
Submitted:
26 September 2023
Posted:
28 September 2023
You are already at the latest version
Abstract
Antimicrobial peptides (AMPs) have emerged as a promising class of bioactive molecules with the potential to combat infections associated with medical implants and biomaterials. This review article aims to provide a comprehensive analysis of the role of antimicrobial peptides in medical implants and biomaterials, along with their diverse clinical applications. The incorporation of AMPs into various medical implants and biomaterials has shown immense potential in mitigating biofilm formation and preventing implant-related infections. We review the latest advancements in biomedical sciences and discuss the AMPs that were immobilized successfully to enhance their efficacy and stability within the implant environment. We also highlight successful examples of AMP coatings for the treatment of surgical site infections (SSIs), Contact lenses, Dental Applications, AMP-Incorporated Bone Grafts, Urinary tract infections (UTIs), Medical Implants, etc. Additionally, we discuss the potential challenges and prospects of AMPs in medical implants, such as effectiveness, instability and implant-related complications. We also discuss strategies that can be employed to overcome the limitations of AMP-coated biomaterials for prolonged longevity in clinical settings.
Keywords:
Antimicrobial Peptides
; Bio-films
; Biomedical Applications
; Sepsis
1. Introduction
Antimicrobial resistance (AMR) poses a significant global threat to public health, as it renders conventional antibiotics ineffective against bacterial infections [1]. The overuse and misuse of antibiotics have accelerated the emergence of drug-resistant strains, leading to increased morbidity, mortality, and healthcare costs. In the search for alternative therapeutic options, antimicrobial peptides (AMPs) have emerged as promising candidates due to their unique properties and ability to overcome cellular resistance mechanisms [2]. This resistance can be intrinsic, acquired through genetic mutations or horizontal gene transfer, and can spread rapidly within and between species [3]. The misuse of antibiotics in clinical and agricultural settings, inadequate infection control measures, and poor surveillance contribute to the proliferation of resistant strains [4]. The consequences of AMR are far-reaching and affect both individual patients and global healthcare systems [5]. Infections caused by drug-resistant pathogens are associated with higher rates of treatment failure, prolonged illness, and increased mortality. Additionally, the economic burden of AMR is substantial, with increased healthcare costs, extended hospital stays, and a reduction in productivity. If left unaddressed, AMR could potentially reverse the progress made in modern medicine, leading to a future where common infections become life-threatening once again. Another challenge is that there is an intricate relationship between AMR and some life-threatening diseases such as sepsis, highlighting the implications of AMR in sepsis management and the urgent need for comprehensive strategies to combat this dual crisis. Resistant bacteria, such as methicillin-resistant Staphylococcus aureus (MRSA) or extended-spectrum beta-lactamase (ESBL)-producing Enterobacteriaceae, are common culprits in sepsis cases [6].
Sepsis is a critical condition that occurs as a result of an uncontrolled systemic inflammatory response to an infection. Lipopolysaccharide (LPS), also known as endotoxin, initially binds to the LPS binding protein (LBP). LPS-LBP complex then binds to Toll-like receptor 4 (TLR4) and CD14, initiating a series of signaling events, leading to the activation of transcription factors, including nuclear factor-kappa B (NF-κB) [7]. Activation of these transcription factors results in the production and release of pro-inflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α), interleukin-1 beta (IL-1β), and interleukin-6 (IL-6)[8,9,10]. (Figure 1A).
AMPs hold promise as a potential treatment for sepsis as they not only possess direct antimicrobial activity but also exhibit anti-cancer and immunomodulatory properties[11]. They can modulate the immune response by enhancing the function of immune cells, promoting the clearance of pathogens, and regulating the release of pro-inflammatory cytokines. Many AMPs directly bind to LPS and inhibit the downstream signaling [12] (Figure 1B). Interestingly, numerous antimicrobial peptides also interact with cancer cell membranes in a manner akin to their bacterial membrane targeting, owing to the anionic properties of cancer cell membranes[13,14](Figure 1C).
1.1. How AMPs Can Overcome Antimicrobial Resistance?
AMPs exhibit broad-spectrum activity, meaning they can target a wide range of microorganisms including bacteria, fungi, viruses, and drug-resistant strains. Humans are in continuous contact with microorganisms in a microbial-containing environment. At the time of birth, skin and mucous layers are first exposed to microorganisms and these microbes constitute the normal flora[15]. Our body hosts more bacterial cells than human cells [16]. These bacteria play a vital role in continuing the life of humans. However, the bacteria from the outside are mostly pathogenic and breach the defenses of the host and cause various infections which sometimes lead to sepsis[17].
Unlike traditional antibiotics that often focus on specific bacterial species, AMPs offer a broader action, making it challenging for microorganisms to develop resistance. AMPs employ multiple modes of action to kill or inhibit the growth of microorganisms. They can disrupt the microbial cell membrane, leading to cell lysis, and interfere with intracellular processes like protein synthesis and DNA replication. This multifaceted approach makes it difficult for microorganisms to develop resistance since they would need to overcome several barriers simultaneously. One notable advantage of AMPs is their ability to rapidly kill bacteria. This fast bactericidal activity effectively eliminates bacteria. Moreover, studies have shown that the development of resistance to AMPs is relatively low compared to traditional antibiotics, further emphasizing their potential in combating antimicrobial resistance.
AMPs can also synergize with existing antibiotics when used in combination[18]. By disrupting bacterial cell membranes, AMPs can enhance the penetration of antibiotics, allowing them to reach their targets more effectively. This synergistic effect improves the efficacy of antibiotics and reduces the likelihood of resistance development. [19]. Moreover, AMPs target unique sites in microorganisms. For instance, they can bind to and disrupt bacterial biofilms, which are protective structures contributing to antibiotic resistance[20]. By targeting bacterial biofilms, AMPs provide an alternative approach to combating resistant microorganisms.
1.2. Discovery of AMPs
The discovery of AMPs has been a fascinating journey spanning several decades. It began with early observations of antimicrobial properties in natural substances used by ancient civilizations. In the 1980s, researchers stumbled upon a class of cationic peptides called defensins in rabbit leukocytes, marking the first identification of AMPs[21,22]. One of the earliest defensins to be discovered was human neutrophil peptide 1 (HNP-1), also known as alpha-defensin 1, was identified in 1985 from human neutrophils, [23]. Shortly after the discovery of HNP-1, several other defensins were identified in different organisms. For example, plant defensins were discovered in the late 1980s, and insect defensins were discovered in the early 1990s. Scientists then embarked on systematic screening efforts, exploring various sources including humans, animals, plants, and microorganisms to identify new AMPs. Advances in molecular biology techniques, such as cloning and DNA sequencing, allowed for the characterization of genes encoding AMPs and the production of large quantities of peptides for further study. Drosocin and Melittin are examples of peptides that were identified using the above-mentioned approaches[24]. Drosocin was identified when researchers studying Drosophila melanogaster noticed that the flies had a robust immune response to bacterial infections [25]. They then extracted peptides from fly samples and later purified them using chromatography and mass spectrometry. The purified peptides were then subjected to further characterization to determine their chemical structure, activity, and mode of action [26,27]. A similar approach was followed by Melittin as well[28]. After the confirmation of the antimicrobial and cytolytic activity in the bioassays, the active fraction was further purified using techniques such as high-performance liquid chromatography (HPLC) and mass spectrometry. The purified melittin was tested again for its antimicrobial, cytolytic, or other activities to confirm that it matched the initial observations and demonstrated the characteristic effects associated with melittin[29,30].
Bioinformatics, genomics and computational tools are also pivotal to predict and analyze peptide sequences with antimicrobial potential. Moreover, natural product peptide libraries and high-throughput screening methods have contributed to the discovery of novel AMPs as well. The latest in silico approaches to discovering AMPs include several computational methods that play a crucial role in the discovery of AMPs[31]. Researchers use various software tools and algorithms to predict and screen for potential antimicrobial peptides based on their sequence, structure, physicochemical properties, and known AMP databases. Examples of such tools include CAMPR3, AntiBP2, and CAMP2.0[32,33]. Machine learning and artificial intelligence are other advanced tools to discover novel antimicrobial peptides. In advanced machine learning techniques, deep learning and artificial neural networks, are being increasingly employed to identify new AMPs[34]. These models are trained on large datasets of known AMPs and non-AMPs to recognize patterns and make predictions. Screening large libraries of peptides or peptide fragments is another strategy for identifying potential AMPs. Techniques like peptide arrays, phage display, and combinatorial chemistry are also commonly employed for this purpose. Other approaches include DNA sequencing technologies and structural biology approaches. In the former approach, the genetic information is analyzed and by utilizing that knowledge, the researchers identify potential AMP-encoding genes and subsequently validate the antimicrobial activity of the corresponding peptides[35].In structural biology approaches, the three-dimensional structure of AMPs can provide valuable insights into their mechanism of action and help optimize their activity. Techniques such as X-ray crystallography, nuclear magnetic resonance (NMR) spectroscopy, and cryo-electron microscopy (cryo-EM) are used to elucidate the structures of AMPs and their interactions with microbial targets[36]. Antimicrobial and other bioactive peptides have also been derived from naturally occurring proteins and have been reported to possess both immunomodulatory and anti-cancer properties [37,38,39]. Table 1 presents a comprehensive overview of all the discussed AMPs in this review. Various factors, such as net charge and hydrophobicity, govern the activity of AMPs [40]. The antimicrobial peptide database reveals that the majority of AMPs possess distinct levels of cationicity and hydrophobicity (Figure 2).
1.3. Harnessing Antimicrobial Peptides for Advanced Biomaterials
1.3.1. Next-Level Surgical Innovation: Antimicrobial Peptide-Enhanced Sutures
Surgical site infections (SSIs) pose a significant challenge in modern healthcare, leading to increased patient morbidity, extended hospital stays, and substantial healthcare costs[41]. Antimicrobial peptides (AMPs) have emerged as a potential solution to combat SSIs due to their broad-spectrum antimicrobial activity and unique mechanisms of action. In recent years, researchers have explored the incorporation of AMPs into surgical sutures to create antimicrobial peptide-impregnated sutures (AMPIS). The incorporation of AMPs into sutures offers several advantages in the prevention of SSIs: Firstly, AMPs used in sutures are originally selected for their minimal cytotoxicity and immunogenicity, ensuring biocompatibility and compatibility with wound healing processes [42]. Besides, AMPs due to their broad-spectrum activity on drug-resistant strains, make them suitable for combating polymicrobial infections. Besides this, AMPs can disrupt biofilm formation, which gets embedded within a protective matrix[43]. By preventing biofilm formation, AMPIS can impede bacterial adhesion and subsequent colonization. This also reduces the infection rates. Studies have demonstrated that the use of (AMPIS), can significantly reduce the incidence of SSIs compared to traditional sutures.
LL-37 is a naturally occurring human cathelicidin that possesses broad-spectrum antimicrobial activity against bacteria, fungi, and viruses. It also exhibits immunomodulatory properties and promotes wound healing. Its incorporation into sutures can prevent surgical site infections [44]. Several members of the β-defensin family, such as HBD-2 and HBD-3, have been utilized in antimicrobial peptide-impregnated sutures. Other examples include the incorporation of Protegrin-1 and Histatins. While the former enhances the antimicrobial efficacy and helps prevent postoperative infections, the latter exhibits activity against various oral pathogens, including Candida albicans, Streptococcus mutans, and Porphyromonas gingivalis. The histatin-impregnated sutures are particularly useful in oral and maxillofacial surgeries [45,46]. Ranalexin is another antimicrobial peptide that was originally isolated from the skin secretions of the northern leopard frog. It possesses antimicrobial activity against various bacteria, including antibiotic-resistant strains. Ranalexin-based coatings have been investigated for their ability to inhibit wound and systemic methicillin-resistant Staphylococcus aureus (MRSA) infections [47].
Pexiganan is another example of an AMP that can be utilized for the incorporation of AMPs into sutures. It is a synthetic analog of the AMP magainin 2 and displays potent antimicrobial activity against both Gram-positive and Gram-negative bacteria, including methicillin-resistant Staphylococcus aureus (MRSA). Pexiganan-infused collagen matrices have been shown to facilitate wound healing in rat models of Infection [48]. Similarly, HNP-1-impregnated sutures have shown efficacy in preventing surgical site infections in various surgical procedures [49].
1.3.2. Antimicrobial Peptide-Based Contact Lenses: The Future of Eye Care
Contact lenses have revolutionized vision correction, providing a convenient and comfortable alternative to traditional eyeglasses. However, despite their numerous advantages, contact lenses pose a risk of infection due to microbial colonization on their surfaces[50]. This concern has led to significant efforts in developing novel contact lens materials with built-in antimicrobial properties.
Contact lens-associated microbial keratitis (CLMK) is a serious condition that can lead to vision loss if left untreated[51]. The risk of CLMK arises from bacterial adhesion, biofilm formation, and subsequent infection on the lens surface. Traditional contact lenses, although effective in vision correction, lack inherent antimicrobial properties, making them susceptible to microbial colonization. Therefore, there is a pressing need for contact lenses that actively combat microbial growth to minimize the risk of infections. Researchers have focused on developing strategies to immobilize AMPs onto contact lens surfaces, ensuring sustained release and prolonged antimicrobial efficacy. One such approach involves the covalent attachment of AMPs to the lens material, allowing for controlled release over time. Another method utilizes hydrogels with AMPs encapsulated within, enabling a slow release of the peptide. These approaches not only provide antimicrobial properties but also maintain the biocompatibility and optical properties required for comfortable vision correction (Figure 3).
By inhibiting biofilm formation, AMPs prevent the accumulation of pathogens, ensuring a healthier lens surface. Furthermore, AMPs can combat multidrug-resistant microorganisms, which are becoming increasingly prevalent in healthcare settings.
Further advancements in AMP design, formulation, and delivery systems will undoubtedly enhance their antimicrobial efficacy and biocompatibility. Additionally, the integration of AMPs with other innovative technologies, such as smart materials or drug-delivery systems, may offer new possibilities for multifunctional contact lenses. As we move forward, extensive research, preclinical and clinical trials, and collaborations between scientists, engineers, and ophthalmologists are necessary to bring antimicrobial peptide-based contact lenses from the lab to the market. While challenges remain, ongoing research and technological advancements hold great promise for the development of safe and effective antimicrobial peptide-based contact lenses, benefiting countless individuals who rely on contact lenses for vision correction. α-MSH: α-Melanocyte-stimulating hormone (α-MSH) is a naturally occurring peptide with immunomodulatory and antimicrobial properties. It has been incorporated into contact lenses to provide antimicrobial activity and promote ocular surface healing[52]. Another example of successful use of an AMPs in ocular health are Temporins. Temporins are originally a group of antimicrobial peptides derived from the skin of frogs. Various temporin peptides have been investigated for their antimicrobial activity and have been incorporated into contact lenses to inhibit the growth of bacteria and fungi. Besides naturally occurring AMPs, there are certain synthetic peptides that were designed and synthesized and have been used for AMP based contact lenses. A synthetic peptide, melimine was produced by combining portions of the antimicrobial cationic peptides mellitin and protamine has been studied for its potential in promoting wound healing and preventing infections when incorporated into contact lenses [53]. Pexiganan, has been utilized in antimicrobial peptide-based contact lenses to enhance their antibacterial properties and prevent microbial colonization [54]. Apart from this,some cyclic peptide antibiotics such as Bacitracin have shown promise in their usage as antimicrobial formulations [55]. It has been used in contact lens solutions, due to its efficacy against Gram-positive bacteria. On similar lines,Dermcidin ,an AMP found in human sweat has been explored for its potential in contact lens applications to prevent microbial contamination and infection [56].
1.3.3. Antimicrobial Peptide-Conjugated Nanoparticles for Dental Applications: A Promising Approach for Combatting Oral Infections
Dentistry, as a branch of medicine, is no exception to antimicrobial resistance challenges [57]. Oral infections, such as dental caries and periodontal diseases, pose significant challenges to dental health worldwide. Conventional treatment methods often fall short in effectively targeting and eliminating microbial pathogens associated with these infections. In recent years, the development of antimicrobial peptide (AMP)-conjugated nanoparticles has emerged as a promising strategy for enhancing the antimicrobial efficacy and delivery of therapeutic agents in the field of dentistry[58] The nanoparticle carrier enhances the antimicrobial efficacy of AMPs by increasing their local concentration at the site of infection, allowing for sustained release and reducing the required therapeutic dose. The nanoparticles can also facilitate the internalization of AMPs into microbial cells, leading to increased disruption of microbial membranes and inhibition of intracellular processes (Figure 4A and B). LL-37 has been investigated for its antimicrobial activity against oral pathogens and its potential application in dental materials, including composites[58]. Dermaseptins are a group of antimicrobial peptides found in the skin secretions of amphibians. They possess potent antimicrobial activity against a broad spectrum of microorganisms and have been utilized in dental applications as peptide-conjugated nanoparticles and fatty acids[59]. Nisin is a naturally occurring antimicrobial peptide produced by certain strains of Lactococcus lactis. It has potent antimicrobial activity against Gram-positive bacteria, including oral pathogens. Nisin has been conjugated to nanoparticles for dental applications to enhance their antimicrobial efficacy[60]. Indolicidin-coated silver nanoparticles have shown potent antibacterial activities in oral diseases[61]. The defensins, viz. hBD-1,3,5 exert antibacterial activity against microbes involved in root canal infections, including Enterococcus faecalis, Fusobacterium. nucleatum, Tannerella forsythia, Eikenella corrodens and C. albicans[62,63,64]. Human β-defensin 3 (HBD3) peptide exhibits more antibacterial activity against mature multispecies biofilms containing A. naeslundii, L. salivarius, and E. faecalis, than either calcium hydroxide or 2% chlorhexidine solution [64].
Cateslytin is another AMP that is derived from the venom of the Brazilian scorpion Tityus catesbeianus. It exhibits antimicrobial activity against oral pathogens and can be used in dental applications as peptide-conjugated nanoparticles for its antibacterial effects [65]. GH-12 peptide polymers have been studied for the treatment of secondary caries and the enhanced durability of dental composite restorations[66]. Myxinidin is an antimicrobial peptide isolated from the mucus of hagfish. It exhibits potent antimicrobial activity against various bacteria and fungi including oral pathogens like Pseudomonas aeruginosa (Gram-negative bacteria) and Candida albicans[67]. Chen et al demonstrated a method to construct antimicrobial titanium implants of HHC36 peptide that displaced remarkable antibacterial activity against both Staphylococcus aureus and Escherichia coli only after 2.5 hours of incubation [68].
1.3.4. Antimicrobial Peptide-Incorporated Bone Grafts: Revolutionizing Orthopedic Treatment
In the field of orthopedic surgery, bone grafts play a pivotal role in promoting bone regeneration and restoring skeletal integrity. However, the risk of postoperative infections remains a significant concern, leading to prolonged hospital stays, increased healthcare costs, and potential complications. To address this challenge, researchers have turned to antimicrobial peptides (AMPs) as a promising solution. Bone grafts are widely used in orthopedic surgeries to repair fractures, promote bone healing, and reconstruct skeletal defects. They can be classified into autografts (taken from the patient's own body), allografts (harvested from a donor of the same species), and synthetic grafts. While these grafts provide structural support and scaffolding for new bone formation, they are not inherently antimicrobial. Infection is a major concern following orthopedic surgeries, particularly in the context of bone grafts. Bacterial contamination can lead to implant-associated infections, which are challenging to treat due to the formation of biofilms and limited antibiotic penetration. Traditional strategies such as prophylactic antibiotic use have shown limitations, including antibiotic resistance and side effects. Therefore, there is a need for innovative approaches to combat infection. To harness the antimicrobial properties of AMPs, researchers have explored their incorporation into bone grafts. This involves either directly immobilizing AMPs onto graft surfaces or incorporating them into the graft matrix during the manufacturing process (Figure 4C and D). The goal is to create a localized, sustained release of AMPs to prevent infection while promoting bone healing. Incorporating AMPs into bone grafts offers several advantages. Firstly, it provides an immediate defense against potential pathogens at the surgical site, reducing the risk of infection. Secondly, AMPs have been shown to exhibit synergistic effects with conventional antibiotics, enhancing their efficacy and potentially reducing the required dosage. Thirdly, AMPs can aid in the promotion of bone regeneration by modulating inflammatory responses and enhancing angiogenesis. Several studies have demonstrated the efficacy of AMP-incorporated bone grafts in preclinical models. For example, one study showed that AMP-coated titanium implants reduced bacterial colonization and enhanced bone healing in a rat model [69]. As host cells and bacterial cells vie for control over the implant surface, the introduction of antimicrobial peptides onto the implant surfaces can tip the scales, thwarting implant infections. Yucesoy et al. pioneered a groundbreaking chimeric peptide designed to enhance the functionality of implant materials. This innovative peptide boasts a dual-purpose design, with one segment binding to the surface of a titanium alloy implant through a titanium-binding domain, while the other segment exhibits potent antimicrobial properties[70].
KLD peptides have also been studied for their bone-healing properties. It has been shown that the addition of arginine at the N-terminus converts them into an antimicrobial peptide[71]. Magainin-derived peptides such as E14LKK have also been investigated for their potential use in total-joint replacement prostheses[72]. Further research is needed to optimize AMP selection, develop reliable delivery systems, and evaluate long-term outcomes. Additionally, exploring the potential of AMPs in combination with other bioactive agents, such as growth factors or stem cells, could enhance bone regeneration and infection prevention synergistically.
1.3.5. Antimicrobial Peptide-Based Scaffolds: Enhancing Tissue Regeneration with Antimicrobial Properties
Scaffolds play a crucial role in tissue engineering and regenerative medicine by providing a supportive framework for tissue regeneration. In recent years, the integration of antimicrobial peptides (AMPs) into scaffolds has emerged as a promising approach to enhance the antimicrobial properties of these constructs. By incorporating AMPs into scaffolds, researchers aim to prevent infections that may hinder successful tissue regeneration. The scaffolds serve as three-dimensional templates for cell attachment, proliferation, and differentiation in tissue engineering. They mimic the extracellular matrix (ECM) and provide structural support to regenerate damaged tissues. However, scaffolds alone are vulnerable to microbial colonization, which can lead to infections and impede the healing process. The incorporation of AMPs into scaffolds offers a synergistic approach by combining the regenerative capabilities of the scaffold with the antimicrobial properties of AMPs. Covalent attachment, physical adsorption, electrostatic interactions, and peptide amphiphiles are among the techniques utilized to ensure the retention and sustained release of AMPs from the scaffold structure. These strategies aim to maintain an effective concentration of AMPs at the scaffold-tissue interface, preventing microbial colonization without compromising cell viability or function. Antimicrobial peptide-based scaffolds offer several advantages over traditional scaffolds in tissue engineering. Firstly, they provide a controlled release of AMPs, ensuring a localized antimicrobial effect while minimizing systemic exposure. Secondly, AMPs can be tailored to exhibit selective antimicrobial activity, targeting specific pathogens without disrupting the commensal microbiota. Additionally, the incorporation of AMPs into scaffolds can facilitate faster wound healing, reduce the risk of post-operative infections, and enhance the success of tissue regeneration procedures. In skin tissue engineering, they have been employed to combat wound infections and promote re-epithelialization. Also, AMP-based scaffolds show potential in addressing chronic wounds, where infections can severely impede healing progress. Dermaseptin-S4 (S4) and its analogues have been studied for their potent antimicrobial activity and have been investigated as scaffolds for the development of novel AMP-based scaffolds[73]. Similarly thanatin has been used as a scaffold for the development of peptide-based scaffolds with enhanced antimicrobial activity and stability[74]. Ib-AMPs (Insect-derived antimicrobial peptides) are a diverse group of AMPs found in insects. Various ib-AMPs have been studied and used as scaffolds for the development of antimicrobial peptide-based scaffolds. LL-37 has also been used as a scaffold to design novel AMP-based scaffolds with enhanced antimicrobial activity and tissue injury healing [75]. LLKKK18, an engineered variant of the LL-37 peptide, designed to enhance its antimicrobial potency. It has been utilized in wound healing scaffolds for its broad-spectrum antimicrobial activity [76].Similarly DPK-060, a synthetic antimicrobial peptide has been incorporated into wound healing scaffolds [77,78].Another example of AMPs that are used in scaffolds include SMAP-29 (Sheep myeloid antimicrobial peptide 29).SMAP-29 due to its strong antimicrobial activity against bacteria and fungi can be used in wound healing scaffolds[78]. Abdel-Sayed et al. revealed the capacity of polycationic dendrimers to exert anti-angiogenic effects on burn wounds. They synthesized peptide dendrimers with three generations using amino acid residues L-lysine (G3KL) and L-arginine (G3R) distributed within the branches, resulting in two antimicrobial polycationic dendrimers (AMPDs). These AMPDs, specifically G3KL and G3R, were safely employed in combination with biological bandages comprising progenitor skin cells. This innovative approach effectively thwarted Pseudomonas aeruginosa infections and significantly enhanced wound healing in keratinocytes and endothelial cells. [79].MSI-78, a Magainin 2 analog also possesses antimicrobial activity and has been used in wound healing scaffolds for its antimicrobial properties [80]. Wang and colleagues explored the formation of nanoparticles using MSI-78 and methoxy poly(ethylene glycol)-b-poly(α-glutamic acid). Their findings indicated a reduction in the hemolytic activity of the peptide on human red blood cells within the nanoparticle structure, while the peptide's antibacterial effectiveness remained intact.[81]. With continued advancements in this field, antimicrobial peptide-based scaffolds have the potential to revolutionize the field of tissue engineering, facilitating the development of safer and more effective therapeutic strategies for tissue regeneration.
1.3.6. Antimicrobial Peptide-Coated Urinary Catheters: An Approach to Prevent Catheter-Associated Infections
Urinary tract infections (UTIs) are a prevalent healthcare-associated infection, and catheter-associated urinary tract infections (CAUTIs) contribute significantly to their occurrence[82]. UTIs are responsible for substantial morbidity, increased healthcare costs, and prolonged hospital stays. The emergence of antimicrobial resistance has limited the effectiveness of conventional antibiotics, necessitating the exploration of alternative strategies. CAUTIs are primarily caused by the introduction of bacteria into the urinary tract through the insertion of a catheter [83]. The longer the duration of catheterization, the greater the risk of infection. Several techniques have been developed to coat urinary catheters with AMPs, including physical adsorption, covalent binding, and incorporation into polymer coatings (Figure 5A and B). The choice of coating technique depends on factors such as stability, durability, and release kinetics of AMPs. Li et al. demonstrated that arginine/lysine/tryptophan-rich antimicrobial peptides possess broad-spectrum antimicrobial properties and salt-tolerant characteristics on silicone surfaces, addressing the issue of catheter-associated urinary tract infections (CAUTIs)[84]. Through a process confirmed by X-ray photoelectron spectroscopy and water contact angle analyses, the researchers immobilized these peptides onto polydimethylsiloxane and urinary catheter surfaces using an allyl glycidyl ether (AGE) polymer brush interlayer. The resulting peptide-coated silicone surfaces exhibited remarkable antimicrobial efficacy against bacteria and fungi present in urine and phosphate-buffered saline solution. This effect was enhanced by the synergistic actions of the AGE polymer brush and AMPs, which not only prevented biofilm formation but also repelled cell adhesion. Importantly, the peptide-coated surface demonstrated no toxicity towards smooth muscle cells.[84]. Bac2A is a synthetic antimicrobial peptide derived from the cathelicidin family. It has been evaluated for its antimicrobial efficacy and potential use in preventing biofilm formation on urinary catheters [85,86]. Monteiro et al studied the peptide Chain201D (KWIVWRWRFKR) where they bound the peptide to ((1-mercapto-11-undecyl) -(tetra(ethylene glycol) (EG4)) terminated self-assembled monolayers (SAMs), (EG4-SAMs), activated by 1,1′-Carbonyldiimidazole (CDI) at different concentrations. The study showed the potential of utilizing Chain201D peptide for the development of antimicrobial urinary catheters[87].
Yu et al showed the potential of E6 (RRWRIVVIRVRRC) peptide in treating CAUTIs.The E6 peptide, featuring a cysteine label at its C-terminus, was harnessed to coat polyurethane (PU) surfaces, revealing expansive antimicrobial capabilities[88]. This approach effectively thwarted catheter-associated infections in a mouse urinary infection model. On similar lines, Yao et al. explored an AMP (RWRWRWC–NH2) and incorporated a thiol group into the cysteine residue, integrating it into a Cu2+-coordinated polydopamine coating on PU ureteral stents. The stents featuring the AMP coating demonstrated remarkable suppression of bacterial growth and biofilm formation, all while exhibiting negligible toxicity [89]. A synthetic peptide CD4-PP, constructed through dimerization and backbone cyclization of the antimicrobial region from human cathelicidin LL-37, effectively inhibited uropathogens such as Escherichia coli, Pseudomonas aeruginosa and Klebsiella pneumoniae. Additionally, catheter fragments coated with saline fluid enriched with CD4-PP showcased reduced E. coli attachment and dissolution of mature biofilm produced by these pathogens [90]. Thus, compared to traditional catheters, AMP-coated catheters have shown superior efficacy in reducing the risk of CAUTIs. They significantly inhibit bacterial adhesion, colonization, and biofilm formation, leading to a reduced incidence of infections. Antimicrobial peptide-coated urinary catheters hold great promise in preventing catheter-associated infections. Their unique mechanism of action, broad-spectrum activity, and low propensity for resistance make them an attractive alternative to conventional antibiotics. Despite challenges in selecting appropriate AMPs, overcoming resistance, and ensuring cost-effectiveness, the development and implementation of AMP-coated catheters offer significant potential to reduce the burden of CAUTIs and improve patient outcomes. Continued research and collaboration among scientists, clinicians, and regulatory authorities are crucial for the successful translation of this innovative technology into clinical practice.
1.3.7. The Hurdles Ahead: Constraints of Antimicrobial Peptide Biomaterials
While AMP-coated biomaterials offer promising solutions, it is important to recognize their limitations and challenges that may hinder their widespread use and effectiveness. Some AMP-coated biomaterials suffer from a limited spectrum of activity. While they may be highly effective against certain strains of bacteria, their spectrum of activity is often limited. Antimicrobial peptides may be ineffective against certain bacteria or even become less potent over time due to the development of bacterial resistance. This limitation raises concerns regarding the reliability and long-term efficacy of AMP-coated biomaterials, as they may fail to protect against a wide range of microbial threats. Nisin, for example, exhibited potent activity against Gram-positive bacteria, including various strains of Staphylococcus and Streptococcus [91,92]. However, its effectiveness against Gram-negative bacteria is limited due to the outer membrane barrier possessed by these organisms[91]. Another major concern is with peptide stability and degradation. AMPs are susceptible to enzymatic degradation and can be inactivated by various factors present in the physiological environment. The instability of these peptides can significantly impact their antimicrobial activity and longevity on the biomaterial surface. Factors such as pH, temperature, and the presence of proteases can cause peptide degradation, leading to a decrease in their effectiveness over time [93,94]. This limitation poses challenges for maintaining sustained antimicrobial activity on the coated biomaterials throughout the intended duration of use. Several proline-rich peptides, such as Bac7 and Bac5, have shown antimicrobial activity. However, their stability and vulnerability to proteolytic degradation have posed challenges in developing AMP-coated biomaterials[95]. Biofilm formation is another common challenge associated with the use of biomaterials, and antimicrobial peptide coatings may not be effective in preventing or eradicating biofilms. While AMP coatings may have some impact on preventing initial bacterial adhesion, their effectiveness against established biofilms remains limited. Besides this, concerns with antimicrobial peptide-coated biomaterials are the potential for cytotoxicity and immunogenic reactions. Some AMPs can exhibit toxicity towards host cells, impairing the healing process and causing adverse effects on surrounding tissues. Additionally, the immune response to the presence of antimicrobial peptides may lead to inflammation or hypersensitivity reactions. Moreover, the lack of standardized testing methods and criteria for evaluating antimicrobial activity and long-term performance poses hurdles in assessing and comparing the effectiveness of different coatings. These regulatory challenges can delay the translation of antimicrobial peptide-coated biomaterials into clinical practice. Formulating stable and bioactive AMP-based formulations suitable for administration represents a significant hurdle, as issues like aggregation and loss of bioactivity during formulation, storage, and administration can compromise their efficacy. Furthermore, the diverse physicochemical properties of AMPs influence their interactions with delivery vehicles and biological systems, necessitating meticulous optimization to balance therapeutic potency with delivery efficiency. Achieving efficient penetration of AMPs through barriers such as tissues, cellular membranes, and biofilm matrices adds further complexity to their effective delivery.
1.3.8. From Resistance to Resilience: Innovative Strategies to Overcome Limitations of Antimicrobial Peptide Coating Biomaterials
The therapeutic Efficacy of AMPs can be achieved through chemical modifications in constituent amino acids [96,97]. Some common strategies include incorporating D-amino acids instead of L-amino acids, introducing non-natural amino acids, or substituting specific amino acids known to be prone to degradation or enzymatic cleavage [98,99]. The sulfide bridges can provide structural stability to AMPs [100]. By introducing cysteine residues into the peptide sequence and allowing them to form disulfide bonds, the stability of the peptide can be enhanced. This method can be particularly effective for cyclic peptides. Cyclization is another chemical modification that reduces susceptibility to proteolysis and enhances resistance to chemical degradation [101]. Adding lipid moieties to AMPs can enhance their stability and membrane-binding properties. Lipidation can be achieved by attaching fatty acid chains or lipid-like groups to the peptide sequence, improving the peptide's resistance to enzymatic degradation and increasing its overall amphipathicity [102]. Identification of new structural motifs present in AMPs can be used to immobilize them on the biomaterial surfaces [103,104]. Introducing non-natural amino acids into the peptide sequence can enhance stability and resistance to proteolytic degradation. Unnatural amino acids with modified side chains or increased chemical stability can be utilized to improve peptide characteristics [105]. Some stabilizing agents can be incorporated into AMP formulations to enhance their stability. For example, protease inhibitors, metal chelators, or antioxidants can be added to protect AMPs from degradation by enzymes or reactive oxygen species [106]. Optimization of pharmacokinetic properties and the method validation of peptides can also help to prevent AMP-coating failures and degradation in clinical applications [107]. Materials with properties such as high surface area, stability, and compatibility should be used in coating applications. Examples of potential materials include metal oxides (e.g., silver, copper), carbon-based nanomaterials (e.g., graphene oxide), or polymer-based nanoparticles. Nanoparticles have effectively demonstrated their capacity to encapsulate established drugs used in the treatment of specific diseases, including certain types of cancer[108]. The interactions between the AMPs and chosen materials/nanoparticles should be well characterized and optimized (Figure 6).
In 2017, Casciaro et al. documented the first report of a covalent linkage between the antimicrobial peptide, esculentin-1a (which exhibits potent activity against Pseudomonas aeruginosa bacteria and soluble gold nanoparticles (AuNPs) via a polyethylene glycol (PEG) linker. The resultant compound, AuNPs@Esc(1-21), displayed nearly 15-fold increased antipseudomonal activity compared to Esc(1-21) alone, without inducing toxicity in human cells[109]. Chaudhari et al. conducted an assessment of the toxicity and antimicrobial efficacy of various antimicrobial peptides (TP359, TP226, and TP557) when incorporated onto silver-coated carbon nanotubes (CNTs). This investigation focused on combating Staphylococcus aureus infection within a three-dimensional human skin model of full thickness[110]. Nordström et al. showed that significant quantities of cationic antimicrobial peptides LL-37 and DPK-060 can be encapsulated within anionic poly(ethyl acrylate-co-methacrylic acid) microgels. These microgels demonstrated the ability to shield the enclosed peptides from degradation by infection-associated proteases, especially when a high microgel charge density was utilized[77]. Zetterberg et al explored the application of PEG-stabilized liposomes as carriers for antimicrobial peptides (AMPs). They examined the susceptibility of the melittin liposome to proteases and its antimicrobial efficacy in comparison to free melittin. Through repeated exposure to E. coli, the melittin liposomes displayed notable bactericidal activity upon secondary exposure, outperforming free melittin. This effect was attributed to the time-dependent release of AMP from the liposomes. Interestingly, the liposome-encapsulated melittin was completely shielded from trypsin degradation, highlighting its enhanced stability [111]. Likewise, in various other investigations, cyclodextrins, hydrogels, and dendritic systems have been employed as carriers for antimicrobial peptides.
2. Conclusions
In conclusion, AMPs represent a promising avenue for combating infectious diseases and addressing the growing issue of antibiotic resistance. These small molecules, derived from natural sources, have shown potent antimicrobial activity against a wide range of pathogens, including bacteria, fungi, and even some viruses. By integrating AMPs onto the surface of biomaterials such as medical implants, wound dressings, or catheters, we can effectively inhibit the colonization and growth of harmful microorganisms, reducing the risk of infections. The use of AMP-coated biomaterials offers several advantages over traditional antimicrobial strategies. Furthermore, AMP-coated biomaterials have shown promising results in terms of biocompatibility and reduced cytotoxicity. Studies have demonstrated that AMPs can selectively kill pathogens while preserving host cells, minimizing the risk of adverse reactions or tissue damage. This makes them suitable for long-term use in medical devices and implants, where compatibility with the human body is crucial. Although there are still challenges to overcome, such as optimizing the stability and delivery of AMPs, as well as addressing potential manufacturing and regulatory hurdles, the development of AMP-coated biomaterials holds great promise. Continued research and innovation in this field will pave the way for new and effective strategies in combating infections and improving patient outcomes. In conclusion, antimicrobial peptides and their integration into AMP-coated biomaterials offer a compelling solution to the global challenge of antimicrobial resistance. Through further advancements and collaborations between researchers, healthcare professionals, and industry partners, we can harness the power of these naturally occurring molecules to develop innovative therapies and biomedical devices that effectively combat infections and enhance human health.
Author Contributions
A.K.T. and J.S. wrote the different sections of the review, R.T. and P.R. included diagrams. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data supporting the findings of this study are available from the corresponding authors upon reasonable request.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Abushaheen, M.A.; Muzaheed; Fatani, A.J.; Alosaimi, M.; Mansy, W.; George, M.; Acharya, S.; Rathod, S.; Divakar, D.D.; Jhugroo, C.; et al. Antimicrobial resistance, mechanisms and its clinical significance. Dis Mon 2020, 66, 100971. [CrossRef]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L.; et al. The value of antimicrobial peptides in the age of resistance. Lancet Infect Dis 2020, 20, e216-e230. [CrossRef]
- Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Environmental factors influencing the development and spread of antibiotic resistance. FEMS Microbiol Rev 2018, 42. [CrossRef]
- Christaki, E.; Marcou, M.; Tofarides, A. Antimicrobial Resistance in Bacteria: Mechanisms, Evolution, and Persistence. J Mol Evol 2020, 88, 26-40. [CrossRef]
- Ferri, M.; Ranucci, E.; Romagnoli, P.; Giaccone, V. Antimicrobial resistance: A global emerging threat to public health systems. Crit Rev Food Sci Nutr 2017, 57, 2857-2876. [CrossRef]
- Wieler, L.H.; Ewers, C.; Guenther, S.; Walther, B.; Lubke-Becker, A. Methicillin-resistant staphylococci (MRS) and extended-spectrum beta-lactamases (ESBL)-producing Enterobacteriaceae in companion animals: nosocomial infections as one reason for the rising prevalence of these potential zoonotic pathogens in clinical samples. Int J Med Microbiol 2011, 301, 635-641. [CrossRef]
- Ai, M.; Lin, S.; Zhang, M.; Wu, T.; Yang, N.; Li, Y.; Li, L. Cirsilineol attenuates LPS-induced inflammation in both in vivo and in vitro models via inhibiting TLR-4/NFkB/IKK signaling pathway. J Biochem Mol Toxicol 2021, 35, e22799. [CrossRef]
- Chen, X.L.; Wang, Y.; Peng, W.W.; Zheng, Y.J.; Zhang, T.N.; Wang, P.J.; Huang, J.D.; Zeng, Q.Y. Effects of interleukin-6 and IL-6/AMPK signaling pathway on mitochondrial biogenesis and astrocytes viability under experimental septic condition. Int Immunopharmacol 2018, 59, 287-294. [CrossRef]
- Ushio, N.; Wada, T.; Ono, Y.; Yamakawa, K. Sepsis-induced disseminated intravascular coagulation: an international estrangement of disease concept. Acute Med Surg 2023, 10, e00843. [CrossRef]
- Srivastava, S.; Kumar, A.; Tripathi, A.K.; Tandon, A.; Ghosh, J.K. Modulation of anti-endotoxin property of Temporin L by minor amino acid substitution in identified phenylalanine zipper sequence. Biochem J 2016, 473, 4045-4062. [CrossRef]
- Tripathi, A.K.; Vishwanatha, J.K. Role of Anti-Cancer Peptides as Immunomodulatory Agents: Potential and Design Strategy. Pharmaceutics 2022, 14. [CrossRef]
- Kumari, T.; Verma, D.P.; Kuldeep, J.; Dhanabal, V.B.; Verma, N.K.; Sahai, R.; Tripathi, A.K.; Saroj, J.; Ali, M.; Mitra, K.; et al. 10-Residue MyD88-Peptide Adopts beta-Sheet Structure, Self-Assembles, Binds to Lipopolysaccharides, and Rescues Mice from Endotoxin-Mediated Lung-Infection and Death. ACS Chem Biol 2022, 17, 3420-3434. [CrossRef]
- Kumar, A.; Tripathi, A.K.; Kathuria, M.; Shree, S.; Tripathi, J.K.; Purshottam, R.K.; Ramachandran, R.; Mitra, K.; Ghosh, J.K. Single Amino Acid Substitutions at Specific Positions of the Heptad Repeat Sequence of Piscidin-1 Yielded Novel Analogs That Show Low Cytotoxicity and In Vitro and In Vivo Antiendotoxin Activity. Antimicrob Agents Chemother 2016, 60, 3687-3699. [CrossRef]
- Tripathi, A.K.; Kumari, T.; Tandon, A.; Sayeed, M.; Afshan, T.; Kathuria, M.; Shukla, P.K.; Mitra, K.; Ghosh, J.K. Selective phenylalanine to proline substitution for improved antimicrobial and anticancer activities of peptides designed on phenylalanine heptad repeat. Acta Biomater 2017, 57, 170-186. [CrossRef]
- Shoaie, S.; Karlsson, F.; Mardinoglu, A.; Nookaew, I.; Bordel, S.; Nielsen, J. Understanding the interactions between bacteria in the human gut through metabolic modeling. Sci Rep 2013, 3, 2532. [CrossRef]
- Tlaskalova-Hogenova, H.; Stepankova, R.; Hudcovic, T.; Tuckova, L.; Cukrowska, B.; Lodinova-Zadnikova, R.; Kozakova, H.; Rossmann, P.; Bartova, J.; Sokol, D.; et al. Commensal bacteria (normal microflora), mucosal immunity and chronic inflammatory and autoimmune diseases. Immunol Lett 2004, 93, 97-108. [CrossRef]
- van Vught, L.A.; Klein Klouwenberg, P.M.; van der Poll, T. Secondary Infection in Patients With Sepsis--Reply. JAMA 2016, 316, 772. [CrossRef]
- Lachica, M.; Anutrakunchai, C.; Prajaneh, S.; Nazmi, K.; Bolscher, J.G.M.; Taweechaisupapong, S. Synergistic effects of LFchimera and antibiotic against planktonic and biofilm form of Aggregatibacter actinomycetemcomitans. PLoS One 2019, 14, e0217205. [CrossRef]
- Duong, L.; Gross, S.P.; Siryaporn, A. Developing Antimicrobial Synergy With AMPs. Front Med Technol 2021, 3, 640981. [CrossRef]
- Abebe, G.M. The Role of Bacterial Biofilm in Antibiotic Resistance and Food Contamination. Int J Microbiol 2020, 2020, 1705814. [CrossRef]
- Patterson-Delafield, J.; Martinez, R.J.; Lehrer, R.I. Microbicidal cationic proteins in rabbit alveolar macrophages: a potential host defense mechanism. Infect Immun 1980, 30, 180-192. [CrossRef]
- Patterson-Delafield, J.; Szklarek, D.; Martinez, R.J.; Lehrer, R.I. Microbicidal cationic proteins of rabbit alveolar macrophages: amino acid composition and functional attributes. Infect Immun 1981, 31, 723-731. [CrossRef]
- Ganz, T.; Selsted, M.E.; Szklarek, D.; Harwig, S.S.; Daher, K.; Bainton, D.F.; Lehrer, R.I. Defensins. Natural peptide antibiotics of human neutrophils. J Clin Invest 1985, 76, 1427-1435. [CrossRef]
- Bulet, P.; Dimarcq, J.L.; Hetru, C.; Lagueux, M.; Charlet, M.; Hegy, G.; Van Dorsselaer, A.; Hoffmann, J.A. A novel inducible antibacterial peptide of Drosophila carries an O-glycosylated substitution. J Biol Chem 1993, 268, 14893-14897.
- Hanson, M.A.; Kondo, S.; Lemaitre, B. Drosophila immunity: the Drosocin gene encodes two host defence peptides with pathogen-specific roles. Proc Biol Sci 2022, 289, 20220773. [CrossRef]
- Mangano, K.; Klepacki, D.; Ohanmu, I.; Baliga, C.; Huang, W.; Brakel, A.; Krizsan, A.; Polikanov, Y.S.; Hoffmann, R.; Vazquez-Laslop, N.; et al. Inhibition of translation termination by the antimicrobial peptide Drosocin. Nat Chem Biol 2023. [CrossRef]
- Uttenweiler-Joseph, S.; Moniatte, M.; Lagueux, M.; Van Dorsselaer, A.; Hoffmann, J.A.; Bulet, P. Differential display of peptides induced during the immune response of Drosophila: a matrix-assisted laser desorption ionization time-of-flight mass spectrometry study. Proc Natl Acad Sci U S A 1998, 95, 11342-11347. [CrossRef]
- Moreno, M.; Giralt, E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: melittin, apamin and mastoparan. Toxins (Basel) 2015, 7, 1126-1150. [CrossRef]
- Vogel, H.; Jahnig, F. The structure of melittin in membranes. Biophys J 1986, 50, 573-582. [CrossRef]
- Asthana, N.; Yadav, S.P.; Ghosh, J.K. Dissection of antibacterial and toxic activity of melittin: a leucine zipper motif plays a crucial role in determining its hemolytic activity but not antibacterial activity. J Biol Chem 2004, 279, 55042-55050. [CrossRef]
- Maccari, G.; Di Luca, M.; Nifosi, R. In silico design of antimicrobial peptides. Methods Mol Biol 2015, 1268, 195-219. [CrossRef]
- Lata, S.; Mishra, N.K.; Raghava, G.P. AntiBP2: improved version of antibacterial peptide prediction. BMC Bioinformatics 2010, 11 Suppl 1, S19. [CrossRef]
- Waghu, F.H.; Barai, R.S.; Gurung, P.; Idicula-Thomas, S. CAMPR3: a database on sequences, structures and signatures of antimicrobial peptides. Nucleic Acids Res 2016, 44, D1094-1097. [CrossRef]
- Veltri, D.; Kamath, U.; Shehu, A. Deep learning improves antimicrobial peptide recognition. Bioinformatics 2018, 34, 2740-2747. [CrossRef]
- Mabrouk, D.M. Antimicrobial peptides: features, applications and the potential use against covid-19. Mol Biol Rep 2022, 49, 10039-10050. [CrossRef]
- Han, M.; Mei, Y.; Khant, H.; Ludtke, S.J. Characterization of antibiotic peptide pores using cryo-EM and comparison to neutron scattering. Biophys J 2009, 97, 164-172. [CrossRef]
- Tripathi, A.K.; Vishwanatha, J.K. Abstract 2184: Short Peptides derived from MIEN1 and their analogs exhibit anti-cancer activity in breast and prostate cancer cells. Cancer Research 2023, 83, 2184-2184. [CrossRef]
- Tandon, A.; Harioudh, M.K.; Ishrat, N.; Tripathi, A.K.; Srivastava, S.; Ghosh, J.K. An MD2-derived peptide promotes LPS aggregation, facilitates its internalization in THP-1 cells, and inhibits LPS-induced pro-inflammatory responses. Cell Mol Life Sci 2018, 75, 2431-2446. [CrossRef]
- Guo, C.; Trivedi, R.; Tripathi, A.K.; Nandy, R.R.; Wagner, D.C.; Narra, K.; Chaudhary, P. Higher Expression of Annexin A2 in Metastatic Bladder Urothelial Carcinoma Promotes Migration and Invasion. Cancers (Basel) 2022, 14. [CrossRef]
- Zhang, Q.Y.; Yan, Z.B.; Meng, Y.M.; Hong, X.Y.; Shao, G.; Ma, J.J.; Cheng, X.R.; Liu, J.; Kang, J.; Fu, C.Y. Antimicrobial peptides: mechanism of action, activity and clinical potential. Mil Med Res 2021, 8, 48. [CrossRef]
- Hou, Y.; Collinsworth, A.; Hasa, F.; Griffin, L. Incidence and impact of surgical site infections on length of stay and cost of care for patients undergoing open procedures. Surg Open Sci 2023, 11, 1-18. [CrossRef]
- Pulat, G.; Muganli, Z.; Ercan, U.K.; Karaman, O. Effect of antimicrobial peptide conjugated surgical sutures on multiple drug-resistant microorganisms. J Biomater Appl 2023, 37, 1182-1194. [CrossRef]
- Yasir, M.; Willcox, M.D.P.; Dutta, D. Action of Antimicrobial Peptides against Bacterial Biofilms. Materials (Basel) 2018, 11. [CrossRef]
- Petkovic, M.; Mouritzen, M.V.; Mojsoska, B.; Jenssen, H. Immunomodulatory Properties of Host Defence Peptides in Skin Wound Healing. Biomolecules 2021, 11. [CrossRef]
- Morroni, G.; Simonetti, O.; Brenciani, A.; Brescini, L.; Kamysz, W.; Kamysz, E.; Neubauer, D.; Caffarini, M.; Orciani, M.; Giovanetti, E.; et al. In vitro activity of Protegrin-1, alone and in combination with clinically useful antibiotics, against Acinetobacter baumannii strains isolated from surgical wounds. Med Microbiol Immunol 2019, 208, 877-883. [CrossRef]
- Khurshid, Z.; Najeeb, S.; Mali, M.; Moin, S.F.; Raza, S.Q.; Zohaib, S.; Sefat, F.; Zafar, M.S. Histatin peptides: Pharmacological functions and their applications in dentistry. Saudi Pharm J 2017, 25, 25-31. [CrossRef]
- Desbois, A.P.; Gemmell, C.G.; Coote, P.J. In vivo efficacy of the antimicrobial peptide ranalexin in combination with the endopeptidase lysostaphin against wound and systemic meticillin-resistant Staphylococcus aureus (MRSA) infections. Int J Antimicrob Agents 2010, 35, 559-565. [CrossRef]
- Gopinath, D.; Kumar, M.S.; Selvaraj, D.; Jayakumar, R. Pexiganan-incorporated collagen matrices for infected wound-healing processes in rat. J Biomed Mater Res A 2005, 73, 320-331. [CrossRef]
- Franco, A.R.; Fernandes, E.M.; Rodrigues, M.T.; Rodrigues, F.J.; Gomes, M.E.; Leonor, I.B.; Kaplan, D.L.; Reis, R.L. Antimicrobial coating of spider silk to prevent bacterial attachment on silk surgical sutures. Acta Biomater 2019, 99, 236-246. [CrossRef]
- Sankaridurg, P.R.; Sharma, S.; Willcox, M.; Naduvilath, T.J.; Sweeney, D.F.; Holden, B.A.; Rao, G.N. Bacterial colonization of disposable soft contact lenses is greater during corneal infiltrative events than during asymptomatic extended lens wear. J Clin Microbiol 2000, 38, 4420-4424. [CrossRef]
- Zimmerman, A.B.; Nixon, A.D.; Rueff, E.M. Contact lens associated microbial keratitis: practical considerations for the optometrist. Clin Optom (Auckl) 2016, 8, 1-12. [CrossRef]
- Niederkorn, J.Y. Mechanisms of immune privilege in the eye and hair follicle. J Investig Dermatol Symp Proc 2003, 8, 168-172. [CrossRef]
- Willcox, M.D.; Hume, E.B.; Aliwarga, Y.; Kumar, N.; Cole, N. A novel cationic-peptide coating for the prevention of microbial colonization on contact lenses. J Appl Microbiol 2008, 105, 1817-1825. [CrossRef]
- Maciejewska, M.; Bauer, M.; Neubauer, D.; Kamysz, W.; Dawgul, M. Influence of Amphibian Antimicrobial Peptides and Short Lipopeptides on Bacterial Biofilms Formed on Contact Lenses. Materials (Basel) 2016, 9. [CrossRef]
- Sharma, S.; Srinivasan, M.; George, C. Acanthamoeba keratitis in non-contact lens wearers. Arch Ophthalmol 1990, 108, 676-678. [CrossRef]
- Ting, D.S.J.; Mohammed, I.; Lakshminarayanan, R.; Beuerman, R.W.; Dua, H.S. Host Defense Peptides at the Ocular Surface: Roles in Health and Major Diseases, and Therapeutic Potentials. Front Med (Lausanne) 2022, 9, 835843. [CrossRef]
- Haque, M.; Sartelli, M.; Haque, S.Z. Dental Infection and Resistance-Global Health Consequences. Dent J (Basel) 2019, 7. [CrossRef]
- Khurshid, Z.; Naseem, M.; Yahya I. Asiri, F.; Mali, M.; Sannam Khan, R.; Sahibzada, H.A.; Zafar, M.S.; Faraz Moin, S.; Khan, E. Significance and Diagnostic Role of Antimicrobial Cathelicidins (LL-37) Peptides in Oral Health. Biomolecules 2017, 7, 80.
- Porat, Y.; Marynka, K.; Tam, A.; Steinberg, D.; Mor, A. Acyl-substituted dermaseptin S4 derivatives with improved bactericidal properties, including on oral microflora. Antimicrob Agents Chemother 2006, 50, 4153-4160. [CrossRef]
- Tong, Z.; Dong, L.; Zhou, L.; Tao, R.; Ni, L. Nisin inhibits dental caries-associated microorganism in vitro. Peptides 2010, 31, 2003-2008. [CrossRef]
- Zannella, C.; Shinde, S.; Vitiello, M.; Falanga, A.; Galdiero, E.; Fahmi, A.; Santella, B.; Nucci, L.; Gasparro, R.; Galdiero, M.; et al. Antibacterial Activity of Indolicidin-Coated Silver Nanoparticles in Oral Disease. Applied Sciences 2020, 10, 1837.
- Ji, S.; Hyun, J.; Park, E.; Lee, B.L.; Kim, K.K.; Choi, Y. Susceptibility of various oral bacteria to antimicrobial peptides and to phagocytosis by neutrophils. J Periodontal Res 2007, 42, 410-419. [CrossRef]
- Lee, S.H.; Baek, D.H. Antibacterial and neutralizing effect of human beta-defensins on Enterococcus faecalis and Enterococcus faecalis lipoteichoic acid. J Endod 2012, 38, 351-356. [CrossRef]
- Lee, J.K.; Park, Y.J.; Kum, K.Y.; Han, S.H.; Chang, S.W.; Kaufman, B.; Jiang, J.; Zhu, Q.; Safavi, K.; Spangberg, L. Antimicrobial efficacy of a human beta-defensin-3 peptide using an Enterococcus faecalis dentine infection model. Int Endod J 2013, 46, 406-412. [CrossRef]
- Mancino, D.; Kharouf, N.; Scavello, F.; Helle, S.; Salloum-Yared, F.; Mutschler, A.; Mathieu, E.; Lavalle, P.; Metz-Boutigue, M.H.; Haikel, Y. The Catestatin-Derived Peptides Are New Actors to Fight the Development of Oral Candidosis. Int J Mol Sci 2022, 23. [CrossRef]
- Xie, S.X.; Song, L.; Yuca, E.; Boone, K.; Sarikaya, R.; VanOosten, S.K.; Misra, A.; Ye, Q.; Spencer, P.; Tamerler, C. Antimicrobial Peptide-Polymer Conjugates for Dentistry. ACS Appl Polym Mater 2020, 2, 1134-1144. [CrossRef]
- Lombardi, L.; Shi, Y.; Falanga, A.; Galdiero, E.; de Alteriis, E.; Franci, G.; Chourpa, I.; Azevedo, H.S.; Galdiero, S. Enhancing the Potency of Antimicrobial Peptides through Molecular Engineering and Self-Assembly. Biomacromolecules 2019, 20, 1362-1374. [CrossRef]
- Chen, J.; Zhu, Y.; Xiong, M.; Hu, G.; Zhan, J.; Li, T.; Wang, L.; Wang, Y. Antimicrobial Titanium Surface via Click-Immobilization of Peptide and Its in Vitro/Vivo Activity. ACS Biomater Sci Eng 2019, 5, 1034-1044. [CrossRef]
- Pihl, M.; Galli, S.; Jimbo, R.; Andersson, M. Osseointegration and antibacterial effect of an antimicrobial peptide releasing mesoporous titania implant. J Biomed Mater Res B Appl Biomater 2021, 109, 1787-1795. [CrossRef]
- Yucesoy, D.T.; Hnilova, M.; Boone, K.; Arnold, P.M.; Snead, M.L.; Tamerler, C. Chimeric peptides as implant functionalization agents for titanium alloy implants with antimicrobial properties. JOM (1989) 2015, 67, 754-766. [CrossRef]
- Tripathi, J.K.; Pal, S.; Awasthi, B.; Kumar, A.; Tandon, A.; Mitra, K.; Chattopadhyay, N.; Ghosh, J.K. Variants of self-assembling peptide, KLD-12 that show both rapid fracture healing and antimicrobial properties. Biomaterials 2015, 56, 92-103. [CrossRef]
- Qi, X.; Poernomo, G.; Wang, K.; Chen, Y.; Chan-Park, M.B.; Xu, R.; Chang, M.W. Covalent immobilization of nisin on multi-walled carbon nanotubes: superior antimicrobial and anti-biofilm properties. Nanoscale 2011, 3, 1874-1880. [CrossRef]
- Sekar, P.C.; Rajasekaran, R. Could Dermaseptin Analogue be a Competitive Inhibitor for ACE2 Towards Binding with Viral Spike Protein Causing COVID19?: Computational Investigation. Int J Pept Res Ther 2021, 27, 1043-1056. [CrossRef]
- Gang, D.; Kim, D.W.; Park, H.S. Cyclic Peptides: Promising Scaffolds for Biopharmaceuticals. Genes (Basel) 2018, 9. [CrossRef]
- Yang, X.; Guo, J.L.; Han, J.; Si, R.J.; Liu, P.P.; Zhang, Z.R.; Wang, A.M.; Zhang, J. Chitosan hydrogel encapsulated with LL-37 peptide promotes deep tissue injury healing in a mouse model. Mil Med Res 2020, 7, 20. [CrossRef]
- Silva, J.P.; Dhall, S.; Garcia, M.; Chan, A.; Costa, C.; Gama, M.; Martins-Green, M. Improved burn wound healing by the antimicrobial peptide LLKKK18 released from conjugates with dextrin embedded in a carbopol gel. Acta Biomater 2015, 26, 249-262. [CrossRef]
- Nordstrom, R.; Nystrom, L.; Andren, O.C.J.; Malkoch, M.; Umerska, A.; Davoudi, M.; Schmidtchen, A.; Malmsten, M. Membrane interactions of microgels as carriers of antimicrobial peptides. J Colloid Interface Sci 2018, 513, 141-150. [CrossRef]
- Patrulea, V.; Borchard, G.; Jordan, O. An Update on Antimicrobial Peptides (AMPs) and Their Delivery Strategies for Wound Infections. Pharmaceutics 2020, 12. [CrossRef]
- Abdel-Sayed, P.; Kaeppeli, A.; Siriwardena, T.; Darbre, T.; Perron, K.; Jafari, P.; Reymond, J.L.; Pioletti, D.P.; Applegate, L.A. Anti-Microbial Dendrimers against Multidrug-Resistant P. aeruginosa Enhance the Angiogenic Effect of Biological Burn-wound Bandages. Sci Rep 2016, 6, 22020. [CrossRef]
- Parreira, P.; Monteiro, C.; Graca, V.; Gomes, J.; Maia, S.; Gomes, P.; Goncalves, I.C.; Martins, M.C.L. Surface Grafted MSI-78A Antimicrobial Peptide has High Potential for Gastric Infection Management. Sci Rep 2019, 9, 18212. [CrossRef]
- Wang, C.; Feng, S.; Qie, J.; Wei, X.; Yan, H.; Liu, K. Polyion complexes of a cationic antimicrobial peptide as a potential systemically administered antibiotic. Int J Pharm 2019, 554, 284-291. [CrossRef]
- Werneburg, G.T. Catheter-Associated Urinary Tract Infections: Current Challenges and Future Prospects. Res Rep Urol 2022, 14, 109-133. [CrossRef]
- Jacobsen, S.M.; Stickler, D.J.; Mobley, H.L.; Shirtliff, M.E. Complicated catheter-associated urinary tract infections due to Escherichia coli and Proteus mirabilis. Clin Microbiol Rev 2008, 21, 26-59. [CrossRef]
- Li, X.; Li, P.; Saravanan, R.; Basu, A.; Mishra, B.; Lim, S.H.; Su, X.; Tambyah, P.A.; Leong, S.S. Antimicrobial functionalization of silicone surfaces with engineered short peptides having broad spectrum antimicrobial and salt-resistant properties. Acta Biomater 2014, 10, 258-266. [CrossRef]
- Hilpert, K.; Elliott, M.; Jenssen, H.; Kindrachuk, J.; Fjell, C.D.; Korner, J.; Winkler, D.F.; Weaver, L.L.; Henklein, P.; Ulrich, A.S.; et al. Screening and characterization of surface-tethered cationic peptides for antimicrobial activity. Chem Biol 2009, 16, 58-69. [CrossRef]
- Nicolas, M.; Beito, B.; Oliveira, M.; Tudela Martins, M.; Gallas, B.; Salmain, M.; Boujday, S.; Humblot, V. Strategies for Antimicrobial Peptides Immobilization on Surfaces to Prevent Biofilm Growth on Biomedical Devices. Antibiotics (Basel) 2021, 11. [CrossRef]
- Monteiro, C.; Costa, F.; Pirttila, A.M.; Tejesvi, M.V.; Martins, M.C.L. Prevention of urinary catheter-associated infections by coating antimicrobial peptides from crowberry endophytes. Sci Rep 2019, 9, 10753. [CrossRef]
- Yu, K.; Lo, J.C.; Yan, M.; Yang, X.; Brooks, D.E.; Hancock, R.E.; Lange, D.; Kizhakkedathu, J.N. Anti-adhesive antimicrobial peptide coating prevents catheter associated infection in a mouse urinary infection model. Biomaterials 2017, 116, 69-81. [CrossRef]
- Yao, Q.; Chen, B.; Bai, J.; He, W.; Chen, X.; Geng, D.; Pan, G. Bio-inspired antibacterial coatings on urinary stents for encrustation prevention. J Mater Chem B 2022, 10, 2584-2596. [CrossRef]
- White, J.K.; Muhammad, T.; Alsheim, E.; Mohanty, S.; Blasi-Romero, A.; Gunasekera, S.; Stromstedt, A.A.; Ferraz, N.; Goransson, U.; Brauner, A. A stable cyclized antimicrobial peptide derived from LL-37 with host immunomodulatory effects and activity against uropathogens. Cell Mol Life Sci 2022, 79, 411. [CrossRef]
- Helander, I.M.; Mattila-Sandholm, T. Permeability barrier of the gram-negative bacterial outer membrane with special reference to nisin. Int J Food Microbiol 2000, 60, 153-161. [CrossRef]
- Boziaris, I.S.; Adams, M.R. Effect of chelators and nisin produced in situ on inhibition and inactivation of gram negatives. Int J Food Microbiol 1999, 53, 105-113. [CrossRef]
- Starr, C.G.; Wimley, W.C. Antimicrobial peptides are degraded by the cytosolic proteases of human erythrocytes. Biochim Biophys Acta Biomembr 2017, 1859, 2319-2326. [CrossRef]
- Hitchner, M.A.; Santiago-Ortiz, L.E.; Necelis, M.R.; Shirley, D.J.; Palmer, T.J.; Tarnawsky, K.E.; Vaden, T.D.; Caputo, G.A. Activity and characterization of a pH-sensitive antimicrobial peptide. Biochim Biophys Acta Biomembr 2019, 1861, 182984. [CrossRef]
- Mattiuzzo, M.; De Gobba, C.; Runti, G.; Mardirossian, M.; Bandiera, A.; Gennaro, R.; Scocchi, M. Proteolytic activity of Escherichia coli oligopeptidase B against proline-rich antimicrobial peptides. J Microbiol Biotechnol 2014, 24, 160-167. [CrossRef]
- D'Souza, A.R.; Necelis, M.R.; Kulesha, A.; Caputo, G.A.; Makhlynets, O.V. Beneficial Impacts of Incorporating the Non-Natural Amino Acid Azulenyl-Alanine into the Trp-Rich Antimicrobial Peptide buCATHL4B. Biomolecules 2021, 11. [CrossRef]
- Lu, J.; Xu, H.; Xia, J.; Ma, J.; Xu, J.; Li, Y.; Feng, J. D- and Unnatural Amino Acid Substituted Antimicrobial Peptides With Improved Proteolytic Resistance and Their Proteolytic Degradation Characteristics. Front Microbiol 2020, 11, 563030. [CrossRef]
- Verma, N.K.; Dewangan, R.P.; Harioudh, M.K.; Ghosh, J.K. Introduction of a beta-leucine residue instead of leucine(9) and glycine(10) residues in Temporin L for improved cell selectivity, stability and activity against planktonic and biofilm of methicillin resistant S. aureus. Bioorg Chem 2023, 134, 106440. [CrossRef]
- Caldwell, M.; Hughes, M.; Wei, F.; Ngo, C.; Pascua, R.; Pugazhendhi, A.S.; Coathup, M.J. Promising applications of D-amino acids in periprosthetic joint infection. Bone Res 2023, 11, 14. [CrossRef]
- Liu, T.; Wang, Y.; Luo, X.; Li, J.; Reed, S.A.; Xiao, H.; Young, T.S.; Schultz, P.G. Enhancing protein stability with extended disulfide bonds. Proc Natl Acad Sci U S A 2016, 113, 5910-5915. [CrossRef]
- Han, Y.; Zhang, M.; Lai, R.; Zhang, Z. Chemical modifications to increase the therapeutic potential of antimicrobial peptides. Peptides 2021, 146, 170666. [CrossRef]
- Kowalczyk, R.; Harris, P.W.R.; Williams, G.M.; Yang, S.H.; Brimble, M.A. Peptide Lipidation - A Synthetic Strategy to Afford Peptide Based Therapeutics. Adv Exp Med Biol 2017, 1030, 185-227. [CrossRef]
- Tripathi, A.K.; Kumari, T.; Harioudh, M.K.; Yadav, P.K.; Kathuria, M.; Shukla, P.K.; Mitra, K.; Ghosh, J.K. Identification of GXXXXG motif in Chrysophsin-1 and its implication in the design of analogs with cell-selective antimicrobial and anti-endotoxin activities. Sci Rep 2017, 7, 3384. [CrossRef]
- Saxena, R.; Vekariya, U.K.; Kumar, P.; Tripathi, A.K.; Ghosh, J.K.; Tripathi, R.K. HIV-1 Nef CAWLEAQ motif: a regulator of monocytes invasion through ENO1 modulation. Mol Cell Biochem 2018, 447, 151-164. [CrossRef]
- Qvit, N.; Rubin, S.J.S.; Urban, T.J.; Mochly-Rosen, D.; Gross, E.R. Peptidomimetic therapeutics: scientific approaches and opportunities. Drug Discov Today 2017, 22, 454-462. [CrossRef]
- Grassi, L.; Cabrele, C. Susceptibility of protein therapeutics to spontaneous chemical modifications by oxidation, cyclization, and elimination reactions. Amino Acids 2019, 51, 1409-1431. [CrossRef]
- Puttrevu, S.K.; Laxman, T.S.; Tripathi, A.K.; Yadav, A.K.; Verma, S.K.; Mishra, A.; Pradhan, R.; Verma, N.K.; Ghosh, J.K.; Bhatta, R.S. Liquid chromatography-tandem mass spectrometry based method development and validation of S016-1271 (LR8P), a novel cationic antimicrobial peptide for its application to pharmacokinetic studies. J Pharm Biomed Anal 2019, 169, 116-126. [CrossRef]
- Lampe, J.B.; Desai, P.P.; Tripathi, A.K.; Sabnis, N.A.; Chen, Z.; Ranjan, A.P.; Vishwanatha, J.K. Cabazitaxel-Loaded Nanoparticles Reduce the Invasiveness in Metastatic Prostate Cancer Cells: Beyond the Classical Taxane Function. Pharmaceutics 2023, 15. [CrossRef]
- Casciaro, B.; Moros, M.; Rivera-Fernandez, S.; Bellelli, A.; de la Fuente, J.M.; Mangoni, M.L. Gold-nanoparticles coated with the antimicrobial peptide esculentin-1a(1-21)NH(2) as a reliable strategy for antipseudomonal drugs. Acta Biomater 2017, 47, 170-181. [CrossRef]
- Chaudhari, A.A.; Joshi, S.; Vig, K.; Sahu, R.; Dixit, S.; Baganizi, R.; Dennis, V.A.; Singh, S.R.; Pillai, S. A three-dimensional human skin model to evaluate the inhibition of Staphylococcus aureus by antimicrobial peptide-functionalized silver carbon nanotubes. J Biomater Appl 2019, 33, 924-934. [CrossRef]
- Zetterberg, M.M.; Reijmar, K.; Pranting, M.; Engstrom, A.; Andersson, D.I.; Edwards, K. PEG-stabilized lipid disks as carriers for amphiphilic antimicrobial peptides. J Control Release 2011, 156, 323-328. [CrossRef]
Figure 1.
Multifaceted functions of (AMPs): (A) Depiction of the signaling pathway involved in Gram-Negative Bacteria-Induced Infection caused by Lipopolysaccharides (LPS). (B) AMPs bind directly to LPS, leading to the inhibition of LPs-mediated inflammation.(C) Cationic AMPs bind to the negatively charged phosphatidyl serine (PS) heads on cancer cell membranes.
Figure 1.
Multifaceted functions of (AMPs): (A) Depiction of the signaling pathway involved in Gram-Negative Bacteria-Induced Infection caused by Lipopolysaccharides (LPS). (B) AMPs bind directly to LPS, leading to the inhibition of LPs-mediated inflammation.(C) Cationic AMPs bind to the negatively charged phosphatidyl serine (PS) heads on cancer cell membranes.
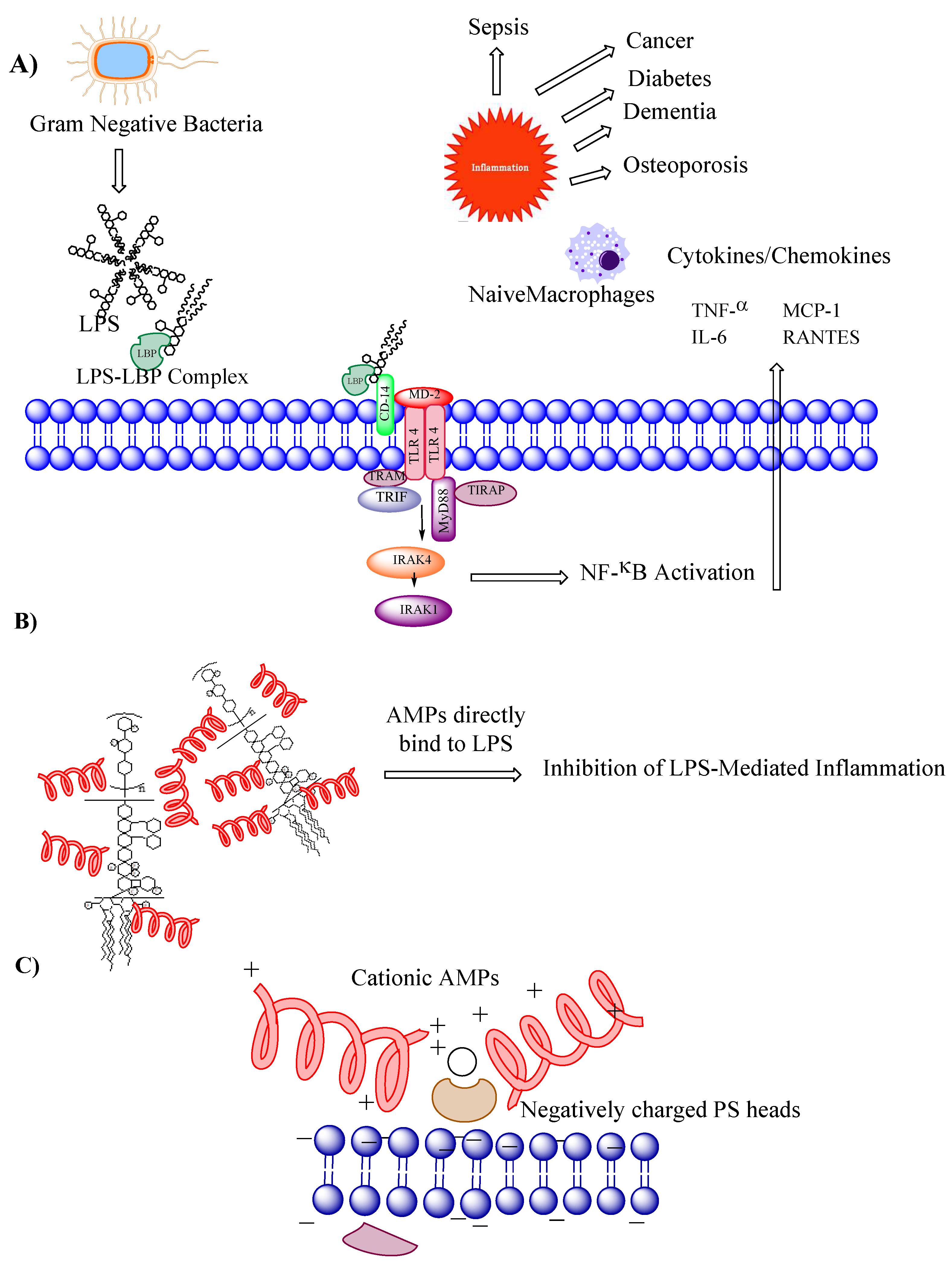
Figure 2.
AMPs count in Antimicrobial Peptide Database at each net positive charge and percent hydrophobicity.
Figure 2.
AMPs count in Antimicrobial Peptide Database at each net positive charge and percent hydrophobicity.
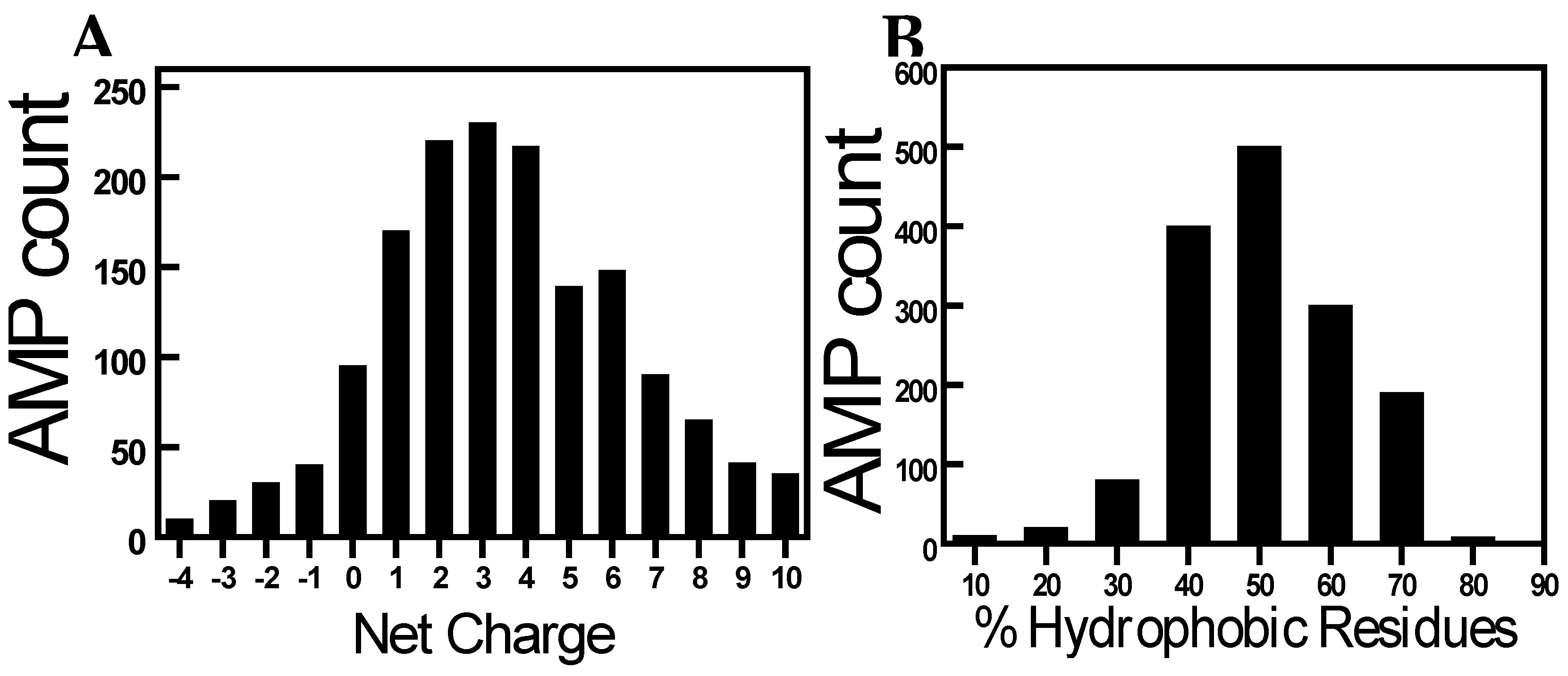
Figure 3.
AMPs to fight against Contact lens-associated microbial keratitis (CLMK). (A): Illustrates the process of microbial attachment to a contact lens, leading to microbial keratitis and eye infections. (B): Demonstrates the application of AMPs immobilized on contact lenses, effectively inhibiting microbial proliferation on the lens surface, thereby contributing to the maintenance of ocular health.
Figure 3.
AMPs to fight against Contact lens-associated microbial keratitis (CLMK). (A): Illustrates the process of microbial attachment to a contact lens, leading to microbial keratitis and eye infections. (B): Demonstrates the application of AMPs immobilized on contact lenses, effectively inhibiting microbial proliferation on the lens surface, thereby contributing to the maintenance of ocular health.
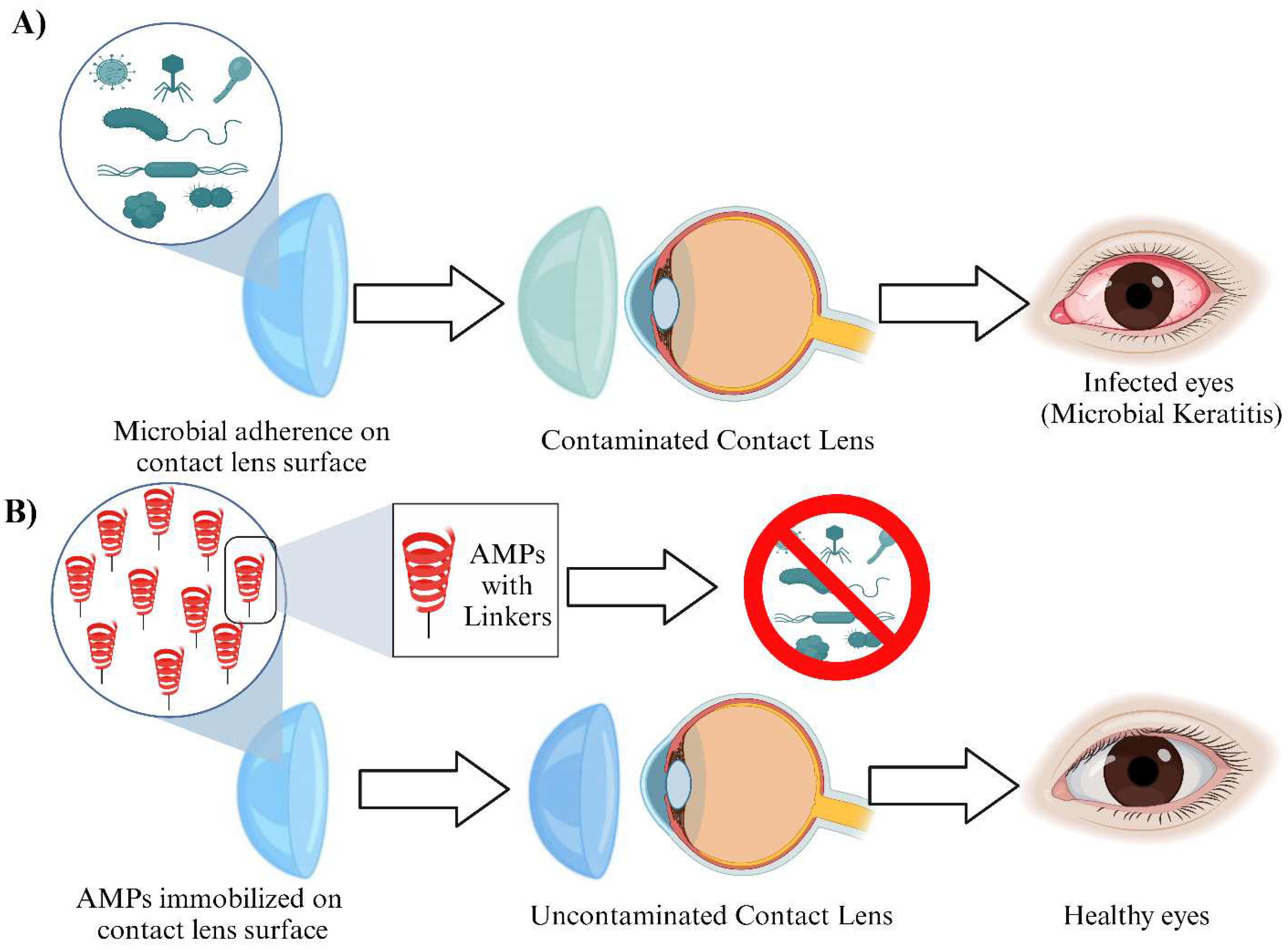
Figure 4.
Applications of AMPs in dental care and bone graft treatment: (A) Conventional implants are unable to repel microbial infections; (B) AMP-coated implants kill bacteria promoting oral health; (C) Tradition bone implants lack inherent anti-bacterial properties; (D) Titanium alloy implants with AMPs inhibit bacterial growth.
Figure 4.
Applications of AMPs in dental care and bone graft treatment: (A) Conventional implants are unable to repel microbial infections; (B) AMP-coated implants kill bacteria promoting oral health; (C) Tradition bone implants lack inherent anti-bacterial properties; (D) Titanium alloy implants with AMPs inhibit bacterial growth.
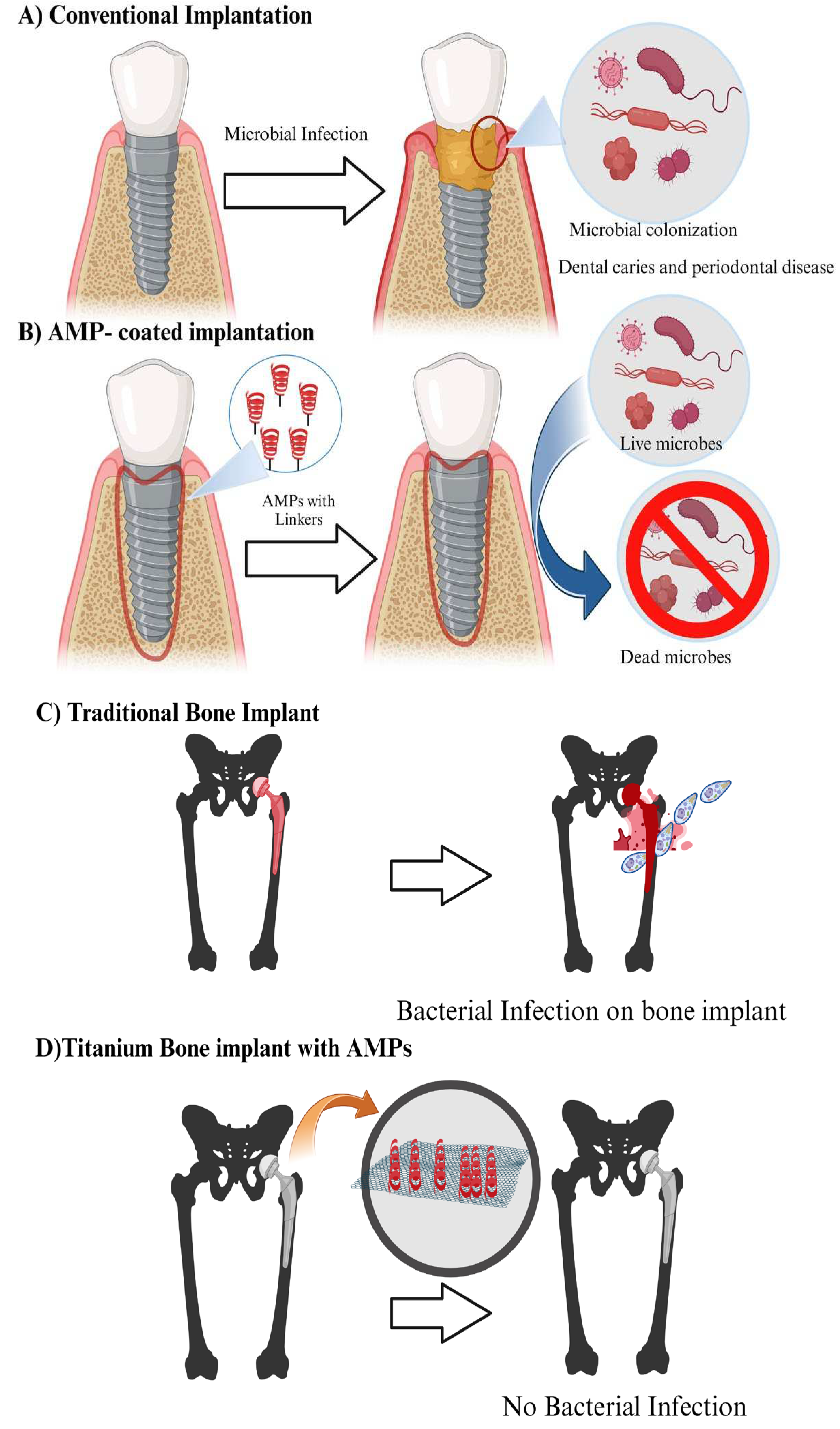
Figure 5.
AMPs to fight catheter-associated urinary tract infections (CAUTIs) (A) Illustration depicting the initiation of bacterial colonization, biofilm development, and subsequent dispersion on an unmodified urinary catheter surface. (B) In contrast, representation displaying anti-bacterial activity on an AMP immobilized urinary catheter surface.
Figure 5.
AMPs to fight catheter-associated urinary tract infections (CAUTIs) (A) Illustration depicting the initiation of bacterial colonization, biofilm development, and subsequent dispersion on an unmodified urinary catheter surface. (B) In contrast, representation displaying anti-bacterial activity on an AMP immobilized urinary catheter surface.
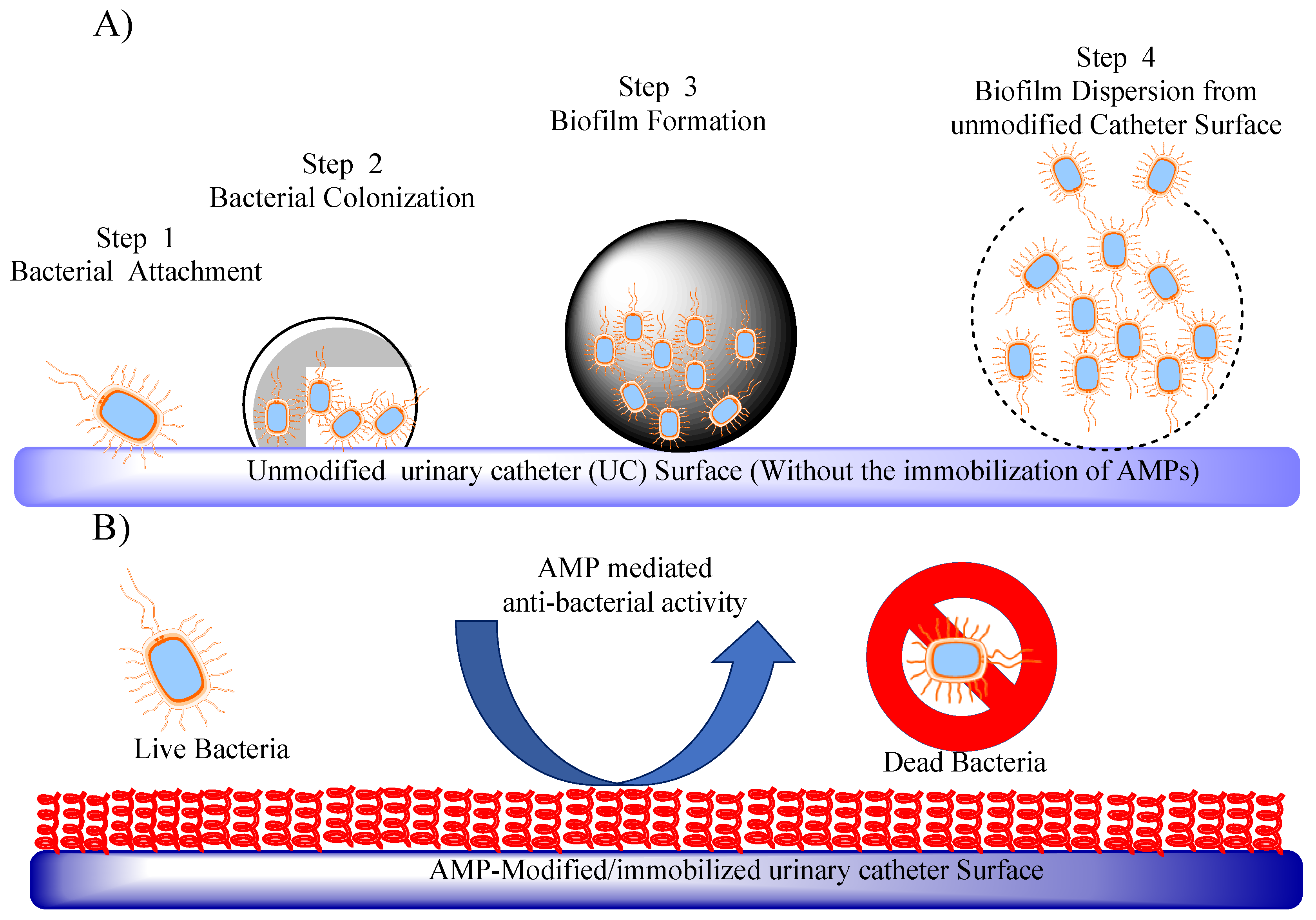
Figure 6.
Diverse Nanocarriers for Encapsulation and Immobilization of Antimicrobial Peptides (AMPs).
Figure 6.
Diverse Nanocarriers for Encapsulation and Immobilization of Antimicrobial Peptides (AMPs).
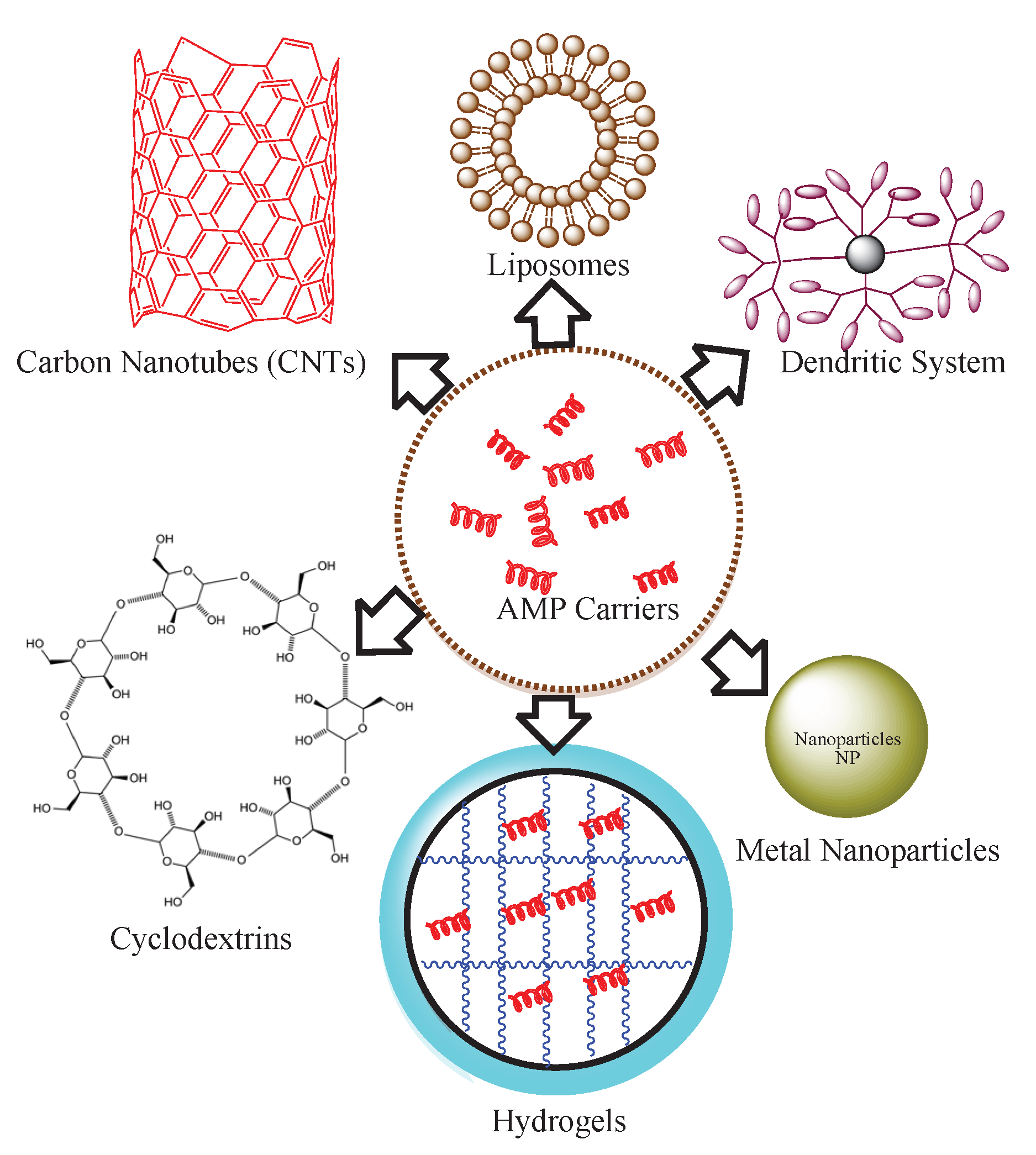

Table 1.
Antimicrobial Peptide Names and Corresponding Sequences Discussed in the Review.
| Sl. No | Peptide Name | Peptide Sequence |
|---|---|---|
| 1 | HNP-1 | ACYCRIPACIAGERRYGTCIYQGRLWAFCC |
| 2 | Drosocin | GKPRPYSPRPTSHPRPIRV |
| 3 | Melittin | GIGAVLKVLTTGLPALISWIKRKRQQ |
| 4 | LL-37 | LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES |
| 5 | HBD-2 | GIGDPVTCLKSGAICHPVFCPRRYKQIGTCGLPGTKCCKKP |
| 6 | HBD-3 | GIINTLQKYYCRVRGGRCAVLSCLPKEEQIGKCSTRGRKCCRRKK |
| 7 | Protegrin-1 | RGGRLCYCRRRFCVCVGR |
| 8 | Ranalexin | FLGGLIKIVPAMICAVTKKC |
| 9 | Pexiganan | GIGKFLKKAKKFGKAFVKILKK |
| 10 | α-MSH | SYSMEHFRWGKPV |
| 11 | Melimine | TLISWIKNKRKQRPRVSRRRRRRGGRRRR |
| 12 | Magainin 2 | GIGKFLHSAKKFGKAFVGEIMNS |
| 13 | Dermcidin | SSLLEKGLDGAKKAVGGLGKLGKDAVEDLESVGKGAVHDVKDVLDSV |
| 14 | Dermaceptin | ALWKTMLKKLGTMALHAGKAALGAAADTISQGTQ |
| 15 | Nisin A | ITSISLCTPGCKTGALMGCNMKTATCHCSIHVSK |
| 16 | Indolicidin | ILPWKWPWWPWRR |
| 17 | HBD-1 | DHYNCVSSGGQCLYSACPIFTKIQGTCYRGKAKCCK |
| 18 | HBD-5 | GLDFSQPFPSGEFAVCESCKLGRGKCRKECLENEKPDGNCRLNFLCCRQRI |
| 19 | Cateslytin | RSMRLSFRARGYGFR |
| 20 | GH-12 | GLLWHLLHHLLH |
| 21 | Myxinidin | GIHDILKYGKPS |
| 22 | HHC-36 | KRWWKWWRR |
| 23 | KLD | KLDLKLDLKLDL |
| 24 | E14LKK | LKLLKKLLKLLKKL |
| 25 | Dermaseptin-S4 | ALWMTLLKKVLKAAAKAALNAVLVGANA |
| 26 | Ib-AMP4 | QWGRRCCGWGPGRRYCRRWC |
| 27 | LLKKK18 | KEFKRIVKRIKKFLRKLV |
| 28 | DPK-060 | GKHKNKGKKNGKHNGWKWWW |
| 29 | SMAP-29 | RGLRRLGRKIAHGVKKYGPTVLRIIRIAG |
| 30 | MSI-78 | GIGKFLKKAKKFGKAFVKILKK |
| 31 | Bac2A | RLCRIVVIRVCR |
| 32 | Chain201D | KWIVWRWRFKR |
| 33 | E6 | RRWRIVVIRVRRC |
| 34 | Yao et al (Unnamed Peptide)[89] |
(RWRWRWC–NH2) |
| 35 | SESB2V | [(RGRKVVRR)2K]2KK |
| 36 | Temporin-1CEa | FVDLKKIANIINSIF |
| 37 | Esc(1-21) | GIFSKLAGKKIKNLLISGLKG-NH2 |
| 38 | 18-mer LLKKK | KLFKRIVKRILKFLRKLV |
| 39 | Thanatin | GSKKPVPIIYCNRRTGKCQRM |
| 40 | Histatins | Sequence Differs Across Subtypes With Conserved Cationic Nature |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



