
Submitted:
27 September 2023
Posted:
28 September 2023
You are already at the latest version
Abstract
Chalcones are a class of flavonoids possessing a variety of biological activities, including antimicrobial and antioxidant effects. Therefore, chalcones and their derivatives have potential use in the control of postharvest diseases. In this work, the effects of 2ʹ,4ʹ-dichloro-chalcone on the in vitro growth and in vivo pathogenicity of F. tricinctum and T. roseum were investigated. The results showed that 100 µM of 2ʹ,4ʹ-dichloro-chalcone strongly inhibited mycelial growth and conidial production of F. tricinctum (32.3%) and T. roseum (65.2%) in vitro. This treatment also significantly inhibited the expansion of potato dry rot from F. tricinctum (48.6%) and apple rot spot from T. roseum (36.2%). The incubation of F. tricinctum and T. roseum conidia with this agent for 2 h increased their cell membrane permeability by 25% and 22.5%, respectively and conidial membrane permeability by 41.7 and 65.4%, respectively. This treatment also significantly inhibited the total respiration rate and activated the cyanide-resistant respiratory pathway in both pathogens. Protein immunoblotting showed that the treatment increased alternative oxidase (AOX) levels after 4 h in F. tricinctum and T. roseum by 52.76% and 39.13%, respectively. Conclusion: 100 µM 2ʹ,4ʹ-dichloro-chalcone significantly inhibited mycelial growth and spore production in F. tricinctum and T. roseum in vitro and reduced their pathogenicity in postharvest potato and apple crops. Further analysis indicated that 2ʹ,4ʹ-dichloro-chalcone disrupted the integrity of cell membranes and the mitochondrial respiratory electron transport chain which leaded to ROS burst and oxidative stress, AOXs were induced directly or indirectly, and the cyanide resistant respiratory pathway was activated. The in vitro growth and pathogenicity of F. tricinctum and T. roseum were inhibited.
Keywords:
Fusarium tricinctum
; Trichothecium roseum
; chalcone
; cyanide-resistant respiration
; alternative oxidase
1. Introduction
Fusarium tricinctum and Trichothecium roseum are important fungal pathogens of various plant diseases worldwide. F. tricinctum infests many cereal crops, vegetables, and traditional Chinese medicinal plants including rice, wheat, barley, maize, potato, lily and licorice. [1,2,3,4,5,6]. T. roseum infests a wide range of fruits and vegetables such as apples, peaches, melons and tomatoes [7,8,9,10], causing severe preharvest and postharvest rots. Infection by F. tricinctum and T. roseum not only leads to a reduction in product quality, but also to increased food safety risks through their production of toxic and carcinogenic mycotoxins, including trichothecenes, zearalenones and fumonisins [11]. Currently, chemical fungicides are commonly used in the postharvest control of these two pathogens, which can have negative impacts on the development of fungal resistance and environmental pollution [12]. Therefore, there is a need for the development of alternative and environmentally safe means for postharvest disease prevention during storage of important crop products.
Chalcones are natural class of open-chain flavonoids that are widely found in edible or medicinal plants [13]. The presence of active α, β-unsaturated carbonyl functional groups (-CO-CH=CH-) in their molecular structure and the delocalized active π-electrons in the aryl ring provide for a variety of biological activities such as antioxidant, antitumor and anti-inflammatory effects [14]. Several studies have found that chalcones have a significant inhibitory activity against fungi and bacteria [15]. The phenolic groups in chalcones show high affinities for some microbial proteins and can therefore selectively inhibit microbial growth and development [16,17]. López et al. (2001) found that chalcone derivatives inhibited the activity of enzymes involved in yeast cell wall synthesis and significantly inhibited yeast growth [18]. López et al. (2020) found that chalcone had a strong inhibitory effect on root rot and fruit blight caused by Oomycetes [19]. Tsukiyama et al. (2002) isolated licorice chalcone A from licorice root and showed its significant inhibitory effect on the growth of Bacillus subtilis [6]. Natural chalcones and chalcone derivatives have emerged as preferred biomolecules that are safe and efficient alternatives to chemical fungicides. The antimicrobial effects of natural chalcones have motivated the study of chalcone structures and the synthesis of new chalcone derivatives for the enhancement of their antibacterial effects [20]. Wang et al. (2023) found that chalcones containing the triazole structure were all inhibitory to bacterial growth, particularly against Staphylococcus aureus [21]. Génesis et al. (2020) found that three allyl-structured chalcones had a strong inhibitory effect on the growth of the fungal pathogen, Mycobacterium avium [19]. Liu et al. (2017) synthesized halogenated chalcone aminothiourea Schiff bases with good inhibitory activities against tyrosinase and fungal growth [22]. Meaningfully, chalcone compounds are easy coatings with film-forming material. 3-Hydroxy-4-methoxy was prone to polymerize with high density polyethylene, low density polyethylene, polypropylene, and polyurethane, polymers form polymeric biofilms which presented significantly decrease of bacterial adhesion to the biofilms, thereby 3-Hydroxy-4-methoxy significantly inhibiting food contamination caused by foodborne pathogenic bacteria [23]. Phloretin containing dihydrochalcone structure was easy to polymerize with polyvinyl alcohol/polyacrylic acid to form a film, which significantly reduces the adhesion of bacteria on fresh cutting pork and has a good antibacterial effects [24]. However, the inhibitory effects of chlorinated chalcones on pathogenic fungi have been less characterized.
The objective of this study was to assess the effects of 2ʹ,4ʹ-dichloro-chalcone on the pathogenicities of F. tricinctum and T. roseum during the postharvest storage of potatoes and apples, respectively, and to provide mechanistic insight into its antifungal action. The results provide a meaningful theoretical basis for chalcone bioactive molecules to be used as film preservation materials.
2. Materials and Methods
2.1. Biological materials and test reagents
The F. tricinctum and T. roseum tested were isolated from potato tubers and apple fruits with typical symptoms of dry rot and mold heart disease, respectively by the post-harvest laboratory of the College of Food Science and Engineering, Gansu Agricultural University. Species identification of fungi was conducted by the ITS DNA sequences. Isolated fungal DNA was extracted with a fungal genomic DNA extraction Kit (Solarbio, Beijing, China) and PCR amplification of their ITS regions were carried out using the ITS gene- rot symptoms and from apples with typical specific primers ITS1 5'-TCCGTAGGTGAACCTGCGG-3', and ITS4 5'-TCCTCCGCTTATTGATATATGC-3' with reference to the method of Schoch et al. (2012) [25]. The sequencing results were queried against the National Center for Biotechnology Information (NCBI) database using BlastP [26], and sequence similarities of > 99% was used to choose the reference sequences. The phylogenetic analysis was conducted using MEGA 11.0 software (https://www.megasoftware.net) using ClustalW for sequence alignment and the neighbor joining (NJ) method for sequence clustering. The phylogenetic tree of F. tricinctum and T. roseum isolates are shown in Figure 1B and 1C. The identified pathogens were cultivated in potato dextrose agar (PDA) medium for future use.
Minitubers of potatoes (Solanum tuberosum L. cv. Atlantic) were purchased from Gansu Yihang Potato Industry Co., LtD in September 2022 and used to obtain mature tubers in a virus-free, mini potato planting greenhouse. Apple fruits tested (Malus Mill cv. Fuji) were purchased from the local market. The potatoes and apples were dried for 4 h, then refrigerated at 4 ± 1℃ with a relative humidity of (80±5)% to test for the development of F. tricinctum or T. roseum, respectively.
The 2ʹ,4ʹ-dichloro-chalcone (CAS No. 19672-60-7; molecular structure is shown in Figure 1A) and was purchased from Shanghai Yuanye Biological Company. Potassium cyanide (analytically pure) was obtained from Professor Shijun Bao (College of Animal Medicine, Gansu Agricultural University).
2.2. Methods
2.2.1. Configuration of 2ʹ,4ʹ-dichloro-chalcone stock solution and dosing medium preparation
A stock solution of 0.1 M 2ʹ,4ʹ-dichloro-chalcone, was prepared by dissolving 0.27 g in 500 μL of DMSO, 100 μL Tween 80 and 400 μL of anhydrous ethanol (Figure 2). The same solution without 2ʹ,4ʹ-dichloro-chalcone was prepared for use as a negative control. Both solutions were stored at 4 ℃ and used within 7 d.
2.2.2. Preparation of spore suspension of F. tricinctum and T. roseum
F. tricinctum and T. roseum were cultured on PDA medium for 7 d at 25℃ and their conidia collected by agitation in 10 mL sterile water for 15 s. The conidial suspension was filtered through two layers of sterile gauze, then counted using a hemocytometer and adjusted to 1 × 106 CFU/mL in sterile water for subsequent experiments.
2.2.3. Determination of the effects of 2ʹ,4ʹ-dichloro-chalcone on F. tricinctum and T. roseum mycelial growth and sporulation
Five μL of 1×106 CFU/mL spore suspension was inoculated onto solid PDA plates containing 0, 1, 10 or 100 µM 2ʹ,4ʹ-dichloro-chalcone and incubated at 28 ℃. The average diameter (n = 3) of each colony was measured every 24 h and the growth inhibition (%) of mycelial growth relative to the control was calculated according to the following formula:
The spore count from plates was determined with a hemocytometer as in section 2.2.2 after 7 d of culture (n = 5). The relative inhibition of sporulation was calculated from the following formula:
2.2.4. The effects of 2ʹ,4ʹ-dichloro-chalcone on hyphal cell membrane permeability
A 1 cm diameter sterile punch was used to collect three mycelial samples (about 1 g of mycelium per sample) that were rinsed in water, incubated in 10 mL deionized water for 0, 30, 60, 90 or 120 min, then centrifuged to collect the supernatant for conductivity measurements with a DS-307A conductivity meter (Shanghai Youke Instrument Co. Ltd., China). The cell membrane permeability was calculated according to the following formula:
2.2.5. Kinetics of oxygen consumption rates in total and cyanide resistant respiration in F. tricinctum and T. roseum
The oxygen consumption rates of total and cyanide resistant respiration were measured essentially according to Xu et al. (2021) [27]. Twenty μL of 106 CFU/mL spore suspensions were added into 30 mL potato dextrose broth (PDB) and cultured in an orbital shaker at 28℃ and 160 rpm (QYC-2102C, Shanghai Shengke Instrument Equipment Co. Ltd, China) at 28℃. After 24 h, 2ʹ,4ʹ-dichloro-chalcone was added to either 1, 10 or 100 µM and incubated further. Two mL of the culture solution was taken after a further 4 and 8 h for measurement of oxygen consumption rates with a Clark oxygen electrode (Oxygraph+, Hansha Scientific Instrument Co. Ltd., UK) at a constant temperature of 25 ℃ and expressed as nmol(O2)/mL/min. The total oxygen consumption rates without inhibitors (RIt) were determined. 1 mM KCN was added to determine the contribution from the cytochrome respiration pathway (RIcyt). 1 mM KCN and 1 mM salicylhydroxamic acid were added to measure the residual oxygen consumption rate (RIres). The rate of cyanide-resistant respiration (RIalt) was subsequently determined from RIcyt–Rires [28].
2.2.6. Protein immunoblotting analysis of AOX expression in F. tricinctum and T. roseum
To facilitate the collection of mycelia, 2 μL of 1×106 CFU/mL spore suspension was placed on cellophane in contact with PDA medium and incubated at 25℃ for 3−4 days. Protein extracts of the mycelium were prepared by incubating the collected mycelia in 50 mM Tris.HCl (pH 6.8) containing x mM -mercepto-ethanol and 2% (w/v) SDS at 40 ℃ for 30 min and clarified by centrifugation at 10000 g for 5 min. Proteins were separated by 12% SDS-PAGE and transferred to nitrocellulose membranes (0.22 mm pore size) using the procedure described by Affourtit [29]. Filters were washed in TBST and incubated for 1 h at room temperature with 1:1,000 anti-AOX and anti-actin monoclonal antibodies (Agrisera, Vännäs, Sweden). Following a wash in TBST, filters were incubated with HRP-labeled secondary antibody (Proteintech, Wuhan Sanying, Hubei, China) and the immunoblots developed with ECLTM Western Blotting Detection Reagents GE Healthcare (Sigma-Aldrich, St. Louis, MO, USA). The intensity of the detected bands was determined from their grey scale values AOX using Image J software (https://imagej.nih.gov/ij/) followed by normalization of AOX intensity with respect to that of the actin reference.
2.2.7. Measurement of ROS production in F. tricinctum and T. roseum
The production of ROS was detected based on the method of Xin et al. (2021) [30]. Briefly, 1 × 106 CFUs were introduced into 1 mL PDB containing either 0, 10 or 100 µM 2ʹ,4ʹ-dichloro-chalcone. After16 h of incubation, the fungal cultures were washed three times in PBS (pH 7) by centrifugation at 5,000 × g for 10 min, then resuspended in 30 µL of 30 µg/mL of dichloro-dihydrofluorescein diacetate (DCFH-DA) and incubated for 15 min at 28 ℃. The hypha were then washed twice by centrifugation, resuspended in 1 mL PBS (pH 7) for 1 h for signal development and the ROS levels determined by fluorescence spectrophotometry (U-LH100-3, Shanghai Yongke Optical Instrument Co. Ltd.) with 488 nm excitation and 500−600 nm emission filters.
2.2.8. Determination of F. tricinctum and T. roseum pathogenicities in vivo
The pathogenicity of F. tricinctum was determined in potato, while that of T. roseum was determined in apple using the methods established by Zhu et al. (2023) [31] and Ge et al. (2015) [32], respectively. Briefly, tubers and fruits of uniform size and no mechanical injuries or symptoms of disease were selected for testing. The selected samples of each were surface sterilized with 2% sodium hypochlorite solution for 2 min, rinsed in in running water then dried naturally. The halved potato and intact apple samples were immersed in sterile water containing 0, 1, 10 or 100 µM 2ʹ,4ʹ-dichloro-chalcone solution for 10 min, then air-dried for 4 h before inoculation with 20 μL of F. tricinctum or T. roseum spore suspension (1×106 CFU/mL), respectively, and incubation in sterile boxes (RH=70%) at room temperature in the dark. The expansion of the mycelial diameter (disease progression) was monitored and the symptoms of disease were photographed. from 2 to 20 d on a daily basis. The inhibition rate of disease diameter was calculated according to the following formula:
2.3. Statistical analysis
All experiments were repeated three times, and all data were calculated as mean and standard error (±SE). ANOVA tests utilized were used to test statistical significance where P< 0.05 was considered as significant.
3. Results
3.1. Effect of 2ʹ,4ʹ-dichloro-chalcone on mycelial growth of F. tricinctum and T. roseum
As shown in Figure 3, 2ʹ,4ʹ-dichloro-chalcone significantly inhibited mycelial growth of both pathogens on PDA medium (A, D). The effects on mycelial growth was low (2.5−7.2%) and insignificant at 1 and 10 µM, but clearly significant at 100 µM, inhibiting F. tricinctum by 32.3% after 7 d and T. roseum by 22.8% after 8d (P < 0.05) (B, C and E, F, respectively).
The hyphae of F. tricinctum cultivated on control PDA plates were slender and relatively highly branched, indicating a strong growth ability (G), while the mycelium grown under 100 µM 2ʹ,4ʹ-dichloro-chalcone (H) consisted of thinner and twisted hyphae with relatively fewer branches which were restricted to the hyphal apices, indicating a poorer growth ability. Under control conditions, T. roseum presented uniformly thin hyphae with a smooth, slender surface and a pointed apical tip (I). However, in the presence of 100 µM 2ʹ,4ʹ-dichloro-chalcone, the hyphae produced showed increased branching, but were of uneven thickness and showed blunt or enlarged apices (J). The results indicated that 2ʹ,4ʹ-dichloro-chalcone inhibited mycelium growth and colony formation in F. tricinctum and T. roseum with substantial effects on the hypha growth and morphology.
3.2. Effect of 2ʹ,4ʹ-dichloro-chalcone on spore production in F. tricinctum and T. roseum
Significant effects (P < 0.05) of 2ʹ,4ʹ-dichloro-chalcone on sporulation were observed at 100 µM for both pathogens, resulting in 64-65% decrease in conidial production from both pathogens (Figure 4). The lower concentrations of 2ʹ,4ʹ-dichloro-chalcone tested (1 or 10 µM) showed smaller (22-36%) and less significant effects.
3.3. Effect of 2ʹ,4ʹ-dichloro-chalcone on membrane permeability of F. tricinctum and T. roseum
The membrane permeability of intact hyphae can be measured through their effect on the conductivity of incubating solutions [33]. Figure 5 shows that 2ʹ,4ʹ-dichloro-chalcone at 100 µM significantly enhanced the membrane permeability (P < 0.05) in the two pathogens, while lower concentrations produced smaller and insignificant effects.
3.4. Effect of 2ʹ,4ʹ-dichloro-chalcone on the oxygen consumption rates in respiratory pathways of F. tricinctum and T. roseum
The total oxygen consumption rate largely reflects the activity of the respiratory pathway in aerobic fungi and the relative contributions from total and cyanide-resistant respiration is reflected in the electron distribution in the mitochondrial respiratory chain. Relative to control conditions, the total oxygen consumption rate in F. tricinctum and T. roseum was slightly reduced by 1 and 10 µM 2ʹ,4ʹ-dichloro-chalcone, but reduced by 32% and 25% at 100 µM, respectively (Figure 6A, D). In contrast, the oxygen consumption rate in the cyanide-resistant respiration pathway was increased by 42−43% in response to 100 µM 2ʹ,4ʹ-dichloro-chalcone treatment in the two pathogens (B, E). The cyanide-resistant respiration rate was significantly induced (P<0.05). For clarity, the ratio of cyanide-resistant respiration to total respiration is shown in panels C and F. The results indicated that 2ʹ,4ʹ-dichloro-chalcone significantly inhibited the total respiration rate but activated an increased contribution from cyanide-resistant respiration in both pathogens.
3.5. Effect of 2ʹ,4ʹ-dichloro-chalcone on the oxygen consumption rates in respiratory pathways of F. tricinctum and T. roseum
AOX is the terminal oxidase of the cyanide resistant respiratory pathway and its expression at the protein level reflects the activity of cyanide resistant respiratory pathway [34]. As shown in Figure 7, AOX was highly increased in response to 10 and 100 µM 2ʹ,4ʹ-dichloro-chalcone in both F. tricinctum and T. roseum. These results are consistent with the increased contribution of the cyanide-resistant respiration pathway to total respiration shown in Figure 6, panels B and E.
3.6. Effect of 2ʹ,4ʹ-dichloro-chalcone on ROS accumulation in F. tricinctum and T. roseum
The levels of intracellular ROS was assayed by fluorescence spectroscopy using the fluorescent probe, DCFH-DA. As shown in Figure 8, Relative to the control group, 100 µM 2ʹ,4ʹ- dichloro-chalcone significantly increased the intracellular ROS levels in F. tricinctum and T. roseum conidia by 41.7 and 65.4%, respectively, whereas 10 µM 2ʹ,4ʹ- dichloro-chalcone induced lesser, but significant increases of 25 and 38.2%, respectively. No significant effects were with 1µM 2ʹ,4ʹ- dichloro-chalcone.
3.7. Effect of 2ʹ,4ʹ-dichloro-chalcone on the pathogenicities of F. tricinctum and T. roseum
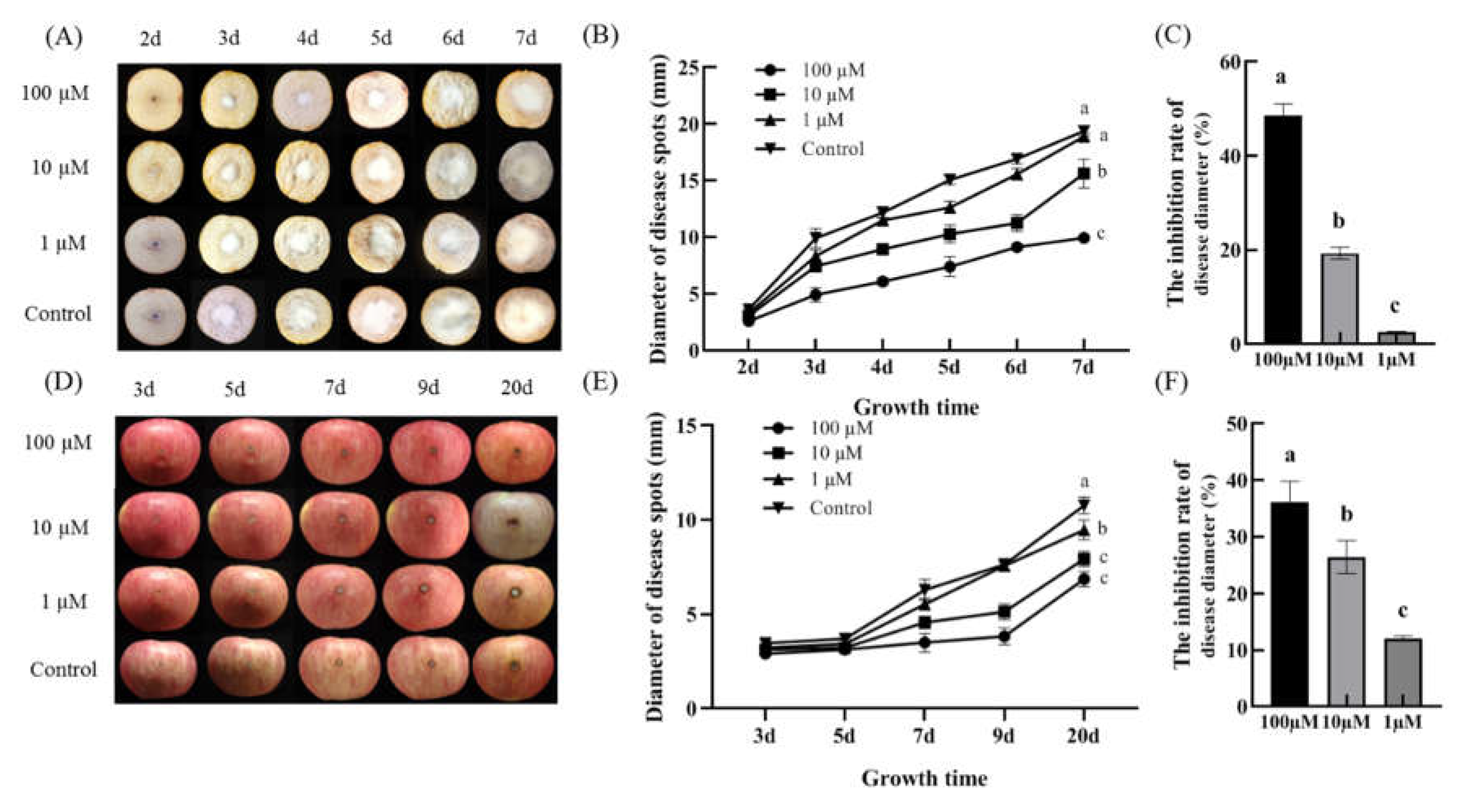
The effect of 2ʹ,4ʹ- dichloro-chalcone on the pathogenicities of F. tricinctum and T. roseum were assessed by colony growth (disease spot diameter) after their inoculation onto the cut surface of potatoes and apples, respectively. Under control conditions, F. tricinctum displayed faster growth kinetics, achieving an average disease spot size of 19.38 mm after 7 d (Figure 9A). In contrast, the development of T. roseum was relatively slower, achieving 10.77 mm after 20 d (D). The pre-inoculation treatment of potatoes and apples with 10 or 100 µM 2ʹ,4ʹ- dichloro-chalcone significantly inhibited disease spot development from both pathogens (A, D). After 7 d, 10 and 100 µM 2ʹ,4ʹ- dichloro-chalcone was observed to inhibit F. tricinctum spot development by 19.3 and 48.6%, respectively (B, C), while after 20 d, T. roseum spot diameter was reduced by 26.4 and 36.3%, respectively (E, F). The results indicate that 100 µM 2ʹ,4ʹ-dichloro-chalcone treatment could effectively reduce product damage from potato dry rot and apple rot spot by inhibiting their pathogenicity.
4. Discussion
Chalcones are members of the flavonoid family, are precursors for the synthesis of flavonoids and isoflavones in plants. Due to their content of active α and β unsaturated carbonyl functional groups, they are chemically very reactive, and have been used as safe and efficient alternative chemical fungicides for the control of postharvest diseases [35]. However, the fungicidal activity of chalcone derivatives with different substituents varies greatly. Previous studies have reported that chalcone derivatives with halogen substituents were highly fungicidal [36,37]. Therefore, the aim of this study was to investigate the inhibitory effect of 2ʹ,4ʹ-dichloro-chalcone on postharvest fungal pathogens, to explore its inhibitory mechanism and to provide a theoretical basis for the use of chlorinated chalcones in the postharvest preservation of fruits and vegetables.
Tests of 2ʹ,4ʹ - dichloro-chalcone against the pathogens F. tricinctum and T. roseum showed it had inhibitory effects on mycelium growth and sporulation (Figure 3 and Figure 4). This is consistent with previous research, which demonstrated that the growth of Candida glabrata and Trichophyton interdigitatum were inhibited by 12.5 µg/mL non-alkylated chalcone derivatives containing 2-bromine or 2-chloride subunits [38]. Kumar et al. (2013) also found that 0.1 mg/mL chalcone compounds containing p-fluorinated substituents in the benzene ring usually showed high antimicrobial activity [39]. However, our results indicated that 2ʹ,4ʹ-dichloro-chalcone showed significant inhibitory effects against F. tricinctum and T. roseum at 100 µM, which is substantially lower than the reported inhibitory concentrations required for these other halogenated chalcone derivatives.
Changes in the permeability of cell membranes in response to treatments can provide an important measure of membrane integrity [28]. In this study, 100 µM 2ʹ,4ʹ- dichloro-chalcone was observed to rapidly increase the cell membrane permeability of F. tricinctum and T. roseum conidia, indicating the cell membrane integrity was substantially compromised (Figure 5). It has been reported that phenolic chalcones can destroy the integrity of the cell membrane of Gram-negative and Gram-positive bacteria [40]. Chalcone compounds have also been reported to show high affinity to the cell membrane of Staphylococcus aureus and that their binding results in loss of cell membrane integrity [41].
In addition to normal respiration via the cytochrome respiratory pathway, the cyanide-resistant respiratory pathway can operate when normal respiration is compromised, in which AOX functions as the terminal mitochondrial oxidase in fungi. Studies have indicated that AOX was induced in unfavorable conditions to activate cyanide-resistant respiration as a survival mechanism [42]. AOX can also regulate the response of fungi to oxidative stress to reduce the oxidative stress damage to cells [43]. In addition, AOX also plays a role in determining the fungal susceptibility to some fungicides [44]. It has been shown that Quinol oxidation-inhibiting fungicides (such as Azoxystrobin, Kresoxim-methyl, Metominostrobin) acted on the fungal mitochondrial respiratory complex III, with inhibition of fungal growth, but that AOX expression affected this response [45]. Xu et al. (2013) reported that the expression of AOX in Sclerotinia sclerotiorum reduced its sensitivity to Azoxystrobin but could increase the sensitivity to Procymidone [46]. Consistent with these earlier reports, our results showed that the total respiratory rate of F. tricinctum and T. roseum was gradually inhibited after treatment with 100 µM 2ʹ,4ʹ- dichloro-chalcone, while the cyanide-resistant respiratory rate and AOX levels were increased (Figure 6 and Figure 7).
ROS are mainly generated by the mitochondrial respiratory chain in fungi, and unregulated ROS accumulation can enhanced the oxidative stress and damage to cell components, resulting in reduced cell viability [47,48]. ROS accumulation can also disrupt mitochondrial functions [49]. Some researchers have found that quinoline chalcone derivatives significantly induced ROS accumulation in Candida albicans, with damage to the mitochondrial membrane and inhibition of growth [50]. The 2-hydroxychalcone treatment can also promote the production of ROS in dermatophytes, leading to fungal cell apoptosis and necrosis [51]. Synthetic coumarin-chalcone was reported to inhibit thioredoxin reductase, induce significant ROS accumulation and activate the mitochondrial apoptosis pathway [52]. In this study, 2ʹ,4ʹ-dichloro-chalcone treatment was also found to promote ROS accumulation in F. tricinctum and T. roseum (Figure 8).
Zhan et al. (2016) found that the host contents of pyrimidinyl chalcones compounds reduced infection by Rhizoctorzia solani, Physolospora piricola, Fusarium graminearum, and Bipolaris maydis vitro [53]. Our results showed that 2ʹ,4ʹ-dichloro-chalcone treatment significantly reduced the pathogenicities of F. tricinctum and T. roseum in potato and apple, respectively (Figure 9).
AOX is the terminal oxidase of the cyanide resistant respiratory pathway in the fungal mitochondrial respiratory chain and can reduce mitochondrial dysfunction caused by the excessive production of ROS [54]. During excessive ROS is accumulation, oxidative and functional damage to the mitochondrial membrane can occur, ultimately resulting depressed respiration and overall physiological status of the cell [55], with a substantial decrease in the fungal growth rate and pathogenicity [56]. Singh et al. (2021) reported that an excessive intracellular ROS accumulation induces the expression of AOX and an increase in the cyanide-resistant respiration rate of Ascochyta rabiei, resulting in inhibition of the mycelial growth, spore production and cell vitality [57], which is consistent with our results. Therefore, 2ʹ,4ʹ-dichloro-chalcone affects the integrity of cell membranes, thereby affecting the mitochondrial respiratory electron transport chain, leading to a large production of ROS, directly or indirectly inducing the expression of AOX, activating the cyanide resistant respiratory pathway, and affecting bacterial growth and pathogenicity. The inhibition mechanism of chlorinated chalcones against pathogen fungi still needs to be further analyzed at the molecular level.
5. Conclusions
Our results showed that 100 µM 2ʹ,4ʹ-dichloro-chalcone significantly inhibited mycelial growth of F. tricinctum and T. roseum in vitro and in vivo. This occurred with significant reductions in sporulation, altered hypha morphology and reductions in cell membrane integrity. This treatment also enhanced intracellular ROS levels, together with higher levels of AOX and the activation of the alternative respiratory pathway, indicating respiratory stress. This study suggested that 2ʹ,4ʹ-dichloro-chalcone has significant antifungal activity against F. tricinctum and T. roseum. characteristics of film formation of 2ʹ,4ʹ-dichloro-chalcone and other chlorinated chalcones should be further investigated as alternative film-forming materials for the postharvest preservation of fruits and vegetables.
Author Contributions
Conceptualization, F.Z. and Y.Z. (Yan Zhu); methodology, W.Z., F.Z. and J.D.; software, W.S. and Y.S.; validation, W.Z. and P.W.; formal analysis, F.C. and Y.Z. (Yuanshou Zhao); investigation, F.Z. and Y.Z. (Yan Zhu).; resources, F.C.; data curation, W.S. and Y.L.; writing—original draft preparation, F.Z. and Y.Z. (Yan Zhu).; writing—review and editing, Y.Z. (Yan Zhu) and W.S.; visualization, W.Z. (Weibing Zhang) and J.D.; project administration, Y.Z. (Yan Zhu); funding acquisition, Y.Z. (Yan Zhu) and F.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation (31760473), the Young Doctor Fund Project of Gansu Provincial Education Department (2022QB-077), and the Excellent Postgraduate Innovation Star project of Gansu Province (2023).
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Dong, F.; Xu, J.H.; Zhang, X.; Wang, S.F.; Xing, Y.J.; Mokoena, M.P.; Olaniran, A.O.; Shi, J.R. Gramineous weeds near paddy fields are alternative hosts for the Fusarium graminearum species complex that causes fusarium head blight in rice. Plant Pathol. 2020, 69, 433–441. [Google Scholar] [CrossRef]
- Amato, B.; Pfohl, K.; Tonti, S.; Nipoti, P.; Prodi, A. Fusarium proliferatum and fumonisin B1 co-occur with Fusarium species causing Fusarium Head Blight in durum wheat in Italy. J Appl Bot Food Qual. 2015, 88, 228–233. [Google Scholar]
- Arino, A.A.; Bullerman, L.B. Fungal colonization of corn grown in nebraska in relation to year, genotype and growing conditions1. J. Food Prot. 1994, 57, 1084–1087. [Google Scholar] [CrossRef] [PubMed]
- Gachango, E.; Hanson, L.E.; Rojas, A.; Hao, J.J.; Kirk, W.W. Fusarium spp. causing dry rot of seed potato tubers in michigan and their sensitivity to fungicides. Plant Dis. 2012, 96, 1767–1774. [Google Scholar] [CrossRef] [PubMed]
- Lakshman, D.; Vieira, P.; Pandey, R.; Slovin, J.; Kamo, K. Symptom development in response to combined infection of in vitro-grown lilium longiflorum with pratylenchus penetrans and soilborne fungi collected from diseased roots of field-grown lilies. Plant Dis. 2017, 101, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Tsukiyama, R.I.; Katsura, H.; Tokuriki, N.; Kobayashi, M. Antibacterial activity of licochalcone A against spore-forming bacteria. Antimicrob. Agents Chemother. 2002, 46, 1226–1230. [Google Scholar] [CrossRef]
- Dai, P.; Jiang, Y.; Liang, X.; Gleason, M.L.; Sun, G. Trichothecium roseum enters 'fuji' apple cores through stylar fissures. Plant Dis. 2019, 104, 1060–1068. [Google Scholar] [CrossRef]
- May-De Mio, L.L.; Negri, G.; Michailides, T.J. Effect of Trichothecium roseum, lime sulphur and phosphites to control blossom blight and brown rot on peach. Can. J. Plant. Pathol. 2014, 36, 428–437. [Google Scholar]
- Gong, D.; Bi, Y.; Li, S.; Li, Y.; Wang, Y. Trichothecium roseum infection promotes ripening of harvested muskmelon fruits and induces the release of specific volatile compound. J Plant Pathol. 2019, 101, 529–538. [Google Scholar] [CrossRef]
- Ghazanfar, M.U.; Hussain, M.; Hamid, M.I.; Ansari, S.U. Utilization of biological control agents for the management of postharvest pathogens of tomato. Pak. J. Bot. 2016, 48, 2093–2100. [Google Scholar]
- Zhu, Y.H.; Shao, Y.Y.; Li, L.; Zhao, L.; Zhang, M.J.; Dong, C.M. The plant growth-promoting endophytic Fusarium oxysporum GG22 enhances Rehmannia glutinosa secondary metabolites accumulation. Ind. Crops Prod. 2022, 182, 114881. [Google Scholar] [CrossRef]
- Rizk, S.A.; Elsayed, G.A.; El-Hashash, M.A. One-pot synthesis, spectroscopic characterization and DFT study of novel 8-azacoumarin derivatives as eco-friendly insecticidal agents. J. Iran. Chem. Soc. 2018, 15, 2093–2105. [Google Scholar] [CrossRef]
- Ouyang, Y.; Li, J.J.; Chen, X.Y.; Fu, X.Y.; Sun, S.; Wu, Q. Chalcone Derivatives: Role in Anticancer Therapy. Biomolecules. 2021, 11, 894. [Google Scholar] [CrossRef] [PubMed]
- Cavusoglu, B.K.; Atli, O.; Gormus, G.; Ozkay, Y.; Kaplancikli, Z.A. Synthesis and Evaluation of Heterocycles Based Chalcone Derivatives as Anti-proliferative Agents. Anti-Cancer Agents Med. Chem. 2018, 18, 1044–1053. [Google Scholar] [CrossRef] [PubMed]
- Parikh, K.; Joshi, D. Antibacterial and antifungal screening of newly synthesized benzimidazole-clubbed chalcone derivatives. Med. Chem. 2013, 22, 3688–3697. [Google Scholar] [CrossRef]
- Zhuang, C.; Zhang, W.; Sheng, C.; Zhang, W.; Xing, C.; Miao, Z. Chalcone: A Privileged Structure in Medicinal Chemistry. Chem Rev. 2017, 117, 7762–7810. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Sood, A.K.; Goyal, K.; Singh, A.; Sharma, V.; Guliya, N.; Gulati, S.; Kumar, S. Chalcone Scaffolds as Anticancer Drugs: A Review on Molecular Insight in Action of Mechanisms and Anticancer Properties. Anti-Cancer Agents Med. Chem. 2021, 21, 1650–1670. [Google Scholar] [CrossRef] [PubMed]
- Lopez, S.N.; Cortes, J.C.; Ribas, J.C.; Castelli, M.V.; Zacchino, S.A.; Dominguez, J.N.; Lobo, G.; Charris-Charris, J.; Devia, C.; Rodriguez, A.M.; et al. In vitro antifungal evaluation and structure-activity relationships of a new series of chalcone derivatives and synthetic analogues, with inhibitory properties against polymers of the fungal cell wall. Bioorgan Med Chem. 2001, 9, 1999–2013. [Google Scholar] [CrossRef] [PubMed]
- Lopez, G.; Mellado, M.; Werner, E.; Said, B.; Godoy, P.; Caro, N.; Besoain, X.; Montenegro, I.; Madrid, A. Sonochemical Synthesis of 2'-Hydroxy-Chalcone Derivatives with Potential Anti-Oomycete Activity. Antibiotics-Basel. 2020, 9, 576. [Google Scholar] [CrossRef] [PubMed]
- Kakati, D.; Sarma, R.K.; Saikia, R.; Barua, N.C.; Sarma, J.C. Rapid microwave assisted synthesis and antimicrobial bioevaluation of novel steroidal chalcones. Steroids. 2013, 78, 321–326. [Google Scholar] [CrossRef]
- Wang, Z.W.; Sun, Y.J.; Zhang, L.; Zhang, P. Synthesis and Antifungal Activity of Chalcones Containing Triazole Moiety. Chin J Synth Chem. 2023, 31, 405–414. [Google Scholar]
- Liu, J.B. Bioactivities of chalcones with halogen atom and their thiosemicarbazide derivatives. Chemistry. 2017, 8, 77–83. [Google Scholar]
- Sivakumar, P.M.; Iyer, G.; Natesan, L.; Doble, M. 3 '-Hydroxy-4-methoxychalcone as a potential antibacterial coating on polymeric biomaterials. Appl. Surf. Sci. 2010, 256, 6018–6024. [Google Scholar] [CrossRef]
- Hu, Z. Preparation and properties of pH-responsive chalcone-loaded PVA/PAA functional films. Hefei, China, 2021.
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc Natl Acad Sci. 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lu, K.H.; Liu, X.; Zhu, Y.P.; Liu, C.H. Comparative Functional Genome Analysis Reveals the Habitat Adaptation and Biocontrol Characteristics of Plant Growth-Promoting Bacteria in NCBI Databases. Microbiol. Spectrum 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Copsey, A.C.; Young, L.; Barsottini, M.R.O.; Albury, M.S.; Moore, A.L. Comparison of the Kinetic Parameters of Alternative Oxidases from Trypanosoma brucei and Arabidopsis thaliana-A Tale of Two Cavities. Front. Plant Sci. 2021, 12, 744218. [Google Scholar] [CrossRef]
- Feng, H.Q.; Sun, K.; Wei, Y.; Wang, R.F.; Jia, L.Y.; Zhang, J.P.; Li, Y. Role of cyanide-resistant respiration during light-induced attraction of predators to herbivore-infested leaves. Photosynthetica. 2013, 51, 583–592. [Google Scholar] [CrossRef]
- Affourtit, C.; Moore, A.L. Purification of the plant alternative oxidase from Arum maculatum: measurement, stability and metal requirement. Acta Biochim. Biophys. Sin. 2004, 1608, 181–189. [Google Scholar] [CrossRef]
- Xin, Y.H.; Yang, D.; Yang, J.; Zhang, T.D.; Wang, Y.; Zhang, J.H.; Shi, Y.F. Effects of chalcone derivatives on mitochondrial structure and function of Aspergillus niger. Mycosystema. 2021, 40, 2144–2122. [Google Scholar]
- Zhu, Y.; Zhu, F.P.; Chen, F.; Shi, G.Y.; Zhao, Y.T.; Zhang, F.; Gong, W.J.; Zhang, W.; Wang, X.J.; Li, Y. C. Isolation, identification and pathogenicity of pathogens causing dry rot of stored potato tubers in Gansu Province. J Food Saf Qual. 2023, 14, 240–248. [Google Scholar]
- Ge, Y.H.; Deng, H.W.; Bi, Y.; Li, C.Y.; Liu, Y.Y.; Dong, B.Y. Postharvest ASM dipping and DPI pre-treatment regulated reactive oxygen species metabolism in muskmelon (Cucumis melo L.) fruit. Postharvest Biol. Technol. 2015, 99, 160–167. [Google Scholar] [CrossRef]
- Liu, J.J.; Hagberg, I.; Novitsky, L.; Hadj-Moussa, H.; Avis, T.J. Interaction of antimicrobial cyclic lipopeptides from Bacillus subtilis influences their effect on spore germination and membrane permeability in fungal plant pathogens. Fungal Biol. 2014, 118, 855–861. [Google Scholar] [CrossRef] [PubMed]
- El-Khoury, R.; Rak, M.; Benit, P.; Jacobs, H.T.; Rustin, P. Cyanide resistant respiration and the alternative oxidase pathway: A from to mammals. Bba-Bioenergetics. 2022, 1863, 148567. [Google Scholar] [CrossRef] [PubMed]
- Yadav, N.; Dixit, S.K.; Bhattacharya, A.; Mishra, L.C.; Sharma, M.; Awasthi, S.K.; Bhasin, V.K. Antimalarial activity of newly synthesized chalcone derivatives in vitro. Chem Biol Drug Des. 2012, 80, 340–347. [Google Scholar] [CrossRef]
- Shakil, N.A.; Singh, M.; Kumar, J.; Sathiyendiran, M.; Kumar, G.; Singh, M.; Pandey, R.P.; Pandey, A.; Parmar, V.S. Microwave synthesis and antifungal evaluations of some chalcones and their derived diaryl-cyclohexenones. J Environ Sci Heal B. 2010, 45, 524–530. [Google Scholar] [CrossRef]
- Shakhatreh, M.A.; Al-Smadi, M.L.; Khabour, O.F.; Shuaibu, F.A.; Hussein, E.I.; Alzoubi, K.H. ; Study of the antibacterial and antifungal activities of synthetic benzyl bromides, ketones, and corresponding chalcone derivatives. Drug Des Dev Ther. 2016, 10, 3653–3660. [Google Scholar] [CrossRef]
- Kucerova-Chlupacova, M.; Vyskovska-Tyllova, V.; Richterova-Finkova, L.; Kunes, J.; Buchta, V.; Vejsova, M.; Paterova, P.; Semelkova, L.; Jandourek, O.; Opletalova, V. Novel halogenated pyrazine-based chalcones as potential antimicrobial drugs. Molecules. 2016, 21, 1421. [Google Scholar] [CrossRef] [PubMed]
- Kumar, C.; Loh, W.S.; Ooi, C.; Quah, C.; Fun, H.K. Heteroaryl chalcones: design, synthesis, x-ray crystal structures and biological evaluation. Molecules 2013, 18, 12707–12724. [Google Scholar] [CrossRef] [PubMed]
- Erguden, B.; Unver, Y. Phenolic chalcones lead to ion leakage from gram-positive bacteria prior to cell death. Arch. Microbiol. 2021, 204, 3. [Google Scholar] [CrossRef]
- Tagousop, C.N.; Tamokou, J.D.; Ekom, S.E.; Ngnokam, D.; Voutquenne-Nazabadioko, L. Antimicrobial activities of flavonoid glycosides from Graptophyllum grandulosum and their mechanism of antibacterial action. Bmc Complem Altern M. 2018, 18, 252. [Google Scholar] [CrossRef]
- Jian, W.; Zhang, D.W.; Zhu, F.; Wang, S.X.; Zhu, T.; Pu, X.J.; Zheng, T.; Feng, H.; Lin, H.H. ; Nitrate reductase-dependent nitric oxide production is required for regulation alternative oxidase pathway involved in the resistance to Cucumber mosaic virus infection in Arabidopsis. Plant Growth Regul. 2015, 77, 99–107. [Google Scholar] [CrossRef]
- Xu, T.; Yao, F.; Liang, W.S.; Li, Y.H.; Li, D.R.; Wang, H.; Wang, Z.Y. Involvement of alternative oxidase in the regulation of growth, development, and resistance to oxidative stress of Sclerotinia sclerotiorum. J. Microbiol. 2012, 50, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; Parr-Dobrzanski, B. The strobilurin fungicides. Pest. Manage. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Banno, S.; Yamashita, K.; Fukumon, F.; Okada, K.; Uekusa, H.; Takagaki, M.; Kimura, M.; Fujimura, M. ; Characterization of QoI resistance in Botrytis cinerea and identification of two types of mitochondrial cytochrome b gene. Plant Pathol. 2009, 58, 120–129. [Google Scholar] [CrossRef]
- Xu, T.; Wang, Y.T.; Liang, W.S.; Yao, F.; Li, Y.H.; Li, D.R.; Wang, H.; Wang, Z.Y. Involvement of alternative oxidase in the regulation of Sclerotinia sclerotiorum sensitivity to fungicides of azoxystrobin and procymidone. J Microbiol. 2013, 51, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Ban, X.; Zeng, H.; He, J.; Chen, Y.; Wang, Y. The mechanism of antifungal action of essential oil from dill (Anethum graveolens L.) on Aspergillus flavus. PLoS One 2012, 7, 30147. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, D.; Kondo, K.; Uehara, N.; Otokozawa, S.; Tsuji, N.; Yagihashi, A.; Watanabe, N. Endogenous reactive oxygen species is an important mediator of miconazole antifungal effect. Antimicrob. Agents Chemother. 2002, 46, 3113–3117. [Google Scholar] [CrossRef]
- Pozniakovsky, A.I.; Knorre, D.A.; Markova, O.V.; Hyman, A.A.; Skulachev, V.P.; Severin, F.F. Role of mitochondria in the pheromone- and amiodaroneinduced programmed death of yeast. J. Cell Biol. 2005, 168, 257–269. [Google Scholar] [CrossRef]
- Chai, N.N.; Sun, A.M.; Zhu, X.H.; Li, Y.P.; Wang, R.R.; Zhang, Y.; Mao, Z.W. Antifungal evaluation of quinoline-chalcone derivatives combined with FLC against drug-resistant Candida albicans. Bioorg Med Chem Lett. 2023, 86, 129242. [Google Scholar] [CrossRef]
- Bila, N.M.; Costa-Orlandi, C.B.; Vaso, C.O.; Bonatti, J.L.C.; Regasini, L.O.; Fontana, C.R.; Fusco-Almeida, A.M. 2-Hydroxychalcone as a Potent Compound and Photosensitizer Against Dermatophyte Biofilms. Front. Cell. Infect. Microbiol. 2021, 11, 679470. [Google Scholar]
- Wang, Y.; Zhang, W.; Dong, J.; Gao, J. Design, synthesis and bioactivity evaluation of coumarin-chalcone hybrids as potential anticancer agents. Bioorg. Chem. 2019, 95, 103530. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.S.; Jin, H.; Tao, T.; Hou, T.P. Synthesis and Anti-fungal Activity of Chalcones with Pyridyls. J. Shangdong Agric. Univ. 2016, 47, 166–171. [Google Scholar]
- Helmerhorst, E.J.; Stan, M.; Murphy, M.P.; Sherman, F.; Oppenheim, F.G. The concomitant expression and availability of conventional and alternative, cyanide-insensitive, respiratory pathways in Candida albicans. Mitochondrion. 2005, 5, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.; Serrano, M.; L'Haridon, F.; Tjamos, S.E.; Metraux, J.P. Reactive oxygen species and plant resistance to fungal pathogens. Phytochemistry. 2015, 112, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Marschall, R.; Tudzynski, P. A new and reliable method for live imaging and quantification of reactive oxygen species in Botrytis cinerea Technological advancement. Fungal Genet Biol. 2014, 71, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.; Nair, A.M.; Verma, P.K. Surviving the odds: From perception to survival of fungal phytopathogens under host-generated oxidative burst. Plant Commun. 2021, 2, 100142. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chemical structure of 2ʹ,4ʹ-dichloro-chalcone (A); Phylogenetic trees of F. tricinctum (A) and T. roseum isolates (B).
Figure 1.
Chemical structure of 2ʹ,4ʹ-dichloro-chalcone (A); Phylogenetic trees of F. tricinctum (A) and T. roseum isolates (B).
Figure 2.
The preparation of the chalcone stock solution and preparation of PDA medium containing the agent.
Figure 2.
The preparation of the chalcone stock solution and preparation of PDA medium containing the agent.
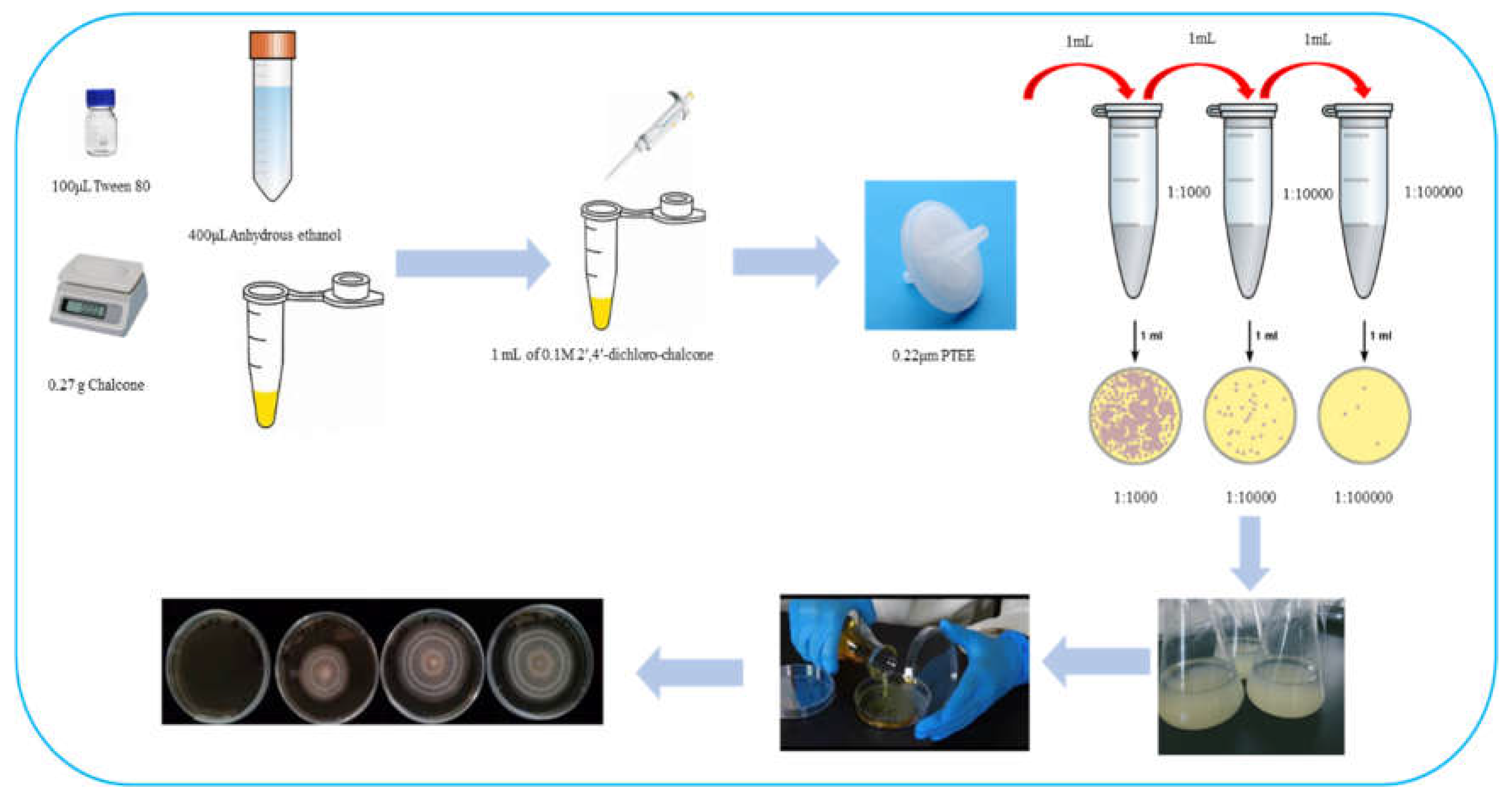
Figure 3.
The effects of 2ʹ,4ʹ-dichloro-chalcone on colony growth and hypha morphology of F. tricinctum (A-C, G, H) and T. roseum (D-F, I, J). (A, D) Representative examples of colony sizes after 7 d growth. (B, E) Mycelial growth curves (diameter). (C, F) The calculated growth inhibition by 2ʹ,4ʹ-dichloro-chalcone. Representative examples of mycelial morphology grown under control conditions (G, I) and with 100 µM 2ʹ,4ʹ-dichloro-chalcone (H, J). The error bars in panels in panels B-F represent the standard error of three replicate assays. Different letters above the bars represent significant differences between treatments (P < 0.05).
Figure 3.
The effects of 2ʹ,4ʹ-dichloro-chalcone on colony growth and hypha morphology of F. tricinctum (A-C, G, H) and T. roseum (D-F, I, J). (A, D) Representative examples of colony sizes after 7 d growth. (B, E) Mycelial growth curves (diameter). (C, F) The calculated growth inhibition by 2ʹ,4ʹ-dichloro-chalcone. Representative examples of mycelial morphology grown under control conditions (G, I) and with 100 µM 2ʹ,4ʹ-dichloro-chalcone (H, J). The error bars in panels in panels B-F represent the standard error of three replicate assays. Different letters above the bars represent significant differences between treatments (P < 0.05).
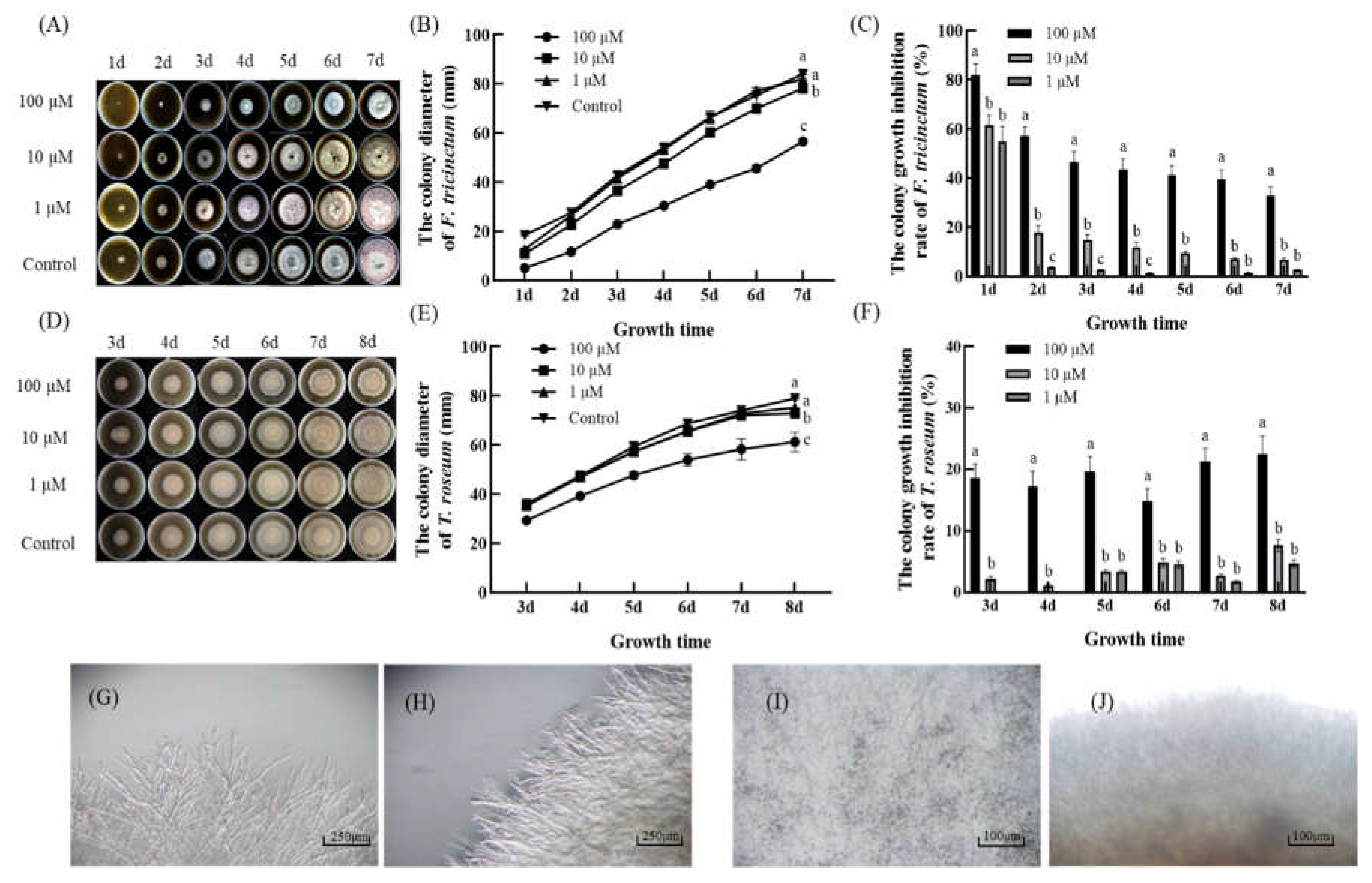
Figure 4.
The effect of 2ʹ,4ʹ-dichloro-chalcone on sporulation in F. tricinctum (A, B) and T. roseum (C, D). (A, C) Spore production capacity under control conditions. (B, D) Inhibition of sporulation (%). Vertical bars indicate the standard error of three replicate assays. The error bars in panels in panels A-D represent the standard error of three replicate assays. Different letters above the bars represent significant differences between treatments (P < 0.05).
Figure 4.
The effect of 2ʹ,4ʹ-dichloro-chalcone on sporulation in F. tricinctum (A, B) and T. roseum (C, D). (A, C) Spore production capacity under control conditions. (B, D) Inhibition of sporulation (%). Vertical bars indicate the standard error of three replicate assays. The error bars in panels in panels A-D represent the standard error of three replicate assays. Different letters above the bars represent significant differences between treatments (P < 0.05).
Figure 5.
The effect of 2ʹ,4ʹ-dichloro-chalcone on membrane permeability in F. tricinctum and T. roseum. (A) F. tricinctum. (B) T. roseum. Vertical bars indicate the standard error of three replicate assays. Different letters above the bars represent significant differences between treatments (P < 0.05).
Figure 5.
The effect of 2ʹ,4ʹ-dichloro-chalcone on membrane permeability in F. tricinctum and T. roseum. (A) F. tricinctum. (B) T. roseum. Vertical bars indicate the standard error of three replicate assays. Different letters above the bars represent significant differences between treatments (P < 0.05).
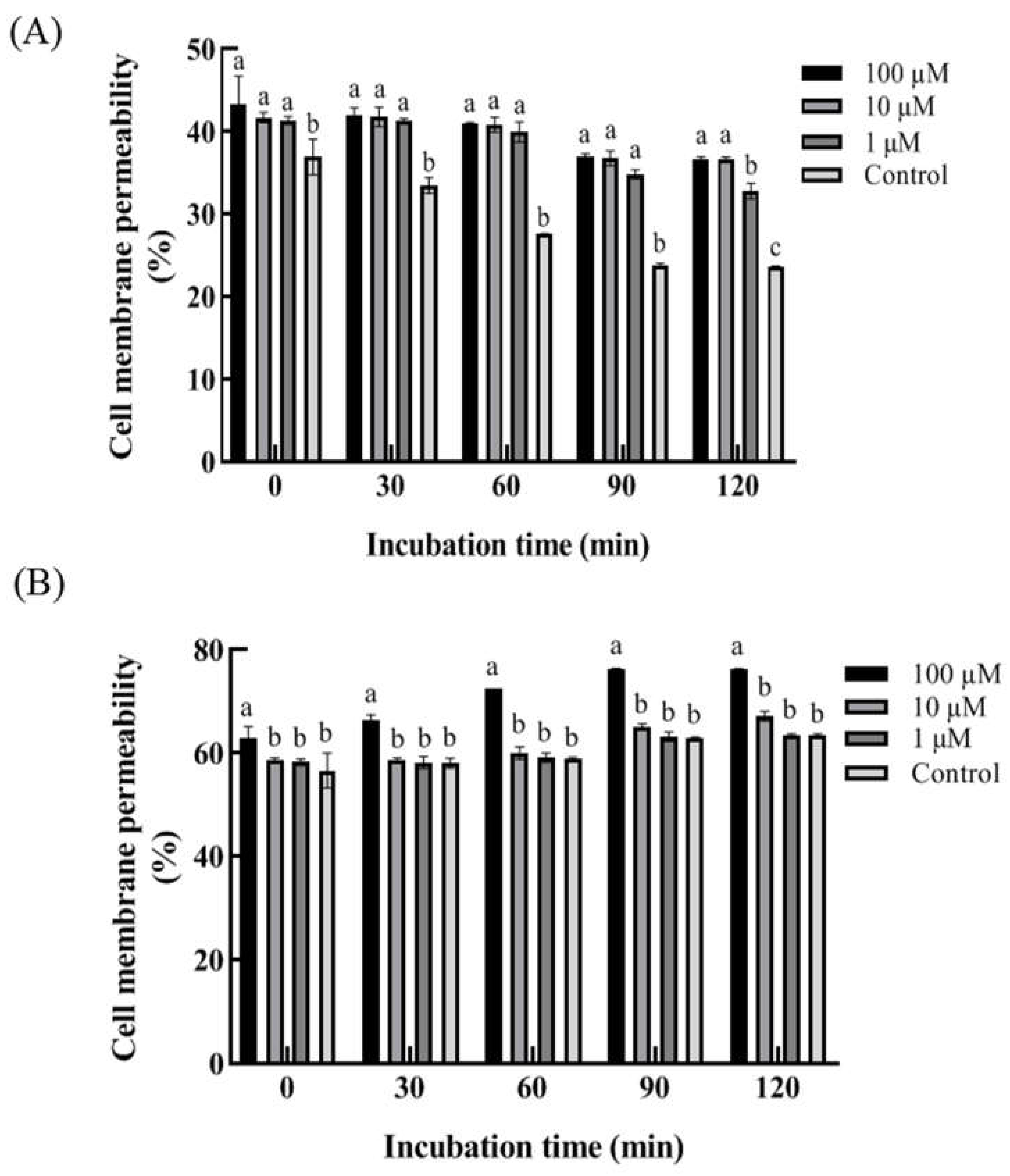
Figure 6.
The effect of 2ʹ,4ʹ-dichloro-chalcone on oxygen consumption rates in total and cyanide-resistant respiration pathways in F. tricinctum (A, D) and T. roseum (B, E). For clarity, the ratios of cyanide-resistant respiration to total respiration in F. tricinctum and T. roseum are shown in panels C and F, respectively. Vertical bars indicate the standard error of three replicate assays. Different letters above the bars represent significant differences between treatments (P < 0.05).
Figure 6.
The effect of 2ʹ,4ʹ-dichloro-chalcone on oxygen consumption rates in total and cyanide-resistant respiration pathways in F. tricinctum (A, D) and T. roseum (B, E). For clarity, the ratios of cyanide-resistant respiration to total respiration in F. tricinctum and T. roseum are shown in panels C and F, respectively. Vertical bars indicate the standard error of three replicate assays. Different letters above the bars represent significant differences between treatments (P < 0.05).
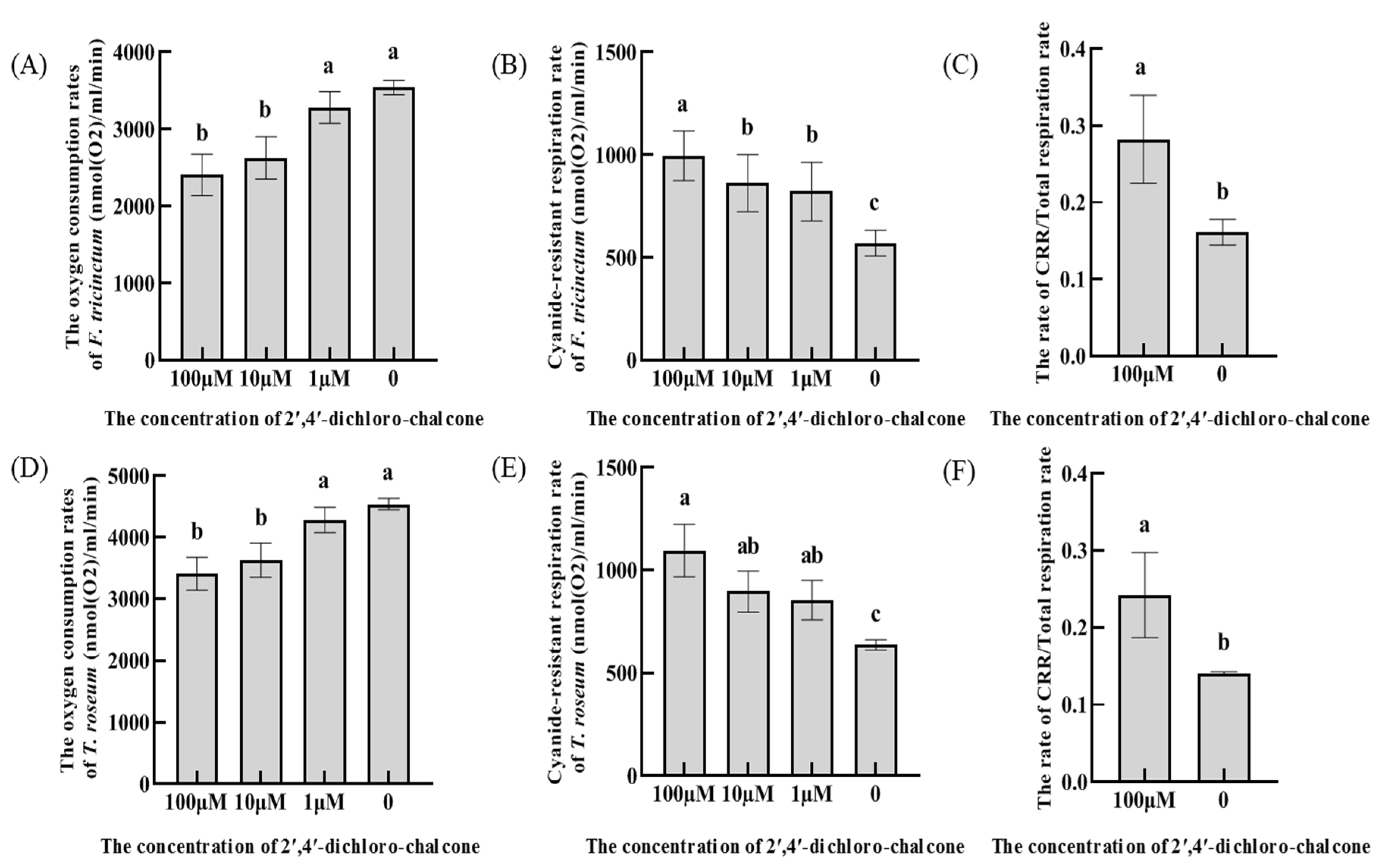
Figure 7.
The effect of 2ʹ,4ʹ-dichloro-chalcone on AOX levels in F. tricinctum (A, B) and T. roseum (C, D). The immunoblots are shown in panels A and C. The grey-scale values of AOX are shown in panels B and D. The values presented were normalized with respect to actin. Vertical bars indicate the standard error of three replicate assays. Different letters above bars indicate significant differences between the treatments (P < 0.05).
Figure 7.
The effect of 2ʹ,4ʹ-dichloro-chalcone on AOX levels in F. tricinctum (A, B) and T. roseum (C, D). The immunoblots are shown in panels A and C. The grey-scale values of AOX are shown in panels B and D. The values presented were normalized with respect to actin. Vertical bars indicate the standard error of three replicate assays. Different letters above bars indicate significant differences between the treatments (P < 0.05).
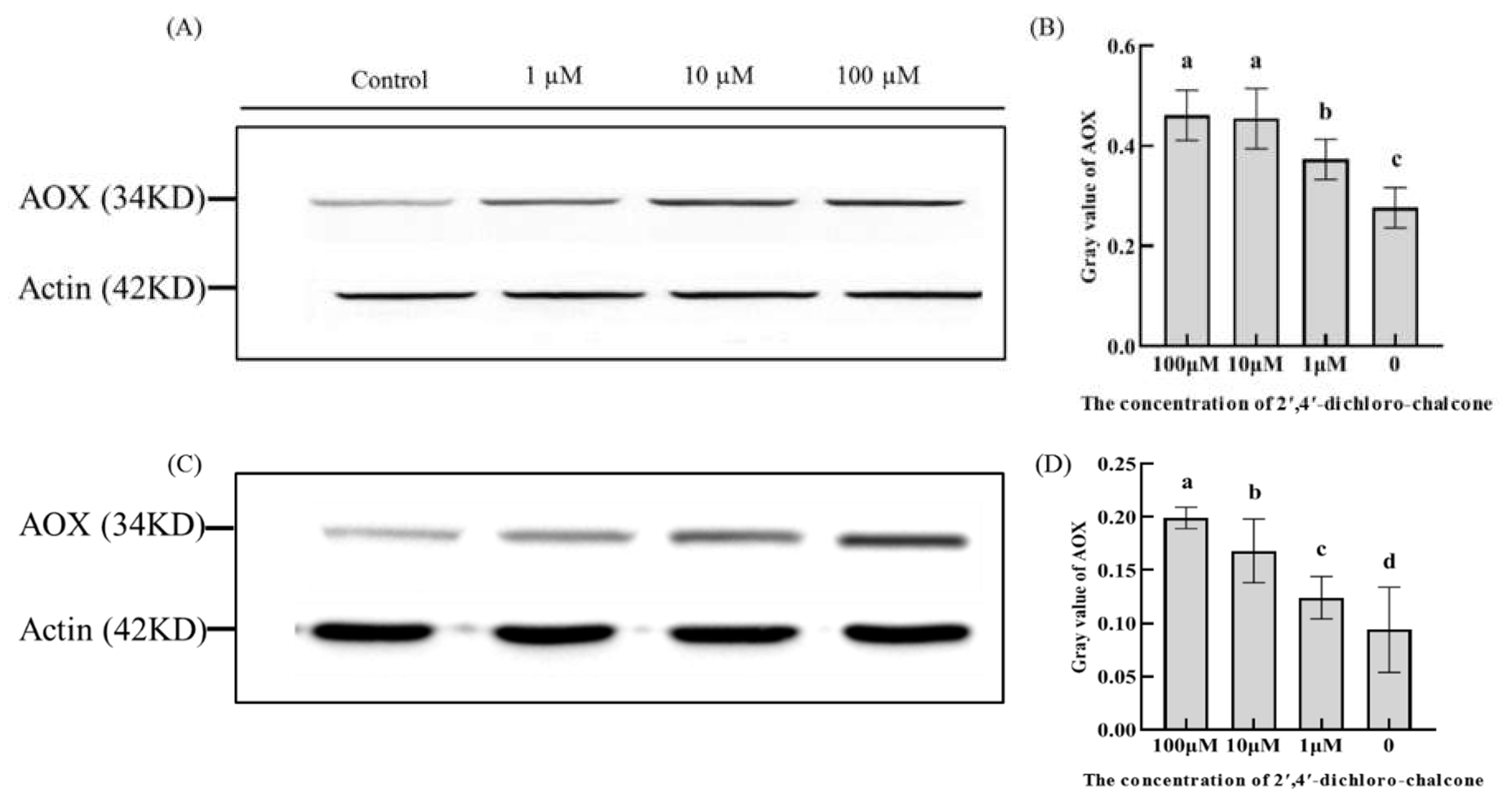
Figure 8.
The effect of 2ʹ,4ʹ- dichloro-chalcone on intracellular ROS levels in conidia of F. tricinctum (A) and T. roseum (B). ROS were detected using DCFH-DA as a fluorescent probe. Vertical bars indicate the standard error of three replicate assays. Differing letters above bars indicate significantly differences between the treatments (P < 0.05).
Figure 8.
The effect of 2ʹ,4ʹ- dichloro-chalcone on intracellular ROS levels in conidia of F. tricinctum (A) and T. roseum (B). ROS were detected using DCFH-DA as a fluorescent probe. Vertical bars indicate the standard error of three replicate assays. Differing letters above bars indicate significantly differences between the treatments (P < 0.05).
Figure 9.
The effect of 2ʹ,4ʹ- dichloro-chalcone treatment on disease spot development from F. tricinctum (A-C) and T. roseum (D-F). (A, D) The diameter of disease spots. (B, E) The inhibition of disease spot development. (C, F) The effect of 2ʹ,4ʹ- dichloro-chalcone treatment on the disease spots. Vertical bars indicate the standard error of three replicate assays. Differing letters above each time point represent significant differences (P < 0.05).
Figure 9.
The effect of 2ʹ,4ʹ- dichloro-chalcone treatment on disease spot development from F. tricinctum (A-C) and T. roseum (D-F). (A, D) The diameter of disease spots. (B, E) The inhibition of disease spot development. (C, F) The effect of 2ʹ,4ʹ- dichloro-chalcone treatment on the disease spots. Vertical bars indicate the standard error of three replicate assays. Differing letters above each time point represent significant differences (P < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



