintaining physiological hemostasis to avoid blood loss and arresting both viral and bacterial infections. However, with discovery of NETs it has been come into light that thrombus is not only for hemostatic purpose stopping bleeding but also takes part in innate immunity. NETs provide large procoagulant surface by activating the contact phase of coagulation [61,62]. Moreover, neutrophils and neutrophils derived micro-particles during NETosis such as DNA, histones and granule proteins provide coagulant activities [61].
1.7. Controversial role of NETs
Infect the intervention of NETs is a landmark progression of science. Now the question is whether NETs are too good or too bad for us. NETs are like double edged sword. While NETs have a physiological role in antimicrobial defenses, if dysregulated they also have various pathological aspects that attracted recent attention (Figure 1). In some conditions when it is excessively generated and present, NETs can do harm to the host. The antimicrobial histones and peptides decorating the NET-DNA are directly cytotoxic to tissue and ineffective clearance of NETs results deleterious inflammation of host tissue. The list of disorders implicated by NETs include sepsis, pulmonary, cardiovascular, hepatic, nephrological, thrombotic, autoimmune, pregnancy, cancer diseases etc. and are gradually increasing.
1.8. Detection of NETs
Uncontrolled release of NETs has been reported in several disease pathophysiologies (discussed later). Therefore, precise spotting of NETs in clinical samples could have great potential in the prognosis of disease progression and subsequent consequences. NETs are made up of cell-free DNA (cf-DNA) and anti-microbial proteins like MPO and NE etc. NET detection is very challenging due to its fragile structure, timing of formation and turnover and frequent presence of DNase. Measuring cf-DNA is not a good approach to quantify NETs as the source could also be of apoptosis and necrosis in addition to NETosis [63]. NETs can be visualized and quantified by fluorescence microscopy [64] and flow cytometry [65] tools in human neutrophils in vitro. However, enzyme-linked immunosorbent assay is the handy, highly sensitive and reliable method to quantitatively measure remnants of circulating NETs in plasma [66,67] as well as in cell culture supernatants in vitro [68].
2. NETs in Clinical Settings
2.1. NETs in Sepsis
Sepsis is a dysregulated response to an infection with deleterious effects in host leading to circulatory shock, multiple organ failure syndromes (MODS) and ultimately death. A common cause of death in sepsis is the overwhelming infection in the blood stream and the resulting complications [69]. During sepsis activation of neutrophil with microbial or inflammatory stimuli occur which results in the expulsion of NETs [70]. NETs are essential antimicrobial defense for pathogen clearance in the blood and tissues during infection but at the same time NETs and NET components exert excessive inflammation resulting host tissue damage [42]. Using animal model of sepsis, circulatory NET in the blood stream became evident [24,45,71] and the biomarkers used to check the presence of NETs, were also increased in septic patients [72,73,74]. NET damages tissue and increase vascular permeability in sepsis. Promotion of neutrophil infiltration occurs in tissues through neutrophil-endothelial cell (EC) interaction resulting increased NET formation [75] and this interaction results excess NET formation which is dependent on activated EC derived IL-8 [76]. This damage is neutralized when incubated with either NAPDH oxidase inhibitors or DNase [76].
Immunothrombosis represents controlled inflammation and coagulation and the major-line of innate immune defense towards intruding infection. In sepsis, dysregulated, sustained and hyper-immunothrombosis leads to DIC complications [77]. Sepsis-induced DIC is detrimental to host and cause organ dysfunctions having a double mortality rate that of septic patients without DIC [78]. Neutrophils diligently participate in thrombosis associated DIC [79]. Neutrophil released NETs contribute excessive thrombin generation due to its early presence at the onset of DIC [80].
Normally platelet remains dormant in the circulation [81]. Growing body of evidences reveal that in sepsis the interaction between neutrophils and activated platelets happen simultaneously and that platelet can rapidly mediate neutrophils to make NET in vivo [24,82]. Activated platelets trigger the neutrophils derived NET release in either P-selectin dependent [83,84] or Toll-like receptor (TLR) 4 dependent manner [24,85] which traps bacteria in expense of endothelial tissue damage. Histones along with fragmented DNA are the abundant components of NETs. In acute systemic inflammatory conditions, including sepsis and trauma, circulating histones aggravates micro-circulatory thrombosis, worsen tissue perfusion and contributes significantly to organ injury [85].
2.2. NETs in Lung Diseases
Excessive activation of neutrophils causes MODS and lung, the most sensitive and important organ in systemic inflammation, is the main target [86,87]. Acute lung injury (ALI) is one of the leading causes of death in the ICU. Lung edema, inflammation, hyaline membrane and alveolar damage are the characteristic morphological features of ALI [88]. Acute respiratory distress syndrome (ARDS) is an acute inflammatory lung injury characterized by hypoxemic respiratory failure as a consequence of increased permeability of the endothelial-epithelial barrier, alveolar damage, and pulmonary edema. ALI and its severest form ARDS or chronic obstructive pulmonary disease remains as an important clinical challenge due to complex and ambiguous pathophysiology [89,90]. A massive influx of activated neutrophils is seen to the lung microvasculature, interstitial, and alveolar space and dysregulated inflammatory neutrophils are the key factor to the progression of ALI/ARDS [91,92,93]. This excessive neutrophil activation and accumulation induces increased formation of NETs along with increased release of proinflammatory mediators [94,95]. NET mediated cytotoxicity on alveolar epithelial cells as well as pulmonary EC is mainly due to protein components of NETs [96]. Histones major components of NETs are too toxic to cells and lung is the most susceptible vital organ to high levels of circulatory histones [97]. NET derived histones were detected from bronchoalveolar lavage fluid (BALF) samples from humans with ARDS [98]. Co-culture of PMA-stimulated neutrophil with EC alter barrier function resulting EC damage which is attributed to NETs and pre-treating EC with DNase, Cl-amidine a PAD 4 inhibitor, and 1-(3-methylbenzoyl)-1H-indazole-3-carbonitrile an NE inhibitor restore the damage [99]. Growing body of evidence suggested that the lung injury in ALI or ARDS is triggered by C5a activated NET release along with histones and enzymes that causes tissue damage [98,100]. NETs also contribute to pathogen induced lung injury in mouse model and human [101].
Cystic fibrosis (CF) is an inherited autosomal recessive disease of the lung due to chronic inflammation of the airways and bacterial colonization leading to death in 90% of patients. CF occurs due to a mutation in CF trans-membrane conductance regulator gene that control the balance of bicarbonate and chloride secretions across the cell surface epithelium of the airways [102] and this mutation increases the susceptibility of these patients to airway bacterial infection mainly by Pseudomonas aeruginosa and Haemophilus influenza and Staphylococcus aureus [103]. Neutrophils are infiltrated to the airway upon bacterial colonization and chronic and show less potency to eliminate microbes instead contributes to lung damage [104]. This colonization induces NET expulsion resulting sputum viscosity and ultimately exacerbating the patient’s respiratory capacity [105]. Neutrophil count and extracellular DNA can be used as severity marker for the assessment of inflammation and lung disease severity in CF [106]. High levels of NET components such as MPO and NE enzymes available in CF sputum and BALF are responsible for the damage of airway epithelium and connective tissues that correlate with lung disease severity [96,107,108]. DNase treatments in CF patients diminished this damage and improve pulmonary function [109].
Asthma is a chronic heterogeneous airway inflammatory disorder with symptomatic features of periodic wheezing, coughing and shortness of breath leading to the deterioration of lung function [110]. Asthmatic inflammation was thought to be attributed mainly by eosinophil but recent researches suggested a greater proportion of neutrophilic involvement having worsened disease severity with poor treatment outcome with traditional glucocorticoids [111]. Extracellular traps have been marked in the airway of atopic allergic asthmatic patients [112]. In asthmatic patients BALF neutrophil count and IL-8 are reported as the most powerful biomarker to differentiate severe and moderate condition [113]. IL-8 is a neutrophil chemoattractant and established agonist of NETosis. Again in asthmatic patient’s plasma activated platelet also rises [114] which is also responsible for induction of NET formation [24].
2.3. NETs in Cardiovascular Diseases
Acute myocardial infarction (AMI) is a life-threatening condition that occurs due to blockade of blood flow to the heart muscle because of thrombus formation occluding one or more coronary artery resulting tissue damage. Neutrophil recruitment has been mentioned to be involved in the development of atherothrombosis [115], coronary thrombi [116] and also used as the prediction of acute coronary events [117]. High levels of NETs are associated with severe coronary atherosclerosis patients [118]. NETs are detected at the advanced atherosclerotic lesion both in human and mice model [119,120] and application of DNase in Ischemia reperfusion injury mice model reduces reperfusion injury [121]. Histone H4 structural components of NET scaffold results atheromatous plaque instability and anti-histone H4 antibody implication results plaque stabilization [122]. A report described that NET could be a potential diagnostic marker in atherosclerosis [123]. Tissue factor is a trans-membrane protein that stimulates coagulation process. NET-associated tissue factor induces thrombogenic potential through platelet activation and increased thrombin generation resulting myocardial infarction [124].
2.4. NETs in Liver Diseases
Liver is the main organ for the clearance of circulating DNA and histones from the body. In Ischemia-reperfusion neutrophils are identified to the site of liver injury and expulsion of damage associated molecular patterns (DAMPs) and extracellular histones from damaged hepatocytes were observed to worsen the hepatic injury through TLR-4 and TLR-9 [125]. These DAMPs activate neutrophils to release NETs which intensifies sterile inflammatory liver injury. Again extracellular histones activate the nucleotide-binding domain, leucine-rich repeat containing protein 3 inflammasome, which further contribute in liver injury [126]. LPS activated platelets and the subsequent NET formation showed disturbed microcirculation and liver damage [24]. In a mouse model of ischemia-reperfusion injury NET mediated amplified inflammation and liver damage found to be restored after DNase or PAD 4 inhibitor treatment [125].
2.5. NETs in Kidney Diseases
The pathophysiology of acute kidney injury involves the renal tubular cell death and auto amplification loop of cell necrosis called necroinflammation [127,128]. Infections, trauma, toxins and ischemia induce a huge neutrophil recruitment in the renal tubule resulting necrosis and apoptosis and release of DAMPs and alarmins. Extracellular free or NET bound or immune complex associated histones are potent mediators of renal epithelial cells necrosis [129,130] and induce further histone release that act as DAMPs [129,131]. These DAMPs and other inflammatory mediators further activate neutrophil to release NETs, which accelerate more surrounding tissue injury. Histones and NETs enhance tubular necrosis and capillary injury [132] and this glomerular injury can be rescued by using PAD-4 inhibitor and anti-histone antibodies [130,133].
2.6. NETs in COVID-19
COVID-19 is a highly contagious respiratory disease caused by SARS-CoV-2 virus and declared as a global pandemic by world health organization. Higher presence of NETs in COVID-19 patients has been marked [134,135]. Neutrophilia, immune dysfunction and hyper-inflammation are the clinical features of severe COVID-19 patients [136,137]. This hyper-inflammation in COVID-19 is attributed by NETs [138]. Excessive production of cytokines termed cytokine storm is the hallmark in the pathogenesis of COVID-19 with augmented plasma levels of CCL 2, IFNγ, IFNγ-inducible protein 10, G-CSF, CCL3, IL-1β, IL-2, IL-6, IL-7, IL-8, IL-10, IL-17, and TNF-α leading to subsequent severe consequences like ALI, ARDS, pulmonary thrombosis and MODS [139,140,141,142,143]. IFN-γ, TNF-α, IL-1β and IL-8 are strong agonist of NET induction [21,22,135]. NETs have been reported to contribute the damage of the alveoli and pulmonary endothelium and immunothrombosis in patients with severe progression of COVID-19 [143,144,145,146,147,148]. Inflammatory microvascular thrombi having NET components have been identified in the lung, kidneys and hearts of COVID-19 patients [149]. NET components such as genomic DNA and citrullinated histones have been marked to initiate coagulation in COVID-19 patients through thrombin generation resulting poor fibrinolysis and dropped anticoagulant factor by binding to factor XII. NET inhibition in COVID-19 patients is found to compensate the NET-induced inflammation and thrombotic tissue damage linked to COVID-19 ARDS and death [143]. Ultimately there is a negative correlation between increased levels of NETs and decreased survivability in COVID-19 patients [150,151].
2.7. NETs in Coagulopathy and Thrombotic microangiopathy
Thrombosis is a principle cause of morbidity and mortality. Vascular occlusion represents deleterious effects of NETs. DIC is a heterogeneous group of disorder featured by widespread activation of intravascular coagulation [70,152]. The interaction between coagulation and innate immunity results immunothrombosis, NET induced immunothrombosis plays an essential physical role in innate immunity by immobilizing and preventing the dissemination of invading pathogens. Nevertheless exaggerated NETs can initiate abnormal thrombogenicity leading DIC. NETs initiate coagulation cascade and high flow circulating NETs aggregate and stick platelets to its own scaffold and form RBC rich thrombus that serve as a template for thrombus formation and adhere to the vascular endothelium in microvasculature [153]. Electrostatic interaction of the negatively charged cells with positively charged histones in NETs probably contribute to construct the scaffold. This scaffold is responsible for deep vein thrombosis (DVT). In fact, this NET-platelet-thrombin axis promotes intravascular coagulation and microvascular dysfunction [154,155]. Moreover, circulating free DNA suppress fibrinolysis either by accelerating the inactivation of tissue plasminogen activator by PAI-I [156] or by thickening the fibers of fibrin clot in combination with histones [157]. Again, there is a recommendation for NETs to be an important factor of excessive and unbalanced thrombin generation because of its early presence at the onset of DIC [80]. Histones also play a key role in the NET mediated coagulopathy. H3 and H4 of NETs stimulate platelets and induce thrombotic reaction in mice model [158]. Again in an animal model NET and its integral part citrullinated histones H3 was found in thrombi and its intravenous administration induced clot formation [159].
Thrombotic microangiopathy (TMA) is life threatening conditions and causes massive microvascular thrombosis with thrombocytopenia, microangiopathic hemolytic anemia and MODS [160]. High level of serum DNA-histone complexes and MPO in TMA patients implies the involvement of NETs in disease severity [161]. In mice model, treatment with DNase and PAD 4 inhibitor blocks DVT which reflects the involvement of NETs as potential agonist of thrombosis [153,162]. NE is a NET component and in an experimental model of NE-deficient mice, thrombosis was found to be ameliorated [163]. Furthermore, low levels plasma of DNase I activity was reported in thrombotic patients which also passively proof the involvement of NET in thrombosis [164].
2.8. NETs in Diabetes
Diabetes mellitus (DM) is an array of metabolic diseases with clinical features of hyperglycemic condition that arise due to impairment in insulin action, secretion or the two. Hyperglycemia is connected priming neutrophils for oxidative burst and ROS generation. Neutrophils and NETs are involved in the pathogenesis of both insulin deficiency type 1 diabetes (T1DM) and insulin resistance type 2 (T2DM) and subsequent complications [165]. In T1DM patients with disease severity of less than 1 year, circulating protein levels and enzymatic activities of proteinase 3 (PR3), NE and MPO-DNA are found significantly higher compared to healthy controls which indicate the amplified NET formation [166]. In experimental model of T1DM, inhibition of neutrophil function and NETosis ablate the progression of diabetes [167]. In T2DM patients elevated level of NETs, neutrophil elastase, mono- and oligonucleosomes and cf-DNA are observed compared to healthy donors [168,169]. NETs are also responsible in pathogenesis of diabetes induced complications. PAD 4, an enzyme taking part in chromatin decondensation during NETosis cascade and citrullinated histones, were markedly elevated in both T1DM and T2DM which implicated poor wound healing in both mice and human and interestingly PAD4 inhibition with Cl amidine and application of DNase restore the wound healing [170,171]. A case control study reported the elevated levels of circulating NET components; DNA-histones and NE are associated with the development diabetes induced retinopathy [172]. NETs are also reported in the sera of diabetic neuropathy [173] and nephropathy [174] patients.
2.9. NETs in Autoimmune Diseases
NETs and histones are strongly associated with autoimmune diseases [175]. NET exposure in the presence of B-cells like immune cells develops antibodies directed against self-nucleic acids and cytoplasmic proteins such as MPO and PR 3 [176]. Prolong presence of NETs trigger the production of anti-neutrophil cytoplasmic antibodies and anti-nuclear antibodies and vice-versa which promotes vasculitis and enhances the autoimmune response [177]. NETs have been implicated in numerous autoimmune diseases like systemic lupus erythematosus (SLE), rheumatoid arthritis (RA)[178,179]. RA is a chronic, deleterious systemic autoinflammatory disease of the synovial joints and SLE is a complex and heterogeneous disease with characteristic feature of the systemic, multi-organ inflammation. Elevated propensity of spontaneous NET release along with high ROS, MPO and NE had been identified from isolated neutrophils in patients with RA and SLE compared to healthy controls along with higher nuclear translocation of PAD 4 [179,180,181]. Moreover, increased levels of NET formation and NET remnants MPO-DNA are also reported in the serum of RA patients [182]. In a rat model of RA, PAD4 inhibition by chloramidine compensated the NET induced inflammation and erosive changes [183]. Another RA model study reports the reduction of joint inflammation and erosion when NETs inhibition was implicated by monoclonal antibodies directed against citrullinated histones [184]. In SLE slow degradation of NETs and complement activation contributes to the disease progression [185]. In a murine model of SLE NET inhibition by DNase I constrain the development of anti-ssDNA and anti-histone antibodies [186] and PAD4 inhibition by chloramidine reduces SLE induced subsequent vascular injury and organ damage [187].
2.10. NETs in Cancer
Inflammation is a trademark of cancer and recent emerging evidences identified presence of neutrophils as infiltrating inflammatory immune cells with tumor [188,189]. Neutrophil plays a controversial dual pro-and anti-tumorigenic role in tumor biology [190,191]. Consequently the effects of NETs with tumor are also found in two reverse regulatory ways: pro-tumor effects that enhance cancer cell proliferation, invasion and metastasis and anti-tumor effects that inhibit proliferation and invasion [192]. In this section we will highlight the pro-tumorigenic role of NETs. Highly elevated levels of NETs deposition is marked in some malignant cancers with Ewing sarcoma [193], Lewis lung carcinoma (LLC) [194], breast cancers [195,196] and lymphoma [197].
Although the detail underlying mechanism is yet to be investigated but several published literature disclosed that NETs promote tumor development by the inhibition of apoptosis and proliferative effects and thus uphold tumor progression and metastasis [198,199]. Circulating tumor cells (CTC) are significantly involved in tumor metastasis. Interestingly due to its sticky mesh like architecture NETs can entrap and adhere CTC and take to an adjacent areas thus expanding tumor cell metastasis. In an animal model of sepsis, significant levels of NET associated tumor cells were clearly pictured upon injection of LLC cells compared to healthy group and cancellation of neutrophils void this effect by lowering CTC adhesion within the liver [200]. A group of researchers further re-confirmed that only NETs but not neutrophils are responsible to augment tumor metastasis [197,201]. Moreover, the interaction of NETs with tumor cells induces a severe cancerous phenotype in cancer cell [192].
NET components also play significant role in tumor progression [202]. NE has been described to have pro-tumorigenic activity and is found to induce proliferation and migration of tumor cells in vitro [192]. Lung adenocarcinoma model showed increased proliferation of tumor cell line when neutrophils are co-cultured with A549 cells and this proliferation is nullified when co-cultured with NE-/- neutrophils [203]. This pro-tumorigenic role of NE is implicated by the degradation of insulin receptor substrate-1 and subsequent activation of phosphatidaylinositol-4,5-bisphosphate 3-kinase [203,204]. In addition, NE also induce the release of pro-tumor factors like transforming growth factor ɑ, vascular endothelial growth factor and platelet-derived growth factor facilitating interaction with their respective receptors and favoring tumor progression [205].
Matrix metalloproteinase 9 (MMP-9) an integral parts of NETs degrades extracellular matrix and support tumor metastasis [5]. MMPs favors pro-tumorigenic course through tumor cell proliferation, impairing apoptosis, increased angiogenesis, invasion and metastasis [206,207,208]. In mice model this invasiveness of tumor is reversed in MMP-9-/- mice compared to wild type [207].
Cathepsin G aiding metastasis by increasing angiogenesis and tumor dispersion [209]. In animal model of breast cancer cathepsin G potentiate tumor aggregation in vasculature and form distal tumor emboli [210]. In patients with hepatocellular carcinoma NETs-associated cathepsin G induces tumor cells invasion [211]
2.11. NETs in Preeclampsia
Preeclampsia is a pregnancy disorder with characteristics of inflammation, hypertension, kidney failure and seizures and significant cause of maternal and neonatal mortality worldwide [212]. Placental micro-particle induced neutrophil extracellular DNA lattices are found in preeclampsia [23]. A huge elevation of circulating cf-DNA is measured from the maternal plasma [213]. A published literature indicates inappropriate NET formation as the source of this cf-DNA [214]. Elevated levels of microparticles from preeclampsia patients could trigger NETs in isolated neutrophils though IL-8 dependent manner [215]. Moreover, NET-components have been detected by immunofluorescence staining in the intervillous space of preeclamptic placentae [23,216].Thus exacerbated NETosis lead to placental tissue damage in preeclamptic patients.
2.12. NETs in Kawasaki Disease
Kawasaki disease (KD) is an acute multisystem vasculitis syndrome with characteristic feature of febrile illness mainly affects infants and children in the developed countries [217,218]. EC activation and injury is observed in patients with KD [219]. Although the etiology and mechanism is not crystal clear, however, KD patients are very susceptible to acquire coronary artery abnormalities and myocardial ischemia [220,221]. The endothelial glycocalyx is a carbohydrate-rich gel-like layer lining buildup of syndecan, hyaluronic acid, chondroitin sulfate, and heparan sulfate etc. [221]. A recent report demonstrates circulating endothelial glycocalyx proteins syndecan-1 and hyaluronan as predictive biomarker of coronary artery lesions in KD [222]. Interestingly NETs also promote cardiovascular EC damages suggesting that enhanced NET generation may participate to the pathogenesis of KD vasculitis [223]. Again, high levels of proinflammatory mediators including TNF-α, IL-1β and IL-8 have been reported in KD patients which are strong inducer of neutrophils to release NETs [22,218]. The activation of neutrophils respiratory burst assessed by flow cytometry assay using dihydrorhodamine is described in the acute phase of KD [224]. Isolated neutrophils from acute phase KD patient’s shows enhanced NET formation compared to healthy controls [225,226]. Moreover, NE plasma level is also higher in acute phase KD [227]. Thus, NET and NET components contribute to KD and subsequent squeals.
3. Evaluation of NETs Inhibition as Therapeutic Targets
NETs are generated from the matured activated neutrophils in response to a wide array of infectious microbial pathogens and sterile stimuli and act as an indispensable protective barrier of innate immunity. Expelled NETs entrap and protect the dissemination of invading microorganisms throughout the body either by killing or confining them in its web-like structure. In contrast, aberrant activation of neutrophils and excessive production of NETs exaggerate inflammatory response that is likely to contribute to different diseases including infectious and non-infectious. Thus, NETs act as double-edged sword due to their dual controversial role. Albeit the microbial infection would be eradicated by using broad-spectrum antibiotics in ICU, the issue of stopping the NET induced systemic effects has come forward. Recent evidence suggests that inhibition of NETosis does not hamper the killing capacity of neutrophils other than NETs [228]. The development of blocking NETs is in progress (Table 1) [229]. Inhibition of NETs by DNase and other inhibitors postulates benefits in the context of thrombosis [230], ischemia reperfusion injury [231], SLE [232], CF [107], AMI, stroke, diabetes [233] and cancer [234].
The therapeutic target and the time of initiation of the therapies are an important concern in NET situation. Because NETs are beneficial for the host in the early phase of the infections and become detrimental and backfire in later stage. So, the fine tuning of NET formation throughout the disease course would be the goal for the development of new NET targeted therapies. Early detection of NETs is essential to start the therapy and perhaps a combined therapeutic approach; inhibition of stimulation and damaged tissue repair would be the best option for treatment.
4. Conclusions
From discovery in 2004, NETs have long been recognized as a novel central mechanism of innate immunity for their beneficial physiological role in host defense. But their pathogenic role has attracted recent attention. Several factor including aberrant activation, dysregulation and excessive generation determines their controversial role. The list of NET implicated diseases are gradually expanding ranges from autoimmune disorder to diabetes to cancer [170,232,236]. Although new novel therapeutic strategies are evolving targeting NETs but still, further and more extensive researches are needed to explore their implication in different diseases.
Author Contribution: All authors (M.M.I., N.T.) have contributed to the original draft preparation, read and agreed to the published version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rosales, C. Neutrophils at the crossroads of innate and adaptive immunity. J Leukoc Biol. 2020, 108, 377–396. [Google Scholar] [CrossRef] [PubMed]
- Pruchniak, MP.; Arazna, M.; Demkow, U. Life of neutrophil: from stem cell to neutrophil extracellular trap. Respir Physiol Neurobiol 2013, 187, 68–73. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types. Front Physiol 2018, 9, 113. [Google Scholar] [CrossRef]
- Phillipson, M; Kubes, P. The neutrophil in vascular inflammation. Nat Med 2011, 17, 1381–1390. [Google Scholar] [CrossRef]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat Rev Immunol 2013, 13, 159–175. [Google Scholar] [CrossRef]
- Brinkmann, V.; Zychlinsky, A. Neutrophil extracellular traps: is immunity the second function of chromatin? J Cell Biol 2012, 198, 773–783. [Google Scholar] [CrossRef]
- Borregaard, N. Neutrophils, from marrow to microbes. Immunity 2010, 33, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Greenlee-Wacker, MC. Clearance of apoptotic neutrophils and resolution of inflammation. Immunol Rev 2016, 273, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Mortaz, E.; Alipoor, S.D.; Adcock, I.M. ; Mumby, S; Koenderman, L. Update on Neutrophil Function in Severe Inflammation. Front Immunol 2018, 9, 2171. [Google Scholar] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; et al. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Wang, X.; Yousefi, S.; Simon, HU. Necroptois and neutrophil-associated disorders. Cell Death Dis 2018, 9, 111. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.I.; Wang, Y.; Liu, Q.; Li, H-R.; Yu, C-M.; Li, P.; Deng, X-M.; Wang, J-F. Dysregulation of neutrophil death in sepsis. Front Immunol 2022, 13, 963955. [Google Scholar] [CrossRef]
- Ganesh, K.; Joshi, M.B. Neutrophil sub-types in maintaining immune homeostasis during steady state, infections and sterile inflammation. Inflamm Res 2023, 72, 1175–1192. [Google Scholar] [CrossRef] [PubMed]
- Adrover, J.M.; Fresno, C.D.; Crainiciuc, G.; Cuartero, M.I.; Casanova-Acebes, M.; Weiss, L.A.; Huerga-Encabo, H.; Silvestre-Roig, C.; Rossaint, J.; Cossío, I.; Lechuga-Vieco, A.V.; García-Prieto, J.; Gómez-Parrizas, M.; Quintana, J.A.; Ballesteros, I.; Martin-Salamanca, S.; Aroca-Crevillen, A. A Neutrophil Timer Coordinates Immune Defense and Vascular Protection. Immunity 2019, 50, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Drifte, G,; Dunn-Siegrist, I. ; Tissières, P.; Pugin, J. Innate immune functions of immature neutrophils in patients with sepsis and severe systemic inflammatory response syndrome. Crit Care Med 2013, 41, 820–832. [Google Scholar] [CrossRef]
- Lipp, P.; Ruhnau, J.; Lange, A.; Vogelgesang, A.; Dressel, A.; Heckmann, M. Less Neutrophil Extracellular Trap Formation in Term Newborns than in Adults. Neonatology 2017, 111, 182–188. [Google Scholar] [CrossRef]
- Saitoh, T. , Komano, J.; Saitoh, Y. et al. Neutrophil extracellular traps mediate a host defense response to human immunodeficiency virus-1. Cell Host Microbe 2012, 12, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Gazendam, R.P.; van de Geer, A.; Roos, D.; van den Berg, T.K.; Kuijpers, T.W. How Neutrophils Kill Fungi. Immunol Rev 2016, 273, 299–311. [Google Scholar] [CrossRef]
- Yufei, H.; Liu, J.; Chen, Y.; Yan, L.; Wu, J. Neutrophil Extracellular Traps in Candida albicans Infection. Front Immunol 2022, 13, 913028. [Google Scholar]
- Babatunde, K.A.; Adenuga, O.F. Neutrophils in malaria: A double-edged sword role. Front Immunol, 2022, 13, 922377. [Google Scholar] [CrossRef]
- Thiam, HR.; Wong, SL.; Wagner, D.D. Waterman CM. Cellular Mechanisms of NETosis. Annu Rev Cell Dev Biol. 2020, 36, 191–218. [Google Scholar] [CrossRef] [PubMed]
- Keshari, R.S.; Jyoti, A.; Dubey, M.; et al. Cytokines induced neutrophil extracellular traps formation: implication for the inflammatory disease condition. PLoS One 2012, 7, e48111. [Google Scholar] [CrossRef]
- Omeljaniuk, W.J.; Jabłońska, E.; Garley, M.; Pryczynicz, A.; Ratajczak-Wrona, W.; Socha, K.; Borawska, M.H.; Charkiewicz, A.E. Biomarkers of neutrophil extracellular traps (NETs) and nitric oxide-(NO)-dependent oxidative stress in women who miscarried. Sci Rep 2020, 10, 13088. [Google Scholar] [CrossRef]
- Su, M.; Chen, C.; Li, S.; Li, M.; Zeng, Z.; Zhang, Y.; Xia, L.; Li, X.; Zheng, D.; Lin, Q.; Fan, X.; Wen, Y.; Liu, Y.; Chen, F.; Luo, W.; Bu, Y.; Qin, J.; Guo, M.; Qiu, M.; Sun, L.; Liu, R.; Wang, P.; Hwa, J.; Tang, W.H. Gasdermin D-dependent platelet pyroptosis exacerbates NET formation and inflammation in severe sepsis. Nat Cardiovasc Res 2022, 1, 698–699. [Google Scholar] [CrossRef] [PubMed]
- Schorn, C.; Janko, C.; Latzko, M.; Chaurio, R.; Schett, G. Herrmann M. Monosodium urate crystals induce extracellular DNA traps in neutrophils, eosinophils, and basophils but not in mononuclear cells. Front Immunol 2012, 3, 277. [Google Scholar] [CrossRef] [PubMed]
- Behnen, M.; Leschczyk, C.; Möller, S.; et al. Immobilized immune complexes induce neutrophil extracellular trap release by human neutrophil granulocytes via FcγRIIIB and Mac-1. J Immunol 2014, 193, 1954–1965. [Google Scholar] [CrossRef]
- Yalavarthi, S.; Gould, T.J.; Rao, A.N.; et al. Release of neutrophil extracellular traps by neutrophils stimulated with antiphospholipid antibodies: a newly identified mechanism of thrombosis in the antiphospholipid syndrome. Arthritis Rheumatol 2015, 67, 2990–3003. [Google Scholar] [CrossRef]
- Ortiz-Espinosa, S.; Morales, X.; Senent, Y.; et al. Complement C5a induces the formation of neutrophil extracellular traps by myeloid-derived suppressor cells to promote metastasis. Cancer Lett 2022, 529, 70–84. [Google Scholar] [CrossRef]
- Park J, Wysocki RW, Amoozgar Z, et al. Cancer cells induce metastasis-supporting neutrophil extracellular DNA traps. Sci Transl Med. 2016, 8, 138. [Google Scholar]
- Rayes, R.F.; Mouhanna, J.G.; Nicolau, I.; et al. Primary tumors induce neutrophil extracellular traps with targetable metastasis promoting effects. JCI Insight 2019, 5, e128008. [Google Scholar] [CrossRef]
- Boone, B.A.; Orlichenko, L.; Schapiro, N.E.; et al. The receptor for advanced glycation end products (RAGE) enhances autophagy and neutrophil extracellular traps in pancreatic cancer. Cancer Gene Ther 2015, 22, 326–334. [Google Scholar] [CrossRef]
- McDonald, B.; Urrutia, R.; Yipp, B.G.; Jenne, C.N.; Kubes, P. Intravascular neutrophil extracellular traps capture bacteria from the bloodstream during sepsis. Cell Host Microbe 2012, 12, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Aube, F.A.; Bidias, A.; Pépin, G. Who and how, DNA sensors in NETs-driven inflammation. Front Immunol 2023, 14, 1190177. [Google Scholar] [CrossRef]
- Ravindran, M.; Khan, M.A.; Palaniyar, N. Neutrophil Extracellular Trap Formation: Physiology, Pathology, and Pharmacology. Biomolecules 2019, 9, 365. [Google Scholar] [CrossRef]
- Azzouz, D.; Khan, M.A.; Sweezey, N.; Palaniyar, N. Two-in-one: UV radiation simultaneously induces apoptosis and NETosis. Cell Death Discov 2018, 4, 51. [Google Scholar] [CrossRef]
- Douda, D.N.; Yip, L.; Khan, M.A.; Grasemann, H.; Palaniyar, N. Akt is essential to induce NADPH-dependent NETosis and to switch the neutrophil death to apoptosis. Blood 2014, 123, 597–600. [Google Scholar] [CrossRef]
- Douda, D. N.; Khan, M. A.; Grasemann, H.; Palaniyar, N. SK3 channel and mitochondrial ROS mediate NADPH oxidase-independent NETosis induced by calcium influx. Proc Natl Acad Sci U S A 2015, 112, 2817–2822. [Google Scholar] [CrossRef] [PubMed]
- Rohrbach, A. S.; Slade, D. J.; Thompson, P. R.; Mowen, K. A. Activation of PAD4 in NET formation. Front Immunol 2012, 3, 360. [Google Scholar] [CrossRef] [PubMed]
- Metzler, K. D.; Goosmann, C.; Lubojemska, A.; Zychlinsky, A.; Papayannopoulos, V. A myeloperoxidase-containing complex regulates neutrophil elastase release and actin dynamics during NETosis. Cell Rep 2014, 2014. 8, 883–896. [Google Scholar] [CrossRef]
- Azzouz, D.; Palaniyar, N. ApoNETosis: discovery of a novel form of neutrophil death with concomitant apoptosis and NETosis. Cell Death Dis 2018, 9, 839. [Google Scholar] [CrossRef]
- Khan, MA.; Palaniyar, N. Transcriptional firing helps to drive NETosis. Sci Rep 2017, 7, 41749. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V. ; Neutrophil extracellular traps in immunity and disease. Nat Rev Immunol 2018, 18, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.S.; O'Brien, X.M.; Johnson, C.M.; Lavigne, L.M.; Reichner, J.S. An extracellular matrix-based mechanism of rapid neutrophil extracellular trap formation in response to Candida albicans. J Immunol 2013, 190, 4136–4148. [Google Scholar] [CrossRef]
- Vorobjeva, N.V.; Chernyak, B.V. NETosis: Molecular Mechanisms, Role in Physiology and Pathology. Biochemistry 2020, 85, 1178–1190. [Google Scholar] [CrossRef]
- Yipp, B.G.; Kubes, P. NETosis: how vital is it? Blood 2013, 122, 2784–2794. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Rizo, V.; Martínez-Guzmán, M.A.; Iñiguez-Gutierrez, L.; García-Orozco, A.; Alvarado-Navarro, A.; Fafutis-Morris, M. Neutrophil Extracellular Traps and Its Implications in Inflammation: An Overview. Front Immunol 2017, 8, 81. [Google Scholar] [CrossRef]
- Liang, C.; Lian, N.; Li. M. The emerging role of neutrophil extracellular traps in fungal infection. Front Cell Infect Microbiol, 2022, 12, 900895. [Google Scholar] [CrossRef]
- Omar, M.; Abdelal, H. NETosis in Parasitic Infections: A Puzzle That Remains Unsolved. Int J Mol Sci 2023, 24, 8975. [Google Scholar] [CrossRef]
- Diaz-Godinez, C.; Carrero, J.C. The state of art of neutrophil extracellular traps in protozoan and helminthic infections. Biosci Rep 2019, 39, BSR20180916. [Google Scholar] [CrossRef]
- Bartneck, M.; Keul, H.A.; Zwadlo-Klarwasser, G.; Groll, J. Phagocytosis independent extracellular nanoparticle clearance by human immune cells. Nano Lett 2010, 10, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Meurer, M.; Öhlmann, S.; Bonilla, M.C.; Valentin-Weigand, P.; Beineke, A.; Hennig-Pauka, I.; Schwerk, C.; Schroten, H.; Baums, C.G.; Köckritz-Blickwede, M.V.; de Buhr, N. Role of Bacterial and Host DNases on Host-Pathogen Interaction during Streptococcus suis Meningitis. Int J Mol Sci 2020, 21, 5289. [Google Scholar] [CrossRef] [PubMed]
- Nel, Jan G. ; Theron, Annette J.; Pool, Roger; Durandt, Chrisna; Tintinger, Gregory R.; Anderson, Ronald. Neutrophil extracellular traps and their role in health and disease. S Afr J Sci 2016, 112, 1–9. [Google Scholar]
- Liao, C.; Mao, F.; Qian, M.; Wang, X. Pathogen-Derived Nucleases: An Effective Weapon for Escaping Extracellular Traps. Front Immunol 2022, 13, 899890. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.D.; Malachowa, N.; DeLeo, F.R. Neutrophils and Bacterial Immune Evasion. J Innate Immun 2018, 10, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Storisteanu, D.M.; M Pocock, J.M.; Cowburn, A.S.; Juss, J.K.; Nadesalingam, A.; Nizet, V.; Chilvers, E.R. Evasion of neutrophil extracellular traps by respiratory pathogens. Am J Respir Cell Mol Biol 2017, 56, 423–431. [Google Scholar] [CrossRef]
- Wartha, F.; Beiter, K.; Albiger, B.; Fernebro, J.; Zychlinsky, A.; Normark, S.; Henriques-Normark, B. Capsule and D-alanylated lipoteichoic acids protect Streptococcus pneumoniae against neutrophil extracellular traps. Cell Microbiol 2007, 9, 1162–1171. [Google Scholar] [CrossRef]
- Branzk, N.; Lubojemska, A.; Hardison, S. E.; Wang, Q.; Gutierrez, M. G.; Brown, G. D.; Papayannopoulos, V. Neutrophils sense microbe size and selectively release neutrophil extracellular traps in response to large pathogens. Nat Immunol 2014, 15, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Low, C. Y.; Rotstein, C. Emerging fungal infections in immunocompromised patients. F1000 Med Rep 2011, 3, 14. [Google Scholar] [CrossRef]
- Zhong, H.; Lu, R.Y.; Wang, Y. Neutrophil extracellular traps in fungal infections: A seesaw battle in hosts. Front Immunol 2022, 13, 977493. [Google Scholar] [CrossRef]
- Marcos-Jubilar, M.; Lecumberri, R.; Páramo, J. Immunothrombosis: Molecular Aspects and New Therapeutic Perspectives. J Clin Med 2023, 12, 1399. [Google Scholar] [CrossRef]
- Iba, T.; Miki, T.; Hashiguchi, N.; Tabe, Y.; Nagaoka, I. Is the neutrophil a 'prima donna' in the procoagulant process during sepsis? Crit Care 2014, 18, 230. [Google Scholar] [CrossRef] [PubMed]
- Stiel, L.; Meziani, F.; Helms, J. Neutrophil Activation During Septic Shock. Shock 2018, 49, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Alcázar, M.; Kim, N.; Fuchs, T. A. Circulating Extracellular DNA: Cause or Consequence of Thrombosis? Semin Thromb Hemost 2017, 43, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Abrams, S.T.; Morton, B.; Alhamdi, Y.; Alsabani, M.; Lane, S.; Welters, I. D.; Wang, G.; Toh, C.H. A Novel Assay for Neutrophil Extracellular Trap Formation Independently Predicts Disseminated Intravascular Coagulation and Mortality in Critically Ill Patients. Am J Respir Crit Care Med 2019, 200, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Fogg, D. K.; Kaplan, M. J. A novel image-based quantitative method for the characterization of NETosis. J Immunol Methods 2015, 423, 104–110. [Google Scholar] [CrossRef]
- Kano, H.; Aminul Huq, M.; Tsuda, M.; Noguchi, H.; Takeyama, N. Sandwich ELISA for Circulating Myeloperoxidase- and Neutrophil Elastase-DNA Complexes Released from Neutrophil Extracellular Traps. Advanced Techniques in Biology & Medicine 2016, 5, 196. [Google Scholar]
- Islam, M.M.; Salma, U.; Irahara, T.; Watanabe, E.; Takeyama, N. Quantifying Myeloperoxidase-DNA and Neutrophil Elastase-DNA Complexes from Neutrophil Extracellular Traps by Using a Modified Sandwich ELISA. J Vis Exp 2023, 195, 64644. [Google Scholar] [CrossRef]
- Sil, P.; Yoo, D. G.; Floyd, M.; Gingerich, A.; Rada, B. High Throughput Measurement of Extracellular DNA Release and Quantitative NET Formation in Human Neutrophils In Vitro. J Vis Exp 2016, 112, 52779. [Google Scholar]
- Lelubre, C.; Vincent, J.L. Mechanisms and treatment of organ failure in sepsis. Nat Rev Nephrol 2018, 14, 417–427. [Google Scholar] [CrossRef]
- Liaw, P. C.; Ito, T.; Iba, T.; Thachil, J.; Zeerleder, S. DAMP and DIC: The role of extracellular DNA and DNA-binding proteins in the pathogenesis of DIC. Blood Rev 2016, 30, 257–261. [Google Scholar] [CrossRef]
- Gardiner, E. E.; Andrews, R. K. Neutrophil extracellular traps (NETs) and infection-related vascular dysfunction. Blood Rev 2012, 26, 255–259. [Google Scholar] [CrossRef]
- Maruchi, Y.; Tsuda, M.; Mori, H.; Takenaka, N.; Gocho, T.; Huq, M.A.; Takeyama, N. Plasma myeloperoxidase-conjugated DNA level predicts outcomes and organ dysfunction in patients with septic shock. Crit Care 2018, 22, 176. [Google Scholar] [CrossRef] [PubMed]
- Stiel, L.; Mayeur-Rousse, C.; Helms, J.; Meziani, F.; Mauvieux, L. First visualization of circulating neutrophil extracellular traps using cell fluorescence during human septic shock-induced disseminated intravascular coagulation. Thromb Res 2019, 183, 153–158. [Google Scholar] [CrossRef]
- Lenz, M.; Maiberger, T.; Armbrust, L.; Kiwit, A.; Von der Wense, A.; Reinshagen, K.; Elrod, J.; Boettcher, M. cfDNA and DNases: New Biomarkers of Sepsis in Preterm Neonates—A Pilot Study. Cells 2022, 11, 192. [Google Scholar] [CrossRef] [PubMed]
- Lerman, Y.V.; Kim, M. Neutrophil migration under normal and sepsis conditions. Cardiovasc Hematol Disord Drug Targets 2015, 15, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Joshi, M.B.; Philippova, M.; Erne, P.; Hasler, P.; Hahn, S.; Resink, T.J. Activated endothelial cells induce neutrophil extracellular traps and are susceptible to NETosis-mediated cell death. FEBS Lett 2010, 584, 3193–3197. [Google Scholar] [CrossRef]
- Iba, T.; Levi, M.; Levy, J.H. Intracellular communication and immunothrombosis in sepsis. J Thromb Haemost, 2022, 20, 2475–2484. [Google Scholar] [CrossRef]
- Ogura, H.; Gando, S.; Saitoh, D.; Takeyama, N.; Kushimoto, S.; Fujishima, S.; Mayumi, T.; Araki, T.; Ikeda, H.; Kotani, J.; Miki, Y.; Shiraishi, S.; Suzuki, K.; Suzuki, Y.; Takuma, K.; Tsuruta, R.; Yamaguchi, Y.; Yamashita, N.; Aikawa, N. ; Epidemiology of severe sepsis in Japanese intensive care units: a prospective multicenter study. J Infect Chemother 2014, 20, 157–162. [Google Scholar] [CrossRef]
- Pfeiler, S.; Stark, K.; Massberg, S.; Engelmann, B. Propagation of thrombosis by neutrophils and extracellular nucleosome networks. Haematologica 2017, 102, 206–213. [Google Scholar] [CrossRef]
- Delabranche, X.; Stiel, L.; Severac, F.; Galoisy, A.C.; Mauvieux, L.; Zobairi, F.; Lavigne, T.; Toti, F.; Anglès-Cano, E.; Meziani, F.; Boisramé-Helms, J. Evidence of Netosis in Septic Shock-Induced Disseminated Intravascular Coagulation. Shock 2017, 47, 313–317. [Google Scholar] [CrossRef]
- Li, Z.; Delaney, M.K.; O'Brien, K.A.; Du, X. Signaling during platelet adhesion and activation. Arterioscler Thromb Vasc Biol 2010, 30, 2341–2349. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, H.; Qu, M.; Nan, K.; Cao, H.; Cata, J.P.; Chen, W.; Miao, C. Review: The Emerging Role of Neutrophil Extracellular Traps in Sepsis and Sepsis-Associated Thrombosis. Front Cell Infect Microbiol 2021, 11, 653228. [Google Scholar] [CrossRef] [PubMed]
- Sreeramkumar, V.; Adrover, J.M.; Ballesteros, I.; Cuartero, M.I.; Rossaint, J.; Bilbao, I.; Nácher, M.; Pitaval, C.; Radovanovic, I.; Fukui, Y.; McEver, R. P.; Filippi, M.D.; Lizasoain, I.; Ruiz-Cabello, J.; Zarbock, A.; Moro, M.A.; Hidalgo, A. Neutrophils scan for activated platelets to initiate inflammation. Science 2014, 346, 1234–1238. [Google Scholar] [CrossRef] [PubMed]
- Etulain, J.; Martinod, K.; Wong, S.L.; Cifuni, S.M.; Schattner, M.; Wagner, D.D. P-selectin promotes neutrophil extracellular trap formation in mice. Blood 2015, 126, 242–246. [Google Scholar] [CrossRef]
- Carestia, A.; Kaufman, T.; Rivadeneyra, L.; Landoni, V.I.; Pozner, R.G.; Negrotto, S.; D'Atri, L.P.; Gómez, R.M.; Schattner, M. Mediators and molecular pathways involved in the regulation of neutrophil extracellular trap formation mediated by activated platelets. J Leukoc Biol 2016, 99, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Szatmary, P.; Huang, W.; Criddle, D.; Tepikin, A.; Sutton, R. Biology, role and therapeutic potential of circulating histones in acute inflammatory disorders. J Cell Mol Med 2018, 22, 4617–4629. [Google Scholar] [CrossRef]
- Potey, P.M.; Rossi, A.G.; Lucas, C.D.; Dorward, D.A. Neutrophils in the initiation and resolution of acute pulmonary inflammation: understanding biological function and therapeutic potential. J Pathol 2019, 247, 672–685. [Google Scholar] [CrossRef]
- Ranieri, V.M.; Rubenfeld, G.D.; Thompson, B.T.; Ferguson, N.D.; Caldwell, E.; Fan, E.; Camporota, L.; Slutsky, A. S. Acute respiratory distress syndrome: the Berlin Definition. JAMA 2012, 307, 2526–2533. [Google Scholar]
- Ramji, H.F.; Hafiz, M.; Altaq, H.H.; Hussain, S.T.; Chaudry, F. Acute Respiratory Distress Syndrome; A Review of Recent Updates and a Glance into the Future. Diagnostics 2023, 13, 1528. [Google Scholar] [CrossRef]
- Bellani, G.; Laffey, J.G.; Pham, T.; Fan, E.; Brochard, L.; Esteban, A.; Gattinoni, L.; van Haren, F.; Larsson, A.; McAuley, D.F.; Ranieri, M.; Rubenfeld, G.; Thompson, B.T.; Wrigge, H.; Slutsky, A.S.; Pesenti, A. ; Epidemiology, Patterns of Care, and Mortality for Patients With Acute Respiratory Distress Syndrome in Intensive Care Units in 50 Countries. JAMA, 2016, 315, 788–800. [Google Scholar] [CrossRef]
- Grommes, J.; Soehnlein, O. Contribution of neutrophils to acute lung injury. Mol Med 2011, 17, 293–307. [Google Scholar] [CrossRef]
- Narasaraju, T.; Yang, E.; Samy, R.P.; Ng, H.H.; Poh, W.P.; Liew, A.A.; Phoon, M.C.; van Rooijen, N.; Chow, V.T. Excessive neutrophils and neutrophil extracellular traps contribute to acute lung injury of influenza pneumonitis. Am J Pathol 2011, 179, 199–210. [Google Scholar] [CrossRef]
- Yang, S.C.; Yung-Fong Tsai, Y.F.; Yen-Lin Pan, Y.L.; Tsong-Long Hwang, T.L. Understanding the role of neutrophils in acute respiratory distress syndrome. BioMed J 2021, 44, 439–446. [Google Scholar] [CrossRef]
- Mikacenic, C.; Moore, R.; Dmyterko, V.; West, T.E.; Altemeier, W.A.; Liles, W.C.; Lood, C. Neutrophil extracellular traps (NETs) are increased in the alveolar spaces of patients with ventilator-associated pneumonia. Crit Care 2018, 22, 358. [Google Scholar] [CrossRef]
- Scozzi, D.; Liao, F.; Krupnick, A.S.; Kreisel, D.; Gelman, A.E. The role of neutrophil extracellular traps in acute lung injury. Front Immunol 2022, 13, 953195. [Google Scholar] [CrossRef]
- Saffarzadeh, M.; Juenemann, C.; Queisser, M.A.; Lochnit, G.; Barreto, G.; Galuska, S.P.; Lohmeyer, J.; Preissner, K.T. Neutrophil extracellular traps directly induce epithelial and endothelial cell death: a predominant role of histones. PLoS One 2012, 7, e32366. [Google Scholar] [CrossRef]
- Abrams, S. T.; Zhang, N.; Manson, J.; Liu, T.; Dart, C.; Baluwa, F.; Wang, S. S.; Brohi, K.; Kipar, A.; Yu, W.; Wang, G.; Toh, C. H. Circulating histones are mediators of trauma-associated lung injury. Am J Respir Crit Care Med 2013, 187, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Bosmann, M.; Grailer, J.J.; Ruemmler, R.; Russkamp, N.F.; Zetoune, F.S.; Sarma, J.V.; Standiford, T. J.; Ward, P.A. Extracellular histones are essential effectors of C5aR- and C5L2-mediated tissue damage and inflammation in acute lung injury. FASEB J 2013, 27, 5010–5021. [Google Scholar] [CrossRef] [PubMed]
- Mori, H.; Huq, M.A.; Islam, M.M.; Takeyama, N. Neutrophil extracellular traps are associated with altered human pulmonary artery endothelial barrier function. European Journal of Inflammation 2021, 19, 1–10. [Google Scholar] [CrossRef]
- Garcia, C.C.; Weston-Davies, W.; Russo, R.C.; Tavares, L.P.; Rachid, M.A.; Alves-Filho, J.C.; Machado, A.V.; Ryffel, B.; Nunn, M.A.; Teixeira, M.M. Complement C5 activation during influenza A infection in mice contributes to neutrophil recruitment and lung injury. PLoS One 2013, 8, e64443. [Google Scholar] [CrossRef]
- Lefrançais, E.; Mallavia, B.; Zhuo, H.; Calfee, C.S.; Looney, M.R. Maladaptive role of neutrophil extracellular traps in pathogen-induced lung injury. JCI Insight 2018, 3, e98178. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Alemán, S.R.; Campos-García, L.; Palma-Nicolas, J.P.; Hernández-Bello, R.; González, G.M.; Sánchez-González, A. Understanding the Entanglement: Neutrophil Extracellular Traps (NETs) in Cystic Fibrosis. Front Cell Infect Microbiol 2017, 7, 104. [Google Scholar] [CrossRef] [PubMed]
- Manzenreiter, R.; Kienberger, F.; Marcos, V.; Schilcher, K.; Krautgartner, W.D.; Obermayer, A.; Huml, M.; Stoiber, W.; Hector, A.; Griese, M.; Hannig, M.; Studnicka, M.; Vitkov, L.; Hartl, D. Ultrastructural characterization of cystic fibrosis sputum using atomic force and scanning electron microscopy. J Cyst Fibros 2012, 11, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S. V.; Bruce, K. D. Bruce, The cystic fibrosis airway microbiome. Cold Spring Harb Perspect Med 2013, 3, a009738. [Google Scholar] [CrossRef]
- Yoo, D. G.; Floyd, M.; Winn, M.; Moskowitz, S.M.; Rada, B. NET formation induced by Pseudomonas aeruginosa cystic fibrosis isolates measured as release of myeloperoxidase-DNA and neutrophil elastase-DNA complexes. Immunol Lett 2014, 160, 186–194. [Google Scholar] [CrossRef]
- Piva, T.C.; Luft, C.; Antunes, K.H.; Marostica, P.J.C.; Pinto, L.A.; Donadio, M.V.F. Extracellular DNA in sputum is associated with pulmonary function and hospitalization in patients with cystic fibrosis. Respir Med 2020, 172, 106144. [Google Scholar] [CrossRef]
- Papayannopoulos, V.; Staab, D.; Zychlinsky, A. Neutrophil elastase enhances sputum solubilization in cystic fibrosis patients receiving DNase therapy. PLoS One 2011, 6, e28526. [Google Scholar] [CrossRef] [PubMed]
- Keir, H.R.; Chalmers, J.D. Neutrophil extracellular traps in chronic lung disease: implications for pathogenesis and therapy. Eur Respir Rev 2022, 31, 210241. [Google Scholar] [CrossRef]
- Rahman, S.; Gadjeva, M. Does NETosis Contribute to the Bacterial Pathoadaptation in Cystic Fibrosis? Front Immunol 2014, 5, 378. [Google Scholar] [CrossRef]
- Kim, H.Y.; DeKruyff, R.H.; Umetsu, D. T. The many paths to asthma: phenotype shaped by innate and adaptive immunity. Nat Immunol 2010, 11, 577–584. [Google Scholar] [CrossRef]
- Hammad, H.; Lambrecht, B.N. The basic immunology of asthma. Cell 2021, 18, 1469–1485. [Google Scholar] [CrossRef]
- Dworski, R.; Simon, H. U.; Hoskins, A.; Yousefi, S. Eosinophil and neutrophil extracellular DNA traps in human allergic asthmatic airways. J Allergy Clin Immunol 2011, 127, 1260–1266. [Google Scholar] [CrossRef]
- Hosoki, K.; Ying, S.; Corrigan, C.; Qi, H.; Kurosky, A.; Jennings, K.; Sun, Q.; Boldogh, I.; Sur, S. Analysis of a Panel of 48 Cytokines in BAL Fluids Specifically Identifies IL-8 Levels as the Only Cytokine that Distinguishes Controlled Asthma from Uncontrolled Asthma, and Correlates Inversely with FEV1. PLoS One 2015, 10, e0126035. [Google Scholar] [CrossRef]
- Pablo-Torres, C.; Izquierdo, E.I.; Tan, T.J.; Obeso, D.; Layhadi, J.A.; Sánchez-Solares, J.; Mera-Berriatua, L.; Bueno-Cabrera, J.L.; Reaño-Martos, M.D.M.; Iglesias-Cadarso, A.; Barbas, C.; Gomez-Casado, C.; Villaseñor, A.; Barber, D.; Shamji, M.H.; Escribese, M.M. Deciphering the role of platelets in severe allergy by an integrative omics approach. Allergy 2023, 78, 1319–1332. [Google Scholar] [CrossRef]
- Bonaventura, A.; Montecucco, F.; Dallegri, F. Cellular recruitment in myocardial ischaemia/reperfusion injury. Eur J Clin Invest 2016, 46, 590–601. [Google Scholar] [CrossRef]
- Mozzini, C.; Pagani, M. Cardiovascular Diseases: Consider Netosis. Curr Probl Cardiol 2021, 47, 100929. [Google Scholar] [CrossRef]
- Distelmaier, K.; Winter, M. P.; Dragschitz, F.; Redwan, B.; Mangold, A.; Gleiss, A.; Perkmann, T.; Maurer, G.; Adlbrecht, C.; Lang, I. M. Prognostic value of culprit site neutrophils in acute coronary syndrome. Eur J Clin Invest 2014, 44, 257–265. [Google Scholar] [CrossRef]
- Borissoff, J.I.; Joosen, I.A.; Versteylen, M.O.; Brill, A.; Fuchs, T.A.; Savchenko, A.S.; Gallant, M.; Martinod, K.; Ten Cate, H.; Hofstra, L.; Crijns, H.J.; Wagner, D.D.; Kietselaer, B.L.J.H. Elevated levels of circulating DNA and chromatin are independently associated with severe coronary atherosclerosis and a prothrombotic state. Arterioscler Thromb Vasc Biol 2013, 33, 2032–2040. [Google Scholar] [CrossRef] [PubMed]
- Megens, R.T.; Vijayan, S.; Lievens, D.; Döring, Y.; van Zandvoort, M.A.; Grommes, J.; Weber, C.; Soehnlein, O. Presence of luminal neutrophil extracellular traps in atherosclerosis. Thromb Haemost 2012, 107, 597–598. [Google Scholar] [CrossRef] [PubMed]
- Oklu, R.; Stone, J.R.; Albadawi, H.; Watkins, M.T. Extracellular traps in lipid-rich lesions of carotid atherosclerotic plaques: implications for lipoprotein retention and lesion progression. J Vasc Interv Radiol 2014, 25, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Zhou, X.; Ji, W.J.; Lu, R.Y.; Zhang, Y.; Zhang, Y.D.; Ma, Y.Q.; Zhao, J.H.; Li, Y. M. Neutrophil extracellular traps in ischemia-reperfusion injury-induced myocardial no-reflow: therapeutic potential of DNase-based reperfusion strategy. Am J Physiol Heart Circ Physiol 2015, 308, H500–H509. [Google Scholar] [CrossRef]
- Silvestre-Roig, C.; Braster, Q.; Wichapong, K.; Lee, E.Y.; Teulon, J.M.; Berrebeh, N.; Winter, J.; Adrover, J.M.; Santos, G.S.; Froese, A.; Lemnitzer, P.; Ortega-Gómez, A.; Chevre, R.; Marschner, J.; Schumski, A.; Winter, C.; Perez-Olivares, L.; Pan, C.; Paulin, N.; Schoufour, T.; … Soehnlein, O. . Externalized histone H4 orchestrates chronic inflammation by inducing lytic cell death. Nature 2019, 569, 236–240. [Google Scholar] [CrossRef]
- Pérez-Sánchez, C.; Ruiz-Limón, P.; Aguirre, M.A.; Jiménez-Gómez, Y.; Arias-de la Rosa, I.; Ábalos-Aguilera, M.C.; Rodriguez-Ariza, A.; Castro-Villegas, M.C.; Ortega-Castro, R.; Segui, P.; Martinez, C.; Gonzalez-Conejero, R.; Rodríguez-López, S.; Gonzalez-Reyes, J.A.; Villalba, J.M.; Collantes-Estévez, E.; Escudero, A.; Barbarroja, N.; López-Pedrera, C. Diagnostic potential of NETosis-derived products for disease activity, atherosclerosis and therapeutic effectiveness in Rheumatoid Arthritis patients. J Autoimmun 2017, 82, 31–40. [Google Scholar] [CrossRef]
- Stakos, D.A.; Kambas, K.; Konstantinidis, T.; Mitroulis, I.; Apostolidou, E.; Arelaki, S.; Tsironidou, V.; Giatromanolaki, A.; Skendros, P.; Konstantinides, S.; Ritis, K. Expression of functional tissue factor by neutrophil extracellular traps in culprit artery of acute myocardial infarction. Eur Heart J 2015, 36, 1405–1414. [Google Scholar] [CrossRef]
- Huang, H.; Tohme, S.; Al-Khafaji, A.B.; Tai, S.; Loughran, P.; Chen, L.; Wang, S.; Kim, J.; Billiar, T.; Wang, Y.; Tsung, A. Damage-associated molecular pattern-activated neutrophil extracellular trap exacerbates sterile inflammatory liver injury. Hepatology 2015, 62, 600–614. [Google Scholar] [CrossRef]
- Huang, H.; Chen, H.W.; Evankovich, J.; Yan, W.; Rosborough, B. R.; Nace, G.W.; Ding, Q.; Loughran, P.; Beer-Stolz, D.; Billiar, T.R.; Esmon, C.T.; Tsung, A. Histones activate the NLRP3 inflammasome in Kupffer cells during sterile inflammatory liver injury. J Immunol 2013, 191, 2665–2679. [Google Scholar] [CrossRef]
- Linkermann, A.; Chen, G.; Dong, G.; Kunzendorf, U.; Krautwald, S.; Dong, Z. Regulated cell death in AKI. J Am Soc Nephrol 2014, 25, 2689–2701. [Google Scholar] [CrossRef]
- Linkermann, A.; Stockwell, B.R.; Krautwald, S.; Anders, H.J. Regulated cell death and inflammation: an auto-amplification loop causes organ failure. Nat Rev Immunol 2014, 14, 759–767. [Google Scholar] [CrossRef]
- Allam, R.; Scherbaum, C.R.; Darisipudi, M.N.; Mulay, S.R.; Hägele, H.; Lichtnekert, J.; Hagemann, J.H.; Rupanagudi, K.V.; Ryu, M.; Schwarzenberger, C.; Hohenstein, B.; Hugo, C.; Uhl, B.; Reichel, C.A.; Krombach, F.; Monestier, M.; Liapis, H.; Moreth, K.; Schaefer, L.; Anders, H.J. Histones from dying renal cells aggravate kidney injury via TLR2 and TLR4. J Am Soc Nephrol 2012, 23, 1375–1388. [Google Scholar] [CrossRef]
- Kumar, S.V.; Kulkarni, O.P.; Mulay, S.R.; Darisipudi, M.N.; Romoli, S.; Thomasova, D.; Scherbaum, C.R.; Hohenstein, B.; Hugo, C.; Müller, S.; Liapis, H.; Anders, H.J. Neutrophil Extracellular Trap-Related Extracellular Histones Cause Vascular Necrosis in Severe GN. J Am Soc Nephrol 2015, 26, 2399–2413. [Google Scholar] [CrossRef]
- Ekaney, M.L.; Otto, G.P.; Sossdorf, M.; Sponholz, C.; Boehringer, M.; Loesche, W.; Rittirsch, D.; Wilharm, A.; Kurzai, O.; Bauer, M.; Claus, R.A. Impact of plasma histones in human sepsis and their contribution to cellular injury and inflammation. Crit Care 2014, 18, 543. [Google Scholar] [CrossRef]
- Nakazawa, D.; Kumar, S.V.; Marschner, J.; Desai, J.; Holderied, A.; Rath, L.; Kraft, F.; Lei, Y.; Fukasawa, Y.; Moeckel, G.W.; Angelotti, M.L.; Liapis, H.; Anders, H.J. Histones and Neutrophil Extracellular Traps Enhance Tubular Necrosis and Remote Organ Injury in Ischemic AKI. J Am Soc Nephrol 2017, 28, 1753–1768. [Google Scholar] [CrossRef]
- Allam, R.; Kumar, S.V.; Darisipudi, M.N.; Anders, H.J. Extracellular histones in tissue injury and inflammation. J Mol Med (Berl) 2014, 92, 465–472. [Google Scholar] [CrossRef]
- Zuo, Y.; Yalavarthi, S.; Shi, H.; Gockman, K.; Zuo, M.; Madison, J.A.; Blair, C.; Weber, A.; Barnes, B.J.; Egeblad, M.; Woods, R.J.; Kanthi, Y.; Knight, J.S. Neutrophil extracellular traps in COVID-19. JCI Insight, 2020, 5, e138999. [Google Scholar] [CrossRef]
- Janiuk, K.; Jabłońska, E.; & Garley, M.; & Garley, M. Significance of NETs Formation in COVID-19. Cells, 2021, 10, 151. [Google Scholar] [CrossRef]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; Tian, D.S. Dysregulation of Immune Response in Patients With Coronavirus 2019 (COVID-19) in Wuhan, China. Clin Infect Dis 2020, 71, 762–768. [Google Scholar] [CrossRef]
- Zhang, B.; Zhou, X.; Zhu, C.; Song, Y.; Feng, F.; Qiu, Y.; Feng, J.; Jia, Q.; Song, Q.; Zhu, B.; Wang, J. Immune Phenotyping Based on the Neutrophil-to-Lymphocyte Ratio and IgG Level Predicts Disease Severity and Outcome for Patients With COVID-19. Front Mol Biosci 2020, 7, 157. [Google Scholar] [CrossRef]
- Torres-Ruiz, J.; Absalón-Aguilar, A.; Nuñez-Aguirre, M.; Pérez-Fragoso, A.; Carrillo-Vázquez, D.A.; Maravillas-Montero, J.L.; Mejía-Domínguez, N.R.; Llorente, L.; Alcalá-Carmona, B.; Lira-Luna, J.; Núñez-Álvarez, C.; Juárez-Vega, G.; Meza-Sánchez, D.; Hernández-Gilsoul, T.; Tapia-Rodríguez, M.; Gómez-Martín, D. Neutrophil Extracellular Traps Contribute to COVID-19 Hyperinflammation and Humoral Autoimmunity. Cells 2021, 10, 2545. [Google Scholar] [CrossRef]
- Mahmudpour, M.; Roozbeh, J.; Keshavarz, M.; Farrokhi, S.; Nabipour, I. COVID-19 cytokine storm: The anger of inflammation. Cytokine 2020, 133, 155151. [Google Scholar] [CrossRef]
- Borges, L.; Pithon-Curi, T.C.; Curi, R.; Hatanaka, E. COVID-19 and Neutrophils: The Relationship between Hyperinflammation and Neutrophil Extracellular Traps. Mediators Inflamm 2020, 2020, 8829674. [Google Scholar] [CrossRef]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. ; COVID-19: consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; Cheng, Z.; Yu, T.; Xia, J.; Wei, Y.; Wu, W.; Xie, X.; Yin, W.; Li, H.; Liu, M.; Xiao, Y.; … Cao, B. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Barnes, B.J.; Adrover, J.M.; Baxter-Stoltzfus, A.; Borczuk, A.; Cools-Lartigue, J.; Crawford, J.M.; Daßler-Plenker, J.; Guerci, P.; Huynh, C.; Knight, J.S.; Loda, M.; Looney, M.R.; McAllister, F.; Rayes, R.; Renaud, S.; Rousseau, S.; Salvatore, S.; Schwartz, R.E.; Spicer, J.D.; Yost, C. C.; Weber, A.; Zuo, Y.; Egeblad, M. Targeting potential drivers of COVID-19: Neutrophil extracellular traps. J Exp Med 2020, 217, e20200652. [Google Scholar] [CrossRef]
- Schönrich, G.; Raftery, M.J.; Samstag, Y. Samstag, Devilishly radical NETwork in COVID-19: Oxidative stress, neutrophil extracellular traps (NETs), and T cell suppression. Adv Biol Regul 2020, 77, 100741. [Google Scholar] [CrossRef]
- Yaqinuddin, A.; Kashir, J. Novel therapeutic targets for SARS-CoV-2-induced acute lung injury: Targeting a potential IL-1β/neutrophil extracellular traps feedback loop. Med Hypotheses, 2020, 143, 109906. [Google Scholar] [CrossRef] [PubMed]
- Yaqinuddin, A.; Kvietys, P.; Kashir, J. COVID-19: Role of neutrophil extracellular traps in acute lung injury. Respir Investig 2020, 58, 419–420. [Google Scholar] [CrossRef]
- Allegra, A.; Innao, V.; Allegra, A.G.; Musolino, C. Coagulopathy and thromboembolic events in patients with SARS-CoV-2 infection: pathogenesis and management strategies. Ann Hematol 2020, 99, 1953–1965. [Google Scholar] [CrossRef]
- Middleton, E.A.; He, X.Y.; Denorme, F.; Campbell, R.A.; Ng, D.; Salvatore, S.P.; Mostyka, M.; Baxter-Stoltzfus, A.; Borczuk, A.C.; Loda, M.; Cody, M.J.; Manne, B.K.; Portier, I.; Harris, E.S.; Petrey, A.C.; Beswick, E.J.; Caulin, A.F.; Iovino, A.; Abegglen, L.M.; Weyrich, A.S.; Rondina, M.T.; Egeblad, M.; Schiffman, J.D.; Yost, C.C. Neutrophil extracellular traps contribute to immunothrombosis in COVID-19 acute respiratory distress syndrome. Blood, 2020, 136, 1169–1179. [Google Scholar] [CrossRef] [PubMed]
- Nicolai, L.; Leunig, A.; Brambs, S.; Kaiser, R.; Weinberger, T.; Weigand, M.; Muenchhoff, M.; Hellmuth, J.C.; Ledderose, S.; Schulz, H.; Scherer, C.; Rudelius, M.; Zoller, M.; Höchter, D.; Keppler, O.; Teupser, D.; Zwißler, B.; von Bergwelt-Baildon, M.; Kääb, S.; Massberg, S.; Pekayvaz, K.; Stark, K. Immunothrombotic Dysregulation in COVID-19 Pneumonia Is Associated With Respiratory Failure and Coagulopathy. Circulation 2020, 142, 1176–1189. [Google Scholar] [CrossRef]
- Ng, H.; Havervall, S.; Rosell, A.; Aguilera, K.; Parv, K.; von Meijenfeldt, F.A.; Lisman, T.; Mackman, N.; Thålin, C.; Phillipson, M. Circulating Markers of Neutrophil Extracellular Traps Are of Prognostic Value in Patients With COVID-19. Arterioscler Thromb Vasc Biol 2021, 41, 988–994. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Pérez, M.P.; Águila, S.; Reguilón-Gallego, L.; de Los Reyes-García, A.M.; Miñano, A.; Bravo-Pérez, C.; de la Morena, M.E.; Corral, J.; García-Barberá, N.; Gómez-Verdú, J.M.; Bernal, E.; Herranz, M.T.; Vicente, V.; Martínez, C.; González-Conejero, R.; Lozano, M. L. Neutrophil extracellular traps and von Willebrand factor are allies that negatively influence COVID-19 outcomes. Clin Transl Med 2021, 11, e268. [Google Scholar] [CrossRef]
- Gando, S.; Levi, M.; Toh, C.H. Disseminated intravascular coagulation. Nat Rev Dis Primers 2016, 2, 16037. [Google Scholar] [CrossRef]
- Fuchs, T.A.; Brill, A.; Duerschmied, D.; Schatzberg, D.; Monestier, M.; Myers, D.D.; Jr, Wrobleski, S. K.; Wakefield, T.W.; Hartwig, J.H.; Wagner, D.D. Extracellular DNA traps promote thrombosis. Proc Natl Acad Sci U S A, 2010, 107, 15880–15885. [Google Scholar] [CrossRef]
- McDonald, B.; Davis, R.P.; Kim, S.J.; Tse, M.; Esmon, C.T.; Kolaczkowska, E.; Jenne, C. N. Platelets and neutrophil extracellular traps collaborate to promote intravascular coagulation during sepsis in mice. Blood, 2017, 129, 1357–1367. [Google Scholar] [CrossRef]
- Tanaka, K.; Koike, Y.; Shimura, T.; Okigami, M.; Ide, S.; Toiyama, Y.; Okugawa, Y.; Inoue, Y.; Araki, T.; Uchida, K.; Mohri, Y.; Mizoguchi, A.; Kusunoki, M. In vivo characterization of neutrophil extracellular traps in various organs of a murine sepsis model. PLoS One, 2014, 9, e111888. [Google Scholar] [CrossRef]
- Komissarov, A.A.; Florova, G.; Idell, S. Effects of extracellular DNA on plasminogen activation and fibrinolysis. J Biol Chem 2011, 286, 41949–41962. [Google Scholar] [CrossRef]
- Longstaff, C.; Varjú, I.; Sótonyi, P.; Szabó, L.; Krumrey, M.; Hoell, A.; Bóta, A.; Varga, Z.; Komorowicz, E.; Kolev, K. Mechanical stability and fibrinolytic resistance of clots containing fibrin, DNA, and histones. J Biol Chem 2013, 288, 6946–6956. [Google Scholar] [CrossRef]
- Fuchs, T.A.; Bhandari, A.A.; Wagner, D.D. Histones induce rapid and profound thrombocytopenia in mice. Blood 2011, 118, 3708–3714. [Google Scholar] [CrossRef]
- Brill, A.; Fuchs, T.A.; Savchenko, A.S.; Thomas, G.M.; Martinod, K.; De Meyer, S.F.; Bhandari, A.A.; Wagner, D.D. Neutrophil extracellular traps promote deep vein thrombosis in mice. J Thromb Haemost 2012, 10, 136–144. [Google Scholar] [CrossRef]
- Iba, T.; Watanabe, E.; Umemura, Y.; Wada, T.; Hayashida, K.; Kushimoto, S. Japanese Surviving Sepsis Campaign Guideline Working Group for disseminated intravascular coagulation, & Wada, H. Sepsis-associated disseminated intravascular coagulation and its differential diagnoses. J Intensive Care 2019, 7, 32. [Google Scholar]
- Fuchs, T.A.; Kremer Hovinga, J.A.; Schatzberg, D.; Wagner, D.D.; Lämmle, B. Circulating DNA and myeloperoxidase indicate disease activity in patients with thrombotic microangiopathies. Blood 2012, 120, 1157–1164. [Google Scholar] [CrossRef]
- Martinod, K.; Demers, M.; Fuchs, T.A.; Wong, S.L.; Brill, A.; Gallant, M.; Hu, J.; Wang, Y.; Wagner, D.D. Neutrophil histone modification by peptidylarginine deiminase 4 is critical for deep vein thrombosis in mice. Proc Natl Acad Sci U S A, 2013, 110, 8674–8679. [Google Scholar] [CrossRef]
- Massberg, S.; Grahl, L.; von Bruehl, M.L.; Manukyan, D.; Pfeiler, S.; Goosmann, C.; Brinkmann, V.; Lorenz, M.; Bidzhekov, K.; Khandagale, A.B.; Konrad, I.; Kennerknecht, E.; Reges, K.; Holdenrieder, S.; Braun, S.; Reinhardt, C.; Spannagl, M.; Preissner, K.T.; Engelmann, B. Reciprocal coupling of coagulation and innate immunity via neutrophil serine proteases. Nat Med 2010, 16, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Alcázar, M.; Napirei, M.; Panda, R.; Köhler, E.C.; Kremer Hovinga, J.A.; Mannherz, H.G.; Peine, S.; Renné, T.; Lämmle, B.; Fuchs, T.A. Impaired DNase1-mediated degradation of neutrophil extracellular traps is associated with acute thrombotic microangiopathies. J Thromb Haemost 2015, 13, 732–742. [Google Scholar] [CrossRef] [PubMed]
- Njeim, R.; Azar, W.S.; Fares, A.H.; Azar, S. T.; Kfoury Kassouf, H.; Eid, A.A. NETosis contributes to the pathogenesis of diabetes and its complications. J Mol Endocrinol 2020, 65, R65–R76. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiao, Y.; Zhong, L.; Ye, D.; Zhang, J.; Tu, Y.; Bornstein, S.R.; Zhou, Z.; Lam, K.S.; Xu, A. Increased neutrophil elastase and proteinase 3 and augmented NETosis are closely associated with β-cell autoimmunity in patients with type 1 diabetes. Diabetes 2014, 63, 4239–4248. [Google Scholar] [CrossRef] [PubMed]
- Diana, J.; Simoni, Y.; Furio, L.; Beaudoin, L.; Agerberth, B.; Barrat, F.; Lehuen, A. Crosstalk between neutrophils, B-1a cells and plasmacytoid dendritic cells initiates autoimmune diabetes. Nat Med 2013, 19, 65–73. [Google Scholar] [CrossRef]
- Menegazzo, L.; Ciciliot, S.; Poncina, N.; Mazzucato, M.; Persano, M.; Bonora, B.; Albiero, M.; Vigili de Kreutzenberg, S.; Avogaro, A.; Fadini, G.P. NETosis is induced by high glucose and associated with type 2 diabetes. Acta Diabetol 2015, 52, 497–503. [Google Scholar] [CrossRef]
- Carestia, A.; Frechtel, G.; Cerrone, G.; Linari, M.A.; Gonzalez, C.D.; Casais, P.; Schattner, M. NETosis before and after Hyperglycemic Control in Type 2 Diabetes Mellitus Patients. PLoS One 2016, 11, e0168647. [Google Scholar] [CrossRef]
- Wong, S.L.; Demers, M.; Martinod, K.; Gallant, M.; Wang, Y.; Goldfine, A.B.; Kahn, C.R.; Wagner, D.D. Diabetes primes neutrophils to undergo NETosis, which impairs wound healing. Nat Med 2015, 21, 815–819. [Google Scholar] [CrossRef]
- Fadini, G.P.; Menegazzo, L.; Rigato, M.; Scattolini, V.; Poncina, N.; Bruttocao, A.; Ciciliot, S.; Mammano, F.; Ciubotaru, C.D.; Brocco, E.; Marescotti, M.C.; Cappellari, R.; Arrigoni, G.; Millioni, R.; Vigili de Kreutzenberg, S.; Albiero, M.; Avogaro, A. NETosis Delays Diabetic Wound Healing in Mice and Humans. Diabetes 2016, 65, 1061–1071. [Google Scholar] [CrossRef]
- Park, J.H.; Kim, J.E.; Gu, J.Y.; Yoo, H.J.; Park, S.H.; Kim, Y.I.; Nam-Goong, I.S.; Kim, E.S.; Kim, H.K. Evaluation of Circulating Markers of Neutrophil Extracellular Trap (NET) Formation as Risk Factors for Diabetic Retinopathy in a Case-Control Association Study. Exp Clin Endocrinol Diabetes 2016, 124, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhou, X.; Yin, Y.; Mai, Y.; Wang, D.; Zhang, X. Hyperglycemia Induces Neutrophil Extracellular Traps Formation Through an NADPH Oxidase-Dependent Pathway in Diabetic Retinopathy. Front Immunol 2018, 9, 3076. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, A.; Yamada, M.; Shida, H.; Nakazawa, D.; Kusunoki, Y.; Nakamura, A.; Miyoshi, H.; Tomaru, U.; Atsumi, T.; Ishizu, A. Circulating Neutrophil Extracellular Trap Levels in Well-Controlled Type 2 Diabetes and Pathway Involved in Their Formation Induced by High-Dose Glucose. Pathobiology 2016, 83, 243–251. [Google Scholar] [CrossRef]
- Alarcon, M.F.; McLaren, Z.; Wright, H.L. Neutrophils in the Pathogenesis of Rheumatoid Arthritis and Systemic Lupus Erythematosus: Same Foe Different M.O. Front Immunol 2021, 12, 649693. [Google Scholar] [CrossRef]
- Kessenbrock, K.; Krumbholz, M.; Schönermarck, U.; Back, W.; Gross, W.L.; Werb, Z.; Gröne, H.J.; Brinkmann, V.; Jenne, D.E. Netting neutrophils in autoimmune small-vessel vasculitis. Nat Med 2009, 15, 623–625. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Zhang, Y.; Yin, S.W.; Gao, X. J.; Shi, W.W.; Wang, Y.; Huang, X.; Wang, L.; Zou, L.Y.; Zhao, J.H.; Huang, Y.J.; Shan, L.Y.; Gounni, A.S.; Wu, Y.Z.; Zhang, J.B. Neutrophil extracellular trap formation is associated with autophagy-related signalling in ANCA-associated vasculitis. Clin Exp Immunol 2015, 180, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Khandpur, R.; Carmona-Rivera, C.; Vivekanandan-Giri, A.; Gizinski, A.; Yalavarthi, S.; Knight, J.S.; Friday, S.; Li, S.; Patel, R.M.; Subramanian, V.; Thompson, P.; Chen, P.; Fox, D.A.; Pennathur, S.; Kaplan, M.J. NETs are a source of citrullinated autoantigens and stimulate inflammatory responses in rheumatoid arthritis. Sci Transl Med 2013, 5, 178ra40. [Google Scholar] [CrossRef]
- Garcia-Romo, G. S.; Caielli, S.; Vega, B.; Connolly, J.; Allantaz, F.; Xu, Z.; Punaro, M.; Baisch, J.; Guiducci, C.; Coffman, R.L.; Barrat, F.J.; Banchereau, J.; Pascual, V. Netting neutrophils are major inducers of type I IFN production in pediatric systemic lupus erythematosus. Sci Transl Med 2011, 3, 73ra20. [Google Scholar] [CrossRef]
- Papadaki, G.; Choulaki, C.; Mitroulis, I.; Verginis, P.; Repa, A.; Raptopoulou, A.; Boumpas, D.; Sidiropoulos, P. Enhanced release of neutrophil extracellular traps from peripheral blood neutrophils in patients with rheumatoid arthritis. Annals of the Rheumatic Diseases 2012, 71, A79. [Google Scholar] [CrossRef]
- Sur Chowdhury, C.; Giaglis, S.; Walker, U. A.; Buser, A. , Hahn, S.; Hasler, P. Enhanced neutrophil extracellular trap generation in rheumatoid arthritis: analysis of underlying signal transduction pathways and potential diagnostic utility. Arthritis Res Ther 2014, 16, R122. [Google Scholar] [CrossRef]
- Wang, W.; Peng, W.; Ning, X. Increased levels of neutrophil extracellular trap remnants in the serum of patients with rheumatoid arthritis. Int J Rheum Dis 2018, 21, 415–421. [Google Scholar] [CrossRef]
- Willis, V. C.; Gizinski, A. M.; Banda, N. K.; Causey, C. P.; Knuckley, B.; Cordova, K. N.; Luo, Y.; Levitt, B.; Glogowska, M.; Chandra, P.; Kulik, L.; Robinson, W. H.; Arend, W. P.; Thompson, P. R.; Holers, V. M. N-α-benzoyl-N5-(2-chloro-1-iminoethyl)-L-ornithine amide, a protein arginine deiminase inhibitor, reduces the severity of murine collagen-induced arthritis. J Immunol 2011, 186, 4396–4404. [Google Scholar] [CrossRef]
- Chirivi, R. G. S.; Jenniskens, G. J.; Raats, J. M. H, Anti-Citrullinated Protein Antibodies as Novel Therapeutic Drugs in Rheumatoid Arthritis. J Clin Cell Immunol 2013, S6, 006. [Google Scholar]
- Leffler, J.; Martin, M.; Gullstrand, B.; Tydén, H.; Lood, C.; Truedsson, L.; Bengtsson, A. A.; Blom, A. M. Neutrophil extracellular traps that are not degraded in systemic lupus erythematosus activate complement exacerbating the disease. J Immunol 2012, 188, 3522–3531. [Google Scholar] [CrossRef] [PubMed]
- Manderson, A. P.; Carlucci, F.; Lachmann, P. J.; Lazarus, R. A.; Festenstein, R. J.; Cook, H. T.; Walport, M. J.; Botto, M. The in vivo expression of actin/salt-resistant hyperactive DNase I inhibits the development of anti-ssDNA and anti-histone autoantibodies in a murine model of systemic lupus erythematosus. Arthritis Res Ther 2006, 8, R68. [Google Scholar] [CrossRef] [PubMed]
- Knight, J. S.; Subramanian, V.; O'Dell, A. A.; Yalavarthi, S.; Zhao, W.; Smith, C. K.; Hodgin, J. B.; Thompson, P. R.; Kaplan, M. J. Peptidylarginine deiminase inhibition disrupts NET formation and protects against kidney, skin and vascular disease in lupus-prone MRL/lpr mice. Ann Rheum Dis 2015, 74, 2199–2206. [Google Scholar] [CrossRef]
- Smith, H. A.; Kang, Y. The metastasis-promoting roles of tumor-associated immune cells. J Mol Med (Berl) 2013, 91, 411–429. [Google Scholar] [CrossRef]
- Zhao, J.; Jin, J. Neutrophil extracellular traps: New players in cancer research. Front Immunol 2022, 13, 937565. [Google Scholar] [CrossRef]
- Fridlender, Z. G.; Albelda, S. M. Tumor-associated neutrophils: friend or foe? Carcinogenesis 2012, 33, 949–955. [Google Scholar] [CrossRef]
- Piccard, H.; Muschel, R. J.; Opdenakker, G. On the dual roles and polarized phenotypes of neutrophils in tumor development and progression. Crit Rev Oncol Hematol 2012, 82, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, L. The pro-tumor effect and the anti-tumor effect of neutrophils extracellular traps. Biosci Trends 2020, 13, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Berger-Achituv, S.; Brinkmann, V.; Abed, U. A.; Kühn, L. I.; Ben-Ezra, J.; Elhasid, R.; Zychlinsky, A. A proposed role for neutrophil extracellular traps in cancer immunoediting. Front Immunol 2013, 4, 48. [Google Scholar] [CrossRef]
- Chen, CJ.; Wu, C.C.; Chang, C.Y.; Li, J.R.; Ou, Y.C.; Chen, W.Y.; Liao, S.L.; Wang, J.D. Metformin Mitigated Obesity-Driven Cancer Aggressiveness in Tumor-Bearing Mice. Int J Mol Sci 2022, 23, 9134. [Google Scholar] [CrossRef]
- Demers, M.; Krause, D. S.; Schatzberg, D.; Martinod, K.; Voorhees, J. R.; Fuchs, T. A.; Scadden, D. T.; Wagner, D. D. Cancers predispose neutrophils to release extracellular DNA traps that contribute to cancer-associated thrombosis. Proc Natl Acad Sci U S A 2012, 109, 13076–13081. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Franco, M. M.; Leon-Rodriguez, E.; Torres-Ruiz, J. J.; Gómez-Martín, D.; Angles-Cano, E.; de la Luz Sevilla-González, M. Neutrophil Extracellular Traps Associate with Clinical Stages in Breast Cancer. Pathol Oncol Res 2020, 26, 1781–1785. [Google Scholar] [CrossRef] [PubMed]
- Najmeh, S.; Cools-Lartigue, J.; Rayes, R. F.; Gowing, S.; Vourtzoumis, P.; Bourdeau, F.; Giannias, B.; Berube, J.; Rousseau, S.; Ferri, L. E.; Spicer, J. D. Neutrophil extracellular traps sequester circulating tumor cells via β1-integrin mediated interactions. Int J Cancer 2017, 140, 2321–2330. [Google Scholar] [CrossRef]
- Mishalian, I. , et al., Tumor-associated neutrophils (TAN) develop pro-tumorigenic properties during tumor progression. Cancer Immunol Immunother, 2013. 62(11): p. 1745-56.
- Sangaletti, S.; Tripodo, C.; Vitali, C.; Portararo, P.; Guarnotta, C.; Casalini, P.; Cappetti, B.; Miotti, S.; Pinciroli, P.; Fuligni, F.; Fais, F.; Piccaluga, P. P.; Colombo, M. P. Defective stromal remodeling and neutrophil extracellular traps in lymphoid tissues favor the transition from autoimmunity to lymphoma. Cancer Discov 2014, 4, 110–129. [Google Scholar] [CrossRef]
- McDonald, B.; Spicer, J.; Giannais, B.; Fallavollita, L.; Brodt, P.; Ferri, L. E. Systemic inflammation increases cancer cell adhesion to hepatic sinusoids by neutrophil mediated mechanisms. Int J Cancer 2009, 125, 1298–1305. [Google Scholar] [CrossRef]
- Spicer, J. D.; McDonald, B.; Cools-Lartigue, J. J.; Chow, S. C.; Giannias, B.; Kubes, P.; Ferri, L. E. Neutrophils promote liver metastasis via Mac-1-mediated interactions with circulating tumor cells. Cancer Res 2012, 72, 3919–3927. [Google Scholar] [CrossRef]
- Cools-Lartigue, J.; Spicer, J.; Najmeh, S.; Ferri, L. Neutrophil extracellular traps in cancer progression. Cell Mol Life Sci 2014, 71, 4179–4194. [Google Scholar] [CrossRef] [PubMed]
- Houghton, A. M.; Rzymkiewicz, D. M.; Ji, H.; Gregory, A. D.; Egea, E. E.; Metz, H. E.; Stolz, D. B.; Land, S. R.; Marconcini, L. A.; Kliment, C. R.; Jenkins, K. M.; Beaulieu, K. A.; Mouded, M.; Frank, S. J.; Wong, K. K.; Shapiro, S. D. Neutrophil elastase-mediated degradation of IRS-1 accelerates lung tumor growth. Nat Med 2010, 16, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Gregory, A. D.; Hale, P.; Perlmutter, D. H.; Houghton, A. M. Clathrin pit-mediated endocytosis of neutrophil elastase and cathepsin G by cancer cells. J Biol Chem 2012, 287, 35341–35350. [Google Scholar] [CrossRef] [PubMed]
- Deryugina, E.; Carré, A.; Ardi, V.; Muramatsu, T.; Schmidt, J.; Pham, C.; Quigley, J.P. Neutrophil Elastase Facilitates Tumor Cell Intravasation and Early Metastatic Events. iScience 2020, 23, 101799. [Google Scholar] [CrossRef]
- Alaseem, A.; Alhazzani, K.; Dondapati, P.; Alobid, S.; Bishayee, A.; Rathinavelu, A. Matrix Metalloproteinases: A challenging paradigm of cancer management. Semin Cancer Biol 2019, 56, 100–115. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L. M.; Tinkle, C. L.; Hanahan, D.; Werb, Z. MMP-9 supplied by bone marrow-derived cells contributes to skin carcinogenesis. Cell 2000, 103, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, S.; Koran, S.; AlOmair, L. Insights Into the Role of Matrix Metalloproteinases in Cancer and its Various Therapeutic Aspects: A Review. Front Mol Biosci 2022, 9, 896099. [Google Scholar] [CrossRef]
- Wilson, T. J.; Nannuru, K. C.; Futakuchi, M.; Singh, R. K. Cathepsin G-mediated enhanced TGF-beta signaling promotes angiogenesis via upregulation of VEGF and MCP-1. Cancer Lett 2010, 288, 162–169. [Google Scholar] [CrossRef]
- Morimoto-Kamata, R.; Mizoguchi, S.; Ichisugi, T.; Yui, S. Cathepsin G induces cell aggregation of human breast cancer MCF-7 cells via a 2-step mechanism: catalytic site-independent binding to the cell surface and enzymatic activity-dependent induction of the cell aggregation. Mediators Inflamm 2012, 2012, 456462. [Google Scholar] [CrossRef]
- Guan, X.; Lu, Y.; Zhu, H.; Yu, S.; Zhao, W. ; Chi,X. ; Xie, C.; Yin, Z. The Crosstalk Between Cancer Cells and Neutrophils Enhances Hepatocellular Carcinoma Metastasis via Neutrophil Extracellular Traps-Associated Cathepsin G Component: A Potential Therapeutic Target. J Hepatocell Carcinoma 2021, 8, 451–465. [Google Scholar]
- Redman, C.W.; Sargent, I.L. Sargent, Latest advances in understanding preeclampsia. Science 2005. 308, 1592–1594.
- Miranda, M.L.; Macher, H.C.; Muñoz-Hernández, R.; Vallejo-Vaz, A.; Moreno-Luna, R.; Villar, J.; Guerrero, J.M.; Stiefel, P. Role of circulating cell-free DNA levels in patients with severe preeclampsia and HELLP syndrome. Am J Hypertens 2013, 26, 1377–1380. [Google Scholar] [CrossRef]
- Gupta, A.; Hasler, P.; Gebhardt, S.; Holzgreve, W.; Hahn, S. Occurrence of neutrophil extracellular DNA traps (NETs) in pre-eclampsia: a link with elevated levels of cell-free DNA? Ann N Y Acad Sci 2006, 1075, 118–122. [Google Scholar] [CrossRef]
- Hasler, P.; Giaglis, S.; Hahn, S. Neutrophil extracellular traps in health and disease. Swiss Med Wkly 2016, 146, w14352. [Google Scholar] [CrossRef]
- Marder, W.; Knight, J.S.; Kaplan, M.J.; Somers, E.C.; Zhang, X.; O'Dell, A.A.; Padmanabhan, V.; Lieberman, R.W. Placental histology and neutrophil extracellular traps in lupus and pre-eclampsia pregnancies. Lupus Sci Med 2016, 3, e000134. [Google Scholar] [CrossRef]
- Saguil, A.; Fargo, M.; Grogan, S. Diagnosis and management of kawasaki disease. Am Fam Physician 2015, 91, 365–371. [Google Scholar]
- McCrindle, B.W.; Rowley, A.H.; Newburger, J.W.; Burns, J.C.; Bolger, A.F.; Gewitz, M.; Baker, A.L.; Jackson, M.A.; Takahashi, M.; Shah, P.B.; Kobayashi, T.; Wu, M.H.; Saji, T.T.; Pahl, E. Diagnosis, Treatment, and Long-Term Management of Kawasaki Disease: A Scientific Statement for Health Professionals From the American Heart Association. Circulation 2017, 135, e927–e999. [Google Scholar] [CrossRef]
- Tian, J.; Lv, H.T.; An, X.J.; Ling, N.; Xu, F. Endothelial microparticles induce vascular endothelial cell injury in children with Kawasaki disease. Eur Rev Med Pharmacol Sci 2016, 20, 1814–1818. [Google Scholar]
- Newburger, J.W.; Takahashi, M.; Burns, J.C. ; Kawasaki Disease. J Am Coll Cardiol 2016, 67, 1738–1749. [Google Scholar] [CrossRef]
- Richter, J.R.; Sanderson, R.D. The glycocalyx: pathobiology and repair. Matrix Biol Plus 2023, 17, 100128. [Google Scholar] [CrossRef]
- Ohnishi, Y.; Yasudo, H.; Suzuki, Y.; Furuta, T.; Matsuguma, C.; Azuma, Y.; Miyake, A.; Okada, S.; Ichihara, K.; Ohga, S.; Hasegawa, S. Circulating endothelial glycocalyx components as a predictive marker of coronary artery lesions in Kawasaki disease. Int J Cardiol 2019, 292, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Zhang, Y.; Yang, X.; Yan, C.; Feng, Y. Recent Insights into Neutrophil Extracellular Traps in Cardiovascular Diseases. J Clin Med 2022, 11, 6662. [Google Scholar] [CrossRef]
- Hu, J.; Qian, W.; Yu, Z.; Xu, T.; Ju, L.; Hua, Q.; Wang, Y.; Ling, J.J.; Lv, H. Increased Neutrophil Respiratory Burst Predicts the Risk of Coronary Artery Lesion in Kawasaki Disease. Front Pediatr 2020, 8, 391. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Takeshita, S.; Kawamura, Y.; Kanai, T.; Tsujita, Y.; Nonoyama, S. Enhanced formation of neutrophil extracellular traps in Kawasaki disease. Pediatr Res 2020, 87, 998–1004. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Ding, M.; Fu, J.; Xiao, Y.; Chen, X.; Zhang, Q. Neutrophil extracellular trap from Kawasaki disease alter the biologic responses of PBMC. Biosci Rep 2020, 40, BSR20200928. [Google Scholar] [CrossRef] [PubMed]
- Zandstra, J.; van de Geer, A.; Tanck, M.W.T.; van Stijn-Bringas Dimitriades, D.; Aarts, C.E.M.; Dietz, S.M.; van Bruggen, R.; Schweintzger, N.A.; Zenz, W.; Emonts, M.; Zavadska, D.; Pokorn, M.; Usuf, E.; Moll, H.A.; Schlapbach, L.J.; Carrol, E.D.; Paulus, S.; Tsolia, M.; Fink, C.; Yeung, S.; Shimizu, C.; Tremoulet, A.; Galassini, R.; Wright, V.J.; Martinón-Torres, F.; Herberg, J.; Burns, J.; Levin, M.; Kuijpers, T.W.; EUCLIDS Consortium, PERFORM Consortium and UK Kawasaki Disease Genetics Study Network. Biomarkers for the Discrimination of Acute Kawasaki Disease From Infections in Childhood. Front Pediatr 2020, 22, 355. [Google Scholar] [CrossRef]
- Martinod, K.; Fuchs, T.A.; Zitomersky, N.L.; Wong, S.L.; Demers, M.; Gallant, M.; Wang, Y.; Wagner, D.D. PAD4-deficiency does not affect bacteremia in polymicrobial sepsis and ameliorates endotoxemic shock. Blood 2015, 125, 1948–1956. [Google Scholar] [CrossRef]
- Sorvillo, N.; Cherpokova, D.; Martinod, K.; Wagner, D.D. Extracellular DNA NET-Works With Dire Consequences for Health. Circ Res 2019, 125, 470–488. [Google Scholar] [CrossRef]
- Martinod, K.; Wagner, D.D. Thrombosis: tangled up in NETs. Blood 2014, 123, 2768–2776. [Google Scholar] [CrossRef]
- Savchenko, A.S.; Borissoff, J.I.; Martinod, K.; De Meyer, S.F.; Gallant, M.; Erpenbeck, L.; Brill, A.; Wang, Y.; Wagner, D.D. VWF-mediated leukocyte recruitment with chromatin decondensation by PAD4 increases myocardial ischemia/reperfusion injury in mice. Blood 2014, 123, 141–148. [Google Scholar] [CrossRef]
- Hakkim, A.; Fürnrohr, B.G.; Amann, K.; Laube, B.; Abed, U.A.; Brinkmann, V.; Herrmann, M.; Voll, R.E.; Zychlinsky, A. Impairment of neutrophil extracellular trap degradation is associated with lupus nephritis. Proc Natl Acad Sci U S A 2010, 107, 9813–9818. [Google Scholar] [CrossRef]
- Wong, S.L.; Wagner, D.D. Wagner, Peptidylarginine deiminase 4: a nuclear button triggering neutrophil extracellular traps in inflammatory diseases and aging. FASEB J 2018, 32, fj201800691R. [Google Scholar] [CrossRef]
- Patutina, O.; Mironova, N.; Ryabchikova, E.; Popova, N.; Nikolin, V.; Kaledin, V.; Vlassov, V.; Zenkova, M. Inhibition of metastasis development by daily administration of ultralow doses of RNase A and DNase I. Biochimie 2011, 93, 689–696. [Google Scholar] [CrossRef] [PubMed]
- De Meyer, S.F.; Suidan, G.L.; Fuchs, T.A.; Monestier, M.; Wagner, D.D. Extracellular chromatin is an important mediator of ischemic stroke in mice. Arterioscler Thromb Vasc Biol 2012, 32, 1884–1891. [Google Scholar] [CrossRef] [PubMed]
- Cools-Lartigue, J.; Spicer, J.; McDonald, B.; Gowing, S.; Chow, S.; Giannias, B.; Bourdeau, F.; Kubes, P.; Ferri, L. Neutrophil extracellular traps sequester circulating tumor cells and promote metastasis. J Clin Invest 2013, 123, 3446–3458. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Controversial role of NETs. NET is a powerful weapon of innate immunity and provides protection to the host through clearing the invading pathogens (right part). On the contrary aberrant NET formation causes pathology in numerous ways and attributed to autoimmune, infectious, and non-infectious diseases (left part).
Figure 1.
Controversial role of NETs. NET is a powerful weapon of innate immunity and provides protection to the host through clearing the invading pathogens (right part). On the contrary aberrant NET formation causes pathology in numerous ways and attributed to autoimmune, infectious, and non-infectious diseases (left part).
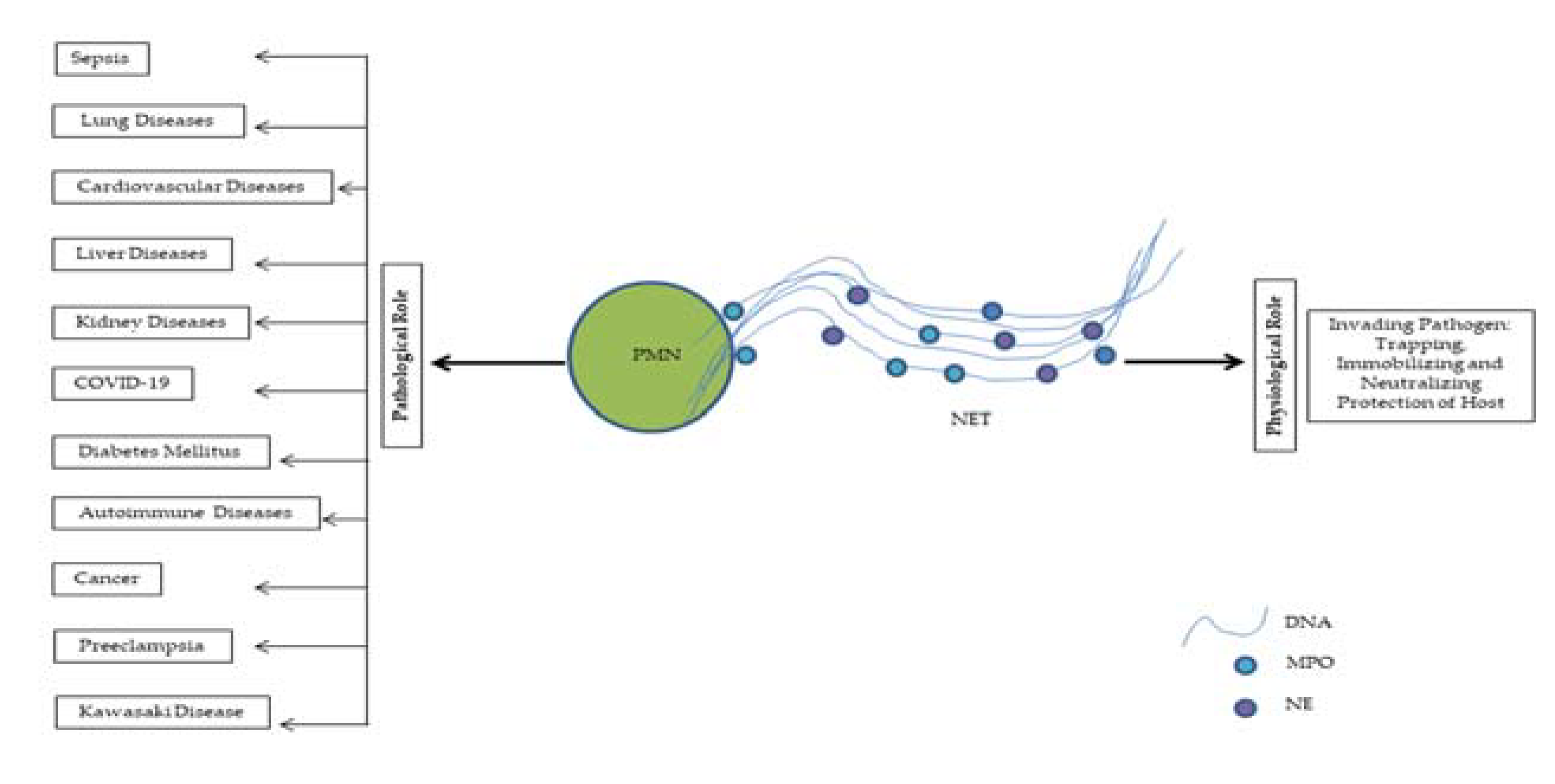

Table 1.
Inhibition of NETs as therapeutic approach
| References | Year | Results |
|---|---|---|
| Martinod et al [230] | 2014 | NET inhibition by DNase I administration showed protective role against thrombosis in vivo murine model of ischemic stroke, myocardial infarction and DVT. |
| Brill et al [163] | 2012 | Infusion of DNase I protected mice from DVT through inhibition of NETs. |
| De Meyer et al [235] |
2012 | NET inhibition by DNase I improves ischemic stroke outcome in mice model. |
| Savchenko et al [231] |
2014 | DNase I therapy against NETs in mice model exerts cardioprotective effects and improve cardiac contractile function. |
| Papayannopoulos et al [107] | 2011 | DNase I therapy enhances sputum solubilization in CF patients |
| Wong et al[233] | 2018 | Elevated NETs contribute to diabetes complications and NET disruption by using DNase I therapy improves wound healing. |
| Patutina et al [234] | 2011 | DNase I treatment inhibits tumor progression and dissemination. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



